
UNIVERSIDADE DE LISBOA
FACULDADE DE FARMÁCIA
ROLE OF APOPTOSIS FACTORS
DURING MOUSE NEURAL STEM CELL
DIFFERENTIATION
Márcia Maria de Almeida Aranha
DOUTORAMENTO EM FARMÁCIA
BIOQUÍMICA
2010


UNIVERSIDADE DE LISBOA
FACULDADE DE FARMÁCIA
ROLE OF APOPTOSIS FACTORS
DURING MOUSE NEURAL STEM CELL
DIFFERENTIATION
Márcia Maria de Almeida Aranha
Tese de Doutoramento em Farmácia (Bioquímica), apresentada à
Universidade de Lisboa através da Faculdade de Farmácia
Research advisor:
Prof. Doutora Cecília M. P. Rodrigues
Lisboa
2010

The studies presented in this thesis were performed at the Research Institute for
Medicines and Phamaceutical Sciences (iMed.UL), Faculty of Phamacy, University of
Lisbon under the supervision of Professor Cecília M. P. Rodrigues, and at the
Departments of Medicine, Genetics, Cell Biology, and Development, and the Department
of Neurosurgery, Stem Cell Institute, University of Minnesota Medical School,
Minneapolis, Minnesota, USA, in collaboration with Professor Clifford Steer and
Professor Walter C. Low.
Márcia Maria de Almeida Aranha was the recipient of a Ph.D. fellowship
(SFRH/BD/28429/2006) from Fundação para a Ciência e Tecnologia (FCT), Lisbon,
Portugal. This work was supported by grants PTDC/BIA-BCM/67922/2006 and
PTDC/SAU-FCF/67912/2006 from FCT and FEDER.
De acordo com o disposto no ponto 1 do artigo nº41 do Regulamento de Estudos
Pós-Graduados da Universidade de Lisboa, deliberação nº 93/2006, publicada em Diário
da República – II Série nº 153 – 5 de Julho de 2003, o Autor desta dissertação declara que
participou na concepção e execução do trabalho experimental, interpretação dos
resultados obtidos e redacção dos manuscritos.

Aos meus queridos pais


Resumo


Resumo
ix
A utilização de células estaminais para tratamento de doenças neurológicas é,
actualmente, objecto de intensa investigação. Para que seja possível desenvolver terapias
regenerativas, a partir destas células, é fundamental compreender os seus processos
biológicos básicos, nomeadamente o seu mecanismo de diferenciação.
Estudos recentes sugerem que alguns factores específicos do processo apoptótico,
como as caspases, a proteína p53, a família Bcl-2 e, mais recentemente, os microRNAs
(miRNAs ou miRs), possam estar envolvidos na diferenciação celular. Assim,
formulámos a hipótese de que algumas moléculas associadas à apoptose desempenhem
um papel importante na diferenciação de células estaminais neurais.
Avaliou-se, inicialmente, o envolvimento da p53 e caspase-3 na diferenciação de
células estaminais neurais. Observámos que, tanto a clivagem da caspase-3, como a
fosforilação da proteína p53 e a sua capacidade de ligação ao DNA aumentaram
significativamente ao longo da diferenciação. A inibição das caspases e o silenciamento
de p53 retardaram, de forma diferencial, a neurogénese e a gliogénese. Os nossos
resultados sugerem que a p53 e a caspase-3 desempenham um importante papel na
diferenciação neural, que parece envolver a modulação da via do FOXO3A/Id1.
Seguidamente quisemos determinar se os miRNAs associados à apoptose estariam,
igualmente, envolvidos no processo de diferenciação de células estaminais neurais. A
determinação dos perfis de expressão dos miRNAs, juntamente com a validação dos
níveis de expressão por PCR em tempo real, mostraram que miRNAs pró-apoptóticos,
tais como o miR-16, o let-7a e o miR-34a, são modulados durante a diferenciação de
células estaminais neurais. Resultados similares foram observados na diferenciação de
células estaminais embrionárias e de células PC12 e NT2N, o que sugere o envolvimento
destes miRNAs no desenvolvimento neural dos mamíferos.
Por fim, modularam-se os níveis de expressão do miR-34a em células estaminais
neurais e concluiu-se que este miRNA regula, de forma positiva, o aparecimento de
neurónios pós-mitóticos. Experiências adicionais sugeriram que o miR-34a regula,
também, a diminuição da SIRT1 e o aumento da capacidade de ligação da p53 ao DNA.
Este trabalho clarifica o papel que algumas moléculas do processo apoptótico têm
na diferenciação de células estaminais neurais e confirma a sua importância na definição
do destino celular, proporcionando indicações úteis ao desenvolvimento de novas
estratégias terapêuticas.
Palavras chave: Apoptose – Caspase-3 - Células estaminais neurais – Diferenciação –
miRNAs – miR-34a – p53


Abstract


Abstract
xiii
Neural stem (NS) cells are under active consideration as a source of donor tissue for
neuronal cell therapy. Therefore, understanding basic processes of NS cell biology,
including cell differentiation, is essential to develop regenerative therapies from the
promise of stem cells.
Several studies have recently provided a persuasive argument that specific factors of
apoptosis pathways, such as caspases, p53, Bcl-2 family members, and microRNAs
(miRNAs or miRs) are involved in the differentiation process. Therefore, we
hypothesized that specific apoptosis-associated molecules play a critical role in NS
differentiation.
In initial studies we evaluated the involvement of p53 and caspase-3 during the
differentiation process of NS cells. Our results revealed a significant increase in caspase-
3 activation, p53 phosphorylation, and p53 DNA-binding activity during differentiation.
Pharmacologic inhibition of caspases delayed neuronal and glial differentiation, while
down-regulation of p53 affected only the neuronal lineage. Therefore, our results suggest
that p53 and caspases play a role in the differentiation of NS cells and advance the
potential modulation of FOXO3A/Id1 signaling in this cellular context.
Next, we were interested in determining whether miRNAs linked to apoptosis might
be involved in the differentiation of NS cells. miRNA expression profiles and
quantitative real time-PCR revealed that pro-apoptotic miRNAs, such as miR-16, let-7a
and miR-34a, were modulated during NS cell differentiation. Similarly, upregulation of
pro-apoptotic miRNAs in the differentiation of embryonic stem cells, PC12 and NT2N
cells implicates these specific miRNAs in mammalian neuronal development.
Finally, we further investigated the effect of miR-34a modulation in NS cells
differentiation. We reported that miR-34a positively regulated the appearance of post-
mitotic neurons of mouse NS cells. Additional experiments indicated that miR-34a-
mediated differentiation involved SIRT1 down-regulation and was associated with
increased p53-DNA binding activity. These results suggested that miR-34a was required
for proper neuronal differentiation.
In conclusion, these studies further clarify the role of apoptosis-associated
molecules in the differentiation process of NS cells. Our findings may prove useful in the
development of novel therapeutic strategies to improve long term-survival and
differentiation of transplanted stem cells.
Keywords: Apoptosis – Caspase-3 – Differentiation – miRNAs – miR-34a – Neural stem
cells – p53


xv
Acknowledgements
Começo por agradecer àquela que foi a mentora de todo o trabalho desenvolvido
nesta tese, a Professora Cecília Rodrigues. Agradeço ter acreditado em mim e apostado
no meu trabalho, ainda no início da minha formação enquanto investigadora. Agradeço
todos os ensinamentos que me transmitiu e que foram determinantes para a minha
formação e crescimento profissional. Agradeço, sobretudo, a ajuda nos momentos de
maior hesitação. Obrigada pelo acompanhamento constante no trabalho e pelas
oportunidades que me proporcionou ao longo destes anos. A sua enorme capacidade de
trabalho e disponibilidade total, aliados à sua preocupação constante com o trabalho
desenvolvido pelos seus alunos, definem-na como uma excelente orientadora.
Part of the research studies leading to this Ph.D. thesis were conducted in the
University of Minnesota, in Minneapolis, at Professor Clifford Steer’s and Professor
Walter Low’s laboratories. I would like to express my gratitude for kindly receiving me
and for all the support during my stay in the U.S.A. I am also thankful for all the critical
comments and suggestions that contributed immensely to the studies included in this
thesis. My thanks to Phil Wong, Carol Bruzzone, Dr. Betsy Kren, and everybody else I
met, and worked with at the lab, and at the University. I would like to thank Dr. Yan
Zeng, Jing Xiao and Crusoe for all the help and teaching.
Um agradecimento muito especial à Susana Solá, com quem iniciei este trabalho e
com quem partilhei as frustrações e conquistas de cada resultado obtido. Agradeço o seu
interesse e acompanhamento constantes que foram, sem dúvida, fundamentais para o
desenvolvimento deste trabalho. Acima de tudo, obrigada pela sua amizade.
Agradeço também ao Rui, à Rita, à Joana, ao Pedro, à Filipa, ao Ricardo, à
Benedita, à Daniela, ao Duarte e à Joana, meus queridíssimos amigos e colegas de
trabalho. Foi um privilégio ter trabalhado ao lado de vocês. Mesmo sem darem conta,
vocês foram determinantes na minha vida. Obrigada! Agradeço ainda à Isabel Moreira da

