
Indian Journal of Experimental Biology
Vol. 49, June 2011, pp. 446-454
Sub-chronic diclofenac sodium induced alterations of alkaline phosphatase
activity in serum and skeletal muscle of mice
Shalini Chouhan & Sushma Sharma*
Department of Biosciences, H P University, Summer Hill, Shimla 171 005, India
Received 29 June 2010; revised 1 March 2011
The present study has been carried on changes in activity of alkaline phosphatase in serum and gastrocnemius muscle
of mice after sub-chronic use of diclofenac. Mice in experimental group received diclofenac (10 mg/kg body wt /day) for 30
days while control group received normal saline. Alkaline phosphatase was assayed in muscle and serum and its activity was
localized histochemically in muscle. Results showed that diclofenac induced changes in specific activity of alkaline
phosphatase at different periods of treatment variably compared to control group. Specific activity of alkaline phosphatase
decreased significantly in gastrocnemius initially (48.74%), increased thereafter (132.96%) and slight decrease (13.97%)
was noticed after 30 days. In serum, the specific activity of alkaline phosphatase decreased slightly after 10 days (18.78%),
increased in the middle of the treatment period (132.04%) as well as showed increase (109.09%) compared to control group
after 30 days stage of investigation. These findings were also confirmed by electrophoretic studies in muscle.
Keywords: Alkaline phosphatase, Diclofenac, Gastrocnemius, Serum, Sub-chronic
Diclofenac residues were found to be the cause for
rapid population decline of Gyps vultures in southern
Asian countries1. The toxic effects of diclofenac were
known however the mechanism of action is still not
fully understood. Nonsteroidal anti-inflammatory
drugs (NSAIDs) continue to be prescribed
as
analgesics for patients with healing fractures even
though these drugs diminish bone formation, healing,
and remodeling2. Diclofenac is a widely used NSAID
for treatment of a variety of inflammatory conditions
such as rheumatoid arthritis, osteoarthritis and
acute muscle aches. Diclofenac inhibits both
cyclooxygenase (COX) isoenzymes, COX-1 and
COX-2, by blocking arachidonate binding resulting in
analgesic, antipyretic and anti-inflammatory
pharmacologic effects3. The enzymes COX-1 and
COX-2 catalyze the conversion of arachidonic acid to
prostaglandin G2 (PGG2), the first step of the
synthesis of prostaglandins and thromboxanes that are
involved in rapid physiological responses. Delayed
fracture healing and no body weight gain was
reported in rats after long term treatment with
diclofenac4. Diclofenac sodium causes a rare but
potentially fatal hepatotoxicity that may be associated
with the formation of reactive metabolites and
subsequent adverse hepatitis effects may arise in
certain individuals5. NS-398, a COX-2 specific
inhibitor has been reported to interfere with the
healing of injured skeletal muscle6. An inhibition
of protein synthesis in rat skeletal muscle with
three different cyclooxygenase inhibitors (aspirin,
indomethacin and meclofenamate) was also reported7.
The elevation in the serum level of alkaline
phosphatase has been used as diagnostic index for
skeletal muscle disorders8. Alkaline phosphatase
showed an association with a number of muscle
diseases including neuromuscular disorders,
myopatheis or other wasting conditions9. The
objective of the study was to investigate diclofenac
induced alterations of alkaline phosphatase activity in
mice skeletal muscle.
Alkaline phosphatase (APase; E.C.3.1.3.1)
represents a group of isozymes that are membrane-
bound glycoproteins which catalyze the hydrolysis of
inorganic and organic monophosphate esters at
alkaline pH in vitro10
. APase is ubiquitous in nature
located in basal membrane of various tissues and also
found in the serum11
. Soluble forms of APase exist in
the serum. Alkaline phosphatases are members of a
rather diverse group of membrane proteins which are
anchored to lipid bilayers in cell membranes by a
phosphatidylinositol-glycan moiety attached to the
——————— *Correspondent author
Telephone: 91-177-2830946
Fax: 91-177-2830495
E-mail: [email protected]; [email protected]

CHOUHAN & SHARMA: DICLOFENAC SODIUM & ALKALINE PHOSPHATASE ACTIVITY
447
carboxy terminus of the protein12
. Serum APase is a
dimer, whereas the membrane-bound forms of APase
are probably tetramers13
. Studies reported earlier
have established that APase does form an important
entity in the muscle cell14
. Patients suffering from
hypophosphatasia, a genetic deficiency in APase
activity, commonly suffer from severe rickets and
osteomalacia15
.
