
1
PROYECTO DE MAESTRÍA
TÍTULO: “EL ROL DE LAS HORMIGAS TEJEDORAS (OECOPHYLLA SMARAGDINA)
Y LA ESTRUCTURA DE SUS NIDOS EN SU REGULACIÓN MICROCLIMÁTICA”
GIOVANNI M. RAMÓN C.
UNIVERSIDAD JAMES COOK – ESCUELA DE BIOLOGÍA MARINA Y TROPICAL
TOWNSVILLE, QUEENSLAND, AUSTRALIA
2013

2
Giovanni Ramón
Student No. 12632085
Minor project report
Word count: 6101
The role of green tree ants (Oecophylla smaragdina) and the structure of
their nests in the microclimatic regulation
Abstract
The climate of a region is fundamental in determining the local species
composition and distribution. On smaller scales, organisms experiment
microclimates different from the bigger scale macroclimate. The environmental
temperature and humidity are important in determining the life history,
distribution and abundance of organisms, including insects. The particularities of
this group make them more prone to respond to changes in these abiotic factors.
Ants, an ecologically important, diverse and widely distributed group of social
insects, are also influenced by temperature and humidity. The capacity of ants to
build nests, however, enables them to be active even when external conditions are
not favorable and it has allowed them to colonize many environments. There has
been a lack of research in tropical canopy-nesting ant species and how they
regulate the temperature and humidity inside their nests. In this study, the
microclimate inside the nests of Oecophylla smaragdina, a conspicuous arboreal
species, and the factors determining it are investigated. We installed data loggers
inside nests of O. smaragdina to record their temperature and humidity. Results
showed that high levels of humidity were maintained, regardless of the external
environmental conditions. The high levels of humidity were caused mainly by the
presence of the ants inside the nests, as confirmed by an experimental

3
manipulation. Threshold temperatures, in which development stops, were not
reached although it is possible that these temperatures are reached during the dry
season. With base on our results and previous findings, a mechanism that O.
smaragdina would use to cope with low temperature and humidity is proposed.
Implications of our findings for the ecology of O. smaragdina and their nests are
discussed. Finally, we discuss the implications of our findings for other arboreal
ant species and at a bigger scale.
Introduction
The climate of a given region is fundamental in determining the species
composition and distribution of a given region. However, on a smaller scale
organisms experiment microclimates that can vary widely in time and space
(Kearney and Porter 2009; Sears et al. 2011) and differ from the general
macroclimate (Geiger et al. 2009). Ultimately the life history, distribution and
abundance of species reflect the operation of ecological interactions and metabolic
processes in the context of varying environments, especially temperature and
water availability (Andrewartha and Birch 1954). These two abiotic factors are
also fundamental in determining the abundance and distribution of a diverse
group: insects (Chown and Nicolson 2004), mainly because of two reasons. First,
because they are small they have a high surface area to volume ratio that makes
them more vulnerable than bigger animals to stressful environmental conditions.
They loss and gain heat more readily (Hood and Tschinkel 1990; Chown and
Nicolson 2004). Secondarily, because insects are ectotherms they rely on external
heat sources to perform their basic functions such as locomotion, growth and

4
reproduction (Deutsch et al. 2008). Therefore the incorporation of the effects of
these two important factors is essential in the study of the ecology of insects.
Because most insects are poikilothermic ectotherms, they allow their temperature
to vary considerably and are not able to generate their own metabolic heat. Their
activities perform optimally only if their body temperatures fall within a range that
permits the necessary biochemical, developmental, physiological, and behavioral
functions (Harrison et al. 2012). This range is determined by individual age, size,
color, water reserves and other factors (Schowalter 2011) and for most insects
these ranges are narrow. For example, for the fire ant Solenopsis invicta its range
has been determined to go from 22 to 32°C (Porter 1988) outside which colony
development stops or, at extreme temperatures, ants die. Insects must respond,
physiologically or behaviorally, to changes in environmental temperature in order
to stay within their optimal temperature range (Chown and Nicolson 2004).
Because of these, insects are more vulnerable to environmental change in
temperature than bigger or endotherm organisms that can generate their own
heat.
The maintenance of physiologically adequate water levels is also a constant
challenge for small organisms. As noted by Harrison et al. (2012) dehydration can
increase ion concentrations affecting membrane potentials and causing the
denaturation of proteins. Other insects face the opposite problem (e.g. aquatic
insects, sap feeders) of having too much water, in which case blood dilution can
destroy cells, change ion concentration and increase the insects volume and weight
(Benoit and Denlinger 2010). Insects acquire water by drinking, from food and by

5
the aerobic respiration and they loss it by excretion, via the cuticle and through the
spiracles (Buckley et al. 2010). As with temperature, insects must maintain
adequate water levels of humidity in their bodies in order to perform their
activities (Chown 2002). For example, a study by Zachariassen (1991) in the
routes of water loss in the beetle Phrynocolus petrosus, that inhabits African dry
savannas, found that cuticular water permeability was considerably lower than
related species that live in more benign habitats. Similar results have been found
for many other insect species (Zachariassen 1996; Klok and Chown 1998; Addo-
Bediako et al. 2001; Chown and Nicolson 2004), showing the importance of
maintaining adequate levels of humidity for this group.
Enclosed in one of the biggest insect orders, Hymenoptera, the family of ants
represents the most diverse and ecologically important group of social insects
(Hölldobler and Wilson 1990; Grimaldi and Engel 2005). With 12500 species
described (Bolton et al. 2007), ants have colonized almost all terrestrial habitats.
Since their first appearance about 140 million years ago (Brady et al. 2006;
Moreau et al. 2006), ants have played important ecological roles as soil turners,
scavengers and predators of many plants and animals. They also engage in a
myriad of symbiotic relationships with other insects, plants and bacteria
(Hölldobler and Wilson 1990; Ness et al. 2010). The understanding of the
adaptations this insects have developed to become such a cosmopolitan group is
essential to understand their biology.
Ants as most insects are poikilothermic ectotherms, although there are a few
species that can generate their own metabolic heat (e.g. Rosengren et al. 1987;

6
Franks 1989). Some ant species have managed to adapt to habitats with extreme
temperature conditions. In Western Australia, for example, the ant Melophorus
bagoti ants start foraging when temperatures reach 50°C and can survive
temperatures of up to 70°C (Christian and Morton 1992). On the other extreme of
the spectrum Camponotus obscuripes workers can survive temperatures of -20°C
for short periods of time (Ohyama and Asahina 1972). Most ant species, however,
are comfortable at temperatures ranging from 10 to 40°C (Kaspari 2000) and they
have evolved many adaptations to maintain themselves within their optimal
temperature ranges (Hölldobler and Wilson 1990). Temperature then, is one of the
most important factors limiting the distribution and abundance of ants at large and
small scales.
The maintenance of physiologically adequate water levels is also a constant
challenge for ants. As with temperature, different ant species have different
moisture preferences. For example, a study of carpenter ant queens, Camponotus
modoc and C. vicinus, found that colony founding (the establishment of a new
colony) in one species was less sensitive to wood moisture content, suggesting that
this is better adapted to low moisture conditions (Mankowski and Morrell 2011).
Brood development is also strongly dependent on nest temperature and humidity,
with an ideal range between 20 to 35°C outside which colony growth is inhibited
(Crozier et al. 2010). On the other extreme high air humidity, fog, rain and
waterlogging may limit foraging activities of ants and even destroy their colonies
(Brown 1959; Skinner 1980; Milford 1999; Kaspari 2000; Ballinger et al. 2007).
That is why it is important for ants to keep adequate levels of humidity, not only
when they are foraging for food but also inside their nests.