Acknowledgements
xvi
Silva pela atenção e generosidade, assim como às restantes pessoas fantásticas do Centro
de Patogénese Molecular com quem tive a sorte de me cruzar e partilhar bons momentos.
Obrigada pelos conselhos, pela ajuda e pelo amparo nos momentos difíceis. Obrigada
sobretudo, pela alegria e boa disposição que transmitem e que foram fundamentais ao
longo destes anos. Os momentos que passámos juntos serão recordados com muita
saudade e alegria.
Agradeço à minha amiga de todas as horas, a Gui, por estar sempre ao meu lado,
pela confiança e pelo apoio que me deu ao longo destes anos. Agradeço ainda à Verinha,
pela sua amizade e aos amigos Barroca, Ricardo, Hugo, Clara e Sara por terem partilhado
comigo o entusiasmo pela ciência. Obrigada à Maíra, Daniele, Patrícia, Paula e Giuliana,
pela amizade duradoura que se tem mantido, apesar da distância que nos separa.
Agradeço-te a ti, Tiago, por me ter trazido paz e serenidade nesta recta final.
Agradeço o apoio incondicional e a força que me transmitiu em todos os momentos.
Obrigada pelo carinho; obrigada por me fazer tão feliz!
À Ana Paula e à Mariá, minhas irmãs queridas do coração, um muito obrigada!
Agradeço cada mensagem, cada conversa, cada gesto de carinho e preocupação que
tornaram mínima a distância física que nos separa. A nossa ligação e cumplicidade
ultrapassaram barreiras e deram-me a força necessária para seguir sempre em frente.
Obrigada por terem estado sempre ao meu lado nesta jornada. Agradeço, ainda, às minhas
sobrinhas Amaya e Lila; duas estrelas que iluminam e enchem a minha vida de alegria e
boa disposição. Obrigada pelo carinho, tão sincero e genuíno, que transmitem em cada
telefonema e de cada vez que nos encontramos.
Por fim, agradeço aos meus pais, a quem dedico esta tese, por tudo o que me
transmitiram ao longos destes anos e que fizeram de mim a pessoa que sou hoje. Obrigada
pela vossa dedicação e por tudo o que me proporcionaram até agora. Sem o vosso esforço
nada disto teria sido possível. Obrigada pela força, preocupação, mas acima de tudo,
obrigada pelo vosso carinho.

xvii
Abbreviations
bFGF basic fibroblast growth factor
CDK6 cyclin dependent kinase 6
CNC central nervous system
EGF epidermal growth factor
ES cells embryonic stem cells
GFAP glial fibrillary acidic protein
GFP green fluorescent protein
Id1 inhibitor of differentiation 1
iPS cells induced pluripotent stem cells
MEK1 mitogen-activated protein kinase kinase 1
miRNAs or miRs microRNAs
MMP mitochondrial membrane permeabilization
NeuN neuronal nuclei
NGF nerve growth factor
NS cells neural stem cells
NT2N Ntera2/D1 neuron-like
PC12 rat adrenal pheochromocytoma
pre-miRNA precursor miRNA
RISC miRNA-induced silencing complex
RMS rostral migratory stream
SGZ subgranular zone
SIRT1 silent information regulator 1
Sox2 SRY (sex determining region Y)-box 2
SVZ subventricular zone
TNF tumor necrosis factor
TNFR1 TNF-receptor type I
TNFR2 TNF-receptor type II
TUDCA tauroursodeoxycholic acid


Publications
The present thesis was mostly based on work that has been published, or is in
preparation for publication, in international peer-reviewed journals:
Aranha MM, Solá S, Low WC, Steer CJ, Rodrigues CMP. Caspases and p53 temporally
modulate FOXO3A/Id1 signaling during mouse neural stem cell differentiation. Journal
of Cellular Biochemistry 2009; 107 (4): 748-758.
Aranha MM, Santos DM, Xavier JM, Low WC, Steer CJ, Solá S, Rodrigues CMP.
Apoptosis-associated microRNAs are modulated in mouse, rat and human neural
differentiation. BMC Genomics 2010, 11: 514.
Aranha MM, Solá S, Santos DM, Low WC, Steer CJ, Rodrigues CMP. miR-34a
regulates mouse neural stem cell differentiation. 2010 (In preparation).
The following manuscripts have also been published during the Ph.D. studies:
Solá S, Amaral JD, Borralho PM, Ramalho RM, Castro RE, Aranha MM, Steer CJ,
Rodrigues CMP. Functional modulation of nuclear steroid receptors by
tauroursodeoxycholic acid reduces amyloid β-peptide-induced apoptosis. Molecular
Endocrinology 2006; 20 (10): 2292-2303.
Solá S, Amaral JD, Aranha MM, Steer CJ, Rodrigues CMP. Modulation of hepatocyte
apoptosis: cross-talk between bile acids and nuclear steroid receptors. Current Medicinal
Chemistry 2006; 13 (25): 3039-3051.
Aranha MM, Borralho PM, Ravasco P, Moreira da Silva IB, Correia L, Fernandes A,
Camilo ME, Rodrigues CMP. NF-κB and apoptosis in colorectal tumourigenesis.
European Journal of Clinical Investigation 2007; 37 (5): 416-424.

Publications
xx
Borralho PB, Moreira da Silva IB, Aranha MM, Albuquerque C, Leitão CN, Steer CJ,
Rodrigues CMP. Inhibition of Fas expression by RNAi modulates 5-fluorouracil-induced
apoptosis in HCT116 cells expressing wild-type p53. Biochimica et Biophysica Acta
2007; 1772 (1): 40-47.
Solá S, Aranha MM, Steer CJ, Rodrigues CMPR. Game and players: mitochondrial
apoptosis and the therapeutic potential of ursodeoxycholic acid. Current Issues in
Molecular Biology 2007; 9 (2): 123-138.
Aranha MM, Cortez-Pinto H, Costa A, Moreira da Silva IB, Camilo ME, Carneiro de
Moura M, Rodrigues CMP. Bile acid levels are increased in the liver of patients with
steatohepatitis. European Journal of Gastroenterology and Hepatology 2008; 20 (6):519-
525.
Ravasco P, Aranha MM, Borralho PM, Moreira da Silva IB, Correia L, Fernandes A,
Rodrigues CMP, Camilo M. Colorecta cancer: can nutrients modulate NF-κB and
apoptosis? Clinical Nutrition 2009; 29 (1):42-46.
Parry GJ, Rodrigues, CM, Aranha MM, Hilbert SJ, Davey C, Kelkar P, Low WC, Steer
CJ. Safety, tolerability and cerebrospinal fluid penetration of ursodeoxycholic acid in
patients with amyotrophic lateral sclerosis. Clinical Neuropharmacology 2010; 33(1): 17-
21.
Solá S, Xavier JM, Santos DM, Aranha MM, Jepsen K, Rodrigues CMP. A new
regulatory mechanism involving p53 in JMJD3-mediated neurogenesis in mouse neural
stem cells. 2010 (Submitted).

Table of Contents
Resumo ..................................................................................................................... vii
Abstract ..................................................................................................................... xi
Acknowledgements .................................................................................................. xv
Abbreviations ......................................................................................................... xvii
Publications ............................................................................................................. xix
1 General Introduction ............................................................................................... 1
1.1 Neural stem cells ..................................................................................... 3
1.1.1 Neural stem cells during developing CNS .............................................. 4
1.1.2 Adult neurogenesis ................................................................................. 6
1.1.3 In vitro culturing of NS cells .................................................................. 9
1.1.4 Potential therapeutic uses of NS cells ..................................................... 9
1.2 Apoptosis-associated proteins in cell differentiation ............................ 10
1.2.1 The tumor suppressor p53 .................................................................... 13
1.2.2 Caspases ................................................................................................ 19
1.2.3 Bcl-2 family members .......................................................................... 24
1.2.4 Others apoptotic players in cell differentiation .................................... 27
1.3 microRNAs ........................................................................................... 28
1.3.1 Overview of miRNA biogenesis and function ...................................... 28
1.3.2 Apoptosis-associated miRNAs as regulators of differentiation and cell
fate decision .......................................................................................... 30
Objectives ................................................................................................................. 35
2 Caspases and p53 modulate FOXO3A/Id1 signaling during mouse neural stem
cell differentiation ................................................................................................ 37

Table of Contents
xxii
2.1 Abstract ................................................................................................. 39
2.2 Introduction ........................................................................................... 40
2.3 Methods ................................................................................................ 42
2.3.1 Mouse NSC Culture and Differentiation .............................................. 42
2.3.2 Immunocytochemistry .......................................................................... 43
2.3.3 Evaluation of Apoptosis ....................................................................... 43
2.3.4 Total, Cytosolic, and Nuclear Protein Extraction ................................. 44
2.3.5 Immunoblotting .................................................................................... 44
2.3.6 p53-DNA Binding ELISA Assay ......................................................... 45
2.3.7 Inhibition of Caspase Activity .............................................................. 45
2.3.8 Short Interference RNA and Transfection ............................................ 45
2.3.9 RNA Isolation and RT-PCR ................................................................. 46
2.3.10 Densitometry and Statistical Analysis .................................................. 46
2.4 Results ................................................................................................... 46
2.4.1 The Akt/p-FOXO3A/Id1 Pathway is Down-Regulated during
Differentiation ....................................................................................... 46
2.4.2 Apoptosis Factors are Increased during Differentiation ....................... 49
2.4.3 Caspases Inhibit p-FOXO3A/Id1 Signaling during Differentiation ..... 50
2.4.4 Caspase Inhibition Delays Differentiation ............................................ 52
2.4.5 p53 Represses p-FOXO3A/Id1 Signaling in Early Stages of
Differentiation ....................................................................................... 53
2.4.6 Caspases and p53 Act in a Synergistic Manner to Modulate
Neurogenesis ......................................................................................... 54
2.5 Discussion ............................................................................................. 56
3 Apoptosis-associated microRNAs are modulated in mouse, rat and human neural
differentiation ....................................................................................................... 61
3.1 Abstract ................................................................................................. 63
3.2 Introduction ........................................................................................... 64