Materials and Methods
Animals―Healthy male mice of Balb-c strain (36)
weighing about 24-27 g were procured from Central
Research Institute (CRI), Kasauli, (Himachal Pradesh),
India. Animals were maintained in hygienic conditions
in a well ventilated room of animal house of Department
of Bio-Sciences of H P University, Shimla with D:L
cycle of (12:12) h and a temperature of 25 ± 2°C.
Animals were provided with commercial feed
(Hindustan Lever Ltd., New Delhi, India) and water ad
libitium. All procedures, including the maintenance of
the animals had the approval of Institutional Animals
Ethics Committee of the University (IAEC approval no:
IAEC/Bio/2009/6-HPU).
Drug administration―Diclofenac sodium was
purchased from Sigma Aldrich Co., USA. Stock
solution was made in distilled water. Further dilutions
were done according to the body weight records of the
animals. All the chemicals used were of analytical
grade. To investigate the changes in the specific
activity of APase in control and experimental group,
mice (18) received diclofenac intramuscularly at
the dose rate of 10 mg/kg body wt. /day for 10, 20 and
30 days while 18 of the control group received normal
saline. The drug dose chosen was higher than the
recommended therapeutic dose (8 mg/kg) for mice.
Mice were sacrificed by cervical dislocation, the next
day at the end of the experiment. Gastrocnemius
muscle was excised and serum was collected at each
stage of the investigation.
Histological study―Bouin fixed gasrocnemii were
embedded in paraffin wax after dehydrating in
ascending grades of alcohol and clearing in xylene.
Further, 7-8 µm thin sections were processed for
Haematoxylin-Eosin staining. Lastly, after clearing in
xylene, sections were mounted in DPX and
photographed.
Histochemical localization―APase was localized
histochemically in cryostat cut sections of the
gastrocnemius muscle by Gomori’s calcium cobalt
sulphide method16-18
with slight modifications.
Briefly, tissues were fixed overnight in neutral
calcium formol and next day, processed for
histochemical staining. The cryostat cut sections were
preincubated in 100 mM Tris-maleate buffer
containing 1% MgCl2 overnight at room temperature.
Thereafter, the preincubated sections were incubated
in the substrate solution (pH 8.9) containing sodium
β-glycerophosphate for half an h at 37ºC and preceded
according to Gomori’s method16
. These were quickly
dehydrated in ascending grades of alcohol and
mounted in Canada balsam and photographed.
Biochemical assay―The muscle homogenate was
prepared in ice cold distilled water which was
centrifuged at 1664 × g for 20 minutes at 4°C. The
supernatant was used for the enzyme assay. Serum was
collected and centrifuged at 2599 × g for 15 min at 4°C.
Muscle and serum APase was assayed by the method of
Weil and Russel19
. Breifly, sodium β-glycerophosphate
was used as substrate. The homogenates were incubated
with the substrate solution (pH 8.9) for one h at 37°C.
The reaction was stopped by 10% TCA. Finally, the
solution was centrifuged at 936 × g at 4ºC and the
supernatant was collected for determination of enzyme
activity. The color intensity was read at 650 nm in a
Hitachi 150-ophrey Double Beam Spectrophotometer.
The amount of phosphate released was plotted against
known concentrations of KH2PO4. Protein content was
estimated according to method of Lowry et al.20
. The
specific activity of alkaline phosphatase was calculated.
Homogenization of the tissue―Muscle homogenate
was prepared according to the method previously
described21-22
with slight modifications. Tissue was
homogenized in 5 volumes of Tris buffer, 10 mM, pH
7.4 containing 1% triton X-100, 2 mM MgCl2 and
0.025 mM ZnCl2. Homogenate was centrifuged at
15,000 × g for 20 min. The pellet was rehomogenized,
centrifuged at 15,000 × g for another 20 min.
Supernatants were combined and again centrifuged at
20,000 × g for 30 min. All the steps in this procedure
were carried out at 4°C.
Electrophoresis—It was conducted according to the
method of Epstein et al.23
with some modifications.
Supernatants were used for electrophoretic studies.
Proteins were estimated according to Lowry et al.20
.
The enzyme was resolved on 7% separating and 5%
stacking gel. Equal amount of protein was loaded in
each lane. The enzyme was localized using activity
staining technique earlier described by some
workers24
with certain modifications. Gels were
analyzed densitometrically.

INDIAN J EXP BIOL, JUNE 2011
448
Statistical analyses―Values are represented as
mean ± SEM. Results were analysed using Student’s
‘t’ test.
Results
Toxic effects of diclofenac treatment were
observed in mice gastrocnemius in terms of altered morphological and biochemical levels. The histopathological changes pointed towards massive tissue damage. Diclofenac treatment caused numerous pathological changes during different stages of investigation as compared to the control group.
