
Natural Selection

DNA encodes information that interacts with the environment to influence phenotype
Among The Traits That Can Be Influenced By Genetically Determined Responses to the Environment Are:
1. The Viability in the Environment2. Given Alive, the Mating Success in the
Environment3. Given Alive and Mated, Fertility or
Fecundity in the Environment.

Viability
Hb- Locus In Africa:
A/A A/S S/S
Non-Malarial Area
No Anemia No Anemia Anemia
Viability: High High Low
Malarial AreaNot Resistant
to MalariaResistant Anemia
Viability: Low High Low
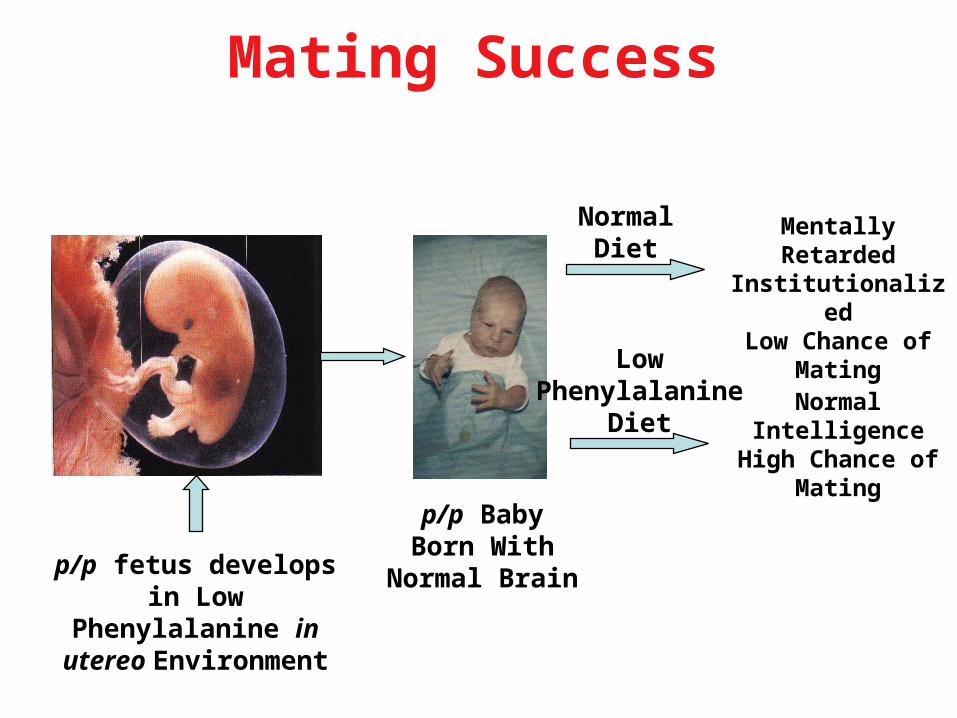
p/p fetus develops in Low
Phenylalanine in utereo
Environment
Mentally Retarded
Institutionalized
Low Chance of Mating
p/p BabyBorn With
Normal Brain
NormalDiet
LowPhenylalanine
Diet
Mating Success
Normal Intelligence
High Chance of Mating
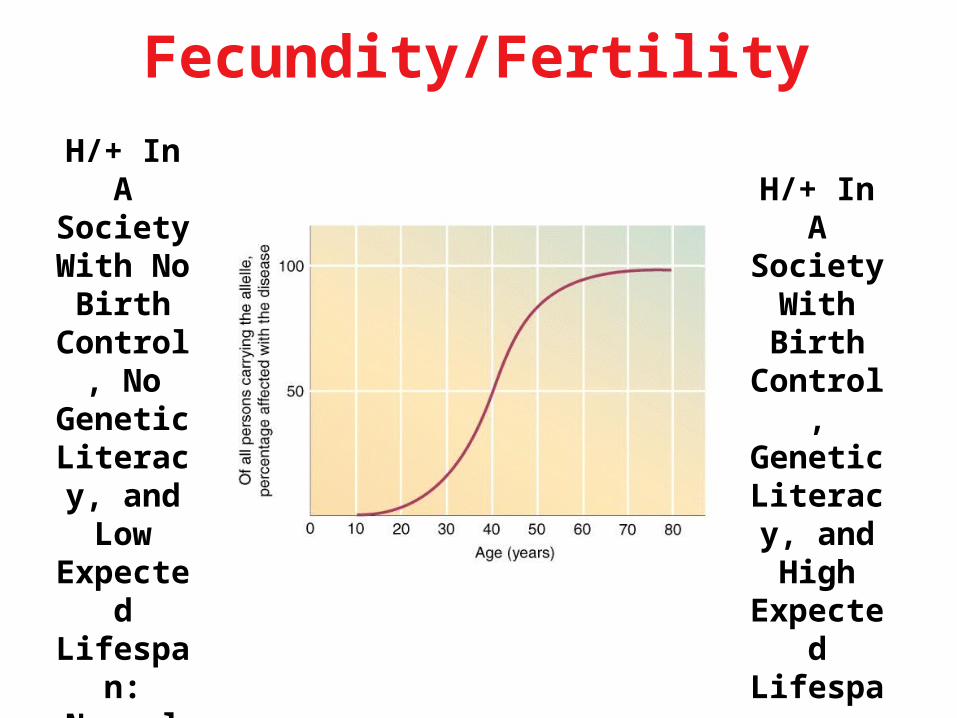
Fecundity/Fertility
H/+ In A Society
With No Birth
Control, No
Genetic Literacy, and Low Expected Lifespan:Normal
Fecundity
H/+ In A Society
With Birth Control, Genetic
Literacy, and High Expected Lifespan:
Low Fecundity

Why Are Viability, Mating Success, and Fecundity/Fertility
Important Phenotypes in Evolution?
Because All Of These Phenotypes Influence The Chances For
Successful DNA Replication

Physical Basis of Evolution
• DNA can replicate
• DNA can mutate and recombine
• DNA encodes information that interacts with the environment to influence phenotype

Physical Basis of Evolution
• DNA can replicate
• DNA can mutate and recombine
• DNA encodes information that interacts with the environment to influence phenotype
ViabilityMating Success
Fecundity/Fertility

Physical Basis of Evolution
• DNA can replicate
• DNA can mutate and recombine
• DNA encodes information that interacts with the environment to influence phenotype
ViabilityMating Success
Fecundity/Fertility
These Are Combined Into A SinglePhenotype of Reproductive Success
Or FITNESS

DNA can replicateDNA can mutate and recombine
Genotypic VariationIn Demes and Species
Environment
PhenotypicVariationIn Fitness
HeritableVariationIn Fitness

Natural Selection Is Heritable Variation In Fitness
That Is, The Genes Borne By A Gamete Influence The
Probability of That Gamete Being Passed On To The Next
Generation.
THINK LIKE A GAMETE!