7
The ecological success of ants is principally due to their capacity to build nests that
allow them to be active when other non-social insects need to seek refuge and also
to raise their brood under stable conditions. While foragers are directly affected by
the pre-existing ambient temperature (Malsch et al. 2008) and humidity (Kaspari
and Weiser 2000), ants in the colony can control the microenvironment by
modifying the structure or location of the nest (e.g. digging deeper nests) or by
clustering together to retain metabolic heat and moisture (Hölldobler and Wilson
1990). At high latitudes most ant species nest under the ground and create
networks of underground galleries that allow them to create a gradient of
microclimates (Mikheyev and Tschinkel 2004). While we approach to the tropics
we see that as conditions become milder and more stable more ants species nest
and forage above ground and over the vegetation (Hood and Tschinkel 1990; Ward
2000). Most studies however, have focused on the regulation of temperature and
humidity in ground-nesting high-latitude species (see Jones and Oldroyd 2007 and
references therein) and invasive species (e.g. Hartley and Lester 2003; Xu et al.
2009). More research is needed in tropical canopy-nesting ant species in order to
understand how they cope with changes in temperature and humidity at this
microhabitat in which they reach high densities (Davidson and Patrell-Kim 1996).
The green tree ant, Oecophylla smaragdina, is widely distributed in India, across
tropical Asia, the Solomon Islands and northern Australia (Holldobler 1983). They
are known among entomologists for the conspicuous nests they build at treetops,
by gluing leaves together with larval silk, and by their aggressive behavior (Crozier
et al. 2010; Newey et al. 2010). They form large colonies that are usually spread
among many nests across their territories, can dominate up to 50 to 75% of the

8
trees and host more than half a million workers (Lokkers 1990). Lokkers
(1986,1990) proposed that the distribution of this species in Australia is primarily
determined by temperature and humidity and even produced a formula to predict
the likeliness of this species presence at a given site. However, there have not been
studies of the microclimatic conditions inside the nests of this conspicuous species,
or of any other ant species, and the main factors determining it.
In this study I investigate how temperature and humidity vary inside green tree
ant nests, in comparison with the external environment, to determine if the extent
to which green tree ants nests may buffer the colony against changes in
temperature and humidity. I also examine how any microclimate buffering may be
influenced by the elements that comprise the nest itself: the nest walls made of
living leaves, nest walls made of dead leaves and/or the presence of living ants.
Methods
Study Site
The study was conducted at the campus of James Cook University, Townsville,
Australia (19°19’40’’S, 146°45’31’’E; 39m). A total of 52 green ant, Oecophylla
smaragdina, colonies were located at trees and bushes around the university
campus. From these, 36 were randomly selected. A temperature/humidity Ibutton
(model DS1923-F5, Maxim Integrated ™) enclosed by a rubber cover was placed
inside each nest, along with a second Ibutton within 30 cm outside of the nest
opening (Figure 1).

9
Thirty-six Ibuttons were deployed to record temperature and relative humidity at
20 minutes intervals from February 25th to March 19th. Because of some
eventualities (e.g. lost Ibuttons on the field, data corrupted in Ibuttons, nests
abandoned by ants during the study period, nests withering and falling off) the
number of effective nests usable for our analysis was reduced to 28 (Active Green
treatment=9, Empty Green=5, Empty Withered=4, Displacement=10, see
explanation of treatments below). This period coincided with the late wet season
(Lokkers 1990).
Nest selection and field study design
To test the whether nests buffered the ants from variations in temperature and
humidity 12 nests with green leaves that contained active colonies were randomly
selected (Active Green nests, n=9) (Fig. 2A). Previous studies have shown that
sometimes these ants abandon their nests and that the leaves that form them keep
the nest shape (Lokkers 1990). Our experimental design took advantage of this
phenomenon. To explore further how much of the variation in temperature and
humidity was explained either by the presence of the ants or the leaves that
formed the nest 6 nests with green leaves but without ants were selected (Empty
Green nests, n=5) (Fig. 2B). 6 nests with withered leaves and without ants as a
control were also selected (Empty withered nests, n=4) (Fig.2C).
To explore how the environmental buffering effect changes within a nest after ants
abandon it and understand better the role of ants in determining the internal
microclimate, an experimental manipulation was performed. Twelve nests were
selected with active colonies and green leaves and were given a treatment after the

10
first week of data recording (Displacement nests, n=10) in order to make the ants
abandon them. Many treatments to displace ants were tested initially: inserting
naphthalene pellets inside the nests, spraying the nests with insect repellant,
poking the nests, etc. None of these treatments effectively displaced ants out.
Naphthalene returned mostly mixed results, repellant too, with the disadvantage
that it killed the plants leaves, and poking was the most ineffective of all as it just
triggered a defensive response by ants. Using smoke allowed us to get ants to
evacuate the nest. To produce the smoke 3 to 4 A4 paper printing sheets were
burned inside a small cylindrical metallic container (13*11 cm) and then some dry
vegetation from the ground was added to produce smoke. The nests were poked,
while holding the container about 20 cm away from the nest, to make sure that
ants left. This process went for 4 to 7 minutes in each nest.
To explore the influence of some relevant environmental factors on the variations
of temperature and humidity the length and width of every nest were measured,
across their longest two axis. These two measures were added in order to obtain
an approximation of the nest size. As canopy cover is important in determining the
microclimate in the understory (Marquis et al. 1986) 5 measures of the canopy
cover were taken, one just above the nest and other four two meters apart from
the center of the nest forming a cross with 90° angles. To take these measures a 10
cm. diameter tube with a mirror attached to the bottom and a square grid at the
top, a densiometer, was used. All the measurements were taken by the same
person and at the same height above ground (≈ 1.76 cm).

11
Statistical analysis
Because they were taken along time, data points at each nest were not
independent (e.g. they were auto correlated). To avoid this drawback we
calculated the mean temperature and humidity in every nest and comprised all the
data points into a single number (e.g. mean temperature and humidity values for
each nest were calculated). Also, because sometimes data inside data loggers was
corrupted for one of the variables the total number of colonies available was not
the same for each variable (Temperature=24 colonies, AG=7, EG=5, EW=4, D=8;
Humidity=18, AG=3, EG=4, EW=4, D=7). Shapiro tests were performed to
determine if data was normally distributed and used analysis of variances, if data
was normally distributed, or non-parametric Kruskall-Wallis test to determine if
there were differences in temperature and humidity between the treatments and
between the treatments and the outside. Because sample size was unbalanced
between treatments type-III sum of squares were used in the ANOVAs.
Planned post-hoc comparisons were performed on ANOVA results when
significant differences were found. The Fisher least significant differences method
was used to compare treatments with the conditions outside used as a control and
using the ‘comparison-wise’ error type, which gives the narrowest confidence
intervals for the difference between the two means. Displacement colonies were
not included in this analysis.
Maximum and minimum temperature and humidity values were not used, as a
more conservative approach, and to avoid the influence of outliers, we calculated
the 5th and 95th percentiles for each colony and each variable. We used this values