Lightmicrographs of control mice gastrocnemius depicted normal fascicles surrounded by perimysium encasing group of myofibers having relatively uniform pattern. Individual muscle fibers showed endomysium around them, each one having normal peripheral arrangement of the nuclei (Fig. 1A and
1B). Diclofenac treatment for 10 days on the other hand resulted in structural disorganization in terms of degenerating fibers, atrophied fibers and inflammatory cell infiltration (Fig. 1C). After 20 days of drug treatment, merging of fibers was noticed which had irregular limiting membranes, many
fibrolysing as well as abnormally hypertophied fibers were also observed. Previous peripheral arrangement of the nuclei delineated which occupied central position now; (Fig. 1D). More disorganized structure of the muscle was observed after 30 days of diclofenac treatment. Chains of darkly stained nuclei
were noticed in interfibrillar regions. Muscle fibers lost proper shape and uniformity due to loss of perimysial and endomysial boundaries, many atrophied fibers were noticed to be without nuclei, degeneration continued as well as merged fibers were also noticed (Fig. 1E and 1F).
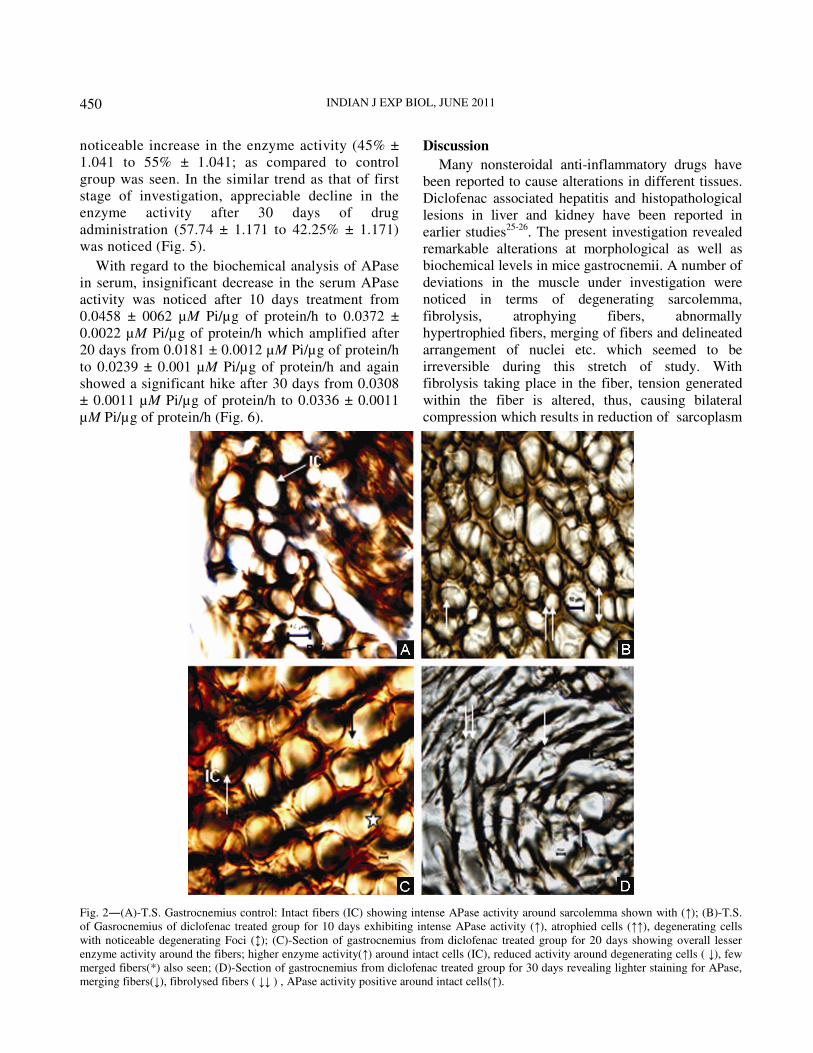
With regard to histochemical studies, the
micrographs of gastrocnemius muscle exhibited
darker staining with intense APase activity around the
sarcolemmal regions in the control group. Further,
section from control group showed normal fibers with
intact boundaries having a uniform pattern (Fig. 2A);
whereas the intensity of enzyme positive areas was
found to be lighter in the section from treated group
after 10 days of treatment. Degenerating fibers and
merging sarcolemma were also noticed. The enzyme
activity was found to be more pronounced around the
smaller fibers as compared to the hypertrophied ones
where enzyme activity seemed to be less (Fig. 2B).
