
Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 955
_____________________________ _____________________________
The Expression and Regulation of Hyaluronan Synthases and Their Role
in Glycosaminoglycan Synthesis
BY
JONAS BRINCK
ACTA UNIVERSITATIS UPSALIENSISUPPSALA 2000

2
Dissertation for the Degree of Doctor of Philosophy (Faculty of Medicine) in MedicalBiochemistry presented at Uppsala University in 2000
ABSTRACT
Brinck, J. 2000. The Expression and Regulation of Hyaluronan Synthases and Their Role inGlycosaminoglycan Synthesis. Acta Universitatis Upsaliensis. Comprehensive Summaries ofUppsala Dissertations from the Faculty of Medicine 955. 63 pp. Uppsala. ISBN 91-554-4806-2.
The glycosaminoglycan hyaluronan is an essential component of the extracellular matrix in allhigher organisms, affecting cellular processes such as migration, proliferation and differentiation.Hyaluronan is synthesized by a plasma membrane bound hyaluronan synthase (HAS) which existsin three genetic isoforms. This thesis focuses on the understanding of the hyaluronan biosynthesisby studies on the expression and regulation of the HAS proteins.
In order to characterize the structural and functional properties of the HAS isoforms wedeveloped a method to solubilize HAS protein(s) while retaining enzymatic activity. The partiallypurified HAS protein is, most likely, not asscociated covalently with other components. Cellstransfected with cDNAs for HAS1, HAS2 and HAS3 were studied and all three HAS isozymeswere able to synthesize high molecular weight hyaluronan chains in intact cells. The regulation ofthe hyaluronan chain length involves cell specific elements as well as external stimulatory factors.HAS3 transfected cells with high hyaluronan production exhibit reduced migration capacity andreduced amounts of a cell surface hyaluronan receptor molecule (CD44) compared to wild-typecells.
The three HAS isoforms were studied and shown to be differentially expressed and regulated inresponse to external stimuli. Platelet derived growth factor (PDGF-BB) and transforming growthfactor (TGF-ß1) are important regulators of HAS at both the transcriptional and translational level.The HAS2 isoform is the isoform most susceptible to external regulation.
The role of the UDP-glucose dehydrogenase in mammalian glycosaminoglycan biosynthesiswas assessed. The enzyme is essential for hyaluronan, heparan sulfate and chondroitin sulfatebiosynthesis, but does not exert a rate-limiting effect.
Key words: hyaluronan, glycosaminoglycans, CD44, growth factors, UDP-glucose dehydrogenase.
Jonas Brinck, Department of Medical Biochemistry and Microbiology, Biomedical Centre, Box582, SE-751 23 Uppsala, Sweden
© Jonas Brinck 2000
ISSN 0282-7476ISBN 91-554-4806-2
Printed in Sweden by Lindbergs Grafiska, Uppsala 2000

3
non scholae sed vitae discimusSeneca († 65)

4
This thesis is based on the following papers, which are referred to in the text by their Roman
numerals:
I Asplund, T., Brinck, J*., Suzuki, M., Briskin, M. and Heldin, P. (1998) Characterization
of hyaluronan synthase from a human glioma cell line. Biochim Biophys Acta. 1380:377-
388* The two first authors contributed equally to the work.
II Brinck, J. and Heldin, P. (1999) Expression of recombinant hyaluronan synthase (HAS)
isoforms in CHO cells reduces cell migration and cell surface CD44. Exp Cell Res.
252:342-351
III Jacobson, A., Brinck, J., Briskin, M., Spicer, A. and Heldin, P. (2000) Expression of
human hyaluronan synthases in response to external stimuli. Biochem J. 348:29-35
IV Brinck, J., Roman, E*., Heldin, P. and Kusche-Gullberg, M. (2000) The role of UDP-
glucose dehydrogenase in mammalian glycosaminoglycan biosynthesis. Manuscript.* The two first authors contributed equally to the work
Published manuscripts have been reprinted with permission from the respective journals.

5
TABLE OF CONTENTS
BACKGROUND………………………………………………………………… 9
1 The extracellular matrix – the social context of the cells…………………………….…. 9
2 Glycosaminoglycans……………………………………………………………….…… 9
2.1 Hyaluronan (HA)………………………………………………………….…… 10
2.1.1 Structure and properties…………………………………………………10
2.1.2 Tissue distribution……………………………………………….….… 10
2.1.3 Functions…………………………………………………………….… 11
2.2 Sulfated glycosaminoglycans……………………………………………………11
2.2.1 Structures and properties…………………………………………….… 11
2.2.2 Functions……………………………………………………………… 12
3 HA binding proteins…………………………………………………………………… 13
3.1 Link module containing proteins………………………………………………. 13
3.1.1 Hyalectans………………………………………………………………14
3.1.2 CD44……………………………………………………………………14
3.1.2.1 Structure and ligand binding…………………………………… 14
3.1.2.2 CD44 and HA in cell migration…………………………………16
3.1.3 LYVE-1………………………………………………………………… 17
3.2 BX7B containing proteins…………………………………………………….… 17
3.3 Novel HA binding proteins…………………………………………………..… 18
4 Biosynthesis of glycosaminoglycans………………………………………………….…18
4.1 UDP-sugars…………………………………………………………………… 19
4.1.1 Synthesis of UDP-sugar pools……………………………………….…19
4.1.2 Regulatory role………………………………………………………… 21
4.2 Biosynthesis of HA………………………………………………………………22
4.2.1 Hyaluronan synthase (HAS) gene family……………………………… 22
4.2.2 HAS protein………………………………………………………….… 23
4.2.2.1 Enzyme properties…………………………………………..… 23
4.2.2.2 Kinetics…………………………………………………..…… 25
4.2.3 Regulation of HA biosynthesis………………………………………… 26
4.2.3.1 Turnover of HAS protein……………………………………… 26
4.2.3.2 Effects of growth factors……………………………………… 27
4.2.3.3 Involvement of protein kinases…………………………..…… 28
4.2.4 Biosynthetic directionality……………………………………………… 28

6
4.3 Biosynthesis of sulfated glycosaminoglycans…………………………….…… 29
4.3.1 Linkage region………………………………………………………… 30
4.3.2 Polymerization and modification……………………………….……… 30
5 Degradation of HA………………………………………………………………………31
5.1 Tissue turnover………………………………………………………….……… 31
5.2 Hyaluronidases………………………………………………………….……… 31
PRESENT INVESTIGATION…………………………………………..……… 33
6 Aims of the study…………………………………………………………….………… 33
7 HAS enzyme can be solubilized in an active form (Paper I)………………….………… 34
7.1 Solubilization and purification………………………………………..………… 34
7.2 Antibodies towards HAS protein……………………………………….……… 35
8 Is HA synthesized from the reducing end? (Paper I)…………………………….………35
9 Expression of HAS affects cell behavior (Paper II)……………………………..……… 37
9.1 Synthesis and accumulation of HA……………………………………..……… 37
9.2 HA chain length………………………………………………………………… 38
9.3 Cell migration…………………………………………………………..……… 39
10 HAS isoforms show specificity in gene regulation (Paper III)………………………… 39
10.1 Effects of culture conditions…………………………………………..……… 39
10.2 Effects of external stimuli………………………………………………………40
11 The role of UDP-glucose dehydrogenase in mammalian glycosaminoglycan
biosynthesis (Paper IV) ……………………………………………………….……… 40
12 Conclusions…………………………………………………………………………… 42
13 Future perspectives……………………………………………………………….…… 43
ACKNOWLEDGEMENTS…………………………………………………….… 45
REFERENCES………………………………………………………….…….… 46

7
Abbreviations
aa amino acids
CHO Chinese hamster ovary (cells)
CS chondroitin sulfate
DS dermatan sulfate
EGF epidermal growth factor
GAG glycosaminoglycan
Gal D-galactose
GalNAc N-acetyl D-galactosamine
Glc D-glucose
GlcA D-glucuronic acid
GlcNAc N-acetyl D-glucosamine
GPI glycosylphospatidylinositol
HA hyaluronan
HAS hyaluronan synthase
HS heparan sulfate
IdoA L-iduronic acid
KS keratan sulfate
PDGF platelet derived growth factor
PMA phorbol 12-myristate 13-acetate
TGF transforming growth factor
TSG-6 protein product of tumor necrosis factor-stimulated gene-6
UDP uridine 5'-diphosphate
xlHAS-rs Xenapus laevis hyaluronan synthase related sequence
Xyl D-xylose

8

9
BACKGROUND
1 The extracellular matrix – the social context of the cellsIt is a well-known fact that the human body is composed of different kinds of cells carrying out the
specialized functions for which they have been designed. Cells with similiar or complementary
functions assemble into tissues and organs that constitute functional entities and make the biological
organism Homo sapiens “work”. One difference between the human body and single cell
organisms such as bacteria is the fact that the individual cells in the body exist in a self-created
social environment, in contrast to the bacteria where the cell wall constitutes the barrier towards the
exterior world. The biological name for this social habitat is the extracellular matrix.
The cells synthesize the components of the extracellular matrix to fit their needs, which
means that structure and function of the matrix is almost as specialized and unique as the cells
themselves. It is the extracellular matrix that gives the tissues their tensile strength or elasticity, and
serves as a scaffold for cell adhesion and cell movement. The components of the matrix also help to
transmit signals that regulate cellular functions and are able to enhance or diminish these signals.
The extracellular matrix consists of a wide variety of protein and sugar molecules entangled
in a complex network that is constantly turned over. Among these are the linear carbohydrate
polymers, glycosaminoglycans, with essential functions both in the matrix and attached to the
surfaces of cells. It is of great importance to understand their metabolism since imbalances or
disorders in this system eventually will lead to disease. The main topic of this thesis is to give new
insights to the synthesis and regulation of the glycosaminoglycan hyaluronan.
2 GlycosaminoglycansThe members of the glycosaminoglycan (GAG) family are essential components in all higher
organisms and are found both on the surfaces of cells and in the extracellular matrix. GAGs are
unbranched carbohydrate polymers consisting of repeating disaccharide units that can be subject to
extensive modifications, thereby rendering highly diverse polysaccharide chains. The individiual
members of the GAG family are defined according to their disaccharide structure and modifications.

10
2.1 Hyaluronan (HA)2.1.1 Structure and properties
The GAG hyaluronan (HA)(reviewed in (106)) was first isolated from the vitreous body of the eye
by Meyer and Palmer in the 1930’s (123). HA consists of repeating disaccharide units of [GlcAß1-
3GlcNAcß1-4]n, where n can be up to twentyfive-thousand. The contour length of an HA chain of
Mr 4 x 106 is 10 µm (44). The HA chain is polymerized by a plasma membrane bound enzyme and
is not subjected to any type of covalent modification during its synthesis.
Nuclear magnetic resonance studies of the shape of the HA chain by Scott (165) have
shown the existence of internal hydrogen bounds that stabilize the chain in a stiffened helical
conformation. HA chains have the capacity to self-aggregate in aqueous solutions which can be
visualized in electron microscopy of rotary-shadowed preparations of HA (166). The aggregation of
two anti-parallel HA molecules is promoted by hydrogen bonds between the acetoamido group on
one chain and the carboxylate on the other (167). Preparations of high molecular weight HA in
concentrations of 5-10 mg/ml are therefore viscous.
Figure 1. An HA pericellular matrix excluding fixed red blood cells from the plasma membrane of a
Chinese hamster ovary cell.
2.1.2 Tissue distribution
HA is synthesized by almost all members of the animal kingdom as well as by certain bacteria and
algae viruses (106)(26). HA is found mainly in the extracellular space where it accumulates, but the
polymer can also be bound to the cell surface or be located intracellularly around in the nucleus and
in the lysosomes (153)(36)(108). The largest storage of HA in the human body is in the skin,
constituting about 50 % of the total HA supply (152).

11
Table 1 HA concentrations in various human organs and fluids
Organ or fluid Concentration (µg/g)
aqueous humour 0.3-2.2
brain 35-115
dermis 200
plasma (serum) 0.01-0.1
synovial fluid 1400-3600
thoracic lymph 8.5-18
umbilical cord 4100
urine 0.1-0.3
vitreous body 140-340
Data taken from (48).
2.1.3 Function
High molecular weight HA have been ascribed an essential structural role in the extracellular matrix
of the connective tissue where the polymers create a mesh-like structure (106)(166). The viscous
hydrated HA gel provides resistance to compressive forces and acts as a biological lubricant. During
embryonic development the deposite of HA creates a cell free area by expansion of the extracellular
space which facilitates cell migration (202). Besides being a structural element, HA has been
reported to be involved in several specific processes such as lympocyte extravasation (32), cancer
metastasis (171) and angiogenesis (158) probably through interactions with HA binding proteins.
Since the HA polymer itself does not exhibit any structural diversity, its function is in part
due to the chain length. The inductive role of HA in angiogenesis could be ascribed to HA
oligosaccharides (4-25 disaccharides)(221) whereas high molecular weight HA exerted an
inhibitory effect (43). HA fragments have also been reported to evoke an inflammatory response in
macrophages (121)(80).
2.2 Sulfated glycosaminoglycans2.2.1 Structure and properties
The sulfated GAGs can be subdivided into three groups based on the disaccharide composition:
heparan sulfate (HS)/heparin, chondroitin sulfate (CS)/dermatan sulfate (DS) and keratan sulfate
(KS) (Table 2)(96). During their synthesis in the Golgi, the chains are heavily modified by
12
deacetylation, C5-epimerization, N- and O-sulfations generating a high degree of diversity (see
section 4.3). The chain lengths vary between 5-70 kDa.
Table 2 The structure of the sulfated GAGs
GAG disaccharide unit modifications
heparan sulfate (HS)/
heparin
[GlcA/IdoAß/α1-4GlcNAcα1-4] N-deacetylation, N-sulfation
C5-epimerization of GlcA
C2-sulfation on GlcA/IdoA
C3,C6-sulfation on GlcNAc
chondroitin sulfate (CS)/
dermatan sulfate (DS)
[GlcA/IdoAß/α1-3GalNAcß1-4] C5-epimerization of GlcA
C2-sulfation on GlcA/IdoA
C4,C6-sulfation on GlcNAc
keratan sulfate (KS) [Galß1-4GlcNAcß1-3] C6-sulfation on Gal/GlcNAc
The sulfated GAG chains appear as proteolglycans, i.e. covalently linked to proteins, where the
protein core can be of a different nature: transmembrane, glycosylphosphatidylinositol (GPI)-
anchored or soluble. A proteoglycan can have several GAG chains of different types attached to it as
exemplified by aggrecan, which consists of a 200 kDa protein core and about 130 CS and KS
chains, with a total Mr of about 3 x 106 (66).
The expression pattern of proteoglycans is specific; heparin is synthesized only by
connective tissue mast cells in the form of the proteoglycan serglycin whereas HS is found on
several types of proteoglycans on virtually all cells in the body (96). The structural diversity of the
GAG chains is also precisely regulated as shown by recent studies with monoclonal antibodies
directed against structurally different epitopes of HS (212)(90). Different structures of the kidney
and muscle basal lamina were investigated and shown to contain a subset of different HS species.
2.2.2 Functions
The proteoglycan superfamily consists of over 30 members with a wide variety of biological
functions (82). They play an important role in the extracellular matrix organization, influence cell
growth and tissue maturation, and participate in the regulation of matrix turn-over by the binding
and inactivation of protease inhibitors. Proteoglycans act also as biological sieves by being
structural components in the glomerular basal laminae.
Many of the specific functions of the proteoglycans are due to the highly modified diverse
saccharide sequences of the GAG chains. For example, the anticoagulant activity of heparin has

13
been known for a long time (77). A unique pentameric structure present only in a fraction of the
total heparin has the capacity to interact with antithrombin-III, thus forming a potent inhibitor of
thrombin and thereby blood coagulation (12). Furthermore, proteoglycans are important modulators
of growth factor activities (159). The binding of growth factors to the GAGs protect ligands from
proteolytic degradation and GAGs have therefore been suggested to act as a growth factor storage
facility. HS has been shown to be required for binding of basic fibroblast growth factor to its
receptor and the growth factor mediated effects (228)(149).
3 HA binding proteinsHA is unique in the GAG family by lacking covalent linkages to proteins thus not being able to
form proteoglycans. The only exception to this rule is the ester linkage between the C-terminal
aspartic acid of the heavy chains of pre-α-trypsin inhibitor/inter-α-trypsin inhibitor and the C6-
hydroxyl group of an internal GlcNAc residue in the HA chain (234), although the functional role
of this linkage is not clear.
The fact that HA can form several specific interactions with proteins, serves as indirect evidence that
HA is more than a structural component of the extracellular matrix. These HA binding proteins are
mainly found extracellularly, but can also be bound to the cell surface or be located intracellularly.
New HA binding proteins are currently under characterization and the list is likely to get longer.
Below is some of the best characterized HA binding proteins presented, subdivided according to
their HA binding motif.
3.1 Link module containing proteinsThe Link module, about 100 amino acids (aa), is the structural HA binding motif shared by the Link
module superfamily, which comprise extracellular matrix molecules (link protein, hyaluronectin,
versican, aggrecan, neurocan, brevican, TSG-6) and the cell surface receptors (CD44 and LYVE-1)
(25).The three-dimensional structure of the human TSG-6 Link module, expressed in Escherichia
coli, has been resolved by nuclear magnetic resonance spectroscopy (100). It comprises two α-
helices and two triple-stranded anti-parallel ß-sheets arranged around a large hydrophobic core,
identical to the fold of the C-type lectin domain. The recombinant expressed Link module of TSG-6
has the capacity to bind HA (100), compared to CD44 which requires additional 60 aa of the
receptor in order to acquire the correct fold and HA binding capacity (6). The Link module of TSG-
6 can also bind CS at a position which is overlapping with the HA binding surface (136).