12
to test if there were differences in maximum and minimum temperature and
humidity using the same procedure described above.
To verify the changes in the microclimatic conditions inside the displacement
nests, after the ants were forced to abandon them, the mean humidity inside each
nest at the start (from the second to the third day) and at the end of the study
period (from the twentieth to the twenty-second day) were calculated and then a
two-sample equal variances t-Test was performed. Finally, Pearson´s correlation
tests were performed between each colony mean temperature, humidity and
colony size and mean canopy cover, all colonies were included in these analyses.
All analyses were performed on S-Plus 8.2 (TIBCO Software Inc.).
Results
The temperature inside the nests varied from 18.4°C to 45.3°C with a mean of
26.96±3.32°C. Outside nests mean temperature was 26.77±3.02°C with a minimum
of 18.3°C and a maximum of 37°C (Table 1). No differences could be detected in
mean temperature neither between treatments nor between inside and outside
nests (ANOVA, F2,30=1.16; p=0.34). Similar results were found regarding the
minimum temperature (e.g. 5th temperature percentile)(ANOVA, F2,24=1.54;
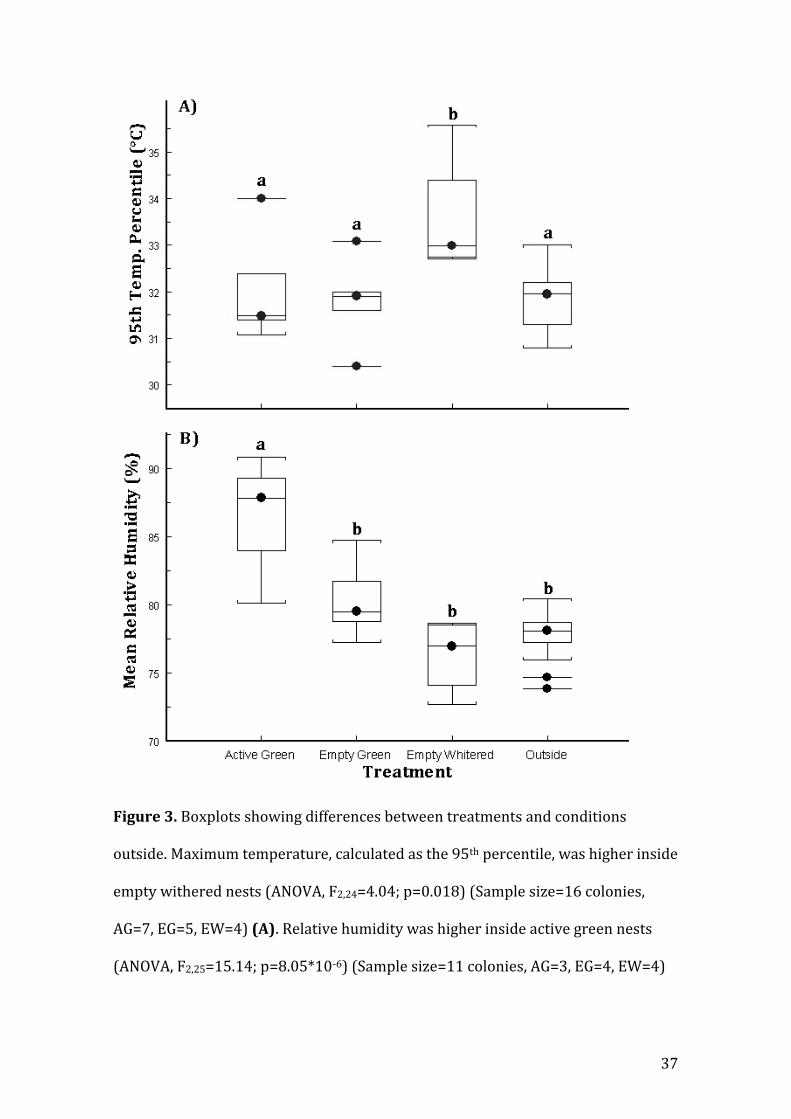
p=0.23). However, significant differences in maximum temperature (e.g. 95th
temperature percentile) between treatments were found (ANOVA, F2,24=4.04;
p=0.018). Fisher comparisons showed that EW nests had significantly higher
maximum temperatures in comparison to the other treatments and conditions
outside (Fig. 3A).

13
Relative humidity inside nests varied from 24.6% to a 100%, with a mean of
81.78±13.08%. Outside nests mean humidity was 78.11±14.02% with a minimum
of 29% and reaching 100% (Table 2). No differences could be detected in
maximum humidity (e.g. 95th humidity percentile) between treatments or between
inside and outside nests (Kruskal-Wallis rank test, X2=3.42, df=3, p=0.33).
However, significant differences in minimum (e.g. 5th humidity percentile)
(ANOVA, F2,24=14.9; p=0.00001) and mean humidity (ANOVA, F2,25=15.14;
p=8.05*10-6) between treatments were found (Figure3B). Both minimum and
mean humidity were significantly higher inside active green nests. To explore how
the differences in humidity varied in each treatment the difference between the
95th and 5th humidity percentiles for each colony was calculated. Significant
differences were found (Kruskal-Wallis rank test, X2=14.32, df=3, p=0.0025, Fig. 4).
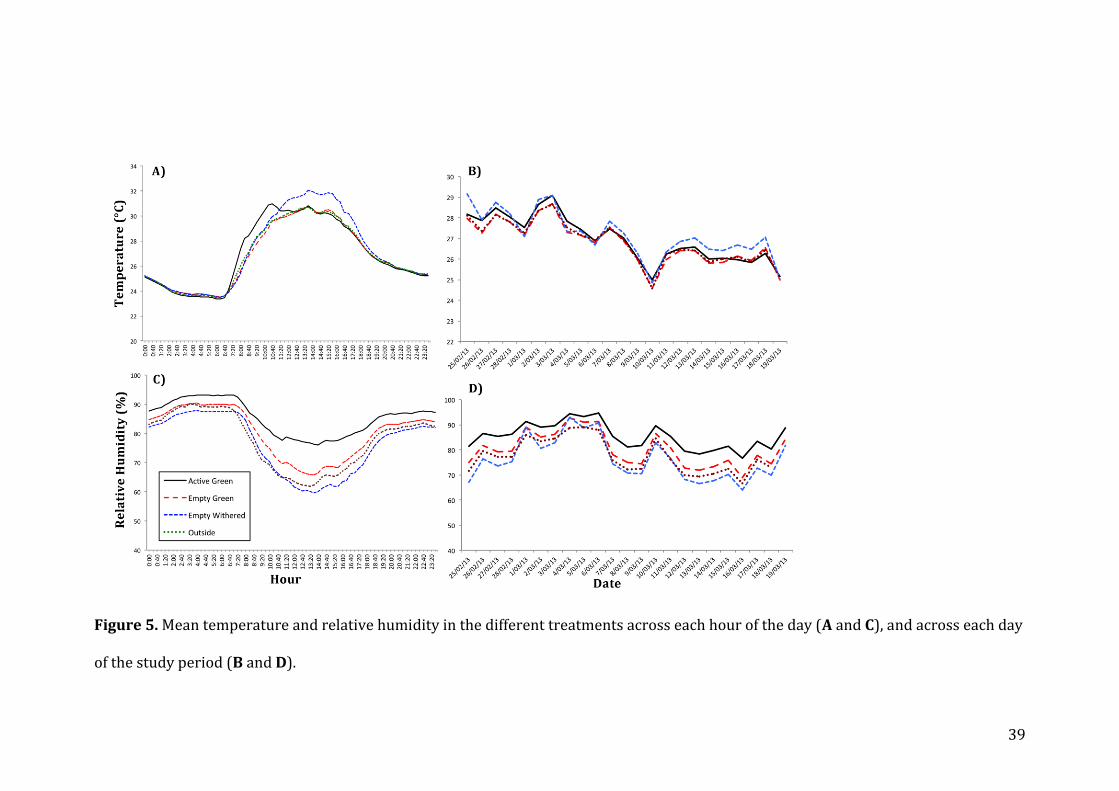
From the visual inspection of the temperature and humidity data along time it was
found that temperature inside nests during the day did not differ considerably
from the other treatments, and in the night and noon these differences disappear
completely (Fig. 5A). Additionally, it was observed that active green nests heated
earlier in the morning than the other treatments (Fig. 5A). Temperature tended to
decrease for all treatments as the study progressed (Fig. 5B). Humidity was clearly
higher inside active green nests at every hour of the day (Fig. 5C) when compared
to the other treatments and it also varied less when comparing the cold hours of
the day with the warmer ones. A similar pattern could be seen along all the study
period (Fig 5D).

