
STUDIES ON INTERACTION OF ROOT-KNOT NEMATODES AND ROOT-INFECTING FUNGI
ON AIR POLLUTION STRESSED LEGUMINOUS CROPS
DISSERTATION SUBMITTED IN PARTIAL FULFILMENT OF THE REQUIREMENTS
FOR THE AWARD OF THE DEGREE OF
jWas^ter of ^jjilosfopljp IN
AGRICULTURE
SYED ABBAS KOSSEINI-NBGAD
AGRICULTURE CENTRE ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA)
1991

^UNTU l.Mi' ''
•;:r/ . ACC/NO.
:k '"^-^
'7(.f.,
. ^ ? /
DS2061

Decficatecf
to
Mjj Parents

Dr. M. Uajid Khan H . S c , Ph.D., F.L.S., F.P.S.I.
Reader
AGRICULTURE CENTRE DEPARTMENT OF BOTANY ALI6ARH HUSLIM UNIVERSITY ALIGARH-202002 (U.P.) INDIA
Certificate
This i s to certify that Mr. Syed Abbas Hosselni Negad
has worked in this centre as a Research Scholar under my
guidance. His dissertation on Studies on interaction of root-knot
nematodes and root-infecting fungi on air pollution stressed
leguminous crops is original and upto-date. He i s allowed to
submit this dissertation for consideration of the award of the
degree of Master of Philosophy in Agriculture (Plant Pathology).
(M. WAJID KHAN) RESEARCH SUPERVISOR

ACKNOWLEDGEMENTS
Submission of the present dissertation provides me the
best opportunity to express profound sense of gratitude and
sincere thanks to my supervisor Dr.M. Wajid Khan, Reader,
Agriculture Centre, Department of Botany, Aligarh Muslim
University, Aligarh for suggesting the problem,
encouragement, inexhaustible source of inspiration, his able
guidance and supervision and valuable suggestion during the
preparation of this manuscript. It has been indeed a proud
privilege to learn from him as a research supervisor.
The cooperative attitude of Prof. Syed Israr Husain,
the present and Prof. Khalid Mahmood, the Ex-Coordinator of
the Agriculture Centre, A.M.U., Aligarh in providing
necessary facilities is greatly appreciated.
Thanks are also due to Prof. Abrar M.Khan, Professor
Emeritus, Department of Botany, A.M.U., Aligarh for his
encouragement and valuable suggestions.
I would like to place my thanks to Dr. Saad Ashraf, Dr.
Abrar Ahmad Khan, Dr. Mujeebur Rahman Khan, Dr. Nisar Ahmad
Ansari and Mr. Kamal Singh for their help and cooperation.
I offer my heartiest respect to my most affectionate
parents who have always been a source of inspiration,
encouragement, patience and support.
yLz-A^ SYED ABBAS HOSSEINI-NEGAD

TABLE OF CONTENTS
PAGE
INTRODUCTION 1
LITERATURE REVIEW 17
Air Pollution and Plants 19
Effects of Air-borne Sulphur Pollutions 20 on Plants
Susceptibility and Resistance of 34 Plants to SOg
Physiological and Biochemical effects 35 of SOg on Plants
Nematode Parasites of pulses 40
Pulses and Soil-borne Fungal Pathogens 42
Interaction of Sulphur Dioxide and Plant 49 Parasites
Fungal Plant Pathogens and Sulphur Dioxide 49
Plant Parasitic Nematodes 55
Plant Pathogenic and Nodule-forming Bacteria 60
Interaction of Sulphur Dioxide with other 62 Air pollutants
Interaction of Nematodes with other Soil-borne 64 Plant Pathogens
Nematode and Fungal Plant Pathogens 66
(a) Interaction with Wilt causing Fungi 69
(b) Interaction with Root-rot Fungi 81
Interaction of Nematode and Fungal Plant 90 Pathogens under Pollution Stress
MATERIAL AND METHODS 93
1. Root-knot Nematode Population 93
(a) Collection of Field Populations 93

(b) Identification of Species
(i) Perineal Pattern Method
(ii) North Carolina Host Differential test
(c) Maintenance of Inoculum
(d) Inoculation
Inocula of F. oxysporum f.sp. ciceris and R. bataticola
(a
(b
Po
(a
(b
(c
(d
(e
(f
(g
(h
(i
Maintenance and Pure Culturing
Preparation of Inoculum
lutant (Sulphur Dioxide)
Exposure Chamber
Generation of SOg Gas
Exposure and Doses
Air Pollution Symptoms
Sampling
Analysis of Sulphur Dioxide
Standardisation of Sodium Thiosulphate
Standardisation of Metabisulphite Solution
Working Standard Metabisulphite solution in Absorbing Media
(j) Calibration of Standard Curve
(k) Estimation
4. Pathogenicity Tests
Pathogenicity of M. incognita
Sensitivity to SOg
5. Interaction Studies
(a) Interaction of M. incognita with wilt and root-rot fungi
93
93
94
95
95
96
96
96
S<
CL
99
100
100
100
101
103
104
104
105
106
106
108
109
110
110

(b) Interaction of SOg with wilt and root-rot 111 fungi
(c) Interaction of SOg with M. incognita 112
(d) Interaction between SOg, wilt and root-rot 112 fungi and M. incognita
6. Laboratory Experiments 113
(a) Effect of SOg on Hatching and Mortality of 113 M. incognita
(b) Effect of Acidified Soil on Hatching and 114 Mortality of M. incognita
7. Measurement of Soil pH. 116
8. Raising of Seedlings 115
9. Rhizobium inoculation 115
10. Parameters 116
11. Root Nodule Bacteria 116
LITERATURE CITED 117

LIST OF TABLES
TABLE No. PAGE
1. Maximal Permissible Concentrations of Harmful 21
Substances in the Air over Polluted Areas
2. Response to Sulphur Dioxide-Acute effects 25
3. Response to Sulphur Dioxide-Chronic effects 29
4. Response to Sulphur Dioxide-Subtle effects 31 5. Differentiation between three types of Plant 32
effects caused by SOp
6. Interaction of Sulphur Dioxide and Plant 50 Pathogenic Fungi
7. Effect of Air Pollution on Plant Parasitic 56 Nematodes
8. Interaction of Sulphur Dioxide (SOg) and Plant 61 Pathogenic Bacteria and Viruses
9. Effect of Sulphur Dioxide Mixture with other 65 Pollutants on soybean
10. Interaction of Root-knot Nematodes with Wilt 83 causing Fusaria and Root-rot Fungi
11. North Carolina Differential Host Test Reaction 97 Chart

INTRODUCTION

Pulses are one of the most important sources of protein
in human dietary system. Their importance becomes more
appreciable for vegetarian populations particularly in India,
otherwise they would suffer from protein deficiency. Pulses
are used as a food in a variety of forms. The most common is
the soup, the split pulse grain being boiled in spiced water,
grains (seeds) may be ground and boiled, roasted or fried.
Pulses are also grown as fodder crops. Their yrainr ^rc
used also as concentrates to enrich the diet of the animals.
Some pulse crops grown also as green-manure crops and are
excellent for this purpose. They fix atmospheric nitrogen
symbiotically in association with root nodules bacteria and
show no dependence on nitrogen fertilizers. Their ability to
thrive under a wide range of soil and climatic conditions,
specially with their deep root system and ability to with
stand dry conditions more successfully than most other
commonly grown crops are excellent. Therefore, some of the
pulse crops are suitable for dry farming and can figure
prominently in soil conservation programmes.
One of the most important pulse crop grown in India is
gram {Cicer arietinum L.). It is a multipurpose pulse crop
grown throughout the country for various uses. Its leaves are

as a vegetable, spiced and cooked. In general, it is a rich
source of protein, fat, carbohydrate and vitamins.
Soybean, Glycine max (L.) Merr., is another Important
legume grown mainly as a food crop in various parts of the
world. In recent years, soybean has attained great importance
in the U.S.A which shows the highest acreage under this crop.
The cultivation of this crop in India is also getting
significance. Because of its high nutritional values and good
quality of oil, soybean seeds have been put to a wide rang*
of uses through-out the world. Seeds of soybean are consumed
green, dry or sprouted, besides making various products such
as soymilk, soysauce, cheese called 'Tofu' and curd. The
soybean milk can be used for feeding infants. In the U.S.A.
although it is grown essentially as an oil seed crop and more
than 90 percent of the produce is annually crushed for oil,
it has vast diversification of industrial ulitization. The
commercially important by-products of oil industry include
lecithin, soap stock and oil meal for livestock and poultry;
and the edible ones comprise margarines, shortening and salad
oils. The non-edible outlets are paints, varnishes, inks, oil
cloth, enamel and linoleum.
Recently attention of the Indian Council of
Agricultural Research was drawn to the problem of pulses in
India and an integrated All India Coordinated Programme for

the improvement of pulse crops has been undertaken. The work
is being done in collaboration with U.S. Department of
Agriculture at different centres in India.
The nutritive quality of the pulse grain is an
important aspect of research work which is considered by the
plant breeders from the beginning of the programme. The plant
breeders has also considering the disease and pests problems
which causes severe damage to the crops.
Like many other crops pulses are attacked by a numtcr
of plant pathogen. Fungi, bacteria, viruses and nematodes are
common pathogens that cause an array of diseases of economic
consequences. The most important fungi attacking the aerial
parts of the crops are Ascohyta rabiei causing blight,
Uromyces phaseoTi (rust), Alternaria spp. (leaf blotch),
Botrytis spp. (chocolate spot), Erysiphe polygoni (powedy
mildew), Stemphylium spp. (Chlorosis of leaves), Cercospora
spp., Pleospora herbarum, Colletotrichum limdemuthianum f.
sp. phaseoli (anthracnose). Colletotrichum destructivum,
Leviuellula taurica Lev. (powdery mildew), Oidium spp. and
Helminthosporium spp. The fungal pathogen of roots of pulses
are Rhizoctonia bataticola, R. solani, Fusarium oxysporum, F.
lateritium, F. culmorum, F. avenaceum, Phythium spp.,
Phialophora spp. Peyronelleae spp. and Sclerotium rolfsii.
Among the bacterial diseases of pulses, bacterial blight
caused by Pseudomonas glycinea Coerper, bacterial pustule

caused by Xanthomonas phaseoli and w i l d f i r e caused by
Pseudomonas tabaci (Wolf and Foster) F.L. Steven, are the
most important ones. Pulses a lso s u f f e r from v i r a l diseases
t o o . Some examples are Bean Common Mosaic V i rus (8MV), Common
Aphid t r a n s f e r r e d Mosaic v i r u s (CAMY), Bean Yellow Mosaic
V i rus (BYMV), Bean Mosaic V i rus (BMV), and Pea Mosaic V i rus
(PMV).
Among the f unga l d isease o f c h i c k - p e a and soybean ,
Fusarium oxysporum f . sp. ciceris (Padwick) Chattopedhyay ar"-'
Sen Gupta and Rhizoctonia bataticola ( T a u b . ) Bi. 1 .
(.=Sc1erotium bataticola Taub.) causing w i l t and r o o t - r o t of
t h e i r r e s p e c t i v e hosts have been f o u n d t o be t h e most
s e r i o u s i n I n d i a and a r e o f t e n r e s p o n s i b l e f o r " s o i l
s ickness" problem when the s o i l becomes heav i l y contaminated
w i t h them due t o cont inuous crop c u l t i v a t i o n .
A v a r i e t y o f p l a n t p a r a s i t i c nematodes have been
found i n a s s o c i a t i o n w i t h p u l s e s . Spec ies o f Heterodera,
MeToidogyne, RotylenchuTus, Tylenchorhynchus, Aphelenchoides,
Xiphinema, He!icotylenchuTus, Hemicriconemoides. Paraphelen-
chus, Paratrichodorus, Hoplolamis, HirschmanieTIa, Macro-
posthoni a, Pratyl enchus, Cri conemoi des, Di tylenchus,
Diptherophora, Tylenchus, Caloosia, Telotylenchus, Aglenchus,
and Orisitylus are recorded on pulses (Singh and G i l l , 1990).

Among a l l m e n t i o n e d above , r o o t - k n o t n e m a t o d e s ,
MeToidogyne s p p . w i t h a w o r l d - w i d e d i s t r i b u t i o n and
d i v e r s i f i e d host range are a ser ious pest of pu lses. They
a t t a c k a lmost a l l t he pu lses and t h e i r p a t h o g e n i c i t y have
been es tab l i shed . Meloidogyne incognita causes a s i g n i f i c a n t
r e d u c t i o n i n g r o w t h p a r a m e t e r s o f mo thbean , Vigna
aconitifolius Jacq. (Mishra and Gaur, 1981a), f rench bean,
PhaseoTus vulgaris (Singh e t a1. , 1981), black gram, Vigna
mungo (Mishra and Guar, 1981 b ) , c l u s t e r bean, i.y^.monfis
tetragonaloba (Verma et a / . , 1 9 8 1 ) , pea , Pisum sativum
(Upadhyay and Kusum Dwivedi 1987), cowpea, Vigna unguiculata
( V a r s h n e y et al. , 1 9 8 1 ) , w inged bean , Psophocarpus
tetragonabola (Nayak e t a / . , 1987), soybean, Glycine max
( N a l i n i e t a / . , 1985; K i n l o c h , 1980 ) , c h i c k - p e a , Cicer
arietinum (Mani and S e t h i , 1987; Kumar e t al., 1988 ) . M.
Javanica a lso a t t acks pulses e . g . chick-pea (Goel and Gupta
1984, 1986), pigeonpea, Cajanus cajan (Ngauma and Saka, 1986;
Abdul Salam and Khan 1986), b lackeye bean, Vigna sinensis
(Thomason, 1958). M. arenaria a lso i s a l i m i t i n g f a c t o r i n
p roduct ion of soybean (K in loch e t al., 1987; Rodriguez-Kabana
and W i l l i a m s , 1981) . Soybean c y s t nematode. Heterodera
glycines has been repor ted t o be one of the most widespread
and damaging nematodes of soybean (K in loch , 1980; Wrather and
Anand, 1988).

Females o f r o o t - k n o t nematodes a re s e d e n t a r y
endoparasi tes. Ce l l s around the head and neck of the female
nematodes becomes mul t inuc leated and c e l l wal l i r r e g u l a r l y
th ickened. Usual ly 6-12 of such c e l l s are found around the
neck of each female which f u n c t i o n as t r a n s f e r c e l l s (Jones
and No r thco te , 1972b) and female nematodes o b t a i n t h e i r
n u t r i t i o n from them. Vascu lar t i s s u e s o f t he p l a n t a re
a l t e r e d e x t e n s i v e l y due to i n v a s i o n o f r o o t s by r o o t - k n o t
nematodes which impair the absorpt ion and t rans 1 j c a t t o r o*
w a t e r and n u t r i e n t s t o v e g e t a t i v e p a r t s o f t h e p l a n t .
Suppression o f p l a n t growth and appearance o f d e f i c i e n c y
symptoms on leaves are due t o biochemical changes induced by
the nematodes in the host and a l t e r e d physiology due t o the
anatomical t rans fo rmat ions .
The second s t a g e j u v e n i l e ( J 2 ) o f t h e nematode
penetrates i n t o the root and moves through the cor tex i n t e r -
and i n t r a c e l l u l a r l y t i l l i t reaches c lose t o the p e r i c y c l e .
The f e m a l e become s a c c a t e and s e d e n t a r y and m a i n l y
r espons ib l e f o r pathogenic damages caused t o t h e h o s t
p l a n t s . Root-knot nematode i n t e r a c t s w i t h s o i l - b o r n e p l a n t
pathogens. Synerg is t i c i n t e r a c t i o n s of roo t - kno t nematodes
and severa l s o i l - b o r n e f u n g i a re w e l l documented P o w e l l ,
1971, 1979; Khan, 1984; T a y l o r , ( 1979 , 1991) . R o o t - k n o t
nematodes suppress the format ion of r oo t -nodu la t i on i n pulses
(Dhanger and Gupta, 1983: Rau t i , 1980; Mani & Sethi 1984,

1987), as a r e s u l t o f which the t o t a l b e n e f i t f r o m t h e
n i t rogen f i x i n g bac te r i a i s reduced.
Atmospheric p o l l u t i o n i s a major problem of the modern
w o r l d . The f a s t i n c r e a s i n g human p o p u l a t i o n and q u e s t f o r
improvement i n l i v i n g s tandard have r e s u l t e d i n t o r a p i d
i n d u s t r i a l i s a t i o n , u rban isa t ion , excessive use of f e r t i l i z e r s
and pes t i c i des on major a g r i c u l t u r a l crops and p roduc t ion of
n u c l e a r e n e r g y . C o n s e q u e n t l y , e v e r y component o f t h e
biosphere i . e . the a i r and the s o i l , i s becnmin; t rcme r r'r i^ely
p o l l u t e d by day and n i g h t . A c c o r d i n g t o t h e ^n- 'mates o f
Steve Van Matre (1984), i n the present wor ld on ly 20% of the
a i r i s breathable and only 10X of the land i s capable o f
being e x p l o i t e d f o r producing food. Astan in and Blagosklonov
(1983) r epo r t ed t h a t hundreds o f m i l l i o n s t o n s o f h a r m f u l
gases and dus ts are em i t t ed i n t o t h e e a r t h ' s a tmosphere
every year.
The release of harmful chemicals and t o x i c ma te r i a l s
from i n d u s t r i e s i n t o the environment ( a i r , water , s o i l ) cause
damaging e f f e c t s on both fauna and f l o r a . These t o x i c wastes
i n excess, exceeding the normal s e l f - r e g u l a t i n g capac i t y of
atmosphere, lead t o p o l l u t i o n . Weber (1982) d e f i n e d a i r
p o l l u t i o n as ' t h e presence o f subs tances i n t h e amb ien t
atmosphere, r e s u l t i n g f rom the a c t i v i t y o f man or f r o m
n a t u r a l p rocesses , causing adverse e f f e c t s t o man and t he

environment". In 1966, the committee on pollution, National
Academy of Sciences, U.S.A. defined the pollution as an
undesirable change in the physical, chemical or biological
characteristics of our air, land and water, that may or will
harmfully affect human life or that of desirable species, our
industrial processes, living conditions and cultural assets,
or that may or will waste our raw material resources
(Anonymous, 1966). In other word, any undesirable change in
the physico-chemical and biological properties o^ air, wate-
and soil, which may cause harm to man, other organisms, -.-r t
cultural and natural elements of man's environment, is
pollution.
Environmental pollution includes many kinds of
pollutions such as soil pollution, water pollution, noise
pollution and air pollution. However, the last one probably
has attracted the attention of the world most. Wood (1968)
classified the toxic substances responsible for air pollution
"air pollutants" basically in two categories based on their
origin. Primary air pollutants, those originating at the
source in the form of toxic to living organisms which may be
in gaseous or particulate forms. Gaseous air pollutants are
sulphur dioxide (SOg), oxides of nitrogen (NO^^), hydrogen
fluoride (HF), ammonia (NHg), ethylene (Cg Hg) etc. and
particulate air pollutants are coal dust, cement dust, fly
ash, suspended particulate matter (SPM) etc. The second

category of pollutants are secondary air pollutants which
originates from the reaction between primary air pollutant
that originate from the source e.g. photochemical pollutants
like peroxyacetyl nitrate (PAN) and ozone (O3). SO2 and NOj.
in high humid conditions are converted into acid (Hg SO^ and
HNO3) which fall on the ground in form of 'acid rain' during
the atmospheric precipitation (Oden, 1968).
Sulphur dioxide is one of the most damaging air
pollutant for plants. Sulphur dioxide make entry throLph
stomata to reach the mesophyll of the leaves and t'lut hfeviriu
come in contact with water, change in H2 SO3 or HC03~ if the
— ? — ? medium is acidic, or SO3 - 80^ when alkaline, which
damages to adjoining tissues making for chlorosis and drying
of leaf tips, etc.
P l a n t s a r e a f f e c t e d by t h e p resence o f gaseous
p o l l u t a n t s i n t he a i r . The pe r fo rmance o f t he p l a n t s i s
i n f l u e n c e d d i r e c t l y o r i n d i r e c t l y by i n h i b i t i o n o r
a c c e l e r a t i o n of the p lan t metabol ism, which may in f luence
t h e i r p r o d u c t i v i t y . Many s i g n i f i c a n t and somet imes
devas ta t ing e f f e c t s of a i r p o l l u t i o n of vegetat ion have come
t o l i g h t d u r i n g the c u r r e n t c e n t u r y . Some e a r l i e r r e p o r t s
have revea led the d e v a s t a t i n g e f f e c t o f SOg, f l u o r i d e ,
hydrogen c h l o r i d e and other p o l l u t a n t on p lant popu la t i on .
Recently Temmerman (1982) descr ibed the e f f e c t of ammonia on
v e g e t a t i o n , f o l l o w i n g an a c c i d e n t a l r e l e a s e o f t h e gas.

1 O
A s t a n i n and Blagosklonov (1983) conc luded t h a t v a r i o u s
a f f l i c a t i o n s of p l a n t leaves appear under the i n f l uence of
a i r p o l l u t a n t s and the p lan t growth i s s tun ted. Leaves are
probably the most s e n s i t i v e p lan t par ts to a i r p o l l u t i o n s .
They f i r s t d e v e l o p c e r t a i n p r o t e c t i v e a d a p t a t i o n and
u l t i m a t e l y f a l l prey to the harsh a tmosphere. Stomata and
t r i chomes u s u a l l y undergo changes p e r t a i n i n g t o s i z e and
f r e q u e n c y . P h o t o s y n t h e s i s , a p r o c e s s r e s p o n s i b l e f o r
sustenance of the whole p lan t body i s murh vu lnerab le '• p-'r
p o l l u t i o n damage. The i n f l u e n c e of t o x i c 9ast. . on
a g r i c u l t u r a l crops i s l i t t l e s t u d i e d . However, i n some
i n d u s t r i a l i z e d coun t r ies a i r p o l l u t i o n i s known t o reduce the
y i e l d of many crops near i n d u s t r i a l areas. Plants a f f e c t e d by
su lphu r d i o x i d e are poor i n p r o t e i n ; the sugar bea t s and
potatoes show reduc t ion in sugar and s tarch r e s p e c t i v e l y . The
g r e a t e s t harm t o U.S. a g r i c u l t u r e i s i n f l i c t e d by p h o t -
o x i d a n t s . For example, i n the Los Ange les , V a l l e y f a r m e r s
were fo rced to cease growing c e r t a i n vegetables (sp inach ,
l e t t u c e ) most s u s c e p t i b l e t o p h o t o - o x i d a n t s (ozone and
peroxyacety le n i t r a t e ) , and the c i t r u s f r u i t y i e l d decreased
by 50% (Astan in and Blagosklonov, 1983).
Deposi t ion of p a r t i c u l a t e a i r p o l l u t a n t on lea f sur face
b locks the stomatal c a v i t i e s which hampers t r a n s p i r a t i o n and
checks the t ransmiss ion of so la r r ad ia t i on (Dar ley , 1966).

1 1
Their deposition reduces plant vigour, growth and
productivity.
In agricultural field plots, plants are exposed to a
variety of biotic and abiotic pathogens, especially roots,
which are constantly exposed to many soil microorganisms.
According to Richards (1976), one square meter of highly
fertile field soil may contain as many as 3 x 10 bacterial
cells (300g), 5x10® protozoa (39g), 1x10^ nematodes (12g),
and 400g of fungi. However, this can not generalized for a"ii
soils, and many microbes present are saprophytic or have weak
pathogenic potential to plants. Many investigators have
concluded that the majority of root diseases have a complex
etiology (Grogan 1981; Wallace 1978). Fawcett (1931) stated
that nature does not work with pure cultures and that most
plant diseases, particularly root diseases, are influenced by
associated micro-organisms. Root infections by one pathogen
may modify the host response to subsequent infections by
pathogen or saprophytes.
Nematodes and soil-borne micro-organisms thrives best
under moist soil condition. A number of excellent reviews on
the interactions of plant-parasitic nematodes with other
plant pathogens have been written by Bergeson, (1972)
Pitcher (1963, 1965, 1978), Powell (1963a, 1963b, 1971a,
1971b, 1979), Taylor, (1979, 1991) and Wallace, (1978). The

1 2
rev iews analyse many s p e c i f i c i n t e r a c t i o n s and i n most
i n t e r a c t i o n s i nvo l v i ng fungi and nematodes, nematode was not
essen t ia l f o r the establ ishment and development of the fungal
p a t h o g e n . The nematode u s u a l l y a s s i s t e d t h e f u n g u s by
a l t e r i n g the inc idence and speed o f deve lopment o f t h e
pathogen and thus increas ing the disease s e v e r i t y . Thus, the
i n t e r a c t i o n between the nematode and the fungal pathogen i s
o f ten i n d i r e c t and occurs owing t o induced m o d i f i c a t i o n s i n
the host p l a n t . I n case o f e c t o p a r a s i t i c nematodes, h o s t -
p l a n t m o d i f i c a t i o n may be m i n i m a l , whereas •« n s e d e n t a r y
endoparas i t i c nematodes such as r o o t - k n o t nematodes, these
m o d i f i c a t i o n s are ex tens i ve and c o m p l e x . Bowman et al.,
(1966) r e p o r t e d both l o c a l i z e d and s y s t e m i c h o s t - t i s s u e
m o d i f i c a t i o n s t h r o u g h t h e p r o d u c t i o n o f t r a n s l o c a b l e
m e t a b o l i t e s by nematodes. However, t h e nematodes and t h e
fungal pathogens may occas iona l l y ac t d i r e c t l y on each o t h e r .
Another type of mu l t i p le -pa thogen i n t e r a c t i o n i s the
i n t e r a c t i o n between b i o t i c and a b i o t i c ( a i r p o l l u t a n t s )
pathogens. A i r p o l l u t a n t s are emi t ted t o the atmosphere from
the po in t sources cont inuously and spread i n a l l d i r e c t i o n s
depending upon d i r e c t i o n and v e l o c i t y of the wind, a f f e c t i n g
p lan ts growing in the v i c i n i t y . Therefore on a i r p o l l u t i o n s
s t ressed p l a n t s , associated microorganisms i nc l ud i ng p l an t
pa ras i tes are l i k e l y to be a f f e c t e d . At p resen t , very l i t t l e
i s known of the e f f e c t of p o l l u t a n t s on p a r a s i t i c diseases

-I ?
of plants and host parasite relationships. Some studies
however, have been undertaken to determine this aspect of air
pollution impact. Theoretically, parasitism may be affected
by air pollutants in different ways. Parasitism may be
increased or decreased through a direct effect of pollutant
on the parasite, or the effects may be indirect through
pollutant - induced changes in the host plant. Sulphur
dioxide may act directly upon the fungi or indirectly' throuf*-
some effect upon living or dead plant tissue, soil cr waLf
Studies undertaken show that fungi differ in their reaction
to SO2; rust diseases and wood-destroying fungi are sensitive
to SOg. Wheat stem rust, Puccini a graminis was less in
industrialized area than unindustrialized areas (Johanson,
1954). Scheffer and Hedgcock (1955) found decreased
parasitism by species of Cronarti urn. Coleospori um,
Melampsora, Peridermium, Pucciniastrum, and Puccinia where
trees were injured by SOg. Sulphur dioxide affects fungal
growth too. Hyphae of different species vary in sensitivity
to SOp exposures (MacCallan et al. , 1940).
Pollutants also affect the nematode parasitism.
Reproduction of Heterodera glycines and Paratrichodorus minor
was decreased by Og alone or in mixture of SOg, but that of
Belonolaimus longicaudatus and Pratylenchus penetrans will
not altered (Weber et al. , 1979). Reproduction of P.
penetrans on tomato was slightly suppressed by O3 at 0.20 ppm

1 -4-
but not by SOg a t 0.80 ppm (Brewer 1979). Kozlowska (1981)
o b s e r v e d t h a t i n d u s t r i a l d u s t has h a r m f u l e f f e c t on
Panagrolaimus rigidus. B isessar e t al., (1984) found t h a t
r oo t - kno t nematode (Af. hapTa) i n fec ted tobacco p lan ts were
more s e n s i t i v e t o ambient ozone. However, non- in ju r ious doses
o f SO2 f a i l e d t o a f f e c t a d v e r s e l y t h e p l a n t - p a r a s i t i c
n e m a t o d e s . R e p r o d u c t i o n o f Praty lenchus penetrans was
inc reased when p l a n t s were exposed t o SO2 b e f o r e nematode
i n o c u l a t i o n . Th i s dose o f SO2 was no t enough t o i n j u r e
leaves or s i g n i f i c a n t l y t o a f f e c t p l an t growth. Singh (19£;s)
observed t h a t i nc idence o f i n f e c t i o n o f c h i c k - p e a by M.
incognita and M. Javanica were less around the p o l l u t i o n
sou rce . The i nc i dence o f d i s e a s e was decreased w i t h t h e
inc reas ing d is tance from the p o l l u t i o n source. He observed
t h a t a i r p o l l u t i o n i n general i n h i b i t e d the roo t -kno t disease
on pulse crops in ambient c o n d i t i o n . The extent of i n h i b i t i o n
was c o r r e l a t e d w i t h the ambient l e v e l s o f p o l l u t a n t s . He
noted t h a t j u v e n i l e hatching of both species were adversely
a f f e c t e d by SO2, O3 and t h e i r m i x t u re . Higher concent ra t ions
o f each p o l l u t a n t s i n d i v i d u a l l y o r i n m i x t u r e were more
i n h i b i t o r y t o j u v e n i l e hatch ing and M. incognita was more
s e n s i t i v e than M. javanica t o p o l l u t a n t s . However, much i s
n o t known a b o u t such i n t e r a c t i o n and needs more
i n v e s t i g a t i o n . Although some s tud ies have been undertaken to
determine the e f f e c t of a i r p o l l u t i o n on nematodes or fungal
d isease, the impact of a i r p o l l u t i o n s t ress on development of

1 a
f u n g u s - n e m a t o d e comp lexes on c r o p p l a n t s i s y e t t o be
i n v e s t i g a t e d .
The main o b j e c t i v e of the proposed work i s t o ace r t a i n
the impact o f a i r p o l l u t i o n caused by SOg on r o o t - k n o t -
fusar ium w i l t i n t e r a c t i o n on ch ick-pea and r o o t - k n o t - r o o t r o t
i n t e r a c t i o n on soybean.
E x p e r i m e n t s w i l l be c a r r i e d o u t under a r t i f i c i a l
t r e a t m e n t c o n d i t i o n s i . e . u s i n g exposure cha.oer ^ M . ' V,
t rea tments . The f o l l o w i n g aspects are proposed t o be s tud ied
t o achieve main o b j e c t i v e of the s tudy.
1 . P a t h o g e n i c i t y t e s t of r o o t - k n o t nematode, Meloidogyne
incognita on chick-pea and soybean c u l t i v a r s .
2. Pathogen ic i ty t e s t of w i l t fungus (Fusarium oxysporum f .
sp. ciceris) on chick-pea c u l t i v a r s and r o o t - r o t fungus,
Rhizoctonia bataticola on soybean c u l t i v a r s .
3 . E f f e c t o f s u l p h u r d i o x i d e on c h i c k - p e a and soybean
c u l t i v a r s .
4 . E f f e c t o f M. incognita i n f e c t i o n t o w i l t o f c h i c k - p e a
caused by F. oxysporum f sp. cicers and r o o t - r o t of
soybean caused by R. bataticola.

1 6
5. Interaction of M. incognita and F. oxysporum f. sp.
ciceris on chick-pea and M. incognita and /?. bataticola
on soybean.
6. Interaction of root-knot nematode, M. incognita and SOp
on chick-pea and soybean.
7. Interaction of SOg with F. oxysporum of sp. ciceris on
chick-pea and /?. bataticola on soybean.
8. Impact of SOg on interaction of M. incognit wth P.
oxysporum f. sp. ciceris and /?. £jatat7co7a on their
respective hosts i.e. chick-pea and soybean.
These aspects can be suitably modified depending upon
the progress of the work and results obtained.

LITERATURE REVIEW

1 7-
Total land under the cultivation of pulse crops in
India is about 24 million hectare, with approximate
production of 12 million tons annually. According to Vavilov
(1951) India and Middle East form the primary centre of
origin of most of the important legumes.
Chick-pea (Cicer arietinum L.) is the most important
pulse crop grown in India which occupies an area of abo, t :.:-
million acres, which is about three-fourths the wheat acreage
of India. The chick-pea is perhaps indigenous to the South
east Europe. It is supposed that it was cultivated in Egypt
from the early earliest time of Christian era where it was
introduced from Greece and Italy. Its introduction into India
is of a more early date. There is a Sanskrit name and several
other names in modern Indian languages. It is considered to
have originated in the tract lying between Caucasus and the
Himalayas where from it has spread to the southern Europe,
Persia (Iran), Egypt and India. It has also been introduced
into parts of the central and southern America, Australia and
parts of Africa. Cicer includes 22 species (Index Kewensis,
1913) distributed in the Mediterranean and West and Central
Asia. In Asia minor and Egypt, they occur only in the wild
state. Cicer soongaricum is cultivated in parts of the
Western Himalayas. Grains of the two wild species C.