DNA can replicateDNA can mutate and recombine
Genotypic VariationIn Demes and Species
It’s the Environment, stupid!
PhenotypicVariationIn Fitness
HeritableVariationIn Fitness
NATURAL SELECTION IS NOT CIRCULAR

Natural Selection At A Single Locus in A Randomly Mating Deme
AA
p2
Aa
2pq
aa
q2
Zygotic Frequencies
VAA VAa VaaViabilities
aa
q2 Vaa
Aa
2pqVAa
AA
p2 VAA
Environment
Adult Frequencies
CAA CAa CaaMating Prob.Environment
aa
q2 VaaCaa
Aa
2pqVAaCAa
AAp2 VAACAA
Mated Adult Frequencies
bAA bAa baaAve. No. OffspringEnvironment
aaq2 VaaCaabaa
Aa
2pqVAaCAabAa
AAp2 VAACAAbAA
Mated Adult FrequenciesWeighted By No. of Off.

Let WAA = VAACAAbAA; WAa = VAaCAabAa; Waa = VaaCaabaa
AA
p2
Aa
2pq
aa
q2
WAA
Zygotic Frequencies
WAa WaaFitness
AA
p2 WAA
Aa
2pqWAa
aa
q2 Waa
Environment
Mated Adult FrequenciesWeighted By No. of Off.
aa
q2Waa/W
Aa
2pqWAa/WAA
p2 WAA/WMated Adult Frequencies
Convert to Freq. By Dividing by = W = p2WAA+2pqWAa+q2Waa
1 1/21/2
aq’= q2 Waa/W + pqWAa/W
Ap’= p2 WAA/W + pqWAa/W
1Meiosis
Gene Pool
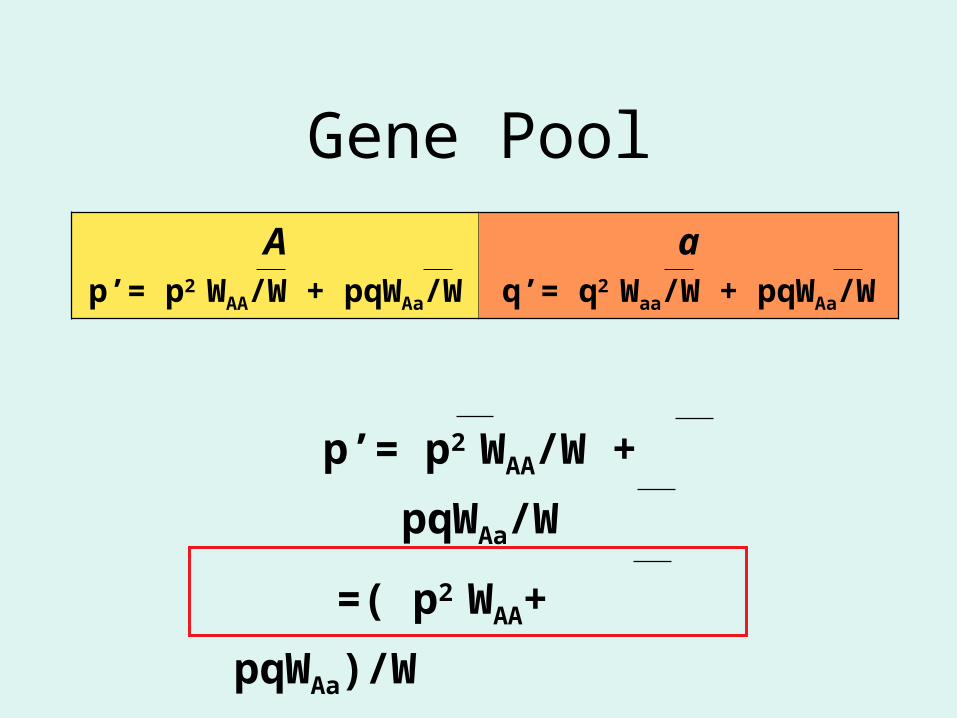
Ap’= p2 WAA/W + pqWAa/W
aq’= q2 Waa/W + pqWAa/W
Gene Pool
p’= p2 WAA/W + pqWAa/W
=( p2 WAA+ pqWAa)/W
p’ = p(pWAA+ qWAa)/W

Does Evolution Occur?
p = p’ - p
= p(pWAA+ qWAa)/W - p
= p[pWAA+ qWAa)/W - 1]
p = p[pWAA+ qWAa- W]/W

Does Evolution Occur?Note, W = W(p+q)=pW+qW
p = p[pWAA+ qWAa- W]/W
=p[p(WAA-W)+ q(WAa-W)]/W
Since p and W are always > 0,This is the only part of the equationThat Can Change Sign and Hence
Determine the Direction of EvolutionUnder Natural Selection.

Does Evolution Occur?What is:
p(WAA-W)+ q(WAa-W)?
Mean Phenotype of Fitness

Does Evolution Occur?What is:
p(WAA-W)+ q(WAa-W)?
Genotypic Deviations for thePhenotype of Fitness

Does Evolution Occur?What is:
p(WAA-W)+ q(WAa-W)?
This is the Average Excess of the A Allelefor the Phenotype of Fitness

Does Evolution Occur?
p = paA/W

Does Evolution Occur?
p = paA/W
Natural Selection is An Evolutionary ForceWhenever p ≠ 0 or p ≠ 1 (that is, there isGenetic variation) and when aA ≠ 0 (that is,When there is heritable variation in thePhenotype of fitness).

To Understand Natural Selection
THINK LIKE A GAMETE!

Sickle Cell Anemia In Africa
An Example of Natural Selection

The Sickle Cell Mutation

Infection of a Red Blood Cell By a Malarial
Parasite• Sickle-Cells Are Filtered Out
Preferentially by the Spleen• Malaria Infected Cells Are Often Filtered
Out Because of Sickling Before the Parasite Can Complete Its Life Cycle
• The Sickle Cell Allele is Therefore an Autosomal, Dominant Allele for Malarial Resistance.