14
3.1.1 Hyalectans
The hyalectans are a family of proteoglycans interacting with HA and lectins and have currently four
members: versican, aggrecan, neurocan and brevican (83). They share a common tridomain structure
consisting of an N-terminus that binds HA, a central domain that carries GAG chains and a C-
terminal that binds to lectins (82).
Versican is the largest member of the hyalectan family with the core protein of 265-370 kDa
which is modified by 10-30 CS/DS chains. Versican binds HA with a Kd of about 4 nM (109) but
can also bind simple monosaccharide sugars as well as HS and heparin (209). The versican gene
can be upregulated by a variety of growth factors such as platelet derived growth factor (PDGF),
epidermal growth factor (EGF) and tumor necrosis factor (TNF) (195). Versican has been
implicated in regulation of neural crest cell migration by exerting a barrier role for the axonal
outgrowth (103).
Aggrecan consists of a core protein of about 220 kDa modified by CS and KS chains (up
to 130). Aggrecan secreted by chondrocytes aggregates extracellularly but can also bind to the cell
surface (94)(176). Aggrecan, link protein and HA forms a ternary complex (66) which, inside
cartilage, occupies a large hydrodynamic volume making the tissue elastic to compressive forces.
The affinity of the aggrecan link complex for HA is quite high (Kd≤1 nM).
Neurocan (protein core 140 kDa) was first cloned from rat brain (150) and is synthesized
by neurons, and probably astrocytes, in the brain (218). Neurocan binds neural adhesion molecules
Ng-CAM and N-CAM with high affinity and can thereby inhibit their homophilic interactions and
block neurite outgrowth (58)(51). Neurocan is developmentally regulated, and its interactions with
its ligands may be confined to restricted areas and a relatively brief stage in development (126).
Brevican is the smallest member of the hyalectans (protein core 100 kDa). The molecule can
be synthesized in a soluble extracellular form or as a GPI-anchored protein (168), which localizes to
the plasma membrane. This latter splice variant is deprived of its lectin binding domain, but still has
the HA binding domain and sites for glycosylation. Brevican is one of the most abundant CS
containing proteoglycans in the adult brain and its expression is highly specific (224). Brevican has
been reported to inhibit neurite outgrowth in vitro thus maybe controlling infiltration of axons and
dendrites in vivo (222).
3.1.2 CD44
3.1.2.1 Structure and ligand binding
CD44 is the best characterized HA receptor to date and its structure and function have been covered
in several excellent reviews (113)(95)(112). Besides HA, the CD44 receptor can bind a number of
other extracellular matrix components such as collagen I (42), fibronectin (89), HS, CS and
heparin (174). CD44 is widely expressed, and can be found on most hematopoetic cells,

15
keratinocytes, chondrocytes, many epithelial cell types and some endothelial and neural cells (112).
The many functions attributed to the receptor include T-cell extravasation (32), myelopoeisis (129)
and tumor migration (59)(237), although the mechanisms behind this are not always clear.
CD44 is a type-1 transmembrane glycoprotein and the standard isoform (CD44H; originally
found on hematopoetic cells (23)) consists of ~270 aa. The amino terminal part (~180 aa) of the
extracellular domain is characterized as the conserved region and exhibits 85 % homology between
mammalian species. This region harbors the Link module, five N-glycosylation sites and a BX7B
motif (see section 3.2). The proximal part of the extracellular domain (non-conserved region) is
characterized by alternative splicing of at least ten variant exons of the CD44 gene which allows
insertion of extra aa (87)(200)(237). The alternative splicing occurs only in particular cell types and
under certain conditions and is therefore considered to be precisely regulated. This proximal region
also contains O-glycosylation sites and sites for attachment of GAG chains. The CD44 protein,
including the splice variants, have been shown to be decorated with KS, HS and CS chains
(14)(72)(188). The transmembrane domain (21 aa) is 100 % conserved between mammalian
species. The cytoplasmic domain (~72 aa) seems to be able to interact with several intracellular
proteins such as ankyrin (91) and the ERM-protein family (ezrin, radixin and moesin) (206)(160)
thus potentially creating a linkage between the receptor and the actin cytoskeleton. The cytoplasmic
domain can also be phosphorylated at several tyrosine and serine residues but only two serine
residues are phosphorylated in vivo (130).
CD44 is the principle cell surface receptor for HA (1) by binding of HA to the Link module
(100)(4); the contribution of the BX7B motif (225) to HA binding is probably minor (112). The
smallest HA molecule that can bind to CD44 is a hexasaccharide to decasaccharide (210)(2)(189).
However, the existence of CD44 molecules on the cell surface does not automatically convey HA
binding but depends on the ”activation state” of the cell (113)(95). Three CD44 activation states
have been described: 1) CD44+ cells that bind HA constitutively, 2) CD44+ cells that can be induced
(by monoclonal antibodies or cytokines) to bind HA and 3) CD44+ cells that do not bind HA at all.
A lot of work has been put in to clarify what regulates the binding of HA to CD44. N-
glycosylation of CD44 seems to inhibit HA binding, since active HA binding cells have less N-
glycosylation than non-binding cells and treatment of these non-binding cells with N-glycosylation
inhibitors rendered them active (64)(92). O-glycosylation of CD44 has also been reported to have a
negative effect on HA binding in some cases (24) but not in all (235). Modification of the receptor
by sulfation leads to increased HA binding (118).
The transmembrane and intracellular domain of CD44 contribute to HA binding. Evidence
that the transmembrane domain is involved in dimerization of the receptor by disulfide bonding
(116) and palmitoylation (11) has been presented, and a correlation between ligand binding capacity
and dimerization has been shown (173). The cytoplasmic part of CD44 is important for ligand

16
binding, since cells expressing this truncated form failed to bind HA if they were not first exposed
to CD44-specific antibodies (111). The cytoplasmic part is also involved in heterogeneous
localization and polarization of receptors on the cell surface in pre-B cells and epithelial cells
(131)(216). Taken together, this strengthens the role of receptor-clustering as a positive factor for
HA binding. Phosphorylation of the intracellular domain of CD44 does however not seem to
promote or reglulate ligand binding (138)(208).
3.1.2.2 CD44 and HA in cell migration
Cell migration and cell locomotion involves a series of complex interactions between cytoskeleton,
cell surface receptors and matrix components (105)(170). Several examples from the field of
embryonic development implicate HA and CD44 as positive elements in cell migration. The
migration of cells into the developing chick embryo cornea and heart coincided temporally and
spatially with the synthesis of HA and the migration decreased as the hyaluronidase activity in the
tissues increased (204)(135). During the development of the soft palate, Spicer and co-workers have
shown with studies of HAS knock-out mice that the cells of the cranio-facial mesenchyme are
dependent on HA for migration and normal organization of the tissue (178). The embryonic
development of the cardiac septum and valves were also dependent on HA as shown in HAS2
knock-out mice (16); when the cardiac valve cells were cut out and grown on collagen gels they
could not migrate unless they were rescued with HAS2 expression or addition of exogenous HA. In
another study, Thomas et al. (194) showed that melanoma cells enhanced their migration rates on
HA coated dishes after transfection with CD44 compared to non-transfected cells, while antibodies
against the receptor inhibited migration. Phosphorylation of the intracellular serine residues of
CD44 is important for the CD44 mediated migratory effect, since melanoma cells expressing
phosphorylation mutants of CD44 were not able to migrate (138).
Attempts to explain the mechanism of the CD44 – HA interaction in cell migration have
been made. The deposit of HA in the extracellular matrix leads to an overall expansion of the tissue
(202)(122), thereby creating hydrated routes that facilitates cell migration according to Toole (202).
For example, HA indirectly promoted glioblastoma cell migration in 3-dimensional fibrin gels by
modulation of the fibrin fiber architecture thus increasing the pore sizes in the gels (65). In such a
model, CD44 could be the link between the cell’s actin cytoskeleton and the scaffold that the HA-
gel provide. The mechanism of migration might also comprise a turnover of the extracellular
components and cell surface receptors (105). Fibroblasts, macrophages and chondrocytes have the
capacity to internalize and degrade HA by a CD44 mediated endocytosis (22)(81), although the
amount of HA degraded by the cells of mesenchymal origin seems to be limited (71).
Knudson and Knudson (97) have presented an hypothesis where the assembly of HA
containing pericellular matrices around cells (first visualized by Claris and Fraser (18)) prevent

17
them from forming necessary interactions for cell migration with a decreased migration rate as an
effect. In vitro these matrices consist either of newly synthesized HA chains, still attached to the
HAS protein (69), or of exogeneously added HA which is bound to HA receptors (CD44)(99)(98).
The matrix formation can be promoted by HA binding proteoglycans, which also increase their
density and stability (70). Experimental evidence supporting this hypothesis has been presented
(140)(141) where neural crest cell migration was inhibited by HA matrix promoting chondroitin
sulfate proteoglycans (i.e. aggrecan).
3.1.3 LYVE-1
LYVE-1 is a type-1 transmembrane protein and a new member of the Link module superfamily
being a homologue of the CD44 receptor with an overall similarity to CD44 of 41 % (7). Of the
nine key amino acids that have been shown to be important in HA binding to CD44 (137)(4), only
three are conserved in LYVE-1 (7). The receptor is almost exclusively expressed in lymph vessels
where it co-localizes with HA on the luminal side. LYVE-1 is not, unlike CD44, expressed in blood
vessels. Since the major part of the HA turnover and clearance takes place via the lymphatic system
(see section 5.1) the receptor could be an important regulator of these events.
3.2 BX7B containing proteinsThe α-helical peptide sequence BX7B, where B is a basic aa and X is any non-negatively charged aa,
have been shown to bind HA (226)(225). At least three proteins (RHAMM, IHABP and CD34)
have such a functional HA binding motif, although it exists in several other proteins, including
members of the Link module superfamily. It is not known if the BX7B sequence can confer HA
binding capacity in these contexts.
RHAMM (acronym for Receptor for Hyaluronan-Mediated Motility) was first characterized
by Turley in 1992 from a 3T3 cDNA expression library (63) and contains two adjacent BX7B
motifs which contribute equally to HA binding. The 52 kDa RHAMM protein proved to be similar
to two of the proteins (52 kDa and 58 kDa) previously reported to be members of the Hyaluronan
Receptor Complex (HARC) involved in ras-transformed cell locomotion (207). However, the
cDNA clone isolated (63) was not of full length and additional exons, including splice variants, have
been described (14 exons in total)(40)(217) coding for a protein of 70-73 kDa.
The RHAMM receptor was first reported to be cell surface associated/extracellular (63) but
recent studies revealed that it also exists in the cytoplasm and in the nucleus (233)(39). RHAMM
has been implicated as a regulator of several important cell functions. Overexpression of the
RHAMMv4 (splice variant 4) in murine fibroblasts is transforming and causes spontaneous
metastases in the lung (60). RHAMM has been postulated to act down-stream of Ras on the MAP
kinase - ERK (extracellular-regulated kinase) signaling pathway (233). There is also a correlation

18
between RHAMM overexpression and overexpression of MAP kinase, which could serve as a
prognostic indicator of breast carcinoma progression (217).
In 1998 Hofmann et al. (76) identified IHABP (acronym for Intracellular Hyaluronic Acid
Binding Protein) which has over 90 % similarity to the RHAMM protein and which shares the HA
binding BX7B motifs found in RHAMM. Fieber et al. (45) could later show that RHAMM is a N-
truncated form of IHABP and that the full length RHAMM/IHABP gene encodes 18 exons in total.
IHABP is expressed as a 95 kDa intracellular protein in a wide variety of tissues. The function of
IHABP remains to be elucidated but the intracellular localization gives potentially new biological
roles for HA. The physiological role of RHAMM is under evaluation in the light of the new data
about IHABP (75).
3.3 Novel HA binding proteinsThe liver is important for HA turnover by the uptake and degradation of the HA in liver endothelial
cells (see section 5). The putative receptor(s) responsible for receptor-mediated endocytosis of HA
on these cells have been described but not cloned (107)(120)(227)(236). The receptor reported by
McCourt et al. (120) is possibly a part of a protein complex of three (related) proteins (two 170-
180 kDa and one 270 kDa proteins) and shows functional similarities to the scavenger receptor
family. Polyclonal antisera towards the 270 kDa protein blocked HA binding and degradation in
liver endothelial cells. The receptor reported by Zhou et al. (236) indicate a 300 kDa protein
complex as the functional unit for the receptor. Complete cloning and sequencing remains to be
done for both of these receptors to determine if they are related and to elucidate the molecular basis
of their HA binding.
Recently, three putative HA binding proteins have been identified on cDNA level by search
in an expressed sequence tag (EST) database (205). Based on the original tissues from which these
cDNAs were derived, namely white fat, bone marrow and osteoblast, they have received the names
WF-HABP, BM-HABP and OE-HABP, respectively. The functional roles of these molecules as
HA binding proteins remain to be assessed.
4 Biosynthesis of glycosaminoglycansThe biosynthesis of the members of the GAG family is done by partly unique pathways and involve
interactions between enzymes in different subcellular compartments. In a first common step, high
energy nucleotide sugar donors are synthesized in the cytoplasm. These UDP-sugars can be
directly utilized by the plasma membrane bound hyaluronan synthases (HAS) for HA
polymerization. The sulfated GAGs are synthesized on a protein core as proteoglycans in the Golgi
and the UDP-sugars must therefore first be transported into the Golgi compartments by an

19
antiporter system. The Golgi biosynthetic machinery include chain polymerization and various
modifications which, in concert, generate unique chain structures.
4.1 UDP-sugars4.1.1 Synthesis of UDP-sugar pools
The main part of the cell’s glycosylation reactions occur in the Golgi apparatus, which includes N-
and O-glycosylation of proteins and lipids, synthesis of ceramide linked oligosaccharides and
glycophospholipid anchors and the polymerization of GAG chains (214). A common feature for
these reactions is the transfer of monosaccharide units to proteins/lipids from high-energy
nucleotide sugar donors, UDP-sugars, by the action of specific glycosyltransferases.
The biosynthetic pathways for UDP-sugar have been studied and are now established (50).
Practically all of them occur in the cytoplasm of the cell and include unique pathways and
interconversion of UDP-sugars from other pathways (Figure 2). The starting material is often the
cytoplasmatic glucose pool, but monosaccharide units salvaged from lysosomal degradation of
glycoproteins and GAGs (117) can also act as precursors. Rome and Hill (157) have shown that
hexosamines in the lysosomes can be reutilized by over 50 % after being transported back into the
cytoplasm by specific transporters. Thus the transporters may give an important contribution to the
UDP-sugar synthesis in the cell.
UDP-GalNAcUDP-GalNAc-1-PGalNAc UDP-GlcNAc
Fru-6-P
GlcN-6-P
GlcNAc-6-P
GlcNAc-1-P
UDP-Glc
UDP-GlcA
UDP-Xylose
UDP-Gal
Gal
Gal-1-P
Glc
Glc-6-P
Glc-1-P GlcN
Figure 2. Schematic view of UDP-sugar biosynthesis.
Adapted from (50).
20
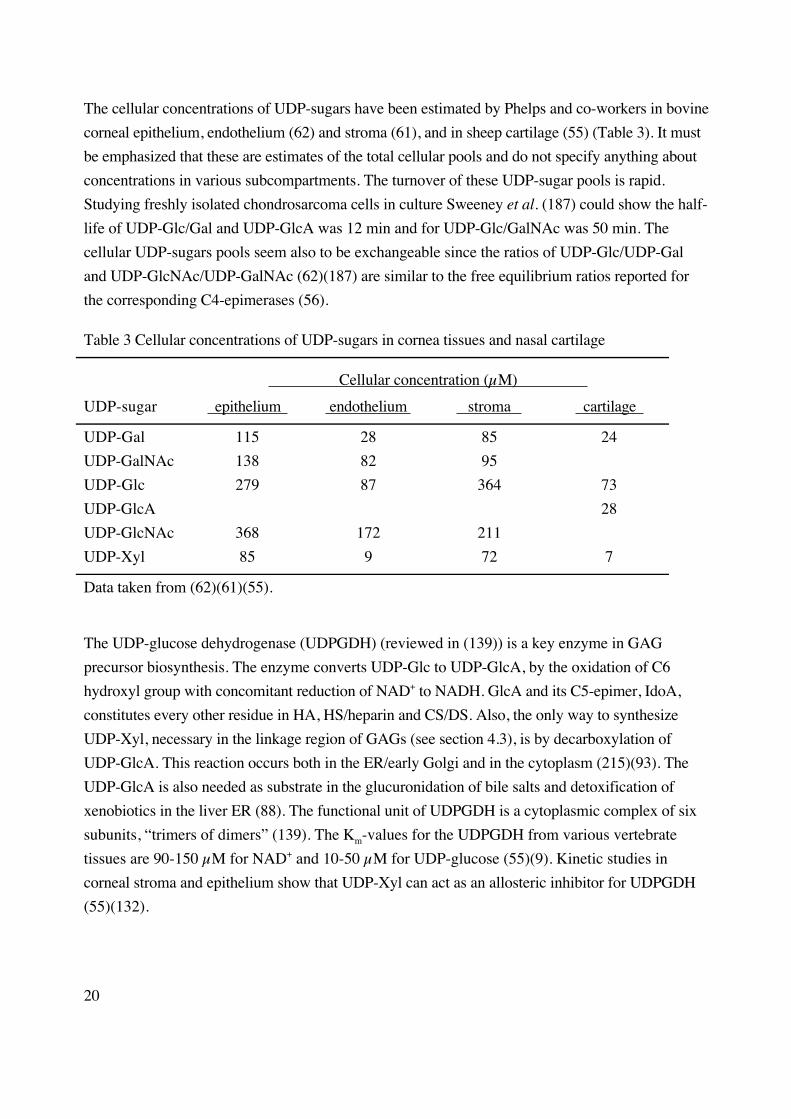
The cellular concentrations of UDP-sugars have been estimated by Phelps and co-workers in bovine
corneal epithelium, endothelium (62) and stroma (61), and in sheep cartilage (55) (Table 3). It must
be emphasized that these are estimates of the total cellular pools and do not specify anything about
concentrations in various subcompartments. The turnover of these UDP-sugar pools is rapid.
Studying freshly isolated chondrosarcoma cells in culture Sweeney et al. (187) could show the half-
life of UDP-Glc/Gal and UDP-GlcA was 12 min and for UDP-Glc/GalNAc was 50 min. The
cellular UDP-sugars pools seem also to be exchangeable since the ratios of UDP-Glc/UDP-Gal
and UDP-GlcNAc/UDP-GalNAc (62)(187) are similar to the free equilibrium ratios reported for
the corresponding C4-epimerases (56).
Table 3 Cellular concentrations of UDP-sugars in cornea tissues and nasal cartilage
Cellular concentration (µM)
UDP-sugar epithelium endothelium stroma cartilage
UDP-Gal 115 28 85 24
UDP-GalNAc 138 82 95
UDP-Glc 279 87 364 73
UDP-GlcA 28
UDP-GlcNAc 368 172 211
UDP-Xyl 85 9 72 7
Data taken from (62)(61)(55).
The UDP-glucose dehydrogenase (UDPGDH) (reviewed in (139)) is a key enzyme in GAG
precursor biosynthesis. The enzyme converts UDP-Glc to UDP-GlcA, by the oxidation of C6
hydroxyl group with concomitant reduction of NAD+ to NADH. GlcA and its C5-epimer, IdoA,
constitutes every other residue in HA, HS/heparin and CS/DS. Also, the only way to synthesize
UDP-Xyl, necessary in the linkage region of GAGs (see section 4.3), is by decarboxylation of
UDP-GlcA. This reaction occurs both in the ER/early Golgi and in the cytoplasm (215)(93). The
UDP-GlcA is also needed as substrate in the glucuronidation of bile salts and detoxification of
xenobiotics in the liver ER (88). The functional unit of UDPGDH is a cytoplasmic complex of six
subunits, “trimers of dimers” (139). The Km-values for the UDPGDH from various vertebrate
tissues are 90-150 µM for NAD+ and 10-50 µM for UDP-glucose (55)(9). Kinetic studies in
corneal stroma and epithelium show that UDP-Xyl can act as an allosteric inhibitor for UDPGDH
(55)(132).