14
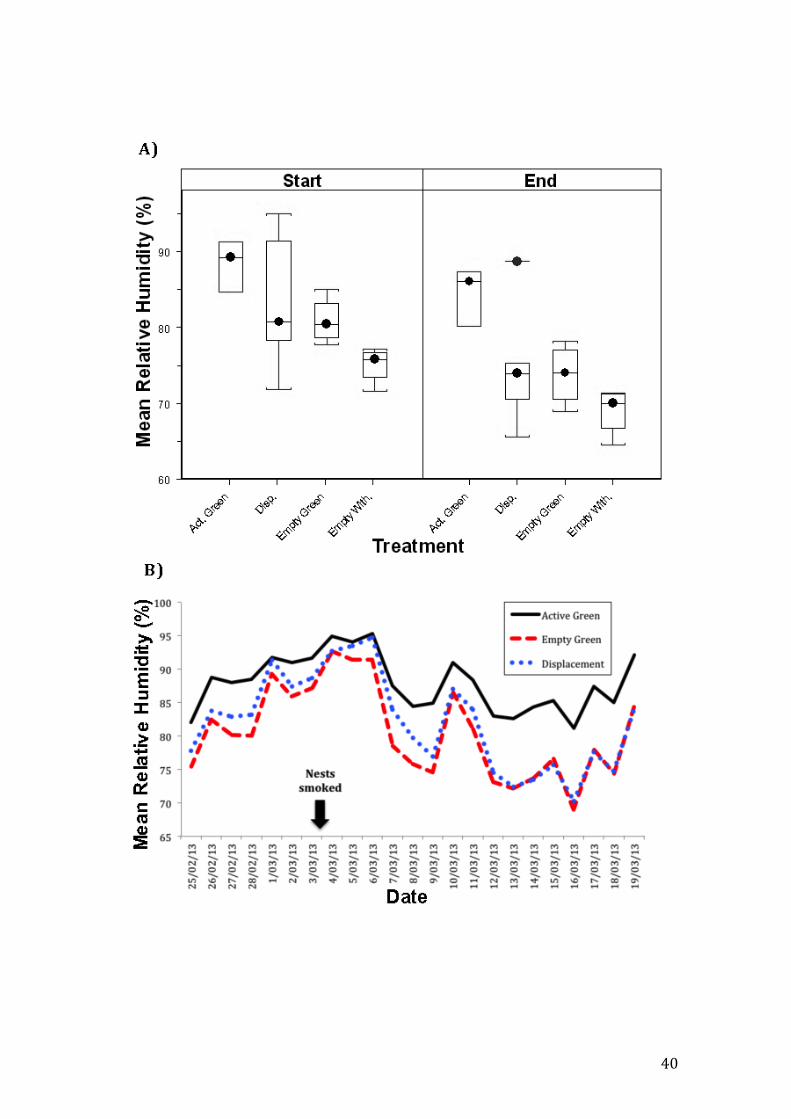
In the displacement colonies, the internal microclimate at beginning of the
experiment was similar to that of active green nests and after the smoking
treatment that displaced the ants, conditions inside became similar to those of
empty green nests (t2,34=-3.07, p=0.004, Fig. 6). The average nest length was
26.08±9.27 cm., the average width 19.34±5.64 cm. and the mean canopy cover was
76.62±18.38% for all the 28 colonies measured. A significant (negative)
correlation was found between temperature inside nests and canopy cover
(Pearson’s correlation, r=-0.47, t2,23 =-2.5282, p=0.0019, Fig. 7), no significant
correlation was found between nest humidity and canopy cover (Spearman’s rank
correlation, rs=0.07, z=0.34, p=0.73). No significant correlation was found between
temperature (r=0.24, t(2)23 =1.17, p=0.25) or humidity (r=-0.12, t(2)20 =-0.54,
p=0.59) and nest size.
Discussion
Previous studies by Lokkers (Lokkers 1986,1990) showed that temperature and
humidity are the main factors that determine the distribution of green tree ants.
However, studies of the microclimate inside the nests of this species were lacking.
In this study we found that throughout the wet season the nests of Oecophylla
smaragdina maintain high levels of humidity, regardless of the external
environmental conditions. These high levels of humidity are caused by the
presence of the ants inside the nests, although the transpiration of the leaves
partially contributes to increase the humidity. In contrast, inside empty withered
nests, humidity varies in correspondence to environmental humidity. This shows
that humidity is not only important in determining the big scale distribution of this

15
species but that it is also important at the microhabitat scale inside green tree ant
nests.
Not only did ants maintained high levels of humidity inside their nests throughout
the whole period but also the variation in humidity (the difference between
maximum and minimum humidity) was less pronounced than the other
treatments. Although studies on the maintenance of stable humidity conditions are
scarce in comparison to those studying variations in temperature, some have
found similar results. The distribution of the Argentine ant Linepithema humile, for
example, is limited primarily by the soil moisture content (Walters and Mackay
2003) because it builds its nests near the surface and it can not cope with
variations in humidity (Newell and Barber 1913). Neotropical leaf-cutting ants,
which maintain several fungus inside their nests as a food source, need to maintain
an appropriate climate inside their nest to maximize the growth of the fungus
(Farji-Brener 2000). Interestingly a study that investigated the responses of
workers of the leaf-cutting ant Acromyrmex heyeri to the conflicting demands of
temperature and humidity conditions inside their nests found that they traded off
the response related to thermoregulation (i.e. they tolerated higher temperatures)
for the maintenance of appropriate minimum levels of internal nest humidity
necessary for the growth of their fungus and brood (Bollazzi and Roces 2010).
Similar results have been found for Holarctic ants (Banschbach et al. 1997). Our
results, in addition to those from previous studies, show that the maintenance of
high and stable levels of humidity might be also important for ant species that nest
over the vegetation and in the canopy of tropical regions.

16
During the sampling period the minimum threshold temperatures were not
reached. Temperature fluctuated from 18.9°C to 41.4°C. Previous studies by
Lokkers (1990) determined that one of the most important factors limiting the
growth of green tree ant colonies is minimum temperature. Specifically, in
laboratory trials he determined the minimum growth temperature of the different
developmental stages and found that while eggs and pupae can withstand
relatively low temperatures (10 and 7°C respectively), larvae would stop growing
at temperatures lower to 17°C. While this temperature was not reached at any
time during our study, it has been observed that during the dry season
temperatures can go below 12°C in this area (Lokkers 1990). Lokkers (1990) also
determined that larvae are produced during all year, so ants can build and repair
the nests; more studies on how larvae are protected from low temperatures during
the dry season are needed although a possible mechanism is discussed in following
paragraphs.
There are several potential explanations for the high levels of humidity recorded
inside green tree ant nests. While the evapotranspiration of the leaves that are
used to build the nest account for a good proportion of the humidity present inside
nests, it was the presence of the ants that significantly increased these levels. The
source of this humidity from ants could be the loss through their respiratory water
loss. It is known that the need to obtain oxygen from the air posses a problem for
insects, as they need to prevent water loss to the usually drier atmosphere (Chown
2002). Studies on North American dessert ant species, adapted to low humidity
conditions, have shown that respiratory transpiration ant cuticular water loss
account only for a small proportion of total water loss (Lighton and Feener Jr

17
1989; Lighton 1992; Quinlan and Lighton 1999). However in species like green
tree ants, which have a thinner cuticle and are not adapted to live in low humidity
conditions, respiratory transpiration and cuticular water loss might be more
significant. Not only would workers contribute to increase the humidity but eggs
and larvae would also contribute to increase these levels, mainly because they
have a higher surface area to volume ratio (Jones and Oldroyd 2007). Another
possible source of humidity would be that produced from the respiratory water
loss and cuticular transpiration of the associated insects that live inside green tree
ant nests (see discussion below). Mores studies are needed to identify the most
important sources for the humidity inside green tree ant nests.
It has been observed that the size and density of green tree ant nests vary
seasonally in the wet tropics. During the wet season, when temperature and
humidity are high, green tree ant colonies nests tend to be dispersed in many small
nests across the whole territory. During the dry season on the other hand, when
temperature and humidity are lower, colonies tend to contract into the core areas
of their territories into fewer but bigger nests (Lokkers 1990). Although this
phenomenon might be partially explained by the deciduousness of some trees, it
suggests that during the dry season bigger nests, which would have a lower
surface area to volume ratio, might buffer ants from lower temperature and
humidity. The modification of the nest architecture for microclimatic regulation
has also been observed for Solenopsis invicta (Cassill et al. 2002). The migration of
ants between nests has been observed in ant species of the Palearctic region (Brian
and Brian 1951; Pontin 1960) that relocate their nests into sites with greater
thermal isolation. In the tropics, this movement would also be related to the