The SickleCell Anemia Phenotype

Most Deaths Due to Sickle Cell Anemiaand Due to Malaria Occur Before
Adulthood. Viability Is The Phenotypeof Living To Adulthood
• In a non-Malarial Environment, The S Allele is a Recessive Allele For Viability Because Only the Homozygotes Get Sickle Cell Anemia.
• In a Malarial Environment, The S Allele is an Overdominant Allele For Viability Because Only the Heterozygotes Are Resistant to Malaria And Do Not Get Sickle Cell Anemia.
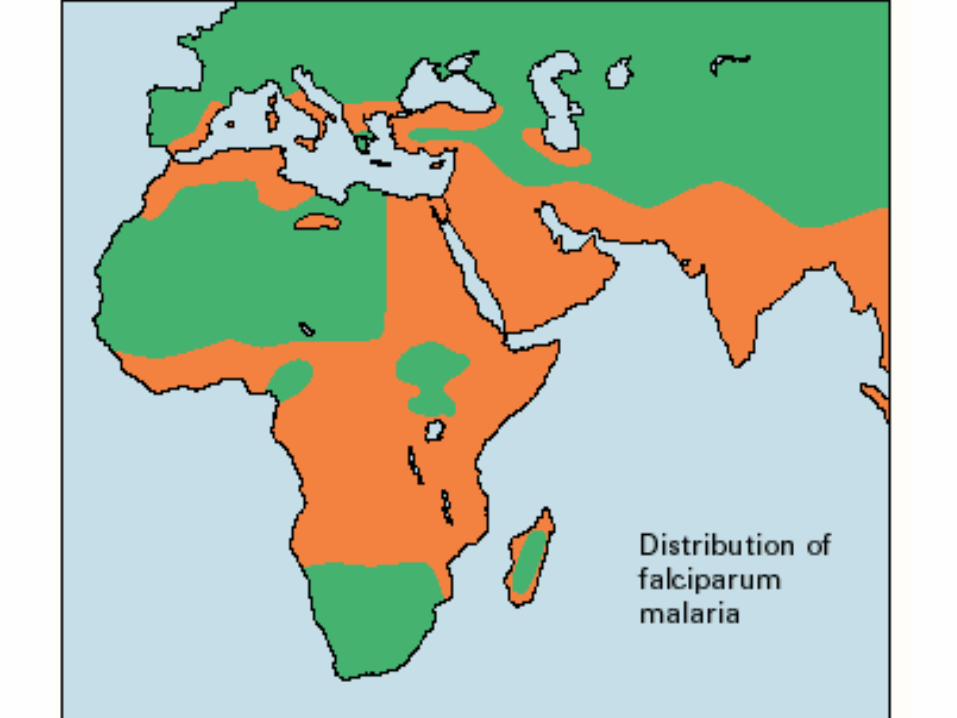
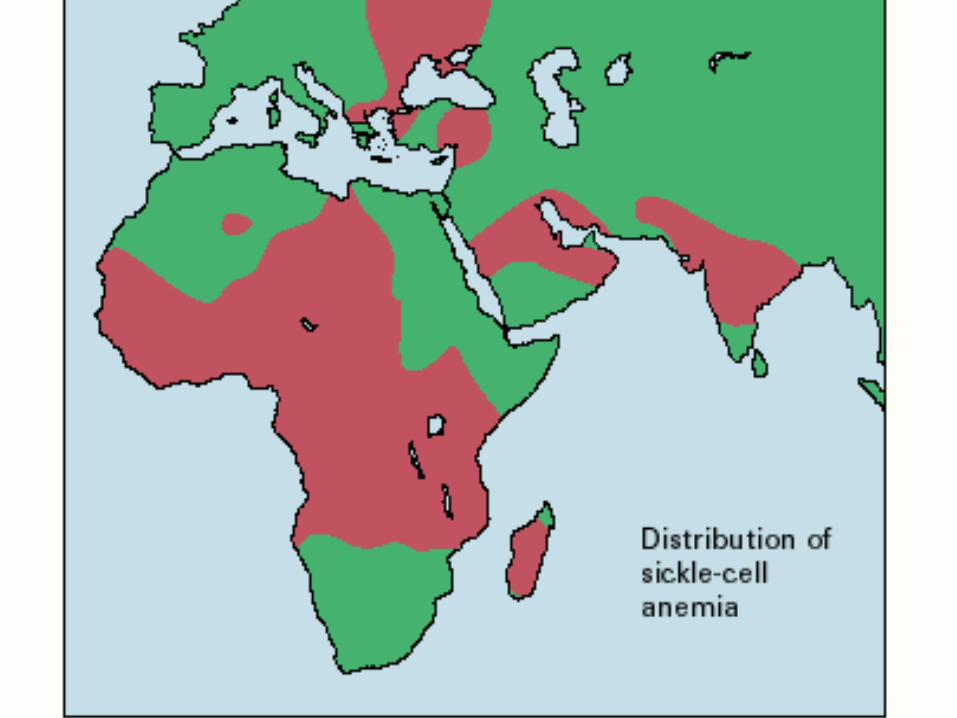