After 20 days of treatment with diclofenac, the
increased number of hypertrophied as well as
degenerating fibers with degeneration foci were
observed (Fig. 2C). Also lesser enzyme activity
around the merging sarcolemma was seen as
compared to control. After 30 days, weaker staining
for APase was observed; many fibers were found
without sarcolemma being merged with each other
along with somewhat larger and intact fibers showing
more enzyme activity in comparison to other
degenerating fibers. However, some atrophic fibers
also had darker staining for APase. Fibrolysis was
also noticed along with lesser enzyme positive areas
(Fig. 2D).
These histochemical findings were also confirmed from biochemical assay where APase activity in gastrocnemius decreased significantly as compared to control after first stage of investigation. Activity of APase in gastrocnemius showed a decrease from 0.0143 ± 0.0014 µM Pi/µg of protein/h to 0.0073 ± 0.0003 µM Pi/µg of protein/h after 10 days of treatment. Significant increase was observed in alkaline phosphatase activity after 20 days of treatment from 0.00364 ± 0.00016 µM Pi/µg of protein/h to 0.00484 ± 0.00024 µM Pi/µg of protein/h. After 30 days of treatment, slight decrease in APase activity [0.0022 ± 0.0001 µM Pi/µg of protein/h to 0.0019 ± 0.00014 µM Pi/µg of protein/h; was found (Fig. 3)].
The biochemical findings were further supported
by electrophoretic study (Fig. 4). An image analysis
of gels showed that in treated group, alkaline
phosphatase activity decreased significantly from
61.62 ± 1.227 to 38.37% ± 1.227 after 10 days of
treatment, while at the second stage of investigation,
Fig. 1―(A)-Lightmicrograph of control mice gastrocnemius showing normal fascicles (NF) and individual muscle fibers with peripheral
arrangement of the nuclei (NU); H & E × 200; (B)-T.S. gastrocnemius of control mouse showing perimysium (PM) around individual
fiber bundle, Endomysium (EnM) surrounds muscle fiber; peripherally placed nuclei (NU) are clearly visible (H & E; × 400); (C)- T.S. of
gastrocnemius of diclofenac treated mice for 10 days showing loss of cellular architecture (←), atrophied fiber (AF), blood vessel ( BV)
is also seen (H & E × 400); (D)-Histopathological changes like irregular limiting membrane, exhibiting merged adjacent fibers (MF),
centrally placed nuclei (*), fibrolysed fibers (F Fb) and hypertrophied fibers (HF) observed in gastrocnemius after 20 days diclofenac
treatment; chains of nuclei is noticed; H & E × 400; (E)-Diclofenac treatment for 30 days resulted in aggravated structural disorganization
in terms of fiber degeneration (*), atrophied fibers showing loss of nuclei (AF) and infiltration of inflammatory cells (•) in mice
gastrocnemius; H & E × 200; (F)-Photomicrograph from 30 days diclofenac treated gastrocnemius muscle exhibiting degenerating fibers
(DF), atrophied fiber (AF), merged fibers (MF); clumped inflammatory cells also visible H & E × 400.

CHOUHAN & SHARMA: DICLOFENAC SODIUM & ALKALINE PHOSPHATASE ACTIVITY
449
INDIAN J EXP BIOL, JUNE 2011
450
noticeable increase in the enzyme activity (45% ±
1.041 to 55% ± 1.041; as compared to control
group was seen. In the similar trend as that of first
stage of investigation, appreciable decline in the
enzyme activity after 30 days of drug
administration (57.74 ± 1.171 to 42.25% ± 1.171)
was noticed (Fig. 5).
With regard to the biochemical analysis of APase
in serum, insignificant decrease in the serum APase
activity was noticed after 10 days treatment from
0.0458 ± 0062 µM Pi/µg of protein/h to 0.0372 ±
0.0022 µM Pi/µg of protein/h which amplified after
20 days from 0.0181 ± 0.0012 µM Pi/µg of protein/h
to 0.0239 ± 0.001 µM Pi/µg of protein/h and again
showed a significant hike after 30 days from 0.0308
± 0.0011 µM Pi/µg of protein/h to 0.0336 ± 0.0011
µM Pi/µg of protein/h (Fig. 6).
Discussion
Many nonsteroidal anti-inflammatory drugs have
been reported to cause alterations in different tissues.