21
To be utilized for the biosynthesis of the sulfated GAGs in the Golgi, UDP-sugars synthesized in
the cytoplasma have to overcome their subcellular mislocation. The transport into the Golgi is
mediated through a group of hydrophobic transmembrane proteins and has been extensively studied
by Hirschberg (reviewed in (74)(73)). The proteins are antiporters, meaning that for each nucleotide
sugar transported into the Golgi, the corresponding nucleoside monophosphate must be transported
back to the cytoplasm. Each nucleotide sugar has its specific transporter and there is little, if any,
non-specificity in the system. The transport does not require ATP but is thought to be driven by the
down-concentration for the nucleoside monophosphate being transported out from the lumen into
the cytosplasm.
Studies on reconstituted antiporters in proteoliposomes have revealed useful information
regarding transport kinetics (127)(128). The Km-values for the UDP-Gal, UDP-Xyl and UDP-GlcA
transporters were determined to be between 2-5 µM which was very close to the Km-values for intact
Golgi vesicles. Preloading the proteoliposomes with UMP, a putative antiporter for the nucleotide
sugars, increased the nucleotide sugar transport by 2-3-fold. Thus the antiporters have the potential
to concentrate the nucleotide sugars between 50- to 100-fold relative to the incubation medium (73).
4.1.2 Regulatory role
The question of whether UDP-sugars exert any regulatory effect on mammalian GAG biosynthesis
is important, but there are few definite answers up to date. Balduini et al. (5) could show a decrease
CS synthesis in cornea when UDP-Xyl was added in vitro. They concluded that the level of
regulation was at the UDPGDH enzyme, presumably due to the negative allosteric effect of UDP-
Xyl. The turn-over of the UDP-sugars is rapid as reported by Mason and co-workers (187). After
stimulation of chondrosarcoma cells with serum or insulin the [35S]sulfate and [3H]leucin
incorporation into macromolecules increased 2-3-fold although the UDP-sugar pool sizes remained
constant. In another study (230), articular explant cultures could increase their UDP-GlcA pool
upon stimulation with serum concomitantly with increased production of GAGs. Thus the increased
demand for UDP-sugars in cells can be met by an increased fluctuation of the pools or by an
expansion of the pool size. They concluded that the UDP-sugars pools are unlikely to become rate-
limiting in GAG biosynthesis (187).
A Madin-Darby canine kidney (MDCK) cell line, 98 % deficient in UDP-Gal transport into
the Golgi lumen, had marked reduced galactosylation of glycoproteins and glycosphingolipids (13)
and reduced amounts of KS (the only GAG containing Gal in the polymer) (201). However, the
biosynthesis of HS and CS in these cells was not affected although Gal is needed in the linkage
region between the core protein and the GAG polymer (201). The authors speculated that the Km-
values for the glycosyltransferases involved in the transfer of Gal in the linkage region are lower

22
than that of the transferases involved in polymerization of KS, thereby explaining the different
effects on KS and HS/CS, although they could not rule out the possibility of subcompartementation
of the two processes.
Several reports, including studies of Km-values of the nucleotide antiporters in
proteolysomes, have suggested that the rate-limiting step of the Golgi located posttranslational
modifications (e.g. glycosylation) is the transport of UDP-sugars into the Golgi by the antiporter
system and not the glycosyltransferase activities (73).
4.2 Biosynthesis of HA4.2.1 Hyaluronan synthase (HAS) gene family
In 1992 van de Rijn and co-workers identified a gene responsible for HA production in group A
Streptococcus (33)(211). It soon became clear that the synthase was part of a hyaluronan synthase
(HAS) operon consisting of three members and that all encapsulated Group A Streptococci
possessed this operon (20). The gene was characterized (31)(35) and received the acronym hasA.
The hasB gene was characterized as a UDPGDH (34), the enzyme that converts UDP-Glc to UDP-
GlcA. The hasC gene encodes a UDP-glucose pyrophosphorylase responsible for the production of
UDP-Glc from glucose 1-phosphate and UTP (19).
The vertebrate HAS proteins belong to a gene family with four members (180). The first
three isoforms termed HAS1, HAS2 and HAS3 were cloned from human and mouse origins in a
short period of time between 1996 and 1998 (85)(172)(219)(177)(182). The fourth member, termed
HAS-related sequence (xlHAS-rs), was identified in Xenopus leavis (African green frog) with no
mammalian orthologue found so far (180). The first HAS gene was actually submitted to the gene
bank already in 1983 (164) but under the name DG42 (Differentially expressed at Gastrulation), a
protein found to be predominantly expressed during X. laevis gastrulation. The function of the
protein was at that point in time unknown. Later, a controversy about its catalytic properties arose
(213) which now has been resolved (see section 4.2.2.1).
The four members of the vertebrate gene family have probably arisen from a common
ancestral gene (180) after three gene duplications with subsequent divergence. Data that support this
hypothesis are common intron-exon boundaries and high sequence similarities between different
orthologues (Figure 3). The HAS gene family seems to have developed early in evolution since the
three first HAS genes are located on three separate autosomes (183). The tight coupling between the
HAS protein and the precursor producing enzymes seen in bacteria is not seen in the vertebrates; the
UDPGDH gene is located on a separate chromosome from the HAS genes (179).
The identification of a HAS gene in 1997 in the Paramecium bursaria Chlorella virus
(PBCV-1) genome (30) was a big surprise. When infecting chlorella-like green algae, a HAS
enzyme is translated from the open reading frame A98R. The origin of the viral HAS gene is still an

23
enigma. The enzyme shows about 30 % similarity on aa level with both vertebrate and bacterial HAS
with a slight predominance for the vertebrate protein. However, the viral genome also carries two
genes for HA precursors synthesis; a UDPGDH and a glutamine:fructose-6-phosphate
amidotransferase for synthesizing UDP-GlcA and glucosamine, respectively (104), which show a
higher degree of homology with their prokaryotic counterparts than their vertebrate ones (26).
However, the three genes do not function as an operon in the algae as they do in bacteria (30).
HAS1
HAS2
HAS3
xl HAS-rs 47
41
53
44
52
58
44
53
67
46
46
44
xl HAS-rs HAS1 HAS2 HAS3
Amino acid sequence similarity (%)
DNA sequence similarity (%)
xl HAS-rs
HAS1
HAS2
HAS3
ancestralHAS gene
Figure 3. The evolution of the vertebrate HAS gene family and the sequence similarities
between its members.
Data taken from (180).
The Gram-negative bacteria Pasteurella multocida is able to synthesize an HA capsule. Utilizing
transposon insertional mutagenesis a new type of HAS enzyme was cloned in the bacterial strain by
DeAngelis (29). This enzyme does not show any major resemblance to the other HAS genes,
deviating by size, predicted membrane topology and polymerization mechanism. DeAngelis has
suggested a classification of the HAS enzymes based on these characteristics (26). As of now, all
the HAS proteins cloned including vertebrate, prokaryotic and viral HAS belong to class I, with the
P. multocida HAS as the only member of class II.
4.2.2 HAS protein
4.2.2.1 Enzyme properties
The structure and topology of the HAS protein is to a large extent unknown. Philipson and
Schwartz could, with rigid experimental data, show that the vertebrate enzyme co-purified with the
plasma membrane fraction of mouse oligodendroglioma cells when fractionated by centrifugation

24
(142). Previously, Dorfmann and co-workers had shown that the bacterial HAS resides in the
membrane in group A Streptococci (119).
The size of the vertebrate HAS proteins is about 65 kDa, deduced from their aa sequences.
In a review of the HAS enzymes (220) a seven transmembrane or membrane associated domain
topology of the enzyme was suggested based on hydrophobicity plots of the aa sequence
(Figure 4); only a very small portion of the polypeptide chain is exposed to the outer surface in this
model. The structure has partly been supported by experimental data putting both the N- and C-
terminus in the cytosol (178). A basic aa cluster in the C-terminal part of the enzyme has been
proposed to participate in the regulation of the shedding of the negatively charged HA chain from
the cell surface. The large intracellular loop has motifs that are highly homologous between different
HAS members (180)(26) indicating this as the catalytic domain. Recently, site-directed mutagenesis
of HAS1 revealed potential sites for ß1-3GlcA and ß1-4GlcNAc transferase activities in the central
loop (229). The suggested topology is shared by both the prokaryotic and eukaryotic members of
the class I HAS family (220) with the difference that the prokaryotic proteins are shorter by a lack
of two C-terminal membrane spanning domains. This leaves only the P. multocida HAS (class II)
with a different structure (30) which probably only has two transmembrane domains or is soluble
(26).
NH2
COOH
Cytoplasm
Extracellular matrix
Figure 4. Suggested topology of the HAS enzyme (class I).
Adapted from (220). Star and circles show positions of aa important in ß1-3GlcA and ß1-4GlcNAc
transferase activities, respectively. Box depicts basic aa cluster suggested to be involved in shedding
of the HA chain from cell surface.
The long lasting enigma about the enzymatic activity of the DG42 protein, now termed Xenopus
laevis HAS1, seems to be resolved. In 1996 Semino et al. (169) claimed that DG42 synthesizes

25
chitin oligomers [GlcNAcß1-4GlcNAc]n n=4-5, while Meyer and Kreil (124) showed HA
polymerization; the dispute was commented on by Ajit Varki (213)). Kimata and co-workers have
recently shown that both activities reside in the purified enzyme when assayed in vitro (229).
The structure and topology of the HAS enzyme has been studied in greater detail in the
bacterial system. Weigel and co-workers have suggested that the HAS protein forms a pore in the
plasma membrane through which the polymer can be translocated concomitantly with chain
elongation (199)(197). The association of about 16 cardiolipin molecules, which induced enzyme
activity in vitro, seems to assist in the formation of a pore like structure. Other lipids have been
shown to enhance enzymatic activity, while others completely abolish it. (197). Less is known about
the vertebrate HAS enzymes and their lipid dependence and association. Several laboratories,
including ours, have solubilized HAS protein from different cell lines with sustained activity using
(preferably) digitonin as detergent (Paper I)(133). Kimata and co-workers have recently shown that
the detergent CHAPS was able to restore activity and kinetics of solubilized HAS protein to that of
the membrane bound enzyme (229).
To synthesize HA the following enzymatic activities are needed: ß1-3GlcA transferase, ß1-
4GlcNAc transferase and HA translocation (through the membrane). Does the HAS protein display
all of these activities by itself or is it aided by an accessory component? Three reports give rigid
evidence supporting the one polypeptide hypothesis. Weigel and co-workers (199) could show that
the size of the functional Streptococcal HAS complex expressed in E. coli was the same as the HAS
protein itself in irradiation studies, thereby excluding additional proteins. DeAngelis (28)
transformed the yeast strain Saccharomyces cerevisiae with the cDNA for Xenopus laevis HAS1
which gave the yeast the capacity to synthesize HA polymers in vitro. Wild-type yeast lacks the
ability to synthesize any GAGs due to lack of UDP-GlcA in the cells. The yeast cell should
therefore completely lack the machinery for GAG synthesis but the HAS polypeptide gives it that
capacity. Kimata and co-workers (229) managed to purify mouse HAS1 to homogeniety and
showed that it could synthesize HA in vitro.
4.2.2.2 Kinetics
The enzyme kinetics of synthases of both prokaryotic (198) and eukaryotic (86) origin have been
studied with regard to their substrates UDP-GlcA and UDP-GlcNAc. The data are summarized in
Table 4. The UDP-GlcNAc utilization of recombinant Streptococcus pyogenes (spHAS) and
Streptococcus equisimilis (seHAS) expressed in E. coli were studied and shown to be sigmoidal
for spHAS in a Michaelis-Menten plot. The indicated cooperativity for UDP-GlcNAc was seen
both for membrane fractions of spHAS (198) and for the solubilized form in the presence of
cardiolipin (197). The utilization of the UDP-GlcA was non-cooperative for both the bacterial
strains. Stoolmiller and Dorfman have also reported a non-hyperbolic plot for UDP-GlcNAc

26
utilization by Streptococcus (185). The enzyme kinetics displayed by spHAS may reflect an
evolutionary adaptation to ensure that cell growth is not imparied by the production of the HA
capsule (198).
The three vertebrate HAS proteins were assessed in a similar manner to the bacterial forms
(86). The enzymes displayed similar Km- and Vmax-values and showed no sign of cooperativity for
either substrate.
Table 4 Enzyme kinetics of prokaryotic and eukaryotic HAS enzymes
Source
Km for UDP-GlcA †
(µM)
Km for UDP-GlcNAc ††
(µM)
HA polymerization rate
(monosaccharide/min)
spHAS 27 125
seHAS 51 30
HAS1 53 501 1256
HAS2 33 331 1014
HAS3 35 223 174
Data taken from (198)(86). Membrane fractions of recombinant Streptococcal and mouse HAS
proteins were used to determine Km-values.† Concentration of UDP-GlcNAc was 500 µM†† Concentration of UDP-GlcA was 100 µM for bacterial HAS and 200 µM for mouse HAS
4.2.3 Regulation of HA biosynthesis
The HA biosynthesis can be regulated on different levels. As described in section 4.1.2 the access to
UDP-sugars can potentially constitute a regulatory mechanism for GAG synthesis. Several studies
have also been conducted on the growth factor stimulation of HA synthesis and the involvement of
protein kinases.
4.2.3.1 Turnover of the HAS protein
The turnover of the HAS proteins have only been studied indirectly and the results are
contradictory. Calabro and Hascall (15) studied HA and CS synthesis in chondrosarcoma cells
during brefaldin A treatment. Brefaldin A is a fungal metabolite that interferes with vesicular
transport causing disassembly of the Golgi complex and could thus indirectly interfere with the
delivery of the newly synthesized HAS proteins to the cell membrane. However, the HA synthesis
remained remarkably constant for a period of 8 hours (15). Bansal and Mason (8) came to different
conclusions using the same cell line when they added cycloheximide, an eukaryotic protein

27
synthesis inhibitor, to the cultured cells. They could see a 50 % reduction in HA production after
less than 2 hours treatment. Other studies have also indicated a more rapid turnover of HAS protein
using cycloheximide (68).
An explanation to the contradictory results of HAS protein turnover could be that different
HAS isozymes are influenced by the cell type that they are expressed in, by the cell culture
conditions and/or that the HAS isoforms exhibit different turnover rates. Itano et al. (86) have
shown that the three enzymes deviated from each other in terms of their catalytical stability in vitro,
HAS1 having the narrowest window of activity (1 hour) while the HAS2 (4 hours) and HAS3
(8 hours) windows were broader.
4.2.3.2 Effects of growth factors
Platelet derived growth factor (PDGF) is a growth factor with broad specificity which stimulates
proliferation in connective-tissue cells (67). The transforming growth factor (TGF) is a family of
growth factors acting on a broad range of cells exhibiting stimulatory and inhibitory effects in a cell
specific manner (143). Heldin and co-workers (68) demonstrated that PDGF-BB stimulation of
human mesothelial cells led to increased production of HA and that antibodies towards PDGF-BB
partly inhibited this increase (3). The authors showed that mesothelioma cells, the transformed
counterpart of mesothelial cells, could generate stimulatory signals for HA production.
Mesothelioma cells do not produce HA by themselves; rather they secrete factors that stimulate HA
synthesis in fibroblasts and mesothelial cells (3). Furthermore, inhibition of the de novo protein
synthesis with cycloheximide diminished the effect of PDGF in mesothelial cells, suggesting that
PDGF controls the HA synthesis at the transcriptional level (68). In another study, PDGF-BB and
TGF-ß1 were shown to stimulate HA synthesis in human foreskin fibroblasts (186). These signals
were also dependent on protein synthesis since cycloheximide inhibited in part the stimulatory
effects.
Schor and collegues have been studying HA production in skin fibroblasts in relation to
growth substratum and growth factors. These studies revealed that fetal and adult fibroblasts
upregulate their HA synthesis when cultured on a collagen substratum compared to plastic dishes,
and that TGF-ß1 stimulated HA synthesis in confluent fetal cells growing on plastic substratum, but
inhibited HA synthesis when cells were growing on collagen (38). In 3-dimensional collagen gels
fetal fibroblast HA synthesis was unaffected by PDGF, epidermal growth factor (EGF) and
fibroblast growth factor (FGF), but was inhibited by TGF-ß1. Adult fibroblast HA synthesis was
stimulated by the same factors, but was unaffected by TGF-ß1 (37).
More recently, the promotor sequence of mouse HAS1 have been cloned and partly
analyzed (223). Using the Internet Transfac program several potential regulatory cis-elements have

28
been identified, such as AP-2, CREB, MyoD, SRY and Sox-5, which in future might help to predict
and determine the importance of various growth stimuli for HA production.
4.2.3.3 Involvement of protein kinases
Several reports have indicated that protein kinases are involved in the stimulatory signals leading to
increased HA production, including direct phosphorylation of the HAS protein (125)(146). After
the cloning and characterization of the HAS enzyme family (220) caution is advised about the
conclusions drawn in these former reports.
Studies by Heldin and co-workers revealed that the stimulatory effect of PDGF on HA
biosynthesis in mesothelial cells and fibroblasts was partly dependent on protein kinase C and
partly dependent on de novo protein synthesis. Interestingly, the phorbol ester PMA, an activator of
protein kinase C, stimulated HAS proteins through a pathway which was not dependent on de novo
synthesis (68)(186). Treatment of mesothelial cells with phosphotyrosine phosphatase inhibitors
have also been shown to stimulate HA production (68)(134).
Stimulation of mesothelial cells from rabbit pericardial cavity with EGF and insulin-like
growth factor (IGF) gave a cooperative enhancement of HA biosynthesis, which could be
suppressed by a tyrosine kinase inhibitor (78). This result suggested that the inhibitory effect was
on the tyrosine kinase activity of the cytoplasmic domains of the IGF-I and EGF receptors, rather
than a direct influence on the HAS protein. Similar results with IGF-I were also seen in peritubular
rat testis cells (193). The HA production in pericardial mesothelial cells could be stimulated by
prostaglandin E2 through a cAMP-mediated signal, which could also be seen when the intracellular
level of cAMP was raised by addition of synthetic analogues or stimulators of cAMP production to
the cell culture media (79). They could also show that the signal transduction pathway from cAMP
was to protein kinase A and not protein kinase C in these cells.
Salustri and co-workers have shown that the cumulus cells increase their HA synthesis
during the expansion of the cumulus oophorus in the mammalian preovulatory follicle (163) in
response to follicle stimulating hormone (FSH) and a soluble oocyte factor(s) (162). FSH has been
shown to increase the intracellular cAMP levels, which correlated with the net increase of HA
production in mouse cumulus cells, while tyrosine kinase inhibitors suppressed the effect of FSH
partially (196). The HA synthesis was abolished after treatment with RNA synthesis inhibitors,
indicating that the induction of HA production is primarily controlled at the transcriptional level.
4.2.4 Biosynthetic directionality
A question that still remains unclear is the biosynthetic directionality of the HAS enzyme, i.e.
whether monosaccharides are added to the reducing end or the non-reducing end of the growing
HA polymer. In nature the predominant biosynthetic directionality for carbohydrate polymers