18
conservation of water in the colder, but also dryer, seasons. However, other factors
like vegetation density and the degree of connectedness of the canopies, limit the
distribution of green tree ants at local scales (Lokkers 1990; Peng et al. 1997). The
study of the interaction of these factors on the distribution and size of the nests of
green tree ants and how they vary across seasons represent a promising field of
study.
A similar behavior has been observed for the sister species of green tree ants: O.
longinoda. In cocoa trees in Zanzibar, nests concentrated in the southern side of
the trees during winter and in the northern side during summer. This migration
was related to the position of the sun. Vanderplank (1960) suggested that this
might help ants to increase insolation levels throughout the seasons, while Way
(1954) proposed that this might also help ants to shelter from the desiccating
prevailing winds. A similar behavior in which workers move larvae and pupa
through the nest in order to rear them under appropriate conditions has been
observed in many ant species (Jones and Oldroyd 2007). Fire ants, Solenopsis
invicta for example, build mounds over their nests with tunnels and galleries inside
(Hölldobler and Wilson 1990). On cool mornings S. invicta ants move their brood
up into the mound to the side receiving the most sunlight. When temperature
exceeds optimal they move their brood inner into the nest (Penick and Tschinkel
2008). For green tree ants it is known that brood is moved from and to different
nests at night (Lokkers 1990), probably to avoid environmental stress. The
movement of brood from one portion of the nest to another to sites with an
adequate microclimate is a widespread behavior in ants, and it is also very likely to
be used by green tree ants.

19
Interestingly, it was found that temperatures reached their higher values inside
empty withered nests. Many factors could account for this (e.g. the loss of the
impermeable waxy cuticle, the loss of protective pubescence) but probably the two
most important factors are the complete loss of water from the leaves and the
change into darker colors, that might increase the heat absorbance of the nest
surface (Aston and Van Bavel 1972). Interestingly, even after withering some nests
continued to host major workers. In the field it was also noticed that these nests
might also serve as a refugee for other insects after ants abandon them,
particularly cockroaches (Blattaria). With base on these observations it seems like
even after the nests of green tree ants wither and lost their buffering capacity they
might still provide green tree ants on the periphery of the territory, and probably
other insects, with a refugee from other factors (e.g. predators, parasites).
Lokkers (1990) found that daily activity patterns are correlated to temperature at
the end of the wet season and during the dry season. This circadian pattern tends
to disappear as wet season starts and conditions are more benign. This has also
been observed in other arboreal ant species (Medeiros and Oliveira 2009; Camargo
and Oliveira 2012). Although a significant buffering effect of temperature inside
nests was not found, the activity of foragers outside nests is influenced by
temperature. Foraging activity is reduced by extreme temperatures and reaches a
maximum peak between 28 and 33° (Lokkers 1990), which falls within the range
of temperature recorded inside the nests. The activity of workers inside green tree
ant nests therefore should resemble that outside nests. Our results also showed
that while during at night and dawn temperature inside green active nests is not
different when compared with the outside, during the first hours of the mornings

20
active green nests heat faster than empty green or withered nests (Fig. 5A). This
would probably allow ants to start performing their activities earlier although only
studies correlating the activity of ants with the temperatures inside the nests
would allow to verify this.
Non-significant correlations were found for most of the covariables studied. No
significant correlation was found between canopy cover and nest humidity. This
can be explained by the fact that, as previously mentioned, green tree ants prefer
to build their nests under thick vegetation (Peng et al. 1997) where moisture levels
would be expected to be high in comparison to open areas. The lack of significance
between nest size and any of the two environmental variables could be due to the
fact that most of the nests would have been small in size. It is probable that during
the dry season, when nests tend to be bigger (Lokkers 1990), an effect of nest size
over temperature or humidity would be significant. It is also possible, however,
that this is just a sampling artifact due to our sample size that was not big enough
to detect an effect. Future studies must try to include bigger sample sizes to
improve the statistical power and verify these observations.
A significant negative correlation, however, was found between mean canopy
cover and the temperature inside nests. In tropical regions with seasonal climates
it has been generally found that the increase in vegetation cover, with the
consequent reduction in temperature, generally result in a local increase in the
relative abundance of the most common species (Retana and Cerdá 2000). In other
ecosystems, however, increasing habitat complexity and canopy cover may lead to
high diversity and reduced dominance in ants (Perfecto and Snelling 1995;

21
Perfecto and Vandermeer 1996). At our study site, however, the threshold
temperature below which the development of green tree ant stops (Lokkers 1990),
was never reached and, therefore, low temperatures would not limit the growth of
this species in the wet season. Moreover, it has been noted that O. smaragdina
prefers sites with a thick vegetation cover (Lokkers 1990). These results however
cannot be interpolated to the dry season when there are lower temperatures that
could potentially limit the growth of this species.
Another interesting field that needs to be researched is how the microclimatic
conditions inside green tree ant nests influence the arthropods associated with
them. It is well known now that ant nests represent environmentally buffered
habitats for many organisms (Hughes et al. 2008). Only for the neotropical Army
ant Eciton burchellii for example, 80 obligate associate organisms have been
described at their temporary nests (Rettenmeyer et al. 2011). Similarly, inside the
complex nests of leaf-cutting ants many organisms have been described; only for
the group of beetles for example 411 species have been found (Navarrete-Heredia
2001). Green tree ants with their woven nests are considered one of the great
achievements of organic evolution and many organisms living in association with
their nests have been described (e.g. salticid spiders, Allan and Elgar 2001; Nelson
et al. 2005; lycaenid butterflies, Saarinen 2006; beetles, Bily et al. 2008; moths and
flies, Leong and D'rozario 2012). More research is needed to determine the
influence of the microclimate inside green tree ant nests and how it influences
these organisms.

22
Green tree ants are also known for establishing mutualisms with sap-sucking
hemipterans (e.g. Blüthgen and Fiedler 2003; Schwartz and Cassis 2003; Crozier et
al. 2010) in which ants protect them in return for honeydew, a carbohydrate-rich
substance. Green tree ants have been found to build leaf nests around these insects
(Pers. Obs.). Besides benefiting from the protection from predators, hemipterans
might benefit from the humid microclimate inside green tree ant nests. Indirect
evidence for this comes from observations in Aphis clerodenroni in Australia. This
aphid is known for producing galls that deform the leaves of its host plant; where
the shoots are galled the leaves close around each other forming an almost
completely concealed refugee (Carver et al. 2003) that to a certain degree
resemble the leaf nests of green tree ants . This aphids may receive some of the
benefits that other hemiptera receive inside nests of green tree ants (e.g. humid
microclimate, protection from predators, etc.). Verifying this observation would
add another function of adaptive value to the already remarkable nests of green
tree ants.
During the field study we notice there were variations in the proportion of larval
silk used to build the nests. The physical properties of the silk of Oecophylla have
been described by Crewe and Thompson (1979); they found that the silk is
designed to withstand the numerous mechanical stresses to which it is subject in
holding the leaves of the nest together. It has also been proposed that the larval
silk may provide an hygienic substrate from which to build nests and that it might
even have antimicrobial properties that benefit weaving ants (Fountain and
Hughes 2011). In this study and on previous studies by Lokkers (1990), it has been
noticed that nests vary in the proportion of larval silk they have and in some cases