Diclofenac associated hepatitis and histopathological
lesions in liver and kidney have been reported in
earlier studies25-26
. The present investigation revealed
remarkable alterations at morphological as well as
biochemical levels in mice gastrocnemii. A number of
deviations in the muscle under investigation were
noticed in terms of degenerating sarcolemma,
fibrolysis, atrophying fibers, abnormally
hypertrophied fibers, merging of fibers and delineated
arrangement of nuclei etc. which seemed to be
irreversible during this stretch of study. With
fibrolysis taking place in the fiber, tension generated
within the fiber is altered, thus, causing bilateral
compression which results in reduction of sarcoplasm
Fig. 2―(A)-T.S. Gastrocnemius control: Intact fibers (IC) showing intense APase activity around sarcolemma shown with (↑); (B)-T.S.
of Gasrocnemius of diclofenac treated group for 10 days exhibiting intense APase activity (↑), atrophied cells (↑↑), degenerating cells
with noticeable degenerating Foci (↕); (C)-Section of gastrocnemius from diclofenac treated group for 20 days showing overall lesser
enzyme activity around the fibers; higher enzyme activity(↑) around intact cells (IC), reduced activity around degenerating cells ( ↓), few
merged fibers(*) also seen; (D)-Section of gastrocnemius from diclofenac treated group for 30 days revealing lighter staining for APase,
merging fibers(↓), fibrolysed fibers ( ↓↓ ) , APase activity positive around intact cells(↑).

CHOUHAN & SHARMA: DICLOFENAC SODIUM & ALKALINE PHOSPHATASE ACTIVITY
451
and the myofibrils27
leading to atrophy. COX-2
activity is essential for efficient repair after muscle
injury6 as well as recovery from atrophy
28. NS-398, a
specific COX-2 inhibitor was documented to inhibit
the proliferation and differentiation of myogenic cells
in vitro6. Furthermore, it decreased the regeneration of
injured muscle by delaying the maturation of
regenerating myofibers, promoted fibrosis by
upregulating transforming growth factor (TGF-β1)
expression in mice gastrocnemius. It is speculative
that the observed hisopathological changes in
gastrocnemius might be due to diclofenac
administration which is also a partial COX-2
inhibitor. Rows of nuclei in interfibrillar region and
their displacement from peripheral positions to centre
of the muscle fiber were indicative of some
alterations. Such changes in the nuclear morphology
are apparently on account of loss of nucleoplasmic
substances29
. The administration of different classes
of COX inhibitors like aspirin, nimesulide and
celecoxib when used in animal studies at the clinically
safer doses have also caused alterations in the
biochemical and biophysical state of the intestinal
brush border membrane30
while diclofenac was
reported to have negative effect on colon anastomotic
healing in rats31
. Tubular epithelial cell degeneration
and necrosis in kidney and acute hepatitis has been
earlier reported in rats on administration of the high
dose of diclofenac32
that may be reversible or
irreversible.
The present study revealed that diclofenac induces
remarkable changes in specific activity of alkaline
phosphatase in serum and mice muscle as well.
Alkaline phosphatase has been categorized as cell
activity biomarker33
as well as hydrolytic enzyme
functioning at alkaline pH optima by a number of
workers earlier34
. Biochemically, it was found that
specific activity of APase in gastrocnemius decreased
to 48.74% and increased to 132.96% after 10 and 20
Fig. 3―APase activity in mice gastrocnemius in µM Pi /µg of
protein / h [***P <0.01,**P<0.02, *P<0.05; n=6].
Fig. 4―APase activity, analyzed by native PAGE in mice
gastrocnemius: D10 control group (lane 1), D10 diclofenac treated
group (lane 2), Lane 3 ,4 depicting enzyme activity in control
group and diclofenac treated group for 20 days (D20 group)
respectively, enzyme activity in control and drug treated group for
30 days (lane 5 and 6).
Fig. 5―Alkaline phosphatase activity (%) in mice gastrocnemius
[**P<0.001; *P<0.01; n=7 each].
Fig. 6―APase activity in serum of mice in µM Pi /µg of protein /
h [**P<0.001, *P<0.01; n=6].

INDIAN J EXP BIOL, JUNE 2011
452
days respectively. These altered patterns of the
enzyme activity were further confirmed by
densitometric analysis of gels where APase activity
declined (37.73%) which augmented to 122.2% after
20 days drug administration. The lytic process and
stimulation of alkaline phosphatase has been
documented in skeletal muscle by some workers35
earlier. In another study, enhanced fibrolysis was
documented with parallel rise in APase activity in
sciatectomized chick gastrocnemii36
.
