
Molecular properties of plasmids
Presented by:
Mahboube shahrabi,MSc student of medical biotechnology,IUMS

The size and copy number of plasmids
Bacterial plasmids in general exist within the cell as circular DNA molecules with a very compact conformation
Many of them are small molecules, just a few kilobases
But in some cases:
Like, members of the genus Pseudomonas, plasmids up to several hundred kilobases are common

2 types of Ecoli plasmids
1-ColE1
relatively small (usually less than 10 kb)
are present in multiple copies within the cell
Their replication is not linked to the processes of chromosomal replication and cell division
Although there are some controls on plasmid replication

2-F plasmid
larger (typically greater than 30 kb; F itself is about 100 kb)
Are present in only one or two copies per cell
their replication is controlled in essentially the same manner as that of the chromosome
These large plasmids are able to promote their own transfer by conjugation

High copy number imposes a size constraint
Replication of a plasmid imposes a metabolic burden that is related to the size and copy number of the plasmid
It is logical that high copy number plasmids will also be small
for example:
ColE1, is 6.4 kb in size. If there are 30 copies per cell, this represents about 4 per cent of the total DNA of the cell
The F plasmid is 100 kb, if it were to be present at a similar copy number, would add nearly 70 per cent to the total DNA content
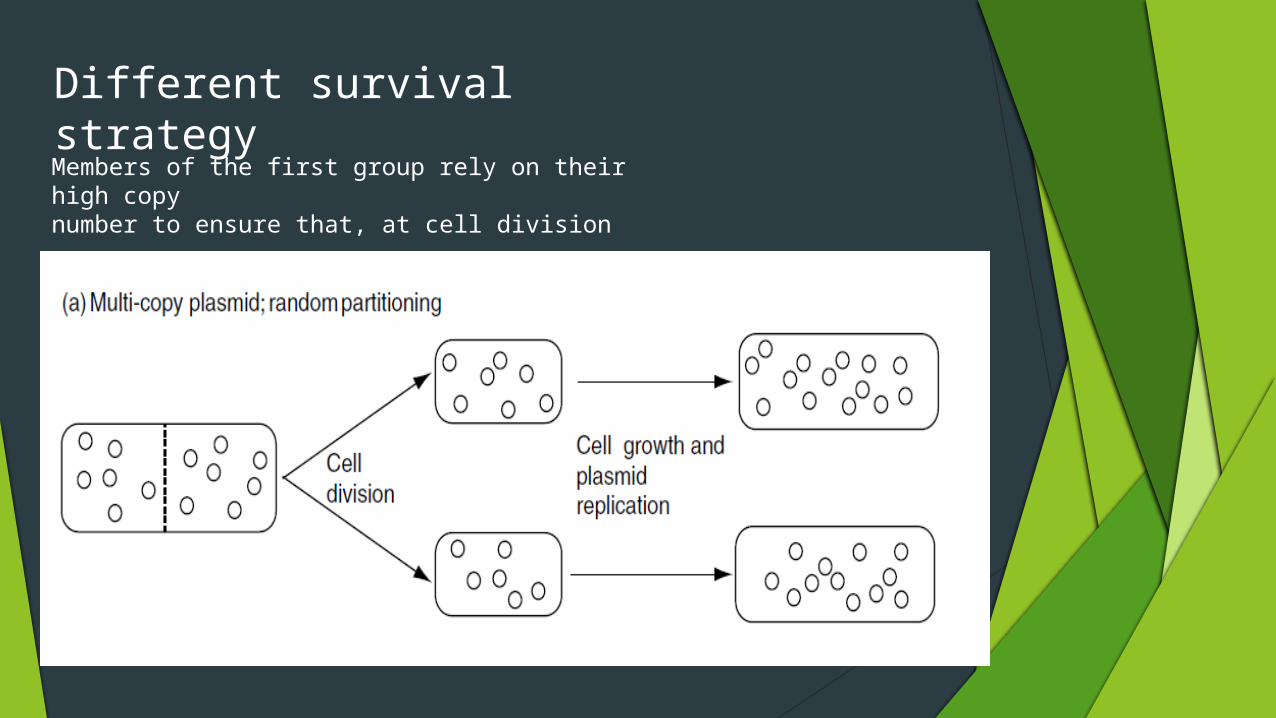
Different survival strategy
Members of the first group rely on their high copynumber to ensure that, at cell division
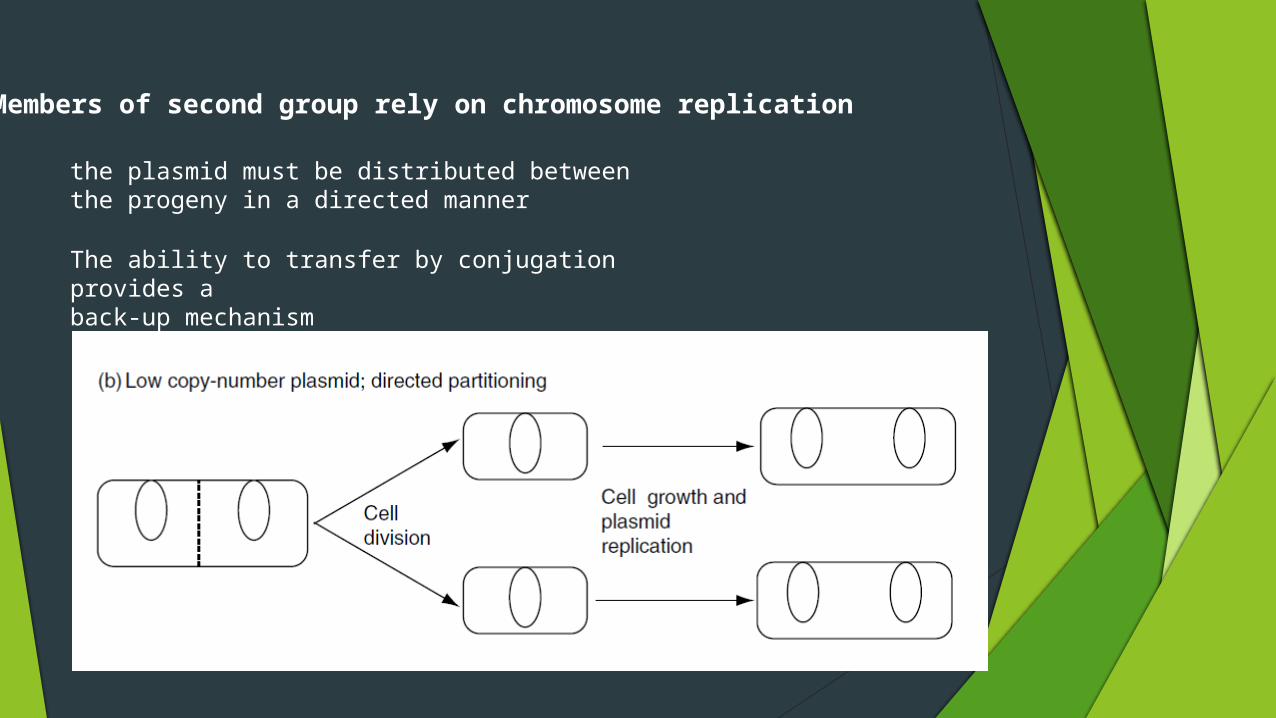
Members of second group rely on chromosome replication
the plasmid must be distributed betweenthe progeny in a directed manner
The ability to transfer by conjugation provides aback-up mechanism

Plasmid replication and control
Many plasmids are replicated as doublestranded circular molecules
The over all picture of their replication is similar to chromosome replication
Replication starts at a fixed point known as oriV
proceeds from this point, either in one direction or in both directions simultaneously
However there are some aspects of replication that differ from that of the chromosome, especially for the multicopy plasmids