Two Complications to This Simple Story in Africa:
1. Epidemic Malaria is Recent to Most of Wet, Tropical Africa and the Process of Adaptation to Malaria in Africa Is Still Not in Equilibrium.
2. There is a Third Allele, Hemoglobin C, Involved in the Adaptation to Malaria in Africa.
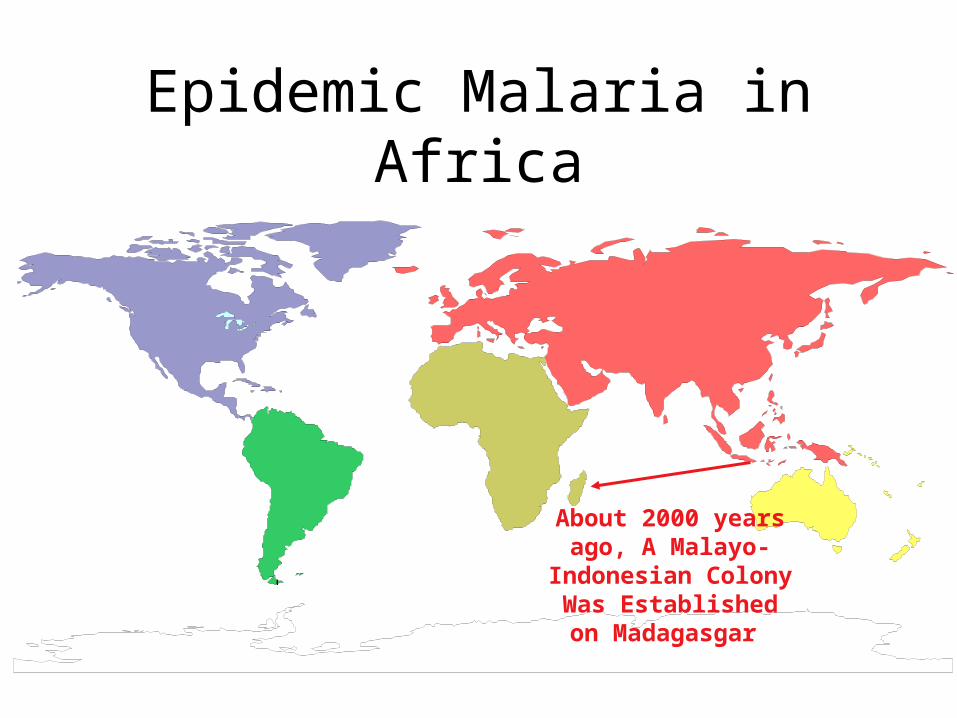
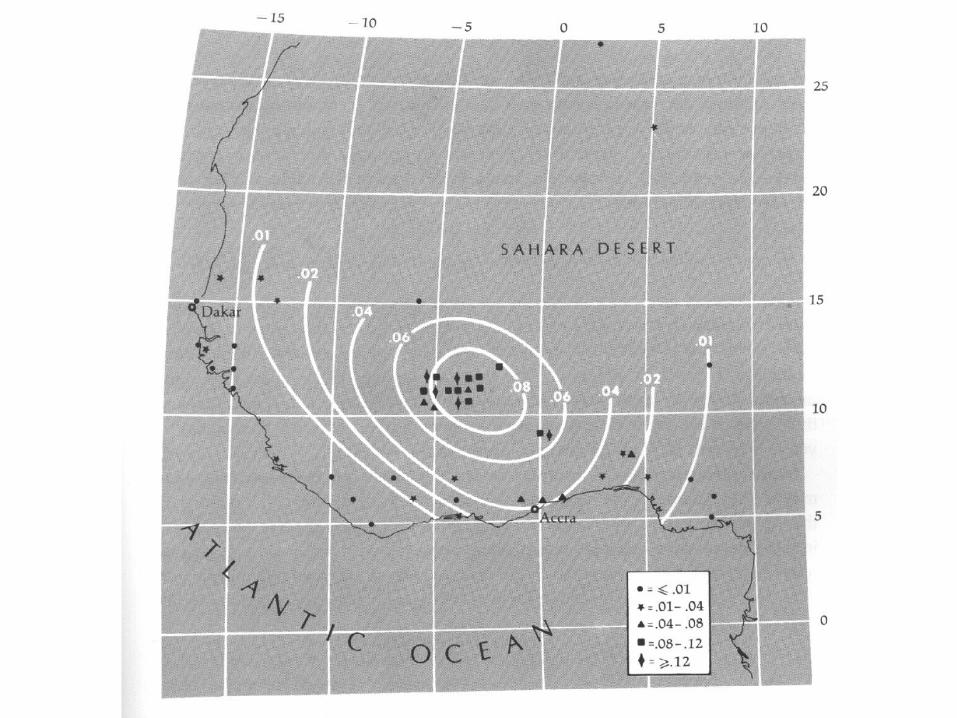
Epidemic Malaria in Africa
MADAGASCAR
ICELAND
About 2000 years ago, A Malayo-Indonesian
Colony Was Established on Madagasgar

Epidemic Malaria in Africa
This Colony Introduced The
Malaysian Agricultural
Complex into This Region

Epidemic Malaria in Africa
This Agricultural Complex Was
Taken Up By Bantu-Speaking Peoples,
Followed by A Large Expansion of the Bantu In Africa About 1500 years
Ago.

The Malaysian Agricultural Complex In Africa
• Is associated with slash-and-burn agriculture: Provides habitat and breeding sites for Anopheles gambiae, the primary mosquito vector for falciparum malaria.
• Results in the high local densities of human populations that are necessary to establish and maintain malaria as a common disease.

Epidemic Malaria in Africa
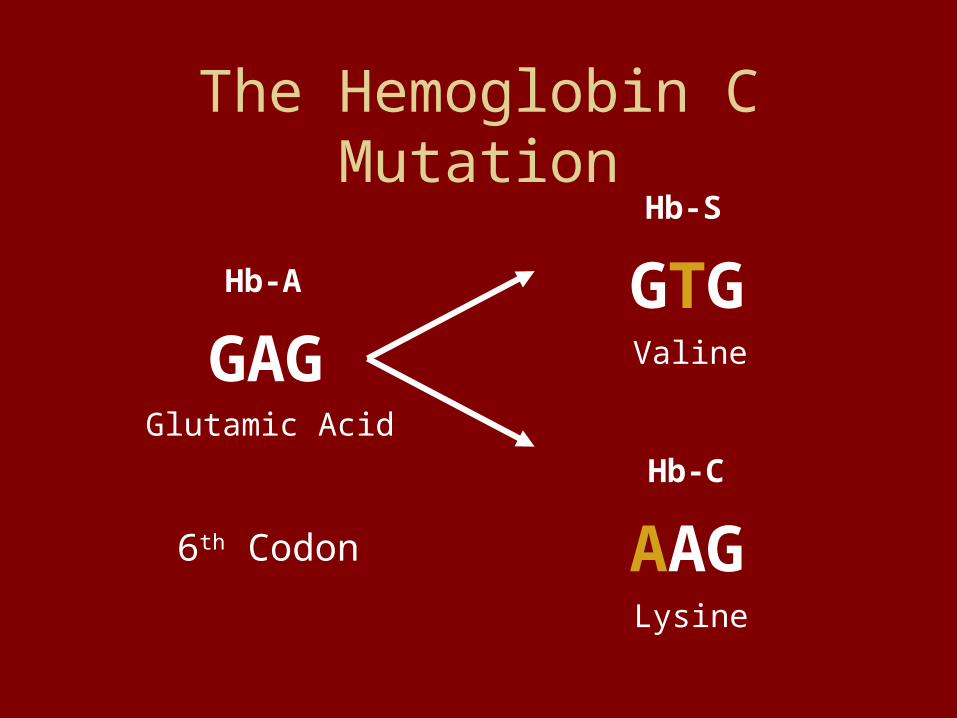
The Hemoglobin C Mutation
GAGGlutamic Acid
Hb-A GTGValine
Hb-S
AAGLysine
Hb-C
6th Codon