In the treated spleen, a significant increase in the activity of APase was observed in the foamy cells and macrophages as well after use of diclofenac in rabbit
37. After 30 days of treatment, activity of APase
decreased to 13.97% in gastrocnemius in biochemical assay while elctrophoretic analysis also recorded parallel results (26.83% diminution). In previous studies with aspirin and nimuslide on rat intestine, the increase in the alkaline phosphatase enzyme activity was indicative of intestinal dystrophy
30. The decline
observed in APase activity during initial stage could be due to the cytoprotective role of non-steroidals to some extent. However, when the drug was continued for another 10 days, enhanced APase activity was noticed which further diminished after 30 days (13.97% diminution). From these varied trends of
APase activity, it is hypothesized that by lowering APase, diclofenac has played role of anti-inflammation protecting the tissue while in 2
nd stage
of the study, trend of enzyme reversal may be due to an adverse effect of the drug. Higher levels of alkaline phosphatase activity in diseased human muscle has
been associated with lytic role of enzyme and also leading to proliferation of non-contractile connective tissues
9. It is noteworthy that the role of alkaline
phosphatase varies differently in different tissues. APase has been proposed to be a function related marker in the renal proximal epithelia
38 having
protective effect on proximal tubule damage of the kidney
39. However, after aspirin treatment, 80%
decrease in APase activity in rat heart was documented to play cardiovascular protective role by preventing calcification of the system
40.
Advanced age and long-term physical exercise
cause changes in the activity of APase in rat muscle41
.
The nonspecific increase in the activity of alkaline
phosphatase by diclofenac exposure may be a result
of the incorporation of the drug in the place of zinc
atoms leading to an increase in the activity of this
enzyme as APase being a metaenzyme37
. In serum,
APase specific activity decreased to 18.78% and
showed 132.04% increase after 10 and 20 days and
again after 30 days of diclofenac treatment, increase
(109.09%) was noticed. Previous studies have shown
that cataflam (potassium diclofenac) significantly
increased the activity of alkaline phosphatase in
rat serum42
which is indicative of skeletal and
muscular disorders8.
The trends of APase in gastrocnemius and serum
in the present investigation exhibited unexpected similar decline in the enzyme activity after 10 days of drug treatment and hypothesized that decrease in the serum APase could be accounted for its hike in other tissues. However, increase in the serum APase during the last two stages definitely hints towards
some deleterious effects of the drug on different tissues including gastrocnemius for which many morphological evidences have been proposed. Increased transmembrane transport of diclofenac has been suggested where APase is involved in the absorption and transportation across these
membranes43
. The involvement of alkaline phosphatase in active transport
44 was reported by a
number of workers. Histochemical studies on gastrocnemius muscle
revealed lesser APase activity around sarcolemma initially. It was noticed that atrophied (narrow) fibers
showed greater activity as compared to the normal fibers after 10 days of treatment. It has been reported earlier that increased fiber alkaline phosphatase positivity is correlated with an increased incidence of degeneration and regenerative changes, fibrosis and atrophy in skeletal muscles
45-46. An association
of APase with pathological conditions has also been reported for skeletal muscle of chicks and rats, in which striated muscles revealed increased activity in pathological conditions such as denervation
47.
Declined levels of APase later on suggested that
the system may show adaptive responses towards
otherwise cytotoxic effects of the drug. However, the
level of serum APase was found to be higher in last
two stages of investigation suggesting some kind of
deleterious effects caused due to diclofenac
treatment. An increase in the activity of serum
alkaline phosphatase after aspirin treatment was
reported earlier and this increase was linked with the
hepatotoxic effects caused by aspirin48
. Although,
there remains a question of future research due to
limitation of this study as to look for other possible
reasons of raised levels of serum APase taking other
vital organs such as liver, kidney, bone and intestine
etc. into consideration. It is concluded from

CHOUHAN & SHARMA: DICLOFENAC SODIUM & ALKALINE PHOSPHATASE ACTIVITY
453
the present study that altered APase activity after
sub-chronic diclofenac treatment can cause
histopathological and physiological changes in
skeletal muscle and may affect other tissues as well.
References
1 Oaks J L, Gilbert M, Virani M J, Watson R T, Meteyer C U,
Rideout B A, Shivaprasad H L, Ahmed S, Chaudhury M J,
Arshad M, Mahmood S, Ali A & Khan A A, Diclofenac
residues as the cause of vulture population decline of white-
backed vultures in Pakistan, Nature, 427 (2004) 630.
2 Nair P, Kanwar S S & Sanyal S N, Effects of non steroidal
anti-inflammatory drugs on the antioxidant defense system
and the membrane functions in the rat intestine, Nutr Hosp,
21 (2006) 638.
3 Brune K & Hinz B, Selective cyclooxygenase-2 inhibitors:
Similarities and differences, Scand J Rheumatol, 33 (2004) 1.
4 Muller S S, Curcelli E C, Sardenberg T, Zuccon A, De
Crudis J L Jr & Padovani C R, Clinical and biomechanical
analysis of the effect of Diclofenac sodium in tibial fracture
healing in rats, Acta ortho bras, 12 (2004) 197.