1 a
pinnatifidum Jaub & Spach (Avdulov, 1937) and C. montbreii
Jaub & Spach are t i n y ; those of former have angular seeds
w i t h minute sp ines on the t e s t a ; w h i l e the g r a i n s of t h e
l a t t e r have a smooth t es ta ( lynegar , 1935),
Soybean {Glycine max ( L . ) Me r r . i n b e l i e v e d t o have
o r i g i n a t e d from south-east As ia , der ived from a twiny slender
p l a n t G. ussuriensis Regal and Macck, commonly growing i n
t h i s reg ion . According to Fukuda (1933), Manchuria should be
the centre of o r i g i n since i t shows d i v e r s i t y in forms i n
t h i s count ry . Engelbert Kaemfer, a German bo tan is t whc wac ir.
Japan dur ing 1690-92, was the f i r s t t o in t roduce soybean t o
European Wor ld . The f i r s t ment ion o f soybean i n Amer ican
l i t e r a t u r e i s by Mease in 1804. General ly three species o f
Glycine are recognized, v i z . , G. ussuriensis ( w i l d soybean),
G. max ( the c u l t i v a t e d fo rm) , and G. gracilis ( in te rmed ia te
t y p e between t h e f i r s t t w o ) . I t i s b e l i e v e d t h a t G.
ussuriensis, fo rmer ly known as 6. soja, i s the progeni tor o f
t h e c u l t i v a t e d soybean ( P i p e r and Mo rse , 1 9 1 0 ) . I t i s
procumbent type w i t h t w i n i n g stems and narrow l e a f l e t s .
Seeds are sooty dark in colour and range from 1-2g. per 100
seeds, wh i le in the c u l t i v a t e d soybean the p lan t i s e rec t or
sem i -e rec t and the seed co lou r v a r i e s f rom y e l l o w , g r e e n ,
brown t o b lack. The seed weight ranges from 5-35 g. per 100
seeds.

-I e
Air Pollution and Plants
All living organisms including plants are dependent on
their environment. The environment constantly influences and
is influenced by its inorganic and organic surroundings. The
various gases and inorganic substances obtained from air,
water and soil one converted into complex organic molecules
by leaves of the plant. Presence of gaseous pollutants in the
air affect the performance of plant directly or indirectly by
inhibiting or accelerating the plant metabolism whic • may
influence its productivity.
A considerable amount of gaseous air pollutants
originating from different activities of man are released
into air which impures atmosphere. Sulphur dioxide (802)1
ozone -(O3), oxides of nitrogen (NO ) are recognised as the
most important toxic gaseous air pollutants and harmful to
the plants. Cameron (1874) observed that factory smoke which
caused disease in plants contained SOg. According to Heck et
al. (1982) crop losses due to air pollutants may reach upto
905($. They suggested that it conducting agricultural
research, air quality-agricultural plant ecosystem
interrelationship should be taken into the consideration.
Heagle (1973) reported that air pollutants alter the
susceptibility of plants to biotic pathogens like fungi,
nematodes, bacteria, etc.

2 0
It is now realised that the pollution damage is
obvious, severe and inescapable unless the violation of the
environment safety is not checked. Astanin and Blagosklonov
(1983) have given a chart for maximum permissible
concentration of harmful substances in the air over polluted
area (Table I).
Effects of airborne sulphur pollutants on plants
Airborne sulfur pollutants which affect plant 1if€ ?•
sulphur dioxide (SOg); hydrogen sulfide (HgS), artd sulj-' . c
acid (Hg SO4), and acid precipitation. The information is
mostly available on SOg, and the effects of this gas on
forest trees, horticultural plants, and lichens have been
documented. Effects of various sulphur pollutants have been
reviewed by Thomas (1952), Daines (1968), Barrett and
Benedict (1970), the US Environmental Protection Agency
(1973), Heck and Brandt (1975), Varshney and Garg (1979),
Amundson (1983), Bender et a1 (1986), Fluckinger and Younger
(1986), Benwart (1987), Bytnerowicz et al., (1987) and Kumar
and Yadav (1988).
Sulphur is an essential element for plants. It is a
constituent of the amino acids cysteine, and methionine. It
is also a constituent of the plant vitamins, thiamine and
biotin, and of other biochemical constituents such as
glutathione, coenzyme A, and cytochrome C. Normally, sulphur
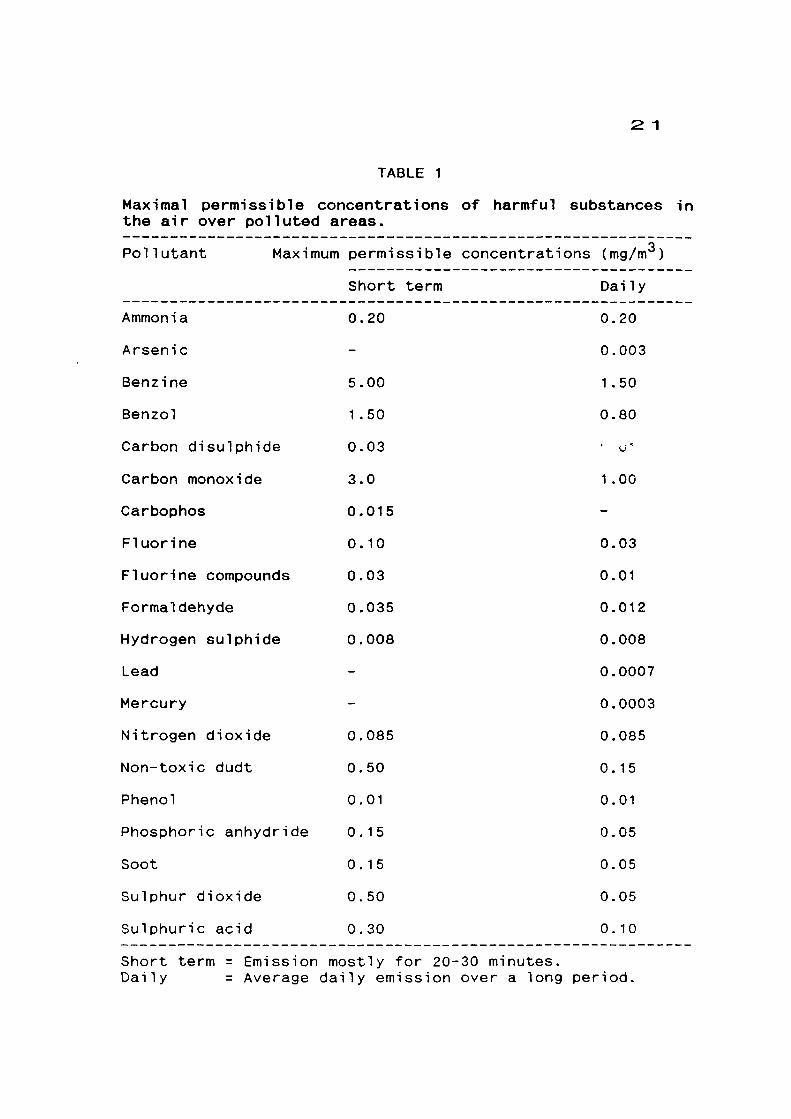
2 1
TABLE 1
Maximal permissible concentrations of harmful substances in the air over polluted areas.
Pollutant Maximum permissible concentrations (mg/m )
Short term Daily
Ammonia 0.20
Arsenic
Benzine 5.00
Benzol 1.50
Carbon disulphide 0.03
Carbon monoxide 3.0
Carbophos 0-015
Fluorine 0.10
Fluorine compounds 0.03
Formaldehyde 0.035
Hydrogen sulphide 0.008
Lead
Mercury
Nitrogen dioxide 0.085
Non-toxic dudt 0.50
Phenol 0.01
Phosphoric anhydride 0.15
Soot 0.15
Sulphur dioxide 0.50
Sulphuric acid 0.30
0.20
0.003
1 .50
0.80
' o"
1 .00
0.03
0.01
0.012
0.008
0.0007
0.0003
0.085
0.15
0.01
0.05
0.05
0.05
0. 10
Short term = Emission mostly for 20-30 minutes. Daily = Average daily emission over a long period,

2 2
is taken up by plants from soil in the sulphate form and
assimilated into various compounds usually after being
chemically reduced. Sulfur dioxide absorbed from the air has
been shown to rapidly undergo oxidation to sulphates inside
plant tissue. De Cormis (1969) found that tomato plants soon
after exposure to SO2 had 98S5 of the S in the form of
sulphates.
Sulphur dioxide is emitted from various industrial
sources and the adverse effects on vegetation 11 uht
vicinity of these sources have been well documented. Some of
the industerial sources are -
1. Zinc and lead Smelters
2. Nickel and Copper Smelters
3. Iron concentrators
4. Petroleum refineries
5. Pulp and paper mills
6. Fossil fuel thermal generating stations
Estimation of the cost of air pollution damage to crops
have varied considerably. Annual loss of $ 500 million have
been estimated in the United States from air pollution injury
to crop plants with approximately one third of this total
loss attributable to SOj injury.

2 3
The first visible evidence of SOp injury to plants is
discernible in the foliage. The stem, bud, and reproductive
parts of the plants are visibly more resistant to SOg. The
effects of airborne sulphur pollutants on plants may be
divided into three categories: acute, chronic, and subtle
(physiological and biochemical). Sulphur dioxide enters
leaves mainly through the stomata and is toxic to the
metabolic processes taking place in the mesophyll cells.
Acute injury is caused by a rapid accumulation of b-.sulp' "'t'
and sulphite. When the oxidation product, sulphate,
accumulates beyond a threshold value that the plant cells can
tolerate, chronic injury occurs. It is estimated that
sulphate is about 30 times less toxic than sulphite (Thomas,
1951 ).
Acute injury is macroscopic necrotic injury to plant
tissue variable within hours or days after exposure to short-
term (less than 24 h) high concentrations of SOg. Acute
injury to broad leaves takes the form of lesions on both
surfaces, usually appearing between veins, and often more
prominent toward the petiole and injury is local. The
metabolic processes are completely disrupted in the dead or
necrotic areas with the surrounding green tissue remaining
functional. In some cases, injury can occur on the margins of
the leaves. Young leaves rarely display necrotic markings,
whereas fully expanded leaves are most sensitive to acute SO2

24-
injury. The anatomical effects of acute injury on foliage can
be seen under the microscope. Initially water-soaked, flaccid
areas of diffuse, gayish-green colouration appear and the
chlorophyll appears to have diffused from the chloroplasts
into the cytoplasm. This is followed by desiccation and
shrinkage of the affected cells. The green pigments are
decomposed, and the affected cells. The green pigments are
decomposed, and the affected leaf area assumes a bleached,
ivory, tan, orange-red, reddish-brown, or otown appee
Sulphur dioxide concentrations that caused injury to
vegetation in short-term periods are listed in Table 2.
Acute injury to forest trees in the field has been
attributed to SO2 doses of 0.95 ppm for 1 h, 0.55 ppm for 2
h, 0.35 ppm for 4 h and 0.25 ppm for 8 h (Dreisinger, 1965).
For acute injury to occur, other environmental and plant
factors are important. These includes sunlight, moderate
temperature, high relative humidity, adequate soil moisture,
and plant genotype and the stage of growth. Katz and McCallum
(1939) subjected various plant species to different SOg
concentrations over a number of exposure periods. The
threshold for injury was found to be 0.3 ppm SO2 for 8 h for
western larch {Larix occidental is) and 0.14 ppm SO2 for 12 h
for Douglas fir {Pseudotsuga taxifolia). In a 1-hr
experimental exposure Zimmerman and Crocker (1934) reported
that 0.66 ppm SO2 injured buck-wheat {Fagopyrum esculentum).

2 5
TABLE 2
Responses to sulfur dioxide - acute effects
SOp Exposure Plant response References cone, period (ppm) (h)
A-Responses to low SO2 doses
0.03 1 Injury to sensitive eastern Costonis (1971) white pine
0.025 6 Injury to sensitive eastern Houston (1974) white pine
B- Responses to medium SO2 ooses
0.25 1 Injury to eastern white pine Berry (1967)
0.25 2 Injury to eastern white red Berry (1971) and jack pines
0.25 2 Injury to Virginia, short Berry (1974) leaf, slash,and loblolly pines
0.25 4 Injury to broccoli Tingey et a1. (1973)
0.66 1 Injury to buckwheat Zimmerman and Crocker (1934)
0.95 1 Injury to foliage of forest Dreisinger trees (1965)
0.70 1 Injury to forest trees under Dreisinger and sensitive environment McGovern (1970)
0.3 8 Injury to weastern larch Katz and McCallum (1939)
C-Responses to high SO2 doses
2.0 2 Injury to 10 of 87 native Hill et a1. desert species (1973)
2.0 4 Injury to 10 weed species Benedict and Breen (1955)
3.0 1 Injury to Acacia pruinosa O'Conner et al ( 1974 ) .

2 6
Acute injury was produced on foliage of trembling aspen
{PopuTus tremuToides) in artificial fumigation with 0.35 ppm
SOg for a period of 3 h. Moderate to severe injury to several
Kentucky bluegrass cultivars (Poa pratensis) in artificial
fumigation of 0.2 ppm SOg for 2 h was observed
The acute effects referred to above may be considered
responses to medium doses of SOg. In the literature there are
reports of responses of plants to much lower arc to
substantially higher SOg doses. In artificial fumigation
experiments SO2 doses as low as 0.3 ppm for 1 h (Costonis,
1971) have been reported to injure extremely sensitive
strains of eastern white pine; and conversely SO2 doses as
high as 2.0 ppm for 2 h (Hill et al., 1973), have been found
to be required in order to injure tolerant plant species as
Acacia pruinosa, several native desert plant species, and
several chrysanthemum varieties respectively. Khan (1989)
found browning and chlorosis in leaves of egg plant, tomato
and okra induced by the SOp specially at 0.2 ppm.
Chronic injury is macroscopic chlorotic injury
(sometimes changing to necrotic injury) to plant tissue
usually developing over a long period of time (from over a
day to 1 or more years) from exposure to variable
concentration of SO2. Long term chronic effects on trees are
related to variety of SOg exposures which includes short-term
high concentration, short term and long-term periods of

2 T
sublethal concentrations, and even SOp-free periods when the
plant life can recuperate by translocating and assimilating
accumulated sulphur. To correlate chronic effects on forest
trees with atmospheric SOg levels, it is preferrable to use
the average concentration for the total period of exposure,
rather than the average of SO2 fumigation periods only. The
U.S. Environmental Protection Agency (1973) has concluded
that many short-term high concentration episodes were
responsible for the injury and growth reductions to occur in
the vicinity of an iron ore concentrator that emitted onl>
about 10 tons of SO2 daily. Although vegetation injury was
severe in the immediate vicinity of the plant, SOg damage
extended for only a little more than 2 km from the source
(Guderian and Stratmann, 1962). Linzon (1958a) observed that
three large nickel and copper smelters that discharged about
6000 tons of SO2 daily into the atmosphere, caused severe
damage to pine forest with both acute and chronic effects
occurring to a distance of about 25 km miles. Beyond this
distance the condition of white pine improved rapidly when
the concentration of SOg were lower than 0.25 ppm. Chemical
analyses of vegetation in the area showed the accumulation
and build-up of sulphur in foliage in vegetation growing
close to SOp sources. Both high and low concentration
fumigations of SOp over varying periods of time contributed
to the accumulation of sulphur in conifers which lead to the
development of chronic injury in older needles. In

2e
Czechoslovakia occurrence of moderate chronic injury to
foliage of spruce trees, under the influence of an average
concentration of SOg of 0.019 ppm was observed. Epiphytic
lichens which are perennial and evergreen are extremely
sensitive to SOg because they are continually exposed to the
gases in a polluted environment. In a study conducted by
LeBlanc et al. (1972), the number of epiphytes found growing
on balsam poplar (PopuTus balsamifera) trees was drastically
reduced in zones where the growing season mean levels o-f Op
were over 0.02 ppm and slightly reduced in zones where tht
mean levels of SO2 were over 0.01 ppm. Similarly Skye (1964)
found that the survival of lichens was less in areas with an
annual SO2 concentration of approximately 0.015 ppm.
Chronic injury appears as a yellowing or chlorosis of
the leaf, sometimes from lower to upper surfaces on broad
leaves. Occasionally only a bronzing or silvering will occur
on the under surface of the leaves. The rate of metabolism is
reduced in leaves displaying chronic injury. Chronic injuries
develop slowly on coniferous perennial foliage, with the
greatest increase injuries in the Sudbury sulphur-fume
effects area occurring on the 1-year-old needles of eastern
white pine trees. Continued chronic injury to perennial
foliage of coniferous trees results in premature needle
abscission, reduced radial and volume growth, and early death
of the trees. Table 3 lists the SOp concentrations causing
chronic injuries.

2 9
TABLE 3
Responses to sulphur dioxide - chronic effects
Plant response References SO2 Cone. (ppm)
Exposure period
0.07 3 days
0,
0,
0,
0,
0,
0,
0,
.076
.05
.067
.035
.008
.015
.016
14 days
24 days
26 weeks
Anuual avr. (5 months growing period)
10 year avr. (5 m. growing season)
Annual avr.
Annual avr.
Injury to sensitive eastern white pine
Injury to barley
Injury to beech
Depressed yields of ryegrass of 52X
Injury to eastern white pine in field
Banfield (1972)
Mandl et a7. (1975) Keller (1976)
Bell and '"'•c t 9I1 (1973)
Roberts (1976)
Slight injury to old foliage of eastern white pine trees in forest
Reduction in lichens and bryophytes in field
Skye (1964)
Very low diveristy in Gilbert (1969) lichen and bryophyte species in the field
0.02 Growing season avr,
Epiphyte species drastically reduced in forest.
Le Blanc et a7, (1972).

3 0
S u b t l e e f f e c t s a re measured by p h y s i o l o g i c a l o r
b iochemica l changes, and/or r e d u c t i o n i n p l a n t g row th or
y i e l d i n t h e absence o f m a c r o s c o p i c i n j u r y . E a r l y
i n v e s t i g a t o r s s tudy ing the e f f e c t s of SOg on vege ta t ion in
Europe concluded t h a t i n v i s i b l e i n j u r y , o r p h y s i o l o g i c a l
d is turbances or e f f e c t s on the growth or y i e l d s of p lan ts
cou ld occur i n the absence o f v i s i b l e mark ings ( S t o k l a s a ,
1923). Th is theory was r e f u t e d by s e v e r a l •* r v e s t i g e f ' o n s
c a r r i e d out i n North America in the 1930s (H- 1 a id " HomHt
1933; Katz e t a7 . , 1939). These i n v e s t i g a t o r s found t h a t non-
mark ing c o n c e n t r a t i o n s o f SO2 e m i t t e d under c o n t r o l l e d
c o n d i t i o n s f o r long p e r i o d o f t i m e d i d no t a f f e c t ca rbon
d i o x i d e a s s i m i l a t i o n , s t o m a t a l b e h a v i o u r , c h e m i c a l
compos i t i on or the r a t e o f g rowth o f t he exposed p l a n t s .
Davis (1972) found no y i e l d loss from soybean unless there
were v i s i b l e man i fes ta t i on r e s u l t i n g from SOg fum iga t i ons .
Subt le e f f e c t s w i thou t microscopic v i s i b l e i n j u r y t o p lan t
t i s s u e s , have been repor ted i n other i n v e s t i g a t i o n s , Bennett
and H i l l (1974) found t h a t subnecrot ic p o l l u t a n t exposures
could repress pho tosyn the t i c ra tes . Table 4 l i s t s the r e s u l t s
o f exper iments conducted t o d e t e r m i n e t he o c c u r r e n c e o f
sub t l e e f f e c t s on p lan ts in the presence of c o n t r o l l e d SOg
doses. Table 5 d i f f e r e n t i a t e s t h e t h r e e t y p e s o f p l a n t
e f f e c t s (acute , chron ic and sub t l e ) caused by SO2•

3 1
TABLE 4
Responses to sulfur dioxide - subtle effects
SOg Exposure Plant response References Cone. period (ppm)
0.25 1 hr. 2% depression in COg Bennett and Hill uptake by alfalfa (1974)
0.5 1 hr. 2^% Depression in COg uptake by alfalfa
0.7 1 hr. Increased amino acids, Godzik & Linskens decreased protein synth- (1974) esis in bean.
0.71 1 hr. 55% inhibition of pollen Masaru et a7. tube length of lily (1976)
1.0 30 min Reduced chlorophyll in Constantinidou cotyledons of red pine et a7. (1976)
0.5 2 hr. Reduced chlorophyll in cotyledons and decreased dry weight of primary needles of red pine
Ambient 187 days Ryegrass grown in water Bleasdale(1973). air scrubbed air with
98-100% of SOg removed had 48% more dry weight than plants grown in polluted air.
0.067 26 weeks Decreased yields of Bell and Clough ryegrass by 52% (1973). (Chlorosis of leaves but no lesions)
0.05 31-60 Decreased photosynthe- Keller (1976) days sis in Scot pine
0.05 141 days Decreased pollen germination in white fir.
0.05 9 months Increased peroxidase activity in spruce needle homegenates.

TABLE 5
3 2
Differentiation between three types of plant effects caused by SOg
Acute effects caused by high concentrations of SOg during short-term exposure (less than 24 h)
Chronic effects caused by variable cone, of SOg.Usually over a long period of time.
Subtle effects caused by variable concentrations of SOo.
Necrotic lesions on foliage of the current year
Chlorotic markings on foliage, sometimes developing to necrosis, on perennial conifers usually on foliage older than current year
No foliar markings
Folair lesions appear within hours or days after exposure
Foliar injury usually May be possible to develops slowly over measure effects on a long period of time physiological
processes photosynthes i s, respiration, and transpiration; on biochemical processes-enzyme activities and chemical compositions; on pollen tube elongatin; or on pollen germination
May cause reduction in growth and loss in yield
Rarely result in plant mortality unless from recurring acute fumi gations
May cause reduction growth and loss in yield
May cause reduction in growth and loss in yield
May result in plant mortality from slowly developing injuries (especially on perennial conifers and lichens)
No plant mortality

3 3
Studies conducted in England repor t losses i n y i e l d s of
S23 ryegrass [Lolium perenne) reported t h a t ryegrass p lan ts
exposed to ambient a i r in one glasshouse weighed between 16
and 575S less than p l a n t s grown in s i m i l a r a i r i n a n o t h e r
glasshouse tha t had 98 to 100^ of the SOj removed by water
scrubb ing. Ke l l e r (1976) introduced the term ' l a t e n t ' f o r
sub t l e e f f e c t s and reported the resu l t s of long- term exposure
to a low concent ra t ion of 0.05 ppm SO2. The e f f e c t s inc luded
decreased photosynthesis of Scots pine ('^ je sy''^•est f ic],
decreased po l len germinat ion in white f i r {Abies alba), and
inc reased perox idase a c t i v i t y of need les homogenates o f
spruce iPicea excelsa). Godzik and Linskens (1974) found an
inc rease i n the t o t a l amount o f f r e e amino a c i d s and a
concurrent reduct ion in p ro te i n synthes is i n pr imary leaves
o f Widusa bean {Phaseolus vulgaris) when a r t i f i c i a l l y
fumiga ted w i t h 0.70 ppm SOg • Some o f t he e f f e c t s were
o b s e r v e d a f t e r o n l y 1 h o f f u m i g a t i o n and b e f o r e t h e
occurrence of v i s i b l e i n j u r y . Subtle e f f e c t s of SOp have been
repor ted on po l len tube e longat ion in v i t r o s t u d i e s .
Reiner t et al. (1980) found t ha t growth of soybean was
i n h i b i t e d by exposure to S02 at the concent ra t ion of 25 ppm,
when p lan ts were exposed f o r 4 h three t imes per; week f o r 11
weeks. The main e f f e c t of SO2 was a reduc t ion of shoot dry
weight at 7th week and t o t a l p lant growth a t 11th week. Kress
et al. (1986) exposed soybean p l a n t s t o i n t e r m i t t e n t SO2

3 ^
fumigations (4 h a day, 3 days a week) from shortly after
emergence untill maturity. About 7% yield losses due to 0.10
ul/L of SO2 in intermittent exposures was observed. Foliage
of many cultivars of soybean were sensitive to SOg, but the
amount of injury varied with cultivars. The point of attack
of sulphur dioxide in grasses (Benedict or Breen, 1955b;
Brennan et al., 1970; Bell and Clough, 1973; Cowling and
Lockyer, 1976; Bell, 1982) and in pines (Karpen, 1970; Berry,
1971; Roberts, 1976) is the tip of foliage w»'ich becomes
reddish brown followed by necrosis in contrast to the broad
leaved plants where the symptoms may appear anywhere on the
leaf.
Susceptibility and resistance of plants to SOg
Different plant species and varieties, and individuals
of the same species, may vary considerably in their
sensitivity or tolerance to SOp. Crop plants are classified
into three resistance groups according to their tolerance
limits. Clover-type fodder plants were most sensitive to SOg;
wheat, leafy vegetables (excluding cabbage), beans,
strawberries, and roses were moderately sensitive; and root
crops and cabbage were least sensitive. Ruston (1914)
reported that SOp caused a marked increased in soil acidity,
reducing the number of soil bacteria. Jones Eustace (1952)
observed the disappearance of lichens in heavily polluted
atmosphere which was another early report of disturbed plant

3 5
population due to air pollution. Miller et al. (1974) found
that foliage of many cultivars of soybean were sensitive to
SOrt but the amount of injury varied with the cultivar.
Reinert et al . (1980) found the growth of soybean was
inhibited by exposure to 25 ppm of SO2 when plants were
exposed for 4 h, three times per week for 11 weeks. The main
effect of SOg were a reduction of shoot dry weight at 7th
week and total plant growth at 11th week. SOg contributed to
the reduced growth in soybean in the absence of visible SO,-,
injury. Sprugel et al. (1980) observed that soybean plant
were relatively sensitive to elevated SO2 concentrations that
occurred near a point source. Reinert et al. (1982) found
that marigold shoot and root dry weights were decreased when
plants were exposed to 0.3 ppm SOg. It reduced dry flower
weight too. Sheng et al. (1988) exposed soybean cultivars to
0.7 ppm SOg which resulted in reduction in net photosynthesis
and stomatal conductance and all the cultivars developed
typical SO2 symptoms during or after fumigation.
Physiological and biochemical effects of SOg on plants
The manner in which SOp affects the metabolism of the
plant is not well understood. At the level of the plant, or
the plant leaf, the metabolic process of primary interest is
photosynthetic carbon dioxide fixation. Since 90 to 95% of
the dry weight of plants is derived from this process
(Zelitch, 1975), the impairment of photosynthesis has been a
popular explanation for plant growth reduction caused by SOg.

3 6
Different plant species have different capacities to remove
SO2 from the surrounding air (Jenes et a1., 1976). Lucerne
was found to be susceptible to SOg only if the stomata were
open. This has been confirmed by numerous investigations
since then and it is generally accepted that the primary
factor controlling SO2 uptake by plant leaves is the degree
of opening of the stomata. Generally, drought is considered
to be protective of the plant since the stomata close during
this condition. An initial increase in transpi rat .o- has .le
noticed during SOg exposure Weigl et al., 1962. Sulphur
dioxide may also react with chemical compounds making up the
cuticle and thus change the cuticular resistance. It has
been suggested that SO2 stimulates the production of waxes.
From scanning electron micrographs it cah be seen that SOg
alters the surface of confier needles. The inhibition of
photosynthesis is often regarded as the first stage of SO2
action on plants. In short-term fumigation the most sensitive
lichen species yet studied is Lobaria pulmonaria, whose
photosynthesis was inhibited after exposure to 0.5 ppm SO2
for 14 h. SO2 injury increases respiration rate of the
affected plants, the rate of respiration increases initially
in pine and spruce plants exposed to SO2.
Combustion of sulphur-containing fuels yields SO2 in
quantities 40 to 80 times that of SO3 (U.S. Dept. of Health,
Education and Welfare, 1969). Sulphur dioxide is oxidized in

3 T
the atmosphere catalytically or photochemically. The photo-
oxidation rate of SOg in air and sunlight is between 0.1 and
0.2% h. The SO3 molecules combine with water vapour in the
air to form sulphuric acid (H2S0^) aerosols. Acid smog injury
to vegetation observed in Los Angeles was described by Thomas
et a 7. (1952). Spots were formed on the upper surface of the
leaves which later extended throughout the leaf. In the
Scandinavian countries, the north-eastern United States, acid
rain has been found to have had serious implications with
respect to increasing the acidity of lakes and reducing f-'sh
populations. It has been postulated that decreased
productivity of forests in susceptible region in Sweden is
attributable to natural rainfall acidified by industrial
emissions. Wood and Bormann (1974, 1975) and Wood (1976)
reported foliar tissue damage of yellow birch (Setu/a
al leghaniensis) seedlings, using artificial acid mist of pH
3.0, and significant growth decrease at pH 2.3; increased
foliar leaching of potassium, magnesium, and calcium from
sugar maple (Acer saccharum) and pinto bean seedling using
artificial acid mists of pH 4.0, and tissue damage at pH 3.0;
and a 205K increase in growth of eastern white pine seedling
subjected to artificial acid rain at a pH of 2.3 compared to
rains with higher pH. However, soil acidity increased, and
magnesium, calcium and potassium cations were leached from
the soil at the low pHs.

3a
Sulphur dioxide after its absorption through stomata
accumulates into intracellular spaces. A large amount of SO2
can be absorbed by the cells of the leaf without any injury
if the absorption is slow and the concentration of the gas is
low. However, SO2 accumulates faster than it can be oxidised
and assimilated thus, exceeding the threshold level, and a
phytotoxic concentration presumably develops in the
intercellular spaces of the leaves, causing injury (Katz,
1952; Thomas, 1961). The excess SOg, accumulatea in ,n».
intercellular spaces, comes in contact with the water bathed
cell walls of the surrounding tissue to form sulphurous acid
and sulphates, leading to plasmolysis and collapse of the
cell (Brandt and Heck, 1968). Haselhoff and Lindau (1903)
believed that the gas reacts with aldehydes and sugars to
form secondary products which slowly release sulphurous or
sulphuric acid causing injury to the cell. However, the idea
could not last long, since the leaves are more sensitive to
SO2 in morning when the sugar content is low (Treshow and
Pack 1970). SO2 inactivates iron in the chloroplast causing
interference with its catalytic properties in assimilation.
This promotes secondary processes and break down the
chlorophyll, ultimately killing the cells. Dorries (1932)
observed that the absorbed acidity could decomposed
chlorophyll, liberating magnesium and forming pheophytin;
which may lead to chlorosis and reduced photosynthesis.
According to Thomas et al. (1943), the toxicity of sulphurous

3 9
ac id i s r e l a ted p r i m a r i l y t o i t s reducing p roper t i es ra the r
than t o i t s a c i d i t y because i t i s 30 f o l d s more t o x i c than
s u l p h u r i c a c i d . Excess of sulphate may i n t e r f e r e w i t h ion
a b s o r p t i o n , l e a d i n g t o d i s r u p t i o n i n n u t r i e n t b a l a n c e .
B leasda le (1952) found t h a t n o r m a l l y an e q u i l i b r i u m i s
mainta ined between su lphydry l group and more ox id ized su lphur
compounds such as s u l p h i t e s . Any i m b a l a n c e i n t h i s
e q u i l i b r i u m , as might be caused by an excess o f o x i d i z e d
s u l p h u r compounds, would upset s u l p h u r u t •»'"'• ?.t-c.- and
p r o t e i n syn thes i s . Sulphur d iox ide would reo-v^e t «= s u l p h i t e s
t o s u l p h y d r y l , caus ing an a c c u m u l a t i o n o f t he l a t t e r and
d i s t u r b i n g the r a t i o between su lphydry l t o ox id ized sulphur
compounds. The su lphydry l compounds thus becomes a products
o f a d i s t u r b e d su lphu r me tabo l i sm, and t h e t o l e r a n c e o f
p l a n t s t o SOg may be a f u n c t i o n o f t h e s t a b i l i t y o f
s u l p h y d r y l (Dekok et al., 1982) . Benne t t e t al. (1973)
observed t h a t r a te of uptake of gaseous p o l l u t a n t s i n t o a
l e a f depends on seve ra l p h y s i c a l f a c t o r s . Be fo re b e i n g
capable to cause i n j u r y w i t h i n the p lan t c e l l , the p o l l u t a n t
has f i r s t t o enter the s o l u t i o n in the e x t r a - c e l l u l a r water
conta ined in the c e l l w a l l . The simple s o l u b i l i t y in water i s
t h e r e f o r e , l i k e l y t o assume importance wh i le cons ider ing two
p o l l u t a n t s which d i f f e r markedly in s o l u b i l i t y and are taken
up a t d i f f e r e n t r a t e s by the p l a n t s (Law and M a n s f i e l d ,
1982). MacCormick (1968) observed tha t the e f f e c t of SOg in
" d a i l y c y c l e " i s more in the morning (7-10 A.M.) and i n the

-4-0
evening (7-10 P.M.). This diurnal change in sensitivity
corresponds to the diurnal changes of stomatal aperture in a
number of plants. During the "seasonal cycle" the effect of
SOg is more during winter than in summer (MacCormick, 1968).
Experiments under two light regimes depicting winter and
summer have proved that 505»i reduction in shoot dry weight was
obvious when SO2 fumigation accompanied winter lights
(Davies, 1980).
Nematode Parasites of Pulses
Pulses has been found to be attacked by a number of
nematodes and their pathogenicity have been proved (Gupta et
a/., 1986; Kinloch, 1980;2 Mujib et al., 1988; Patel et a/.,
1988: Chavda et al., 1988; Nath et al., 1979; Mani et al.,
1984; Rodriguez-Kabana et al. , 1981; Herman et al., 1988;
Routary et al., 1986; Wrather et al., 1988; Kinloch et al. ,
1987; Sobun et al., 1979; Dhanger and Gupta 1983; Srivastava
et al., 1974, Upadhyay and Swarup, 1981; Singh and Gill,
1990). The important nematodes which are usually found in
association with the roots of soybean and check-pea are
Heterodera glycines, Pratylenchus brachyurus, Tylenchorhyn-
chus spp., Tylenchus spp., He 1icoty1enchus spp.,
Hirschmanie 1 1 a spp., Roty1enchu 1 us reniformis, and
Meloidogyne spp.
Kinloch (1980) reported that the most widespread and
damaging nematodes in soybean production are Meloidogyne

A- 1
incognita ( K o f o i d & W h i t e ) C h i t w o o d and soybean c y s t
nematode, Heterodera glycines Ich inode, Rodriguez-Kabana et
al. (1981) observed tha t M. arenaria (Neal) Chitwood and M.
incognita p r e v a l e n t i n sandy s o i l s can s e v e r e l y l i m i t
soybean, Glycine max ( L . ) Mer r . y i e l d s . Wrather and Anand
(1988) found t h a t soybean seed l ing growth was i n h i b i t e d by H.
glycines i n f e c t i o n and soybean s e n s i t i v i t y t o the nematode
seemed t o d i m i n i s h w i t h the age o f p l a n t . K i n l o c h et al.
(1987) conducted a f i e l d t r a i l w i t h 39 soybean cu l t -var .9 and
5 breeding l i n e s a t s i t e s i n f es ted w i th M. arenaria. Y ie las
o f a l l c u l t i v a r s were too low t o j u s t i f y t h e i r p l a n t i n g in
s i t e s h e a v i l y i n f e s t e d w i t h M. arenaria. Herman et al.,
(1988) observed t ha t M. incognita suppressed shoot growth of
soybean even t h a t which was c l a i m e d t o be r e s i s t a n t t o
nematode.
Mani and Sethi (1984) s tud ied the pa thogen ic i t y of M.
incognita on ch i ck -pea c u l t i v a r Pusa-209 w i t h 5 i nocu lum
l e v e l s , namely, 0 .5 , 1.0, 2 .0 , 4 . 0 , and 8.0 larvae per gram
of s o i l . They found t h a t there was a progressive decrease i n
p lan t growth as the inoculum leve l of the nematode increased.
Nath e t al. (1979) s t u d i e d the e f f e c t o f M. incognita on
ch ick -pea. They found tha t increase in the leve l of l a r v a l
i n o c u l u m o f t h e nematode r e s u l t e d i n t h e p r o p o r t i o n a l
decrease in p l an t growth, f l o w e r i n g , f r u i t i n g and b a c t e r i a l
nodu la t ion i n ch ick-pea.