Table of Contents
xxiii
3.3 Methods ................................................................................................ 65
3.3.1 Cell Lines .............................................................................................. 65
3.3.2 RNA isolation and semiquantitative reverse-transcriptase-polymerase
chain reaction (RT-PCR) ...................................................................... 67
3.3.3 Preparation of labeled RNA and array hybridization ........................... 68
3.3.4 Evaluation of miRNAs expression levels by quantitative Real Time-
PCR ....................................................................................................... 69
3.3.5 Immunoblotting .................................................................................... 69
3.3.6 Immunocytochemistry .......................................................................... 70
3.3.7 Assessment of apoptosis ....................................................................... 70
3.3.8 Flow Cytometry .................................................................................... 71
3.3.9 Statistical Analysis ................................................................................ 72
3.4 Results ................................................................................................... 72
3.4.1 NS cells have both neurogenic and gliogenic potential in vitro ........... 72
3.4.2 Apoptosis-associate miRNAs are differentially modulated during
mouse neural stem cell differentiation .................................................. 74
3.4.3 Expression of apoptosis-related miRNAs is not associated with
increased cell death during mouse NS cell differentiation ................... 78
3.4.4 Apoptosis-associated miRNAs are upregulated in different models of
neural differentiation ............................................................................ 80
3.5 Discussion ............................................................................................. 85
4 miR-34a regulates mouse neural stem cell differentiation .................................. 89
4.1 Abstract ................................................................................................. 91
4.2 Introduction ........................................................................................... 92
4.3 Methods ................................................................................................ 93
4.3.1 Cell Culture ........................................................................................... 93
4.3.2 Small interfering RNA transfections, overexpression/antisense
experiments and cell treatments ............................................................ 94

Table of Contents
xxiv
4.3.3 Evaluation of miR-34a expression levels by quantitative Real Time-
PCR ....................................................................................................... 94
4.3.4 Flow cytometry analysis of Nestin, β-III Tubulin, NeuN and GFAP ... 95
4.3.5 Immunoblotting .................................................................................... 95
4.3.6 Immunocytochemistry .......................................................................... 96
4.3.7 Nuclear protein extraction .................................................................... 96
4.3.8 Electrophoretic mobility shift assay ..................................................... 96
4.3.9 Cytofluorometric analysis of apoptosis ................................................ 97
4.3.10 Statistical Analysis ................................................................................ 97
4.4 Results ................................................................................................... 97
4.4.1 miR-34a modulates the proportion of postmitotic neurons .................. 97
4.4.2 SIRT1 expression is modulated by miR-34a during neural
differentiation ....................................................................................... 99
4.4.3 miR-34a regulates neuronal differentiation through a SIRT1-mediated
mechanism .......................................................................................... 100
4.4.4 Astrogliogenesis can be modulated by miR-34a through a SIRT1-
independent mechanism ...................................................................... 102
4.4.5 miR-34a upregulation increases p53-DNA binding activity .............. 104
4.5 Discussion ........................................................................................... 106
5 Concluding Remarks .......................................................................................... 109
References .............................................................................................................. 117

Table of Contents
xxv
Figures
Figure 1.1. Location of NS cells niches ..................................................................... 3
Figure 1.2. NS cells in the developing brain .............................................................. 4
Figure 1.3. Neurogenesis in the SVZ. ........................................................................ 7
Figure 1.4. Neurogenesis in the dentate gyrus ........................................................... 8
Figure 1.5. Intrinsic and extrinsic pathways of apoptosis in mammals ................... 12
Figure 1.6. Schematic overview of miRNA biogenesis ........................................... 29
Figure 1.7. Involvement of members of miR-34 family in p53 network ................. 33
Figure 2.1. Time course of neurogenesis and gliogenesis ....................................... 47
Figure 2.2. The Akt/p-FOXO3A/Id1 signaling pathway is downregulated during
mouse NSC differentiation .................................................................... 48
Figure 2.3. p53 activity and caspase-3 cleavage are modulated during mouse NSC
differentiation. ....................................................................................... 50
Figure 2.4. Pan-caspase inhibition modulates p53 activity and interferes with the
Akt/p-FOXO3A/Id1 pathway during mouse NSC differentiation ........ 51
Figure 2.5. Inhibition of caspase activation delays neuronal and glial differentiation
of mouse NSCs ...................................................................................... 52
Figure 2.6. Silencing of p53 decreases p-FOXO3A and Id1 expression levels during
mouse NSC differentiation, without potentiating z-VAD.fmk
effects .................................................................................................... 54
Figure 2.7. Downregulation of p53 delays neuronal and glial differentiation of
mouse NSCs .......................................................................................... 55
Figure 3.1. Mouse NS cells have both neurogenic and gliogenic potential
in vitro ................................................................................................... 73
Figure 3.2. Apoptosis-associated miRNAs are modulated during mouse NS cell
differentiation ........................................................................................ 77
Figure 3.3. Differentiation of mouse NS cells is not associated with increased cell
death ...................................................................................................... 79
Figure 3.4. Inhibition of apoptosis by TUDCA was not associated with a decrease
in proapoptotic miRNAs expression...................................................... 80
Figure 3.5. miR-16, let-7a and miR-34a are increased during mouse ES cell
differentiation ........................................................................................ 81

Table of Contents
xxvi
Figure 3.6. Differentiation of PC12 and NT2N cells were associated with
modulated levels of miR-16, let-7a and miR-34a expression ............... 83
Figure 3.7. miR-34a overexpression increases the ratio of postmitotic neurons in
mouse NS cells ...................................................................................... 85
Figure 4.1. miR-34a modulates the proportion of NeuN-positive cells ................... 98
Figure 4.2. SIRT1 expression levels decrease after miR-34a overexpression ....... 100
Figure 4.3. miR-34a promotes the appearance of post-mitotic neurons by
downregulating SIRT1 expression ...................................................... 101
Figure 4.4. miR-34a positively regulates the astrocytic population under resveratrol
treatment .............................................................................................. 103
Figure 4.5. miR-34a overexpression increases p53-DNA binding activity. .......... 105
Tables
Table 1.1. Function of mammalian caspases in cell differentiation. ........................ 20
Table 3.1. Proapoptotic miRs expression during mouse NS cell differentiation ..... 75
Table 3.2. Antiapoptotic miRs expression during mouse NS cell differentiation .... 76

1 General Introduction


General Introduction
3
1.1 Neural stem cells
Neural stem (NS) cells are self-renewing multipotent cell populations present in the
developing and adult mammalian central nervous system (CNS) (Temple, 2001a). NS
cells generate neurons and glia of the developing brain, and account for neuronal
plasticity in the adult brain.
The process of generating functionally integrated neurons from progenitor cells,
termed neurogenesis, was traditionally believed to occur only during embryonic stages in
the mammalian CNS (Ramon y Cajal, 1913). However, almost forty years ago,
pioneering work by Altman and Das suggested the existence of continuing neurogenesis
throughout adulthood (Altman and Das, 1965). Significant progress has been made over
the past few years and investigators have now firmly established that new neurons are
indeed born in restricted regions of the adult mammalian CNS (Alvarez-Buylla and
Garcia-Verdugo, 2002; Gage, 2000; Kempermann and Gage, 2000). Since then, stem
cells have been isolated from many regions of embryonic and adult nervous systems (Fig.
1.1). The discovery of adult neurogenesis not only provides a unique model system to
understand basic mechanisms of neural development in the mature CNS, but also may
lead to strategies for neural tissue repair and cell-based replacement therapies of the
nervous system.
Figure 1.1. Location of NS cells niches. Principal regions of embryonic and adult nervous
systems, from which neural stem cells have been isolated. SVZ, subventricular zone; PNS,
peripheral nervous system. Adapted from Temple et al. 2001.
Chapter 1
4
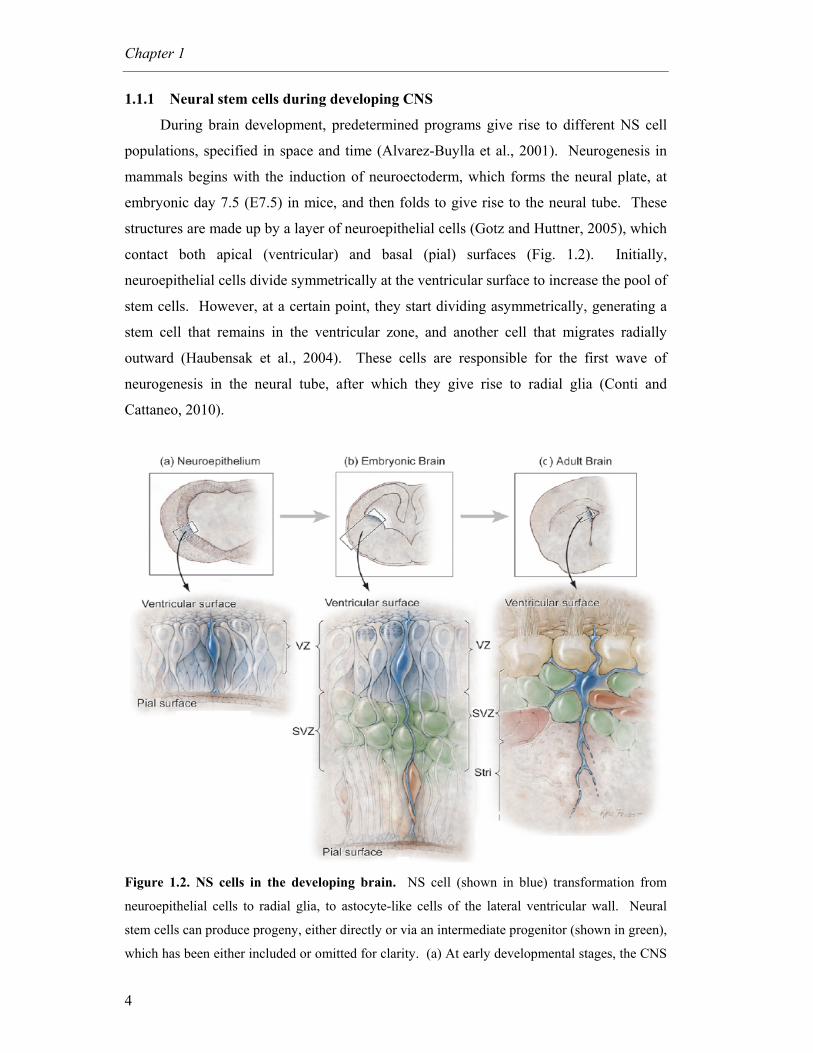
1.1.1 Neural stem cells during developing CNS
During brain development, predetermined programs give rise to different NS cell
populations, specified in space and time (Alvarez-Buylla et al., 2001). Neurogenesis in
mammals begins with the induction of neuroectoderm, which forms the neural plate, at
embryonic day 7.5 (E7.5) in mice, and then folds to give rise to the neural tube. These
structures are made up by a layer of neuroepithelial cells (Gotz and Huttner, 2005), which
contact both apical (ventricular) and basal (pial) surfaces (Fig. 1.2). Initially,
neuroepithelial cells divide symmetrically at the ventricular surface to increase the pool of
stem cells. However, at a certain point, they start dividing asymmetrically, generating a
stem cell that remains in the ventricular zone, and another cell that migrates radially
outward (Haubensak et al., 2004). These cells are responsible for the first wave of
neurogenesis in the neural tube, after which they give rise to radial glia (Conti and
Cattaneo, 2010).
Figure 1.2. NS cells in the developing brain. NS cell (shown in blue) transformation from
neuroepithelial cells to radial glia, to astocyte-like cells of the lateral ventricular wall. Neural
stem cells can produce progeny, either directly or via an intermediate progenitor (shown in green),
which has been either included or omitted for clarity. (a) At early developmental stages, the CNS