The Hemoglobin C Mutation
Hb-C Is A “Recessive”Allele for Malarial
Resistance
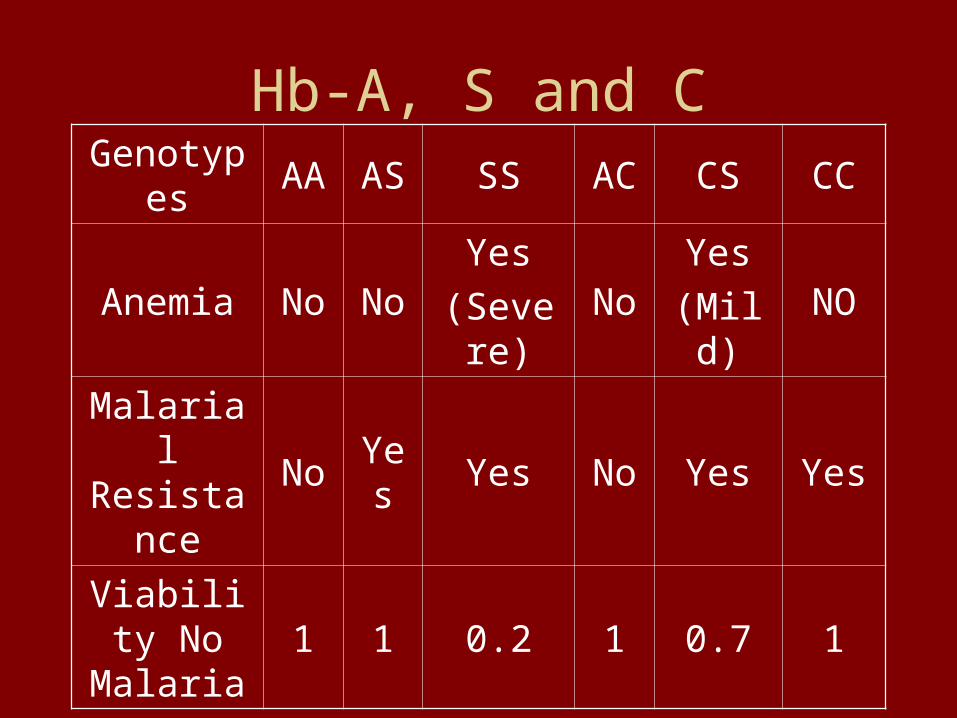
Hb-A, S and CGenotypes AA AS SS AC CS CC
Anemia No NoYes
(Severe)No
Yes
(Mild)NO
Malarial Resistance
No Yes Yes No Yes Yes
Viability No Malaria
1 1 0.2 1 0.7 1

Hb-A, S and CGenotypes AA AS SS AC CS CC
Anemia No NoYes
(Severe)No
Yes
(Mild)NO
Malarial Resistance
No Yes Yes No Yes Yes
Viability No Malaria
1 1 0.2 1 0.7 1
The A and S Alleles Define An Autosomal Recessive Genetic Disease: Selection Will Insure it is Rare But Difficult to
Eliminate in a Random Mating Population.

Hb-A, S and CGenotypes AA AS SS AC CS CC
Anemia No NoYes
(Severe)No
Yes
(Mild)NO
Malarial Resistance
No Yes Yes No Yes Yes
Viability No Malaria
1 1 0.2 1 0.7 1
The A and C Alleles Define A Set of Neutral Alleles in a Non-malarial Environment: Their Frequencies Are
Determined by Genetic Drift and Mutation.

Hb-A, S and CGenotypes AA AS SS AC CS CC
Anemia No NoYes
(Severe)No
Yes
(Mild)NO
Malarial Resistance
No Yes Yes No Yes Yes
Viability No Malaria
1 1 0.2 1 0.7 1
Viability Malaria
0.9 1 0.2 0.9 0.7 1.3
Observed Relative Viabilities In Western Tropical Africa

Hb-A, S and C• CC is the Fittest Genotype By Far• If Natural Selection is “Survival of the
Fittest”, Then Natural Selection Should Increase the Frequency of the C allele and the CC Genotype.
• Contrary to Rumor, Natural Selection is Not “Survival of the Fittest.”
• Natural Selection Is Heritable Variation in Fitness, so Think Like A Gamete: Which Gamete Has the Highest Average Excess of Fitness?

Initial Gene Pool Before Malaria
A
pA = 0.99
pS=.005 pC=.005

Initial Ave. Fitness After Transition to Malaysian Agricultural Complex
A
pA = 0.99
pS=.005 pC=.005
Under Random Mating, theMean Phenotype = W = 0.901

Initial Phenotypes After Transition to Malaysian Agricultural Complex
Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
Genotypic Deviation (W = 0.901)
-.001 .099 -.701 -.001 -.201 .399
A
pA = 0.99
pS=.005 pC=.005

Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
Genotypic Deviation (W = 0.901)
-.001 .099 -.701 -.001 -.201 .399
Initial Phenotypes After Transition to Malaysian Agricultural Complex
aA = -0.0005aS = 0.0935aC = 0.0000

Initial Phenotypes After Transition to Malaysian Agricultural Complex
The Initial AdaptiveResponse To A MalarialEnvironment MediatedBy Natural Selection IsTo Decrease A, IncreaseS, and Leave C The Same px = px(ax)/W)
aA = -0.0005aS = 0.0935aC = 0.0000

Gene Pool After Several Generations of Selection Under
A Malarial EnvironmentA
pA = 0.95S
pC = 0.005
W = 0.907pS = 0.045

Gene Pool After Several Generations of Selection Under
A Malarial Environment pC=.005
A
pA = 0.95S
.045
Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
Genotypic Deviation (W = 0.907)
-.007 .093 -.707 -.007 -.207 .393

Gene Pool After Several Generations of Selection Under
A Malarial EnvironmentAfter the Initial AdaptiveResponse To A MalarialEnvironment, NaturalSelection Continues to Decrease A, Increase S, but Now It Also Decreases C Because aC= -0.014.
aA = -0.003aS = 0.055aC = -0.014

As pS increases in frequency, W increases and theseGenotypic Deviations Become Increasingly Negative.Therefore, Natural Selection Eliminates the C Allele.
Gene Pool After Several Generations of Selection Under
A Malarial EnvironmentA
pA 1-pS
S
pS
pC 0
Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3