Replication of ColE1
Genetic map of the plasmid ColE1
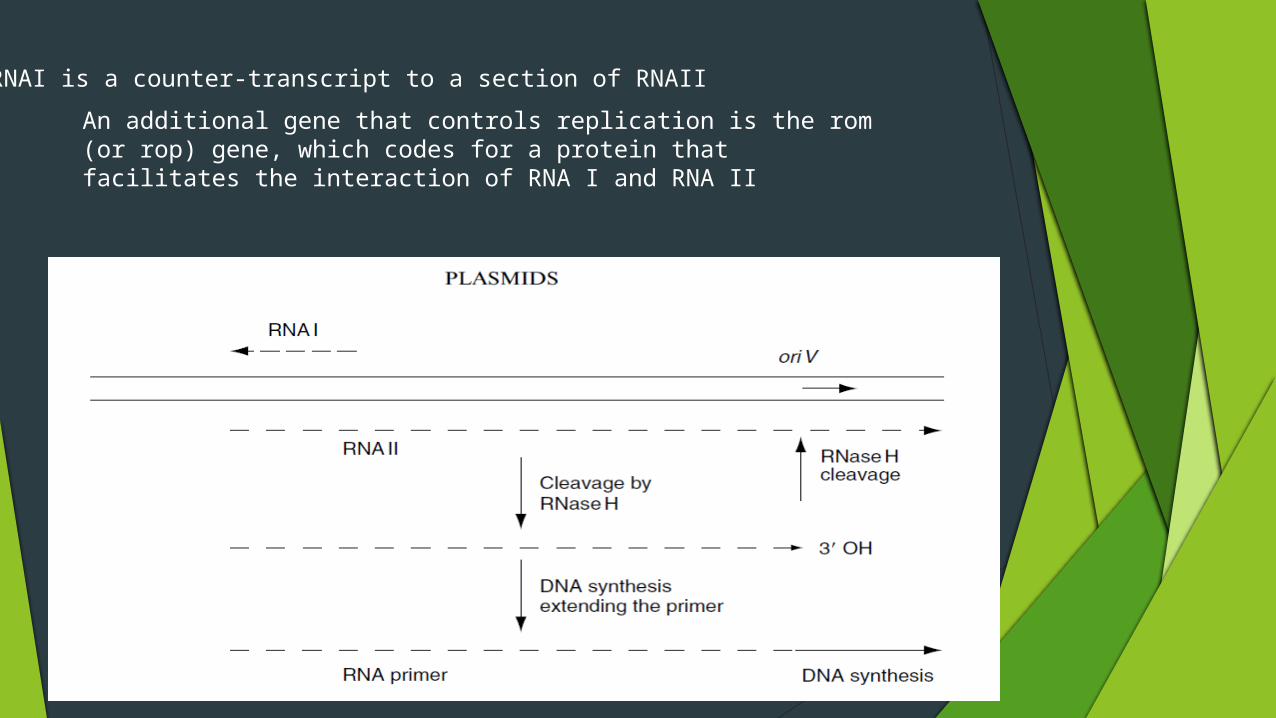
An additional gene that controls replication is the rom (or rop) gene, which codes for a protein that facilitates the interaction of RNA I and RNA II
RNAI is a counter-transcript to a section of RNAII

Replication of R100
R100 is a low copy number
conjugative
resistance plasmid
contains about 89 kb of DNA
resistance to four different antibiotics (tetracycline, chloramphenicol, streptomycin and sulphonamides), as well as to mercury salts

Genetic map of the conjugative E. coli plasmid R100

a gene known as repA (adjacent to oriV) is required for the initiation of replication from oriV
Plasmid copy number is controlled by two genes that regulate the production of the RepA protein
1-cop A
copA, regulates expression of RepA
2-cop B
copB, codes for a protein that represses transcription of the repA gene

The RepA protein is needed for initiation of replication. Transcription of repA is repressed by CopB and translation of the repA mRNA is inhibited by the antisense copA RNA
Replication control of the plasmid R100

par locus
Plasmids like R100 also contain a region known as the par locus (for partitioning) that is necessary for accurate partitioning of plasmid copies at cell division
for ensuring accurate partitioning:
all low copy number plasmids have a par sequence
While,
plasmids such as ColE1 rely primarily on high copy number to ensure that each daughter cell receives a copy of the plasmid