23
it can represent a very high proportion of the nest walls. It is possible that this
affects the isolating capacities of green tree ant nests and ultimately their interior
microclimate.
Overall, the results reported here show that high levels of humidity are maintained
inside nests of green tree ants, Oecophylla smaragdina, throughout the wet season.
Our study design allowed us to discriminate between the effects of the structure of
the nest itself (empty withered nests), the nest and the green leaves (empty green
nests) and the combined effects of all the factors plus the presence of the ants
inside (active green nests). Additionally, an experimental manipulation that
displaced ants out of their nests showed that after ants left, conditions inside
became similar to those of empty green nests, with lower levels of humidity.
Therefore, it can be confidently conclude that the high levels of humidity observed
inside nests are caused mostly by the presence and activity of ants inside the nests.
These would be another of the adaptations (Hölldobler and Wilson 1990; Crozier
et al. 2010) that enable green tree ants to rear their brood all year round and
dominate up to 50 to 70% of the trees of a given area, depending on the season
(Lokkers 1990). Not only are nests of green tree ants remarkable for the way in
which they are constructed but also because they represent environmental
‘fortresses’ (Wilson 1968) for their occupants.
The nests of social insects have been described as one of the pinnacles of biological
evolution. Many studies in high latitudes, have shown that nests have an important
function of thermal buffering and that this is the main factor that has allowed
many ant species to colonize those ecosystems (e.g. Farji-Brener 2000; Mikheyev

24
and Tschinkel 2004; Bollazi et al. 2008). In the tropics however, and specially for
arboreal ants, where species live in ecosystems with generally stable temperatures
(Diamond et al. 2012) humidity probably represents a more important factor at a
local scale. The nests of the so-called weaver ants, a complex of ants that includes
green tree ants and species from several genera that use larval silk to build their
nests over vegetation (Hölldobler and Wilson 1990), probably help them to
maintain high levels of humidity to perform their activities. This needs to be tested
for other species and in different seasons especially because arboreal ants have
been identified as one of the most vulnerable groups to changes due to climate
change (Diamond et al. 2012).
Acknowledgements
Simon Robson and Justin Welbergen helped to design this study, analyze the
results and provided valuable comments that helped to improve the quality of this
manuscript. Ben Phillips, Betsy Roznik and David Pike kindly loaned their Ibuttons.
An Australian Research Council Discovery Grant founded this study.
Literature cited
Addo-Bediako, A., Chown, S.L. and Gaston, K.J. 2001. Revisiting water loss in
insects: a large scale view. Journal of Insect Physiology 47: 1377-1388.
Allan, R.A. and Elgar, M.A. 2001. Exploitation of the green tree ant, Oecophylla
smaragdina, by the salticid spider Cosmophasis bitaeniata. Australian Journal of
Zoology 49: 129-137.
Andrewartha, H.G. and Birch, L.C. 1954. The distribution and abundance of
animals. The University of Chicago Press, Chicago, IL.

25
Aston, A. and Van Bavel, C. 1972. Soil surface water depletion and leaf temperature.
Agronomy Journal 64: 368-373.
Ballinger, A., Lake, P.S. and Nally, R.M. 2007. Do terrestrial invertebrates
experience floodplains as landscape mosaics? Immediate and longer-term effects
of flooding on ant assemblages in a floodplain forest. Oecologia 152: 227-238.
Banschbach, V.S., Levit, N. and Herbers, J.M. 1997. Nest temperatures and thermal
preferences of a forest ant speies: is seasonal polydomy a thermoregulatory
mechanism. Insectes Sociaux 44: 109-122.
Benoit, J.B. and Denlinger, D.L. 2010. Meeting the challenges of on-host and off-host
water balance in blood-feeding arthropods. Journal of Insect Physiology 56: 1366-
1376.
Bily, S., Fikacek, M. and Sipek, P. 2008. First record of myrmecophily in buprestid
beetles: immature stages of Habroloma myrmecophila sp. nov.(Coleoptera:
Buprestidae) associated with Oecophylla ants (Hymenoptera: Formicidae). Insect
Systematics & Evolution 39: 121-131.
Blüthgen, N. and Fiedler, K. 2003. Disentangling a rainforest food web using stable
isotopes: dietary diversity in a species-rich ant community Oecologia 29: 418-429.
Bollazi, M., Kronenbitter, J. and Roces, F. 2008. Soil temperature, digging
behaviour, and the adaptive value of nest depth in South American species of
Acromyrmex leaf-cutting ants. Oecologia 158: 165-175.
Bollazzi, M. and Roces, F. 2010. Leaf-cutting ant workers (Acromyrmex heyeri)
trade off nest thermoregulation for humidity control. Journal of Ethology 28: 399-
403.

26
Bolton, B., Alpert, G., Ward, P.S. and Naskrecki, P. 2007. Bolton's catalogue of ants
of the world. Harvard University Press, Cambridge, MA. CD-ROM.
Brady, S.G., Schultz, T.R., Fisher, B.L. and Ward, P.S. 2006. Evaluating alternative
hypotheses for the early evolution and diversification of ants. Proceedings of the
National Academy of Sciences of the United States of America 103: 18172-18177.
Brian, M.V. and Brian, A.D. 1951. Insolation and ant population in the west of
Scotland. Transactions of the Royal Entomological Society of London 102: 303-330.
Brown, E.S. 1959. Immature nutfall of coconuts in the Solomon Islands. II. Changes
in ant populations and their relation to vegetation. Bulletin of Entomological
Research 50: 523-558.
Buckley, L.B., Davies, T.J., Ackerly, D.D., Kraft, N.J.B., Harrison, S.P., Anacker, B.L.,
Cornell, H.V., Damschen, E.I., Grytnes, J.A. and Hawkins, B.A. 2010. Phylogeny, niche
conservatism and the latitudinal diversity gradient in mammals. Proceedings of the
Royal Society B: Biological Sciences 277: 2131-2138.
Camargo, R.X. and Oliveira, P.S. 2012. Natural history of the neotropical arboreal
ant, Odontomachus hastatus: nest sites, foraging schedule, and diet. Journal of
Insect Science 12: 1-9.
Carver, M., Blüthgen, N., Grimshaw, J.F. and Bellis, G.A. 2003. Aphis clerodendri
Matsumura (Hemiptera: Aphididae), attendant ants (Hymenoptera: Formicidae)
and associates on Clerodendrum (Verbenaceae) in Australia. Australian Journal of
Entomology 42: 109-113.
Cassill, D., Tschinkel, W. and Vinson, S. 2002. Nest complexity, group size and
brood rearing in the fire ant, Solenopsis invicta. Insectes Sociaux 49: 158-163.
Chown, S. 2002. Respiratory water loss in insects. Comparative biochemistry and
physiology-part A: Molecular & Integrative Physiology 133: 791-804.

