
Structure and Function of
Cytochromes P450 2B and 3A:
From Mechanism-Based
Inactivators to X-Ray Crystal
Structures and Back
James R. HalpertSkaggs School of Pharmacy & Pharmaceutical
Sciences
University of California, San Diego

To determine the structural basis of mammalian P450 specificity in order to rationalize and predict metabolism of xenobiotics
Tools (1979) Tools (2010)
Purified rat and rabbit hepatic P450s Heterologously expressed
Chemical modification mammalian P450s
Absorbance spectroscopy Site-directed mutagenesis
Functional assays X-ray crystallography
NMR
Homology models
Molecular dynamics simulations
Pressure-perturbation spectroscopy
Isothermal titration calorimetry
Hydrogen-deuterium exchange
FRET
Long-term Objective

Goals of Predictive Drug
Metabolism
• Overcome metabolic lability of a new
chemical entity (NCE)
• Avoid drug interactions caused by an
NCE
• Predict functional consequences of
SNP's (individuals, populations)
• Predict pharmacokinetics of an NCE

Overview
1. Species, strain, and individual
differences in P450 2B function
2. Conformational plasticity of cytochromes
P450 2B
3. Multiplicity of cytochromes P450 3A in
various species
4. Cooperativity of human P450 3A4
5. Lessons learned

Species, Strain, and Individual
Differences in P450 2B Function
• Mechanism-based inactivation
• Steroid hydroxylation
• PCB oxidation

P450 2B Enzymes
1. Highly inducible by phenobarbital in a number of species (human, rat, rabbit, mouse, dog)
2. Among the first P450s purified and characterized (LM2, PB-B/P450b)
3. Metabolize angular, medium-sized neutral or basic compounds
4. Exhibit marked species and strain differences in function

• Chloramphenicol is converted by P450 2B1 to an oxamyl chloride, which binds to the ε-amino group of one or more lysine residues in the enzyme.
• The modified enzyme is unable to undergo enzymatic reduction by NADPH-cytochrome P450 reductase.
Halpert, J. (1981). Covalent modification of lysine during the suicide inactivation
of rat liver cytochrome P-450 by chloramphenicol. Biochem. Pharmacol.
30:875-881.
TIME (min)
14
C(c
pm
)
Halpert, J., Miller, N., and Gorsky, L. (1985). On the mechanism of the
inactivation of the major phenobarbital-inducible isozyme of rat liver
cytochrome P-450 by chloramphenicol. J. Biol. Chem. 260:8397-8403.

• Chloramphenicol inactivates three (2B1, 2C6, 2C11) of eight
major rat liver microsomal cytochrome P450 enzymes.
Halpert, J., Balfour, C., Miller, N.E., Morgan, E.T., Dunbar, D., and Kaminsky,
L.S. (1985) Isozyme-selectivity of the inhibition of rat liver cytochromes P-450
by chloramphenicol in vivo. Mol. Pharmacol. 28:290-296.

• N-(2-p-nitrophenethyl)chlorofluoroacetamide is a
selective inactivator of P450 2B1 in vitro and in vivo.
Halpert, J., Jaw, J.-Y., Balfour, C., and Kaminsky, L.S. (1990). Selective
inactivation by chlorofluoroacetamides of the major phenobarbital-inducible
form(s) of rat liver cytochrome P-450. Drug Metab. Dispos. 18:168-174.