-4-2
Root-knot nematodes are one of the most destructive
plant parasitic nematodes with a wide host range and thriving
in a greatly diverse habitats. Gupta et al. (1986) screened
219 varieties / lines of mung bean for their susceptibility
to root-knot nematode, M. Javanica and found none as
resistant. Mujib et al. (1988) studied effect of M. incognita
on carpet legume (Dolichos Tablab v. Tignosus), They observed
an decrease in the length, fresh weight of shoot and root as
the inoculum density of M. incognita increased and
significant reduction in growth occurred with 1000 nematode
per 500 cc soil. Out of 60 cowpea lines/germplasm screened by
patel et al. (1988) for pathogenicity of mixed population of
M. javanica and M. incognita none was found to be resistant.
Chavda et al. (1988) studied reaction of 27 varieties/lines
of greengram to a mixed population of M. incognita and M.
javanica and found that all the tested cultivars were either
moderately or highly susceptible.
Pulses and Soil-borne Fungal Pathogens
Pulses are known to suffer from number of soil micro
organisms among which fungal pathogens causing wilt and root-
rot are of the main limiting factors in their production. The
most important ones causing severe damage to pulses are
Fusarium oxysporum f.sp. ciceris (Padwick) Chattopadhyay,
Rhizoctonia bataticola (Taub.) Butl. {=Sc]erotium bataticola
Taub, Macrophomina phased ina (Tassi) Goid.), Fusarium solani

^ 3
f . s p . pi si, F. eumartii, Pythium ultimum, Rhizoctonia sol ani,
Sclerotium rolfsii, Operculel la padwickii, Verticil Hum albo-
atrum, and Botrytis cinera.
Mani and Seth i (1985) founded f i v e p a t h o g e n i c f u n g i
v i z . Fusarium oxysporum f . s p . ciceris, F. sol ani f . s p . pisi,
Phythium ultimum, Rhizoctonia sol ani, and Macrophomi na
phaseol ina i n a s s o c i a t i o n w i t h r o o t - r o t s and w i l t o f
c h i c k p e a . Of t h e s e , F. solani i s r e p o r t e d t o cause
a p p r e c i a b l e l o s s t o c h i c k - p e a under D e l h i c o n d i t i o n s .
I n i t i a l l y the diseased p lan ts showed ye l lowing of leaves and
the s ide shoot s t a r t e d dry ing and showed c h a r a c t e r i s t i c brown
d i s c o l o r a t i o n . Uproo t ing o f such p l a n t s , showed brown
co loured , i r r e g u l a r t o round les ions on the c o l l a r reg ions .
In advanced stages, the c o l l a r got const ra ined and the p l a n t
t a p p l e d down. B h a t t i et al. (1987) observed t h a t i s o l a t e s
from w i l t e d Cicer arietinum p lan ts included Fusarium spp.
(42%), R. solani (28%). Macrophomi na phaseol ina (1756), and V.
albo-atrum (4%). Westerlaud et al. (1974) observed t h a t F.
solani f . s p . pisi, and F. oxysporum f . s p . ciceris causes
s i m i l a r ye l low ing and w i l t i n g of the chickpea shoots, but the
former caused d i s t i n c t i v e black roo t les ions and the l a t t e r ,
vascu la r d i s c o l o r a t i o n e x t e n d i n g t o the t op o f t he s h o o t .
They observed t h a t F. oxysporum f . s p , ciceris may requ i re
wounding fo r e f f i c i e n t i n f e c t i o n , but F. solani f . s p . pisi
does not . I t was found tha t F. oxysporum f . sp. ciceris was

A-A-
not seed-borne. There were evidences which showed F. solani
f. sp. pi si was seed borne.
W i l t o f ch ick-pea which i s caused by several s o i l -
borne fungal pathogens inc lud ing F. oxysporum f. sp. ciceris,
Sclerotium rolfsii, Rhizoctonia bataticola (. Macrophomina
phaseolina), and Opercalella padwicki has been repor ted by
Kotasthane e t al. (1980) t o be a l i m i t i n g f a c t o r i n i t s
p roduct ion in Madhya Pradesh. M o r t a l i t y i n the e a r l y seedl ing
stage was c h i e f l y due to S. rolfsii wh i le a t f l owe r i ng F.
oxysporum f . s p . ciceris was the major pathogen. Alvarez and
B r i n g e r (1987) i s o l a t e d F. solani f r o m c h i c k - p e a p l a n t s
showing reduced g row th , l e a f y e l l o w i n g , d e f o l i a t i o n and
premature death. There were c o l l a r and roo t l e s i o n s , but roo t
vascu lar d i s c o l o r a t i o n . Trapero-Casas and Jimenez-Diaz (1985)
i n a disease survey in 1979-81 observed t h a t ch ick-peas in
southern Spain were severely a f f ec ted by a w i l t and r o o t - r o t
complex. Symptoms included vascular w i l t or y e l l o w i n g , non
vascular ye l l ow ing , c o l l a r and r o o t - r o t s or c o r t i c a l c o l l a r
and r o o t n e c r o s i s , and ye l low s t u n t . They f o u n d t h a t F.
oxysporum, F. soTani and M. phased ina were associated w i th
the w i l t and r o o t - r o t complex. Salmon-pigmented i s o l a t e s of
F. oxysporum induced vascular w i l t or ye l l ow ing , and redd ish-
p i g m e n t e d i s o l a t e s i nduced n o n - v a s c u l a r y e l l o w i n g and
c o r t i c a l c o l l a r and roo t necros is . F. solani and F. eumartii
induced non-vascu lar y e l l o w i n g and b l a c k c o l l a r and r o o t

-4-5
rot, M. phaseolina induced dry collar and root.~ro't.. Si may
(1989) detected Botrytis cinerea from chick-pea seedlings
showing pod and seedling-rot symptoms. Wilt of chick-pea
caused by F. oxysporum f. sp. ciceris for the first time in
India was observed in 1940 by Padwick (1941), which causes
about 10% losses in yield annually (Singh and Dahiya, 1973).
Murumkar and Chavan (1985) observed that infection of
chick-pea by F. oxysporum f.sp. ciceris resulted in reduction
in chlorophyll and increase in organic acids, polyphenols and
carbohydrates. Toxins isolated from culture filtrate of
chick-pea wilt pathogens F. oxysporum f.sp. ciceris by
Kaur et a1. (1987), caused browning and inhibition of callus
growth, and reduced free proline and soluble protein content
of cell. Wilt resistant varieties were less sensitive to
toxin. Toxin did not result in bursting of protoplast.
Primary site of action of toxins was inhibition of RNA
synthesis. Murthy and Bazyaraji (1980) observed that the
root nd shoot protein of resistant varieties contained
higher amounts of total alkaloids than the susceptible
cultivar. Alkaloids were fungitoxic and accumulated at the
site of wound,
F. oxysporum f. sp. ciceris and, at a lower rate, F.
solani f.sp. pi si were detected inside chick-pea seeds. F.
oxysporum f. sp. ciceris reaches seeds by systemic pathway
varying for each studied line, while F. solani f.sp. pisi

was traced only to the stem and plant collar (Conci et a/.,
1985).Sharma and Gupta (1986) noted that chlamydospores of
F. oxysporum f.sp. ciceris remained viable throughout the
high temperature of the summer month during the monocropping
period in naturally infected roots of Cicer arietinum at soil
depths of 5,10 and 15 cm. The fungi did not survive in roots
placed on the soil surface.
The ubiquitous sclerotial fungus Rhizoctonia bataticola
in its pycnidial state is known as Macrophomina phaseolina.
The fungus causes complex disease syndromes like charcoal rot
of stem, root-rot, seedling blight, foliage blight, tuber
decay, dry rot, fruit rot, pod nd seed rot in several crops.
In absence of host plant, it survives or over seasons
predominantly as small black sclerotia in diseased plant
parts or in soil. /?. t>atat7co7a from different host species
differ in their morphological and cultural characters and
even differences occur in the isolates from various parts of
the same host (Jain et a7., 1973; Dhingra and Sinclair,
1973; Ghosh and Sen, 1973; Grover and Sakhuja, 1981; Hooda
and Grover, 1982; Byadgi and Hedge, 1985). M. phaseolina is
an important pathogen of many hosts like lima bean (Andrus,
1938), Jute (Varada Rajan and Patel, 1946), black gram (Tarr,
1954), sesame (Meiri and Solel, 1963), soybean (Gangopadhyay
et al. , 1970) and sweet potato (Stevens et al. , 1987). The
location of pathogen in the seeds decides the disease

-4-7
severity. Firm establishment of the pathogen within the seed
tissue is pre-requisite for its effective transmission.
Widespread losses at different stages of soybean growth
are caused by species of Col 1etotrichum, Rhizoctonia,
ScTerotium, Macrophomina, and Fusarium (Perera, 1985). Sharma
and Bhowmik (1987) observed that M. phaseoT i na (=f?.
batati col a) infection caused both quantitative and
qualitative damage to groundnut seeds. It rerultcc' •'. r
characteristic discoloration of pod and kernel, anc a mar»•
reduction in pod and kernel yield, shelling percentage and
oil content. Prinsloo (1986) noted that M. phaseolina caused
symptom of black root-rot on chicory only when roots were
tooth-pick-inoculated in the laboratory. M. phaseolina caused
symptoms only when the roots were injured by Thielaviopsis
basicola. All the cotton cultivars test by Lee et al. (1986)
were susceptible to isolates of M. phaseolina from Phaseolus
vulgaris, cotton and groundnut. The groundnut isolate (M14)
was the most virulent on cotton and mechanical wounds were
not necessary for infection. The stele portion of the tap
root and pith in the lower portion of the main stem and root
were discolored, varying from rust-brown to black. Sclortia
of the pathogen were found in the pith and the organism could
be isolated from the stele. All the soybean cultivars tested
by Garica et al. (1985) were found to be susceptible to
Sclerotium r o l f s i i , Fusarium spp., R. solani, and M.

-4-3
phaseol ina. Anahosus and Naik (1986) observed that M.
phaseolina colonizes in root and stalk of sorghum leading to
disintegration of their tissues.
Isolates of M. phaseolina obtained from different plant
species differed in cultural characters, growth rate,
morphology of the sclerotia, pycinidia and pycinidiospores,
and virulence. Bean, chick-pea and cowpea isolates were the
most virulent (Byadgi and Hegde, 1985). Raut and Ingle (1989)
observed variation between the isolates of R. batat ico'
collected from different cultivated crops belonging to seven
families. Differences were abundant even between the isolates
of crops belonging to same family. Chitima et a1. (1987) also
observed differences in morphology, growth rate and
pathogenicity among isolates of M. phaseolina and also to
some extent between individual conidia originating from a
single pycnidium.
Prasad (1986) concluded that incidence of root and stem
rot disease of jute caused by M. phaseolina was inversely
proportional to K levels. Maximum disease was observed at 0
Kg Potash/ha. and minimum of 70 Kg potash/ha. In pot culture
experiments conducted by Sharma and Chaudhary (1985) it was
observed that application of 120, 80 and 40 Kg/ha. of N, P2
Og and Kg 0 respectively were the best for the reducing
mortality of cauliflower seeding suffering from wilt and root
rot caused by R. solani and R. bataticola {M. phaseol ina).

^ 3
Losses caused by s o i l - b o r n e f u n g i , p a r t i c u l a r l y M.
phaseol ina and F. oxysporum f . s p . vasinfectum, were
est imated a t cost $ 481593 over 27000 ha. of co t ton i n 1982
(Delgado and Agurto, 1984).
In teract ion of Sulphur Dioxide and Plant Parasites
Ef fec t s of SOg on the r e l a t i o n s h i p between p lan t host
and v a r i o u s p l a n t pathogens have been examined bo th i n
ambient cond i t i ons and a r t i f i c i a l t reatments . Hegale (1973,
1982) Treshow, (1965) summarized the interact ions- oetween
severa l a i r p o l l u t a n t s i nc l ud ing sulphur d i ox ide , and hos t -
p a r a s i t e r e l a t i o n s h i p s .
The re a r e a number o f p o s s i b l e h o s t - p a r a s i t e
i n t e r a c t i o n e f f e c t s t h a t may occu r when a p o l l u t a n t i s
i n t r o d u c e d i n t o a p a r t i c u l a r h a b i t a t . The s e v e r i t y o f t h e
p a r a s i t i s m may be increased or decreased as a r e s u l t o f the
a c t i o n o f the p o l l u t a n t on the v i ru lence of the pa ras i t e or
on the s u s c e p t i b i l i t y of the hos t .
Fungal Plant Pathogens and Sulphur Dioxide
E f f e c t s o f SOg on f u n g a l d i s e a s e o f p l a n t s a r e
summarised in Table 6. Fungi may be a f fec ted by SO2 d i r e c t l y
or i n d i r e c t l y through some e f f e c t upon l i v i n g or dead p lan t
t i s s u e s , s o i l , o r w a t e r . S u l p h u r d i o x i d e r e s u l t s i n
a c c u m u l a t i o n o f s u l p h u r , i n c r e a s e d a c i d i t y and o t h e r
a l t e r a t i o n s in p lan t physio logy and b iochemist ry .

TABLE 6
Interaction of sulphur dioxide and plant pathogenic fungi
5 0
Fungal plant pathogens Host Dose
Puccinia graainis Wheat ftsbient
Effect Reference
Johanson,i954
Scheffer and HedgcQck, 1955
Scheffer and Hedgcock, 1955
Kundela and Novakovj. 1962
Skye, 19tn'
Scheffer and Hedgcock, 1955
Jancarik, 1961
Chiba and Tanaka, 1968
Sevan and Sreenhalgh, 1976
Linzon, 1958; Scheffer and Hedgcock, 1955 Sharp, 1967
Oonaubauer, 1968 Jancarik, 1961; Kundela and Novakova, 1962
Linzon, 1958; Scheffer and Hedgcock, 1955
Puccinia sp.
Hypodersella sp.
Lophoderaiua piceae
Hysteriue pulicare
Helaapsora sp.
Traaetes serialis and r. heteroaopha
Trees Aabient
Aabient
Aabient
Alder Aibient and Brich
Atbient
Trees Anbient
Rhiiosphaera kalkhoffii Red pine Aabient
Rhytisaa ocerinua
Cronatriua sp.
Puccinia striiforais
Sycaiarea Aabient
Trees Aibient
Atbient
f^raillaria aellea Trees Aabient
Coleosporiua sp. Trees Aabient
Less disease incidence
Decrease in disease incidence
Decrease in disease incidence
Increase in disease incidence
Decrease in disease incidence
Increase in disease incidence
Increase in disease incidence
Increase in disease incidence
Decrease in disease incidence
Decrease in disease incidence
Decrease in geraination of uredospores
Increase in disease incidence
Decrease in disease incidence

Table 6 Contd.
5 -I
Fungal plant pathogens Host Dose Effect Reference
Uroayces phaseoli Bean 0.13 ppi for 24 h/d Decrease in pustule Weidensaul (on 8 days before or nuiber, spore size and Darling, 1979 7 days after inocula- spare size and geri-tion) ination
Scirrhh acicola Scots 0.20 ppi for 6 h 5 pine days after inocula
tion
Increase in lesion Weidensaul nunber and Darling, 1979
S. acicola
Puccinia graainis
ledius
Mheat
1.0 ppa for 4 h Nortal groHth Hai, 1971
0.10 ppa for 100 h Decrease in lesion Laurence et ah, froB 2 days frot nuiber 1979 inoculation
mternaria sp. 50 ppa for 2 ainutes 60^ decrease in Couey, 1965 at aoist condition of spore geraination 100 ppa for 24 ainutes at 981; RH
Oiplocarpon rosae Rose 0.01 and 0.04 ppa for Diseased leaflet area Saunders, 1966 2 days after inoculation slightly increased at
at 0.01 ppa Bore than the 0.04 ppa
Penicilliua sp.
Helaintho sporiua eaydis Maize
90 ppa SO, in nutrient Slight stiaulation in Saunders, 1966 solution groHth
0.15 ppa for 14 h/d 8 Decrease in lesion Laurence et ah, days before inoculation nuaber 1979
nicrosphaera alni Lilac 0.30-0.40 ppa for 24-72 Decrease in conidial Hibben and

5 2
Genera l l y , excess o f SO2 i n the atmosphere t ends t o
d e c r e a s e t h e s e v e r i t y o r i n c i d e n c e o f h o s t - p a r a s i t e
r e l a t i o n s h i p . Rus t d i s e a s e s appear t o be p a r t i c u l a r l y
s e n s i t i v e to SOg. Johanson (1954) reported less wheat stem
rus t caused by Puccini a graminis i n an i n d u s t r i a l i z e d area of
Sweden t h a n i n non i n d u s t r i a l i z e d a r e a s . S c h e f f e r and
Hedgcock (1955) found decrease of a number of rus t diseases
caused by species o f Cronartium, Coleosporium, Melampsora,
and Peridermium, where t rees were i n j u red by SO2. We-ipctein
et al. (1975) found t h a t SOg reduced the i n c i d e n c e and
s e v e r i t y of bean r u s t disease caused by Uromyces phasedi on
p i n t o bean. Continuous exposure t o 0.13 ppm of SOg on the 8
days before i nocu la t i on or on the 7 days a f t e r i n o c u l a t i o n
caused f o l i a r i n j u r y and decreased the number of pus tu les and
the s i z e and percentage g e r m i n a t i o n o f spores produced on
exposed leaves. In genera l , ob l i ga te fungal p lan t pathogens
are more sens i t i ve t o SO2 as compared to other fungal p l a n t
p a t h o g e n s . Fungal d i s e a s e s on p l a n t f o l i a g e have been
repor ted to be reduced in incidence by atmospheric SOg i n the
v i c i n i t y of i ndus t r i es in several coun t r i es . These lea f fung i
i n c l u d e s HypodermeT 1 a laricis, Lophodermium pinastri,
Hypodermella sp. (Schef fer and Hedgcock, 1955), Lophodermium
Juniperi, Rhytisma acerinum (A i r p o l l u t i o n , 1968), Hysterium
pulicare (Skye, 1968), and Venturia inaequalis ( P r z y b y l s k i ,
1967). Kock (1935) repor ted the absence of oak powdery mildew
caused by Microsphaera alni near a paper m i l l in A u s t r i a .

5 3
Saunders (1966) observed that rose black spot disease caused
by Diplocarpon rosae was rarely present in areas where the
daily average SOg concentration was greater than 0.04 PPM.
Khan and Kulshrestha (1991) studied effect of SO2 on conidial
germination of eight powdery mildew-fungi species viz.,
Sphaerotheca fuliginea (Schlecht. ) Poll., S. cassiae Pandotra
& Ganguly, Erysiphe cichoracearum D C , E. trifolii Grev., E.
pisi D C , Microsphaera alphitoides Griff & Maubl . f. sp.
zizyphi and PhyHactinia dalbergiae Pr ioz. They observed
that all the conidia tested were sensitive to SO2 and its
germination was increasingly inhibited with an increase in
the concentration of SOg.
Effect of SO2 on parasitism by non-obligate fungal
pathogens show cases of inhibition, stimulation, or no
response. Exposures that decreased parasitism of bean by
Uromyces phaseoli did not affect parasitism of tomato leaves
by Alternaria soTani. Laurence et al. (1979) observed
decrease in number of Helminthosporum maydis lesion by 38%
when maize plants were exposed to SO2 on the 8 days before
inoculation (0.15 ppm for 14 h daily) if the exposure
occurred on the 8 days before and on the 2 days after
inoculations, the number of lesions decreased by only 13-16%,
Spores of most fungi tested appear to be very resistant to
direct exposure to SO2 and moist spores apparently are more
sensitive to than the dry ones. Couey and Uota (1961)

5-4-
observed that only 20* of the conidia of Botrytis cinerea
germinated when they were exposed to 36 ppm SO2 for about
1/2 h. But Hibben (1966) observed no effect on germination of
spores of 10 saprophytic and parasitic fungi when they were
exposed on agar to 10 ppm SO2 for 1-6 h. Couey (1965) found
that germination of wet conidia of ATternaria sp. decreased
60X from exposure to 50 ppm SO2 for 24 min but 100 ppm SO2
was required to produce a similar decrease in the dry
spores at 985K RH. Conidia of Di plocarpon rosae g' rmi n^ted
abnormally and hyphal growth was reduced in aqueoL SJIS. e
solutions that contained the equivalent of more than 35 ppm
SO2. Spores of other fungi such as Aspergillus niger and
Alternaria brassicicola were also affected but were more
resistant than D. rosae (Saunders, 1966). Germination of
Scirrhia acicola conidia suspended in water was not affected
by exposure to 0.90 ppm SOg for 6 h (Ham, 1971). Khan and
Kulshrestha (1991) found that SO2 exposure inhibited conidial
germination of a number of powdery mildews. SO2 concentration
and exposure duration were determinants of inhibition.
Some researchers have found that SOg can increase the
severity of foliar diseases. Tanaka et al. (1982) reported
that sooty leaf mold disease, Rhizosphaera klkhoffi i, was
more severe on Pinus densiflora trees that were inoculated
and transplanted to areas with high concentrations of
atmospheric SOg than on trees in areas with no air
pollution. Weidensaul and Darling (1979) observed more

5 5
les ions of Scirrhia acicola on needles of scots pine seedings
when they were exposed t o 0 .20 ppm SO2 a t 5 days a f t e r
i n o c u l a t i o n t h a n on unexposed p l a n t s . The e f f e c t was
s i m i l a r , but not s i g n i f i c a n t , when the 6 h. exposure ended 30
min . before i n o c u l a t i o n .
P lan t P a r a s i t i c Nematodes
The e f f e c t o f s u l p h u r d i o x i d e on h o s t - p a r a s i t e
i n t e r a c t i o n s i nvo l v ing roo t -kno t nematodes, has gained l i t t l e
s tudy. But, there are few evidences i n d i c a t i n g the e f f e c t of
a i r p o l l u t i o n on p lan t p a r a s i t i c nematodes (Table 7 ) . Bassus
(1968) found occurrence o f fewer numbers bu t a g r e a t e r
v a r i e t y o f nematode species i n s o i l s where t rees were damaged
by SOg. Khan (1989) s tud ied the e f f e c t of SOg, O3 and SOg-Og
m i x t u r e o f d i f f e r e n t c o n c e n t r a t i o n s on t o m a t o p l a n t s
inocu la ted w i t h M. incognita race 1. He observed t h a t SO2 and
O3 (both p a r t i c u l a r l y at 0.2 ppm) and M. incognita, ac t i ng
alone s i g n i f i c a n t l y reduced p lan t growth and y i e l d parameters
and lea f pigment content of tomato. Synergism was observed
when p lan ts were exposed t o 0.2 ppm SO2 and 0.1 or 0.2 ppm Og
and g r e a t e r r educ t i on was observed than t h e sum o f t h e i r
i n d i v i d u a l e f f e c t s . SO2 and O3 a l s o i n d i v i d u a l l y a c t e d
s y n e r g i s t i c a 1 1 y w i t h t h e nematode and caused g r e a t e r
reduc t ion i n growth parameters than the reduc t ion in nematode
inocu la ted unexposed p lants or un inoculated exposed p l a n t s .
When tomato p lan ts inoculated w i t h M. incognita race 1 were

5 6
IABLE-7
Effect of air pollution on pltnt parasitic neaatodes
Treataent
SO2
SO2
SO2
Oj and
OT and
0, and
Oj and
SO2 ' h
Oj and
0, and sBj + O3
0.2 ug O3/ lit., 0.2/ ug S02/lit. singly or in coabination
0.2 ug Oj/L + 0.8 ug SOp/L
80 ppB O3
Neaatode
Pratylenchus penetrans
Heleidogyne incognita Race 1
fi. incognita & M. javanica
Heterdora glycines
Heterodera glycicss^Rhizobiut
Beloaolaiaus longicaudatus
Paratrichodorus tinor
8. longicaudatus ^Rhiiobiut
Aphelenchoides fragariae
P. penetrans
P. penetrans
H. hapla
Host
Soyabeen
Toaato cv. Pusa Ruby
Lentil, chickpea
; Soyabeen
Soyabeen 1
Soybeen
Soybeen
Soyabean
Begonia
Toaato
Tomato
Tobacco
Effect
Enhanced reproduction of neaatode
SO2 interacted synergis-tically Mith the neaatode
Root galling suppressed
Inhibition in proouctio! and developaent or nesatode
Severe inhibition in nodulation
Unaffected
Reproduction and developaent of neaatode inhibited
Inhibition in nuaber of nodules
Decreased folior injury by neaatode
Synergistic interaction
ftntagonistic interaction
Increase in disease
References
Weber et al.,
Khan, 1989
Singh, 1989
Meber ai.,
Weber et al.,
Neber et al.
Meber et al.
Weber et al.
Weber et al.
Shew et al..
Shew et a l ,
1979
' ] -
, 1979
, 1979
, 1979
, 1979
, 1979
1982
1982
Bisessar and Plaaer 1984

Table 7 Contd.
5 -7
Treattent Nesatode Host Effect References
0.2 ug Oj/L + 0.8 9. penetrans ug SO2 L
Tosato Antagonistic interaction Shew et a l . , 1982
80 ppB Oj (anbient) H. hapla
Simulated of pH 6.0 tines Heel
h SO3+O3
SO2+O3
acid rain or 3.2, 3 dy
t
H. hapla
fi. incognita
H. incognita Race 1
H. incognita
Acid rain at pH 6.6 H. incognita before inoculation Race 1
Acid rain at pH 3.2 H. incognita after inoculation Race 1
Acid rain at pK 5 and 6
H. incognita H. javanica
Tobacco Increase in disease incidence
Kidney Decreased neiatode bean infection of pH 3.2
Bisessar and Pla«er 1984
Shriner, 1978
Lentil Root galling suppressed Singh, i
To«ato Growth of neeatode reduced Khan, 1989
Chickpea Juvenile hatching Singh, 1989 suppressed
Totato Root penetration and Khan, 1989 reproduction increased significantly
Toiato Root penetration and Khan, 198<) reproduction decreased significantly
Lentil Juvenile hatching Singh, 1989 chickpea suppressed

se
exposed to SOg + O3 mixture at 0.2 + 0.2 ppm or 0.2 + 0.1 ppm
of each pollutant respectively, synergistic reduction in
growth of nematode was observed. Hatching of juveniles of M.
incognita and M. Javanica was suppressed by exposing to SOg,
O3 and their mixture. Higher concentration of pollutants were
more suppressive than the lower concentrations, and the
effect of pollutant mixture was greater than their individual
effects. M. incognita was more sensitive to pollutant mixture
than M. javanica. Juvenile hatching of M. incognita and M.
Javanica were adversely affected by the acidity of the media
and decreased with the increasing acidity. At pH 6, greater
suppression in juvenile hatching of M. incognita was observed
whereas in acidic range (from pH 5 onwards) the suppression
in juvenile hatching of M. javanica was more than M.
incognita (Singh, 1989). Khan (1989) observed a synergistic
interaction between M. incognita race 1 and simulated acid
rain at pH 3.2 on tomato plants. They caused greater
reduction on tomato plants than the sum of reductions induced
by each individually. Root penetration and production of M.
incognita race 1 and disease intensity were significantly
increased at pH 6.6, but at pH 3.2 significant decrease were
recorded.
Weber et al. (1979) observed that non-injurious doses
of SOp failed to affect adversely four nematode parasites of
soybean viz., Pratylenchus penetrans, Be 1 one 1 aimus

5 9
longicaudatus, Heterodera glycines, and Paratri chodorus
minor, o r one nematode p a r a s i t e i n l e a v e s o f b e g o n i a ,
Aphelenchoides fragariae. However, r e p r o d u c t i o n o f P.
penetrans was increased when soybean p lants were exposed to
SO2 on the 4 days before i nocu la t i on and on 3 days per week
f o r 13 weeks a f t e r i nocu la t i on (0.25 ppm fo r 4 h d a i l y ) . This
dose of SOp was not enough to i n j u r e leave or to a f f e c t p l an t
g r o w t h . Exposure of soybean p l a n t s t o SOg enhanced t he
r e p r o d u c t i o n o f P. penetrans compared w i t h t h a t i n p ' 'an ts
exposed t o the charcoal f i l t e r e d a i r c o n t r o l . Brewer (1979)
observed t h a t r e p r o d u c t i o n o f P. penetrans on tomato was
s l i g h t l y suppressed by O3 a t 0.2 ppm but not by SOg a t 0.80
ppm (3 h/day on 2 days per week, f o r 8 weeks). However, the
mix tu re o f 0.20 ppm of O3 and 0.80 ppm of SOg caused
increased reproduc t ion of P. penetrans. Shew et a7. (1982)
conducted a s e r i e s of experiments to understand the poss ib le
i n t e r a c t i o n of P. penetrans and Oo and SOg on tomato p l a n t s .
Tomato p lan t s c u l t i v a r 'Wa l te r ' were inoculated w i th i n i t i a l
popu la t ion d e n s i t i e s of P. penetrans ranging 0-4000 nematodes
per po t and were repeated ly exposed f o r 3 h t o OQ doses
ranging 0.0 - 0.4 u l / 1 of a i r (1 ul O3/I of a i r = 1.960 ug
Og/m of a i r ) . I n another e x p e r i m e n t s , tomato p l a n t s ,
un inocu la ted or inocula ted w i th P. penetrans were exposed (4
h per exposure) 15 times to 0.2 ul O3/I of a i r , or 0.2 ul
SOg/l (1 ul SOg/l of a i r = 2620 ugSOg/m"^ of a i r ) , or bo th ,
or were exposed (3 h per exposure) to 0.2 ul O3 / I of a i r or

eo
0.8 U1S02/1 of air or both. Exposures to charcoal-fi1tered
air served as control. Decrease in dry weights of plant
parts excised from tomato plants exposed to 0.2 ulOg/l of air
added to the decrease in dry weight, caused by exposure to
SO2 at 0.2 ul/1 of air adequately predicated the decrease in
dry weight of tomato plants caused by exposure to 0.2 ulOg +
0.2 ulSOg per liter of air. When 0.2 UIO3 and 0.8 ulSOg per
liter of air were present in mixture, they acted
antagonistically and caused less change in leaf and shoot dry
weight than could be predicated by the main affects of 0^ and
SOg.
Plant Pathogenic and Nodule-forming Bacteria
Laurence and Aluiso (1981) observed that SO2 effects
the parasitism of bacteria. Table 8 shows the influence of
SO2 on some plant bacterial pathogens. It has been observed
that Corynebacterium nebraskense was inhibited when maize
plants were exposed continuously to 0.20 ppm SOg for 24
h/day, 5 days before inoculation, 2 days after inoculation
and both pre and post inoculations. However, maximum
inhibition of bacteria occurred at 2 days post-inoculation
exposure. They also observed that Xanthomonas phaseoli was
inhibited when soybean plants were exposed the 0-10 ppm SO2
for 24 h 5 days before inoculation, 5 days after inoculation
and both 5 days before and after inoculation exposures.