27
Chown, S.L. and Nicolson, S. 2004. Insect physiological ecology: mechanisms and
patterns. Oxford University Press, New York.
Christian, K.A. and Morton, S.R. 1992. Extreme thermophilia in a central Australian
ant, Melophorus bagoti. Physiological Zoology 65: 885-905.
Crewe, R. and Thompson, P. 1979. Oecophylla silk: functional adaptation in a
biopolymer. Naturwissenschaften 66: 57-58.
Crozier, R.H., Newey, P.S., Schluns, E.A. and Robson, S.K.A. 2010. A masterpiece of
evolution - Oecophylla weaver ants (Hymenoptera: Formicidae). Myrmecological
News 13: 57-71.
Davidson, D.W. and Patrell-Kim, L. 1996. Tropical arboreal ants: why so abundant?
In: Gibson, A.C. (Ed.) Neotropical biodiversity and conservation. UCLA Botanical
Garden, Los Angeles, California, pp. 127-140.
Deutsch, C.A., Tewksbury, J.J., Huey, R.B., Sheldon, K.S., Ghalambor, C.K., Haak, D.C.
and Martin, P.R. 2008. Impacts of climate warming on terrestrial ectotherms across
latitude. Proceedings of the National Academy of Sciences 105: 6668.
Diamond, S.E., Sorger, D.M., Hulcr, J., Pelini, S.L., Del Toro, I., Hirsch, C., Oberg, E. and
Dunn, R. 2012. Who likes it hot? A global analysis of the climatic, ecological, and
evolutionary determinants of warming tolerance in ants. Global Change Biology
18: 448-456.
Farji-Brener, A.G. 2000. Leaf-cutting ant nests in temperate environments:
mounds, mound damages and nest mortality rate in Acromyrmex lobicornis. Studies
on Neotropical Fauna and Environment 35: 131-138.
Fountain, T. and Hughes, W. 2011. Weaving resistance: silk and disease resistance
in the weaver ant Polyrhachis dives. Insectes Sociaux 58: 453-458.

28
Franks, N.R. 1989. Thermoregulation in army ant bivouacs. Physiological
Entomology 14: 397-404.
Geiger, R., Aron, R.H. and Todhunter, P. 2009. The climate near the ground.
Rowman & Littlefield Publishers.
Grimaldi, D. and Engel, M.S. 2005. Evolution of the insects. Cambridge University
Press, Cambridge.
Harrison, J.F., Woods, H.A. and Roberts, S.P. 2012. Ecological and environmental
physiology of insects. Oxford University Press, Oxford.
Hartley, S. and Lester, P.J. 2003. Temperature-dependent development of the
argentine ant, Linepithema humile (Mayr)(Hymenoptera: Formicidae): a degree-
day model with implications for range limits in New Zealand. New Zealand
Entomologist 26: 91-100.
Holldobler, B. 1983. Territorial Behavior in the green tree ant (Oecophylla
smaragdina). Biotropica 15: 241-250.
Hölldobler, B. and Wilson, E. 1990. The ants. Belknap Press.
Hood, W. and Tschinkel, W. 1990. Desiccation resistance in arboreal and terrestrial
ants. Physiological Entomology 15: 23-35.
Hughes, D.P., Pierce, N.E. and Boomsma, J.J. 2008. Social insect symbionts:
evolution in homeostatic fortresses. Trends in Ecology & Evolution 23: 672-677.
Jones, J. and Oldroyd, B. 2007. Nest thermoregulation in social insects. Advances in
Insect Physiology 33: 153-191.

29
Kaspari, M. 2000. A primer on ant ecology. In: Agosti, D., Majer, J.D., Alonso, L.,
Schultz, T.R. (Eds.) Ants: Standard methods for measuring and monitoring
biodiversity. Smithsonian Institution Press, Washington & London.
Kaspari, M. and Weiser, M. 2000. Ant activity along moisture gradients in a
neotropical Forest. Biotropica 32: 703-711.
Kearney, M. and Porter, W. 2009. Mechanistic niche modelling: combining
physiological and spatial data to predict species' ranges. Ecology Letters 12: 334-
350.
Klok, C.J. and Chown, S.L. 1998. Interactions between desiccation resistance, host-
plant conatact and the thermal biology of a leaf-dwelling sub-antarctic caterpillar,
Embryonopsis halticella (Lepidoptera: Yponomeutidae). Journal of Insect
Physiology 44: 615-628.
Leong, T.M. and D'Rozario, V. 2012. Mimicry of the weaver ant, Oecophylla
smargadina by the moth caterpillar, Homodes bracteigutta, the crab spider,
Amyciaea lineatipes, and the jumping spider , Myrmarachne plataleoides. Nature in
Singapore 5: 39-56.
Lighton, J. 1992. Direct measurement of mass loss during discontinuous ventilation
in two species of ants. Journal of Experimental Biology 173: 289-293.
Lighton, J.R. and Feener Jr, D.H. 1989. Water-loss rate and cuticular permeability in
foragers of the desert ant Pogonomyrmex rugosus. Physiological Zoology: 1232-
1256.
Lokkers, C. 1986. The distribution of the weaver ant, Oecophylla smaragdina
(Fabricius) (Hymenoptera: Formicidae) in Northern Australia. Australian Journal
of Zoology 34: 683-687.

30
Lokkers, C. 1990. Colony dynamics of the green tree ant (Oecophylla smaragdina
Fab.) in a seasonal tropical climate. PhD thesis, James Cook University of North
Queensland, Townsville, Queensland.
Malsch, A.K.F., Fiala, B., Maschwitz, U., Mohamed, M., Nais, J. and Lisenmair, K.E.
2008. An analysis of declining ant species richness with increasing elevation at
Mount Kinabalu, Sabah, Borneo. Asian Myrmecology 2: 33-49.
Mankowski, M.E. and Morrell, J. 2011. Role of relative humidity in colony founding
and queen survivorship in two carpenter ant species. Journal of Economic
Entomology 104: 740-744.
Marquis, R.J., Young, H.J. and Braker, H.E. 1986. The influence of understory
vegetation cover on germination and seedling establishment in a tropical lowland
wet forest. Biotropica 18: 273-278.
Medeiros, F. and Oliveira, P. 2009. Season-dependent foraging patterns case study
of a neotropical forest-dwelling ant (Pachycondyla striata; Ponerinae). In: Jarau, S.,
Hrncir, M. (Eds.) Food Exploitation by Social Insects: Ecological, Behavioral, and
Theoretical Approaches. CRC Press, pp. 81-95.
Mikheyev, A. and Tschinkel, W. 2004. Nest architecture of the ant Formica
pallidefulva: structure, costs and rules of excavation. Insectes Sociaux 51: 30-36.
Milford, E.R. 1999. Ant communities in flooded and unflooded riparian forest of the
middle Rio Grande. Southwest Naturalist 44: 278-286.
Moreau, C.S., Bell, C.D., Vila, R., Archibald, S.B. and Pierce, N.E. 2006. Phylogeny of
the ants: diversification in the age of angiosperms. Science 312: 101-104.
Navarrete-Heredia, J.L. 2001. Beetles associated with Atta and Acromyrmex ants
(Hymenoptera: Formicidae: Attini). Transactions of the American Entomological
Society 127: 381-429.

31
Nelson, X.J., Jackson, R.R., Edwards, G. and Barrion, A.T. 2005. Living with the
enemy: jumping spiders that mimic weaver ants. Journal of Arachnology 33(3):
813-819.
Ness, J., Mooney, K. and Lach, L. 2010. Ants as mutualists. In: Lach, L., Parr, C.L.,
Abbott, K.L. (Eds.) Ant Ecology. Oxford University Press, New York, pp. 97-114.
Newell, W. and Barber, T.C. 1913. The argentine ant. Bureau Entomological
Bulletin 122: 1-98.
Newey, P.S., Robson, S.K. and Crozier, R.H. 2010. Weaver ants Oecophylla
smaragdina encounter nasty neighbors rather than dear enemies. Ecology 91:
2366-2372.
Ohyama, Y. and Asahina, É. 1972. Frost resistance in adult insects. Journal of Insect
Physiology 18: 267-282.
Peng, R., Christian, K. and Gibb, K. 1997. Distribution of the green ant, Oecophylla
smaragdina (F.)(Hymenoptera: Formicidae), in relation to native vegetation and
the insect pests in cashew plantations in Australia. International Journal of Pest
Management 43: 203-211.
Penick, C.A. and Tschinkel, W. 2008. Thermoregulatory brood transport in the fire
ant, Solenopsis invicta. Insectes Sociaux 55: 176-182.
Perfecto, I. and Snelling, R. 1995. Biodiversity and the transformation of a tropical
agroecosystem: ants in coffee plantations. Ecological Applications 5: 1084-1097.
Perfecto, I. and Vandermeer, J. 1996. Microclimatic changes and the indirect loss of
ant diversity in a tropical agroecosystem. Oecologia 108: 577-582.
Pontin, A.J. 1960. Field experiments of colony foundation by Lasius niger (L.) and L.
flavus (F.) (Hym., Formicidae). Insectes Sociaux 7: 227-230.