29
seems to be at the non-reducing end, exemplified by the sulfated GAGs (96), but elongation in the
reducing end has also been reported (polymerization of dextran (155)). The existence of two classes
of HAS enzymes, probably with different evolutionary origin (see section 4.2.1), makes it possible
that the different classes exhibit different polymerization mechanisms. Several attempts using
different approaches have been made to clarify the polymerization directionality of class I Has but
conclusive results are still lacking. However, the polymerization directionality of class II Has
(Pasturella multocida) has been determined.
Stoolmiller and Dorfman in 1969 showed experimental data that HAS protein from
Streptococcus polymerized the HA chain at the non-reducing end (185). The study was based on an
in vitro pulse experiment with UDP-[14C]GlcA alone, with subsequent digestion of the labeled HA
chain with Streptococcal hyaluronidase. The hyaluronidase digests ß1-4 linkages in the HA chain
by an eliminase reaction rendering predominantly unsaturated disaccharides (∆di-HA), and a smaller
portion of saturated disaccharides (di-HA), which are formed from the non-reducing terminal
disaccharide of the HA chain. The majority (88 %) of the radioactivity was found in di-HA after the
pulse experiment, indicating chain growth in the non-reducing end.
Prehm obtained the opposite result in 1983 when assaying differentiated teratocarcinoma
cells by pulse-chase experiments (144). HA polymerization was done with a pulse of UDP-
[14C]GlcA and UDP-GlcNAc followed by a chase with unlabeled UDP-GlcA. The pulse-chase
labeled HA was then degraded from the non-reducing end with exo-enzymes (ß-glucuronidase and
ß-N-acetylglucosaminidase). Prehm concluded that HA synthesis occurs at the reducing end since
the [14C]GlcA residues were cleaved off the HA chains directly in the presence of the digestive
enzymes; if the HA chain had been polymerized from the non-reducing end there would have been a
time lag before a drop in radioactivity could have been seen.
DeAngelis has studied the unique Pasturella multocida HAS expressed in the yeast
S. cerevisiae in terms of its polymerization directionality (27). The system has the advantage of
being devoid of nascent HA chains, since yeast cells are not able to synthesize UDP-GlcA. This
means that all HA produced in the in vitro HA polymerization assay is synthesized de novo. Under
these experimental conditions, DeAngelis was able to show addition of a monosaccharide unit to a
HA tetrasaccharide which could be cleaved off with exoglucosidases from the non-reducing end.
This was also the first time that a HA primer, other than UDP-GlcA or UDP-GlcNAc, could be
used in HA biosynthesis. In summary, rigid evidence that the P. multocida HAS elongates HA from
the non-reducing end was provided.
4.3 Biosynthesis of sulfated glycosaminoglycansThe three groups of sulfated GAGs, HS/heparin, CS/DS and KS, are synthesized as proteoglycans
in the Golgi. A linkage region, connecting the protein core with the GAG chain, initializes the chain

30
polymerization before the chain is further polymerized and modified by an elaborate biosynthetic
machinery.
4.3.1 Linkage region
The linkage region, connecting HS/heparin and CS/DS chains to a serine residue on the protein
core, consists of the tetrasaccharide GlcAß1-3Galß1-3Galß1-4Xylß1-Serine (156) and is initiated
by a unique transferase adding Xyl from a UDP-Xyl donor. The addition of a α-GlcNAc and ß-
GalNAc residue after the terminal GlcA residue of the linkage region constitutes the bifurcation of
HS/heparin and CS/DS biosynthesis, respectively. The aa sequence in the vicinity of the serine
residue partly influences the nature of the GAG chain that is going to be synthesized; repetitive
serine-glycine residues together with an acidic aa cluster and a hydrophobic patch promotes HS
polymerization (232)(231).
The linkage region in KS is different from the tetrasaccharide described above. KS chains
can either be linked to an asparagine or a serine/threonine thus resembeling the linkage of N- and
O-linked glycans, respectively (57).
4.3.2 Polymerization and modification
The heparin synthesis is the best studied and understood and is referred to as the default
modification (161). It begins with alternating transfer of GlcA and GlcNAc from corresponding
UDP-sugars to the linkage region, followed by the action of the first modification enzyme N-
deacetylase/N-sulfotransferase (NDST) that replaces the N-bound acetyl group on the GlcNAc
residue with a sulfate group. The subsequent enzymatic modifications are partly coupled to the
action of the NDST enzyme and therefore cannot take place before N-sulfation has occured. The
C5-epimerase converts GlcA to IdoA which promotes the 2-O-sulfation of the IdoA residue,
although GlcA residues can be modified in the same way. The last step in the chain synthesis is the
6-O-sulfation and selective 3-O-sulfation on GlcNAc residues. The default modification results in a
highly sulfated polymer with a typical disaccharide structure -IdoA(2-OSO3)-GlcNSO3(6-OSO3)-;
this disaccharide constitutes about 80 % of the disaccharides in heparin.
HS is structurally more diverse than heparin because the HS chains escape the default
modification pathway for reasons that are not fully understood. Only about 10 % of the
disaccharides in HS are default modified (161). Four different NDST isozymes have now been
cloned and seem to contribute to the different N-sulfation patterns seen in HS and heparin.
Transfection of NDST-2, an isozyme predominantly expressed in heparin-producing mast cells, into
HS producing cells increased their N-sulfation (17), which was not the case when more widely
distributed NDST-1 was overexpressed (84). Several isoforms of the 2-O-, 3-O and 6-O-

31
sulfotransferases exhibiting different substrate specificities in vitro have been cloned, that could
contribute to generation of specific sulfation patterns in vivo (115).
The polymerization of CS/DS and KS is less well understood (96). The GalNAc residues
remain acetylated in CS/DS chains rendering the chains less structurally diverse than HS. KS can
undergo modifications through selective 6-O-sulfation on both Gal and GlcNAc residues which are
non-randomly distributed along the chain.
5 Degradation of HA5.1 Tissue turnoverHA is constantly being turned over in the body. In a series of experiments performed by Laurent
and Fraser (108)(49)(46) it has been estimated that about one third of the total HA content (11-17 g
for a 70 kg man) is degraded per day. The degradation takes place both locally in the tissue and in
central organs such as the liver, kidney, spleen and bone marrow. The local tissue turnover rate of
HA has been estimated to t1/2=12 hours in the skin studying injected [3H]HA (151). The HA that
escapes degradation locally is transported via the lymph to the lymph nodes (191) where 50-90 %
of the HA is degraded (47). The blood receives HA from lymph nodes (10-100 mg/day), where it
disappears within minutes (t1/2= 2-6 min). The liver takes up about 90 % of the HA that reaches the
blood by a receptor mediated process (41). The receptor(s) have been studied functionally and
biochemically by several groups and their cloning seems to be at hand (120)(236).
5.2 HyaluronidasesThe HA that is taken up by the cells is degraded inside the lysosomes by hyaluronidases and the
exoenzymes ß-glucuronidase and ß-N-acetylglucosaminidase (108). The hyaluronidases are a
diverse group of enzymes isolated from different origins such as vertebrates, leeches and bacteria,
and the enzymes exhibit different kinds of activities (102). The vertebrate enzymes are endo-ß-N-
acetyl-D-hexosaminidases that degrade HA (and CS to a lesser extent) yielding tetrasaccharides and
hexasaccharides as final products.
Up to now six paralogue human hyaluronidase genes have been identified (21)(184) which
are arranged in two tightly linked triplets on chromosomes 3p21.3 (genes HYAL1, HYAL2,
HYAL3) and 7q31.3 (genes HYAL4, SPAM1, HYALP1) and codes for the proteins Hyal-1, Hyal-2,
Hyal-3 and Hyal-4, PH-20 respectively; SPAM1 is a pseudogene with no protein product. HYAL1,
HYAL2 and HYALP1 are widely expressed, whereas HYAL3 and HYAL4 are differentially
expressed in bone marrow and testis, and placenta and skeletal muscle, respectively. The genes for
human hyaluronidases are mapped to loci which have been described as candidate tumor suppressor
loci (101).

32
It is not known if the functional implications of the hyaluronidases are restricted to HA degradation
in the lysosomes due to the fact that the enzymes are only active in acidic pH (52); the role of
hyaluronidases remains largely to be elucidated. An exception is PH-20, a 64 kDa GPI-anchored
hyaluronidase acting at neutral pH, found in the sperm membrane (102). The protein enables the
sperm to penetrate the cumulus cell layer surrounding the egg during the fertilization process (114).
Hyaluronidases have been implicated in tumorigenesis. Frost et al. (54) showed that head
and neck squamous cell carcinomas had inactivated their HYAL1 gene by aberrant splicing of pre-
mRNA, which rendered the cells devoid of hyaluronidase activity. Additional pieces of information
suggesting a broader function of the hyaluronidases in HA metabolism have been presented. The
presence of Hyal-1 in human plasma and urine (10)(53) and the novel specificity of Hyal-2,
hydrolysing HA to 20 kDa fragments (50-60 disaccharide units) instead of tetrasachharides (110),
opens up the field for further investigations.

33
PRESENT INVESTIGATION
6 Aims of the studyAt the start of this study the knowledge about the molecular mechanisms that regulate HA
biosynthesis was limited. Purification of proteins involved in HA polymerization as well as studying
the expression and regulation HAS proteins were general topics of interest. We were also interested
in investigating the effects of HA overproduction for cell function and cell behavior. The specific
aims of the thesis were to study:
• if HA is synthesized by a functional multi-protein complex. Are there proteins associated with
the HAS protein that might assist in the HA polymerization process or regulation of it? If yes, is
it possible to co-purify these accessory proteins from solubilized lysates of HA producing cells?
For these studies, we wanted to develop immunoprecipitating antibodies towards the HAS
protein(s).
• the polymerization directionality of HA synthesis. Is HA polymerized in the same way as the
sulfated GAGs, where monosaccharides are added to the non-reducing end of the polymer or
does it have a unique way of polymerization from the reducing end?
• the overexpression of HAS proteins. Do the HAS isozymes synthesize the same product and are
they equally efficient in HA polymerization? What are the consequences of HA overproduction
for cell function?
• if the HAS isoforms are differentially expressed and regulated. Which signals and growth
factors are involved in upregulation of the HAS genes? Are there signals that can down-regulate
HAS gene and protein expression and thereby decrease HA production?
• the role of UDPGDH in GAG biosynthesis. How is the GAG production affected in cells
overexpressing the UDPGDH and/or HAS enzymes? Is UDPGDH a regulatory or rate-limiting
enzyme in mammalian GAG biosynthesis?

34
7 HAS enzyme in glioma cells can be solubilized in an active form
(Paper I) The knowledge about the enzyme(s) involved in HA biosynthesis was limited when this project was
initiated; none of the HAS isozymes were purified to homogeneity or cloned due to the difficulties
to study membrane associated proteins. We were interested in purifying the HAS protein and
identifying possible accessory proteins involved in the HA polymerization process or regulation of
it. During the progress of the project, our research group was involved in the characterization of the
cloned HAS1 gene (172), which made it possible for us to develop peptide antibodies against the C-
terminal part of the protein.
7.1 Solubilization and purification The glioma cell line U-118 MG produces large amounts of HA and was therefore a potentially
useful source to try to purify and characterize HA polymerizing protein(s). It was important to have
maximal enzymatic activity at the start of the solubilization and purification process, since the
method available to monitor the HAS protein was a functional in vitro assay. We discovered that
when glioma cells were pretreated with the phorbol ester PMA in combination with testicular
hyaluronidase prior to cell harvesting, the in vitro enzymatic activity was increased 23-fold. A
battery of different detergents was tested for their ability to solubilize HAS activity. Only digitonin
extracted an appreciable amount of the enzymatic activity (26 % of non-solubilized protein),
compared to other detergents such as CHAPS, Triton X-100 and Thesit (2-6 %). We concluded that
the HAS protein, despite its intimate association with the plasma membrane, can be solubilized in
active form.
The enzymatically active HAS protein was submitted to gel chromatography on a Superdex-
200 column. The eluted fractions from the column were tested for protein content, HA synthase
activity and immunoblotting. The fractions eluted just after the the void volume (about 600 kDa)
contained the highest HA synthase activity and also exhibited the strongest bands in Western
blotting. These bands were around 66 kDa, under both reducing and non-reducing conditions. A
12-fold purification measured in specific enzymatic activity was achieved after the chromatography.
In summary, we could show that the active HAS enzyme was eluted in the high molecular
weight fractions from a Superdex-200 column, suggesting that the HAS protein(s) forms a
complex with other components. Gel electrophoreses and immunoblotting indicate that such a
complex, if it exists, is not brought together with covalent disulfide bonds, but probably weaker
ionic interactions. In the literature, several reports have indicated that HA is polymerized by a single
polypeptide chain with no additional components (199)(147)(229)(see section 4.2.2). An
explanation to our result from the chromatography, where the HAS protein is eluted in the 600 kDa

35
fraction, is that partly synthesized HA chains are still connected to the HAS proteins during the
purification process thereby increasing the apparent size of the eluted proteins. The cell preparations
used for solubilization have a high HA content (about 2 mg/mg protein) which supports this
explanation (Marianne Svarvare, unpublished observations).
7.2 Antibodies towards HAS proteins The cloning of HAS1 (172) and HAS2 (177) led us to develop peptide antibodies towards different
epitopes of HAS in our laboratory. The GVR-antibody was produced against the C-terminal portion
of HAS1 (peptide sequence GVRRLCRRRTGGYRVQV) and the TIY-antibody against the C-
terminal portion of HAS2 (peptide sequence TIYKESKKPFSESKQT). The specificity of the
antibodies was tested against protein preparations from recombinant expressed HAS (Paper I) or
from cell lines with specific HAS mRNA expression and high HA production (Paper III). This
approch was complicated by the finding that some cell lines have low HA production but a more
pronounced mRNA expression of HAS genes (Jonas Brinck, unpublished observations) and the
difficulty to find native HAS null cells. In paper I, the GVR-antibody was used in immunoblotting
of the HAS protein in glioma cells, a cell line which later showed to express mainly HAS2
(Paper III). The explanation for this could be two-fold. First, the specificity of polyclonal peptide
antibodies is not absolute and there is a similarity of about 30 % between the HAS1 and HAS2 and
HAS3 protein sequences at the epitope site of the GVR-antibody. Second, glioma cells may exhibit
a low HAS1 expression that we did not detect with Northern blots (Paper III).
To be able to identify and study a possible functional HAS protein complex we decided to
develop immunoprecipitating antibodies against HAS using a GST (glutathione-S-transferase)
fusion protein system. Four GST-HAS constructs were developed against conserved and unique
parts of the intracellular domain of various HAS isoforms, expressed in a low protease active
Escherichia coli strain (BL21) and the purified proteins were injected into rabbits for antisera
production. Initial attempts to use these antibodies for immunoprecipitations of HAS proteins have
been unsuccessful.
8 Is HA synthesized from the reducing end? (Paper I) The polymerization directionality of the HAS enzyme had been adressed before (185)(144)(145)
resulting in contradictory conclusions. Therefore we addressed this question by further developing
the pulse-chase experiments performed by Prehm (144).
HAS containing glioma cell membranes were incubated in two different pulse-chase
experiments (see Fig. 5). In the first approach (A), HA was polymerized during a pulse period in the
presence of UDP-[14C]GlcA and UDP-GlcNAc, followed by a chase period where the UDP-
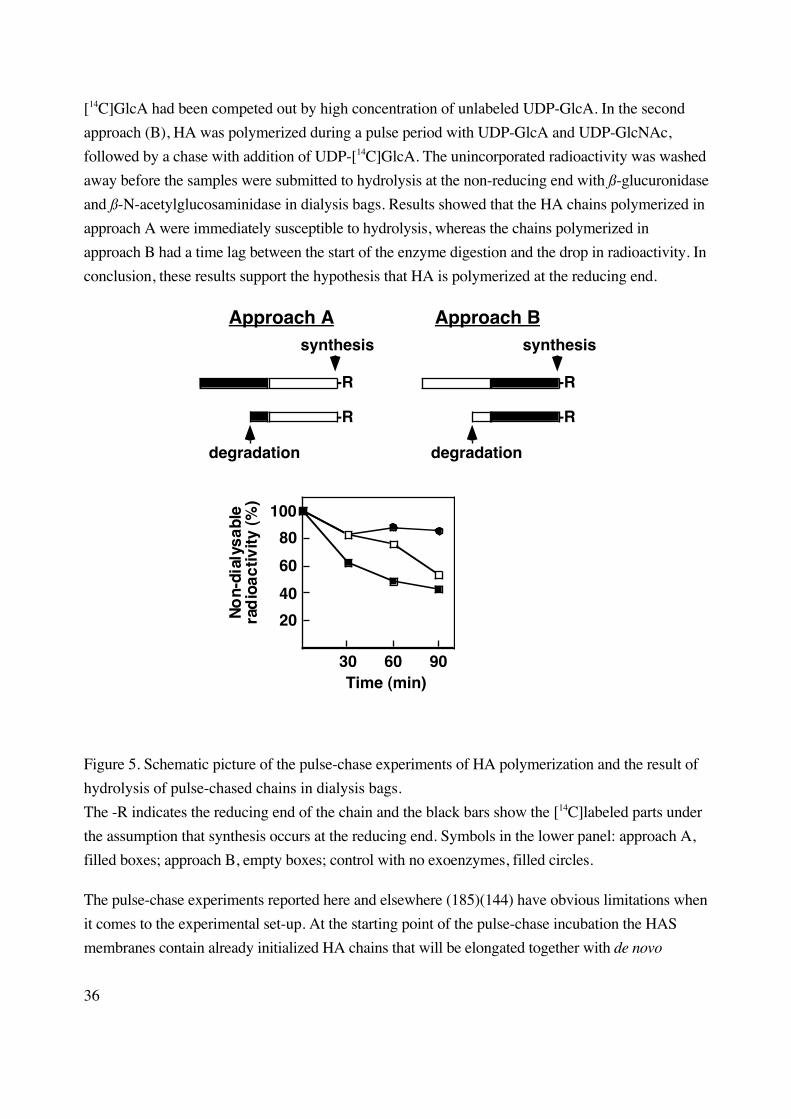
36
[14C]GlcA had been competed out by high concentration of unlabeled UDP-GlcA. In the second
approach (B), HA was polymerized during a pulse period with UDP-GlcA and UDP-GlcNAc,
followed by a chase with addition of UDP-[14C]GlcA. The unincorporated radioactivity was washed
away before the samples were submitted to hydrolysis at the non-reducing end with ß-glucuronidase
and ß-N-acetylglucosaminidase in dialysis bags. Results showed that the HA chains polymerized in
approach A were immediately susceptible to hydrolysis, whereas the chains polymerized in
approach B had a time lag between the start of the enzyme digestion and the drop in radioactivity. In
conclusion, these results support the hypothesis that HA is polymerized at the reducing end.
synthesis
-R
degradation
-R
Approach Asynthesis
-R
degradation
-R
Approach B
100
80
60
40
20
30 60 90
No
n-di
alys
able
rad
ioac
tivity
(%)
Time (min)
Figure 5. Schematic picture of the pulse-chase experiments of HA polymerization and the result of
hydrolysis of pulse-chased chains in dialysis bags.
The -R indicates the reducing end of the chain and the black bars show the [14C]labeled parts under
the assumption that synthesis occurs at the reducing end. Symbols in the lower panel: approach A,
filled boxes; approach B, empty boxes; control with no exoenzymes, filled circles.
The pulse-chase experiments reported here and elsewhere (185)(144) have obvious limitations when
it comes to the experimental set-up. At the starting point of the pulse-chase incubation the HAS
membranes contain already initialized HA chains that will be elongated together with de novo