• A Gly-478 to Ala substitution in P450 2B1 from Wistar-Munich rats is responsible for the low androgen 16 β-hydroxylase activity and lack of susceptibility to inactivation by N-(2-p-nitrophenethyl)-chlorofluoroacetamide.
Kedzie, K.M., Balfour, C.A., Escobar, G.Y., Grimm, S.W., He, Y.-A., Pepperl, D.,
Regan, J., Stevens, J.C., and Halpert, J.R. (1991). Molecular basis for a
functionally unique cytochrome P450IIB1 variant. J. Biol. Chem. 266:22515-22521.
WM IIB1
1616
% 1
6-H
ydro
xyla
se A
ctivity
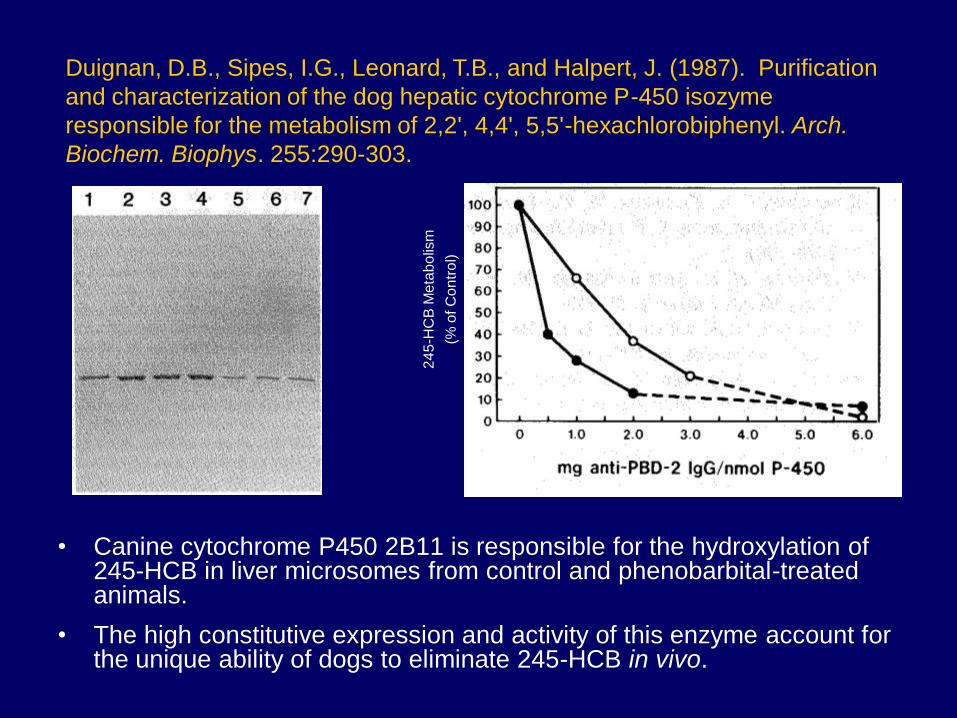
• Canine cytochrome P450 2B11 is responsible for the hydroxylation of 245-HCB in liver microsomes from control and phenobarbital-treated animals.
• The high constitutive expression and activity of this enzyme account for the unique ability of dogs to eliminate 245-HCB in vivo.
Duignan, D.B., Sipes, I.G., Leonard, T.B., and Halpert, J. (1987). Purification
and characterization of the dog hepatic cytochrome P-450 isozyme
responsible for the metabolism of 2,2', 4,4', 5,5'-hexachlorobiphenyl. Arch.
Biochem. Biophys. 255:290-303.
24
5-H
CB
Me
tab
olis
m
(% o
f C
on
tro
l)
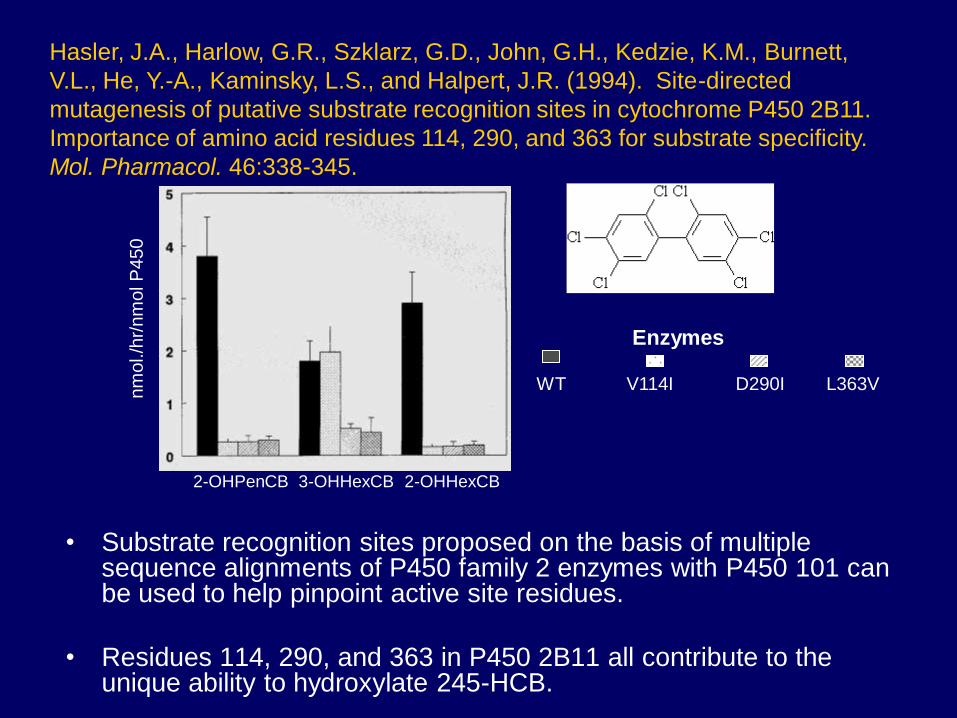
• Substrate recognition sites proposed on the basis of multiple sequence alignments of P450 family 2 enzymes with P450 101 can be used to help pinpoint active site residues.
• Residues 114, 290, and 363 in P450 2B11 all contribute to the unique ability to hydroxylate 245-HCB.
Hasler, J.A., Harlow, G.R., Szklarz, G.D., John, G.H., Kedzie, K.M., Burnett,
V.L., He, Y.-A., Kaminsky, L.S., and Halpert, J.R. (1994). Site-directed
mutagenesis of putative substrate recognition sites in cytochrome P450 2B11.
Importance of amino acid residues 114, 290, and 363 for substrate specificity.
Mol. Pharmacol. 46:338-345.
WT V114I D290I L363V
Enzymes
2-OHPenCB 3-OHHexCB 2-OHHexCB
nm
ol./h
r/nm
ol P
450