32
Porter, S.D. 1988. Impact of temperature on colony growth and developmental
rates of the ant, Solenopsis invicta. Journal of Insect Physiology 34: 1127-1133.
Quinlan, M.C. and Lighton, J. 1999. Respiratory physiology and water relations of
three species of Pogonomyrmex harvester ants (Hymenoptera: Formicidae).
Physiological Entomology 24: 293-302.
Retana, J. and Cerdá, X. 2000. Patterns of diversity and composition of
mediterranean ground ant communities tracking spatial and temporal variability
in the thermal environment. Oecologia 123: 436-444.
Rettenmeyer, C., Rettenmeyer, M., Joseph, J. and Berghoff, S. 2011. The largest
animal association centered on one species: the army ant Eciton burchellii and its
more than 300 associates. Insectes Sociaux 58: 281-292.
Rosengren, R., Fortelius, W., Lindström, K. and Luther, A. 1987. Phenology and
causation of nest heating and thermoregulation in red wood ants of the Formica
rufa group studied in coniferous forest habitats in southern Finland. Annales
Zoologici Fennici 24: 147-155.
Saarinen, E.V. 2006. Differences in worker caste behaviour of Oecophylla
smaragdina (Hymenoptera: Formicidae) in response to larvae of Anthene emolus
(Lepidoptera: Lycaenidae). Biological Journal of the Linnean Society 88: 391-395.
Schowalter, T.D. 2011. Insect ecology: an ecosystem approach. Elsevier.
Schwartz, M.D. and Cassis, G. 2003. New genus and new species of
myrmecomorphic plant bug from Australia (Heteroptera: Miridae: Mirini).
Australian Journal of Entomology 42: 254-259.
Sears, M.W., Raskin, E. and Angilletta, M.J. 2011. The world is not flat: defining
relevant thermal landscapes in the context of climate change. Integrative and
Comparative Biology 51: 666-675.

33
Skinner, G. 1980. The feeding habits of the wood-ant, Formica rufa (Hymenoptera:
Formicidae), in limestone woodland in north-west England. The Journal of Animal
Ecology 49: 417-433.
Vanderplank, F.L. 1960. The bionomics and ecology of the red tree ant, Oecophylla
sp, and its relationships to the coconut bug Pseudotheraptus wayi. Journal of
Animal Ecology 29: 15-33.
Walters, A. and Mackay, D. 2003. An experimental study of the relative humidity
preference and survival of the Argentine ant, Linepithema humile (Hymenoptera,
Formicidae): comparisons with a native Iridomyrmex species in South Australia.
Insectes Sociaux 50: 355-360.
Ward, P.S. 2000. Broad scale patterns of diversity in leaf-litter ants communities.
In: Agosti, D., Majer, J.D., Alonso, L., Schultz, T.R. (Eds.) Ants: Standard methods for
measuring and monitoring biodiversity.Washington & London, pp. 99-121.
Way, M.J. 1954. Studies of the life history and ecology of the ant Oecophylla
longinoda. Bulletin of Entomological Research 44: 669-691.
Wilson, E.O. 1968. The ergonomics of caste in social insects American Naturalist
102: 41-66.
Xu, Y.J., Zeng, L., Lu, Y.Y. and Liang, G.W. 2009. Effect of soil humidity on the
survival of Solenopsis invicta Buren workers. Insectes Sociaux 56: 367-373.
Zachariassen, K. 1996. The water conserving physiological compromise of desert
insects. European Journal of Entomology 93: 359-368.
Zachariassen, K.E. 1991. Routes of transpiratory water loss in a dry habitat
tenebrionid beetle. Journal of Experimental Biology 157: 425-437.

34
Tables and Figures
Position Treatment Number of
colonies Mean SD Maximum Minimum
Inside Active Green 7 26.97 3.24 39.7 19.9
Empty Green 5 26.78 2.95 37 20.3
Empty Withered 4 27.18 3.51 41.4 18.9
Displacement 8 26.89 3.57 45.3 18.4
Total/Mean 24 26.96 3.32 40.85 19.375
Outside Active Green 7 26.81 3.00 35.6 19.8
Empty Green 5 26.72 2.78 35.5 20.2
Empty Withered 4 26.82 3.13 37 18.6
Displacement 8 26.73 3.18 36.5 18.3
Total/Mean 24 26.77 3.02 36.15 19.225
Grand Total/Mean 24 26.86 3.17 38.50 19.30
Table 1. Summary of temperatures (°C) recorded inside and outside nests across
the different treatments. Underlined numbers represent maximum and minimum
values. SD=Standard deviation.
Position Treatment Number of
colonies Mean SD Maximum Minimum
Inside Active Green 3 88.00 8.70 100 43.1
Empty Green 4 82.21 15.10 100 28.8
Empty Withered 4 80.62 13.16 100 32
Displacement 7 76.31 15.35 100 24.6
Total/Mean 18 81.78 13.08 100 32.1
Outside Active Green 3 79.18 13.73 100 35.8
Empty Green 4 77.83 14.83 100 30.9
Empty Withered 4 78.05 13.37 100 31.4
Displacement 7 77.38 14.15 100 29
Total/Mean 18 78.11 14.02 100 31.8
Grand Total/Mean 18 79.76 14.37 100 24.6
Table 2. Summary of the relative humidity (%) recorded inside and outside nests
across the different treatments. Underlined numbers represent maximum and
minimum values. SD=Standard deviation.

35
Figure 1. Picture of an active green nest on the field, the square shows the Ibutton
used to record temperature and humidity (A). Detail of the Ibutton inside the nest.
Notice how ants incorporated it into their nest (B). Picture of an Ibutton inside the
protective cases. Notice the plastic mesh used to protect the surface of the Ibutton
(C).

36
Figure 2. Photographs of the nests used for the study. Active Green nests were
defined as those formed by green leaves and occupied by ants (A). Empty green
nests were formed by green leaves but contained no ants (B) and empty withered
nests were those with withered brown leaves and containing no ants (C).
37
Figure 3. Boxplots showing differences between treatments and conditions
outside. Maximum temperature, calculated as the 95th percentile, was higher inside
empty withered nests (ANOVA, F2,24=4.04; p=0.018) (Sample size=16 colonies,
AG=7, EG=5, EW=4) (A). Relative humidity was higher inside active green nests
(ANOVA, F2,25=15.14; p=8.05*10-6) (Sample size=11 colonies, AG=3, EG=4, EW=4)

38
(B). Letters over the boxplots (a,b) represent groupings of significantly different
treatments as revealed by Fisher LSD comparisons.
Figure 4. Maximum (95th percentile) minus minimum (5th percentile) relative
humidity differences for each treatment. A Kruskal-Wallis test found significant
differences between treatments (Kruskal-Wallis rank test, X2=14.32, df=3,
p=0.0025).
39
Figure 5. Mean temperature and relative humidity in the different treatments across each hour of the day (A and C), and across each day
of the study period (B and D).
40

41
Figure 6. Mean humidity for each treatment inside nests at the start and at the end
of the study period (A). Mean relative humidity data for active green, empty green
and displacement treatments across the study period. Arrow signals the date of
nests smoking (B). A two-sample equal-variances t-test revealed significant
differences in humidity at the start of the study period in comparison with
humidity at the end (t2,34=-3.07, p=0.004, Fig. 6). After displacement nests were
smoked relative humidity became similar to that in empty green nests.
Figure 7. Scatter plot showing the correlation between temperature inside nests
and canopy cover (r=-0.47, t2,23 =-2.5282, p=0.019).