General Introduction
5
is constituted by the neural tube, which is composed by neuroepithelial cells. These cells can
divide either symmetrically, at the ventricular surface, to expand the stem cell pool, or
asymmetrically, to generate neurons. (b) Radial glial cells arise from neuroepithelial cells, and
divide to generate neurons or glia, either directly or via an intermediate progenitor in the SVZ.
The radial processes of radial glial cells support the migration of neuroblasts (shown in red). (c)
In the adult brain, radial glia have transformed into astrocyte-like cells that have the capacity to
generate olfactory bulb interneurons. The ventricular zone (VZ) also disappears, but SVZ
remains into adulthood in some areas. Stri, striatum. Adapted from Merkle et al. 2006.
Radial glia arise in the neural tube at about E9.5 in mice. These cells originate from
neuroepithelial cells at the beginning of neurogenesis, and represent the main cell type in
the developing brain, where they act both as neural progenitors and as scaffolding for
migrating newborn neurons (Conti and Cattaneo, 2010). Like neuroepithelial progenitors,
radial glia are a transient population in the developing brain that exhibits definite
morphological hallmarks. Their soma are located in the ventricular zone, and they extend
a long process to the pial surface. In addition, radial glia exhibit several glial
characteristics, such as glycogen granules (Choi and Lapham, 1978), and the expression
of numerous glial antigens, including brain lipid binding protein (Feng et al., 1994), the
glutamate transporter GLAST (Shibata et al., 1997), tenascin (Yuasa, 2001), and glial
fibrillary acidic protein (GFAP) in some species (Choi and Lapham, 1978; Levitt and
Rakic, 1980). In vivo, radial glia have the ability to generate different neural lineages, in
a time-controlled fashion (Anthony and Heintz, 2008; Ogawa et al., 2005). A similar
heterogeneity has also been observed in vitro. In mammals, radial glial cells disappear
from the brain soon after birth, but in many species they persist into the adulthood
(Merkle and Alvarez-Buylla, 2006). In mammals, the role of NS cells is filled by a
closely related astrocyte-like cell.
Similarly to radial glia, adult mammalian NS cells have many astrocytic
characteristics, such as the expression of GFAP (Doetsch et al., 1999; Garcia et al., 2004;
Imura et al., 2003). In the adult brain, these astrocyte-like cells are mainly found in two
regions: the subventricular zone (SVZ) of the lateral wall of the lateral ventricle, and the
subgranular zone (SGZ) of the dentate gyrus of the hippocampus. The astrocytic nature
of NS cells has been confirmed when GFAP+ cells were conditionally ablated in the adult
mouse brain, resulting in an almost complete loss of neurogenesis (Garcia et al., 2004;
Imura et al., 2003).

Chapter 1
6
Since radial glial cells and SVZ astrocytes share many properties, it has been
hypothesized that they belong to the same lineage (Tramontin et al., 2003). In particular,
radial glial cells of the neonatal lateral ventricular wall occupy the same region as
astrocytic stem cells of the adult SVZ. In summary, current evidence suggests that NS
cells gradually transform from neuroepithelial cells to radial glial cells, to astrocyte-like
cells (Merkle and Alvarez-Buylla, 2006).
1.1.2 Adult neurogenesis
New neurons are continuously added to neural circuits in the adult vertebrate brain
(Doetsch, 2003). In mammals, including humans, neurons are added to restricted brain
regions, the olfactory bulb and the hippocampus (for a review, see (Zhao et al., 2008a).
The addition of new neurons to these regions represents a process by which the brain can
make changes to its own functional circuitry, in addition to molecular, synaptic or
morphological alterations in individual cells. Indeed, this cell-level renovation is not
static or merely restorative, rather constitutes an adaptive response to challenges imposed
by the environment and/or internal state of the animal (Lledo et al., 2006).
The identity of NS cells and adult neurogenesis have been most clearly defined in
the SVZ of adult mice. The SVZ is a layer of dividing cells that extends along the lateral
walls of the lateral ventricle. Neurons are born throughout the SVZ, and feed into a
network of chains of tangentially migrating neuroblasts (Doetsch and Alvarez-Buylla,
1996) that coalesce to form the rostral migratory stream (RMS), leading to the olfactory
bulb (Fig. 1.3). Here, neuroblasts, differentiate into two kinds of olfactory interneurons,
granule and periglomerular cells (Doetsch and Scharff, 2001). The SVZ is separated
from the lateral ventricle by a single layer of multiciliated ependymal cells.
Chemorepulsive factor gradients produced by these cells guide neuroblast migration in
the adult brain (Sawamoto et al., 2006). More than 30,000 neuroblasts exit the rodent
SVZ for the RMS, each day (Alvarez-Buylla et al., 2001).
Three types of precursor cells exist in the SVZ: type B GFAP+ progenitors, type C
transit amplifying cells, and type A migrating neuroblasts (Fig. 1.3). Type B cells are
slowly dividing radial-glia like progenitors, which have been hypothesized to represent
the primary NS cells in vivo (Mu et al., 2010). They are thought to generate rapidly
dividing progenitors, type C cells, which typically have either no or very short processes,
and are characterized by the expression of Dlx2, Mash1, and epidermal growth factor

General Introduction
7
receptor (EGFR). The majority of these intermediate progenitors subsequently give rise
to type A cells that express both doublecortin (DCX) and poly-sialated neural cell
adhesion molecule (PSA-NCAM). These cells migrate into the olfactory bulb, through
the RMS, and differentiate into GABA-producing and dopamine-producing interneurons.
Interestingly, the differentiation potential of SVZ progenitor cells appears to be limited,
as the fate of the progeny is determined by the positional information established during
early development of the CNS (Merkle et al., 2007).
Figure 1.3. Neurogenesis in the SVZ. Progenitor cells (A–C) in the SVZ are located next to the
ependymal cell (E) layer lining the lateral ventricles. A subset of slowly dividing GFAP+ radial
cells (type B cells) has the potential to serve as adult NS cells and generate rapidly dividing,
transit-amplifying nonradial NS cells (type C cells), which in turn give rise to neuroblasts (type A
cells) that migrate through the rostral migratory stream (RMS) toward the olfactory bulb (OB).
Newborn neurons are integrated as granule neurons, in the granule cell layer (GCL), and as
periglomerular neurons (not shown), in the glomerular layer (GL). Mi, mitral cell layer; EPL,
external plexiform layer. Adapted from Zhao et al. 2008.
The hippocampus is other major site of neurogenesis in adult mammals. Granule
neurons in the dentate gyrus are born locally in the SGZ, which lies between the granule
cell layer and the hilus. In contrast to the extensive tangential migration undertaken by
olfactory bulb neurons, hippocampal granule neurons move only a short distance into the
granule cell layer (Fig. 1.4). On a superficial level, describing adult hippocampal
neurogenesis is very straightforward. A dividing precursor cell gives rise to daughter
cells, which migrate away from the site of division (Kuhn et al., 1996), and start to
differentiate into neurons (Kempermann et al., 2003). They first extend dendrites toward
the molecular layer, and later an axon that projects to area CA3 (Stanfield and Trice,