37
synthesised chains, which will create heterogenity in the synthesized HA species. The digestion of
the pulse-chase labeled HA chains by exoglucosidases cannot be achieved completely giving
substantial levels of non-degradable radioactivity. One explanation to these residual levels could be
that the part of the HA chain closest to the HAS protein is less accessible for enzymatic degradation.
In summary, rigid evidence to the question of polymerization directionality of HA is still lacking.
9 Expression of HAS affects cell behavior (Paper II) After the identification and cloning of HAS in 1996 (i.e. HAS1) the cloning of two other isoforms
(HAS2, HAS3) was done during 1996-1997 by separate groups, thus revealing a potentially new
possibility for the regulation of the HA biosynthesis. The aim of this project was to study the
functional expression of the three HAS isoforms and the effects of overproduction of HA on cell
behavior.
9.1 Synthesis and accumulation of HA The Chinese hamster ovary (CHO) cell line was chosen for HAS expression due to its relatively
high genomic stability and high expression levels. Cells were transfected with cDNAs from human
HAS1, mouse HAS2 and mouse HAS3, and stabile expressing clones were established. The
sequence similarities between the mouse and human HAS orthologues is high, about 98 % (180),
indicating a conserved mechanisms of action between the species; we concluded that we could
compare the expression of the different cDNAs in the same system. Screening the clones for HA in
the cell culture media displayed a difference in HA synthesizing capacity between the different HAS
isoforms. The HAS3 transfected clones exhibited the highest HA synthesizing capacity (at
maximum 1.2 µg/105 cells/24 hours) compared to 0.07 µg and 0.6 µg per 105 cells/24 hours for the
most efficiently transfected HAS1 and HAS2 clones, respectively. The increase seen in the HAS3
clone corresponds to a 1000-fold increase in HA production compared to wild-type CHO cells. The
HA content in the culture media of the transfected cells (up to 1.2 µg/105 cells/24 hours) is in the
same range as in media of human mesothelial cells and glioma cells (Paper III). This indicates that
the HA amounts produced are within the physiological range, although not for the CHO cell line.
We also wanted to investigate if overexpression of HAS enzymes could lead to increased
amounts of cell associated HA. A classical way of visualizing pericellular HA since the 1960’s has
been the in vitro particle exclusion assay (18), where the cell surface associated HA can be seen as a
halo between the cell membrane and excluded particles (e.g. erythrocytes). In our CHO cell
model, we could show that whereas wild-type cells did not have the capacity to form HA matrices,

38
their transfected counterparts attained this ability. The HA synthesizing capacity needed to generate
such matrices was about 1 µg of HA per 105 cells and 24 hours. This accumulation of HA around
transfected cells could be confirmed by comparing cell associated [3H]HA amounts from low and
high HA producing clones.
9.2 HA chain length The chain length of HA synthesized by HAS transfected CHO cells were studied both after in vitro
and in vivo synthesis. The in vitro assay is based on HA polymerization by membrane preparations
incubated with UDP-[14C]GlcA,UDP-GlcNAc and MgCl2, while HA synthesized in intact cells in
the presence of [3H]glucosamine mimics in vivo conditions. Results showed that only HAS2
produced high molecular weight HA (Mr >3.9 x 106) when assayed in vitro, compared to HAS1
(average Mr 0.12 x 106) and HAS3 (Mr 0.12-1 x 106). However, under in vivo conditions, all three
isozymes generated HA of high molecular weight (Mr >3.9 x 106). As a comparison, HA chains
synthesized by mesothelial cells (cells with native expression of HAS1, HAS2 and HAS3;
Paper III) were of high molecular weight both in vitro and in vivo.
We also studied the HA chain length after solubilization of the HAS protein from glioma
cells (Paper I) and after stimulation of mesothelial cells with various growth factors and other
compounds (Paper III). Glioma cells, which predominantly express the HAS2 isoform, produced
high molecular weight HA when assayed in vitro. Solubilization of the glioma membranes with
digitonin prior to the assay rendered a more polydisperse product, but still HA polymers were still
produced. Treatment of mesothelial cells with growth factors such as PDGF-BB and TGF-ß led to
HA synthesis of high molecular weight in vivo. Interestingly, phorbol ester PMA pretreatment
induced synthesis of HA chains of polydisperse character (Mr 0.25 x 106 to over 3.9 x 106).
The results show that all three HAS isoforms, whether recombinantly expressed in CHO
cells or native in human mesothelial cells, have the capacity to synthesize high molecular weight HA
in vivo. We also show that the HA chain length is dependent on the cell type where it is synthesized.
Taken together with the result that the phorbol ester PMA can influence the chain size in mesothelial
cells, interesting questions are raised about the molecular mechanims behind the regulation of HA
chain length, which needs further investigation. The matter is important since the function of HA
polymers is partly dependent on their size. Large HA polymers are important structural elements in
the extracellular matrix (106) whereas short HA fragments can induce angiogenesis (221)(148) and
inflammatory reponses (121)(80).

39
9.3 Cell migration The migration capacity of wild-type CHO cells and HAS3 transfectants exhibiting different HA
synthesizing capacities were quantified using a Boyden chamber assay. The results revealed that the
migration capacity was inversely correlated to the HA synthesizing capacity. When excessive
amounts of exogenous HA was added to the migration assay, the migration was inhibited for cells
producing low amounts of HA, but not for high producing cells. Another finding was that the
amounts of cell surface CD44 (as detected by flow cytometry) decreased in high HA producing
cells. For the HAS3 clone producing the highest amounts of HA, a 75 % decrease in migration rate
and a 50 % reduction in cell surface CD44 was observed. When an antibody towards CD44,
blocking the HA binding site, was included in the assays the migration was inhibited up to 50 %.
In conclusion, our results suggests that the overproduction and accumulation of pericellular
HA impairs normal cell function and reduces cell migration in HAS transfected cells. The
concomitant down-regulation of CD44 molecules on the surface of these cells explains partly the
reduction in their migratory ability. The total amount of CD44 does not seem to reduce significantly
when visualized in Western blotting of total cell lysates (Jonas Brinck, unpublished observations),
suggesting a receptor mediated endocytosis of the receptor; such a mechanism has been described
in the literature (22). Knudson and Knudson (97) have presented the hypothesis where the
establishment of a pericellular HA matrix would interfere with and disrupt normal cell migration.
This model has to some extent been experimentally supported by Perris et al. (141), where HA
binding proteoglycans that promote pericellular matrix formation was shown to inhibit migration in
neural crest cells.
10 HAS isoforms show specificity in gene regulation (Paper III) Under pathological conditions such as chronic inflammation and carcinogenesis an upregulation of
HA production is seen, which could be of importance for the pathological development of the
disease state (190)(203). To understand the regulation of the HAS genes we therefore investigated
the expression HAS genes in response to growth conditions and external stimuli.
10.1 Effects of culture conditions Subconfluent and confluent cultures of glioma cells, mesothelioma cells, mesothelial cells, skin
fibroblasts and lung fibroblasts were tested for their ability to express the three HAS isoforms. The
culture conditions affected the HAS expression levels by reducing all HAS genes when the cell
density increased, indicating a cell-cell contact as a negative factor for HAS expression. Northern
blots revealed that only mesothelial cells expressed all three HAS isoforms. The mRNA levels of
HAS in the various cell cultures were well correlated with their HA production. Mesothelial cells

40
displayed the highest HA synthesizing capacity (approx. 18 ng HA per 103 cells after 24 hours of
culture). Interestingly, mesothelioma cells had a different HAS mRNA expression pattern compared
to mesothelial cells; mesothelioma cells expressed HAS3 isoform and synthesized only minute
amounts of HA.
10.2 Effects of external stimuli Mesothelial cells were chosen for studying the differential expression of the HAS genes in response
to external stimuli since they have native expression of all three isoforms. We stimulated mesothelial
cells with the PDGF-BB and TGF-ß1, which are known to be important regulators of HA synthesis
in cells of mesenchymal origin (68)(186). Stimulation of the cells with PDGF-BB increased the
mRNA level of HAS2 to a peak at 6 hours whereas TGF-ß1 treatment attenuated it. The HAS2
mRNA increase observed after PDGF-BB stimulation was accompanied by increased protein levels
and production of high molecular weight HA. The effects of growth factors on HAS1 and HAS3
mRNA expression levels were less pronounced as for HAS2. Treatment of the mesothelial cells
with TGF-ß1 increased HAS1 isoform at both the mRNA and protein level whereas HAS3 isoform
was not markedly affected by either PDGF-BB or TGF-ß1.
Treatment of mesothelial cells with hydrocortisone (125 µg/ml) suppressed HAS2 mRNA
expression considerably (about 80 % suppression compared to normal levels), whereas HAS1 and
HAS3 mRNA levels only decreased marginally or not at all. The down-regulatory effect of
hydrocortisone at HAS2 mRNA level was also seen at the protein level and on the production of
HA. Approximately a 40 % decrease in HA production was observed after 6 hours of treatment with
hydrocortisone compared to control treated cells.
In conclusion, we have found that the three HAS genes are differentially regulated in
different cell types in response to growth conditions and external stimuli. It seems that the growth
factors mediate their major stimulatory effects through the HAS2 isoform, and therefore HAS2
could be the most important enzyme for HA production in the body. Such an hypothesis is in line
with the fact that only HAS2 knock-out mice are embryonically lethal compared to HAS1 and
HAS3 knock-outs (181)(16). In addition, the functional inhibition of HA production seen, after
down-regulation of HAS2 caused by hydrocortisone treatment, could constitute a strategy for
pharmacological treatment of disease in a future perspective.
11 The role of UDP-glucose dehydrogenase in mammalianglycosaminoglycan biosynthesis (Paper IV) The UDPGDH enzyme converts UDP-Glc to UDP-GlcA, which is an essential building block in
mammalian GAG biosynthesis. The enzyme has been postulated to play a key regulatory role in the
cell wall polysaccharide biosynthesis in plants (192)(154) and is indispensable for vertebrate

41
embryonic development (179)(181) by being the sole producer of UDP-GlcA. The aim of this
study was to investigate the role of UDPGDH in mammalian GAG biosynthesis.
We have created an experimental cell system where the rate of synthesis and consumption of
UDP-GlcA can be modulated in human embryonic kidney (293T) cells by transient overexpression
of UDPGDH and HAS3. The dehydrogenase and HA synthase activities were measured in vitro
and were found to be over 400-fold greater in favor of the dehydrogenase activity in 293T cells;
even after the cells were transfected with UDPGDH and/or HAS3 the overcapacity of the
dehydrogenase enzyme persisted. The mRNA expression of UDPGDH was also investigated in
CHO cells stably transfected with HAS3. The basal mRNA expression level of UDPGDH in wild-
type cells was sufficient for HA production in HAS3 transfected cells exhibiting high HA
production; the levels of mRNA in HAS3 expressing cells did not increase after transfection.
The amounts of synthesized GAGs were determined after UDPGDH and/or HAS3
transfection. Overexpression of UDPGDH increased the production of CS about 1.5-fold, but did
not affect the amount of synthesized HA and HS. HAS3 transfection on the other hand, led to a 20-
fold increase of HA production. The overproduction of HA was in part at the expense of HS
production which was reduced to 50 % compared to mock transfected cells (CS amounts were
normal). Co-expression of UDPGDH with HAS3 did not rescue HS production back to normal
levels or increase HA or CS production. The isolated HA, HS and CS chains from the UDPGDH
and/or HAS3 transfected cells did not display any major structural changes, in terms of their chain
lengths or their the degree of sulfation (HS and CS), compared to GAG chains from mock
transfected cells.
In conclusion, our findings indicate that the activity of UDPGDH is not rate limiting in
mammalian GAG biosynthesis. The diverse effects seen on the HS and CS production in response
to UDPGDH and HAS3 transfection could suggest that the biosynthetic machineries responsible
for HS and CS synthesis are located in separate Golgi compartments, or that they have different
affinities for the substrates (Km-values). Studies are currently underway to determine the
concentrations and the fluctuations in the UDP-sugar pools, which will assist in the interpretation of
the effects seen on HA, HS and CS production.

42
12 Conclusions• We have shown that the HAS protein(s) can be solubilized in active form. Results indicate that
the partially purified HAS isozyme(s) is most likely not asscociated covalently with any
accessory components that may be needed in the regulation of HA biosynthesis.
• We have results supporting the hypothesis that HA is polymerized at the reducing end of the
polymer thereby being unique in the polymerization directionality in the family of GAGs.
• We have shown that all three HAS isoforms are able to synthesize high molecular weight HA in
intact mammalian cells and that the regulation of the HA chain length involves cell specific
elements as well as external stimulatory factors.
• We have shown that overexpression of HAS in CHO cells leads to increased HA production and
accumulation of pericellular HA and that these cells exhibit a reduced migration capacity and
reduced amounts of cell surface CD44 molecules.
• We have shown that the HAS1, HAS2 and HAS3 genes are differentially expressed and
regulated in response to culture conditions and external stimuli. PDGF-BB and TGF-ß1 are
important regulators of HAS at both transcriptional and translational level.
• We have results indicating that UDPGDH is not a rate-limiting enzyme in mammalian GAG
biosynthesis.

43
13 Future perspectives The sequencing of the whole human genome is soon complete (175), revealing the genetic code of
man, whereas the work with discovering the function of many of these genes is still in its infancy.
Although the three genes involved in the biosynthesis of HA were sequenced and cloned during the
time period of this thesis study, the action and regulation of the HAS enzymes are still topics of
scientific interest and many questions remain to be answered.
The mechanism of how small HA fragments, found in extracellular matrix of various
mammalian tissues under physiological and pathological conditions, are formed is still not resolved.
Reports by us and others (Paper II, (180)(86)) indicating that HA can be synthesized as shorter
chains can be an in vitro finding with little or no physiological relevance. Two main stand-points can
be taken when trying to explain the origin of these fragments. The first conjecture is that HA is
always synthesized as full size polymers (Mr 5-10 x 106) in vivo. In such a model, the production of
low molecular weight HA could instead be due to hyaluronidase activity. The fact that the human
hyaluronidase gene family now comprise six paralogous genes corresponding to five different
hyaluronidase proteins strengthens their role in HA metabolism (21)(184). Specific growth factors
could thereby regulate hyaluronidase expression and promote or decrease the formation of HA
oligosaccharides. However, before these enzymes (PH-20 excluded) can be ascribed an essential
role in HA chain regulation rigid evidence for hyaluronidase activity in the extracellular matrix at
physiological pH must first be provided.
The second conjecture is that the regulation of the HA chain length is controled by the HAS
molecule itself, or an accessory component, during the process of synthesis. Preliminary results by
Spicer and co-workers show that a positive aa cluster in the C-terminal part of the HAS protein
displays chain length regulation capacity by binding to the nascent intracellular part of the HA chain
(178). More suggestive evidence, based on the fact that HA polymerization is regulated in a cell
specific manner (Paper II)(180)(86), indicates that an additional component might be involved. The
nature of this component, if it exists, remains to be elucidated.
In several pathological conditions such as carcinogenesis and chronical inflammation
elevated levels of HA can be detected. Is this increased production of HA a primary or secondary
effect of the pathological condition and how are the other GAGs affected by the shift in HA
synthesis? In paper IV we investigated the potentially regulatory role of UDP-sugars and
UDPGDH for GAG biosynthesis. Our results indicate that the UDPGDH enzyme does not exert a
rate-limiting effect on mammalian GAG biosynthesis. However, the HS production was markedly
affected in cells overproducing HA, suggesting that shifts in UDP-sugar fluxes can influence their
GAG production quantitatively. It will be of importance to investigate the susceptibility of GAG
biosynthetic machineries to changes in UDP-sugar concentrations, since it could constitute a
mechanism of regulation. The functional diversity of the three HAS isoforms in terms of their HA

44
coat forming capacity (86) could be an example of this kind of regulation. Also, the observed
vulnerability of the HS production in HA overproducing cells could lead to an impairment in the
affected cells’ growth factor signaling, which is known to be mediated through HS containing
proteoglycans (159).