• A homology model of P450 2B1 based on three bacterial P450 x-ray crystal structures can be used to explain alterations in steroid hydroxylation upon side-chain substitution at 10 putative active site positions.
Szklarz, G.D., He, Y.A., and Halpert, J.R. (1995). Site-directed
mutagenesis as a tool for molecular modeling of cytochrome P450 2B1.
Biochemistry 34:14312-14322.
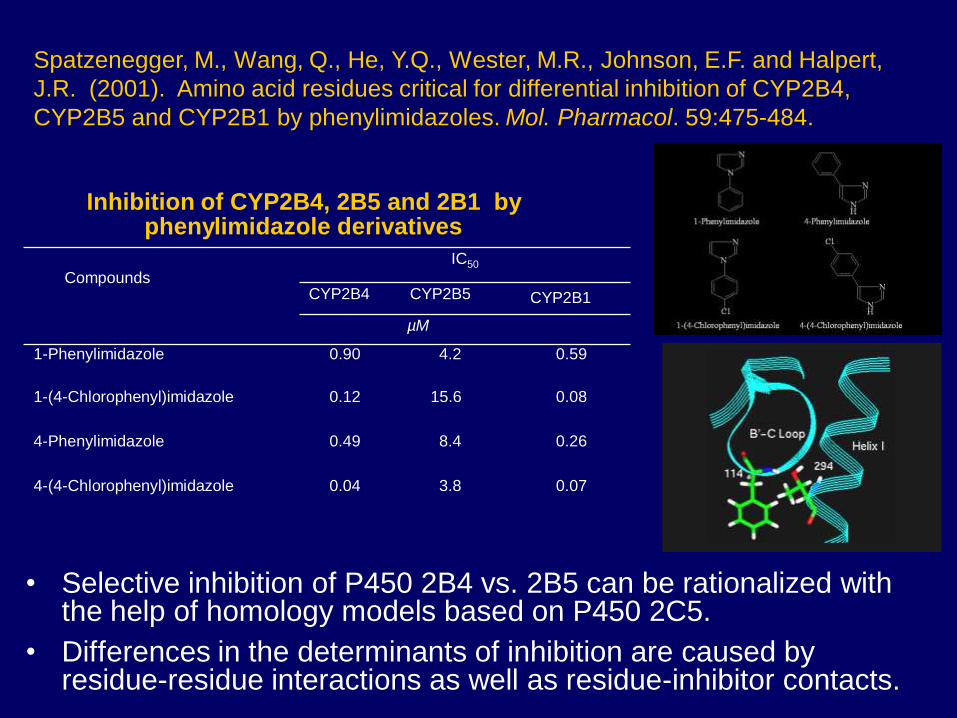
• Selective inhibition of P450 2B4 vs. 2B5 can be rationalized with the help of homology models based on P450 2C5.
• Differences in the determinants of inhibition are caused by residue-residue interactions as well as residue-inhibitor contacts.
Inhibition of CYP2B4, 2B5 and 2B1 by phenylimidazole derivatives
Compounds
IC50
CYP2B4 CYP2B5 CYP2B1
µM
1-Phenylimidazole 0.90 4.2 0.59
1-(4-Chlorophenyl)imidazole 0.12 15.6 0.08
4-Phenylimidazole 0.49 8.4 0.26
4-(4-Chlorophenyl)imidazole 0.04 3.8 0.07
Spatzenegger, M., Wang, Q., He, Y.Q., Wester, M.R., Johnson, E.F. and Halpert,
J.R. (2001). Amino acid residues critical for differential inhibition of CYP2B4,
CYP2B5 and CYP2B1 by phenylimidazoles. Mol. Pharmacol. 59:475-484.