6 1
TABLE
Interaction of sulphur dioxide (SO.) and plant pathogenic bacteria and viruses.
Treatient Bacteria/Viruses Host Effect Reference
0.20 ppi for 24 h/day 5 days Corynebacteriut nebcaskense before or 2 days after inoculation or both
Haize Inhibitory Laurence and Aluisio, 1981
0.10 ppa for 24 h/day for Xanthoeonas phaseoli 5 days before or 5 days after inoculation
Soybean Inhibitory
0.10 ppn for 24 h for 7 days Southern bean aosaic virus Bean
Laurence and Aluisio, 1981
Increase in virus Laurence et a h , 196 conceni ratio'-
0.10 ppii for 24 h for 7 days Maize d«arf mosaic i/irus Maize IncrfcaSfc id \iruii taurciice ii, a;., i9S concentration, infection and severity of syaptoas
61

6 2
Sulphur dioxide increases the soil acidity which can
conceivably alter the progress of plant-parasite
interactions. Shriner (1974) found that kidney been and
soybean exposed over 8 weeks in the field or 5 weeks in the
greenhouse to pH 3.2 developed an average of 75X fewer
nodules than plants exposed to pH 6 treatments. Overall, 74%
of the plants that failed to nodule at all had received pH
3.2 treatments. In a time-related study, nodulation was
inhibited most by daily exposure to pH 3.2 treatments applied
15-21 days after plants were inoculated with Rhizobium.
Interaction of Sulphur Dioxide with other Air Pollutants
In nature, plants are rarely exposed to the influence
of only one air pollutants. Before 1966 some observations and
experiments had indicated that air pollutant mixture might
influence plants differently from the action of a single
pollutant.
Menser and Heggestad (1966) found that three varieties
of tobacco plant suffered from 25 to 38% leaf damage upon
exposure to a combination of 0.24 ppm SO^ and 0.027 ppm Og
for 2 h., whereas either pollutant alone at approximately the
same concentration and for the same time period caused no
injury to any of the three tobacco varieties. They suggested
that the injury to the tobacco plants was caused by a
synergistic effect of SOg and O3. The potential harm that may

6 3
be caused by mixture of low levels of SO2 and NOg was
demonstrated by Tingey et a7. (1971). A gaseous mixture of
0.10 ppm SO2 and 0.10 ppm NOg caused syngergistic effect with
greater than 5% leaf injury being induced on five of six
plant species treated in 4 h exposure periods. Bel W-3
tobacco plant displayed S% leaf injury after a 4-h exposure
to a mixture of 0.05 ppm SOg and 0.10 ppm NOg. Tingey et al.
(1973b) found that foliar injury on broccoli showed an
additive response to a mixture of 0.25 ppm SO^ • MG C 'O po-r.
O, for 4 h, whereas Bel W-3 tobacco showed & synergistic
response under the same regime. Low concentrations of SOg
and/or O3 to injured foliage of peanut plants in controlled
experiments. The response to a mixture of the two gases was
synergistic. Either 0.35 ppm SO2 alone for 3 h or 0.05 ppm O3
alone for 3 h injures the foliage of trembling aspen, a
combination of 0.20 ppm SOg and 0.05 ppm O3 for 3 h causes
more injury than the sum of the individual pollutants. White
et al (1974) reported a synergistic response of alfalfa to a
mixture of 0.15 ppm SO2 and 0.15 ppm NO2 in a 2-hr exposure
which depressed the apparent photosynthetic rate by 1%,
whereas 0.25 ppm of SOg alone or 0.40 ppm NO2 alone was
required to cause a 2% depression.
Mandl et al. (1975) studied effect of SO2, HF alone and
in mixture on barley, sweet corn, and bean plants. A
synergistic response was found to a mixture of low

6 4
concent ra t ions of two gases. Barley leaves d isp layed greater
i n j u r y in a mixed gas experiment than the sum of the i n j u r i e s
caused by the i n d i v i d u a l p o l l u t a n t i n a 27-day exposure
p e r i o d . The concent ra t ions of the po l l u tan t s u t i l i z e d were
approximately 0.08 ppm SO2 and 0.6 ug/m HF. I n t e r a c t i o n o f
su lphur d iox ide w i t h other p o l l u t a n t s on soybean are shown in
Table 9.
I n t e r a c t i o n of Nematodes w i t h o ther S o i l - b o r n e P l a n t
Pathogens
The number of s o i l microorganisms present in zhe •oo t
zone o f any crop p l a n t p rov ide s u f f i c i e n t o p p o r t u n i t y f o r
d i v e r s e i n t e r a c t i o n s . M a j o r i t y o f r o o t d i seases have a
complex e t i o l o g y (Grogan, 1981; Wal lace, 1978). Microorganism
i n the root zone may e x h i b i t cond i t ions l i k e synergism and
antagonism and may i n t e r a c t w i t h each o t h e r and w i t h t h e
hos t . Fawcett (1931) s ta ted t h a t "nature does not work w i t h
pure c u l t u r e s " and t h a t most p lan t diseases, p a r t i c u l a r l y
roo t d iseases, are in f luenced by associated microorganisms.
Root i n f e c t i o n by one pathogen may modify the host response
t o subsequent i n f e c t i o n by pathogens or saprohytes.
The t e r m i n t e r a c t i o n i s used q u a n t i t a t i v e l y and
q u a l i t a t i v e l y in descr ib ing the i n t e r r e l a t i o n s h i p between two
or more f a c t o r s i n v o l v e d i n p l a n t d i s e a s e s . Synerg ism and
a n t a g o n i s m a r e b e s t c o n s i d e r e d as t e r m s d e s c r i b i n g

6 5
TABLE 9
Effect of sulphur dioxide eixture Hitb other pollutants on soybean.
Mixture Host Treataent Effect Reference
SO.tO, Soybean 0.08 or 0.10 ppa 0^, 0.20 or Foliar injury and reduced Pratt et a l . , 1983 0.40 ppa SOji 2 or ^ h/day, chlorophyll synergistic 1-5 days
Soybean 0.067 ppa 0,, 0.30 ppa SO,; Synergistic reduction in Le Sueur-Bryaer and 7.5 h/day, 5 days COj exchange rate (CER), no Oraood, 1984
change in CER with Oj and only slight with SOj
Soybean 0.25-1.0 ppa Oj, 0.50-1.5 ppa Foliar injury and reduced Heagle and Johnstor,. (2 cultivars) SOj; 0.75, 1.5 or 3 hours shoot fresh weight additive, 197">
antagonistic or synergistic, depending upon concentration and tiae
SOj+NOj Soybean 0.2, 0.4, 0.6 ppa of each gases 2; hours
Soybean 0.18 or 0.5 ppa SO, alone (2 cultivars) or Nith 0.06 ppa NOj; 4 h/day, of NOj at loH SOj, I"
Photosynthesis: synergistic, Carlson, 1983 stoaatal conductance: synergistic, respiration: additive
Yield: no additional effect Aaundson, 1983
but 14 days
Soybean 0.13-0.42 ppa SO,, 0.06-0.40 ppa NO2; 3-hours/exposure, 10 days during pod fill (2 year study, aabient air with O3)
increase in reduction at high SO.; not designed to deteraine aixture effects
Chlorophyll reduction, synergistic; yield reduced froa 9 to 25 ; in 2 years, synergistic
Irving and Miller, 1984

6 6
q u a n t i t a t i v e p l an t disease i n t e r a c t i o n s and t h a t the combined
e f f e c t of a p l a n t p a r a s i t i c nematode and a n o t h e r p l a n t
disease organism i n e i t h e r g rea te r or less than the sum of
the e f f e c t s of the i n d i v i d u a l organisms.
Nematodes , f u n g i , b a c t e r i a , s o i l - b o r n e v i r u s e s ,
act inomycetes, e t c . are s o i l i n h i b i t i n g microorganisms and
t h e i r pa thogen ic i t y to many crop p lan ts are we l l documented.
P l a n t p a r a s i t i c nematodes a r e c a p a b l e o f c a u s i n g ' w e l l
man i fes ted d iseases on t h e i r own, bu t i n a s s o c i a t i o n w i t h
o t h e r p a t h o g e n s t h e y cause much g r e a t e r damage due to
s y n e r g i s t i c a c t i o n of i n t e r a c t i n g organisms. There are many
s t a t e m e n t s a v a i l a b l e w h i c h s u g g e s t t h a t s y n e r g i s t i c
i n t e r a c t i o n s are common and cons tan t l y degrade p l a n t h e a l t h .
Nematode and fungal plant pathogens
Observa t ions of A t k i n s o n (1892) on t h e i n t e r a c t i o n
between Meloidogyne spp. Fusarium w i l t of co t ton probably
tend t o researches on d isease comp lexes . He observed t h a t
Fusarium w i l t o f c o t t o n was a l w a y s more s e v e r e i n t h e
presence of r o o t - k n o t nematodes. Many i n v e s t i g a t o r s have
observed t h a t Fusarium w i l t o f c o t t o n caused by Fusarium
oxysporum Sch lec t . f . sp. vasinfactum ( A t k . ) Synd & Hans, was
g r e a t l y increased in presence o f Meloi dogyne i ncognita
(Mar t i n et a / . , 1956; Smith, 1948; 1948; Nor ton, 1960; White,
1962; Brod ie and Cooper, 1964; M a i e r , 1964; M i n t o n and

e 7
Min ton , 1966). P o r t e r and Powel l (1967) observed t h a t
p re inocu la t i on w i th nematodes caused a s i g n i f i c a n t increase
i n fungal i n f e c t i o n . Many i n v e s t i g a t o r s have s t u d i e d t h e
poss ib le i n t e r a c t i o n of nematodes w i t h other s o i l i n h a b i t i n g
microorganisms (Powell 1963, 1971a, 1971b; P i t che r , 1965;
Bergeson, 1972; Khan, 1984; Webs te r , 1985; Mai and Abawi
1987; Tay lor , 1991 ).
The mechanism by which M. incognita predispose co t ton
p lan ts to disease has not been f u l l y exp la ined . M. incognita
may t r a n s p o r t the Fusarium w i l t f ungus i n t o r o o t s o r t h e
fungus may invade th rough wounds made when M. incognita
enters the roots (Smi th , 1954). Caquil and Shepherd (1970)
concluded t h a t s t r ess and d e b i l i t a t i o n of seedl ings caused by
M. incognita damage may be m a j o r f a c t o r i n t h e
p r e d i s p o s i t i o n , and M. incognita i nduced g i a n t c e l l s may
a t t r a c t and/or be more e a s i l y p a r a s i t i z e d by f u n g i t han
normal c e l l s . Sidhu and Webster (1977) proposed t h a t M.
incognita may induce p roduc t ion of a t r ans loca tab le f a c t o r
or f ac to rs t h a t can predispose p l an t pa r ts d i s t a n t from the
s i t e of M. incognita i n f e c t i o n . Root s p l i t s are caused by M.
incognita (McClure et al. 1973; Shepherd, 1970) , and the
s p l i t may be avenues through which the Fusarium oxysporum f .
sp. vasinfectum and other d isease-causing organisms invade
roots (Shepherd, 1970).

6 S
Some fung i are seed-borne, t h i s i nd i ca tes tha t i n these
cases the fungus i s the f i r s t ac t i ve pathogen. S i m i l a r l y i n
t h e s e cases where f u n g i c i d a l seed t r e a t m e n t s a r e n o t
e f f e c t i v e or when non-fumigant nematicides t h a t have systemic
a c t i v i t y are used, fungus become the pr imary , and nematode
the secondary pathogen i n s e q u e n t i a l i n t e r a c t i o n s . The
inc rease i n nematode p o p u l a t i o n d e t e c t e d i n a number o f
s t u d i e s where t h e f u n g u s was t h e p r i m a r y p a t h o g e n
demonstrate the importance o f t h i s t ype o f i n t ^ e r a c t i o n s
(Mountain and Mckeen, 1962; Nordmeyer and S iko ra , 1983).
I n t e r a c t i o n s tud ies i nvo l v i ng nematodes have generally
f o l l o w e d the s e q u e n t i a l d isease complex approach wh i ch
invo lves a pr imary pathogen, the nematode, and a secondary
pathogen, the fungus or bacterium e t c . The use of sequent ia l
i n o c u l a t i o n s c o u l d mask o r p r e v e n t i n t e r a c t i o n f r o m
deve lop ing and a l t e r those a f f e c t e d by t h e r h i z o s p h e r e
p h y s i o c h e m i c a l e n v i r o n m e n t near t h e r o o t t h a t change
d r a s t i c a l l y w i t h p l a n t age (Tay lor , 1979; Webster, 1985).
Change i n root exudat ion pat te rns t ha t develop w i th p l a n t
age could a l t e r the environment and a f f e c t i n t e r p r e t a t i o n
of r e s u l t s . A c t i v i t i e s of microorganisms in the rh izosphere
and r h i z o p l a n e are g r e a t l y i n f l u e n c e d by t he q u a l i t y and
quan t i t y of root exudat ion and s loughed-of f t i ssues (Cook
and Baker, 1983). I t has been o f t e n sugges ted t h a t r o o t
exudates are a l t e r e d by i n f e c t i o n of host p lan ts by p l a n t -

6 9
p a r a s i t i c nematodes, p a r t i c u l a r l y roo t - kno t nematodes. Van
Gundy et a 7. (1977) observed t ha t exudates from M. incognita
i n f e c t e d tomato roo ts a t t r a c t e d hyphae of Rhizoctonia sol ani,
enhanced i n i t i a l s c l e r o t i a l f o r m a t i o n ; and i n c r e a s e d
s e v e r i t y o f r o o t d e c a y . Van Gundy e t a7 . ( 1 9 7 7 ) and
Melakebrahan et a 7. (1985) reported t h a t root exudates from
r o o t - k n o t i n f e c t e d r o o t s were f o u n d t o c o n t a i n h i g h e r
concent ra t ions of Ca, Mg, Na, K, Fe and Cu, as compared to
those f rom n o n - i n f e c t e d r o o t s . Root exuda tes g e n e r a l l y
accumulate near the roo t t i p , and thus the s o i l m i c r o f l o r a
are a l s o abundant and c l ose t o t h e l o c a t i o n o f e n t r y f o r
la rvae o f M. incognita and hyprae of fungus. This sec t ion of
the review w i l l concent ra te on i n t e r a c t i o n s tud ies i n v o l v i n g
nematodes e s p e c i a l l y roo t knot nematodes and w i l t causing and
roo t r o t fung i t h a t would be re levant to the proposed work.
a. In teract ion with w i l t causing fungi
The i n t e r a c t i o n between Meloidogyne spp . and w i l t
c a u s i n g f u s a r i a have been s t u d i e d more t h a n any o t h e r
nematode-fungus combinat ions (Powel l , 1963; Mai and Abawi,
1987). Mar t in et a1. (1956) were the f i r s t who conducted
t h e i r experiments i n greenhouses or growth chambers using
p a s t u r i z e d , s t e r i l i z e d , or f u m i g a t e d s o i l t o s t u d y t he
i n t e r a c t i o n between Fusarium w i l t fungus and Meloidogyne s p . .
I n t e r a c t i o n between Meloidogyne spp . and f u n g i c a u s i n g
Fusarium w i l t have been s tud ied d i f f e r e n t i n v e s t i g a t o r s on
a l f a l f a ( G r i f f i n , 1986; McGuire et al., 1958); banana (Loos,

T O
1959; Newhall , 1958); beans (R iber io and Ferraz, 1984; Singh
et al. , 1981); Cabbage ( F a s s u l i o t i s and Rau, 1969); ca rna t i on
( S c h i n d l e r e£ al., 1 9 6 1 ) ; ch rysan themum ( J o h n s o n and
L i t t r e l l , 1 9 6 9 ) ; c o t t o n ( A t k i n s o n , 1892 ; C a u q u i l and
Shepherd, 1970; Cooper and Brodie, 1933; Garber et al., 1979
Mar t in et al., 1956; Minton and Minton, 1966; Nor ton, 1960)
cowpea (Thomason et al., 1 9 5 9 ) ; mimosa ( G i l l , 1958)
muskmelon (Bergeson, 1975); pea (Davis and Jenk ins , 1963)
tobacco (Melendez and Powell , 1967; Por ter and Powel l , "^96'
and tomato (Abawi and Baker, 1984; B inde r and H u t c h i n s o n ,
1959; Bowman and Bloom, 1966; F a t t a h and Webs te r , 1983;
Goode and McGuire, 1967; Hirano et al., 1979; Jenkins and
Coursen, 1957; Jones et al. 1976; Kawamura and Hi rano, 1967;
1968; Or ion and Hoes t ra , 1974; p i t c h e r , 1978; S idhu and
Webster, 1974, 1977, 1979, 1983; Young, 1939, 1940).
Much work has been done on t h e i n t e r a c t i o n o f r o o t
i n f e c t i n g fungi w i t h endoparas i t ic nematodes i n comparison t o
semi-endoparas i t ic or e c t o p a r a s i t i c nematodes (Khan, 1984).
The i n t e r a c t i o n o f nematodes w i t h w i l t c a u s i n g f u n g i
espec ia l l y Fusarium spp. have drawn much a t t e n t i o n . (Powe l l ,
1971a, 1971b). McGuire et al (1958) s tud ied the e f f e c t of
roo t - kno t nematodes on the development of Fusarium w i l t i n
Bu f fa lo v a r i e t y of a l f a l f a . At t e r m i n a t i o n , the percentage of
p lan ts i n fec ted w i t h Fusarium w i l t observed by them are as
f o l l o w s : Meloidogyne hapla & Fusar i urn oxysporum f . spp .

7 1
vasinfectum 95, M. Javanica & F. oxysporum f. sp. vasinfectum
60, M. incognita & F. oxysporum f. sp. vasinfectum 60, Af.
incognita & F. oxysporum f. sp. vasinfectum 50, Af. arenaria &
F. oxysporum f. sp. vasinfectum 50, W. incognita acrita and
F. oxysporum f. sp. vasinfectum 15, Meloidogyne spp. alone
and check 0, percentage of dead plants in the above
treatments was 40, 5, 10, 4, 0, 3, 0 respectively. Kumar et
a7. (1988) observed that inoculation of either of M.
incognita or F. oxysporum f. sp. ciceris reduced growth of
chick-pea and highest reduction were obtained v»'nth
simultaneous inoculation of both pathogens followed by the
treatment in which the nematode inoculation preceded the
fungus by 10 days. Multiplication of nematodes and galling
was reduced in the presence of fungus. Mani and Sethi (1987)
studied the effect of the combined incoula of M. incognita,
Fusarium oxysporum or F. soTani on growth of chick-pea which
was found to be additive in nature. However, when nematode
was established earlier than the two fungi, the resultant
effect was more than simple additive. All the three organisms
affected the rhizobial nodulation considerably. Occurrence
of M. incognita in combination with F. oxysporum f.sp.
ciceris and F. solani, not only increased the severity of
disease but also shortened the incubation period for disease
expression. F. oxysporum f. sp. ciceris did not affect the
nematode population significantly. Patel et a7. (1987) also
observed increase in incidence of chick-pea wilt due to F.

7 2
oxysporum f. s p . ciceris i n p resence o f M. incognita.
Simultaneous i nocu la t i on of both pathogens resu l ted i n more
w i l t percentage. Mart in et al. (1956) s tudied i n t e r a c t i o n of
Fusarium oxysporum f . s p . vasinfectum causing w i l t o f co t ton
w i t h f i v e nematodes v i z . , M. incognita, M. incognita acrita,
Trichodours s p . , TyTenchorhynchus sp. and Helicotylenchus sp.
Of these nematodes, only M. incognita and M. incognita acrita
s i g n i f i c a n t l y increased incidence of w i l t i n the both w i l t
suscep t ib le and w i l t r e s i s t a n t v a r i e t i e s . The e f f e c t of M.
incognita Race 3 and F. oxysporum f . sp. vasr-^ectum on i-- -c
expression of co t ton was s tud ied by Ibrahim et al. (1982).
They o b s e r v e d t h a t p r e s e n c e o f nematode enhanced t h e
development and seve r i t y of w i l t . Expression of w i l t symptoms
was observed 10 days e a r l i e r i n the case of i n f e c t i o n w i t h
both organisms than w i th the fungus alone and reduc t ion in
p lan t growth were almost more than two- fo lds when the two
pathogens were present toge ther . Palmer and MacDonald (1974)
s tud ied maize roo t -kno t complex caused by Fusarium spp. i n
the presence o f va r i ous p l a n t p a r a s i t i c nematode s p e c i e s
i n c l u d i n g M. i ncogni ta, Pratylenchus penetrans, P.
herincissus, P. scribneri, P. nanus and Tylenchorhynchus
martini. Average dry root and shoot weight of maize seed l ing
i n o c u l a t e d w i t h M. incognita and F. mani 1 iforme were l e s s
than those of seed l i ng i n o c u l a t e d w i t h e i t h e r o rgan i sms
a lone. Goel and Gupta (1984) studied i n t e r a c t i o n between M.
Javanica and F. oxysporum f. sp. cieris. They f ound t h a t

7 3
shoot length was minimum when the fungus was inoculated a
week after nematode inoculation, whereas root length, fresh
shoot and root weights were significantly less when fungus
were inoculated a week before nematode inoculation. After
two years in the same field of study they observed maximum
and significant reduction in shoot length and fresh weight
when both the pathogens were inoculated simultaneously. A
maximum and significant reduction in number of galls was
recorded when nematode and fungus were present together, " he
reduction in the presence of F. oxysporum f. sp. cicerit wat
attributed either to poor and delayed nematode development or
to inhibitory effect of fungus on hatching of egg masses
(Goel and Gupta, 1986). In pot experiments, Upadhyay and
Dwivedi (1987) observed that wilt symptoms were greatest in
chick-pea plants inoculated simultaneously with both M.
javanica and F. oxysporum f. sp. ciceris, followed by
severity by inoculations of the nematode preceding the fungus
and the fungus preceding the nematode. The maximum number of
root galls was recorded on roots inoculated with the nematode
alone and the minimum number on roots where inoculation of
the fungus preceded that of the nematode. The maximum
reduction in shoot weight occurred where inoculation of the
nematode preceded that of the fungus.
Padila et al., (1980) observed that Meloidogyne spp.
alone caused severe stunting, F. oxysporum alone caused no
detrimental effect, whereas concomitant inoculation of the

7-4-
nematode and fungus at planting caused the death of the
plants after 45 days.
Fungi non-pathogenic to a host plant may become
pathogenic in presence of nematodes. F. oxysporum f. sp.
lycopersici was not pathogenic to cucumber plant, but wilt
symptoms developed when M. incognita was in association of
fungus, but F. oxysporum f. sp. cucumerinum did not produce
symptom on tomato plants even when M. incognita was present
(Pelez et al., 1983). Smits and Noguera (1982) observed that
F. oxysporum f. sp. melon is and F. oxysporum f. sp.
lycopersici were pathogenic to only those eggplant seedings
that were infected with M. incognita. Lopes and Lordello
(1979) studied the wilt of Piper nigrum and observed that
more damage was done, when M. incognita and F. sol ani f. sp.
piperi were present together.
Nogurea and Smits (1982) suggested that root exudates
of tomatoes infected with M. incognita stimulate soil
microflora reproduction and account for the greater number of
colonies (per gram soil) of bacteria and fungi in infected
than in healthy plant. This increase, lowered the number of
actinomycetes antagonistic to F. oxysporum f. sp. lycopersici
and enhanced its pathogenic effect. Nogurea (1980) observed
that germination and growth of F. oxysporum f. sp.
lycopersici in root extracts of a Fusarium resistant tomato
was significantly greater when tomatoes had been infected

T 5
wi th M. incognita. A f te r four weeks of nematode i n f e c t i o n ,
the amount of amino acids in the roots had increased by 509
and ca rbohydra tes by 112% and t h i s was assumed t o be t h e
reason f o r the breaking of res is tance to Fusarium. I n f l uence
of M. incognita on the l o c a l i z a t i o n of F. oxysporum f . sp.
lycopersici i n tomato was s t u d i e d by Noguera ( 1 9 8 3 ) . He
observed t h a t funga l p e n e t r a t i o n was no t r e s t r i c t e d t o
nematode s i t e but occur red t h r o u g h o u t t he r o o t c o r t e x and
advanced i n t o t h e stem v i a t h e x y l e m . Nematode a', ". - c -
appeared t o cause systemic change i n t h e p l a n t t i s s u e .
Mon ta l ro and Melendez (1986) s t u d i e d h i s t o p a t h o l o g y o f
i n t e r a c t i o n between F. oxysporum f . sp. dioscoreae and two
nematode species on yam. Examination of roo t t i s s u e s showed
t h a t fungal c o l o n i z a t i o n occurred i n d i s c r i m i n a t e l y , e i t h e r
i n t e r - o r i n t r a c e l l u l a r l y . C o l o n i z a t i o n i n t h e stem was
r e s t r i c t e d t o t h e v a s c u l a r s y s t e m , w i t h a b u n d a n t
p r o l i f e r a t i o n o f hyphae, ma in l y i n t he noda l t i s s u e s .
Apparent ly M. incognita and the fungus co lon ized i n d i f f e r e n t
sec tors of the invaded t i s s u e s , though abundant and v igour
hyphae were p resen t in t i s s u e s m o d i f i e d by M. incognita,
i n d i c a t i n g a p o s i t i v e i n t e r a c t i o n . Pratylenchus coffeae was
sometimes observed wi th the fungus w i t h i n the same t i s s u e s ,
but t h i s assoc ia t i on had no e f f e c t on s e v e r i t y or inc idence
o f w i l t . K le ineke -Borche rs and Wyss (1982) o b s e r v e d t h a t
mixed i n f e c t i o n o f M. incognita and F. oxysporum f . s p .
lycopersici on tomato p l a n t s r e s u l t e d i n an i n c r e a s e i n

y 6
indole-acetic acid which may be responsible for quicker
appearance of typical wilt symptoms. Nogurea (1982) observed
that purified extracts of tomato roots infected with M.
incognita did not show rishitin, an anti-fungal substance
normally present in healthy plants. Nematode free plants also
showed the formation of several well developed tyose in the
xylem vessels of stem whereas in infected plants these were
absent or very reduced. They suggested that the loss o"*"
resistance of this tomato variety '.'~ '". oxysporum ^. c.
lycopersici infection was associated with the nematode
infection which interfered with rishitin and tyose
production. Morrell and Bloom (1981) studied effect of
temperature on expression of wilt symptom on tomato plants
inoculated with M. incognita and F. oxysporum f. sp.
lycopersici and observed that wilt severity was increased at
21°C but not at 16°C. Hillocks (1986) studied localised
effect of nematode on initial infection and a systemic effect
on subsequent disease development of plants. To achieve this,
cotton seedlings were grown with a divided root system and
inoculated with root-knot nematode, M. incognita and F.
oxysporum f. sp. vasinfectum on the same or opposite halves
of the root system. Wilt indices were increased by the
nematode, but only in plants where both organisms were
together on the same parts of the root system. However, in a
second experiments in which plants growing in Meloidogyne
infected soil were stem inoculated with the wilt pathogen.

7 7
the nematode caused an increase in w i l t s e v e r i t y , despite the
phys ica l separat ion of the two organisms.
Root -kno t nematodes are found a s s o c i a t e d w i t h o t h e r
w i l t causing fung i a l so . Shoemaker and Barker (1979) in a
m i c r o p l o t exper iment observed t h a t M. incognita had no
s i g n i f i c a n t e f f e c t on Vertici11ium dahaliae w i l t of tomato.
Goswami e t al. (1970) r e p o r t e d h i g h degree o f synerg ism
between M. incognita and Sclerotium rolfsii in b r i n j a l when
i n o c u l a t e d t o g e t h e r . P rasad et al. ( 1980 ) s t s j o i ec '-he
a s s o c i a t i o n o f Corticium rolfsii and M. incognita i n w i l t
complex of Solanum khasianum, a medicinal p l a n t . They found
t h a t 42% o f the p l a n t s w i l t e d , where t he fungus and t h e
nematodes were combined, but 12.5X w i l t e d w i t h t h e fungus
alone and no w i l t occurred w i th the nematode a lone. Venkata
Rao e t al. (1973) reported an assoc ia t ion of M. incognita
var . acrita on Phytophthora parasitica var . nicotianae on
p i p e r - b e t e l r oo t s , Azam et al. (1977) observed t h a t presence
o f M. incognita cons ide rab l y i n c r e a s e s t he i n c i dence o f
d isease caused by Rhi zoctoni a sol an i, Py^
Col letotrichum atrametarium, on eggplant , wj^c A<fi<M<fS
Synergism and antagonism have been oJj&feVved betij/r^'e^y
f ung i and o the r e n d o p a r a s i t i c nematodes t han r o o t - k n o t
nematodes. Jorgenson (1970) observed tha t i n t e r a c t i o n between
Heterodera schachtii and Fusarium oxysporum was
d isadvantageous t o the nematode and damage t o suge rbee t
. ^

7 3
plants were less when the fungus and nematodes were present
than when only the nematode was present. Edward and Singh
(1979), while studying the interaction between Heterodera
cajani and F. udum on pigeon-pea, found that the nematode
alone caused less damage to var. Type-21 than when
associated with F. udum. Hasan (1984) observed synergism
between H. cajani and F. udum attacking Cajanus cajan. He
found a significant increase in wilting when H. cajani and
F. udum were associated with pigeon-pea. Seinhorst and
Kuniyasu (1971) reported that rate of multiplication of
Rotylenchulus reniformis on pea var. "Rando" was increased by
the presence of F. oxysporum f. pisi race-1. Krishnaprasad
and Padanganur (1980) observed that soil from Verticillium
wilt affected plant harboured more Rotylenchulus reniformis
population compared to that of healthy plants. They concluded
that R. reniformis might be associated with Vertici 11 ium v^^^t
of cotton as evidenced by fairly high population of nematodes
in the rhizosphere and roots of diseased plants. Mountain and
Mckeen (1962), while studying the interaction of lesion
nematode, Pratylenchus penetrans with Verticil 1 ium dahliae on
tomato, observed that reproduction of nematode was enhanced
in such an interaction. In Vertici num-vl^\^X, of potato, the
damage were more when lesion nematode was present (Morsink
and Rich, 1968). Burpee and Bloom (1974) observed that
incubation period of V. albo-atrum was decreased in the
presence of P. penetrans. In another report, they showed that