45
ACKNOWLEDGEMENTS
I wish to express my gratitude to:
Evi Heldin, my supervisor, for sharing her enthusiasm for science with me and teaching me to be a
critical scientist.
Professors Ulf Lindahl and Torvard Laurent for guidance and encouragement in the field of science
in general and glycosaminoglycans in particular.
Marion Kusche-Gullberg and Dorothe Spillman for generous supervision in the field of complex
polysaccharides. All my co-authors and collaborators for their work and advice in the four papers
presented in this thesis.
Marianne Svarvare for always being at hand with excellent technical advice and for teaching me all
the practical things I thought I already knew but apparently did not.
Örjan Zetterqvist, my mentor in everything that has concerned teaching of the graduate students, for
inspiring discussion and for entrusting me with part of the graduate student education. It has been
very stimulating and challenging!
Annica, Johan and Mehdi for creating that day-to-day (year-to-year!) atmosphere that makes it
possible to get through four years of pipetting and cell culture work. Don’t give up – your day will
come soon! Special thanks to Maria for letting me borrow her brother’s “slagborrmaskin” and
many, many other things. Also thanks to all the people in Ulf Lindahl’s group, and especially
Elisabet, for partially adopting me (and “my” polysaccharide) into your group.
All the persons, past and present, at the department for creating a pleasant working atmosphere.
The people in Lena Claesson-Welsh’s group for letting me in on a few secrets in field of cell
biology and for teaching me how to use your (expensive) equipment.
Anna for being at my side.

46
REFERENCES1. Aruffo, A., Stamenkovic, I., Melnick, M., Underhill, C.B. and Seed, B. (1990) CD44 is the
principal cell surface receptor for hyaluronate. Cell. 61:1303-1313.
2. Asplund, T. and Heldin, P. (1994) Hyaluronan receptors are expressed on human malignant
mesothelioma cells but not on normal mesothelial cells. Cancer Res. 54:4516-4523.
3. Asplund, T., Versnel, M.A., Laurent, T.C. and Heldin, P. (1993) Human mesothelioma cells
produce factors that stimulate the production of hyaluronan by mesothelial cells and
fibroblasts. Cancer Res. 53:388-392.
4. Bajorath, J., Greenfield, B., Munro, S.B., Day, A.J. and Aruffo, A. (1998) Identification of
CD44 residues important for hyaluronan binding and delineation of the binding site. J Biol
Chem. 273:338-343.
5. Balduini, C., Brovelli, A. and Castellani, A.A. (1970) Biosynthesis of glycosaminoglycans in
bovine cornea. The effect of uridine diphosphate xylose. Biochem J. 120:719-723.
6. Banerji, S., Day, A.J., Kahmann, J.D. and Jackson, D.G. (1998) Characterization of a
functional hyaluronan-binding domain from the human CD44 molecule expressed in
Escherichia coli. Protein Expr Purif. 14:371-381.
7. Banerji, S., Ni, J., Wang, S.X., Clasper, S., Su, J., Tammi, R., Jones, M. and Jackson, D.G.
(1999) LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for
hyaluronan. J Cell Biol. 144:789-801.
8. Bansal, M.K. and Mason, R.M. (1986) Evidence for rapid metabolic turnover of hyaluronate
synthetase in Swarm rat chondrosarcoma chondrocytes. Biochem J. 236:515-519.
9. Bdolah, A. and Feingold, D.S. (1968) Uridine diphosphate-D-glucose dehydrogenase of hen
oviduct. Biochim Biophys Acta. 159:176-178.
10. Bonner, W.M., Jr. and Cantey, E.Y. (1966) Colorimetric method for determination of serum
hyaluronidase activity. Clin Chim Acta. 13:746-752.
11. Bourguignon, L.Y., Kalomiris, E.L. and Lokeshwar, V.B. (1991) Acylation of the lymphoma
transmembrane glycoprotein, GP85, may be required for GP85-ankyrin interaction. J Biol
Chem. 266:11761-11765.
12. Bourin, M.C. and Lindahl, U. (1993) Glycosaminoglycans and the regulation of blood
coagulation. Biochem J. 289:313-330.
13. Brandli, A.W., Hansson, G.C., Rodriguez-Boulan, E. and Simons, K. (1988) A polarized
epithelial cell mutant deficient in translocation of UDP-galactose into the Golgi complex. J
Biol Chem. 263:16283-16290.

47
14. Brown, T.A., Bouchard, T., St. John, T., Wayner, E. and Carter, W.G. (1991) Human
keratinocytes express a new CD44 core protein (CD44E) as a heparan-sulfate intrinsic
membrane proteoglycan with additional exons. J Cell Biol. 113:207-221.
15. Calabro, A. and Hascall, V.C. (1994) Differential effects of brefeldin A on chondroitin sulfate
and hyaluronan synthesis in rat chondrosarcoma cells. J Biol Chem. 269:22764-22770.
16. Camenisch, T.D., Spicer, A.P., Brehm-Gibson, T. and McDonald, J.A. (1999) Identification
of a hyaluronan synthase2 as a critical source of hyaluronan during mammalian
embryogenesis. Conference abstract in "International Conference on Molecular Interactions
of Proteoglycans", Kanagawa, Japan.
17. Cheung, W.F., Eriksson, I., Kusche-Gullberg, M., Lindhal, U. and Kjellen, L. (1996)
Expression of the mouse mastocytoma glucosaminyl N-deacetylase/ N-sulfotransferase in
human kidney 293 cells results in increased N-sulfation of heparan sulfate. Biochemistry.
35:5250-5256.
18. Clarris, B.J. and Fraser, J.R. (1968) On the pericellular zone of some mammalian cells in
vitro. Exp Cell Res. 49:181-193.
19. Crater, D.L., Dougherty, B.A. and van de Rijn, I. (1995) Molecular characterization of hasC
from an operon required for hyaluronic acid synthesis in group A streptococci.
Demonstration of UDP- glucose pyrophosphorylase activity. J Biol Chem. 270:28676-
28680.
20. Crater, D.L. and van de Rijn, I. (1995) Hyaluronic acid synthesis operon (has) expression in
group A streptococci. J Biol Chem. 270:18452-18458.
21. Csoka, A.B., Scherer, S.W. and Stern, R. (1999) Expression analysis of six paralogous
human hyaluronidase genes clustered on chromosomes 3p21 and 7q31. Genomics. 60:356-
361.
22. Culty, M., Nguyen, H.A. and Underhill, C.B. (1992) The hyaluronan receptor (CD44)
participates in the uptake and degradation of hyaluronan. J Cell Biol. 116:1055-1062.
23. Dalchau, R., Kirkley, J. and Fabre, J.W. (1980) Monoclonal antibody to a human brain-
granulocyte-T lymphocyte antigen probably homologous to the W 3/13 antigen of the rat. Eur
J Immunol. 10:745-749.
24. Dasgupta, A., Takahashi, K., Cutler, M. and Tanabe, K.K. (1996) O-linked glycosylation
modifies CD44 adhesion to hyaluronate in colon carcinoma cells. Biochem Biophys Res
Commun. 227:110-117.
25. Day, A.J. and Parkar, A.A. (1998) The structure of the Link module: a hyaluronan binding
domain. In "The Chemistry, Biology and Medical Applications of Hyaluronan and its
Derivatives". Vol. 72. Laurent, T.C., editor. Portland Press, London. 141-147.

48
26. DeAngelis, P.L. (1999) Hyaluronan synthases: fascinating glycosyltransferases from
vertebrates, bacterial pathogens, and algal viruses. Cell. Mol. Life Sci. 56:670-682.
27. DeAngelis, P.L. (1999) Molecular directionality of polysaccharide polymerization by the
Pasteurella multocida hyaluronan synthase. J Biol Chem. 274:26557-26562.
28. DeAngelis, P.L. and Achyuthan, A.M. (1996) Yeast-derived recombinant DG42 protein of
Xenopus can synthesize hyaluronan in vitro. J Biol Chem. 271:23657-23660.
29. DeAngelis, P.L., Jing, W., Drake, R.R. and Achyuthan, A.M. (1998) Identification and
molecular cloning of a unique hyaluronan synthase from Pasturella multicoda. J Biol Chem.
273:8454-8458.
30. DeAngelis, P.L., Jing, W., Graves, M.V., Burbank, D.E. and Van Etten, J.L. (1997)
Hyaluronan synthase of chlorella virus PBCV-1. Science. 278:1800-1803.
31. DeAngelis, P.L., Papaconstantinou, J. and Weigel, P.H. (1993) Molecular cloning,
identification, and sequence of the hyaluronan synthase gene from group A Streptococcus
pyogenes. J Biol Chem. 268:19181-19184.
32. DeGrendele, H.C., Estess, P. and Siegelman, M.H. (1997) Requirement for CD44 in activated
T cell extravasation into an inflammatory site. Science. 278:672-675.
33. Dougherty, B.A. and van de Rijn, I. (1992) Molecular characterization of a locus required for
hyaluronic acid capsule production in group A streptococci. J Exp Med. 175:1291-1299.
34. Dougherty, B.A. and van de Rijn, I. (1993) Molecular characterization of hasB from an
operon required for hyaluronic acid synthesis in group A streptococci. Demonstration of
UDP- glucose dehydrogenase activity. J Biol Chem. 268:7118-7124.
35. Dougherty, B.A. and van de Rijn, I. (1994) Molecular characterization of hasA from an
operon required for hyaluronic acid synthesis in group A streptococci. J Biol Chem. 269:169-
175.
36. Eggli, P.S. and Graber, W. (1995) Association of hyaluronan with rat vascular endothelial and
smooth muscle cells. J Histochem Cytochem. 43:689-697.
37. Ellis, I., Banyard, J. and Schor, S.L. (1997) Differential response of fetal and adult fibroblasts
to cytokines: cell migration and hyaluronan synthesis. Development. 124:1593-1600.
38. Ellis, I.R. and Schor, S.L. (1996) Differential effects of TGF-beta1 on hyaluronan synthesis
by fetal and adult skin fibroblasts: implications for cell migration and wound healing. Exp
Cell Res. 228:326-333.
39. Entwistle, J., Hall, C.L. and Turley, E.A. (1996) HA receptors: regulators of signalling to the
cytoskeleton. J Cell Biochem. 61:569-577.

49
40. Entwistle, J., Zhang, S., Yang, B., Wong, C., Li, Q., Hall, C.L., A, J., Mowat, M., Greenberg,
A.H. and Turley, E.A. (1995) Characterization of the murine gene encoding the hyaluronan
receptor RHAMM. Gene. 163:233-238.
41. Eriksson, S., Fraser, J.R., Laurent, T.C., Pertoft, H. and Smedsrod, B. (1983) Endothelial cells
are a site of uptake and degradation of hyaluronic acid in the liver. Exp Cell Res. 144:223-
228.
42. Faassen, A.E., Schrager, J.A., Klein, D.J., Oegema, T.R., Couchman, J.R. and McCarthy, J.B.
(1992) A cell surface chondroitin sulfate proteoglycan, immunologically related to CD44, is
involved in type I collagen-mediated melanoma cell motility and invasion. J Cell Biol.
116:521-531.
43. Feinberg, R.N. and Beebe, D.C. (1983) Hyaluronate in vasculogenesis. Science. 220:1177-
1179.
44. Fessler, J.H. and Fessler, L.I. (1966) Electron microscopic visualization of the polysaccharide
hyaluronic acid. Proc Natl Acad Sci U S A. 56:141-147.
45. Fieber, C., Plug, R., Sleeman, J., Dall, P., Ponta, H. and Hofmann, M. (1999) Characterisation
of the murine gene encoding the intracellular hyaluronan receptor IHABP (RHAMM). Gene.
226:41-50.
46. Fraser, J.R., Appelgren, L.E. and Laurent, T.C. (1983) Tissue uptake of circulating hyaluronic
acid. A whole body autoradiographic study. Cell Tissue Res. 233:285-293.
47. Fraser, J.R., Kimpton, W.G., Laurent, T.C., Cahill, R.N. and Vakakis, N. (1988) Uptake and
degradation of hyaluronan in lymphatic tissue. Biochem J. 256:153-158.
48. Fraser, J.R., Laurent, T.C. and Laurent, U.B. (1997) Hyaluronan: its nature, distribution,
functions and turnover. J Intern Med. 242:27-33.
49. Fraser, J.R., Laurent, T.C., Pertoft, H. and Baxter, E. (1981) Plasma clearance, tissue
distribution and metabolism of hyaluronic acid injected intravenously in the rabbit. Biochem
J. 200:415-424.
50. Freeze, H. (1999) Monosaccharide Metabolism. In "Essentials of Glycobiology". Varki, A.,
Cummings, R., Esko, J., Freeze, H., Hart, G. and Marth, J., editors. Coldspring Harbor
Laboratory Press, New York. 69-84.
51. Friedlander, D.R., Milev, P., Karthikeyan, L., Margolis, R.K., Margolis, R.U. and Grumet, M.
(1994) The neuronal chondroitin sulfate proteoglycan neurocan binds to the neural cell
adhesion molecules Ng-CAM/L1/NILE and N-CAM, and inhibits neuronal adhesion and
neurite outgrowth. J Cell Biol. 125:669-680.
52. Frost, G.I., Csoka, T. and Stern, R. (1996) The hyaluronidases: A Chemical, Biological and
Clinical Overview. Trends in Glycoscience and Glycotechnology. 8:419-434.

50
53. Frost, G.I., Csoka, T.B., Wong, T. and Stern, R. (1997) Purification, cloning, and expression
of human plasma hyaluronidase. Biochem Biophys Res Commun. 236:10-15.
54. Frost, G.I., Mohapatra, G., Wong, T.M., Csoka, A.B., Gray, J.W. and Stern, R. (2000)
HYAL1LUCA-1, a candidate tumor suppressor gene on chromosome 3p21.3, is inactivated in
head and neck squamous cell carcinomas by aberrant splicing of pre-mRNA. Oncogene.
19:870-877.
55. Gainey, P.A. and Phelps, C.F. (1972) Uridine diphosphate glucuronic acid production and
utilization in various tissues actively synthesizing glycosaminoglycans. Biochem J. 128:215-
227.
56. Glaser, L. (1959) The Biosynthesis of N-Acetylgalactoseamine. J Biol Chem. 234:2801-
2805.
57. Greiling, H. (1994) Structure and biological functions of keratan sulfate proteoglycans. Exs.
70:101-122.
58. Grumet, M., Flaccus, A. and Margolis, R.U. (1993) Functional characterization of chondroitin
sulfate proteoglycans of brain: interactions with neurons and neural cell adhesion molecules. J
Cell Biol. 120:815-824.
59. Gunthert, U., Hofmann, M., Rudy, W., Reber, S., Zoller, M., Haussmann, I., Matzku, S.,
Wenzel, A., Ponta, H. and Herrlich, P. (1991) A new variant of glycoprotein CD44 confers
metastatic potential to rat carcinoma cells. Cell. 65:13-24.
60. Hall, C.L., Yang, B., Yang, X., Zhang, S., Turley, M., Samuel, S., Lange, L.A., Wang, C.,
Curpen, G.D., Savani, R.C., Greenberg, A.H. and Turley, E.A. (1995) Overexpression of the
hyaluronan receptor RHAMM is transforming and is also required for H-ras transformation.
Cell. 82:19-28.
61. Handley, C.J. and Phelps, C.F. (1972) The biosynthesis in vitro of keratan sulphate in bovine
cornea. Biochem J. 128:205-213.
62. Handley, C.J. and Phelps, C.F. (1972) The concentrations of sugar nucleotides in bovine
corneal epithelium and endothelium. Biochem J. 127:911-912.
63. Hardwick, C., Hoare, K., Owens, R., Hohn, H.P., Hook, M., Moore, D., Cripps, V., Austen, L.,
Nance, D.M. and Turley, E.A. (1992) Molecular cloning of a novel hyaluronan receptor that
mediates tumor cell motility. J Cell Biol. 117:1343-1350.
64. Hathcock, K.S., Hirano, H., Murakami, S. and Hodes, R.J. (1993) CD44 expression on
activated B cells. Differential capacity for CD44-dependent binding to hyaluronic acid. J
Immunol. 151:6712-6722.
65. Hayen, W., Goebeler, M., Kumar, S., Riessen, R. and Nehls, V. (1999) Hyaluronan stimulates
tumor cell migration by modulating the fibrin fiber architecture. J Cell Sci. 112:2241-2251.