• P450 2B4 is an excellent candidate for crystallization trials.
Scott, E.E., Spatzenegger, M., and Halpert, J.R. (2001). A truncation of
2B subfamily cytochromes P450 yields increased expression levels,
increased solubility, and decreased aggregation while retaining function.
Arch. Biochem. Biophys. 395:57-68.
2B P450
% P
450
Extr
acte
d2B1dH 2B4dH 2B6dH 2B11dH

Conformational Plasticity of P450
2B Enzymes
• X-ray crystallography of rabbit P450 2B4
• Hydrogen-deuterium mass exchange
studies of 2B4
• X-ray crystallography of human P450 2B6

highly concentrated protein
solution
(+ salt, detergent, DTT,
EDTA, buffer . . .)
+
precipitant
(EtOH, Na citrate pH 5.5) sitting drop
vapor diffusion
Control of Stability/Solubility
Crystallization
• protein engineering:
1 10 20 30 490
2B4 MALLLAVLLA FLAGLLLLLF RGHPKAHGRL PPGP . . . AR
2B4dH MA KKTSSKGKL PPGP . . . ARHHHH
Underline indicates alteration from the native sequence.• salt • detergent
protein + precipitant
precipitant
• The engineering and purification approaches elaborated for P450
2C5 enable 2B4 crystallization.

Cytochrome P450 2B4 Dimer
molecule 1
molecule 2
• In the absence of exogenous ligand, P450 2B4 crystallized
as a dimer, which allowed trapping of an open form of the
enzyme and helps explain ligand access to the active site.

Location of Active Site Residues in the 2B4 Open
Conformation vs. a Closed 2B5 Structure
2B4 2C5-DMZ
active site
residues

Goal: Substrate/Inhibitor-Bound Complex
Approaches to Discourage Homodimer Formation:
1. Protein engineering: H226Y
2. Choose soluble, high-affinity ligand:
3. Form protein-ligand complex early by adding inhibitor prior to
protein concentration
Cl
N
N
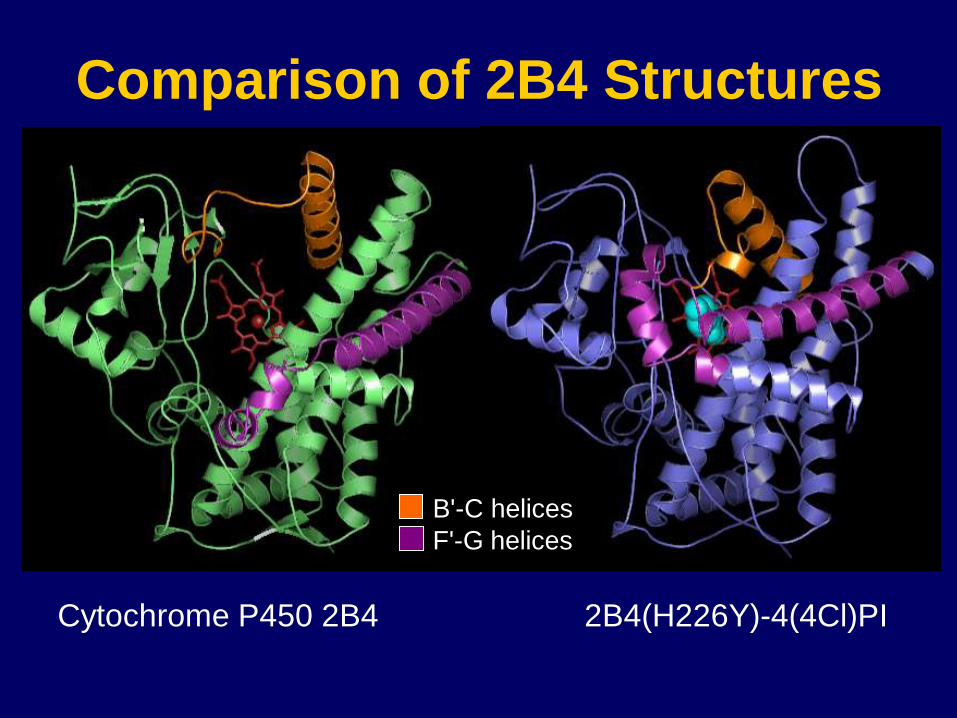
Cytochrome P450 2B4
Comparison of 2B4 Structures
B'-C helices
F'-G helices
2B4(H226Y)-4(4Cl)PI