Control of plasmid replication by DNA repeats (iterons)
Stable maintenance and regulation of some plasmids like,F needs more a much tighter control of replication that is directly linked to the number of copies in the cell in addition to, control of the expression of RepA
Iterons
repeated DNA sequences, 17–22 bp long in the replication initiation regions
The RepA protein binds to these iteron sequences
When there is more than one copy of the plasmid, the RepA protein can bind to iterons on both copies, coupling them togetherCoupling model

Plasmids coupled by RepA bound to iteronsFurther replication is prevented
Coupling model for the control of iteron-containing plasmids

Plasmid replication via single-stranded forms
Many plasmids, especially in Grampositive bacteria, replicate via a single-stranded intermediate
A specific site (the plus origin) on the plus strand of the plasmid is first cut by a plasmid-encoded protein (Rep)
The nicked DNA provides a site for initiation of DNA synthesis using host enzymes which displaces the old plus strand
The Rep protein makes another nick to release the old plus strand and ligates the ends of this molecule to produce an intact single-stranded circular structure
then converted to the doublestranded form
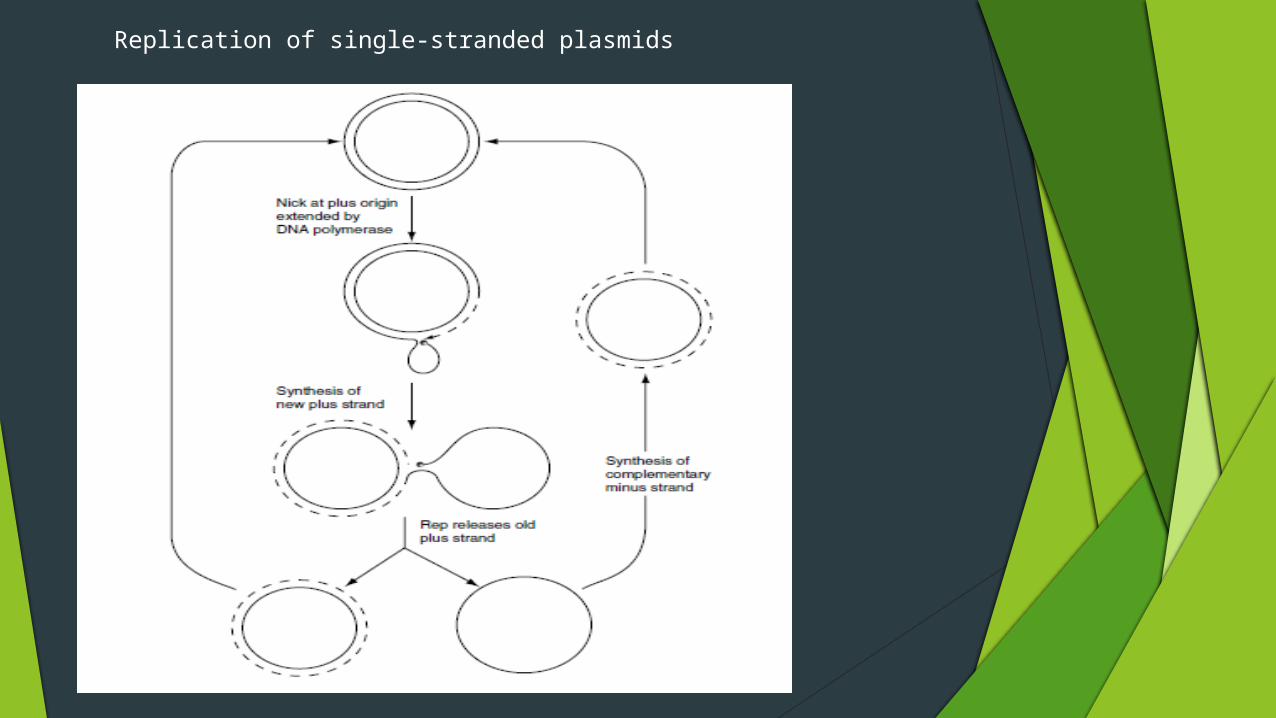
Replication of single-stranded plasmids