51
66. Heinegard, D. and Oldberg, A. (1989) Structure and biology of cartilage and bone matrix
noncollagenous macromolecules. Faseb J. 3:2042-2051.
67. Heldin, C.H. and Westermark, B. (1999) Mechanism of action and in vivo role of platelet-
derived growth factor. Physiol Rev. 79:1283-1316.
68. Heldin, P., Asplund, T., Ytterberg, D., Thelin, S. and Laurent, T.C. (1992) Characterization of
the molecular mechanism involved in the activation of hyaluronan synthetase by platelet-
derived growth factor in human mesothelial cells. Biochem J. 283:165-170.
69. Heldin, P. and Pertoft, H. (1993) Synthesis and assembly of the hyaluronan-containing coats
around normal human mesothelial cells. Exp Cell Res. 208:422-429.
70. Heldin, P., Suzuki, M., Teder, P. and Pertoft, H. (1995) Chondroitin sulfate proteoglycan
modulates the permeability of hyaluronan-containing coats around normal human mesothelial
cells. J Cell Physiol. 165:54-61.
71. Heldin, P., Torres, M., Ytterberg, D. and Bergh, J. (1996) Differential synthesis and binding
of hyaluronan by human breast cancer cell lines: Relationship to hormone receptor status.
Oncology reports. 3:1011-1016.
72. Henke, C.A., Roongta, U., Mickelson, D.J., Knutson, J.R. and McCarthy, J.B. (1996) CD44-
related chondroitin sulfate proteoglycan, a cell surface receptor implicated with tumor cell
invasion, mediates endothelial cell migration on fibrinogen and invasion into a fibrin matrix. J
Clin Invest. 97:2541-2552.
73. Hirschberg, C.B., Robbins, P.W. and Abeijon, C. (1998) Transporters of nucleotide sugars,
ATP, and nucleotide sulfate in the endoplasmic reticulum and Golgi apparatus. Annu Rev
Biochem. 67:49-69.
74. Hirschberg, C.B. and Snider, M.D. (1987) Topography of glycosylation in the rough
endoplasmic reticulum and Golgi apparatus. Annu Rev Biochem. 56:63-87.
75. Hofmann, M., Assmann, V., Fieber, C., Sleeman, J.P., Moll, J., Ponta, H., Hart, I.R. and
Herrlich, P. (1998) Problems with RHAMM: a new link between surface adhesion and
oncogenesis? Cell. 95:591-593.
76. Hofmann, M., Fieber, C., Assmann, V., Gottlicher, M., Sleeman, J., Plug, R., Howells, N., von
Stein, O., Ponta, H. and Herrlich, P. (1998) Identification of IHABP, a 95 kDa intracellular
hyaluronate binding protein. J Cell Sci. 111:1673-1684.
77. Holmer, E., Lindahl, U., Backstrom, G., Thunberg, L., Sandberg, H., Soderstrom, G. and
Anderson, L.O. (1980) Anticoagulant activities and effects on platelets of a heparin fragment
with high affinity for antithrombin. Thromb Res. 18:861-869.
78. Honda, A., Noguchi, N., Takehara, H., Ohashi, Y., Asuwa, N. and Mori, Y. (1991)
Cooperative enhancement of hyaluronic acid synthesis by combined use of IGF-I and EGF,

52
and inhibition by tyrosine kinase inhibitor genistein, in cultured mesothelial cells from rabbit
pericardial cavity. J Cell Sci. 98:91-98.
79. Honda, A., Sekiguchi, Y. and Mori, Y. (1993) Prostaglandin E2 stimulates cyclic AMP-
mediated hyaluronan synthesis in rabbit pericardial mesothelial cells. Biochem J. 292:497-
502.
80. Horton, M.R., McKee, C.M., Bao, C., Liao, F., Farber, J.M., Hodge-DuFour, J., Pure, E.,
Oliver, B.L., Wright, T.M. and Noble, P.W. (1998) Hyaluronan fragments synergize with
interferon-gamma to induce the C-X-C chemokines mig and interferon-inducible protein-10
in mouse macrophages. J Biol Chem. 273:35088-35094.
81. Hua, Q., Knudson, C.B. and Knudson, W. (1993) Internalization of hyaluronan by
chondrocytes occurs via receptor-mediated endocytosis. J Cell Sci. 106:365-375.
82. Iozzo, R.V. (1998) Matrix proteoglycans: from molecular design to cellular function. Annu
Rev Biochem. 67:609-652.
83. Iozzo, R.V. and Murdoch, A.D. (1996) Proteoglycans of the extracellular environment: clues
from the gene and protein side offer novel perspectives in molecular diversity and function.
Faseb J. 10:598-614.
84. Ishihara, M., Guo, Y., Wei, Z., Yang, Z., Swiedler, S.J., Orellana, A. and Hirschberg, C.B.
(1993) Regulation of biosynthesis of the basic fibroblast growth factor binding domains of
heparan sulfate by heparan sulfate-N-deacetylase/N-sulfotransferase expression. J Biol
Chem. 268:20091-20095.
85. Itano, N. and Kimata, K. (1996) Expression cloning and molecular characterization of HAS
protein, a eukaryotic hyaluronan synthase. J Biol Chem. 271:9875-9878.
86. Itano, N., Sawai, T., Yoshida, M., Lenas, P., Yamada, Y., Imagawa, M., Shinomura, T.,
Hamaguchi, M., Yoshida, Y., Ohnuki, Y., Miyauchi, S., Spicer, A.P., McDonald, J.A. and
Kimata, K. (1999) Three isoforms of mammalian hyaluronan synthases have distinct
enzymatic properties. J Biol Chem. 274:25085-25092.
87. Jackson, D.G., Buckley, J. and Bell, J.I. (1992) Multiple variants of the human lymphocyte
homing receptor CD44 generated by insertions at a single site in the extracellular domain. J
Biol Chem. 267:4732-4739.
88. Jakoby, W.B. (1994) Detoxication: Conjugation and Hydrolysis. In "The Liver: Biology and
Pathobiology". Arias, I.M., Boyer, J.L., Fausto, N., Jakoby, W.B., Schachter, D.A. and
Shafritz, D.A., editors. Raven Press Ltd, New York. 429-440.
89. Jalkanen, S. and Jalkanen, M. (1992) Lymphocyte CD44 binds the COOH-terminal heparin-
binding domain of fibronectin. J Cell Biol. 116:817-825.

53
90. Jenniskens, G.J., Oosterhof, A., Brandwijk, R., Veerkamp, J.H. and van Kuppevelt, T.H.
(2000) Heparan sulfate heterogeneity in skeletal muscle basal lamina: demonstration by phage
display-derived antibodies. J Neurosci. 20:4099-4111.
91. Kalomiris, E.L. and Bourguignon, L.Y. (1988) Mouse T lymphoma cells contain a
transmembrane glycoprotein (GP85) that binds ankyrin. J Cell Biol. 106:319-327.
92. Katoh, S., Zheng, Z., Oritani, K., Shimozato, T. and Kincade, P.W. (1995) Glycosylation of
CD44 negatively regulates its recognition of hyaluronan. J Exp Med. 182:419-429.
93. Kearns, A.E., Vertel, B.M. and Schwartz, N.B. (1993) Topography of glycosylation and
UDP-xylose production. J Biol Chem. 268:11097-11104.
94. Kimura, J.H., Hardingham, T.E., Hascall, V.C. and Solursh, M. (1979) Biosynthesis of
proteoglycans and their assembly into aggregates in cultures of chondrocytes from the Swarm
rat chondrosarcoma. J Biol Chem. 254:2600-2609.
95. Kincade, P.W., Zheng, Z., Katoh, S. and Hanson, L. (1997) The importance of cellular
environment to function of the CD44 matrix receptor. Curr Opin Cell Biol. 9:635-642.
96. Kjellen, L. and Lindahl, U. (1991) Proteoglycans: structures and interactions. Annu Rev
Biochem. 60:443-475.
97. Knudson, C.B. and Knudson, W. (1993) Hyaluronan-binding proteins in development, tissue
homeostasis, and disease. Faseb J. 7:1233-1241.
98. Knudson, W., Bartnik, E. and Knudson, C.B. (1993) Assembly of pericellular matrices by
COS-7 cells transfected with CD44 lymphocyte-homing receptor genes. Proc Natl Acad Sci
U S A. 90:4003-4007.
99. Knudson, W. and Knudson, C.B. (1991) Assembly of a chondrocyte-like pericellular matrix
on non-chondrogenic cells. Role of the cell surface hyaluronan receptors in the assembly of a
pericellular matrix. J Cell Sci. 99:227-235.
100. Kohda, D., Morton, C.J., Parkar, A.A., Hatanaka, H., Inagaki, F.M., Campbell, I.D. and Day,
A.J. (1996) Solution structure of the link module: a hyaluronan-binding domain involved in
extracellular matrix stability and cell migration. Cell. 86:767-775.
101. Kok, K., Osinga, J., Carritt, B., Davis, M.B., van der Hout, A.H., van der Veen, A.Y.,
Landsvater, R.M., de Leij, L.F., Berendsen, H.H., Postmus, P.E. and et al. (1987) Deletion of
a DNA sequence at the chromosomal region 3p21 in all major types of lung cancer. Nature.
330:578-581.
102. Kreil, G. (1995) Hyaluronidases--a group of neglected enzymes. Protein Sci. 4:1666-1669.
103. Landolt, R.M., Vaughan, L., Winterhalter, K.H. and Zimmermann, D.R. (1995) Versican is
selectively expressed in embryonic tissues that act as barriers to neural crest cell migration
and axon outgrowth. Development. 121:2303-2312.

54
104. Landstein, D., Graves, M.V., Burbank, D.E., DeAngelis, P. and Van Etten, J.L. (1998)
Chlorella virus PBCV-1 encodes functional glutamine: fructose-6-phosphate
amidotransferase and UDP-glucose dehydrogenase enzymes. Virology. 250:388-396.
105. Lauffenburger, D.A. and Horwitz, A.F. (1996) Cell migration: a physically integrated
molecular process. Cell. 84:359-369.
106. Laurent, T.C. and Fraser, J.R. (1992) Hyaluronan. Faseb J. 6:2397-2404.
107. Laurent, T.C., Fraser, J.R., Pertoft, H. and Smedsrod, B. (1986) Binding of hyaluronate and
chondroitin sulphate to liver endothelial cells. Biochem J. 234:653-658.
108. Laurent, T.C. and Fraser, J.R.E. (1991) Catabolism of Hyaluronan. In "Degradation of
Bioactive Substances: Physiology and Pathology". Henriksen, J.H., editor. CRC Press, Boca
Raton. 249-265.
109. LeBaron, R.G., Zimmermann, D.R. and Ruoslahti, E. (1992) Hyaluronate binding properties
of versican. J Biol Chem. 267:10003-10010.
110. Lepperdinger, G., Strobl, B. and Kreil, G. (1998) HYAL2, a human gene expressed in many
cells, encodes a lysosomal hyaluronidase with a novel type of specificity. J Biol Chem.
273:22466-22470.
111. Lesley, J., He, Q., Miyake, K., Hamann, A., Hyman, R. and Kincade, P.W. (1992)
Requirements for hyaluronic acid binding by CD44: a role for the cytoplasmic domain and
activation by antibody. J Exp Med. 175:257-266.
112. Lesley, J. and Hyman, R. (1998) CD44 structure and function [In Process Citation]. Front
Biosci. 3:D616-630.
113. Lesley, J., Hyman, R. and Kincade, P.W. (1993) CD44 and its interaction with extracellular
matrix. Adv Immunol. 54:271-335.
114. Lin, Y., Mahan, K., Lathrop, W.F., Myles, D.G. and Primakoff, P. (1994) A hyaluronidase
activity of the sperm plasma membrane protein PH-20 enables sperm to penetrate the cumulus
cell layer surrounding the egg. J Cell Biol. 125:1157-1163.
115. Lindahl, U., Kusche-Gullberg, M. and Kjellen, L. (1998) Regulated diversity of heparan
sulfate. J Biol Chem. 273:24979-24982.
116. Liu, D. and Sy, M.S. (1997) Phorbol myristate acetate stimulates the dimerization of CD44
involving a cysteine in the transmembrane domain. J Immunol. 159:2702-2711.
117. Lloyd, J.B. (1996) Metabolite efflux and influx across the lysosome membrane. Subcell
Biochem. 27:361-386.
118. Maiti, A., Maki, G. and Johnson, P. (1998) TNF-alpha induction of CD44-mediated
leukocyte adhesion by sulfation. Science. 282:941-943.

55
119. Markovitz, A., Cifonelli, J.A. and Dorfman, A. (1959) The biosynthesis of HA by group A
Streptococcus. VI. Biosynthesis from uridine nucleotides in cell-free extracts. J Biol Chem.
234:2343-2350.
120. McCourt, P.A., Smedsrod, B.H., Melkko, J. and Johansson, S. (1999) Characterization of a
hyaluronan receptor on rat sinusoidal liver endothelial cells and its functional relationship to
scavenger receptors. Hepatology. 30:1276-1286.
121. McKee, C.M., Lowenstein, C.J., Horton, M.R., Wu, J., Bao, C., Chin, B.Y., Choi, A.M. and
Noble, P.W. (1997) Hyaluronan fragments induce nitric-oxide synthase in murine
macrophages through a nuclear factor kappaB-dependent mechanism. J Biol Chem.
272:8013-8018.
122. Meyer, F.A. (1983) Macromolecular basis of globular protein exclusion and of swelling
pressure in loose connective tissue (umbilical cord). Biochim Biophys Acta. 755:388-399.
123. Meyer, K. and Palmer, J.W. (1934) The polysaccharide of the vitreous humor. J Biol Chem.
107:629-634.
124. Meyer, M.F. and Kreil, G. (1996) Cells expressing the DG42 gene from early Xenopus
embryos synthesize hyaluronan. Proc Natl Acad Sci U S A. 93:4543-4547.
125. Mian, N. (1986) Characterization of a high-Mr plasma-membrane-bound protein and
assessment of its role as a constituent of hyaluronate synthase complex. Biochem J. 237:343-
357.
126. Milev, P., Maurel, P., Haring, M., Margolis, R.K. and Margolis, R.U. (1996) TAG-1/axonin-1
is a high-affinity ligand of neurocan, phosphacan/protein-tyrosine phosphatase-zeta/beta, and
N-CAM. J Biol Chem. 271:15716-15723.
127. Milla, M.E., Clairmont, C.A. and Hirschberg, C.B. (1992) Reconstitution into
proteoliposomes and partial purification of the Golgi apparatus membrane UDP-galactose,
UDP-xylose, and UDP-glucuronic acid transport activities. J Biol Chem. 267:103-107.
128. Milla, M.E. and Hirschberg, C.B. (1989) Reconstitution of Golgi vesicle CMP-sialic acid and
adenosine 3'-phosphate 5'-phosphosulfate transport into proteoliposomes. Proc Natl Acad Sci
U S A. 86:1786-1790.
129. Moll, J., Khaldoyanidi, S., Sleeman, J.P., Achtnich, M., Preuss, I., Ponta, H. and Herrlich, P.
(1998) Two different functions for CD44 proteins in human myelopoiesis. J Clin Invest.
102:1024-1034.
130. Neame, S.J. and Isacke, C.M. (1992) Phosphorylation of CD44 in vivo requires both Ser323
and Ser325, but does not regulate membrane localization or cytoskeletal interaction in
epithelial cells. Embo J. 11:4733-4738.

56
131. Neame, S.J. and Isacke, C.M. (1993) The cytoplasmic tail of CD44 is required for basolateral
localization in epithelial MDCK cells but does not mediate association with the detergent-
insoluble cytoskeleton of fibroblasts. J Cell Biol. 121:1299-1310.
132. Neufeld, E. and Hall, C. (1965) Inhibition of UDP-D-glucos dehydrogenase by UDP-D-
xylose. Biochem Biophys Res Commun. 19:456-461.
133. Ng, K.F. and Schwartz, N.B. (1989) Solubilization and partial purification of hyaluronate
synthetase from oligodendroglioma cells. J Biol Chem. 264:11776-11783.
134. Ohashi, Y., Honda, A., Iwai, T. and Mori, Y. (1988) Stimulatory effect of vanadate on
hyaluronic acid synthesis in mesothelial cells from rabbit pericardium. Biochem Int. 16:293-
302.
135. Orkin, R.W. and Toole, B.P. (1978) Hyaluronidase activity and hyaluronate content of the
developing chick embryo heart. Dev Biol. 66:308-320.
136. Parkar, A.A. and Day, A.J. (1997) Overlapping sites on the Link module of human TSG-6
mediate binding to hyaluronan and chrondroitin-4-sulphate. FEBS Lett. 410:413-417.
137. Peach, R.J., Hollenbaugh, D., Stamenkovic, I. and Aruffo, A. (1993) Identification of
hyaluronic acid binding sites in the extracellular domain of CD44. J Cell Biol. 122:257-264.
138. Peck, D. and Isacke, C.M. (1996) CD44 phosphorylation regulates melanoma cell and
fibroblast migration on, but not attachment to, a hyaluronan substratum. Curr Biol. 6:884-890.
139. Perozich, J., Leksana, A. and Hempel, J. (1995) UDP-glucose dehydrogenase. Structural
characteristics. Adv Exp Med Biol. 372:79-84.
140. Perris, R. and Johansson, S. (1990) Inhibition of neural crest cell migration by aggregating
chondroitin sulfate proteoglycans is mediated by their hyaluronan-binding region. Dev Biol.
137:1-12.
141. Perris, R., Perissinotto, D., Pettway, Z., Bronner-Fraser, M., Morgelin, M. and Kimata, K.
(1996) Inhibitory effects of PG-H/aggrecan and PG-M/versican on avian neural crest cell
migration. Faseb J. 10:293-301.
142. Philipson, L.H. and Schwartz, N.B. (1984) Subcellular localization of hyaluronate synthetase
in oligodendroglioma cells. J Biol Chem. 259:5017-5023.
143. Piek, E., Heldin, C.H. and Ten Dijke, P. (1999) Specificity, diversity, and regulation in TGF-
beta superfamily signaling. Faseb J. 13:2105-2124.
144. Prehm, P. (1983) Synthesis of hyaluronate in differentiated teratocarcinoma cells.
Characterization of the synthase. Biochem J. 211:181-189.
145. Prehm, P. (1983) Synthesis of hyaluronate in differentiated teratocarcinoma cells. Mechanism
of chain growth. Biochem J. 211:191-198.

57
146. Prehm, P. (1989) Identification and regulation of the eukaryotic hyaluronate synthase. In
"The Biology of Hyaluronan". Vol. 143. Evered, D. and Whelan, J., editors. Wiley,
Chichester. 21-31.
147. Pummill, P.E., Achyuthan, A.M. and DeAngelis, P.L. (1998) Enzymological characterization
of recombinant xenopus DG42, a vertebrate hyaluronan synthase. J Biol Chem. 273:4976-
4981.
148. Rahmanian, M., Pertoft, H., Kanda, S., Christofferson, R., Claesson-Welsh, L. and Heldin, P.
(1997) Hyaluronan oligosaccharides induce tube formation of a brain endothelial cell line in
vitro. Exp Cell Res. 237:223-230.
149. Rapraeger, A.C., Krufka, A. and Olwin, B.B. (1991) Requirement of heparan sulfate for
bFGF-mediated fibroblast growth and myoblast differentiation. Science. 252:1705-1708.
150. Rauch, U., Karthikeyan, L., Maurel, P., Margolis, R.U. and Margolis, R.K. (1992) Cloning
and primary structure of neurocan, a developmentally regulated, aggregating chondroitin
sulfate proteoglycan of brain. J Biol Chem. 267:19536-19547.
151. Reed, R.K., Laurent, U.B., Fraser, J.R. and Laurent, T.C. (1990) Removal rate of
[3H]hyaluronan injected subcutaneously in rabbits. Am J Physiol. 259:H532-535.
152. Reed, R.K., Lilja, K. and Laurent, T.C. (1988) Hyaluronan in the rat with special reference to
the skin. Acta Physiol Scand. 134:405-411.
153. Ripellino, J.A., Bailo, M., Margolis, R.U. and Margolis, R.K. (1988) Light and electron
microscopic studies on the localization of hyaluronic acid in developing rat cerebellum. J Cell
Biol. 106:845-855.
154. Robertson, D., Beech, I. and Bolwell, G.P. (1995) Regulation of the Enzymes of UDP-sugar
Metabolism during Differentiation of French Bean. Phytochemistry. 39:21-28.
155. Robyt, J.F., Kimble, B.K. and Walseth, T.F. (1974) The mechanism of dextransucrase action.
Direction of dextran biosynthesis. Arch Biochem Biophys. 165:634-640.
156. Roden, L., Koerner, T., Olson, C. and Schwartz, N.B. (1985) Mechanisms of chain initiation
in the biosynthesis of connective tissue polysaccharides. Fed Proc. 44:373-380.
157. Rome, L.H. and Hill, D.F. (1986) Lysosomal degradation of glycoproteins and
glycosaminoglycans. Efflux and recycling of sulphate and N-acetylhexosamines. Biochem J.
235:707-713.
158. Rooney, P., Kumar, S., Ponting, J. and Wang, M. (1995) The role of hyaluronan in tumour
neovascularization (review). Int J Cancer. 60:632-636.
159. Ruoslahti, E. and Yamaguchi, Y. (1991) Proteoglycans as modulators of growth factor
activities. Cell. 64:867-869.
160. Sainio, M., Zhao, F., Heiska, L., Turunen, O., den Bakker, M., Zwarthoff, E., Lutchman,
M., Rouleau, G.A., Jaaskelainen, J., Vaheri, A. and Carpen, O. (1997) Neurofibromatosis