2B4
Conclusions: Conformational Shift May Accommodate Sequential Substrate Binding and
Catalysis
cleft closed for regio- and
stereoselective metabolism
2B4(H226Y)-4(4Cl)PI
cleft open for substrate
binding/metabolite release

P450 Plasticity
Definition
• Involves the large-scale motions required forsubstrate access and product egress
• Includes the reshaping of the active site to adaptto ligands of different geometry
Questions
• Do the large conformational changes inferredfrom crystal structures occur in solution?
• What are the implications of plasticity forprediction of ligand binding?

Imidazole Inhibitors of P450 2B4
N
N
N
N1-BPI
1-PBI
N
N
1-TI
NCl
N
1-CPI
N
N
1-BI
N
N
Bifonazole
N
N
1-BHI
Cl
N
NH
4-CPI
N
N
Cl
Clotrimazole
N
N
1-(Dibiphenyl)MeI

Comparison of Five 2B4 Structures

Pivoting of the F-G Helix Casette to
Accommodate the Various
Inhibitors

• Remove aliquot for time point
• Quench on ice (0°C)
• Aliquot replicates
• Place all vials in dry ice to flash freeze
• Store samples in -80°C freezer
DX-MS Setup (Exchange)
Garcia, Pantazatos, and Villarreal. Assay and drug development
technologies (2004) 2(1): 81-91.

DX-MS Setup (MS)
Samples quickly thawed to 0°C from -80°C.
Garcia, Pantazatos, and Villarreal. Assay and drug development
technologies (2004) 2(1): 81-91.

2B4dH Exchange (1000s)B’-C Region F-G Cassette
2B4 No Ligand
B’-C Region F-G Cassette
2B4 + 4-CPI
B’-C Region F-G Cassette
2B4 + 1-PBI
B’-C Region F-G Cassette
Difference (4-
CPI vs. No L)
Difference in # of Deuterons

Oxidation by Human P450 2B6 of
Antiplatelet Drugs

CYP2B4:Ticlopidine Complex Ribbon Diagram
F
G
A
NH2
COOH
F’
G’
B’
C
I
H
E
D
K
J
Ticlopidine

T302
E301
F206
I363
A298
F297
S210I209
V477
F115
I101
I114
V367
Ticlopidine Bound in the Active Site of P450 2B4
in the Opposite Orientation Predicted from 2B6

Structure of Ticlopidine and the % Difference
in Paramagnetic Relaxation Rate Relative to
the Slowest Proton

Crystallization of t-BPA Labeled 2B4
• Proposed mechanism of formation of tBPA-2B4 adduct (Zhang, et al. (2009), Mol Pharm)
• Collaboration with Hollenberg Lab (University of Michigan)


Molecular Replacement
• Maps generated using 2B4-1-CPI as a MR search model
• E301 is pointed out of active site.
• Density appears to agree with docking of tBPA by Zhang, et al.
E301
T302

Induced-fit (1) vs. Conformational Selection (2) Models of P450 2B4
DX-MS suggests that an open conformation of ligand-free
enzyme predominates in solution. However, we recently
solved a structure of a closed ligand-free form of P450 2B4.

Properties of Human P450 2B6
– Major catalyst of oxidation of bupropion,
cyclophosphamide, and efavirenz
– Highly polymorphic (Q172H and K262R of
particular importance)
– Represents 1-5% of total hepatic P450
– Along with CYP3A5 one of the few human
drug metabolizing P450s yet to be crystallized
– Low thermal stability