5 Tang W, Stearns R & Banndiera S M, Studies on cytochrome
P-450 mediated bioactivation of Diclofenac in rats and
human hepatocytes: Identification of glutathione conjugated
metabolites, Drug Metabol Dispos, 27 (1999) 365.
6 Shenn W, Li Y, Tang Y, Cummins J & Huard J, Ns-398, a
Cyclooxygenase-2-specific inhibitor, delays skeletal muscle
healing by decreasing regeneration and promoting fibrosis,
Am J Pathol, 167 (2005) 1105.
7 Rodemann H P & Goldberg A L, Arachidonic acid,
prostaglandin E2 and F2α influence rates of protein turnover in
skeletal and cardiac muscle, J Biol Chem, 257 (1982) 1632.
8 Kim W J, Cho H S & Hong S S, Studies on alkaline
phosphatase isoenzymes in the serum and organs of the rat,
Yonsei Med J, 25 (1984) 142.
9 Kar N C & Pearson C M, Alkaline phosphatase in normal
and diseased human muscle, Pro Soc Exp Biol Med, 14
(1972) 4.
10 Pearse A G E, Alkaline Phosphatases, in Histochemistry,
Theoretical and Applied, (J and A Churchill, London)
1968, 495.
11 Wada H, Yagami I, Niwa N, Hayakawa T & Tsuge H,
Distribution and properties of rat intestinal alkaline
phosphatase isoenzymes, Exp Anim, 50 (2001) 153.
12 Harris H, The human alkaline phosphatase: What we know
and what don’t know, Clin Chim Acta, 186 (1989) 133.
13 Safadi A, Livne E, Silbermann M & Reznick A Z, Activity
of alkaline phosphatase in rat skeletal muscle localized along
the sarcolemma and endothelial cell membranes, J
Histochem Cytochem, 39 (1991) 199.
14 Kumar P & Katoch S S, Effects of denervation and nerve
crushing on chicken muscle, Indian J Exp Biol, 32 (1994) 396.
15 Weiss M J, Ray K, Fallon M D, Whyte M P, Fedde K M,
Lafferty N A, Mulivor R A & Harris H, Analysis of
liver/bone/kidney alkaline phosphatase m-RNA, DNA and
enzymatic activity in cultured skin fibroblasts from 14
unrelated patients with severe hypophosphatasia, Am J Hum
Genet, 44 (1989) 686.
16 Gomori G, The distribution of phosphatase in normal organs
and tissues, J Cell Comp Physiol, 17 (1941) 71.
17 Lillie R D, Enzymes, in Histopathologic Technic and
Practical Histochemistry, (McGraw-Hill, Inc., United States
of America) 1965, 312.
18 Miao D & Scutt A, Histochemical localization of alkaline
phosphatase activity in decalcified bone and cartilage, J
Histochem Cytochem, 50 (2002) 333.
19 Weil L & Russel M A, Study on plasma phosphatase activity in
relation to fat metabolism in rats, J Biol Chem, 136 (1940) 9.
20 Lowry O H, Rosebrough N J, Farr A L & Randall R J,
Protein measurements with folin-phenol reagent, J Biol
Chem, 193 (1951) 265.
21 Franki J, Ruuskanen O & Kouvalainen J, Biochemical
characterization of alkaline phosphatase in guinea pig
thymus, Biochim Biophys Acta, 482 (1977) 370.
22 Kornblatt M J, Klugerman A & Nagy F, Biochemical
characterization and localization of alkaline phosphatase
activity in rat testes, Biol Reprod, 29 (1983) 157.
23 Epstein E, Wolf P L, Hortwitz J P & Zak B, An indigogenic
reaction for alkaline phosphatase in disc electrophoresis, Am
J Clin Pathol, 48 (1967) 530.
24 Manchenko G P, Handbook of detection of enzymes
on electrophoretic gels, (CRC Press, Boca Raton, Florida)
2003, 305.
25 Bhogaraju M, Nazeer S, Al-baghdadi Y, Rahman M,
Wrestler F & Patel N, Diclofenac-associated hepatitis, South
Med J, 92 (1999) 711.
26 Reddy N C P, Sivasankari A & Rao K A, Comparative
toxicity studies in birds using nimesulide and diclofenac
sodium, Environ Toxicol and Pharmacol, 22 (2006) 142.
27 Gutman E, The Denervated Muscle (Publishing House of
Czechoslovak, Academy of Sciences, Prague) 1962.
28 Bondesen B A, Mills S T & Pavlath G K, The COX-2
pathway regulates growth of atrophied muscle via multiple
mechanisms. Am J Physiol Cell Physiol, 290 (2006) C1651.