58
2 tumor suppressor protein colocalizes with ezrin and CD44 and associates with actin-
containing cytoskeleton. J Cell Sci. 110:2249-2260.
161. Salmivirta, M., Lidholt, K. and Lindahl, U. (1996) Heparan sulfate: a piece of information.
Faseb J. 10:1270-1279.
162. Salustri, A., Yanagishita, M. and Hascall, V.C. (1990) Mouse oocytes regulate hyaluronic acid
synthesis and mucification by FSH-stimulated cumulus cells. Dev Biol. 138:26-32.
163. Salustri, A., Yanagishita, M., Underhill, C.B., Laurent, T.C. and Hascall, V.C. (1992)
Localization and synthesis of hyaluronic acid in the cumulus cells and mural granulosa cells
of the preovulatory follicle. Dev Biol. 151:541-551.
164. Sargent, T.D. and Dawid, I.B. (1983) Differential gene expression in the gastrula of Xenopus
laevis. Science. 222:135-139.
165. Scott, J.E. (1989) Secondary structures in hyaluronan solutions: chemical and biological
implications. In "The Biology of Hyaluronan". Vol. 143. Evered, D. and Whelan, J., editors.
Wiley, Chichester. 6-20.
166. Scott, J.E., Cummings, C., Brass, A. and Chen, Y. (1991) Secondary and tertiary structures of
hyaluronan in aqueous solution, investigated by rotary shadowing-electron microscopy and
computer simulation. Hyaluronan is a very efficient network-forming polymer. Biochem J.
274:699-705.
167. Scott, J.E. and Heatley, F. (1999) Hyaluronan forms specific stable tertiary structures in
aqueous solution: a 13C NMR study. Proc Natl Acad Sci U S A. 96:4850-4855.
168. Seidenbecher, C.I., Richter, K., Rauch, U., Fassler, R., Garner, C.C. and Gundelfinger, E.D.
(1995) Brevican, a chondroitin sulfate proteoglycan of rat brain, occurs as secreted and cell
surface glycosylphosphatidylinositol-anchored isoforms. J Biol Chem. 270:27206-27212.
169. Semino, C.E., Specht, C.A., Raimondi, A. and Robbins, P.W. (1996) Homologs of the
Xenopus developmental gene DG42 are present in zebrafish and mouse and are involved in
the synthesis of Nod-like chitin oligosaccharides during early embryogenesis. Proc Natl Acad
Sci U S A. 93:4548-4553.
170. Sheetz, M.P., Felsenfeld, D.P. and Galbraith, C.G. (1998) Cell migration: regulation of force
on extracellular-matrix-integrin complexes. Trends Cell Biol. 8:51-54.
171. Sherman, L., Sleeman, J., Herrlich, P. and Ponta, H. (1994) Hyaluronate receptors: key
players in growth, differentiation, migration and tumor progression. Curr. Opin. Cell Biol.
6:726-733.
172. Shyjan, A.M., Heldin, P., Butcher, E.C., Yoshino, T. and Briskin, M.J. (1996) Functional
cloning of the cDNA for a human hyaluronan synthase. J Biol Chem. 271:23395-23399.

59
173. Sleeman, J., Rudy, W., Hofmann, M., Moll, J., Herrlich, P. and Ponta, H. (1996) Regulated
clustering of variant CD44 proteins increases their hyaluronate binding capacity. J Cell Biol.
135:1139-1150.
174. Sleeman, J.P., Kondo, K., Moll, J., Ponta, H. and Herrlich, P. (1997) Variant exons v6 and v7
together expand the repertoire of glycosaminoglycans bound by CD44. J Biol Chem.
272:31837-31844.
175. Smaglik, P. and Butler, D. (2000) Celera turns to public genome data to speed up endgame.
Nature. 403:119.
176. Sommarin, Y. and Heinegard, D. (1983) Specific interaction between cartilage proteoglycans
and hyaluronic acid at the chondrocyte cell surface. Biochem J. 214:777-784.
177. Spicer, A.P., Augustine, M.L. and McDonald, J.A. (1996) Molecular cloning and
characterization of a putative mouse hyaluronan synthase. J Biol Chem. 271:23400-23406.
178. Spicer, A.P., Camenisch, T., Nguyen, T., Haudenschild, D. and McDonald, J. (1999)
Hyaluronan biosynthesis and function in vertebrates. Conference abstract in "Glyco XV",
Tokyo, Japan.
179. Spicer, A.P., Kaback, L.A., Smith, T.J. and Seldin, M.F. (1998) Molecular cloning and
characterization of the human and mouse UDP- glucose dehydrogenase genes. J Biol Chem.
273:25117-25124.
180. Spicer, A.P. and McDonald, J.A. (1998) Characterization and molecular evolution of a
vertebrate hyaluronan synthase gene family. J Biol Chem. 273:1923-1932.
181. Spicer, A.P. and Nguyen, T.K. (1999) Mammalian hyaluronan synthases: investigation of
functional relationships in vivo. Biochem Soc Trans. 27:109-115.
182. Spicer, A.P., Olson, J.S. and McDonald, J.A. (1997) Molecular cloning and characterization
of a cDNA encoding the third putative mammalian hyaluronan synthase. J Biol Chem.
272:8957-8961.
183. Spicer, A.P., Seldin, M.F., Olsen, A.S., Brown, N., Wells, D.E., Doggett, N.A., Itano, N.,
Kimata, K., Inazawa, J. and McDonald, J.A. (1997) Chromosomal localization of the human
and mouse hyaluronan synthase genes. Genomics. 41:493-497.
184. Stern, R. and Csoka, A. (2000) Mammalian Hyaluronidases. Glycoforum Website "Science
of Hyaluronan Today" <http://www.glycoforum.gr.jp>.
185. Stoolmiller, A.C. and Dorfman, A. (1969) The biosynthesis of hyaluronic acid by
Streptococcus. J Biol Chem. 244:236-246.
186. Suzuki, M., Asplund, T., Yamashita, H., Heldin, C.-H. and Heldin, P. (1995) Stimulation of
hyaluronan biosynthesis by platelet-derived growth factor-BB and transforming growth
factor-β1 involves activation of protein kinase C. Biochem J. 307:817-821.

60
187. Sweeney, C., Mackintosh, D. and Mason, R.M. (1993) UDP-sugar metabolism in Swarm rat
chondrosarcoma chondrocytes. Biochem J. 290:563-570.
188. Takahashi, K., Stamenkovic, I., Cutler, M., Dasgupta, A. and Tanabe, K.K. (1996) Keratan
sulfate modification of CD44 modulates adhesion to hyaluronate. J Biol Chem. 271:9490-
9496.
189. Tammi, R., MacCallum, D., Hascall, V.C., Pienimaki, J.P., Hyttinen, M. and Tammi, M.
(1998) Hyaluronan bound to CD44 on keratinocytes is displaced by hyaluronan
decasaccharides and not hexasaccharides. J Biol Chem. 273:28878-28888.
190. Teder, P., Nettelbladt, O. and Heldin, P. (1995) Characterization of the mechanism involved in
bleomycin-induced increased hyaluronan production in rat lung. Am J Respir Cell Mol Biol.
12:181-189.
191. Tengblad, A., Laurent, U.B., Lilja, K., Cahill, R.N., Engstrom-Laurent, A., Fraser, J.R.,
Hansson, H.E. and Laurent, T.C. (1986) Concentration and relative molecular mass of
hyaluronate in lymph and blood. Biochem J. 236:521-525.
192. Tenhaken, K. and Thulke, O. (1996) Cloning of an enzyme that synthesize a key nucleotide-
sugar precursor of hemicellulose biosynthesis from soybean: UDP-glucose dehydrogenase.
Plant Phys. 112:1127-1134.
193. Thiebot, B., Bichoualne, L., Langris, M., Bonnamy, P.J., Barbey, P., Carreau, S. and Bocquet,
J. (1997) IGF-1 stimulates synthesis of undersulfated proteoglycans and of hyaluronic acid
by peritubular cells from immature rat testis. Biochim Biophys Acta. 1358:127-141.
194. Thomas, L., Byers, H.R., Vink, J. and Stamenkovic, I. (1992) CD44H regulates tumor cell
migration on hyaluronate-coated substrate. J Cell Biol. 118:971-977.
195. Tiedemann, K., Malmstrom, A. and Westergren-Thorsson, G. (1997) Cytokine regulation of
proteoglycan production in fibroblasts: separate and synergistic effects. Matrix Biol. 15:469-
478.
196. Tirone, E., D'Alessandris, C., Hascall, V.C., Siracusa, G. and Salustri, A. (1997) Hyaluronan
synthesis by mouse cumulus cells is regulated by interactions between follicle-stimulating
hormone (or epidermal growth factor) and a soluble oocyte factor (or transforming growth
factor beta1). J Biol Chem. 272:4787-4794.
197. Tlapak-Simmons, V., Baggenstoss, B.A., Clyne, T. and Weigel, P. (1999) Purification and
lipid dependence of the recombinant hyaluronan synthases from Streptococcus pyogenes and
streptococcus equisimilis. J Biol Chem. 274:4239-4245.
198. Tlapak-Simmons, V.L., Baggenstoss, B.A., Kumari, K., Heldermon, C. and Weigel, P.H.
(1999) Kinetic characterization of the recombinant hyaluronan synthases from Streptococcus
pyogenes and Streptococcus equisimilis. J Biol Chem. 274:4246-4253.

61
199. Tlapak-Simmons, V.L., Kempner, E.S., Baggenstoss, B.A. and Weigel, P.H. (1998) The
active streptococcal hyaluronan synthases (HASs) contain a single HAS monomer and
multiple cardiolipin molecules. J Biol Chem. 273:26100-26109.
200. Tolg, C., Hofmann, M., Herrlich, P. and Ponta, H. (1993) Splicing choice from ten variant
exons establishes CD44 variability. Nucleic Acids Res. 21:1225-1229.
201. Toma, L., Pinhal, M.A., Dietrich, C.P., Nader, H.B. and Hirschberg, C.B. (1996) Transport of
UDP-galactose into the Golgi lumen regulates the biosynthesis of proteoglycans. J Biol
Chem. 271:3897-3901.
202. Toole, B.P. (1982) Developmental role of hyaluronate. Connect Tissue Res. 10:93-100.
203. Toole, B.P., Biswas, C. and Gross, J. (1979) Hyaluronate and invasiveness of the rabbit V2
carcinoma. Proc Natl Acad Sci U S A. 76:6299-6303.
204. Toole, B.P. and Trelstad, R.L. (1971) Hyaluronate production and removal during corneal
development in the chick. Dev Biol. 26:28-35.
205. Tsifrina, E., Ananyeva, N.M., Hastings, G. and Liau, G. (1999) Identification and
characterization of three cDNAs that encode putative novel hyaluronan-binding proteins,
including an endothelial cell-specific hyaluronan receptor. Am J Pathol. 155:1625-1633.
206. Tsukita, S., Oishi, K., Sato, N., Sagara, J. and Kawai, A. (1994) ERM family members as
molecular linkers between the cell surface glycoprotein CD44 and actin-based cytoskeletons.
J Cell Biol. 126:391-401.
207. Turley, E.A., Austen, L., Vandeligt, K. and Clary, C. (1991) Hyaluronan and a cell-associated
hyaluronan binding protein regulate the locomotion of ras-transformed cells. J Cell Biol.
112:1041-1047.
208. Uff, C.R., Neame, S.J. and Isacke, C.M. (1995) Hyaluronan binding by CD44 is regulated by
a phosphorylation-independent mechanism. Eur J Immunol. 25:1883-1887.
209. Ujita, M., Shinomura, T., Ito, K., Kitagawa, Y. and Kimata, K. (1994) Expression and binding
activity of the carboxyl-terminal portion of the core protein of PG-M, a large chondroitin
sulfate proteoglycan. J Biol Chem. 269:27603-27609.
210. Underhill, C.B., Chi-Rosso, G. and Toole, B.P. (1983) Effects of detergent solubilization on
the hyaluronate-binding protein from membranes of simian virus 40-transformed 3T3 cells. J
Biol Chem. 258:8086-8091.
211. van de Rijn, I. and Drake, R.R. (1992) Analysis of the streptococcal hyaluronic acid synthase
complex using the photoaffinity probe 5-azido-UDP-glucuronic acid. J Biol Chem.
267:24302-24306.
212. van Kuppevelt, T.H., Dennissen, M.A., van Venrooij, W.J., Hoet, R.M. and Veerkamp, J.H.
(1998) Generation and application of type-specific anti-heparan sulfate antibodies using

62
phage display technology. Further evidence for heparan sulfate heterogeneity in the kidney. J
Biol Chem. 273:12960-12966.
213. Varki, A. (1996) Does DG42 synthesize hyaluronan or chitin?: A controversy about
oligosaccharides in vertebrate development. Proc Natl Acad Sci U S A. 93:4523-4525.
214. Varki, A. (1998) Factors controlling the glycosylation potential of the Golgi apparatus.
Trends Cell Biol. 8:34-40.
215. Vertel, B.M., Walters, L.M., Flay, N., Kearns, A.E. and Schwartz, N.B. (1993) Xylosylation is
an endoplasmic reticulum to Golgi event. J Biol Chem. 268:11105-11112.
216. von Andrian, U.H., Hasslen, S.R., Nelson, R.D., Erlandsen, S.L. and Butcher, E.C. (1995) A
central role for microvillous receptor presentation in leukocyte adhesion under flow. Cell.
82:989-999.
217. Wang, C., Entwistle, J., Hou, G., Li, Q. and Turley, E.A. (1996) The characterization of a
human RHAMM cDNA: conservation of the hyaluronan-binding domains. Gene. 174:299-
306.
218. Watanabe, E., Aono, S., Matsui, F., Yamada, Y., Naruse, I. and Oohira, A. (1995) Distribution
of a brain-specific proteoglycan, neurocan, and the corresponding mRNA during the
formation of barrels in the rat somatosensory cortex. Eur J Neurosci. 7:547-554.
219. Watanabe, K. and Yamaguchi, Y. (1996) Molecular identification of a putative human
hyaluronan synthase. J Biol Chem. 271:22945-22948.
220. Weigel, P.H., Hascall, V.C. and Tammi, M. (1997) Hyaluronan synthases. J Biol Chem.
272:13997-14000.
221. West, D.C., Hampson, I.N., Arnold, F. and Kumar, S. (1985) Angiogenesis induced by
degradation products of hyaluronic acid. Science. 228:1324-1326.
222. Yamada, H., Fredette, B., Shitara, K., Hagihara, K., Miura, R., Ranscht, B., Stallcup, W.B. and
Yamaguchi, Y. (1997) The brain chondroitin sulfate proteoglycan brevican associates with
astrocytes ensheathing cerebellar glomeruli and inhibits neurite outgrowth from granule
neurons. J Neurosci. 17:7784-7795.
223. Yamada, Y., Itano, N., Zako, M., Yoshida, M., Lenas, P., Niimi, A., Ueda, M. and Kimata, K.
(1998) The gene structure and promoter sequence of mouse hyaluronan synthase 1. Biochem
J. 330:1223-1227.
224. Yamaguchi, Y. (1996) Brevican: a major proteoglycan in adult brain. Perspect Dev Neurobiol.
3:307-317.
225. Yang, B., Yang, B.L., Savani, R.C. and Turley, E.A. (1994) Identification of a common
hyaluronan binding motif in the hyaluronan binding proteins RHAMM, CD44 and link
protein. Embo J. 13:286-296.

63
226. Yang, B., Zhang, L. and Turley, E.A. (1993) Identification of two hyaluronan-binding
domains in the hyaluronan receptor RHAMM. J Biol Chem. 268:8617-8623.
227. Yannariello-Brown, J., Frost, S.J. and Weigel, P.H. (1992) Identification of the Ca(2+)-
independent endocytic hyaluronan receptor in rat liver sinusoidal endothelial cells using a
photoaffinity cross-linking reagent. J Biol Chem. 267:20451-20456.
228. Yayon, A., Klagsbrun, M., Esko, J.D., Leder, P. and Ornitz, D.M. (1991) Cell surface,
heparin-like molecules are required for binding of basic fibroblast growth factor to its high
affinity receptor. Cell. 64:841-848.
229. Yoshida, M., Itano, N., Yamada, Y. and Kimata, K. (2000) In vitro synthesis of hyaluronan by
a single protein derived from mouse HAS1 gene and characterization of amino acid residues
essential for the activity. J Biol Chem. 275:497-506.
230. Ysart, G.E. and Mason, R.M. (1994) Serum factors, growth factors and UDP-sugar
metabolism in bovine articular cartilage chondrocytes. Biochem J. 303:713-721.
231. Zhang, L., David, G. and Esko, J.D. (1995) Repetitive Ser-Gly sequences enhance heparan
sulfate assembly in proteoglycans. J Biol Chem. 270:27127-27135.
232. Zhang, L. and Esko, J.D. (1994) Amino acid determinants that drive heparan sulfate assembly
in a proteoglycan. J Biol Chem. 269:19295-19299.
233. Zhang, S., Chang, M.C., Zylka, D., Turley, S., Harrison, R. and Turley, E.A. (1998) The
hyaluronan receptor RHAMM regulates extracellular-regulated kinase. J Biol Chem.
273:11342-11348.
234. Zhao, M., Yoneda, M., Ohashi, Y., Kurono, S., Iwata, H., Ohnuki, Y. and Kimata, K. (1995)
Evidence for the covalent binding of SHAP, heavy chains of inter-alpha-trypsin inhibitor, to
hyaluronan. J Biol Chem. 270:26657-26663.
235. Zheng, Z., Cummings, R.D., Pummill, P.E. and Kincade, P.W. (1997) Growth as a solid
tumor or reduced glucose concentrations in culture reversibly induce CD44-mediated
hyaluronan recognition by Chinese hamster ovary cells. J Clin Invest. 100:1217-1229.
236. Zhou, B., Oka, J.A., Singh, A. and Weigel, P.H. (1999) Purification and subunit
characterization of the rat liver endocytic hyaluronan receptor. J Biol Chem. 274:33831-
33834.
237. Zoller, M. (1995) CD44: physiological expression of distinct isoforms as evidence for organ-
specific metastasis formation. J Mol Med. 73:425-438.