Engineering Cytochrome P450 2B6 for Enhanced
Expression and Stability
Temperature (oC)
30 35 40 45 50 55 60 65
% A
cti
vit
y
0
20
40
60
80
100
2B1dH
2B4dH
2B6dH
2B11dHx column 5 vs y column 5
x column 6 vs y column 6
x column 7 vs y column 7
x column 8 vs y column 8
Temperature (OC)
30 35 40 45 50 55 60 65 70 75
% P
450
0
20
40
60
80
100
Temperature (oC)
30 35 40 45 50 55 60 65
% A
cti
vit
y
0
20
40
60
80
100
2B1dH
2B4dH
2B6dH
2B11dHx column 5 vs y column 5
x column 6 vs y column 6
x column 7 vs y column 7
x column 8 vs y column 8
Temperature (oC)
30 35 40 45 50 55 60 65
% A
cti
vit
y
0
20
40
60
80
100
2B1dH
2B4dH
2B6dH
2B11dHx column 5 vs y column 5
x column 6 vs y column 6
x column 7 vs y column 7
x column 8 vs y column 8
P450P450 (HS, LS, P420) Activity
Tm ( C) T50 ( C)
2B1dH 61.3 0.2 52.7 0.3
2B4dH 58.9 0.3 50.8 0.4
2B6dH 48.2 0.2 45.2 0.4
2B11dH 51.4 0.3 48.4 0.2
P450 nmol/L
Rat 2B1dH 800 - 1200
Rabbit 2B4dH 300 - 500
Human 2B6dH 50 - 75
Dog 2B11dH 400 -600

Approaches to Enable
Crystallization of Human P450 2B6
• N-terminal modification and use of
chaperones
• Internal mutations to improve stability
(Y226H) and solubility (K262R)
• Different CM-resin
• Replacement of glycerol by sucrose after
chromatography
• Inclusion of facial amphiphiles during trials

P450 2B6:4-CPI Complex Structure (2 Å)

Structural Alignment of P450 2B4
and 2B6 4-CPI Complexes
G
F
CH
I
G’
AB’
F’
F
G
I
AH
E
D
C
F’
G’
2B6 = Green; 2B4 = Magenta
The RMSD in a Cα overlay of 2B6 and 2B4 is only 0.65 Å.

2B6 Active Site Density/2B6 and
2B4 Active Site Overlay
F115
I101
V104I209
F206
E301
T302
A298
F297
I114
V367
V477
L363/I363
2Fo-Fc map contoured at 1-σ
Only residue 363 and orientation of E301 differ between the active sites.

Summary
1. P450 2B4 exhibits remarkableconformational flexibility with imidazolesof different size and shape as evidencedby X-ray crystallography and solutionmethods.
2. Despite ~100 amino acid differences, theX-ray crystal structures of the 4-CPIcomplexes of P450 2B4 and 2B6 arealmost identical.
3. How do we reconcile these twoseemingly contradictory findings?

Acknowledgments
Dr. Chris Chin Dr. Eric Johnson
Dr. Sean Gay Dr. Dave Stout
You-ai He Dr. Mike Wester
You Qun He Dr. Qinghai Zhang
Dr. Santosh Kumar Dr. Sheng Li
Dr. Hong Liu Dr. Tong Liu
Dr. Keiko Maekawa Dr. Virgil Woods
Dr. B.K. Muralidhara Dr. Paul Hollenberg
Dr. Art Roberts Dr. Haoming Zhang
Dr. Emily Scott ES003619 (JRH)
Dr. Manish Shah ES006676 (JRH)
Dr. Margit Spatzenegger GM31001 (EFJ)
Ling Sun GM59229 (CDS)
Dr. Jyothi Talakad CA016954 (PFH)
Dr. Qinmi Wang
Dr. Mark White
Dr. Ross Wilderman
Dr. Yonghong Zhao