29 Cameron R, Pathological changes in cells, in Cytology and
Cell Pathology, edited by G H Bourne (Academic Press,
New York) 1964.
30 Sood N, Kaushal N & Sanyal S N, Effect of different non-
steroidal anti-inflammatory drugs, aspirin, nimesulide
and celecoxib on the disaccharide hydrolases and
histoarchitecture of the rat intestinal brush border membrane,
Nutr Hosp, 23 (2008) 326.
31 Esen E, Sucullu I, Sinah H, Kucukodaci Z, Filiz A I, Yucel E
& Akin M L, The effects of non-steroid anti-inflammatory
drugs on healing of colonic anastomosis in rats, Eastern J
Med, 13 (2008) 13.
32 Aydin G, Gokcimen A, Cicek E, Karahan N & Gokalp O,
Histopathologic changes in liver and renal tissues induced by
different doses of diclofenac sodium in rats, Turk J Vet Anim
Sci, 27 (2003) 1131.
33 Araujo B de M, Voltarelli F A, Contarteze R V L,
Manchado-Gobatto F de B & Mello M A R de, Oxidative
stress in rats exercised at different intensities, J Chinese Clin
Med, 14 (2009) 11.
34 Pennington R J T, Biochemical aspect of muscle disease, in:
Disorder of voluntary muscle, edited by J N Walton
(Churchill, Livingstone, London) 1974.
35 Malhotra R K & Katoch S S, On the growth metabolism in
chick skeletal muscle. A study on the alterations in proteins,
glycogen and nucleic acid levels, Acta Biol Med Germ, 37
(1978) 1523.

INDIAN J EXP BIOL, JUNE 2011
454
36 Sharma S, Alterations in the lipid profile of chick skeletal muscle under stress conditions, Ph.D. Thesis, H.P.University, Shimla, 1987
37 Taib N T & Jarrar B M, Histochemical alterations in the
spleen of rabbits induced by Diclofenac Sodium (Voltaren), J
King Saud Univ, Science (1), 19 (2006) 21.
38 Hui M & Cheng P T, Tissue non-specific alkaline
phosphatase may be a function-related marker in renal
proximal tubular epithelia and in vascular endothelia as it is
in osteoblasts, Cell Physiol Biochem, 6 (1996) 296.
39 Heemskerk S, Masereeuw R, Moesker O, Bouw M P W J M,
Hoeven J G V D, Peters W H M, Russel F G M & Pickkers
P, Alkaline phosphatase treatment improves renal function in
severe sepsis or septic shock patients, Crit Care Med, 37
(2009) 417.
40 Mota A, Silva P, Neves D, Lemos C, Calhau C, Torres D,
Martel F, Fraga H, Ribeiro L, Alçada M N M P, Pinho M J,
Negrão M R, Pedrosa R, Guerreiro S, Guimaraes J T,
Azevedo I & Martins M J, Characterization of rat heart
alkaline phosphatase isoenzymes and modulation of activity,
Braz J Med Biol Res, 41 (2008) 600.
41 Reznick A Z, Steinhagen-Thiessen E & Silbermann M,
Alkaline phosphatase activity in striated muscle: the effect of
aging and long-term training in female mice, Arch Gerontol
Geniatr, 9 (1989) 59.
42 Ebong P E, Eyong E U & Udosen E O, Effects of aspirin
(acetylsalicylic acid) and cataflam (potassium diclofenac) on
some biochemical parameters in rats, Afr J Med Med Sci, 27
(1998) 243.
43 Sandhir R & Gill K, Effect of lead on lipid peroxidation in
liver of rat, Biol Trace Element Res, 48 (1995) 91.
44 Denielli J F, Structural factors in cell permeability and
secretion, Symp Soc Exp Bio, 6 (1972) 1.
45 Engel W K & Cumsnnimigham G G J, Alkaline phosphatase
positive abnormal muscle fibers of humans, J Histochem
Cytochem, 18 (1970) 55.
46 Cros D, Pearson C & Verity M A, Polymyositis-
Dermatomyositis. Diagnostic and Prognostic Significance of
Muscle Alkaline Phosphatase, Am J Pathol, 101 (1980) 159.
47 Malhotra R K, Dhingra S & Kathoch S S, Alkaline
phosphatase activity in normal and denervated skeletal
muscle, Expenientia, 34 (1978) 1206.
48 Singh H, Chugh J C, Shembesh A M, Ben-Musa A A &
Mehta H C, Hepatotoxicity of high dose salicylates therapy
in acute rheumatic fever, Ann Trop Paediatr, 12 (1992) 37.