7 9
Praty lenchus penetrans i n r o o t s o r r h i z o s p h e r e were
suppressed i n the presence of Vertici11ium (Burpee and Bloom,
1 9 7 8 ) . F e r r a z and Lear ( 1 9 7 6 ) f o u n d a s y n g e r g i s t i c
i n t e r a c t i o n between F. oxysporum f . sp. dianthi and nematodes
such as Criconemoides curvatum, Pratylenchus dianthus and M.
hapla on c a r n a t i o n . G r u j i c i c e t a1. (1984) examined the
lucerne f i e l d and repor ted t ha t damage due to w i l t and r o t
was g r e a t e r when P. penetrans and F. oxysporum v a r ,
medicaginis were p resen t t o g e t h e r , R iede l e t a 7. ( ^ " ^ f ,
r e p o r t e d a d i f f e r e n t i a l i n t e r a c t i o n s o f Praty1enchus
crenatus, P. penetrans and P. scribneri w i th V. dahliae i n
p o t a t o e a r l y dy ing d i sease . S t i n g nematode, Belonolaimus
longicaudatus have been found t o i n f l u e n c e i n c i d e n c e o f
co t ton w i l t caused by F. oxysporum f . sp. vasinfectum (Cooper
and Brod ie , 1963).
R o o t - k n o t nematodes have been f o u n d t o b reak t h e
res is tance of the crop p lan ts against so i l -bo rne pathogens.
Car ter (1978) repor ted enhancement of w i l t sever i t y in the
completely (po l ygen ic ) r e s i s t a n t c u l t i v a r s of tomato as a
r e s u l t of M. incognita and F. oxysporum f . sp. lycopersici
i n t e r a c t i o n . Hadiastono (1981) observed tha t tomato v a r i e t i e s
o r i g i n a l l y r e s i s t a n t to F. oxysporum became suscept ib le when
inocu la ted w i t h Meloidogyne spp. Harr ison and Young (1941)
observed t h a t r o o t - k n o t nematodes reduced the w i l t res is tance
of many v a r i e t i e s of tomatoes. In a glasshouse experiment 20

eo
tomato l i n e s and v a r i e t i e s inocu la ted w i t h a Fusarium p lus
Meloidogyne were found t o be s u s c e p t i b l e i n c l u d i n g those
r e s i s t a n t t o F. oxysporum f. sp . lycopersici. Donald and
Johnson (1973) inocu la ted e igh t c u l t i v a r s of watermelon w i th
known r e a c t i o n s t o Fusarium w i l t w i t h M. incognita and
observed t h a t w i l t symptoms were increased more in r e s i s t a n t
than in suscep t ib le c u l t i v a r s , Johnson and L i t t r e l l (1969)
observed t h a t M. incognita, M. hapla and M. Javanica d id not
break r e s i s t a n c e o f chrysanthemum aga" ">- t F. .--r^^por ••<•.
O'Ereo le et al. (1982) i n an i n v e s t i g a t i o n - r .vo ' .v-ng f o u r
v a r i e t i e s of tomato showing res is tance t o M. incognita and
one not res is tance showed tha t i n f e c t i o n w i t h F. oxysporum f .
sp . lycopersici was g rea te r i n t he nematode s u s c e p t i b l e
v a r i e t y than in o ther more r e s i s t a n t v a r i e t i e s . They observed
s i m i l a r (a l though less conc lus ive) r e s u l t s w i t h Vertici11ium
dahliae. Kle ineke-Borchers and Wyss (1982) noted t h a t p r i o r
i n f e c t i o n o f M. incognita f o l l o w e d by Fusarium oxysporum
i nocu la t i on g r e a t l y enhanced fungal disease in both Fusarium
suscep t ib le and r e s i s t a n t tomato p l a n t s . P i tcher (1974) a lso
proved t h a t i n f e c t i o n of "Pearson VF" tomato wh ich i s
moderate ly r e s i s t a n t t o F. oxysporum f . sp . 1 ycopersici
became suscep t ib le t o t h i s fungus in presence of M. javanica.
Thomason (1958) found tha t w i l t in Vigna sinensis was more
s e v e r e i n p l a n t s grown i n s o i l i n f e s t e d w i t h b o t h F.
oxysporum f . sp. tracheifilum and M. Javanica than in s o i l
i n fes ted w i th fungus alone. McGuire e t al. (1958) observed

a 1
tha t on ly )0% p lan ts showed the w i l t symptoms when i n fec ted
w i th M. incognita acrita and 15% w i th F. oxysporum f. sp.
vasinfectum a lone , whereas 95%, 60% and 50% p lan ts became
w i l t e d when the fungus was present in combination w i t h M.
hap7a, M. Javanica and M. incognita r espec t i ve l y .
D a v i s and J e n k i n s ( 1963 ) o b s e r v e d a s y n e r g i s t i c
response between Meloidogyne spp. and Fusariun^-vi^'St r e s i s t a n t
pea v a r i e t y . An increase i n s e v e r i t y o f Fasar7u/n-wi 11 o f
tomato and increased co lon i za t i on o^ fungus in the vascular
t i ssue were observed when M. incognita was present (Car ter
et al. y 1977). Garber et al., (1979) found t h a t i n s o i l
i n f e s t e d w i t h F. oxysporum f . s p . vasinfectum and M.
incognita, the ex tent of fungal invasion and c o l o n i z a t i o n of
c o t t o n was c o r r e l a t e d w i t h the nematode g a l l i n g . Noguera
(1983) observed t h a t r o o t s o f tomato p l a n t s o n l y showed
c o l o n i z a t i o n by F. oxysporum when in fec ted w i th M. incognita.
b. I n t e r a c t i o n w i t h r o o t - r o t fung i
Many i n v e s t i g a t o r s have not iced t ha t nematodes i n t e r a c t
w i t h r o o t - r o t fung i s y n e r g i s t i c a l l y , hence greater damage t o
p l a n t . Tu and Cheng (1971) found t ha t s e v e r i t y of r o o t - r o o t
i n k e n a f (Hibiscus cannabinus L . ) was i n c r e a s e d when
seed l i ngs were i n f e c t e d s i m u l t a n e o u s l y by Macrophomi na
phased ina and M. javanica. Mishra et al. (1988) observed
tha t M. incognita provided a good s i t e f o r easy en t ry of the
developing hyphae of R. bataticola {M. phased ina) in whi te

3 2
jute {Corchorus capsularis) and thus increased the severity
of the disease. The number of dead plants were increased
significantly when pathogens were inoculated together. Goel
and Gupta (1986) observed that irrespective of time, combined
inoculation of M. Javanica and R. bataticola on chick-pea
reduced growth parameters compared with when inoculated
alone. The number of root galls was significantly reduced
when nematode was inoculated 7 days before the fungus. Bruton
and Heald (1987) studied effect of M. phaseol i na on
cantaloupe in the presence of M. incogni ta and found
reduction in top dry weight of the plant. Apt and Koike
(1962) studied the relationship of M. incognita acrita with
Pythium graminicola on sugarcane. Colin and Powell (1971)
reported an extensive fungal colonization in the root-rot
susceptible cultivar of tomato "Dixie Bright 101" when M.
incognita preceded R. solani by 21 days. Sharma et a 7.
(1980) reported a syngergistic effect on the root-rotting of
okra plants when M. incognita and R. bataticola were
combined together. Al-Hazmi (1985) reported that severity of
root-rot caused by M. phaseolina in root-rot disease complex
of French bean {Phaseolus vulgaris) increased when the
nematode was introduced 2 weeks before the fungus, and if the
fungus was introduced first, then the nematode infection and
reproduction were reduced. Chhabra and Sharma (1981) in a pot
trial with okra and eggplant observed that M. incognita and
R. bataticola significantly reduced the germination of both

8 3
TABLE 10
Interaction of root-knot neiatodes vith Hilt causing Fusaria and root-rot fungi.
Heloidogyne spp. Associated organisi Host Coitents References
H. incognita
H. incognita
H. incognita
H. incognita
H. incognita
H. incognita
H. incognita
H. incognita
H. incognita
Fusariut equiseti
F. oxysporua f. sp. nicotianae
F. oxysporut f. sp. ciceris
F. solani
F. oxysporut f.sp. telonis F. oxysporut f.sp. lycopersici
Fusariut spp.
Fusariut toniliforte
F. oxysporut f.sp. lycopersici
F. oxysporut f.sp. lycopersici
Pea Preponded wilt syiptoi Chahal and Chhabra, appearance 1985
Tobacco Significant infection Porter and PoNell, occurred only when both 1967 pathogens Mere present
Chickpea Increased tiilt severity Hani and Sethi, 1987; and shortened incubation Patel et al.. 1987; period of disease Ki'nar ?: a!., :98f expression
Chickpea Increased Milt severity Hani & Sethi, 1987 and shortened incubation period of disease expression
Bringal Non-pathogenic fungi Stits i Noguera, caused severe vascular 1982 necrosis of the shoots
Hatertelon Increased Nilt severity Donald i Johnson, 1973
Haize Increased root-rot severity
CucuBber Non-pathogenic fungus caused wilting in presence of nenatode
Palier & HacDonald, 1974
Pelez et al., 1983
Toiato Increased Hilt severity Horrell and Blooi, 1981; Kleineke-Borehers and «yss, 1982; Carter et al., 1977.

Table 10 contd..
a^
Heloidogyne spp. Associated organist Host Coiients References
h. incognita
H. incognita
M. incognita
h. incognita
M. incognita
M. incognita
F. oxysporua f.sp. vasinfectua
F. oxysporua f.sp. pisi
F. oxysporua f.sp. vasinfectua
R. bataticola
/?. solani
R. solani
Cotton Increased wilt severity Cooper & Brodie, 1963; Martin et a l . , 1956; Ibrahii et a l . , 1982; Hillocks, 1986
Pea Increased wilt severity Padilla et a l . , 1980
Alfalfa Increased wilt severity McGuire et a l . , 1958
French Increased severity o* ^l-Hazmi, 1985 bean root-rot
Cotton Synergist occurred
Tobacco Worst infection occurred when netatode inoculated 10 or 21 days before fungi
Carter, 1975
Batten and Powell, 1971
M.
n.
M.
H.
H.
n.
H.
incognita
incognita
incognita
incognita
incognita
incognita
incognita acrita
R.
R.
H.
R.
R.
R.
F.
solani
solani
phaseolina
solani
solani
bataticola
Potato
Okra
Hhite jute
Toaato
French bean
Okra
oxysporua f.sp. pisi Pea
Increased disease severity
Sharna and Gill, 1979
Siiultaneous inoculation Chhabra et ai.,1977 caused Hax reduction
Increased disease severity
Increased fungal colonization
Increased disease severity
Increased root-rot
Synergistic response observed, broke resistance
Mishra et al., 1988
Colin and Powell, 1971
Reddy et ai.,1979
Sharea et al., 1980
Davis and Jenkins, 1963

Table 10 contd..
es
Meloidogyne spp. Associated organist Host Conents References
M. incognita acrita R. solani
h. incognita acrita F. oxysporut f.sp. cubense
h. incognita
H. javanica
H. javanica
M. javanica
H. javanica
H. javanica
H. javanica
H. javanica
H. javanica
H. javanica
R. solani
F. oxysporut f.sp. ciceris
F. oxysporut f.sp. tracheiphilut
F. oxysporut f.sp. telonis
F. oxysporut f.sp. lycopersici
F. oxysporut
F. udut
R. bataticola
R. bataticola
R. solani
Cotton Disease increased Reynolds and Hanson, 1957
Banana Incubation period of Loos, 1959 fungus shortened
Toaato Reduction in growth Chahal and Chhabra, paraneters 1984
Chickpea Significant reduction Upadhyay and Kusui in groMth paraieters DHivedi, 1987;
Chhabra et ai..l977; Goel a!\C GuDta. 1984 l%t
CoMpea Increased Nilt severity Thonason et ai.,1959
Huskielon Increased wilt severity, broke resistance
Totato Susceptibility to Milt increased
Chrysan- Early and severe Milt theaui appearance
Pigeon- Susceptibility to Milt pea increased
Chickpea Reduced growth paraieters
To«ato Increased nil ting incidence
CoMpea Plant growth reduced when neaatode inoculation was done 3 weeks prior to the fungus
Orion and Hetzer, 1981
Jenkins and Coursen, 1957; Bergeson et a l . , 1970
Johnsot) and Littrell, 1%9
Abdul Salai and Xhan, 1986
Goel and Gupta, 1986
Goswa«i et al., 1976
Kunar et a l . , 1988

Table 10 contd..
a©
Heloidogyne spp. Associated organisB Host Coiients References
«.
M.
H.
tt.
H.
H.
H.
He
javanica
javanica
hapla
hapla
hapla
arenaria
arenaria
loidogyne spp.
H. phaseolina
R. bataticola
F. oxysporuB f.sp lycopersici
F. oxysporut f.sp vasinfectuB
F. oxysporuB
F. oxysporva f.sp nicotianae
F. solani
F. oxysporua f.sp lycopersici
Kenaf Increased severity of Tu and Cheng, 1971 root-rot
French Reduced gronth para- Reddy et al., 1979 bean leters
Tonato Hilt incidence increased Jenkins and Coursen, 1957
ftlfalfa Severe wilting appeared HcGuire et aJ.,1958
Chrysan- Hilt incidence increasec Johnson ana Liitrel thenun 1969
Tobacco Significant infection Porter and Powell, occurred only Nhen both 1967 pathogens present
Ground Earlier appearance of Patel et al., 1985 nut Milt syiptons
Tonato Resistance broken Aliedia, 1972
Heloidogyne spp. F. oxysporua Toiato Resistance broken Nadiastono, 1981

3 T
spec ies when they were p resent t o g e t h e r i n t h e s u b s t r a t e .
Goswami e t al. (1976) noted t h a t when M. javanica was
inocu la ted onto tomato p l a n t s , 3-weeks before i n f e c t i o n w i t h
R. bataticola, 33.3X w i l t i n g occur red. Approximately 13.3,
6.7 and 6.7% w i l l e y was noted a f t e r simultaneous pathogen
i n o c u l a t i o n ; fungus i n f e c t i o n alone or fungus i n f e c t i o n 3-
weeks before nematode i n o c u l a t i o n s , r e s p e c t i v e l y . Sakhuja
and Sethi (1986) observed t ha t m u l t i p l i c a t i o n of M. Javanica
and the g a l l i n g on groundnut was adversely af-Pet et b:
bataticola and F. solani. Reduction in m u l t i p l i c a t i o n was
maximum wherever one or both t h e f u n g i were i n o c u l a t e d
s imul taneously w i t h the nematode, but R. bataticola proved t o
be more an tagon i s t i c to M. javanica.
Khan and Mul le r (1982) s tud ied the i n t e r a c t i o n between
R. solani and M. hapla on r a d i s h {Raphanus sativus) i n
g n o t o b i o t i c c u l t u r e in p e t r i d i s h e s . This experiment i nd i ca ted
t h a t the phys io log i ca l changes in the ga l l ed reg ion due to
nematode i n f e c t i o n predisposed the roots f o r invas ion and
r a p i d c o l o n i z a t i o n by the f u n g u s . Reddy e t al. (1979)
repor ted t ha t roo t -kno t nematode, M. incognita and R. solani
are o f t e n found assoc ia ted s i m u l t a n e o u s l y w i t h r o o t s o f
f r e n c h bean (Phaseolus vulgaris) under f i e l d c o n d i t i o n s .
Reduct ion in growth parameters were observed when bo th
pathogens were present at a t i m e . Kanwar e t al. (1988)
observed t h a t growth of cowpea was s i g n i f i c a n t l y reduced

when M. javanica was inoculated 3 weeks p r i o r to R. solani.
The number o f b a c t e r i a l n o d u l e s and g a l l s was r e d u c e d
s i g n i f i c a n t l y i n presence of bo th pathogen w i t h maximum
reduc t ion in nematode alone t reatment . Sharma and G i l l (1979)
observed s i g n i f i c a n t r e d u c t i o n i n shoo t w e i g h t o f p o t a t o
p l a n t inocu la ted w i t h M. incognita and / or R. solani and
maximum r e d u c t i o n i n m u l t i p l i c a t i o n o f nematode (66.725K)
occurred when R. solani was inoculated before M. incognita.
Chahal and Chhabra (1984a) reported t h a t M. incognita and R.
solani separate ly as wel l as in combination s i g n i f i c a n t l y
reduced shoot l eng th , shoot and root weights of tomato. They
f u r t h e r observed t h a t i nocu la t i on of tomato seedl ings w i t h M.
incognita and R. solani reduced seed g e r m i n a t i o n t o 18%
compared w i th 48% and 39% in the case of s i ng l e i n o c u l a t i o n
o f M. incognita and R. solani r e s p e c t i v e l y (Chahal and
Chhabra 1 9 8 4 b ) . Chhabra et al. ( 1 9 7 7 ) showed t h a t
i n t e r a c t i o n i n v o l v i n g R. solani and M. incognita on o k r a
s i g n i f i c a n t l y reduced shoot and root lengths and wet and dry
w e i g h t s . H a z a r i k a and Roy ( 1 9 7 5 ) examined t h e
i n t e r r e l a t i o n s h i p between R. solani and M. incognita on
eggplant and found t h a t number of g a l l s and egg masses on
roo ts were s i g n i f i c a n t l y greater in p lan ts inocu la ted w i th
nematode and fungus together than in those inocu la ted w i th
nematode alone. Chhabra et al (1977) observed t h a t maximum
reduc t i on in growth parameters of okra was obtained when M.
incognita and R. solani were i n o c u l a t e d s i m u l t a n e o u s l y .

8 9
Fungus with mechanical injury of roots was more damaging to
plants than the fungus alone, Chhabra et a1. (1978) studied
the influence of soil types on the interaction of R. solani
and M. incognita and found that inoculation of okra with
both the pathogens markedly reduced shoot and root growth.
Nematode population were much denser in sandy loam as compare
to loamy sand or sandy clay loam, but number of galls did not
vary between soil types.
Powell et al. (1971) noticed that soi1-bo»-ne " mg- l--c
Pythium, Curvularia and Botrytis caused extensive root decay
when root-knot nematode preceded the fungi. M. incognita was
found associated in a root-rot with F. concoTar in papaw and
with F. oxysporum in tomato (Saeed et a7, 1972). M. arenaria
appeared to predispose the maize plant to S. rolfsii (Minton
et al., 1975). Welty et al. (1980) found that Phytophthora
megasperma f. sp. medicaginis intensified root-rot severity
in resistant and susceptible lucerne when M. hapla or M.
incognita were present. Mukhtar and Khan (1989) observed
greatest suppression in chick-pea plant when they were
inoculated simultaneously with M. Javanica and S. {Corticium)
rolfsii, followed by the fungus prior to nematode.
Besides root-knot, other nematodes also have been
found to interact with root-rot fungi, Kisiel et al. (1969)
studied the possible interactions between nematodes,
Tylenchus agricola and Tylenchorhynchus claytoni on root-rot

9 0
of corn caused by F, roseum and pythiurn ultimum. Nematode
population increased to a greater degree in plants inoculated
with F. roseum than with P. uTtimum. Penetration of stele of
F. roseum was significantly greater in presence of nematodes.
They concluded that nematode may in some cases increase the
severity of the disease, but the fungus was by so far the
dominant pathogen in this relationship. Littrell and Johnson
(1969) observed an interaction in combined inoculation with
Pythiurn aphanidermatum and BeTonolaimus longicaudatus on
chrysanthemum. Sobun et al. (1979) reported an
interrelationship between Tylenchorhynchus and root-rot
fungus on chick-pea. Upadhyay and Swarup (1981) observed a
significant reduction in growth of maize plant, when both
Tylenchorhynchus vulgaris and F. moniliforme were inoculated
together. Abawi et al. (1985) studied the disease complex of
Phased us vulgaris caused by one or more species of Fusarium,
Rhizoctonia, and Thielaviopsis with lesion nematode,
Pratylenchus spp. Studies on interactions of Vertici11ium
dahliae, Colletotrichum coccodes, Rhi zoctoni a solani and
Pratylenchus penetrans in the early dying syndrome of potato
revealed an additive symptom expression, when P. penetrans
combined with V. dahliae (Kotcon et al., 1985).
Interaction of nematode and fungal plant pathogens under
pollutant stress
Apparently no attempt has been made to determine the
impact of air pollution stress on fungus-nematode interaction

9 1
in disease complexes. Effects of some heavy metals as soil
pollutants have been studied by some workers in nematode-
fungus complexes. Khan and Abul Salam (1990) studied effects
of nickel (Ni) and cobalt (Co) at different concentrations oh
interaction of Meloidogyne Javanica, Fusariurn udum and
Rhizobium on pigeonpea. They observed that M. javanica and F.
udum interacted synergistical 1 y and caused more severe
wilting symptoms than F. udum, but the result was reverse in
the presence of Ni, i.e. wilting was completely in-''--'tet -
root galling was suppressed but it caused reduction in plant
growth and nodulation. They suggested that this may be due to
toxic effect of nickel towards the pathogens. It was found
that cobalt too, suppressed the wilting symptoms on the test
plants by F. udum, and had similar effect as a nickel. Cobalt
increased galling produced by M. javanica and caused a
significant reduction in growth parameters of the plant.
The review of literature shows that in recent years
some studies have been conducted on interactions between
biotic pathogens (including fungi and nematodes) and some
gaseous air pollutants. The ability of some biotic pathogens,
particularly viruses, bacteria and fungi to influence the
susceptibility of the plants to air pollutants has been
Whether the nematodes also influence the plant susceptibility
to air pollutants is yet to be explored fully. The effect of
gaseous air pollutants on the interactions between nematodes

9 2
and fungi has not gained study. Whether the disease i n t e n s i t y
caused by nematodes or f u n g i o r t h e d i s e a s e s e v e r i t y
developing through t h e i r i n t e r a c t i v e e f f e c t in a s y n e r g i s t i c
r e l a t i o n s h i p are i n f l u e n c e d a d v e r s e l y or f a v o u r a b l y or
whether t h i s r e l a t i o n s h i p i s i m p a i r e d on a i r p o l l u t i o n
stressed needs t o be examined. The present study i s planned
to answer some of these quest ions.

MATERIAL AND METHODS

The method t o be employed and steps to be taken f o r
o b t a i n i n g t h e m a t e r i a l s f o r c o n d u c t i n g t h e p roposed
i n v e s t i g a t i o n s w i l l be as f o l l o w s :
1 . Root-knot nematode population
a. Collection of field populations:
F i e l d p l o t s grown w i t h v e g e t a b l e s and pu l ses i n and
around A l i g a r h w i l l be surveyed t o c o l l e c t r o o t samples
i n fec ted w i t h roo t - kno t nematodes. Root samples co l l e c ted in
polythene bags and proper ly marked w i l l be brought t o the
l abo ra to ry for species i d e n t i f i c a t i o n and es tab l i sh i ng of
s i n g l e egg mass popu la t i on .
b. I d e n t i f i c a t i o n of species
The species of roo t - kno t nematodes present in the root
samp les w i l l be i d e n t i f i e d by u s i n g p e r i n e a l p a t t e r n
c h a r a c t e r i s t i c s of mature females (Eisenback et al. , 1981)
and by conduct ing North Caro l ina host d i f f e r e n t i a t e t e s t s
(Tay lor and Sasser, 1978).
( i ) Perineal pattern method
For t h e i d e n t i f i c a t i o n o f s p e c i e s o f Meloidogyne
present in roo t samples, mature females w i l l be d issected out
f rom the g a l l s o f the roo t samples . P e r i n e a l p a t t e r n s o f
f e m a l e s ( 1 0 - 2 0 ) f r o m each sample w i l l be p r e p a r e d and
examined under microscope to study t h e i r c h a r a c t e r i s t i c s . The

species will be identified on the basis of perineal pattern
characteristics (Eisenback et a/., 1981).
(ii) North Carolina host differential test:
The species of Meloidogyne present in the root samples
will be maintained on tomato (cv. Pusa Ruby) or eggplant
(cv. Pusa Kranti ) in pots. Chopped roots from the samples
will be added to pots filled with steamed sterilized field
soil. Tomato or egg plant seedlings will be planted in the
pots. From these field populations, single egg nr, .se ou''. ur,j&
of the species of Meloidogyne will be raised on tomato or
eggplant grown in steam-sterilized soils in pots. For
conducting North Carolina host differential tests to
establish the identity of species, seedings of tomato cv.
Rutgers, tobacco, cv. NC95, pepper cv. California Wounder,
peanut cv. Florrunner, watermelon cv. Charlston Gery and
cotton cv. Delptapine 61 will be grown in clay pots
containing sterlized soils in pots in triplicate. Two
additional pots of tomato will be included to determine the
time of termination of test. Plants will be inoculated with
5,000 freshly hatched second stage juveniles (J2) per pots in
the vicinity of roots of test plants and pots will be kept
at glasshouse benches (temp. 27-30°C). The roots will be
harvested 50-60 days after inoculation and will be washed
thoroughly with tap water and examined for the presence of
galls and egg masses. In case of very light infection, roots

9 5
will be stained with phloxine B to visiulize the minute egg
masses. Galls and egg masses will be counted and GI and EMI
will be rated on 0-5 scale (Taylor and Sasser, 1978).
After rating the root system, result will be compared
with the differential host teet reaction chart (Table 11) to
identify four species of Meloidogyne viz., M. incognita, M.
Javanica, M. arenaria and M. hapla. The results will also
differentiats races in M. incognita and M. arenaria. Results
will be compared with identification made earlier on the
basis of perineal pattern for confirmation of their identity.
c. Maintenance of inoculum
After identification of the species, single egg mass
cultures of the species of root-knot nematodes will be raised
on tomato or eggplant seedlings in greenhouse conditions by
inoculating 10-15 egg masses of single egg mass cultures used
in North Carolina host differential tests. Sub-culturing will
be done by inoculating new seedlings of tomato or eggplant
with at least 15 egg masses, each obtained from the single
egg mass culture in order to maintain sufficient inoculum for
further studies. The results of the host differential tests
will also differentiate races in M. incognita populations.
d. Inoculation
The plants in the proposed study will be inoculated in
the form of second stage juveniles (J2)- Second stage
juveniles will be obtained by incubating egg masses from the

9 e
cultures of nematode in sterilized water in petri dishes at
25°C. The number of juveniles (J2) in the suspension will be
standardized before inoculation of the seedlings in pots. The
described amount of suspension will be taken by micropippet
controller and added near the roots into 4-5 holes made
around the seedlings. The holes will be covered with the soil
after inoculation. The inoculum level (Pi) of 0,10,100,
1000,10000 and 2000 Jg per pot is planned to be used.
2. Inocula of Fusariurn oxysporum f. sp. cicerTs and
Rhizoctonia bataticoTa
Pure cultures of the two fungi, F. oxysporum f. sp.
ciceris (Padwick) Chattopad iyay and Sen Gupta pathogenic on
causing wilt on chickpea and R. batiticola (Taub.) Butl .
causing root-rot on soybean will be obtained from the
Division of Mycology or Plant Pathology, Indian Agricultural
Research Institute, New Delhi.
a. Maintenance and pure cultuning
The fungi will be sub-cultured and maintained on PDA
slants as stock cultures. The fungi will be sub-cultured on
fresh PDA slants at regular intervals.
b. Prepration of inoculum
In order t o ob ta in enough inocu la of the fungi f o r the
expe r imen ts , Czapek's l i q u i d medium w i t h the f o l l o w i n g
cons t i t uen t s w i l l be used as a c u l t u r e medium:

3 7
TABLE 11
North Carolina differential host test reaction chart
Heloidogyne Cotton cv. Tobacco cv. Pepper cv. Naterielon cv. Peanut cv. ToBato cv. species and races Deltapine 16 NC 95 California Charleston florrunner Rutgers
Honder Grey
H. incognita
Race 1
Race 2
Race 3
Race 4
-
-
t
+
-
+
-
t
M. J3vanic3 - +
H. arenaria
Race 1 - +
Race 2 - +
H. hap I a - +
• (0 (*)
B Q
Box indicates key differential host plant Parenthesis indicate that a siall proportion of the population attacks the host + : Susceptible; - : Resistant.

9 3
KNO3 2g
KHg PO4 1 g
Mg SO4 0.5 g
Kcl 0.5 g
Sucrose 30 g
Distilled water 1000 ml.
The prepared medium will be filtered through muslin
cloth. One hundred ml. of the medium will be transfered into
250 ml. flasks and the flasks will be properly plugged with
non-absorbent cotton. Sterilization of the flasks containing
the medium will be done in an autoclave at 15 lbs. presssure
for 15 minutes. With the help of sterilized inoculation
needle, the fungal mycelium will be transfered from the PDA
slants to the flaks at laminar flow bench. Inoculated flasks
will be incubated at 23-25°C for 7 days to allow mycelial
growth of the fungi. At the end of the inoculation period,
the content of the flaks will be poured over in a funnel
lined with Whatman filter paper N0.I. The mycelium mat
collected in the funnel will be washed with distilled water
to remove the traces of the medium and mycelial mat will be
gently pressed between the folds of sterile blotting paper
to remove the excess amount of water. Inoculum in form of
suspension will be prepared by blending 10 g fungal mycelium
in 100 ml. of sterilized distilled water for 30 seconds in a

9 3
Waring blender. Thus, each 10 ml. of this suspension will
contain about 1 g of the fungus,
3. Pollutant (Sulphur dioxide):
a. Exposure chamber
Experimental plants grown in pots for the treatments
receiving SO2 exposure intermittently will be exposed to
sulphur dioxide in air pollutant exposure chamber (Standard
Appliances, Varansi) which is equipped with a full size door,
and an exhaust duct at its top to carry out the .i'if.r -;fous
mixture. The chamber's bottom is double walled, upper side
has opening, and a blower is attached to the lower side.
Voltage supply to blower will be regulated by a voltage
controller which is displayed on a panel meter. The potted
plants which are to be treated will be kept in the exposure
chamber and will be exposed for a desirable length of time.
The concentration of the gas, though regulated by the voltage
supply, will be determined by sampling of the air in chamber
and analysis of the sampled air.
b. Generation of SOg Gas
By the action of sulphuric acid (H2 SO^) on sodium
sulphite (Na2 SO3), SOg gas will be produced in a generator
under contol reaction conditions. The amount of Na2 SO3 and
H2SO4 discharged from the reagent bottles mounted over the
SO2 generator will be determined by collecting the solution
dropping through capillary tube in a graduated cylinder for

1 O O
sometime and e x p r e s s i n g the r a t e i n m l /m i n on the bas i s o f
f l o w r a t e o r s o l u t i o n f e e d i n g r a t e . S o l u t i o n o f s o i d u m
s u l p h i t e (Na2 SOo) and s u l p h u r i c a c i d (10%) w i l l be p repa red ,
t o p r o d u c e r e q u i r e d amount o f SOg g a s / m i n . On c o m p l e t e
r e a c t i o n 1M NagSOg produces 1M. SOg or 126 mg NagSOg produces
64 mg SO2. 10% Hp SO^ s o l u t i o n w i l l be used f o r a l l t h e
w o r k i n g s o l u t i o n s o f Na2S02. The r e a c t i o n i s shown below:
Nag SO3 + HgSO^ > SOg+Nag SO^ + HgO
c . Exposure and doses
Exposure of the plants will be done intermittently.
Seedlings will be exposed to SOg when they are 3-4 weeks old
at alternate days for 3 h. for each exposure. This procedure
will be continued for 75 days. The concentrations of the SOg
proposed to be used for the exposure will be 0.1 and 0.2 ppm.
d. Air pollution symptoms
The experimental plants of chickpea and soybean treated
with SO2 will be regularly examined for SOg induced symptoms.
The symptoms if present will be noted and characterized.
e . Sampl ing
Sampl ing o f t h e a i r w i t h i n t he exposure chambers w i l l
be done by Handy A i r Sampler (K imo to , Japan) a t the r a t e o f
200 m l . a i r per m inu te f o r 3 m in . For sampl ing o f gaseous a i r
p o l l u t a n t s , m e t h o d s b e i n g f o l l o w e d a t t h e N a t i o n a l

1 O 1
Environmental Engineering Research Institute (NEERI), Nagpur,
(Anon, 1986) will be employed.
f. Analysis of sulphur dioxide
Analysis of sulphur dioxide in sample will be done by
Weast and Fack method (being used at NEERI). Some details of
the analysis, preparation of chemicals are described
stepwise, below:
(i) Absorbing reagent (Sodium tetrachloromercurate)
27.2 g meruric chloride and 11.7 g sodiun. en o n o e
will be dissolved in 1 lit. double distlled water (D.D.W.).
This solution will be stored at room temperature and can be
used for two months.
(ii) Sulphamic acid (0.6%)
0.6 g sulphamic acid will be dissolved in D.D.W. to
bring the volume to 100 ml.
(iii) Formaldehyde solution (0.2X, to be prepared fresh).
0.5 ml formaldehyde solution (40%) will be diluted in
D.D.W. to 100 ml.
(iv) Rosaniline hydrochloride solution (0.2%)
a. Step I solution
0.2 g rasaniline hydrochloride will be dissolved in 1 N
hydrochloric acid to bring the volume up to 100 ml. It will
be kept in refrigerator over night.