Chapter 1
8
1988). Over four to seven weeks, these neurons become integrated into the local neuronal
circuits (Jessberger and Kempermann, 2003).
Figure 1.4. Neurogenesis in the dentate gyrus. In the adult SGZ, a population of GFAP+ Sox2+
radial cells corresponds to quiescent NS cells (type 1 cells). They coexist with actively
proliferating, GFAP- Sox2+ nonradial NS cells (type 2 cells) that generate both astrocytes and
neuroblasts. Neuroblasts then migrate into the granule cell layer, and predominantly differentiate
into local glutamatergic dentate granule cells. Progenitor cells in the dentate gyrus are influenced
by local astrocytes (not shown) and by the vasculature (red). GCL, granule cell layer; Mol,
molecular layer. Adapted from Zhao et al. 2010.
Two types of neural progenitors can be identified in the SGZ, according to their
specific morphologies and expression of unique molecular markers (Fig. 1.4). Type 1
hippocampal progenitors act as quiescent NS cells. They have a radial process, spanning
the entire granule cell layer, and ramify in the inner molecular layer. These cells express
nestin, GFAP, and the transcription factor sex-determining region Y-box 2, Sox2 (Fukuda
et al., 2003; Garcia et al., 2004; Suh et al., 2007). Although expressing the astrocyte
marker GFAP, these cells are morphologically and functionally different from mature
astrocytes. They may generate actively self-renewing nonradial progenitors (type 2
cells), expressing Sox2 and nestin, but not GFAP. Type 2 cells, in turn, give rise to
DCX+ neuroblasts that predominantly differentiate into local glutamatergic dentate
granule cells (Mu et al., 2010). A recent study showed that type 2 Sox2+ cells can self-
renew, and that a single Sox2+ cell can give rise to neurons and astrocytes, providing the

General Introduction
9
first in vivo evidence of stem cell properties of hippocampal neural progenitors (Suh et
al., 2007).
1.1.3 In vitro culturing of NS cells
For nearly 20 years, growth factor-based protocols have been developed, leading to
NS cell expansion in both floating and adherent conditions (Kokovay et al., 2008). In
addition, progress in cell culture technologies has enabled researchers to induce the
neuralization of mouse and human embryonic stem (ES) cells, derived from the inner cell
mass of the blastocyst, in vitro (Conti and Cattaneo, 2010). During neural differentiation,
ES cells undergo progressive lineage restrictions, similar to those observed in normal
fetal development (Pankratz et al., 2007), leading to the generation of a range of distinct
neural precursor populations. These can be used to study the molecular and cellular
events that occur during stage-specific transitions between different populations
(Nishikawa et al., 2007).
Neurospheres are free-floating aggregates of neural progenitors, each potentially
derived from a single NS cell (Reynolds et al., 1992). Their generation relies on tissue
microdissection, followed by exposure to growth factors (Chojnacki and Weiss, 2008).
Neurospheres and monolayer culturing systems of NS cells can be generated from ES
cells, induced pluripotent stem (iPS) cells (derived from reprogrammed somatic cells)
and from stem cell niches of the fetal and adult brain (Conti and Cattaneo, 2010). Both
systems have the potential to give rise to neurons, astrocytes and oligodendrocytes. The
different cellular compositions of the neurospheres (a mixed population, with only a
fraction of the cells exhibiting NS cell properties) and monolayer cultures (with
homogeneous composition) results in low and high neurogenic potential, respectively.
Although many efforts are being made to better understand cellular and molecular
events associated with differentiation (Abranches et al., 2009), determining the best
sources for the in vitro derivation of NS cells and optimizing protocols for stable, clonal
proliferation are still central goals of stem cell research.
1.1.4 Potential therapeutic uses of NS cells
Despite the presence of NS cells in the adult mammalian brain, neurons are not
replaced in most regions after injury or pathological conditions. However, evidence of
neurogenic upregulation in the damaged brain has led to interest in therapeutic strategies

Chapter 1
10
that exploit naturally existing responses. Such approaches could prove fruitful, not only
for stroke and trauma patients, but also in patients with degenerative brain diseases, such
as Parkinson’s disease, Huntington’s disease and multiple sclerosis (Curtis et al., 2003;
Jin et al., 2004). Nevertheless, several problems must be solved before the potential
attributed to stem cells becomes a realistic clinical strategy for cellular repair. The
biggest single problem still to be solved is how to direct and control the differentiation of
specific target phenotypes required for replacement and repair in each disease. Selection
of appropriate starting cells and adequate parameters of in vitro manipulation are likely to
be crucial factors in directing appropriate phenotypic differentiation. Further, a more
comprehensive understanding of the molecular pathways controlling stem cell self-
renewal and differentiation, in particular the apparent molecular link between
programmed cell death and cell differentiation, would accelerate efforts to generate
clinically relevant cell types from stem cells.
1.2 Apoptosis-associated proteins in cell differentiation
Apoptosis is a highly regulated form of cell death known to sculpt tissues during
development, and maintain tissue homeostasis by eliminating unnecessary or harmful
cells (Ellis et al., 1991). Apoptosis is a genetically predetermined mechanism. Genetic
analysis of the nematode Caenorhabditis elegans provided the first insights into the
molecular mechanism of apoptosis (Yuan et al., 1993). The canonical apoptotic pathways
are well conserved in all metazoans from invertebrates to vertebrates. The best
characterized and the most prominent are designated by extrinsic and intrinsic pathways
(Kroemer et al., 2007). In the extrinsic pathway, also known as “death receptor
pathway”, apoptosis is triggered by ligand-induced activation of death receptors at the
cell surface. In the intrinsic pathway, also called “mitochondrial pathway”, apoptosis
results from an intracellular cascade of events, mainly characterized by the mitochondrial
membrane permeabilization (MMP).
In the extrinsic pathway of apoptosis, ligation of death receptors causes the
recruitment and oligomerization of the adapter molecule Fas-associating death domain-
containing protein (FADD) within the death-inducing signaling complex (DISC).
Oligomerized FADD binds the initiator caspase-8 and -10, causing their dimerization and
activation (Fig. 1.5) (Debatin and Krammer, 2004). As an alternative, the extrinsic

General Introduction
11
pathway can be activated by the so-called dependency receptors, which are believed to be
connected to rapid caspase activation as well.
Fas-induced apoptosis has been shown to follow almost exclusively one of two
different Fas signaling pathway (Scaffidi et al., 1998), mainly depending on the amount
of active caspase-8 available in the cytosol. In type I cells, Fas-induced apoptosis ensures
enough activation of caspase-8, so that it can go on to activate caspase-3 and -7. In
contrast, in type II cells, apoptosis proceeds by an amplification loop through
mitochondria. In this case, caspase-8 cleaves the inactive cytoplasmic protein Bid, a pro-
apoptotic Bcl-2-family member, thus producing active truncated Bid (tBid) (Li et al.,
1998). The carboxy-terminal fragment of Bid then translocates to the mitochondria
leading to oligomerization of Bax or Bak and the consequent MMP, which in turn results
in caspase-3 activation (Korsmeyer et al., 2000).
Most cell death in vertebrates proceeds via the intrinsic or mitochondrial pathway
of apoptosis (Green and Kroemer, 2004). Here, the executioner caspases are cleaved and
activated by initiator caspase-9, which is activated by multimerization on the adapter
molecule apoptosis protease activating factor-1 (Apaf-1), within a multiprotein complex
called “apoptosome”. Apaf-1 preexists in the cytosol as a monomer, and its activation
depends on the presence of cytochrome c (Cyt c) and ATP/dATP (Cain et al., 2002). The
release of Cyt c, which normally resides only in the inner mitochondrial space where it
functions as an electron shuttle in the respiratory chain (Bernardi and Azzone, 1981), is
rate-limiting for the generation of the apoptosome. Hence MMP is the critical event
responsible for caspase activation in the intrinsic pathway. MMP can also commit a cell
to die when caspases are not activated. This “caspase-independent death” (Chipuk and
Green, 2005; Kroemer and Martin, 2005) can occur by either an irreversible loss of
mitochondrial function or by the mitochondrial release of caspase-independent death
effectors, including apoptosis-inducing factor (AIF) (Susin et al., 1999), endonuclease G
(EndoG) (Li et al., 2001), and others (Chipuk and Green, 2005; Kroemer and Martin,
2005).

Chapter 1
12
Figure 1.5. Intrinsic and extrinsic pathways of apoptosis in mammals. The intrinsic pathway
involves the mitochondria, which acts as an “intracellular death receptor”, receiving a variety of
pro-apoptotic signals that trigger oligomerization of pro-apoptotic proteins (Bcl-2-associated
protein, Bax, and Bcl-2-antagonist killer, Bak, to produce mitochondrial outer membrane
permeabilization. This leads to the release of Cyt c, which activates Apaf-1, induction of
apoptosome formation, recruitment/activation of procaspase-9, and direct processing and
activation of procaspase-3 and -7. In the extrinsic pathway, the Fas receptor ligand (FasL)
triggers the membrane-bound DISC, which recruits procaspase-8 and activates caspase-3 directly.
In some cell types, caspase-8 can also cleave Bid to form tBid, which interacts with Bax/Bak to
trigger mitochondrial outer membrane permeabilization, Cyt c release and apoptosome formation.
The activation of caspase-3 and -7 is antagonized by inhibitor of apoptosis proteins (IAPs), which
in turn can be inhibited by Smac/Diablo and Omi/HtrA2. Activation of caspase-3 and -7
orchestrates the demolition of the cell by cleavage of specific substrates, including poly (ADP)
ribose polymerase (PARP), inhibitor of caspase-3-activated DNase (ICAD) and mammalian
STE20-like kinase 1 (MST1). Adapted from D’Amelio et al. 2010.
A well-known observation in the apoptosis field is that differentiating cells are
more refractory to apoptosis compared with less differentiated cells (Yi and Yuan, 2009).
Next, it will be discussed the emerging evidence for nonapoptotic functions of apoptosis
players in mediating cell fate specification and differentiation.