A Selective Equilibrium Will Only Occur When p = 0 Under Natural Selection For All Alleles.
Genotypes AA AS SS
Viability Malaria 0.9 1 0.2
aA = (1-pS)(0.9-W)+pS(1-W) = 0 = aS = (1-pS)(1-W)+pS(0.2-W)
A
pA = 1-pS
S
pS

A Selective Equilibrium Will Only Occur When p = 0 Under Natural Selection For All Alleles.
A
pA = 1-pS
S
pS
aA = (1-pS)(0.9-W)+pS(1-W) = aS = (1-pS)(1-W)+pS(0.2-W) (1-pS)(0.9)+pS(1) = (1-pS)(1)+pS(0.2)
0.9+0.1pS = 1-0.8pS
0.9pS = 0.1pS = 0.1/0.9 = 0.11
So At Equilibrium, pS = 0.11 and pA=0.89

The Equilibrium Allele Frequencies Are Maintained By Natural Selection, Resulting in a
Balanced Polymorphism A
pA = 0.89
S
pS=0.11
The Balance Occurs Because When pS < 0.11, aS > 0(malarial resistance dominates the average excess)
And When pS > 0.11, aS < 0(anemia dominates the average excess)

The EquilibriumA
pA = 0.89
S
pS=0.11
At Equilibrium, There is Genotypic Variation in Fitness (Broad-Sense Heritability), but No
Heritability (Average Excesses = 0).
AA
0.79
AS
0.20
SS
0.01
WAA = 0.9 WAS = 1WSS
=0.2

Adaptation By Natural Selection Depends Upon History:
Which Mutations Are Present and Their Frequencies. The
course of adaptation is always constrained by the available
genetic variation and proceeds until there is no heritability of
fitness.

Two Possible Responses to Malaria
A
pA
pS 0 pC 0
A
pA = 0.89
S
pS=.11
C
pC = 1
1. The Fittest Genotype is Eliminated. 1. The Fittest Genotype is Fixed.
2. Average Fitness goes from .9 to .91. 2. Average Fitness goes from .9 to 1.3.
3. 20% of the individuals have a relative viability of 1 and 80% have either anemia or malarial susceptibility.
3. 100% of the individuals have a relative viability of 1.3 and none have anemia nor malarial susceptibility.

Two Possible Responses to Malaria
A
pA
pS 0 pC 0
A
pA = 0.89
S
pS=.11
C
pC = 1
1. The Fittest Genotype is Eliminated. 1. The Fittest Genotype is Fixed.
2. Average Fitness goes from .9 to .91. 2. Average Fitness goes from .9 to 1.3.
3. 20% of the individuals have a relative viability of 1 and 80% have either anemia or malarial susceptibility.
3. 100% of the individuals have a relative viability of 1.3 and none have anemia nor malarial susceptibility.
With One Exception

Hb-A, S and CGenotypes AA AS SS AC CS CC
Viability No Malaria
1 1 0.2 1 0.7 1
C is a neutral allele relative to A, so
sometimes the C allele will drift to high
frequencies relative to the A allele.
S is a recessive, deleterious allele relative to A, so
natural selection in the pre-Malarial environment will
keep it rare (no h2).

Suppose There Was A Deme With This Gene Pool Before The Malaysian Agricultural Complex
A
pA = 0.95C
.045
pS=.005
Such a gene pool is likely to evolve in the pre-malarial environment because of the neutrality of A and C
relative to each other.

Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
Genotypic Deviation (W = 0.902)
-.002 .098 -.702 -.002 -.202 .398
Initial Phenotypes After Transition to Malaysian Agricultural Complex
aA = -0.001aS = 0.081aC = 0.015
The Initial Adaptive Response To A Malarial Environment Is To Increase The Frequency of The S and C Alleles.

Gene Pool After Several Generations of Selection Under
A Malarial EnvironmentA
pA = 0.78S
.05
C
0.17
Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
Genotypic Deviation (W = 0.914)
-.01 .09 -.71 -.01 -.21 .39

Gene Pool After Several Generations of Selection Under
A Malarial EnvironmentAfter the Initial AdaptiveResponse To A MalarialEnvironment, NaturalSelection Continues to Decrease A, Increase C, but Now It Also Decreases S Because aS= -0.005.
aA = -0.009aS = -0.005aC = 0.044
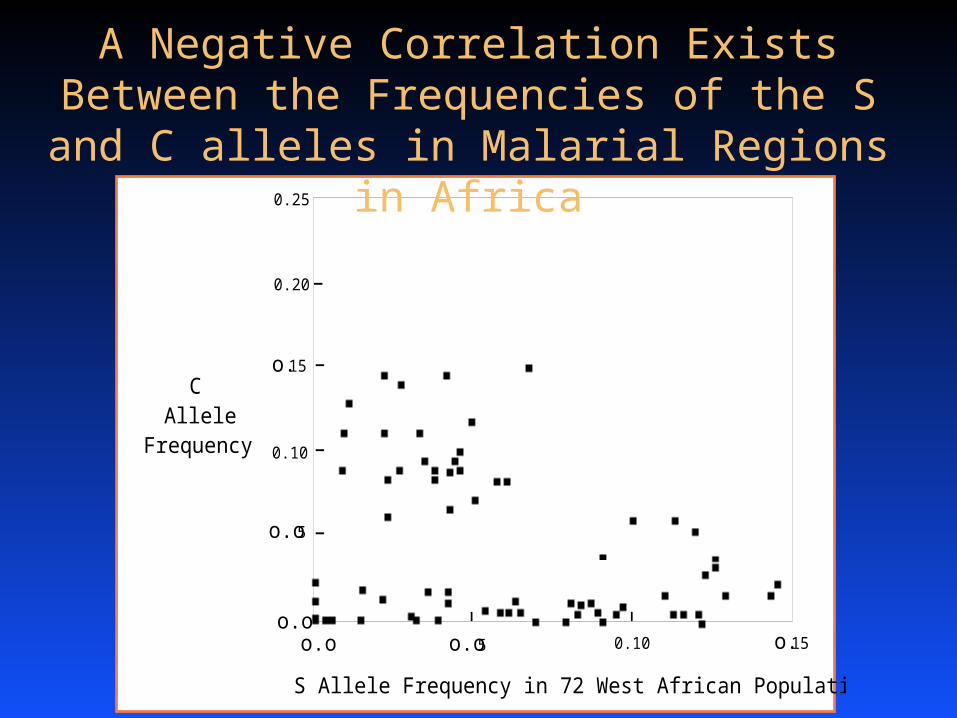
o.o
o.o5
o.15
0.20
0.25
0.10
o.o o.o5 0.10 o.15
S Allele Frequency in 72 West African Populations
CAllele
Frequency
A Negative Correlation Exists Between the Frequencies of the S and C alleles in Malarial
Regions in Africa

Even uniform selective pressures produce divergent adaptive responses because selection
operates upon variation whose creation and initial frequencies are profoundly influenced by
random factors such as mutation and drift.