1 0 2
b. Step II solution
4 ml . of step I and 6 ml concentrated HCl will be
dissolved in D.D.W. to bring the volume to 100 ml.
(v) Potassium dichromate solution (0.1 N)
Exactly 1.226 g dried potassium dichromate will be
dissolved in D.D.W. to 250 ml.
(vi) Sodium thiosulphate (0.1 N)
7.0 g sodium thiosulphate will be disso ecf n 250
D.D.W. For preservation 2 ml. of chloroform will also oe
added.
(vii) Iodine solution (0.1 N)
10 g of potassium iodine will be dissolved in 20 ml
D.D.W. After adding 3 g crystals of iodine, solution will be
stirred. Solution will be kept over-night and dilluted up to
250 ml with D.D.W. Solution will be preserved in amber
coloured bottle in dark.
(viii) Potassium iodine solution (100%)
10 g of potassium iodine will be dissolved in 10 ml of
D.D.W, This may be prepared at the time of titration.
(ix) Starch solution
A paste of 1.25 g soluble starch will be prepared with
D.D.W. and will be poured in 250 ml boiling D.D.W. It will be

1 0 3
boiled for 5 minutes with stirring and allowed to cool on
stand. Fresh and clear supernatent will be used.
(x) Metabisulphite solution
0.3 g sodium metabisulphite will be dissolved in 500 ml
D.D.W. This solution will contain 320 or 400 ug/ml of SOg.
g. Standardisation of sodium thiosulphate
50 ml D.D.W. will be poured into a 250 ml conical flask
and add to it 10 ml of potassium dichromate (0. ' h; •'C
H2 SO. (1:1) and 1 ml of potassium iodine solution wii be
added to it. The flask will be kept in dark for 5 minutes for
reaction.
Above solution will be titrated with sodium thio
sulphate (vi) till a faint yellow colour is obtained. Then 1
ml of starch solution will be added; blue colour will now
appear. The titration will be countinued until faint blue
colour disappears. Final colour will be distinct bluish green
tinge due to chromous ions. Titration reading will be noted.
This will be repeated to get constant reading. Then normality
of sodium thiosulphate will be calculated according to the
following formula.
Normality of dichromate x vol. of dichromate Normality of = thiosulphate Vol. of thiosulphate required

1 OA^
h. Standardisation of metabisulphite solution
25 ml of iodine solution (vii) will be taken in two 250
ml conical flasks (A & B). In A (blank) and B, 25 ml D.D.W.
and 25 ml metabisulphite solution (X) will be added. After
keeping flasks in dark for 5 min for reaction, titration will
be done with sodium thiosulphate till color become faint
yellow. Two ml starch solution will be added and titration
will be continued untill blue colour disappears which was
produced due to addition of starch. Titrstior' e
repeated to get constant readings and normality of
metabibihlphite will be calculated according to the following
formula:
(A-B) X N Normality of metabisulphite solution =
Vm
A = volume of sodium thiosulphate required for blank (flask A)
B = Volume of sodium thiosulphate required for metabisulphite
(flask B).
N = Normality of thiosulphate calculated earlier
Vm = Volume of metabiosulphite solution taken.
i. Working standard metabisulphite solution in absorbing
media
Now s o l u t i o n of metabisulphate w i l l be prepared of such
a s t r e n g t h t h a t 1 ml of s o l u t i o n c o n t a i n s 10 ug o f SO2 •
T o t a l y ug o f SOp in m e t a b i s u l p h i t e s o l u t i o n w i l l be
ca l cu la ted according to f o l l o w i n g formula:

1 0 5
Y = Normal i ty of metab isu lph i te x 32000
where Y i s t o t a l ug of SOg in metab isu lph i te s o l u t i o n :
Now w o r k i n g s t a n d a r d w i l l be p r e p a r e d i n 100 ml
abosr ing media such 1 ml = 10 ug SO2
10 X 100 Z =
Y
Z is the volume of metabisulphite required for dilution
to 100 ml with absorbing media.
j. Calibration of standard curve
Working standard metabisulphite will be pipetted in
graduated amounts (such as 0.1, 0.2, 0.3, 0.5, 0.7, 0.9, 1.0,
1.1, 1.2, 1.5 ml which will contain 1,2,3,5,7,9,10,11,12 and
15 ug SO2 respectively) into a series of impingers or
grauated nessler tubes. Absorbing media will be added to
tubes to make volume 10 ml. In the blanks 10 ml of absorbing
media will be added only. Then 1 ml sulphamic acid (ii), 2.0
ml formaldehyde solution (iii), 5.0 ml rosaniline
hydrochloric solution (iv) will be added one by one, and
after adding each, the solution will be shaken gently. After
it, the volume of each solution will be maintained with
D.D.W. up to 25 ml. After 30 minutes but befor 60 min., the
transmittance will be determined at 550 nm in
spectrophotometer. A standard curve will be drawn between
transmittance and concentration of ug SOg.

1 oe
k. Estimation
10 ml of sampled absorbing medium will be taken in
impinger. The 1 ml sulphemic acid, 2 ml formaldehyde solution
and 5 ml rosaniline will be added one by one and after
adding each, the solution will be shaken gently. After 30 min
but before 60 min transmittance will be determined at 550 nm
in specterophotometer. In the control (blank) in place of
absorbing media non-sampled medium will be taken. The ug of
SOg will be determined by placing of transmittanc^ i%
calibrated standard curve, the corresponding value (ug SOg)
will be found out. SOg ug/m will be calculated according to
following formula:
3 " 3 SO2 SO2 ug/m"* = X 10''
Volume of air sampled (lit)
Volume of air sampled = Flow rate of air x time of sampling
(minutes)
ug of SOg/m^ w i l l be converted i n t o ppm accord ing t o
the f o l l ow ing formula :
ug /m^ X 22400 ppm =
M X 10°
M i s the molecular weight of the p o l l u t a n t .
4 . Pathogenic i ty t e s t s
Pa thogen ic i t y of M. incogni ta, F. oxysporum f . sp .
ciceris, and R. bataticola {-Macrophomina phaseolina) a t the
i nocu l l um l e v e l s of 10,100,1000 and 10000 second s tage

1 O 7
juveniles (J2) and 0.5, 1.0 and 2.0g of mycelial suspension
per pot will be tested on six cultivars of chickpea and
soybean. The relative sensitivity of the cultivars of
chickpea and soybean to 0.1 and 0.2 ppm of SOg will also be
tested.
Plants in pots inoculated with the fungi will be kept
in greenhouse. Plants will be observed regularly for wilt
symptoms appearance and will be noted. The intensity of wilt
symptom, and wilting index will be determined after 15-30-
45-60 days of inoculation according to the following scale.
0 = No symptom
1 = Light symptoms
2 = Moderate symptoms
3 = Heavy symptoms
4 = Servere symptoms (dead) (Sidhu and Webster, 1978)
The relative amount of root-kot on a root system in
each treatment will be determined by scoring the extent of
disease on scale ranging a 0-5 scale (Reddy et al., 1979) as
given below:
0 = No root-rot
1 = 1-20% root-rot
2 = 21-40% root-rot
3 - 41-60% root-rot
4 - 61-80% root-rot
5 - 81-100% root-rot

1 oe
The following treatments will be inocluded for this
study:
T^ = Control
Tg = 0.5 g of mycelial mat per pot
T3 = 1.0 g of mycelial mat per pot
T^ = 2.0 g of mycelial mat per pot
Each treatment will be replicated five times. The same
treatments will be used for each cultivar.
Pathogenicity of M. incognita
Pathogenicity of root-knot nematode, W. incognita will
be tested using, 0, 10, 100, 1000 and 10000 Jg (per pot)
inoculum levels. Three week-old seedlings of six cultivars
of both crops will be inoculated in pots. Inoculated plants
will be kept in greenhouse for 50 days. At the time of
termination, plants will be uprooted gently and roots will be
washed with tap water, with care to avoid loss and injury to
the root system. Growth characters like lengths of root and
shoot; fresh and dry weights of shoot and root will be
recorded. Number of galls and egg masses per root system will
be counted. Gall index and egg mass indices will be rated
according to Taylor and Sasser's scale of 0-5 (Taylor and
Sasser, 1978). After determining GI and EMI, final population
of nematode for each cultivar will be determined by
determining numbers of eggs, female population in roots and

1 O S
soil population of the juveniles and males and Rf will be Pf
determined by Rf = (Oastenbrink, 1966). Treatments Pi
included for this study will be:
T Control
Tg = 10 Jg/pot
T3 = 100 Jg/ pot
T4 = 1000 Jg/ pot
Tg = 10000 J2/pot
The resistant and susceptibility will be allocated to
cultivars according to modified Canto-Saenez scheme (Sasser
et aJ., 1984).
Sensitivity to SOg
Relative sensitivity of all six cultivars of chick-pea
and soybean to SOg will be be tested at 0.1 and 0.2 ppm
SO2. Three-weeks-old seedlings will be exposed to SO2 in
exposure chambers every alternate days for 3 h. Exposing of
plants to SO2 will be continued for 75 days. At the end of
each exposure, plants will be transfered to the green-house.
Treatment are:
T = Control
T2 = 0.1 ppm SO2
T3 = 0.2 ppm SO2
All cualtivars of both crops will be treated and each variety
will be replicated five times.

1 1 O
5. Interact ion studies
a. Interact ion of M. incognita w i l t and root-rot fungi
I n t e r a c t i o n o f M. i ncogn i ta on c h i c k p e a w i t h F.
oxysporum f . sp. ciceris and on soybean w i th R. bataticola
{=M. phaseolina) w i l l be examined f i r s t . One suscept ib le and
one r e s i s t a n t c u l t i v a r t o the r e s p e c t i v e fungus w i l l be
s e l e c t e d c o n s i d e r i n g the r e s u l t s o f p a t h o g e n i c i t y t e s t s .
Three-weeks-o ld seed l i ng of t h e c u l t i v a r s Q-F bo th c r o p s ,
ra ised from sur face s t e r i l i z e d seeds in 30 cm <1 oott :<)
be i n o c u l a t e d w i t h J2 of the nematode. Fungal i n o c u l a t i o n
w i l l be done i n the form of mycel ium s u s p e n s i o n . The
inoculum leve l s o f the pathogens w i l l be se lec ted depending
upon the r e s u l t s of the pa thogen ic i t y t e s t s . The t reatments
w i l l be as f o l l o w
T^ = Control
T2 = Fungus
T3 = Nematode
T4 = F+N (Concomitant)
Tg = F+N (Sequent ia l , nematode 3-4 weeks p r i o r t o the
fungus) .
A f t e r i nocu la t i on of ch ick-pea and soybean c u l t i v a r s ,
the pots w i l l be kept i n greenhouse, and a f t e r a g row ing
per iod of about two months, p lan ts w i l l be removed from pots
and roots w i l l be washed to remove attached s o i l p a r t i c l e s .

1 1 1
Root gall index (GI) and egg mass index (EMI) will be rated
according to Taylor and Sasser's scale (Taylor and Sasser,
1978). In case of wilt fungus alone and the wilt fungus with
the nematode treatments, wilting indices will be recorded
according to Sidhu and Webster's (1978) scale. Root-rot
indices will be rated on 0-5 scale (Reddy et a/., 1979).
b. Interaction of SO2 with wilt and root-rot fungi
Interaction of SOp with F. oxysporum f. sp. ciceris on
chickpea and with R. bataticola {=M. phaseolina) on soybenr,
will be studied. Fungal inoculation of the seedlings will be
carried out as described earlier. Plants will be exposed to
SOg intermittently at the concentration of 0.1 ppm or 0.2
ppm, 3 h per exposures, every alternate days for about 75
days. The same varieties used for nematode-fungus interaction
studies will be used in this experiment as well. Soil pH will
be measured before starting of exposure and the time of
termination of experiments. Treatments for both the fungi
will be as follow:
T = Control (Unexposed, uninoculated)
Tp = SO2 exposed
Tg = Fungus unexposed
T^ = SO2 + Fungus

1 1 2
c. Interact ion of SOg with M. incognita
I n t e r a c t i o n of SO2 wi th M. incognita w i l l be s tud ied on
soybean and ch i ckpea . The p l a n t c u l t i v a r s used i n e a r l i e r
e x p e r i m e n t w i l l be used f o r t h i s s t u d y a l s o . A f t e r
i nocu la t i on the seed l ing , they w i l l be exposed to 0.2 ppm
SO2, 3 h per exposure every a l t e r n a t e day f o r 75 days. So i l
pH w i l l be measured before exposure to SO^ and a t the t ime of
t e rm ina t ion of experiments. Treatments w i l l be as f o l l o w s :
T^ = Contro l (Uninoculated, unexposed)
Tg = M. incognita (Unexposed)
T3 = SO2
T^ = SO2 + M. incognita
d. Interactions between SOg, wilt and root-rot fungi and M.
incognita
Interactions between SO2, F. oxysporum f. sp. ciceri&
and M. incognita on chickpea and between SO2, R. bataticola
and M. incognita on soybean will be studied. The levels of
inocula will be same as described earlier. Plants will be
exposed to SO2 every alternate day 3 h exposure for 75 days.
Soil pH will be measured before starting of exposure and at
the time of termination. Treatments will be as follows:
Unexposed set
T = Control
T2 = Fungus (F)

1 1 3
T^ = Nematode (N)
T4 = F+N (Concomitant)
Tg = F+N (Sequential, nematode 3-4 week prior to
fungus)
Exposed Set
Tg = Control
Ty = Fungus
Tg = Nematode
Tg = F+N (Concomitant)
T^Q = F+N (Sequential, nematode 3-4 weel prio. o
fungus)
In all above mentioned experiments, each treatment will
be replicated five times. An appropriate experimental design
will be used for each data will be analysed accordingly.
6. Laboratory Experiments
a. Effect of SO2 on hatching and mortility of M. incognita
Effect of SOp on hatching and juvenile mortility of M.
incognita will be studied is micro-exposure chambers. Ten egg
masses of relatively same size will be selected and will be
transfered to the small petriplates (3 cm) containing
sterilized. The petriplates containing egg masses will be
placed in micro-exposure chambers and will be exposed to
0.05, 0.1, and 0.2 ppm SO2, 3 h/day for six days. A set of
unexposed petri plates with egg masses will serve as control.
The number of hatched juveniles will be counted and hatching

1 1 ^
in relation to control will be determined. Observations will
be made after 24,48,92 and 184 h, number of motile and dead
will be noted in each observations. pH of water will be
measured before the starting of experiment and after total
exposures.
b. Effect of acidified soil on hatching and roortility of M.
incognita
Effect of acidified soil at pH levels of 7.0, 6.0, 5.0,
4,0, and 3.0 on hatching and mortality of M. incognita wi'l
be studied. Ten egg masses of relatively same size will be
transferred into the paper cups containing approximately 100
g of water washed sterilized sand. 25 ml of acidic solutions
prepared will be added to each cup. Observations will be made
8 days after addition of acidic solution. Number of mobile
and dead nematodes will be noted. Sand pH will be measured
before and after addition of acidic solutions.
7. Measurment of soil pH
To measure soil pH, soil suspension in water will be
prepared in the ratio of 1:10. To achive this, soil sample
will be crushed and mixed thoroughly. Then 10 g of soil will
be poured in 250 ml. Conical flask containing 100 ml. of
double distilled water and will be agitated for one hour. The
suspension will be filtered through filter paper and filtrate
will be used for the measurement of soil pH by glass
electrode.

1 1 5
8. Rais ing of seedl ings
Seeds w i l l be sown i n a mixture of s o i l , sand and farm
yard manure in the r a t i o of 2 :1:1 respec t i ve l y , contained in
30 cm c lay pots which are autocalved f o r s t e r i l i z a t i o n at 20
l b s . f o r 30 minutes. The s t e r i l i z e d pots w i l l be al lowed to
c o o l down a t room t e m p e r a t u r e b e f o r e u s i n g f o r t h e
exper iments.
Before sowing, seeds w i l l be surface s t e r i l i z e d w i th
0 . 1 % m e r c u r i c c h l o r i d e f o r 2 m i n u t e s and washed w i t h
s t e r i l i z e d water t h r i c e , t o remove t he t r a c e o f m e r c u r i c
c h l o r i d e . Seeds w i l l be sown in s t e r i l i z e d s o i l a t the ra te
o f f o u r seeds per p o t . F i f t e e n days a f t e r g e r m i n a t i o n ,
seed l ing w i l l be th inned to one in each pot . Three-weeks-old
s e e d l i n g s (approx . i n 4-5 leaves s t a g e ) w i l l be used f o r
t rea tments . Regular i r r i g a t i o n w i l l be done to keep up the
s o i l mois ture .
9. Rhizobium i n o c u l a t i o n
Seeds in all the experiments related to chickpea and
soybean will be inoculated with their respective strains of
Rhizobium before sowing. Commercial sugar and water will be
added in the soil based culture with thorough mixing. The
seeds of the test crops will be treated with this mixture
followed by the drying in shade for about half an hour before
sowing.

116
10. Parameters
The following parameters will be considered depending
upon the kind of experiments, for the potted plants, related
to interactions, pathogenicity etc.
Fresh and dry weights of shoQt and root
Length of shoot and root
Number of galls/root system
Number of egg masses/ root system
Gall index (GI)
Eggmass index (EMI)
Soil and root population of the nematode
Pf Reproduction factor (Rf = )
Pi
Total number of nodules / plant
Total number of functional nodules/ plant
Number of days for appearence of wilt symptoms
Wilt index at different intervals
Root-rot index (on 0-5 scale)
11. Root nodule bacteria
To assess the impact of selected pathogens, singly or
in combinations on root nodulation, functional, non
functional and total number of nodules per root system in
each experiments will be counted individually. The pinkish,
healthy nodules will be considered as functional and others
as non-functional.

LITERATURE CITED

Abawi, G. S. and K.R. Barker 1984, E f f e c t o f c u l t i v a r , s o i l
t e m p e r a t u r e , and p o p u l a t i o n l e v e l o f Meloidogyne
incognita on r o o t n e c r o s i s a n d Fusarium w i l t o f
tomatoes . P h y t o p a t h o l o g y , 74: 4 3 3 - 4 3 8 .
Abawi, G .S . ; D.C. C r o s i e r and A .C. Cobb 1985. R o o t - r o t o f
snapbean i n New York . New Y o r k ' s Food and L i f e Sc ience
B u l l e t i n 110: 7.
Abdul Salam, M. and M. Wa j i d Khan 1986. R e a c t i o n o f some
c u l t i v a r s o f p i g e o n p e a a g a i n s t Fusarium udum and
Meloidogyne Javanica. I n t e r n a t i o n a l Nematology Network
N e w s l e t t e r , 3 : 16 -17 .
A i r P o l l u t i o n 1 9 6 8 . The e f f e c t o f a i r p o l l u t i o n on
nonvascu la r p l a n t s . P roceed ings F i r s t European Congress
on I n f l u e n c e A i r P o l l u t i o n P l a n t s and A n i m a l s ,
Wagenengen, 2 3 7 - 2 4 1 .
A l -Hazm i , A .S . 1985. I n t e r a c t i o n o f Meloidogyne incognita
and M. phaseol ina i n r o o t - r o t d i s e a s e complex o f f r e n c h
bean. P h y t o p a t h o l o g y , 113; 311 -316 .
A l m e d i a , R . T . 1 9 7 2 . B e h a v i o u r o f v a r i e t i e s o f t o m a t o
{Lycopersicum esculantufn M i l l . ) t o Fusarium x
Meloidogyne complex. R e v i s t o do Soeiedade B r a s i l e n a de
F i t o p a t o l o g i a , 5: 11-17 ,
A l v a r e z , A.M. and G.C. B r i n g e r 1987, R o o t - r o t o f ch i ckpeas
caused by Fusarium solani ( M a r t . ) Sacc . A g r i c u l t u ^ a
T e c m c a , 47 : 78-79 .

1 1 3
Amundson, R.G. 1983. Yield reduction of soybean due to
sulphur dioxide and nitrogen dioxide in combination.
Journal of Environmental Quality, 12: 454-459.
Anahosus, K.H. and S.T. Naik 1986. Changes in sugar and
phenol contests of root and stalk of sorghum due to M. phaseol ina. Indian Phytopathology, 39: 440-441.
Anonymous, 1966. Waste mangment and Control. National
Academic Science (U.S.A.) Publication, No.1400.
Anonymous, 1986. Air quality monitoring: A course Manua'i .
National Environmental Engineering Research Institute,
Nagpur, India.
Apt, J.W. and H. Koike 1962. Pathogenicity of Meloidogyne incognita acrita and its relation with Pythium
graminicola on sugarcane in Hawai. Phytopathology, 52:
1180-1184.
Astanin, L.P, and K.N. Blagosklonov 1983. Conservation of
Nature. Progress Publishers, Moscow (U.S.S.R.), pp.32-
51 .
Atkinson, G.E. 1892. Some disease of cotton. Alabama
Polytechnic Institute of Agriculture Experiment Station
Bulletin, 41: 61-65.
Avdulov, N.P. 1937. The karyotype of Cicer ar i eti num L. Memoirs University Saratov, 1: 30-36.
Azam, M.F., A.M. Khan and S.K. Saxena 1977. Effects of
extracts of roots mfected with root-knot nematodes on
the growth of R. solani, Pythium spp. and Col letotnchum

1 1 9
atramentarium. Indian Journal of Nematology, 7: 182.
Bannfield, W.M. 1972. Ozone synergisimn on eastern while
pine. Phytopathology, 62: 439 (Abstr.).
Barrett, T.W. and H.M. Benedict 1970. In: J.S. Jacobson and
A.C. Hin, eds. Recognition of Air Pollution Injury to
Vegetation: A Pictorial Atlas. Air Pollution Control
Association, Pittsburgh, Pa. Sect. C, pp.17.
Bassus, W. 1968. The effect of industrial emissions on the
populations of nematodes in the soil o-f p-ino •,'•>.-«=:+:s.
Pedobiologia, 8; 289-295.
Bell, J.N.B. 1982. Sulphur dioxide and the growth of
grasses. In: Effects of Gaseous Air Pollution in
Agriculture and Horticulture, (eds. M.H. Unsworth and
D.P. Ormord). Butterworth Scientific, London, pp.225-
246.
Bell, J.N.B. and W.S. Clough 1973. Depression in yield of
rye-grass exposed to SOj. Nature, 241: 47-49.
Bender, J., H.J. Jeager, G. Seufert and U. Arndt 1986.
Metabolic effect of SOp and Oo singly and in combination
on forest trees in open top chambers. ANGEW Botaik, 60:
461-479.
Benedict, H.M. and W.H. Breen 1955a, Development of
standards for evaluating vegetation damage caused by air
pollution. Stanford Research Institute of Technology
Report, pp.11.

1 2 0
Benedict, H.M. and W.H. Breem 1955b. The use of weeds as a
means of evaluating vegetation damage caused by air
pollution. Proc. 3rd National Air Pollution Symposium,
pp.117-190.
Bennelt, J.H. and A.C. Hill 1973. Absorption of gaseous air
pollutant by a standardized plant canopy. Journal of Air
Pollution Control Association, 23: 203-206.
Bennett, J.H. and A.C. Hill 1974. In: Dugger W.M. Jr. Ed.,
Air pollution effects on plant growth. American Chftm-•-.a''
Society, Washington, D.C., Chap. 10, pp.115-1??.
Benwart, W.L. 1987. Response of agricultural crops to acid
rain. Pollution Abstract, 18 (4): 1088.
Bergeson, G.B. 1972. Concepts of nematode—fungus
associations in plant disease complex - a review.
Experimental parasitology, 82: 310-314,
Bergeson, G.B. 1975. The effect of Meloidogyne incognita on
the resistance of four muskmelon varieties to Fusarium
wilt. Plant Disease Reporter, 59: 410-413.
Bergeson, G.B., S.D. Van Gundy and I.J. Thomason 1970.
Effect of M. Javanica on rhizosphere microflora of
Fuserfum wi It of tomato. Phytopathology, 60: 1245-1249.
Berry, C.R. 1967. An exposure chamber for forestry air
pollution studies. Phytopathology, 57 (8): 804 (Abstr.).

1 2 1
Berry , C.R. 1971. Re la t ive s e n s i t i v i t y of red jack and whi te
pine seedings t o O3 and SO2. Phytopathology, 6 1 : 231-
232.
Berry, C.R. 1974. Age of pine seedl ings w i th primary needles
a f f e c t s s e n s i t i v i t y t o ozone and s u l p h u r d i o x i d e .
Phytopathology, 64: 207-209.
Bevan, R.J. and G.H. Greenhalgh 1976. Rhytisma acerincim as
a b i o l o g i c a l i n d i c a t o r of p o l l u t i o n . Env i ronmen ta l
P o l l u t i o n ( A ) , 10 : 2 7 1 - 2 8 5 .
B h a t t i , M.A., S. A l i and I .U . Khan 1987. Pathogenic i ty of
f u n g i a s s o c i a t e d w i t h w i l t o f ch i ckpea w i t h s p e c i a l
re ference t o Verticinium albo-atrum. Pakistan Journal
o f A g r i c u l t u r e Research, 8 ( 1 ) : 39-42.
B i n d e r , E. and M.T. H u t c h i n s o n 1959. F u r t h e r s t u d i e s
c o n c e r n i n g t h e e f f e c t o f t h e r o o t - k n o t nematode ,
Meloidogyne incognita acrita on s u s c e p t i b i l i t y o f t he
Chesapeake t o m a t o t o Fusar i urn w i l t . P l a n t D i s e a s e
Reporter , 43: 972-978,
B isessar , S. and K.T. Palmer 1984. Ozone an t iox idan t spray
and Meloidogyne hepla e f f e c t s on tobacco {Nicotiana
tabacum) c u l t i v a r V i r g i n i a - 1 1 5 . A tmosphere and
Environment, 18 (5 ) : 1025-1028.
Bisessar , S. , R.J. Rinne and J.M. Pot ter 1983. E f fec ts of
heavy metals and Meloidogyne hapla on celery growth on
organic s o i l near n icke l r e f i n e r y . Plant Disease, 6: 11-
14.

1 2 2
Bleasdale, J.K.A. 1952. Air pollution and plant growth. In:
Smokeless Air, report in Journal of Park Administration,
18: 300-301.
Bleasdale, J.K.A. 1973. Effects of coal-smoke pollution
gases on the growth of ryegrass {Lolium perenne L.). Environmental Pollution, 5: 275-285.
Bowman, P. and J.R. Bloom 1966. Breaking the resistance of
tomato varieties to Fusarium wilt by Meloidogyne incognita. Phytopathology, 56: 871 (Abstr.).
Brandat, C.S. and W.W. Heck 1968. Effects of air pollution
on plants. In: Air pollution (ed. A.C. Sterm). Academic
Press, New York, Vol.1, pp.401-443.
Brennan, E., I. Lenon and P.M. Halisky 1970. Response of
turf grass cultivars to ozone and sulphur dioxide in the
atmosphere. Phytopathology, 60: 1544-1546.
Brewer, B.J. 1979. Interaction of ozone, sulphur dioxide and
Pratylenchus species on tomato and soybean. MS Thesis.
North Carolina State University, Raligh, North Carolina.
Brodie, B.B. and W.E. Cooper 1964. Relation of parasitic
nematodes to postemergence sampling-off of cotton.
Phytopathology, 54: 1023-1027.
Bruton, B.D. and C M Heald 1987. Effect of Macrophomina phaseolina and nematodes on cantaloupe. Phytopathology,
77 (12) : 1713.

- 1 2 3
B u r p e e , L . L . a n d J . R . B loom 1 9 7 4 . I n t e r a c t i o n o f
Vertici11ium albo-atrum and Pratylenchus penetrans on
p o t a t o . P h y t o p a t h o l o g y , 64 ( 5 ) : 579.
Burpee, L .L . and J . R . Bloom 1978. I n f l u e n c e of Pratylenchus
penetrans on t h e i nc idence and s e v e r i t y o f Vertici11ium
w i l t o f p o t a t o . Journa l o f Nemato logy, 10 ( 1 ) : 9 5 - 9 9 .
B y a d g i , A . S . and R.K. Hedge 1985 . V a r i a t i o n among t h e
i s o l a t e s o f R. bataticoJa f r om d i f f e r e n t hos t p l a n t s ,
I n d i a n P h y t o p a t h o l o g y , 38 ( 2 ) : 2 9 7 - 3 0 1 .
B y t n e r o w i c z , A . , D.M. O lszyk , G. K a t s , P . J . Dawson, J . Wol f
and C.R, Thompson 1987. E f f e c t o f SO2 p h y s i o l o g y
e l e m e n t a l c o n t e n t and i n j u r y d e v e l o p m e n t on w i n t e r
wheat . A g r i c u l t u r a l Ecosystem and Env i ronment , 20 : 37 -
47 .
Cameron, C.A. 1974. The i n f l u e n c e o f chemica l e x h a l a t i o n s on
a g r i c u l t u r e . Gard . Chron. ( London ) , 1 : 274-275.
Caqj tu i l , J . and R . L . Shepherd 1 9 7 0 . E f f e c t o f r o o t - k n o t
nematode- fung i comb ina t ions on c o t t o n s e e d l i n g d i s e a s e .
P h y t o p a t h o l o g y , 60: 4 4 8 - 4 5 1 .
C a r l s o n , R.W. 1983 . I n t e r a c t i o n b e t w e e n SOg and NOg and
t h e i r e f f e c t s on p h o t o s y n t h e t i c p r o p e r t i e s o f soybean,
Glycine max. Env i ronmenta l P o l l u t i o n , (A) 32: 11 -38 .
C a r t e r , W.W. 1975. E f f e c t of s o i l t e x t u r e on the i n t e r a c t i o n
between Rhi zoctonia sol ani and Meloidogyne incognita on
c o t t o n s e e d l i n g s . Journa l o f Nemato logy, 7: 234-236 .

1 2 - 4 -
C a r t e r , R.W. 1 9 7 8 . The r o l e o f p e r o x i d a s e and
po lypheno l o x i d a s e i n the Meloidogyne Fusanum d i s e a s e
complex o f t o m a t o . D i s s e r t a t i o n A b s t r a c t I n t e r n a t i o n a l ,
39 B ( 2 ) : 466 -467 .
C a r t e r , R.W., G.B. Bergeson and R.S. Green 1977. Enhancement
o f Fusariurn w i l t o f t oma to by Meloidogyne incognita.
P r o c e e d i n g s o f he Amer ican P h y t o p a t h o l o g i c a l S o c i e t y
(1977, p u b l i s h e d 1978), 4 : 124.
C h a h a l , P .P .K . and H.K. Chhabra 1 9 8 4 a . I n t e r a c t i o n o f
Meloidogyne incognita w ^ t h Rhizoctonia on t o m a t c
I n d i a n J o u r n a l o f Nematology, 14 ( 1 ) : 5 6 - 5 7 .
Chaha l , P.P.K. and H.K. Chhabra 1984b. E f f e c t o f Meloidogyne
incognita and Rhizoctonia on t h e emergence and damp ing-
o f f o f t o m a t o s e e d i n g s . J o u r n a l o f R e s e a r c h , P u n j a b
A g r i c u l t u r e U n i v e r s i t y , 21 ( 4 ) : 642-644 .
Chaha l , P.P.K. and H.K. Chhabra 1985. E f f e c t o f Meloidogyne
incognita and Fusarium equiseti on g rowth c h a r a c t e r s and
w i l t i n g o f pea s e e d l i n g s . J o u r n a l o f Research , Pun jab
A g r i c u l t u r e U n i v e r s i t y , 22 ( 4 ) : 684-686 .
Chavada, J . C ; B.A. Pa te l and D . J . Pa te l 1988. R e a c t i o n o f
some greengram v a r i e t i e s t o r o o t - k n o t nematodes. I n d i a n
Jou rna l o f Nemato logy, 18 ( 1 ) : 127.
Chhabra , H.K. and J . K . Sharma 1 9 8 1 . Comb ined e f f e c t o f
Meloidogyne incognita and Rhizoctoma bataticola on p r e -
emergence d a m p i n g - o f f o f o k r a and b r i n g a l . Sc ience and
C u l t u r e , 47 ( 7 ) : 256-257.

1 2 5
C h h a b r a , H.K. A . S . S idhu and I . S i n g h 1 9 7 7 . Meloidogyne
incognita and Rhizoctonia i n t e r a c t i o n on o k r a . I n d i a n
J o u r n a l o f Nematology, 7: 54 -57 .
Chhabra , H.K. ; A . S . Sidhu and I . S ingh 1978. I n f l u e n c e o f
s o i l t ype on t h e i n t e r a c t i o n o f fungus Rhizoctonia and
r o o t - k n o t nematcde , Meloidogyne incognita on o k r a .
Haryana Jou rna l o f H o r t i c u l t u r a l Sc inece , 7 ( 3 / 4 ) : 213-
215.
C h i b a , 0 . and K. Tanaka 1968. The e f f e c t o f su l ph t • ' i ox ide
on t h e deve lopmen t O-P p i n e n e e d l e b l i g h t c a u s e d by
Rhizosphera kalkhoffil Bubok ( I ) . J o u r n a l o f J a p a n
F o r e s t S o c i e t y , 50: 135-139.
C h i t i m a - M a t s i g a , R.T. and T . D . W y l l i e 1 9 8 7 . V a r i a b i l i t y
among s i n g l e c o n i d i a l i s o l a t e s o f Macrophomina
phaseolina. P h y t o p a t h o l o g y , 77 ( 1 2 ) : 1702.
C o l i n , K . B . and N . T . P o w e l l 1 9 7 1 . The Rhizoctonia -
Meloidogyne d i s e a s e complex i n f l u e - c u r e d t o b a c c o .
J o u r n a l o f Nematology, 3 ( 2 ) : 164-169.
Conci , V.C. A. Vasquez and S.F. None 1985. Study o f t h e
fo rm o f t r a n s m i s s i o n o f F. oxysporum f . s p . ciceris on
ch i ckpea {Cicer arietinum). F i t o p a t o l o g i a , 20 ( 2 ) : 57 -
64 .
Cook, R . J . and K.F. Baker 1983. The n a t u r e and p r a c t i c e o f
b i o l o g i c a l c o n t r o l o f p l a n t p a t h o g e n s . S t . P a u l ,
M inneso ta , American P h y t o p a t h o l o g i c a l S o c i e t y , p p . 5 3 9 .

1 2 6
Cooper, W.E. and B.B. Brodie 1963. A comparison of Fusarium-
w i l t i n d i c e s o f c o t t o n v a r i e t i e s w i t h r o o t - k n o t and
s t i n g nematodes as predisposing agents. Phytopathology,
53: 1077-1080.
C o n s t a n t i n i d o u , H. T , T . K o z l o w s k i and K. Jensen 1976.
E f fec ts of sulphur d iox ide on Pinus resinosa seedl ings
i n t h e c o t y l e d o n s t a g e . J o u r n a l o f E n v i r o n m e n t a l
Q u a l i t y , 5: 141-144.
Costonis, A.C, 1971. E f fec ts of sulphur diox-ide anc ozop- on
e a s t e r n w h i t e p i n e i n r u r a l e n v i r o n m e n t .
Phytopathology, 61 : 717-720.
Couey. H.M. 1965. I n h i b i t i o n o f germinat ion of ATternaria
s p o r e s by s u l p h u r d i o x i d e under v a r i o u s m o i s t u r e
c o n d i t i o n s . Phytopathology, 55: 525-527.
Couey, H.M. and M. Uota 1961. E f f e c t o f c o n c e n t r a t i o n ,
exposure t ime , temperature and r e l a t i v e humidi ty on the
t o x i c i t y of sulphur d iox ide t o the spores of Botrytis
cinerea. Phytopathology, 5 1 : 815-819.
Cowling, D.W. and D.R. Lockyer 1976. Growth of perenn ia l
r y e g r a s s (Lolium perenne L . ) exposed t o low
c o n c e n t r a t i o n o f s u l p h u r d i o x i d e . J o u r n a l o f
Experimental Botany, 27: 411-417.
Daines, R.H. 1968. I n : Processings of the Symposium on A i r
Qua l i t y C r i t e r i a . New York, pp.84-92.