General Introduction
13
1.2.1 The tumor suppressor p53
The tumor suppressor p53 is a central protein in mammalian stress response.
Owing to its ability to integrate many different signals controlling cell life and death, p53
has been named the “guardian of the genome” and plays a crucial role in maintaining
genomic stability in somatic cells (Vousden and Prives, 2009). Under normal
physiological conditions p53 remains inactive and unstable, but is rapidly stabilized and
activated by upstream kinases in response to genotoxic and oncogenic stresses, leading to
cell cycle arrest, apoptosis or senescence (Zhao and Xu, 2010). These functions of p53
can protect the genome from accumulating genetic mutations, by allowing the time for
faithful repair of DNA damage or by eliminating cells with excessive DNA damage
(Zhao and Xu, 2010).
p53 functions mainly as a DNA-binding, sequence-specific transcription factor that
activates the expression of multiple genes, including p21, noxa and puma (Vousden and
Prives, 2009), which in turn mediate p53 functions in cell cycle arrest, apoptosis and
senescence. In addition, p53 regulates the expression of proteins that modulate its own
activation and stability, such as Mdm-2, forming multiple positive and negative feedback
loops (Harris and Levine, 2005). Posttranslational modifications of p53 are also
important modulators of its function and stability (Bode and Dong, 2004). In fact,
modifications have been detected on at least 30 different sites on the p53 protein. It has
been proposed that the specific combination of modifications on p53 may provide
selectivity toward a particular cell response.
The role of p53 in controlling genomic stability is crucial in stem cell populations.
The accumulation of unrepaired DNA in ES cells could not only promote tumorigenesis
in multiple cell lineages, but also pass these mutations to the progeny, leading to genetic
instability in differentiated cells (Zhao and Xu, 2010). ES cells must have developed
mechanisms to cope with various stresses, in particular, DNA damage insults. However,
p53 checkpoint pathways are compromised in ES cells, which do not induce p53-
dependent cell cycle arrest and apoptosis after certain types of physiological DNA
damage, although p53 is expressed in abundant quantities in ES cells (Aladjem et al.,
1998).
p53-mediated checkpoint controls can be activated during differentiation. Recent
studies suggest that p53 may maintain genetic stability of ES cells by inducing
differentiation. Following genotoxic stress, p53 can induce differentiation of ES cells by
directly suppressing Nanog expression (Lin et al., 2005), which is the master

Chapter 1
14
transcriptional factor that maintains self-renewal of ES cells. Therefore, the outcome of
p53 activation in DNA-damaged ES cells is to differentiate ES cells into other cells types
that can undergo efficient p53-dependent cell-cycle arrest or apoptosis (Aladjem et al.,
1998; Chao et al., 2000), leading to the maintenance of genomic stability in self-renewing
ES cells. Importantly, p53 was shown to suppress Nanog expression also after treatment
with retinoic acid, suggesting that p53 may be important not only to maintain genomic
stability but also to differentiate ES cells. Given that, p53 should be viewed not as a
tumor suppressor protein, but rather as a crucial decision-maker molecule (Tedeschi and
Di Giovanni, 2009; Vousden and Prives, 2009).
1.2.1.1 Role of p53 during neuronal differentiation
Before the link of p53 and Nanog had been established, mounting evidence was
already suggesting a role for p53 in cell differentiation, including that of neuronal
precursors. Initially, p53 was considered non-essential to development since the majority
of p53-null mice were considered phenotypically normal at birth, with markedly
increased tumor development later in life (Donehower et al., 1992). However, the
discovery of low frequency neural tube defects in p53-/- mice (Armstrong et al., 1995; Sah
et al., 1995) was an apparent paradox, and suggested that the role of p53 in development
may be more complex than initially believed. Determination of p53 mRNA levels during
mouse embryonic development showed that it peaked during the differentiation of several
tissues, including early neuronal precursor cells of the brain, but strongly declines during
terminal differentiation (Louis et al., 1988; Rogel et al., 1985; Schmid et al., 1991). In
addition, evaluation of p53-dependent transcriptional activation during normal
development in vivo indicated that p53 activity is at maximum during neuronal
differentiation, and is clustered in areas that are not correlated with apoptosis (Gottlieb et
al., 1997; Komarova et al., 1997).
Later work supported a possible link between p53 and cell differentiation of NS
cells through the regulation of cell-cycle progression. One report showed that p53
negatively regulates the proliferation and survival of adult NS cells, without affecting
their differentiation potential (Meletis et al., 2006). By comparing the transcriptome of
adult NS cells from p53-null and wild-type mice, the authors identified altered expression
of several cell cycle regulators, including downregulation of p21 expression, in the
absence of p53. Of note, the expression of all well-known neuronal differentiation

General Introduction
15
markers was not affected. However, recent experiments of NS cells extracted from the
olfactory bulb of wild-type and p53-null mouse embryos showed that p53 controls
proliferation, chromosomal stability and differentiation pattern of mouse olfactory bulb
stem cells (Armesilla-Diaz et al., 2009). In fact, p53-null NS cells have increased
proliferation and a bias towards neuronal phenotype, with concomitant reduction of
astrocytes. This effect had been previously reported in SVZ-derived stem cells from adult
mice (Gil-Perotin et al., 2006).
In vitro models of neuronal differentiation also suggest a role for p53 during
differentiation. Studies in neuronal-like PC12 pheochromocytoma and neuroblastoma
cells revealed that p53 gene expression is induced and required during neurotrophin-
dependent neuronal differentiation and maturation (Hughes et al., 2000; Montano, 1997;
Poluha et al., 1997; Zhang et al., 2006). Furthermore, p53 was shown to be crucial for
correct neurite outgrowth and maturation of cortical neurons in vitro (Di Giovanni et al.,
2006; Tedeschi et al., 2009). p53 knockdown reduced spontaneous and directed
differentiation rate in human ES cells (Qin et al., 2007), while activation of p53 with the
small-molecule activator nutlin led to rapid differentiation (Maimets et al., 2008). Still,
although p53 protein levels were reduced after retinoic acid-induced ES cell
differentiation, its transcriptional activity was increased (Lin et al., 2005). Finally, the
involvement of p53 has also been shown in spermatogenesis (Rotter et al., 1993), eye
development (Reichel et al., 1998), renal development (Saifudeen et al., 2009),
osteogenesis (Lengner et al., 2006), immune development (Matas et al., 2004), lung
development (Tebar et al., 2001) and muscle differentiation (Porrello et al., 2000).
1.2.1.2 Mechanism of p53 control of cell differentiation
Despite the supporting data on transcriptional activity of p53 protein in
differentiation, the mechanism through which p53 acts in this cellular context is still a
puzzling question. p53 was reported to play a critical role in nerve growth factor (NGF)-
mediated neuronal differentiation of PC12 cells, in part via regulation of TrkA
expression, which in turn activates mitogen-activated protein kinase (MAPK) pathways
(Browes et al., 2001; Zhang et al., 2006). In the same study, p53 was reported to regulate
cell cycle arrest by inducing p21 expression. Moreover, by genome-wide chromatin
immunoprecipitation (ChIP), Brynczka and colleagues have recently identified new
putative p53 target genes during NGF-mediated PC12 neuronal differentiation, among

Chapter 1
16
which are wnt7b involved in dendritic development, and the tfcp214/grhl3 grainyhead
homolog implicated in ectodermal development (Brynczka et al., 2007). Di Giovanni and
co-workers showed that p53 regulates the expression of both actin-binding protein
Coronin 1b and the GTPase Rab13, which are required for physiological neurite
outgrowth in PC12 cells and dorsal root ganglion neurons (Di Giovanni et al., 2005). In
this sense, within the PC12 neuronal context, p53 may have the ability to regulate two
critical events for neuronal differentiation, cell cycle arrest and neurite outgrowth.
In ES cells, p53 is involved in the rapid down-regulation of Nanog expression
during retinoic acid-induced differentiation (Lin et al., 2005). Furthermore, p53-
dependent expression of p21, Mdm2 and Killer/DR5 were all also markedly induced
indicating again a mechanism of action involving cell cycle regulation. In fact, it has
been shown that p21 can negatively regulate the self-renewal of adult NS cells (Mori et
al., 2001). Similar results were obtained in human ES cells (Maimets et al., 2008).
Integrated transcriptomic profiling, in silico promoter analysis and functional
studies of murine NS cells established that dual, but not singular, inactivation of p53 and
Pten promotes an undifferentiated state, with high self-renewal potential, and drives
increased Myc protein levels (Zheng et al., 2008). Functional studies validated increased
Myc activity as a potent contributor to impaired differentiation and enhanced self-renewal
of NS cells doubly null for p53 and Pten.
Studies in neuronal cells have suggested that the interaction of p53 with the
neuronal specific and pro-differentiation transcription factor Brn-3a facilitates a shift of
p53 transcriptional activity from cell death to neuronal differentiation (Hudson et al.,
2005). In fact, when forming a complex with Brn-3a, p53 is not able to activate pro-cell -
death genes such as Bax and Noxa, but rather shows increased affinity for the pro-
differentiation gene p21. Therefore, Brn-3a might play a crucial role in determining the
pathway taken by p53, when co-expressed during development, and in controlling the cell
fate.
Thus, p53 may generally contribute to neuronal differentiation by targeting cell
cycle regulators, and particularly by controlling the expression of differentiation-specific
genes in a cell-type specific manner. This could be directed by the interaction between
Brn-3a and p53, which facilitates a shift of p53 transcriptional activity from cell death to
neuronal differentiation.