Multiplicity of P450 3A enzymes
• Purification of multiple rat P450 3A
enzymes
• cDNA cloning of two canine P450 3A
enzymes
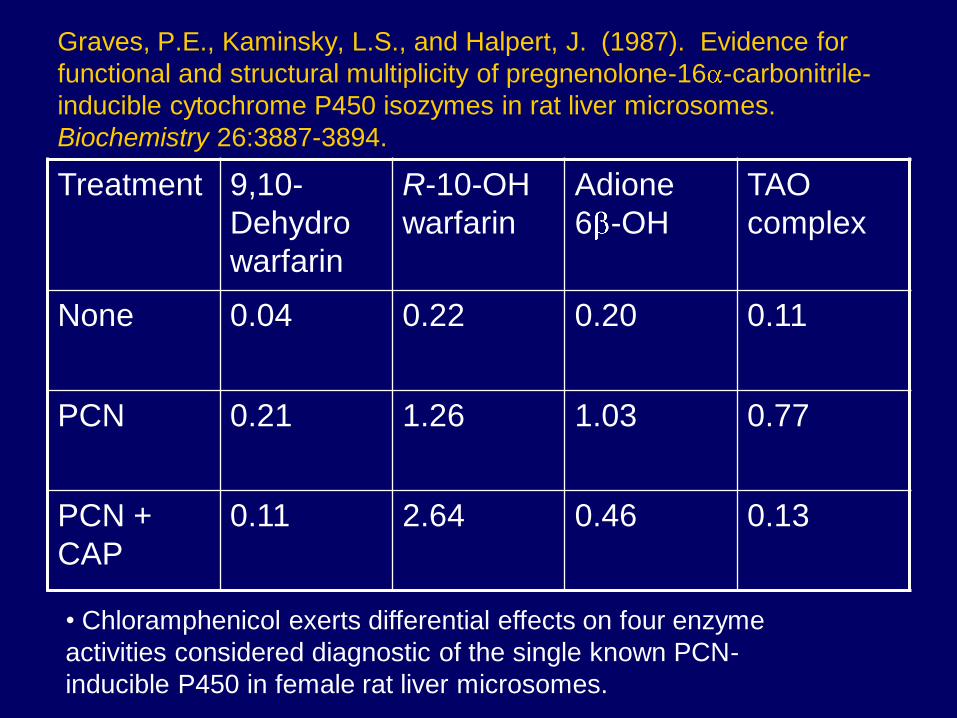
Graves, P.E., Kaminsky, L.S., and Halpert, J. (1987). Evidence for
functional and structural multiplicity of pregnenolone-16 -carbonitrile-
inducible cytochrome P450 isozymes in rat liver microsomes.
Biochemistry 26:3887-3894.
Treatment 9,10-
Dehydro
warfarin
R-10-OH
warfarin
Adione
6 -OH
TAO
complex
None 0.04 0.22 0.20 0.11
PCN 0.21 1.26 1.03 0.77
PCN +
CAP
0.11 2.64 0.46 0.13
• Chloramphenicol exerts differential effects on four enzyme
activities considered diagnostic of the single known PCN-
inducible P450 in female rat liver microsomes.
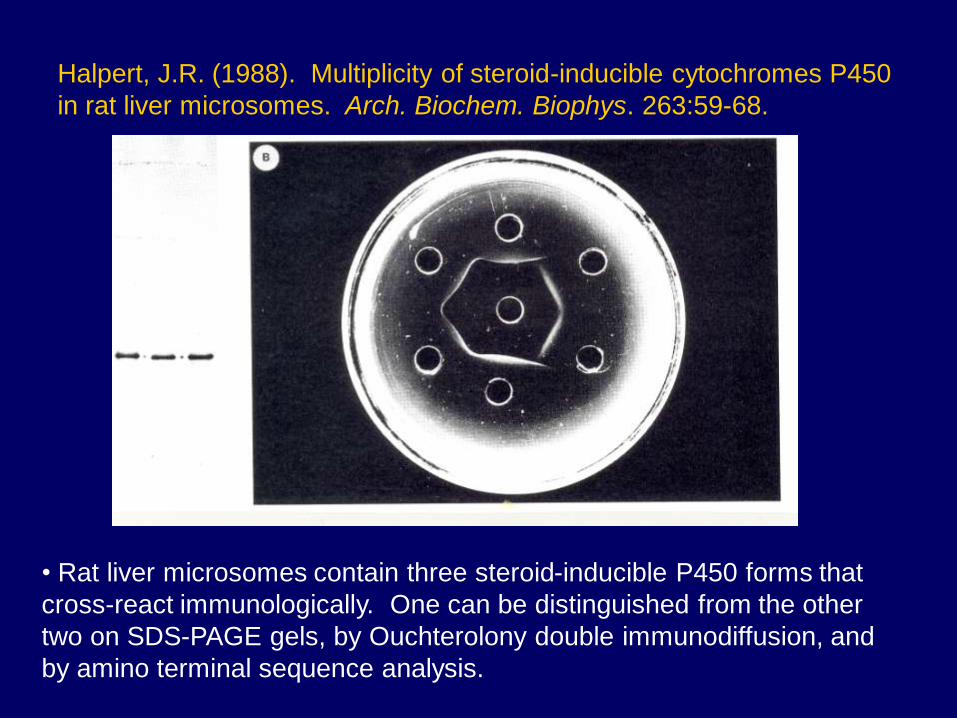
Halpert, J.R. (1988). Multiplicity of steroid-inducible cytochromes P450
in rat liver microsomes. Arch. Biochem. Biophys. 263:59-68.
• Rat liver microsomes contain three steroid-inducible P450 forms that
cross-react immunologically. One can be distinguished from the other
two on SDS-PAGE gels, by Ouchterolony double immunodiffusion, and
by amino terminal sequence analysis.