1 2 y
D a r l e y , E .F . 1966. S t u d i e s on damage t o v e g e t a t i o n f r o m
c e m e n t - k i l n d u s t . J o u r n a l o f A i r P o l l u t i o n C o n t r o l
A s s o c i a t i o n , 16; 761-770.
D a v i s , C.R. 1972. Su lphur d i o x i d e f u m i g a t i o n o f soybeans.
E f f e c t s on y i e l d . J o u r n a l o f A i r P o l l u t i o n C o n t r o l
A s s o c i a t i o n , 22 : 964-966.
D a v i s , T. 1980. Grasses more s e n s i t i v e t o SOg p o l l u t i o n i n
c o n d i t i o n o f low i r r a d i a n c e and s h o r t days . N a t u r e , 284:
483-485 .
D a v i s , R.A. and W.R. Jenk ins 1963. E f f e c t s o f Meloidogyne
spp . and Tylenchorhynchus clay toni on pea w i l t i n c i t e d
by Fusarium oxysporum f . s p , pisi R a c e - 1 . P h y t o p a t h o l o g y ,
53 : 475 .
D e C o r m i s , L. 1969. I n : A i r P o l l u t i o n : P r o c e e d i n g s o f t h e
f i r s t E u r o p e a n C c o n g r e s s on t h e I n f l u e n c e o f A i r
P o l l u t i o n on P l a n t s and A n i m a l s . Wageningen, p p . 7 5 - 7 8 .
Dekok, L . J . , C.R. Thompson and J . B . Mudd 1982. E f f e c t s o f
HgS on w a t e r - e x t r a c t a b l e s u l p h y d r y l c o n t e n t o f c r o p
p l a n t s . I n : E f f e c t s o f g a s e o u s a i r p o l l u t i o n i n
a g r i c u l t u r e and h o r t i c u l t u r e , ( eds . M.H. Unsworth and
D.P. Ormord) . B u t t e r w o r t h S c i e n t i f i c ( London ) , p p . 5 0 1 .
D e l g a d o , M.A. and V .R. A g u r t o 1 9 8 4 . E s t i m a t i o n o f l o s s e s
caused by r o o t and f i b r o v a s c u l a r pathogens by means o f
samp l i ng and p e r i o d i c e v a l u a t i o n o f t he p l a n t p o p u l a t i o n
i n d i f f e r e n t c o t t o n g r o w i n g a r e a s o f P i u r a .
F i t o p a t o l o g i a , 19 ( 1 ) : 27 -38 .

1 2 8
Dhangar , D,S. and D.C. Gupta 1983 . P a t h o g e n i c i t y o f
MeToidogyne Javanica to chickpea in r e l a t i o n to s o i l
t y p e , Rhizobium t r e a t m e n t , s i z e o f p l o t and t i m e
i n t e r v a l . Ind ian Journal of Nematology, 13 ( 2 ) : 161-
170.
D h i n g r a , O.D. and J . B . S i n c l a i r 197 3. L o c a t i o n o f
Macrophomina phaseolina on soybean, r e l a ted to c u l t u r a l
c h a r a c t e r i s t i c and v i ru lence . Phytopathology, 63: 934.
D h i n g r a , O.D. and J . B . S i n c l a i r 19"?^. CU' . ' •.-'•
Macrophomina phoseol ina s c l e r o t i a i n s o i V L ' .-C C j f
s o i l mo is ture , carbon, n i t rogen r a t i o s , carbon sources,
and n i t rogen concent ra t ions . Phytopathology, 65: 236-
240.
Donald, R.S. and A.W. Johnson 1973. E f f e c t o f r o o t - k n o t
nematodes on Fusari urn w i l t o f w a t e r m e l o n .
Phytopathology, 63 ( 7 ) : 857-862.
Donaubauer, E. 1968, Sekundarschaden i n Os te r re i ch i s chen
Rauchschadens gebieten. Schwier igkei ten Der diagnose Und
Bewertung. Niedzynarodowj Konf . Wplyw Z a n i e c z y s z c z e n
Powietraza na Lasy, 6th Katowice, Poland.
D o r r i e s , W. 1932. Uber d ie Brauchbarkei t der spect roscop ic
p h a o p h y t i n p robe i n der R a u c h s h a d e n - D i a g n o s t i k .
Z e i f s c h r i f t f u r Pflanzen Krankheiten Kr. Ga l l enk . , 421 :
257-273.
D re i s i nge r , B.R. and P.C. McGovern 1970. I n : S.N. Linzon Ed.
Proceed ings ; Impac t . o f a i r p o l l u t i o n on v e g e t a t i o n ,
s p e c i a l i t y conference. Toranto, On ta r io , A i r P o l l u t i o n
Contro l Assoc ia t ion , P i t t sbu rgh , Pa. , pp.11-28.

1 2 9
E i s e n b a c k , J . D . , H. H i r s c h m a n n , J . N . Sasse r and A . C .
T r i an taphy l l ou 1981. A guide t o the fou r most common
species of roo t -kno t nematodes (Meloidogyne s p p . ) . North
C a r o l i n a S ta te U n i v e r s i t y G e r a p h i c s , R e l a i g h , N .C . ,
U.S.A., pp.48.
Edward, J .C . and K.P. Singh 1979. I n t e r a c t i o n between
Heterodera cajani and Fusar iurn udum on p i g e o n p e a .
Al lahabad Farmer, 50 ( 1 ) : 23-24.
F a s s u l i o t i s , G. and G.J. Rau 1969. The r e l a t i ont:.N i i. of
Meloidogyne incognita acrita on the inc idence of cabbage
ye l lows . Journal of Nematology, 1 : 219-222.
Fa t tah , F.A. and J.M. Webster 1983. U l t r a s t r u c t u r a l changes
caused by Fusari um oxysporum f . s p . lycopers i ci i n
Meloidogyne Javanica induced g i a n t c e l l s i n Fusarium
r e s i s t a n t and suscept ib le tomato c u l t i v a r s . Journal of
Nematology, 15: 128-135.
Fawcett, H.S. 1931. The importance of i n v e s t i g a t i o n s on the
e f f e c t s of known mixtures of organisms. Phytopathology,
2 1 : 545-550.
Fe r raz , S. and B. Lear 1976. I n t e r a c t i o n o f f o u r p l a n t
p a r a s i t i c nematodes and Fusarium oxysporum f . s p . dianthi
on c a r n a t i o n . Exper ien t ia , 22 (11 ) : 272-277.
F luck inger , R.B. and V.B. Younger 1986. Ozone and sulphur
d i o x i d e e f f e c t s on t a l l f escue I I . A l t e r n a t i o n o f
q u a l i t y cons i t u t en t s . P o l l u t i o n Abs t rac t , 17: 157.

1 3 0
Fukuda, Y. 1933. Cytogenetical studies in the wild and
cultivated Manchurian soybean. Japanese Journal of
Botany, 6: 489-506.
Gangopadhyay, S., T.D. Wyllie and V.D. Luedders 1970.
Charcoal rot disease of soybean transmitted by seeds.
Plant Disease Reporter, 54: 1088-1091.
Garber, R.H. E.G. Jorgenson, S. Smith and A.H. Hyer 1979.
Interaction of population levels of Fusarium oxysporum f.sp. vasinfectum and Meloidogyne incognita on rr^tton
Journal of Nematology, 11: 133-137.
Garica, J.L., L.A. Gonzalez and F. Perez 1985. Incidence of
soil fungi on soybean (Glycine max) plants in tobacco grown zones of Pinar de Rio, Cuba. Ciencias de la
Agricultura, 25: 23-28.
Ghosh, S.K. and C. Sen 1973. Comparative physiological
studies on four isolates of Macrophomina phased ina. Indian Phytopathology, 26: 615-621.
Gilbert, O.L. 1969. In: Air Pollution: Proceedings of the
First European Congress on the Influence of Air
Pollution on Plants and Animals. Wageningen, pp.223-
235.
Gill, D.L. 1958. Effect of root-knot nematodes on Fusarium wilt of mimosa. Plant Disease Reporter, 42: 587-590.
Godzik, S. and H.F. Linskens 1974. Concentration changes of
free amino acid in primary bean leaves after continuous

1 3 1
and i n t e r r u p t e d SO2 f u m i g a t i o n and recove ry .
Environmental Po l lu t ion , 7: 25-38.
Goel, S.R. and D.C. Gupta 1984. Interact ion studies between
Fusarium oxysporum f . s p . ciceris, F. sol an i and
Meloidogyne javanica on chickpea. Proceeding of F i rs t
Internat ional Congress of Nematology, Canada: 36.
Goel, S.R. and D.C. Gupta 1986. Interact ion of Meloidogyne.
javanica and Fusarium oxysporum f . s p . ciceris on
chickpea. Indian Phytopathology. 39 (1)- 112- v ^
Goode, M.J. and J.M. McGuire 1967. Relationship of root-knot
nematodes to pathogenic v a r i a b i l i t y in Fusar i urn
oxysporum f . s p . lycopersici. Phytopathology, 78: 812
(Abs t r . ) .
Gorham, E. and A.G. Gorden 1963. Some effects of smelter
po l lu t ion upon aquatic vegetation near Sudbury, Ontr io.
Canadian Journal of Botany, 41: 371-378.
Goswami, B.K., M.L. Sethi, J.N. Gupta and D.V. Singh 1976.
I n t e r r e l a t i o n s h i p of Meloi dogyne Javani ca and
Rhi zoctonia bataticola in tomato. I n d i a n
Phytopathology, 28 (3) : 387-388.
Goswami, D.V., M.L. Singh and J.N. Gupta 1970. Studies on
association of root-knot nematode, Meloidogyne incognita
and Sclerotium r o l f s i i i n b r i n j a l . I n d i a n
Phytopathology, 23 (3): 587-589.

1 3 5*
G r i f f i n , G.D. 1986. The impor tance o f nematode r e s i s t a n c e
on t h e i n t e r a c t i o n o f Meloi dogyne hapla and Fusar i urn
oxysporum on a l f a l f a . P h y t o p a t h o l o g y , 46 : 843 ( A b s t r . ) .
G r i f f i n , G .D . , and K.H. Asay 1989. P a t h o l o g i c a l r e a c t i o n o f
C r e s t e d W h e a t g r a s s c u l t i v a r s t o f o u r Me 1o i dogyne
Chitwoodi p o p u l a t i o n s . J o u r n a l o f Nematology, 21 ( 4 ) :
446 -452 .
G rogan , R.G, 1 9 8 1 . The s c i e n c e and a r t o f p l a n t d i s e a s e
d i a g n o s i s . Annua"i Review of P h y t o p a t h o l o g y , ' ' 9 : 3C5
351 .
G r o v e r , R.K. and P.K. S a k h u j a 1 9 8 1 . Some p a t h o l o g i c a l
s t u d i e s on Rhi zoctoni a batati col a l e a f b l i g h t o f
mungbean. I n d i a n P h y t o p a t h o l o g y , 34: 24 -29 .
G r u j i c i c , G. , M. M a r t i n o v i c , B. B o r i c and 0 . K r i s t i c 1984.
Pratylenchus penetrans Cobb, and F. oxysporum v a r .
medicaginis Snyder and Hansen on l u c e r n e {Medicago
sativa L . ) . Z a s t i t a B i l j a , 35 ( 4 ) : 3 0 7 - 4 0 1 .
G u d e r i a n , R. and H. Stratmann 1962. F i e l d e x p e r i m e n t s t o
de te rm ine the e f f e c t o f SOo on v e g e t a t i o n . I . Survey o f
method and e v a l u a t i o n o f r e s u l t s . F o r s c h u n g s b e r i c h t e
Landes N o r d r h e i n W e s t f a l e n , 1118: 102.
Gupta , D.C. , I . J . Pa ru th i and K.K. Verma 1986. R e a c t i o n o f
m u n g b e a n g e r m p l a s m and i t s p a t h o g e n i c i t y a g a i n s t
Meloidogyne javanica. I n d i a n Jou rna l o f Nemato logy, 15
( 2 ) : 194-196.

1 3 3
Hadiastono, T. 1981. The influence of Meloidogyne spp. on
tomato damage caused by Fusarium oxysporum. Agririta,
4(3): 19-20.
Ham, D.L. 1971. The biological interactions of sulphur
dioxide and Sacci rhia acicola on loblolly pine, Ph.D.
Thesis, Duke University, Durham, N.C., pp.74.
Harrison, A.L. and P.A. Young 1941. Effect of root-knot
nematodes on tomato wilt. Phytopathology, 31: 749-752.
Hasan, A. 1984. Synergism between Heterodera cajani and
Fusarium udum attacking Cajanus cajan. Nematologia
Mediterranea, 12: 159-162.
Haselhoff, E. and G. Lindau 1903. Die Beschadigung der
vegetation durch Rouch. Brontraeger, Leipzig.
Hazarika, B.P. and A.K. Roy 1975. Effects of Rhizoctonia sol am' on the reproduction of Meloidogyne incognita on
eggplant. Indian Journal of Nematology, 4 (2): 246-248.
Heagle, A.S. 1973. Interactions between air pollutants and
plant parasites. Annual Review of Phytopathology, 11:
365-388.
Heag le , A . S . 1982. I n t e r a c t i o n between a i r p o l l u t a n t s and
p a r a s i t i c p l a n t d i s e a s e s . I n : E f f e c t s o f Gaseous A i r
P o l l u t i o n i n A g r i c u l t u r e and H o r t i c u l t u r e . (Eds . M.H.
U n s w o r t h and D.P. O r m o r d ) . B u t t e r w o r t h S c i e n t i f i c
( L o n d o n / B o s t o n ) , p p . 3 3 3 - 3 4 8 .
H e a g l e , A . S . and J.W. Jhons ton 1979. V a r i a b l e responses o f
s o y b e a n t o m i x t u r e o f o z o n e and s u l p h u r d i o x i d e .

1 34-
Journal of Air Pollution Control Association; 29-729-
732.
Heck, W.W. 1982. Future directions in air pollution
research. In: Effect of Gaseous Air Pollution in
Agriculture and Horticulture. (Eds. M.H. Unsworth and
D.P. Ormord). Butterworth Scientific (London/ Boston),
pp.542.
Heck, W.W. and C.S. Brandt 1975. In; A.C. Stern, Ed., Air
Pollution, Vol. I, 3rd Ed., Academic Press, New York,
Chap. 14.
Heck, W.W.,O.C. Taylor, R. Adams, G. Bingham and J. Miller
1982. Assesment of crop loss from ozone. Journal of Air
Pollution Control Association; 32: 351-361.
Herman, M., R.S. Hussey and R. Boerma 1988. Interaction
between Meloidogyne incognita and Praty 1 enchus
brachyurus on soybean. Journal of Hematology, 20 (1):
79-84.
Hibben, C.R. 1966. Sensitivity of fungal spores to sulphur
dioxide and ozone. Phytopathology 56: 880 (Abstr.).
Hibben, C.R. and M.P. Taylor 1975. Ozone and sulphur dioxide
effects on the lilac powdery mildew fungus.
Environmental Pollution, (A) 9: 107-114.
Hill Jr, G.R. and M.D. Thomas 1933. Influence of leaf
destruction by sulphur dioxide and by clipping on yield
of alfalfa. Plant Physiology, 8: 223-245.

1 3 B
Hillocks, R.J. 1986. Localised and systemic effects of
root-knot nematode on incidence and severity of Fusarium wilt in cotton. Nematologica, 32: 202-208.
Hirano, K. S. Sugiyama and W. lida 1979, Relationship of the
rhizosphere microflora to the occurrence of Fusarium wilt of tomato under presence of root-knot nematodes
{MeToidogyne incognita (Kofoid & White) Chitwood.
Japanese Journal of Nematology, 9: 60-68.
Hooda, I, and R.K. Grover 1982. Studies on different
isolates of Rhi zoctonia bat^ti col a in relation to
disease development in mungbean, Indian Phytopathology,
35: 619-623.
Ibrahim, I.K.A., M.A. Rezk and H.A.A. Khali1 1982. Effects
of Meloidogyne incognita and Fusarium oxysporum f. sp.
vasinfectum on plant growth and mineral content of
cotton, Gossypium barbadense L. Nematologica, 28: 298-
302.
Irving, P.M. and J.E. Miller 1984. Synergistic effect on
field grown soybean from combination of sulphur dioxide
and nitrogen dioxide. Candaian Journal of Botany, 62;
840-846.
Iyengar, N.K. 1935. Cytological investigation on the genus
Cicer. IRFA 1937. Hlth Bulletin NO.23 pp.22-47. Ann.
Botony, 3:271-305.
Jain, N.K, M.N. Khare and H.C. Sharma 1973. Variation among
the isolates of Rhizoctonica bataticola from urid plant

1 3 6
par ts and s o i l I I . Pa thogen ic i t y , morphology and growth
p a t t e r n (M. phoseolina on PhaseoTus mungo). Mysore
Journal of A g r i c u l t u r a l Science, 1: 411-418.
J a n c a r i k , V. 1 9 6 1 . V y s k y t d r e v o k a z n y c h hub V Kourem
Poskozovant Oblas t ; Krusnych hor. L e s n i c t v i , 7: 677-692.
Jenes, K.F. , L.S. Dochinger, B.R. Roberts and A.M. Townsend
1976. I n : J . P . M iksche E d . , P r o c e e d i n g i n L i f e
Science: Modern Methods in Forest Genet ics, Spr inger -
Verlag B e r l i n , pp.189-216.
Jenkins, W.R. and B.W. Coursen 1957. The e f f e c t of r oo t - kno t
nematodes, Meloidogyne incognita acrita and M. hap la on
Fusarium w i l t o f tomato. P lant Disease Reporter , 4 1 ;
182-186.
Johanson, 0. 1954. Rapport over e t t S tud ium av l u f t och
nederbord omkr ing Svenska K l i f f e r o l j e A t t i e b a l o g e t s
an laggningar v i d Kvarn t rop med s p e c i e l l hansyn t i l l
t i s t r i b u t i o n e n av svavel och dess inverkan pa vax ta rna .
L ie . dvh, v i d K. Lantbrukshogskolan, Uppsala.
Johnson, A.W. and R.H. L i t t r e l l 1969. E f f ec t o f Meloidogyne
incognita, M. javanica and M. hapla on the s e v e r i t y of
Fusarium w i l t o f chrysanthemum. Journal of Nematology,
1: 122-125.
Jones, Eustace W. 1952. Some o b s e r v a t i o n s on t he l i c h e n
f l o r a of t ree boles w i th spec ia l reference to the e f f e c t
of smoke. Rev. B roy l . L ichenology, Pa r i s , 2 1 : 96-115.

1 3 7
Jones, J.P. A.J. Overman and P. Grill 1976. Failure of root-
knot nematode to affect Fusariurn wilt resistant of
tomato. Phytopathology, 66: 1339-1341.
Jones, M.G.K. and D.H. Northcote 1972b. Multinucleate
transfer cells induced in coleus roots by the root-knot
nematodes, Meloidogyne arenaria. Protoplasma, 75: 381-
395.
Jorgenson, E.G. 1970. Antagonistic interaction of Heterodera schachtii Schmidt and Fusarium oxysporum (Woll.) on
sugarbeets. Journal of Hematology, 2 (4): 393-399.
Kanwar, R.S. D.C. Gupta and Kum Kum Walia 1988. Effect of
nematode inoculations at different intervals and soil
type on interaction between Meloidogyne Javanica and
Rhizoctonia on cowpea. Indian Journal of Nematology,
18(1): 112.
Karpen, D.N. 1970. Ozone and sulphur dioxide synergism:
Feliar injury to a panderosa pine geographic race
plantation in the puget sound region. Plant Disease
Reporter, 54: 945-948.
Katz, M. 1952. The effect of sulphur dioxide on conifers.
In: Air Pollution (Ed. L.C. McCabe). McGraw Hill Book
Company, New York, Chapter 8.
Katz, M. and G.A. Ledingham 1939. In: Effect of sulphur
dioxide on vegetation. National Research Council,
Bulletin 815, Ottawa, Canada, Chapter XI, pp.262-287.

1 3 8
Kaur, G. , U.S. Singh and G.A. Garg 1987. Mode of action of
toxins isolated from Fusarium oxysporum f.sp. ciceris.
Indian Phytopathology, 40 (1): 76-84.
Kawamura, T. and K. Hirano 1967. Studies on the complex
disease caused by root-knot nematode and Fusarium wilt
fungus in tomato seedings I. Development of wilt
symptoms and differences in disease incidence in tomato
varieties. Technical Bulletin of Faculty of
Horticulture, Chiba University, 15:7-19.
Kawamura, T. and K. Hirano 1968. Studies on the complex
disease caused by root-knot nematode and Fusarium wilt
fungus in tomato seedlings II. Process of infection
associated with disease incidence. Technical Bulletin
of Faculty of Horticulture, Chiba University, 16: 23-25.
Keller, T. 1976. Auswirkungen niedriger SOp- Konzentrationen
auf junge Fichten Schweiz. Zeitschrift fur Forstwirs
chaft, 127: 237-251.
Khan, M.R. 1989. Studies on root-knot nematodes in relation
to environmental pollution on vegetable crops. Ph.D.
Thesis, Aligarh Muslim University, Aligarh, pp.209.
Khan, M. Wajid 1984. Ecological niche of plant nematodes in
soil biotic community. In: Progress in Microbial
Ecology (Eds. K.G. Mukerji, V.P. Agnihotri and R.P.
Singh) pp.165-177. Print House (India), Lucknow.
Khan, M. Wajid and M. Abdul Salam 1990. Interaction of
Meloidogyne Javanica, Fusarium udum and Rhizobium on

1 3 9
pigeonpea i n the presence o f n i c k e l and c o b a l t as
p o l l u t a n t s . Annals of App l ied B io logy , 116: 549-556.
Khan, M. Wajid and Madhu Kulshrestha 1991. Impact of su lphur
d i o x i d e exposure on c o n i d i a l g e r m i n a t i o n o f powdery
mildew f u n g i . Environmental P o l l u t i o n ; 70: 81-88.
Khan, M. Waj id and J . M u l l e r 1982. I n t e r a c t i o n between
Rhizoctonia so l ani and Meloidogyne hap la on r a d i s h
g n o t o b i o t i c c u l t u r e . Liyban Journal of A g r i c u l t u r e , 11 :
133-139.
K i n l o c h , R.A. 1980 . R e v i e w - t h e c o n t r o l o f nematodes
i n j u r i o u s t o soybean. Nematropica, 10:141-153.
K i n l och , R.A.,C.K. Hiebsch and H.A. Peacock 1987. G a l l i n g and
y i e l d of soybean c u l t i v a r s grown in Meloidogyne arenaria
i n f es ted s o i l s . Journal of Nematology, 19 ( 2 ) : 223-239.
K i s i e l , M.JK. Daubert and D.M. Zuckerman, 1969. The e f f e c t
of Tylenchus agricola and Tylenchohbynchus clay torn' on
r o o t - r o t of corn caused by Fusarium roseum and Pythium
ultimum. Phytopathology, 59: 1387-1390.
K le ineke- Borchers, A. and U. Wyss 1982. I n v e s t i g a t i o n on
changes in s u s c e p t i b i l i t y of tomato p lan t s t o Fusarium
oxysporum f . s p . lycopersici a f t e r i n f e c t i o n s by
Meloidogyne incognita. Z e i t s c h r i f t f u r
P f lazenkrankhe i ten und Pf lanzenschutz , 89 ( 1 . 2 ) : 67-68.
Kock, G. 1935 . E i c h e n m e h l t a u und R a u c h a g a s s - s c h a d e n .
Z e i t s c h r i f t f u r P f lenzenkrankhe i ten , XIV: 44-45.

1 ^ O
Kotasthane, S.R., P.S. Agrwal, L.K. Joshi and L. Singh 1980.
Studies on wilt complex in Bengal gram (Cicer arietinum L.). Review of Plant Pathology, 59: 470.
Kotcon, J.B., D.L. Rouse and J.E. Mitechell 1985.
Interaction of Vertici11ium dahTiae, Colletotrichum
coccodes, Rhi zoctonia sol am' and Pratylenchus penetrans
in the early dying syndrom of Russet Burban K
Polutoes.Phytopathology, 75: 68-74.
Kress, L.W., J.E. Miller, H.J. Smith and J.O. Rawlings 1986.
Impact of O3 and SO2 on soybean yield. Environmental
Pollution; (A) 41: 105-123.
Krishnaprasad, K.S. and G.M. Padaganul 1980. Observations on
the association of Roty1enchulus ren i formi s with
Verticmium wilt on cotton. Indian Journal of
Nematology, 10: 91-92.
Kumar, N. and N.K. Yadav 1988. Response of potato {Solanum tuberosum L.) to sulphur dioxide pollution. Pollution
Abstract, 19: 1095.
Kumar, R.,S. Ahmad and S.K. Saxena 1988. Disease complex in
chickpea involving Meloidogyne incognita and Fusarium
oxysporum. International Nematology Network Newsletter,
5 (3): 12-14.
Kundela, M. and E. Novakova 1962. Lensi skudeia skody zveri V
lesich Paskozovanych Kourem Lesnictvi 6: 493-502.
Laurence, J.A. and A.L. Aluiso 1981. Effect of sulphur
dioxide on expansion of lesions caused by
Corynebacterium nebraskense in mazie and by Xanthomonas

14-1
phaseoli Var. sojensis in soybean. Phytopathology, 71:
445-448.
Laurence, J.A., A.L. Aluiso, L.H. Weinstein and D.C. McCune
1981. Effects of sulphur dioxide on southern bean mosaic
and dwarf mosaic. Environmental Pollution, (A) 24: 185-
191 .
Laurence, J.A., A.J. Weinstein, D.C. McCune and A.L. Aluise
1979. Effects of sulphur dioxide on southern corn leaf
blight of maize and stem rust of wheat. Plant Disease
Reporter, 63: 975-978.
Law, R.M. and T.A. Mansfield 1982. Oxides of nitrogen and
the greenhouse atmosphere. In: Effects of Gaseous Air
Pollution in Agriculture and Horticulture. (Eds. M.H.
Unsworth and D.P. Ormord). Butterworth Scientific,
London, Boston, pp. 93-112.
LeBlanc, F., D.N. Rao and G. Comeau 1972. The epiphytic
vegetation of Populus balsamifera and its significance
as an air pollution indicator in Sudbury, Ontario.
Canadian Journal of Botany, 50(3): 519-528.
Lee, C.C., L.S. Bird, M.P. Thaxton and M.L. Howell 1986. The
association of Macrophomina phaseol ina with cotton. Acta
Phytophylactica Sinica, 13(3): 169-173.
Linzon, D.N. 1958a. The influence of smelter fumes on the
growth of white pine in the Sudbury region. Canadian
Department of Agriculture Publication, Ontario,
Department of Lands and Forests, pp. 45.

1 4 - 2
L i t t r e l l , R .H. and A.W. J o h n s o n 1 9 6 9 . P a t h o g e n i c i t y o f
Pythium aphanidermatum t o c h r y s a n t h e m u m i n c o m b i n e d
i n o c u l a t i o n w i t h Be 1 onoTaimus 1 ongi caudatus o r
MeToidogyne incognita. P h y t o p a t h o l o g y , 59 : 115.
Loos , C.A. 1959, Symptom e x p r e s s i o n o f Fusarium w i l t d i s e a s e
o f t h e Gros M iche l banana i n t h e presence o f Rodopholus
similius (Cobb . 1893) T h o r n e , 1949 and MeToi dogyne
incognita acrita C h i t w o o d , 1 9 4 9 . P r o c e e d i n g s
Helminthological Society, Washington, 26: 103-111.
Lopes, E.B. and L.E.G. Lordello 1979. Meloidogyne incognita and Fusarium sol ani f.sp. piperi associated with wilt
of Piper nigrum in Paraiba. Revista de Agricultura,
Piracicaba, Brazil, 54(4): 263-268.
MacCallan, S.E.A. and F.R. Weedon 1940. Toxicity of ammonia,
chlorine, hydrogen cyanide, hydrogen sulphide, and
sulphur dioxide gases, II. Fungi and bacteria.
Contributions Boyce Thompson Institute, 11: 331-343.
MacCormik, R.A. 1968. Air pollution climatology. In: Air
Pollution (Ed. A.C.Stern). Academic Press, New York,
Vol. I: 275-320.
Mai, W.F. and G.S. Abawi 1987. Interaction among root-knot
nematodes and Fusarium v^^^t fungi on host plants. Annual Review of Phytopathology, 25: 317-338.
Maier, C.R. 1964. The importance of Alternaria spp. in the
cotton seedling disease complex in New Mexico. Plant
Disease Reporter, 49: 904-909.

1 ^ 3
Mandl, R.H., L.H. Weinstein and M. Keveny 1975, Effects of
hydrogen fluoride and sulphur dioxide alone and in
combination on several species of plants. Environmental
Pollution, 9: 133-143.
Mani , A. and C.L. Sethi 1984. Plant growth of chickpea as
influenced by initial inoculum levels of Meloidogyne incognita. Indian Journal of Nematology, 14(1): 41-44.
Mani, A. and C.L. Sethi 1985. Reduced lamina, an typical
symptom induced by Fusarium solani on chickpea. Indian
Phytopathology, 38 (3): 542-543.
Mani, A. and C.L. Sethi 1987. Interaction of root-knot
nematode, Meloidogyne incognita with Fusarium oxysporum
f. sp. ciceris and F. solani on chickpea. Indian Journal
of Nematology, 17(1): 1-6.
Martin, W.J., L.P. Newson and J.E. Jones 1956. Relationship
of nematodes to the development of Fusarium wilt in
cotton. Phytopathology, 46: 285-289.
Masaru, N., F. Syozo and K. Saburo 1976. Effects of exposure
to various injurious gases on germination of lily
pollen. Environmental Pollution, 11: 181-188.
McClure, M.A., K.C. Ellis and E.L. Nigh 1973. Plant infection
development and histopathology of Meloidogyne incognita in resistant cotton. Journal of Nematology, 6: 21-26.
McGuire, J.M., H.J. Walters and D.A. Slack 1958. The
relationship of root-knot nematodes to the development
of Fusarium wilt in alfalfa. Phytopathology, 48: 344
(Abstr.).

1 A^A-
Meiri, A. and Z. Sole! 1963. Transmission of charcoal rot by
sesame seeds. Phytopathologia Mediterrania, 29: 90-92.
Melakebrhan, H., R.C. Brooke, J.M. Webster and J.M. D'Auria
1985. The influence of MeToidogyne incognita on the
growth, physiology and nutritent content of Phaseolus vulgaris. Physiology of Plant Pathology, 26: 259-268.
Melendez, P.L. and N.T. Powell 1967. Histological aspects
of the fusarium wilt-root-knot complex in flue-cured
tobacco. Phytopathology, 57: 286-292.
Menser, H.A. and H.E. Heggestad 1966. Ozone and sulphur
dioxide synergism: Injury to tobacco plants. Science,
153: 424-425.
Miller, V.L., R.K. Howell and B.E. Cadwell 1974. Relative
sensitivity of soybean genotypes to Oo and SOg. Journal
of Environmental Quality, 3: 35-37.
Minton, N.A., D.K. Bell and B. Doupink Jr. 1969. Peanut pod
invasion by Aspergillus flavus in the presence of
Meloidogyne hapla. Journal of Nematology, 1: 318-320.
Minton, N.A. and E.B. Minton 1966. Effect of root-knot and
sting nematodes on expression of Fusarium wilt of
cotton in three soils. Phytopathology, 56: 319-322.
Minton, N.A., M.A. Parker and R.N. Flower 1975. Response of
soybean cultivars to Meloidogyne incognita and
Sclerotium rolfsii. Plant Disease Reporter 59(11): 920-
923.

1 4.S
Mishra, C., C. Som and B. Singh 1988. Interaction of
Meloidogyne incognita and Rhizoctonia batati col a in
white jute (Corchorus capsularis). Indian Journal of
Agricultural Science, 58(3): 234-235.
Mishra, S.D. and H.S. Gaur 1981a. Pathogenicity of
Meloidogyne incognita and Rotylenchulus reniformis to
mothbean, Vigna aconitifolius Jacoq. Indian Journal of
Nematology, 11 (1): 85-89.
Mishra, S.D. and H.S. Gaur 1981b. Effect of individual and
concomitant inoculation with Meloidogyne incognita and Rotylenchulus reniformis on the growth of black gram
{Vigna mungo). Indian Journal of Nematology, 11(1): 25-
28.
Montalro, A.E. and P.L. Melendez 1986. Histopathology of
interaction between Fusrium oxysporum f. sp. dioscoreae
and two nematodes species on yam. Journal of Agriculture
of the University of Puerto Rico, 70(4): 245-254.
Morrell, J.J. and J.R. Bloom 1981. Influence of Meloidogyne incognita on Fusarium wilt of tomato at or below the
minimum temperature for wilt development. Journal of
Nematology, 13(1): 57-60.
Mors ink, F. and A.E. Rich 1968. Interaction between
Vertici 11ium albo-atram and Pratylenchus penetrans in
Vertici11 ium wilt of potato. Phytopathology, 58: 401 (Abstr.).
Mountain, W.B. and CD. McKeen 1962. Effect of Vertici11ium dahliae on the population of Pratylenchus penetrans.
Nematologica, 7: 261-266.