General Introduction
17
1.2.1.3 p53 switch and multiple biological functions
Genomic elements regulated by p53 during neuronal differentiation may be unique,
and different from those regulated in genotoxic stress and apoptosis. This is made
possible by the multiple post-translational modifications that target p53 on its N- and C-
termini which are not limited to phosphorylation, but may also include acetylation,
sumoylation, neddylation, and ubiquitination (Lavin and Gueven, 2006). p53 post-
translational modifications directly affect the transcriptional activity of p53 and regulate
its affinity to diverse cofactors, which in turn regulate the occupancy of p53 specific
promoters (Sims and Reinberg, 2008).
The best characterized p53 post-translational modifications are phosphorylation and
acetylation. Kinases phosphorylate several serine and threonine residues, mainly in the
p53 N-terminal region. Phosphorylation at Ser 15, Ser 20, Ser 33, Ser 46 and Thr 18
promotes p53 stabilization by preventing either its nuclear export, or its recruitment to
specific promoters. Importantly, soon after the onset of differentiation of ES cells,
phosphorylation of p53 at Ser 15, Ser 315 and Ser 392 is significantly increased, which is
thought to activate p53 (Xu, 2003). p53 phosphorylation at Ser 315 was also shown to be
particularly important for p53-dependent suppression of Nanog during differentiation of
ES cells (Lin et al., 2005).
Acetylation targets mainly two distinct regions of the p53 C-terminus and, in
neurons, involves the activity of at least two histone acetyltransferases, CREB-binding
protein (CBP)/p300 and p300/CBP-associated protein (P/CAF). CBP/p300 acetylates
Lys370, Lys372, Lys373 and Lys382, whereas P/CAF acetylates only a single residue,
Lys320 (Brooks and Gu, 2003). Importantly, these kinase and acetyltransferase pathways
are activated downstream from NGF and brain-derived nerve growth factor (BDNF)
signaling during neuronal differentiation and axon outgrowth. Interestingly, the
acetylation of p53 at Lys 320 by P/CAF and the highly homologous acetyltransferase
CGN5 leads to increased transcriptional activation of the p21 promoter, which triggers
G1/S arrest and promotes neuronal differentiation in PC12 cells (Wong et al., 2004). In
addition, p53 acetylation in the same residue was reported to be involved in the
promotion of neurite outgrowth (Di Giovanni et al., 2006). Furthermore, acetylated p53
at Lys372, Lys373 and Lys382 was reported to drive axon outgrowth and GAP-43
expression, and to bind specific elements on the neuronal GAP-43 promoter (Tedeschi et
al., 2009).

Chapter 1
18
The ability of p53 to modulate different transcriptional responses in neurons,
depending on specific signaling and transcriptional contexts might be linked not only to
the post-transcriptional modifications of p53, but also to the cross-talk between p53 and
post-transcriptional modifications on histones. Histone modifications influence
chromatin structure, which in turn influences the capacity of transcription factors to bind
to specific promoters. A unique chromatin environment could regulate the affinity of p53
to specific promoters. Specific patterns of p53 codes might lead to different biological
outcomes, depending on the transcriptional context in a given cell or tissue (Murray-
Zmijewski et al., 2008). Understanding the role of such mechanisms in the regulation of
the neuronal phenotype during development undoubtedly represents an exciting challenge
for future investigations.
1.2.1.4 Role of p53 in limiting somatic cell reprogramming
The recent demonstration of in vitro reprogramming of somatic cells into iPS cells
by Yamanaka and colleagues represents a major advance in the field of stem cells. This
discovery raises hope for modeling of human diseases, especially those with complex
genetic traits. In addition, iPS cells could become a renewable source of autologous cells
for transplantation into human patients.
Reprogramming was initially achieved by transduction of four transcription factors
(Oct3/4, Sox2, c-Myc, and Klf-4), but only a minority of donor somatic cells can be
reprogrammed to pluripotency (Takahashi and Yamanaka, 2006). Although alternative
and more efficient reprogramming protocols are being subject of extensive investigation,
it was recently shown that the inhibition of the p53/p21 pathway increased cell division
rate and resulted in accelerated kinetics of iPS cell formation, which was directly
proportional to the increase in cell proliferation (Hanna et al., 2009). By comparing the
reprogramming efficiency of wild-type and p53-deficient mouse and human fibroblasts,
several studies have shown great increase in reprogramming frequency in p53-deficient
cells, indicating that p53 is an effective barrier of induced pluripotency (Hong et al.,
2009; Kawamura et al., 2009; Zhao et al., 2008b). Nevertheless, if inactivation of the p53
pathway is a prerequisite for successful reprogramming, these findings raise concerns on
the genomic stability and tumorigenecity of iPS cells and their derivatives.

General Introduction
19
1.2.2 Caspases
Caspases are a family of cysteinyl aspartate-specific proteases that are highly
conserved in multicellular organisms and function as central regulators of apoptosis. To
date, 11 genes were found in the human genome to encode 11 human caspases, caspase-1
to -10 and caspase-14, whereas 10 genes were found in the mouse genome to encode 10
murine caspases, caspase-1, -2, -3, -6, -7, -8, -9, -11, -12 and -14 (for a review, see Li and
Yuan, 2008). Human caspase-4 and -5 are functional orthologs of mouse caspase-11 and
-12, whereas human caspase-10 is absent in the mouse genome. The remaining caspases
with the same numbers in human and mouse are functional orthologs of each other.
Members of the caspase family are subdivided into upstream “initiator” caspases (in
mammals, caspase-1, -2, -4, -5, -8, -9, -10, -11, and -12), which respond to proapoptotic
signals, and downstream “executioner” caspases (in mammals, caspase-3, -6, -7, and -14)
(Yi and Yuan, 2009). Caspases are synthesized as zymogens and their activation requires
allosteric conformational changes, specific cleavage after a selective aspartate residue, or
both. To date, four pathways leading to caspase activation have been characterized in
vertebrate organisms, including the receptor-mediated pathway, the mitochondria-
mediated pathway, the granzyme B-mediated pathway, and the endoplasmic reticulum-
mediated pathway (Nicholson, 1999). Extensive biochemical and structural analysis
indicate that inflammatory and initiator caspases are recruited to larger complexes that
induce caspase dimerization, autoproteolysis, and protease activation (Shi, 2004).
Following activation, apoptotic initiator caspase-8 and -9 cleave executioner procaspase-
3, -6, and -7. This leads to cleavage of more than 1,000 proteins (Dix et al., 2008;
Mahrus et al., 2008) to produce the characteristic apoptotic phenotypes of membrane
blebbing, nuclear condensation, DNA fragmentation, and ultimately phagocytosis by
immune cells.
Although the critical function of caspases in apoptosis is firmly established, some
pro-apoptotic caspases, namely caspase-3, -8 and -14 also regulate differentiation in
certain cells types (Table 1.1). The cellular processes that stop caspases short of inducing
apoptosis in diverse biological settings remain a mystery.

Chapter 1
20
Table 1.1. Function of mammalian caspases in cell differentiation
Caspase Cell Type
1 Not determined
2 Not determined
3
Epithelial lens, erythroblasts, megakaryocytes, spermatids, NS
and ES cells, osteoclasts, bone marrow stromal and skeletal
muscle cells
4 Not determined
5 Not determined
6 Not determined
7 Not determined
8 Macrophages and trophoblasts
9 Not determined
10 Not determined
11 Not determined
12 Not determined
14 Keratinocytes
Adapted from Yi et al. 2009.
1.2.2.1 Caspase-3
The role of caspase-3 during terminal differentiation has been well established.
Terminal differentiation of certain cell types, such as lens epithelial cells, erythroblasts,
and megakaryocytes, requires the elimination of nuclei. The enucleation process of lens
epithelial cells is prevented after inhibition of caspase activity by the pan caspase
inhibitor, z-VAD.fmk, suggesting that caspases are involved in the specific elimination of
nuclei as opposed to elimination of the whole cell (Ishizaki et al., 1998). The terminal
maturation of erythroblasts into enucleate red blood cells also requires caspase activity.
Caspase-3, together with caspase-9, -7 and -2 are transiently activated during erythroid
differentiation induced by erythropoietin, and caspase inhibition completely inhibits
erythroid differentiation at the basophilic erythroblast stage (Carlile et al., 2004; Zermati