Fraser, D.R., He, Y.Q., Harlow, G.R., and Halpert, J.R. (1999). Use of
chimeric enzymes and site-directed mutagenesis for identification of
three key residues responsible for differences in steroid hydroxylation
between canine cytochromes P-450 3A12 and 3A26. Mol. Pharmacol.
55:241-247.
• Heterologously expressed P450 3A12 and 3A26 can account for
the two proteins in dog liver microsomes recognized by antibodies
to a purifed PB-inducible canine liver P450 (PBD-1).

Complications Presented by CYP3A4• Sigmoidal steady-state kinetics.
aflatoxin B1, amitriptyline,
carbamazepine, diazepam,
17 -estradiol, progesterone,
testosterone.
• Stimulation by some substrates of the oxidation of other substrates.
-naphthoflavone, diazepam, progesterone, quinidine, testosterone.
• Partial or no inhibition in the presence of two substrates.
erythromycin and testosterone, midazolam and terfenadine, aflatoxin B1 and -naphthoflavone, progesterone and 7-benzyloxy-4-trifluoromethyl-coumarin.

Domanski, T.L,, He, Y.-A., Khan, K.K., Roussel, F., Wang, Q., and
Halpert, J.R. (2001). Phenylalanine and tryptophan scanning
mutagenesis of CYP3A4 substrate recognition site residues and effect
on substrate oxidation and cooperativity. Biochemistry 40:10150-
10106.
•The differential effects of site-specific substitutions on
multiple P450 3A4 activities and heterotropic activation by
ANF suggest the presences of three subpockets within a
single large active site.

Multiple Conformers as an Alternative toMultiple Binding Sites within One P450Molecule
Initial concept: P450 is
represented by two
conformers with different
substrate specificity and
different response upon
substrate binding (Koley
et al., 1995, 1997).
Challenge: Static distribution of the enzyme
between two populations suggested by the model
has no obvious basis.

Our Understanding of the Mechanismsof P450 3A4 Cooperativity
P450 3A4 has two binding sites for substrates revealinghomotropic cooperativity (e.g. 1-PB).
Interaction with the high affinity binding site triggers aconformational transition, which promotes binding atthe second, low affinity site and subsequent spinshift.
This second binding event is likely also accompanied by aconformational change.
P450 3A4 in solution and in the membrane is representedby a mixture of (at least) two persistent conformerswith different affinities for substrates and differentposition of spin equilibrium.
This conformational heterogeneity is stabilized by theoligomerization of the enzyme.
ANF can modulate the partitioning of these conformers.

Protein-Protein Interactions and Modulation of Drug
Metabolism, Wednesday 9:00 AM,
Anaheim Convention Center, Room 210AB
• Introduction
T. Tracy, University of Minnesota
• Effect of P450-P450 Complex Formation on Monooxygenase Function
W. L. Backes, Louisiana State University School of Medicine
• Microsomal Monooxygenase as a Multienzyme System: Exploring Protein-Protein Interactions of Cytrochromes P450
D. Davydov, University of California, San Diego
• UGT-CYP Protein Interactions: Role of Conjugating Enzymes in Modulating Oxidative Enzyme Activity
Y. Ishii, Kyushu University
• Lack of Substrate Inhibition in a Monomeric Form of Human Cytosolic Sult2A1 (Abstract 967.5)
I.T. Cook, University of Alabama at Birmingham

Acknowledgments
Dr. Dmitri Davydov
Nadia Davydova
Dr. Harshica Fernando
Dr. Jessica Rumfeldt
Dr. Elena Sineva
Dr. Tamara Tsalkova
Dr. Steve Sligar
GM054995 (JRH)
GM 31756, GM33775 (SGS)

Lessons Learned from Colleagues
• Do what you should do not just what you
can do now (EFJ, RMP).
• It’s better to be scared than bored (DCL).
• Always look out for the next generation
(MJC, RWE, BSM, PFH, AYHL, MRW,
POM)