Although adaptation is often portrayed as “optimizing”
individual or population fitness, only gametic fitness is optimized via natural selection. Individuals or demes with the highest fitness are not necessarily favored and can be actively selected against.

There are many other ways in which human populations have adapted to malaria; e.g. G-6-PD Deficiency:
Plasmodium oxidizes RBC NADPH from the Pentose Phosphate pathway for its metabolism. This results in a deficiency of RBC GSH, most severe in G6PD deficient individuals, leading to peroxide-induced hemolysis which curtails the development of Plasmodium.

There are many other ways in which human populations have adapted to malaria; e.g.
Thalassemia:

Adaptation generally involves many loci with different biochemical, cellular or
developmental functions.
Therefore, we also need to model natural selection as a polygenic
process.

The Fundamental Theorem of Natural Selection
• Fisher was one of the first to model natural selection as a polygenic process.
• Although there are many aspects of his models, the most important results are found in what he termed the “fundamental theorem of natural selection.”

x = phenotypic value of some trait for an individual in a population
f(x) = the probability distribution that describes the frequencies of x in the population.
The mean phenotype is then:
€
= xf(x)dxx∫
w(x) = the fitness of those individuals sharing a common phenotypic value x.
The mean or average fitness of the population is:
€
w = w(x)f(x)dxx∫

w(x)f(x) does not in general define a probability distribution, but w(x)f(x)/ does integrate to one and defines the probability distribution of the selected individuals.
Hence, the mean phenotype of the selected individuals is:
Let h2 = the heritability of the trait. The response to selection is given by R=h2S where S=(s–), R = (o–), and o is the phenotypic mean of the offspring of the selected parents.
€
s =xw(x)f(x)dx
x∫w
€
w

When x = w, w(w) = w by definition, and = .
€
w
€
s =w × wf(w)dw
w∫w
=w2f(w)dw
w∫w
€
s =w2 − w 2( ) + w 2[ ]f(w)dw
w∫w
=w2 − w 2( )f(w)dw + w 2 f(w)dw
w∫w∫w
€
s =w − w ( )
2f(w)dw + w 2
w∫w
=σ 2 + w 2
w
€
S = μ s − μ =σ 2 + w 2
w − w =
σ 2 + w 2 − w 2
w =
σ 2
w

When x = w, the response to selection, R, is . Hence,
€
w
€
R = h2S
Δw =σ a
2
σ 2
⎛
⎝ ⎜
⎞
⎠ ⎟σ 2
w
⎛
⎝ ⎜
⎞
⎠ ⎟
Δw =σ a
2
w
Fundamental Theorem of Natural Selection

Some Implications of FFTNS
• FIRST, natural selection can only operate when there is genetic variation associated with phenotypic variation for fitness in the population.
• SECOND, the only fitness effects that influence the response to natural selection are those transmissible through a gamete.
• THIRD, the adaptive outcome represents an interaction of fitness variation with population structure.

Some Implications of FFTNS
• FOURTH, selective equilibria can only occur when all the average excesses and all the average effects are zero; that is, when all gamete’s have the same average fitness impact. Evolution due to natural selection stops only when there is no heritability for fitness. This in turn means that at a selective equilibrium there is no correlation between the fitness of parents and the fitness of their offspring even when there is genetic variance in the phenotype of fitness.

The EquilibriumA
pA = 0.89
S
pS=0.11
At Equilibrium, There is Genotypic Variation in Fitness (Broad-Sense Heritability), but No
Heritability (Average Effects = 0).
AA
0.79
AS
0.20
SS
0.01
WAA = 0.9 WAS = 1WSS
=0.2

Some Implications of FFTNS
• FIFTH, natural selection acts to increase the average fitness of a population on a per generational basis. Because the additive genetic variation must be greater than or equal to zero, w ≥ 0 under natural selection. Because average fitness can only increase or stay the same under natural selection, the selective equilibria discussed under point four must always correspond to an average fitness local optimum.

Wright’s Concept of An Adaptive Surface or Landscape
AA AS SS
WAA = 0.9 WAS = 1 WSS=0.2
0.0 0.2 0.4 0.6 0.8 1.0p
0.2
0.4
0.6
0.8
1.0Average Fitness
Frequency of S Frequency of S
0.00 0.05 0.10 0.15 0.20p
0.89
0.90
0.91
0.92Average Fitness

Some Implications of FFTNS
• SIXTH, natural selection only takes populations to local adaptive solutions and not necessarily to the adaptive state with the highest average fitness, and indeed may operate to prevent an adaptive state with higher average fitness from evolving.

0.0 0.2 0.4 0.6 0.8 1.0p
0.7
0.8
0.9
1.0Average Fitness
Wright’s Concept of An Adaptive Surface or Landscape
AA Aa aa
WAA = 1 WAa = 0.5 Waa=0.9
Frequency of A
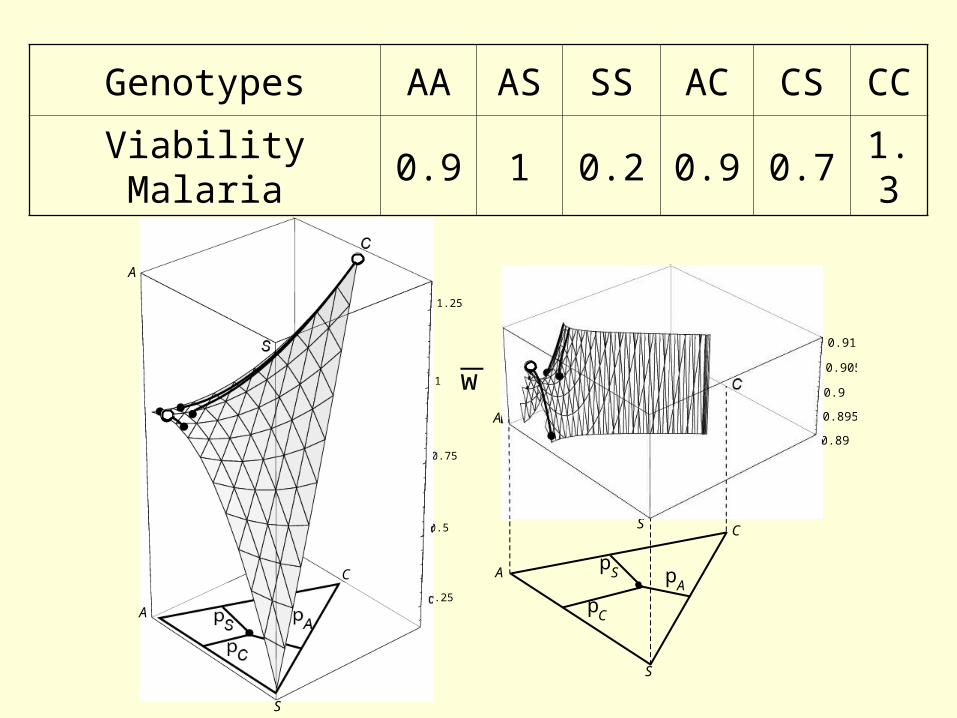
Genotypes AA AS SS AC CS CC
Viability Malaria 0.9 1 0.2 0.9 0.7 1.3
w
A
S
C
A
C
pC
pS pA
S
pC
pS pA
0.89
0.895
0.9
0.905
0.91
A
C
S
0.25
0.5
0.75
1
1.25
A
S
C