Replication of linear plasmids
Linear DNA plasmids have been characterized in Borrelia and Streptomyces
And a problem!
Incomplete replication of linear DNA. Normal modes of replication are unable to replicate the ends of a linear molecule

The solution: In borrelia the ends of the two strands are joined together in a covalently closed
hairpin structure
Bidirectional replication initiating at a central origin of replication (oriC) would lead to a dimeric double-stranded circular molecule in which two copies of the genome are linked by copies of the hairpin loop sequence
This intermediate structure would then be processed by cutting and rejoining the DNA strands at each end to reform the covalently closed hairpin loops.
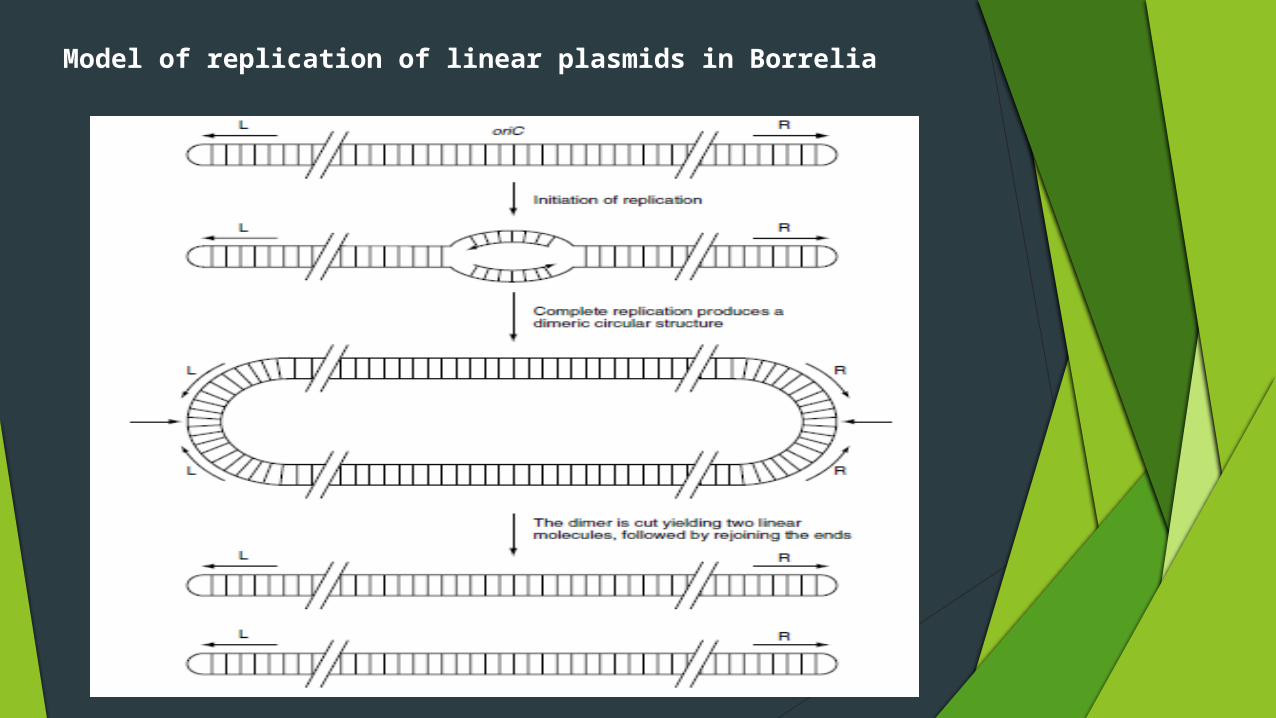
Model of replication of linear plasmids in Borrelia

But in Streptomyces:
A key feature is the presence of a protein (terminal protein, TP), covalently attached to the 50 ends of the DNA
This protein acts as a primer for DNA synthesis, allowing replication of the ends of the linear DNA

Thanks for your attention