1 4-e
Moura, de R.M., E. Echandi and N.T. Powell 1975. I n t e r a c t i o n
o f Corynebacter i urn michiganense and Me 1oi dogyne
incognita on tomato. Phytopathology, 65: 1332-1335.
Mukhtar, J . and A.A. Khan 1989. Disease complex i n chickpea
i n v o l v i n g Meloidogyne Javanica and Sclerotium rolfsii.
I n t e r n a t i o n a l nematology. Network News le t te r , 6 ( 4 ) : 3 1 -
32.
M u j i b , I . Azmi and B.D. P a t i l 1988. E f f e c t o f r o o t - k n o t
nematode on carpet legume, Dolichos lablab v i e lignosus
w i t h a note on sc reen ing o f r e s i s t a n t l i n e s . I n d i a n
Journal of Nematology, 18(1 ) : 104.
M u r t h y , G.S. and D . J , B a g o r z y a r a j a 1980 . F l a v a n o l and
a l k a l o i d content of pigeonpea c u l t i v a r s r e s i s t a n t and
suscep t i b le t o Fusarium udum. I nd ian Phytopathology, 33
( 4 ) : 633-635.
Murumkar, C.V. and P.D. Chavan 1985. Phys io log i ca l changes i n
chickpea leaves w i t h Fusarium w i l t . Biovigyanafn, 11(1) :
118-120.
N a i l i n i , C. G . , P. S ivapa lan and A.M. Dharmasena 1985.
S c r e e n i n g o f d i f f e r e n t c u l t i v a r s o f b l a c k gram
{Phoseolus mungo L . ) , green gram {PhaseoTus aureus
R o b x . ) and cowpea (Vigna catjang W a l p . ) f o r
r e s i s t a n t / s u s c e p t i b i l i t y t o Meloi dogyne s p p .
I n t e r n a t i o n a l Nematology Network News le t te r , 2 ( 4 ) : 22-
25.
Nath, P.P., A.K, Banerjee, M.G. Haider and B.K, Sinha 1979.
Study of the nematodes o f p u l s e c rop i n I n d i a . I .

Pathogen ic i t y of MeToidogyne incognita on gram. Ind ian
Phytopathology, 32 (1 ) : 28-32.
Nayak , D.K. B .N . R o u t a r a y S. Ray and S . N . Das 1987 . Popu la t ion e f f e c t s of Meloidogyne incognita i n winged bean. I n t e r n a t i o n a l Nematology Network News le t te r , 4 ( 1 ) : 22-24.
Newha l l , A.G. 1958. The i n c i d e n c e o f Panama d i s e a s e o f
banana i n t h e p r e s e n c e o f t h e r o o t - k n o t and t h e
b u r r o w i n g nematodes {MeToidogyne incognita and
Radopholus simi1 is). P lan t Disease Repor ter , 42: 853-
856.
Ngauma, B.W. and V.W. Saka 1986. React ions of some loca l
p i g e o n p e a l a n d r a c e s t o r o o t - k n o t n e m a t o d e s ,
Meloidogyne incognita. I n t e r n a t i o n a l Nematology Network
News le t te r , 3 ( 3 ) : 29 -31 .
Nouguera, G.R. 1980. Growth and g e r m i n a t i o n o f spo res o f
Fusarium oxysporum f . s p . lycopersici i n t h e r o o t
e x t r a c t s o f tomato p l a n t s i n f e c t e d w i t h Meloidogyne
incognita. Agronomia T r o p i c a l , 3 0 ( 1 - 6 ) : 305-313.
Noguera, G.R. 1982. A l t e r a t i o n in the p roduc t ion of r i s h i t i n
in the roo ts and t y l oses in stems and the i n t e r a c t i o n of
Meloidogyne - Fusari urn i n tomato p l a n t s . Agronomia
T r o p i c a l , 32 (1 -6 ) : 303-308.
Noguera, G.R. 1983. I n f l uence of Meloidogyne incognita on the
l o c a l i z a t i o n of Fusarium oxysporum f . s p . lycopersici i n
tomatoes. Agronomia T r o p i c a l , 3 3 ( 1 - 6 ) : 103-109.

1 ^ 8
N o g u e r a , G.R. and B.G. S m i t s 1982 . V a r i a t i o n i n t h e r h i z o s p h e r e m i c r o f l o r a o f t o m a t o i n f e c t e d w i t h M. incognita. Agronomia T r o p i c a l , 3 2 ( 1 - 6 ) : 147-154.
N o r d m e y e r , D. and R.A. S i k o r a 1983 . S t u d i e s on t h e
i n t e r a c t i o n be tween Heterodera daverti, Fusar i urn
avenaceum and Fusariurn oxysporum on Trifolium
subterraneum. Rev. Nematologica, 6: 193-198.
Nor ton, D.C. 1960. Meloidogyne hapla as a f a c t o r in a l f a l f a
dec l i ne in Iowa. Phytopathology, 59: 1824-1828,
Oden, S. 1968. The a c i d i f i c a t i o n of a i r and p r e c i p i t a t i o n and
i t s consequences in the na tu ra l environment. B u l l e t i n
No.1 Ecology Committee Swedish Nat iona l Science Research
Counci1, Stokholm.
O 'Ereo le , N. , R. Taeconi and M. S p o r t e l l i 1982. P re l im ina ry
r e s u l t s on the p o s s i b l e a s s o c i a t i o n between v a s c u l a r
f u n g i and Meloidogyne i n some c o m m e r c i a l t o m a t o
v a r i e t i e s . In fo rmator Ag ra r io Verona, 32(30) : 21054.
Oostenbr ink, M. 1966. Major c h a r a c t e r i s t i c s of the r e l a t i o n
be tween nematode and p l a n t s , M e d e d e l i n g e n
Landbouwhogeschool, Wageningen, No. 66-4, 46pp,
O r i o n , D. and H. Hoest ra 1974. The e f f e c t o f r o o t - k n o t
nematodes and E t h r e l l on Fusariurn w i l t o f t o m a t o e s .
Nether land Journal of P lan t Pathology, 80: 28-36.
Or i on , Q and Q. Netzer 1981. Suppressive e f f e c t of the r o o t -
knot nematode on Fusarium w i l t of muskmelon. Revue de
Nematologie, 4 ( 1 ) : 65-70.

1 4 9
Padila, C , R. Lopes and E. Verga 1980. Interaction between
Meloidogyne spp. {M. incognita and M. hapla) and
Fusarium oxysporum f.sp. pi si on pea. Agronomia
Constarriceuse, 4(1): 55-60.
Padwick, G.W. 1941. Report of the Imperial Mycologist, Sci .
Rep. Imperial Agriculture Institute, New Delhi, pp. 94-
101 .
Palmer, L.T. and MacDonald 1974. Interaction of Fusarium spp.
and certain plant parasitic nematodes on maize.
Phytopathology, 64: 14-17.
Patel, B.A., J.C. Chavda; H.V. Patel and D.J. Patel, 1988.
Screening of certain cowpea lines for resistant root-
knot nematodes. Indian Journal of Nematology, 18(1):
128.
Patel, H.R. N.A. Thakur, B.K. Patel and C.C. Patel 1987.
Interaction between Meloidogyne incognita and Fusarium
oxysporum f. sp. ciceris on chickpea variety 'Chaffa'.
Indian Journal of Nematology, 17: 124.
Patel, H.R., M.U. Vaishmav and I.U. Dhruj 1985. Interaction
of Meloidogyne arenar i a and Fusarium solani on
groundnut. Indian Journal of Nematology, 15(1): 98-100.
Pelez, J., K. Skadow and R. Fritzsehe 1983. Influence of
Meloidogyne incognita on the host susceptibility of
cucumber to Fusarium oxysporum f. sp. lycopersici and
tomato to F. oxysporum f.sp. cucumerium. Nemamtologica,
29(4): 443-453.

1 5 0
Perera, E. 1985. Soybean diseases in winter sowing in
Nicaragua. Ciencias de la Agricultura, 25: 14-22.
Piper, C.V. and W.J. and Morse 1910. Soybean. Illus. New
York.
Pitcher, J.R. 1974. Reduction of resistance of tomato to
Fusarium oxysporum f. sp. lycopersici by Meloidogyne
Javam'ca. Journal of Nematology, 5: 148.
Pitcher, R.S., 1963. The role of plant parasitic nematodes in
bacterial diseases. Phytopathology, 53: 35-39.
Pitcher, R.S., 1965. Interrelationship of nematodes and other
pathogens of plants. Helminthological Abstract, 34: 1-
17.
Pitcher, R.S, 1978, Interactions of nematodes with other
pathogens. In: Plant Nematology, (Eds. J.F. Southey) pp.
63-77. Her Majesty Stationery Office, London.
Porter, D.M. and N.T. Powell 1967. Influence of certain
Meloidogyne spp. on Fusarium wilt development in flue-
cured tobacco. Phytopathology, 57: 282-285.
Powell, N.T. 1963. The role of plant parasitic nematodes in
fungal diseases. Phytopathology, 53: 28-35.
Powell, N.T. 1971a. Interaction of plant parasitic
nematodes with other disease causing agents. In: Plant
Parasitic Nematodes (Eds. B.M. Zuckerman; W.F . Mai and
R. A. Rohde). Academic Press, New York and London, Vol.
II, pp. 119-136.

1 5 1
Powell, N.T. 1971b. Interaction between nematodes and fungi
in diseases complexes. Annnual Review of phytopathology,
9: 253-274.
Powell, N.T. 1979. Internal synergisms among organisms
inducing disease. In: Plant Disease. An Advance Treatise
(Eds. J.G. Horsfal1 and E.B. Cowling) Vol.4, pp.113-133,
Academic Press New York.
Powell, N.T., P.L. Melendez and C.K. Batten 1971. Disease
complex in tobacco involving Meloidogyne incognita and
certain soil-borne fungi. Phytopathology, 61: 1332-1337,
Prasad, K.S.K. A.L. Sitarmaiah and R.K. Hedge 1980. Note on
the association of Corticium rolfsii and Meloidogyne
incognita in wilt complex of Solanum khasianum. Indian
Journal of Agriculture Science, 50(5): 545.
Prasad, Y. 1986. Effect of different doses of potash on the
incidence of root and stem rot of jute. Journal of
Research, Assam Agriculture University, 4: 163-164.
Pratt, G.C. K.W. Kromroy and S.V. Krupa 1983. Effects of
ozone and sulphur dioxide on injury and folior
concentrations of SOg and chlorophyll in soybean
{Glycine max). Environmental Pollution (A) 32: 91-99.
Prinsloo, G.C. 1986. Black root-rot of chicory in South
Africa. Phytopathologica, )8(4): 225-226.
Pryzbylski, Z. 1967. Results of observations of the effect
of SO2, SO2 and H2SO4 on fruit trees, and some harmful
insects near the sulphur mine and sulphur processing

1 5 2
p lan t a t Machow near Tarnobrzeg. Postepy Nauk R o l n . , 2 1 :
111-118.
Raut, J.G. and R.W. I ng le 1989. V a r i a t i o n i n i s o l a t e s of
Rhizoctonia bataticola. Ind ian Phytopathology, 42; 506-
508.
Raut, S.P. 1980. E f f e c t o f i n i t i a l i n o c u l u m l e v e l s o f
Meloidogyne. incognita on p l a n t g r o w t h and r h i z o b i a l
nodu la t ion of mungbean. Ind ian Phytopathology, 33(2 ) :
351-353,
Reddy, P.; D.B. Singh and S.R. Shrma 1979. I n t e r a c t i o n of
Meloidogyne. incognita and Rhizoctonia sol ani i n r o o t -
r o t d i s e a s e comp lex o f f r e c h b e a n . I n d i a n
Phytopathology, 32: 651-652.
Re ine r t , R.A. and J .S . Sanders 1982. Growth of rad ish and
m a r i g o l d f o l l o w i n g r e p e a t e d e x p o s u r e t o n i t r o g e n
d i o x i d e , su lphur d iox ide and ozone. P lan t Disease 66:
122-124.
R e i n e r t , R.A. and D.E. Weber 1980. O3 and SO3- i nduced
changes in soybean growth. Phytopathology, 70: 914-916.
Reynolds, H.W. and R.G. Hanson 1957. Rhizoctonia disease of
c o t t o n i n p r e s e n c e o r absence o f c o t t o n r o o t - k n o t
nematode in Ar izona. Phytopathology, 47: 256-261.
R i b e i r o , C.A, and S. Ferraz 1984. The i n t e r a c t i o n between
Meloidogyne javanica and Fusar iurn oxysporum f . s p .
phased i in Phaseolus vulgaris. Fi t o p a t o l o g i a B r a s i l i a
8: 4 3 9 - 4 4 6 .

1 5 3
Richards, B.N. 1976. Introduction of Soil Ecosystem.
Longman, London pp.266.
Riedel, P.M. R.C. Rowe and M.J. Martin 1985. Differential
interactions of Pratylenchus crenatus, P. penetrans and
P. scribneri with Vertici 11ium on potato early dying
disease. Phytopathology, 75: 414-422.
Roberts, B.R. 1976. The response of field grown white pine
seedlings to different sulphur dioxide environments.
Environmental Pollution, 11: 178-180.
Rodriguez-Kabana, R. and S.C. Willians 1981. Assessment of
soybean yield losses caused by Meloidogyne. arenaria. Nematropica, 11: 105-113.
Routaray, B.N.jH. Sahoo and S. N. Das 1986. Evalution of
greengram and balckgram varieties against reniforms
nematode, RotylcnchuTus reniformis. Indian Journal of
Nematology, 16: 27-29.
Ruston, A.G. 1914. The plant as an index of smoke pollution.
Annals of Applied Biology, 7: 392-402.
Saeed, M., M. Ahmad and H.A. Khan 1972. A complex disease of
tomato and papaya caused by nematode-fungi association
in Pakistan. Pakistan Journal of Scientific and
Industrial Research, 15: 312-313.
Sakhuja, P.K. and C.L. Sethi 1986. Multiplication of
Meloidogyne Javanica as affected by Fusarium soTani and
Rhizoctonia bataticola on groundnut. Indian Journal of
Nematology, 16; 1-3.

1 54-
Sasser, J.N. C.C. Carter and K.N. Hartman 1984.
Standardization of host suitability studies and
reporting of resistance to root-knot nematodes. North
Carolina State University, Graphics, Raleigh.
Saunders, P.J.W. 1966. The texicity of sulphur dioxide on
Diplocarpon rosae Wolf. Causing black spot of roses.
Annals of Applied Biology, 58: 103-114.
Scheffer, T.C. and G.G. Hedgcock 1955. Injury to north
western forest trees by sulphur dioxide from smelters.
United States Department of Agriculture Technical
Bulletin, 1117: 49.
Schindler, A.F.; R.N. Stewart and P. Semeniuk, 1961. A
synergistic Fusar7uw-nematode interaction in carnation.
Phytopathology, 51: 143-146.
Seinhorst, J.W. and K. Kuniyasu 1971. Interaction between
Pratylenchus penetrans and Fusarium oxysporum f.sp. pi si
Race-2 and Rotylenchulus reniformis and F. oxysoorum
f.sp. pisi Race-1 on peas. Nematologica, 17: 444-452.
Sharma, R.C. and T.P. Bhowmik. 1987. Effect of Macrophomina
phaseoT ina infection on the physico-chemical component
of groundnut seed. Phytopathology, 118 (2): 181-186.
Sharma, Y. and K.C.Basu Chaudhary 1985. Effect of inorganic
fertitizers on the incidence of wilt and root-rot of
cauliflower. Indian Journal of Plant Pathology, 3 (2):
259-262.

1 5 5
Sharma, N.K. and J.S. Gill 1979. Interaction between
Meloidogyne incognita and Rh izoctonia. solani on potato.
Indian Pathopathology, 32: 277-279.
Sharma, B.L. and R.N. Gupta 1986. Survival of Fusarium oxysporum f.sp, ciceris causing wilt of Bengal gram
during summer months. Indian Phytopathology, 36: 561-
562.
Sharma, J.K., I. Singh and H.K. Chhabra 1980. Observations
on the influence of Meloidogyne incognita and
Rhizoctonia bataticola on okra. Indian Journal of
Nematology, 10: 148-151.
Sharp, E.L. 1967. Atmospheric ions and germinations of the
uredosphores of Pucci ni a str i i formi s. Science, 156:
1359-1360.
Sheng, W.S. and C. Boris 1988. Gas exchange responses of
soybean cultivars to short therm expesure of SOp and SO3
Phytopathology, 78: 1513.
Shepherd, R.L. 1970. Breeding for resistant to root-knot
Fusarium - wilt complex in cotton. Proceeding of the
Beltwide Cotton Production Research Conference, Houston,
TTexas, pp.68.
Shew, B.B., R.A, Reinert and K.R. Baker 1982. Response of
tomatoes to ozone, sulphur dioxide, and infection by
Pratylenchus penetrans. Phytopathology, 72: 822-26.
Shoemaker, P.B. and K.R. Barker 1979. Influence of
Meloidogyne. incognita and Vertici 11 iurn dahliae on

1 5 e
tomato a t d i f f e r e n t inoculum d e n s i t i e s . Phytopathology,
59; 1-9.
Sh r ine r , D.S. 1974. E f f e c t s of s imula ted r a i n a c i d i f i e d w i t h
s u l p h u r i c a c i d on h o s t - p a r a s i t e i n t e r a c t i o n s . Ph .D.
Thes is , North Caro l ina State U n i v e r s i t y , Ra le igh , NC.
Sh r i ne r , D.S, 1978. E f f ec t s of s imula ted a c i d i c r a i n on host
p a r a s i t e i n t e r a c t i o n s in p l a n t d isease. Phytopathology,
63: 213-218.
Sidhu, G.S. and J.M. Webster 1974. Genetics of res is tance in
the tomato to r oo t - kno t nematode-wi I t - fungus complex.
Journal of He red i t y , 65: 153-156.
Sidhu, G.S. and J.M. Webster 1977a. P r e d i s p o s i t i o n o f tomato
t o the w i l t fungus {Fusarium oxysporum lycopersici) by
t h e r o o t - k n o t nematode (Meloidogyne incognita).
Nematologica, 23: 433-442.
S idhu, G.S. and J.M. Webster 1977b. The use of amino ac id
fungus auxotrophs to study the p r e d i s p o s i t i o n phenomena
in the r o o t - k n o t - w i l t fungus disease complex of tomato.
Physiology of P lan t Pathology, 11 : 117-127.
S i d h u , G.S. and J . M . Webs te r 1979 . G e n e t i c s o f t o m a t o
r e s i s t a n t t o Fusarium-VerticiTlium c o m p l e x .
Phys io log i ca l P lant Pathology, 15: 93-98.
Sidhu, G.S. and J.M. Webster 1983. Hor i zen ta l res is tance in
tomato aga ins t the Meloidogyne - Fusarium complex: an
a r t i f a c t of p a r a s i t i c e p i s t a s i s . Crop P r o t e c t i o n , 2 :
205-210.

1 5 T
Simay, E.I. 1989. Fungi observed on chickpea {Ci cer arietinum) in 1985-1988. Novenytermeles, 3(5): 435-442.
Singh, S.K, 1989. Studies on interaction of air pollutants
and root-knot nematodes on some pulse crops. Ph. D.
Thesis, Aligarh Muslim University, Aligarh India, pp.
382.
Singh, K.B. and B.S. Dahiya 1973. Breeding for wilt in
chickpea. Symposium on wilt probelm and breeding for
wilt resistance in Bengal gram. Sept. 1973 at tndian
Agricultural Research Institute, New Delhi, India, pp.
13-14.
Singh, R.V. and J.S, Gill 1990. Nematodes associated with
pulse crops and their control. In: Progress in Plant
Nematology (Eds. S.K. Saxena M.W. Khan, Abdul Rashid
and Rashid M. Khan), pp. 503-514, CBS Publishers and
Distributers, Delhi.
Singh, D.B., P. Parvatha Reddy, V.R. Rao and R. Rajendran
1981. Reaction of some varieties and selection of french
bean to Meloidogyne incognita. Indian Journal of
Nematology, 11: 81-83.
Skye, E. 1964. In: Svensk Naturvetenskap. Statens
Naturvetenskapliga Forskningsrad, Stockholm, pp. 377-
332.
Skye, E, 1968. Lichens and air pollution. Acta Phytogeogr.
Suec., 52: 1-123.

1 5 8
Smith, A.L, 1948. Fusarium and nematodes on cotton. United
States Department of Agriculture year book, Plant
Diseases, 1953, pp. 292-298.
Smith, A.L. 1954. Problems on breeding cotton for resistance
to nematodes. Plant Disease Reporter, Supp., 227: 90-
91 .
Smits, B.G. and R. Noguera 1982. Effect of Meloidogyne incognita on the pathogenicity of different isolates of
Fusarium oxysporum on bringal {SoTamum melongena L,).
Agronomia Tropical, 32(1-6): 285-290.
Sobun, N., K.G. Nema and G.S. Dave 1979. The positive
interrleationship between plant parasitic nematode,
Tylenchorhynchus and a root-rot fungus in gram (Cicer arietinum). Physiology of Parasitism, pp. 451-456.
Sprugel, D.G., J.E. Miller, R.N, Muller, H.J. Smith and P.B.
Xerikos 1980. SO2 effects on yield and seed quality of
field-grown soybean. Phytopathology, 70: 1129-1133.
Srivastava, A.S,, K.D. Upadhyay and G. Singh 1974. Effect of
root-knot nematode, Meloidogyne Javanica on gram crop.
Indian Journal of Hematology, 4: 248-251,
Staklasa, J,, 1923. Die Beschandigung der Vegetation durch
Rauchgase und Fabrisexhalation. Urban und Schwarzenberg.
Springei—Verlag, Berlin, pp. 487.
Steve Van Marte 1984. Need for ecological education. In: The
Times of India 13th July, 1984.

1 5 9
Stevens, C , V. Kahn; J. Lu and A.Y. Tang 1987. The effect of
ultraviolet radiation on the control of storage rots of
sweet potato. Phytopathology, 787 (12): 1698.
Tanaka, K. T. Olsuko and N. Kondo, 1982. Participition of
hydrogen peroxide in the inactivation of Calvin cycle SH
enzyme in SOj fumigated spinach leaves. Plant Cell
Physiology, 23: 1009-1018.
Tarr, S.A.J. 1954. Macrophomina phaseol i na recorded on
Phaseolus mungo var. radiatus. Plant Pathology Report.
Research Division Ministry Sudan, pp. 71-80.
Taylor, A.L, and J.N. Sasser 1978. Biology, identification
and control of root-knot nematodes, Meloidogyne spp.
North Carolina State University Graphics, Raleigh, North
Carolina, pp. 111.
Taylor, C.E. 1979. Meloidogyne interrelationship with
microorganisms. In: Root-knot Nematodes {Meloi dogyne species) systemics. Biology and Control, (Eds. F.
Lamberti and C.E. Taylor), pp.375-398. Academic Press,
London. New York, San Francisco.
Taylor, C.E. 1991, Nematode interaction with other pathogens.
Annals of Applied Biology, 116: 405-416.
Temmerman, L.D. 1982. Acute and subacute damage to vegetation
by ammonia following accidental pollution. In: Effects
of Gaseous Air Pollution in Agriculture and
Horticulture (Eds. M.S. Unsworth and D.P. Ormord), pp.
468-469. Butterworth Scientific, London.

1 eo
Thomas, M.D. 1961 . E f f e c t s o f a i r p o l l u t i o n on p l a n t s . I n :
A i r P o l l u t i o n . W o r l d H e a l t h O r g a n i s a t i o n M o n o g r a p a h
S e r i e s 4 6 . p p . 2 3 3 - 2 7 8 . W o r l d H e a l t h O r g a n i s a t i o n ,
Geneva,
Thomas, M . D . , R.H. H e n d r i c k s , T . R . C o l l i e r and G.R. H i l l
1943. The u t l i z a t i o n o f s u l p h a t e and s u l p h u r d i o x i d e f o r
t h e n u t r i t i o n o f a l f a l f a . P l a n t P h y s i o l o g y , 18: 3 4 5 - 3 7 1 .
T h o m a s , M . D , R . H . H e n d r i c k s a n d G . R . H i l l 1 9 5 2 . Some
i m p u r i t i e s i n t h e a i r and t h e i r e f f e c t s on p l a n t s . I n :
A i r P o l l u t i o n (Ed . McCabe, L . ) , pp . 4 1 - 4 7 . McGraw-H i l l
Book Company, New Y o r k ,
Thomason, I . J . 1958 . The e f f e c t o f r o o t - k n o t n e m a t o d e ,
Meloidogyne Javanica on b l a c k e y e b e a n w i l t .
P h y t o p a t h o l o g y , 48 : 398.
T h o m a s o n , I . J . , D . C . E r w i n a n d M . J . G a r b a r 1 9 5 9 . The
r e l a t i o n s h i p o f t h e r o o t - k n o t n e m a t o d e , Meloidogyne
javanica t o Fusarium w i l t o f cowpea. P h y t o p a t h o l o g y , 49 :
602 -606 .
T i n g e y , D . T . , R.A. R e i n e r t , J . A . Dunning and W.W. Heck 1971 .
V e g e t a t i o n i n j u r y f r o m t h e i n t e r a c t i o n o f n i t r o g e n
d i o x i d e and s u l p h u r d i o x i d e . P h y t o p a t h o l o g y , 6 1 : 1506-
1511 .
T i n g e y , D.T. R.A. R e i n e r t , J . A . Dunning and W.W. Heck 1973b.
F o l i o r i n j u r y r e s p o n s e s o f e l e v e n p l a n t s p e c i e s t o
o z o n e / s u l p h u r d i o x i d e m i x t u r e . A tmosphe r i c E n v i r o n e m n t ,
7: 2 0 1 - 2 0 8 .

1 e 1
Trapero-Casas, A. and R.M. Jimenez-Diaz 1985. Fungal wilt and
root-rot disease of chickpea in southern Spain.
Phytopathology, 75: 1146-1151.
Treshow, M. 1965. Responses of some pathogenic fungi to
sodium fluoride. Mycologia, 57: 216-222.
Treshow, M. 1970. Environmment and plant response. MacGraw-
Hill, New York.
Treshow, M, and M.R. Pack 1970. Fluoride. In: Recognition of
Air Pollution Injury to Vegetation: A Pictorial Atlas
(Eds. J.S. Jacobson and A.C. Hill), Air Pollution
Control Association, Pittsburgh, Pennsylvania.
Tu, C.C. and Y.H. Cheng 1971. Interaction of Meloidogyne Javanica and Macrophomina phaseolina in Kenaf root-rot.
Journal of Nematology, 3: 39-42.
Upadhyay, K.D. and Kusum Dwivedi 1987. Analysis of crop
losses in pea and gram due to Meloi dogyne i ncogni ta. International Nematology Network Newsletter, 4(4): 6-7.
Upadhyay, K.D. and Kusuam Dwiedi 1987. Effect of interaction
between Meloidogyne Javanica and Fusariurn oxysporum
f.sp. ciceris on chickpea. Indian Journal of Nematology,
17(1): 145-146.
Upadhyay, K.D. and G. Swarup 1981. Growth of maize plant in
the presence of Tylenchorhynchus vulgaris Upadhyay et
al. 1973, singly and in combination with Pratylenchus zea Graham 1951, and Fusarium moniliforme Shield. Indian
Journal of Nematology, 11: 29-34.

1 6 2
U.S. Environmental Protection Agency 1973. Revised chapter
5, Air Quality Criteria for Sulphur oxides. Document
EPA-R3-73-030, Research Triangle Park, North Carolina,
PP. 43.
Van Gundy, S.D., J.D.Kirkpatrick and J. Golden 1977. The
nature and role of metabolic leakage from root-knot
nematode gall and infection by Rhi zoctoni a solani. Journal of Nematology, 9: 113-121.
Varda Rajan, B.S. and J.S. Patel 1946. Seed transmission of
stem root of jute and its control. Indian Journal of
Agricultural Science, 16: 193-206,
Varshney, C.K. and J.K. Garg 1979. Plant responses to
sulphur dioxide pollution: CRC Critical Reviews in
Environmental Control, pp. 27-49.
Varshney, V.P., A.M. Khan and S.K. Saxena 1981. Interaction
between Meloidogyne incognita, Rhizoctonia sol ani and
Rhizobium sp. on cowpea. International Nematology
Network Newsletter, 4(3): 11-14.
Vavilov, N.I. 1951. The origin, variation, immunity and
breeding of cultivated plants. (Translated from Russian
by Chester, K.S.). Chronica Botanica, 13: 366.
Venkato Rao, A.V. Narsimhan, P. Rajagopalan, P. Vidyasekharam
and T.E. Muthukrishnan 1973. Efficacy of nematicides for
the control of betel-vine wilt. Indian Journal of
Agriculture Science, 43: 1072-1075.

1 6 3
Verma, A.C., A.K. Pathak and B.S. Yadav 1981. Susceptibility
of certain varieties of cluster bean to root-knot
nematode, Meloidogyne incogni ta. Indian Journal of
Nematology, 11: 120 (Abstr.).
Wallace, H.R. 1978. Diagnosis of plant diseases of complex
etiology. Annual Review of Phytopatholgy, 16: 379-402.
Weber,. E. 1982. Air Pollution: Assessment methology and
modeling. Vol.2, Plenum Press, New York.
Weber, D.E., R.A. Reinert and K.R. Barker 1979. Ozone and
sulphur dioxide effects of selected plant parasitic
nematode. Phytopathology, 69: 624-628.
Webster, J.M. 1985. Interactions of Meloidogyne with fungi on
crop plants. In: Advanced Treatise on Meloidogyne (Eds.
Sasser, J.N. and Carter, C.C.) Vol. I, pp. 183-192.
North Corolina State University Graphics, Raleigh.
Weidensaul, T.C. and S.L. Darling 1979. Effects of ozone and
sulphur dioxide on the host pathogen relationship of
Scots pine and Scirrhia acicola. Phytopathology, 69:
939-941 .
Weigle, J. and Ziegler 1962. Die raumliche verteilung Von S
und die art der markierten verbindungen in
spinatblattern nach begasung mit ^^S02. Planta, 58: 435-
447.
Weinstein, L.H., D.C. McCune, A.L. Aluisio and P. Van Leuken
1975. The effect of sulphur dioxide on the incidence and

1 6 4
severity of bean rust and early blight to tomato.
Environmental Pollution, (A)9: 145-155.
Welty, R.E., K.R. Barker and D.L. Lindsey 1980. Effect of
Meloidogyne hapla and M. incognita on Phytophthora root-
rot of alfalfa. Plant Disease, 64(12): 1097-1099.
Westerlund, F.A., Jr. R.N. Campbell and K.A. Kimble 1974.
Fungal root-rot and wilt of chickpea in California.
Phytopathology, 64(4): 432-436.
White, K.L., A.C. Hill and J.H. Bennett 1974. Synergistic
inhibition of apparent photosynthesis rate of alfalfa by
combinations of sulphur dioxide and nitrogen dioxide.
Environmental Science and Technology, 8: 574-576.
White, L.V. 1962. Root-knot and the seedling disease complex
of cotton. Plant Disease Reporter, 46: 501-504.
Wood, F.A. 1968. Sources of plant pathogenic air pollutants.
Phytopathology, 58: 1075-1084.
Wood, T. 1976. Short-term Effects of simulated Acid Rain
Upon the Growth and Nutrient Relation of Pinus strobus L. In: proceeding 1st International Symposium. Acid
Precipitation and the Forest Ecosystem. (Eds. Dochinger,
L.S. and T.A. Seliga) United States Department of
Agriculture, Northeastern Forest Experimental Station;
Upper Darby, pp. 815.
Wood, T., and F.H. Bormann 1974. The effects of an artificial
acid mist upon the growth of Betula al 1 eghaniensis Britt. Environmental Pollution; 7: 259-268.

1 6 5
Wood, T. and F.H. Bormann 1975. Increase in foliar leaching
caused by acidification of an aritifical mist. Ambio, 4:
169-171 .
Wrather, J.A. and S.C. Anand, 1988. Relationship between time
of infection with Heterodera glycines and soybean yield.
Journal of Nematology, 20: 439-442.
Young, P.A., 1939. Tomato wilt resistance and its decrease by
Heterodera marioni. Pfiytopathology, 29: 871-879.
Young, P.A., 1940. Soil fumigation with chloropicrin and
carbon bisulphide to control tomato root-knot and wilt.
Phytopathology, 30: 860-865.
Zelitich, I., 1975. In: Environmental and Biological Control
of Photosynthesis (Eds. Junke, W. and C. Hague), pp.
251-262.
Zimmerman, P.W. and W. Crocker 1934. Toxicity of air
containing sulphur dioxide. Contribution Boyce Thompson
Institute , 6: 455-470.