Some Implications of FFTNS
• SEVENTH, natural selection generally does not optimize, even in a local sense, any individual trait other than fitness itself, even if the trait contributes to fitness in a positive fashion.

Given a selective equilibrium ( = 0) at a local peak, let feq(x) = the phenotypic distribution of the trait at equilibrium. Then, the average fitness and average trait at equilibrium is:
€
w
€
w eq = w(x)feq(x)dxx∫
€
x eq = xfeq(x)dxx∫
is the “optimal” value of trait X only if
€
x eq
€
w eq = w(xeq )
€
w(x) ≈ w(xeq ) + w'(x eq )(x − x eq ) + 12 w' '(x eq )(x − x eq )2
Use Taylor’s theorem to expand w(x) around :
€
x eq

Take the average value of both sides of the Taylor’s Series approximation by integrating across the equilibrium probability distribution of the trait:
€
w eq = w(x eq ) feq(x)dxx∫ + ′ w (x eq ) x − x eq( )feq(x)dx + 1
2x∫ ′ ′ w (x eq ) x − x eq( )2feq(x)dx
x∫
€
w eq = w(xeq )+ 12 ′ ′ w (xeq )σ eq
2 (x)
€
x eq is an optimal value of trait X that maximizes w(x) when:
1. the trait has no phenotypic variance at equilibrium [2
eq(x) = 0], or 2. the trait is related to fitness in a strictly linear fashion at equilibrium [w”( ) = 0]
€
x eq

Some Implications of FFTNS
• EIGHTH, the process of adaptation can result in the evolution of some seemingly non-adaptive traits. In general, many traits contribute to fitness, not just one. Consider the case in which two traits, say X and Y, contribute to fitness such that w(x,y) is the fitness of those individuals with trait values x and y for the two traits respectively. Then, the two-dimensional requirement for optimality of both traits is:
€
∂2w(xeq ,yeq )
∂x 2σ eq
2 (x) + 2∂ 2w(xeq , yeq )
∂x∂yCoveq (x,y) +
∂ 2w(xeq , yeq )
∂y 2σ eq
2 (y) = 0

E.g., many human populations have adapted to malaria by increasing the frequency of the trait of hemolytic anemia. Here, natural selection favors the increase of a highly deleterious trait.
Such cases are common because of pleiotropy, and indeed most of the people who die or suffer from genetic disease do so because natural selection favored the genes despite one or more pleiotropic deleterious traits.
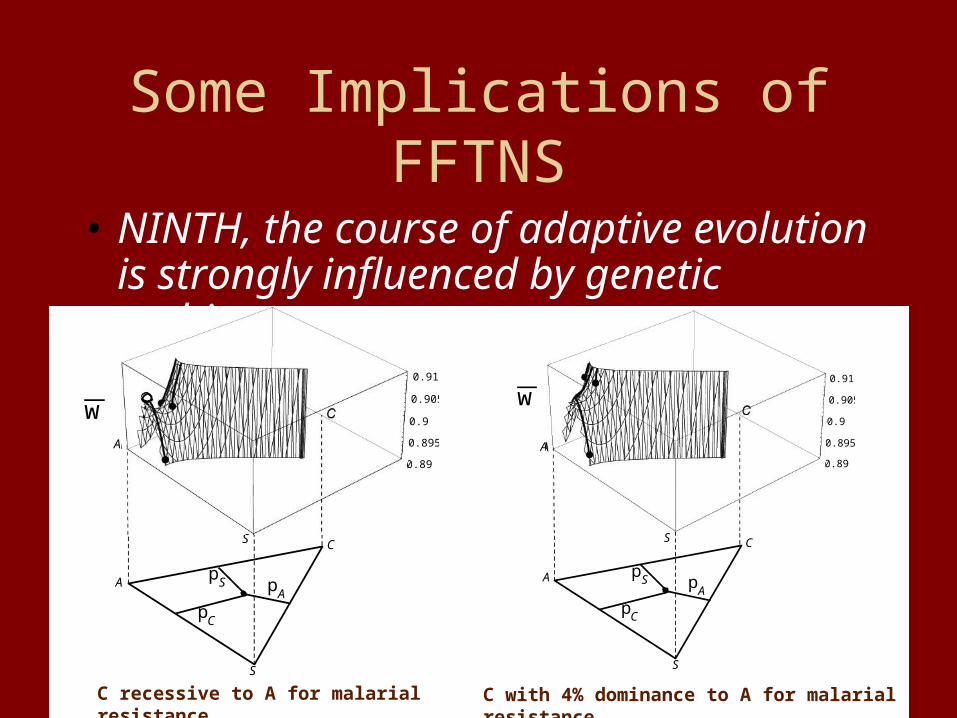
Some Implications of FFTNS
• NINTH, the course of adaptive evolution is strongly influenced by genetic architecture.
wA
C
pC
pS pA
S
0.89
0.895
0.9
0.905
0.91
A
C
S
C recessive to A for malarial resistance
w
A
C
pC
pS pA
S
0.89
0.895
0.9
0.905
0.91
A
C
S
C with 4% dominance to A for malarial resistance

Common theme?
p = paA/W
€
w =σ a
2
w
The Course of Adaptive Evolution Is Determined By the Phenotypic Effects
Assigned to GAMETES, not individuals!