General Introduction
21
et al., 2001). Caspase activation during enucleation is associated with the cleavage of
lamin B and acinus, which are necessary for the maintenance of nuclear integrity and
condensed chromatin, respectively (Zermati et al., 2001). Finally, during the
differentiation of megakaryocytes into mature platelets, cytoplasmic fragmentation of
megakaryocytes leads to the formation of enucleate cells, rather than removal of nucleus,
per se (Radley and Scurfield, 1980). Interestingly, activated caspase-3 was detected in
maturing megakaryocytes, while the pan-caspase inhibitor z-VAD.fmk, as well as more
specific inhibitors of caspase-3 and -9, blocked platelet differentiation (De Botton et al.,
2002).
Localized caspase activity has been also found to play an important role during
sperm development. Dramatic removal of bulk cytoplasm occurs in the terminal
differentiation of spermatid into individual sperm. Multiple fly caspases have been
implicated in this highly specialized cellular event, known as spermatid individualization.
It was found that, during this process, caspase-3 is detected in the individualization
complex, a cytoskeletal membrane complex that moves along the length of the cyst to the
sperm tail (Arama et al., 2003).
The function of caspase-3 in cell differentiation may extend beyond terminal
differentiation. In the CNS, non-apoptotic active caspase-3 expression has been located
in proliferating and differentiating neuronal cells of the VZ and external granular layer of
the developing cerebellar cortex (Oomman et al., 2006). In addition, caspase-3 activation
is also observed in glial cells within the developing cerebellum.
In the past decade, the cell autonomous pro-differentiation effects of caspase-3 have
been noted in diverse cell types including neurons (Rohn et al., 2004), neural and glial
progenitor cells (Fernando et al., 2005; Oomman et al., 2006), and also osteoclasts (Mogi
and Togari, 2003), bone marrow stromal cells (Miura et al., 2004), and skeletal muscle
cells (Fernando et al., 2002). Chemical or genetic blockade of caspase-3 activity leads to
inhibition of differentiation.
Two recent studies implicate caspase-3 in controlling fate of both ES cells and adult
hematopoietic stem cells. One study reported caspase-induced cleavage of Nanog in
differentiating ES cells (Fujita et al., 2008). Unsurprisingly, stem cells lacking the
Caspase-3 gene showed marked defects in differentiation, while forced expression of
caspase cleavage-resistant Nanog mutant in ES cells strongly promoted self-renewal.
This suggests that Nanog is the key caspase substrate for limiting the self-renewing
capacity of ES cells. Curiously, caspase-3 was show to contribute to stem cell

Chapter 1
22
quiescence, by dampening specific signaling events. Alteration in caspase-3 expression
perturbs homeostasis of primitive hematopoietic cells (Janzen et al., 2008). Caspase-3-
deficient stem cells displayed accelerated proliferation and retarded differentiation. At
the molecular level, caspase-3 altered the sensitivity of primary hematopoietic cells to
cytokine stimulation through modulation of specific signaling pathways, such as
extracellular signal-related kinase (ERK) signaling. These data indicate a novel role for
caspase-3 as a governor, downregulating stem cell responsiveness to environmental
signals.
Interestingly, caspase-3 together with caspase-8 was recently shown to be involved
in somatic reprogramming. Induction of iPS cells by transduction of Oct-4, a key iPS
transcription factor, is associated with caspase-3 and -8 activation (Li et al., 2010a).
Inhibition of both caspases in human fibrobast cells prevented the induction of iPS cells,
suggesting that caspase-3 and -8 may play a role in the reprogramming process.
Finally, caspases are involved in dendritic pruning and synaptic plasticity. Two
studies indicate that localized caspase activity is important for the selective removal of
sensory neuron dendrites during fly metamorphosis (Kuo et al., 2006; Williams et al.,
2006). The involvement of caspases in dendritic pruning suggests that caspase activity is
specific to cellular compartments, and does not cause cell death. Furthermore, caspase-3
activity was shown to be regulated at postsynaptic sites in the brain following stimuli
associated with memory (Huesmann and Clayton, 2006). In fact, caspase-3 activity is
necessary to consolidate a persistent physiological trace of the song stimulus.
1.2.2.2 Caspase-8 and -14
Caspase-8 is involved in differentiation of macrophages. Caspase-8 deletion in
cells of the myelomonocytic lineages led to arrest of differentiation into macrophages
(Kang et al., 2004). In fact, caspase-8 activation is observed during differentiation of
monocytes into macrophages, but not in dendritic cells (Netea et al., 2008; Sordet et al.,
2002). An attempt to identify possible caspase substrates during macrophage
differentiation was made, and several proteins associated with cytoskeleton
rearrangements were identified (Cathelin et al., 2006). Expression and activity of
caspase-8 are also necessary for the specialized cell fusion events that are necessary for
the differentiation of human placental villous trophoblas (Black et al., 2004).

General Introduction
23
Caspase-14 is implicated in terminal differentiation of keratinocytes into enucleate
cells that form the skin outer layer. Caspase-14 expression is highly specific for
keratinocytes during the enucleation process (Lippens et al., 2000). Caspase-14-deficient
mice present a unique phenotype of shiny and lichenified skin (Denecker et al., 2007).
Although it was previously though that caspase-14 lacked protease activity (Van de Craen
et al., 1998), caspase-14 was found to directly cleave filaggrin, an abundant protein in the
epidermis, in vitro (Denecker et al., 2007). These findings indicate that caspase-14 plays
a surprising role in maintaining the integrity of skin, although it is uncertain whether it
directly regulates the enucleation process of keratinocytes.
1.2.2.3 Death versus differentiation phenotypes
It is still unclear how differentiating cells restrain active caspases without
compromising cellular integrity. One explanation is that the choice between death and
differentiation derives largely from caspase targeting and activation of specific substrates
or cofactors that are unique to each event. In this regard, it was shown that during
differentiation, but not during apoptosis, the chaperone protein Hsp70 protects the
transcription factor GATA-1, a master regulator of erythroid maturation, from caspase-
mediated proteolysis during the enucleation process of erythroids (Ribeil et al., 2007). It
is unclear, however, whether the selective masking of caspase substrates by Hsp70
provides a universal control mechanism for substrate specificity. Clearly, a systematic
investigation of each caspase substrate is needed to clarify whether these proteins
maintain a regulatory role in both apoptosis and cell differentiation.
In the absence of substrate selectivity, a plausible explanation for death versus
differentiation phenotypes may originate with the timing and intensity of signal pathway
activation. Specifically, the degree of caspase activity required to effectively act on
differentiation signals was lower than that observed to induce apoptosis (Weber and
Menko, 2005). In addition to temporal kinetics, the final choice between death and
differentiation may also depend on subcellular localization of activated caspases and their
constituent regulatory pathways. Finally, negative control by inhibitory proteins of the
inhibitor of apoptosis protein (IAP) family (Plenchette et al., 2004) may account for the
maintenance of cell survival during caspase-mediated differentiation.

Chapter 1
24
1.2.3 Bcl-2 family members
In vertebrates, the Bcl-2 family regulates the mitochondrial pathway of apoptosis by
complex interactions that dictate the integrity of the outer mitochondrial membrane
(Green and Evan, 2002). Bcl-2 family includes over 20 members that are functionally
classified as either anti-apoptotic or pro-apoptotic, and are grouped into three subfamilies
based on the number of homology (BH) domains they share. Most cells express a variety
of anti-apoptotic and pro-apoptotic Bcl-2 proteins, and the regulation of their interactions
dictates survival or commitment to apoptosis.
The anti-apoptotic subfamily of proteins includes Bcl-2, Bcl-xL, Bcl-w, Mcl-1 and
A1/Bfl-1, which possess four BH domains (BH1-4). The next two subfamilies are pro-
apoptotic proteins with three BH (BH1-3) domains, represented by Bax, Bak and Bok,
and BH3-only proteins, which are characterized by the presence of only the BH3 domain,
and include Bik, Egl-1, Bim, Bmf, Noxa, Bid, Bad, Bnip3, and Beclin-1 (Lomonosova
and Chinnadurai, 2008). Importantly, these BH domains serve as mediators for
protein:protein interactions, forming either homo- or heterodimers.
Data implicating Bcl-2 family members during differentiation are scarce and mainly
involve antiapoptotic Bcl-2 and Bcl-xL.
1.2.3.1 Bcl-2
Bcl-2 deficient mice are viable and no severe abnormality in the neuronal system
was encountered (Kamada et al., 1995; Veis et al., 1993). In the peripheral nervous
system, however, more direct evidence from the Bcl-2 knockout mice indicates that Bcl-2
accelerates the differentiation of trigerminal ganglionic neurons (Middleton et al., 1998).
Intriguingly, immunohistochemistry of human and mouse embryonic and fetal tissues has
disclosed high expression of Bcl-2 in organ systems undergoing morphogenesis and
differentiation (LeBrun et al., 1993; Lu et al., 1993), including neural cells of the
developing brain (Abe-Dohmae et al., 1993; Merry et al., 1994). Furthermore, Bcl-2
proteins in the nervous system are subject to developmental regulation, with high
expression levels in neuroepithelial cells of the ventricular zones, and in post-mitotic
neurons before adulthood (Merry et al., 1994). Therefore, it has been suggested that Bcl-
2 might be important, though not crucial, during neuronal cell differentiation. Bcl-2 may
be required for a more effective differentiation during the limited term in embryonic and
post-embryonic development.

General Introduction
25
In vitro, Bcl-2 expression was associated with neural differentiation of human
neural-crest-derived tumor cell line, Paju (Zhang et al., 1996), and with retinoic acid-
mediated differentiation of CNS neurons (Wang et al., 2001). This is in agreement with
the observation that Bcl-2 gene is induced by retinoic acid (Niizuma et al., 2006). In
PC12 cells, Bcl-2 was shown to induce differentiation as a result of prevention of cell
death induced by serum free medium (Sato et al., 1994). In the absence of Bcl-2, NGF-
mediated differentiation resulted in neurite elongation, whereas Bcl-2 overexpression
accelerated neurite formation (Suzuki and Tsutomi, 1998). Importantly, Bcl-2
overexpression was reported to increase neurite extension, but differentiation markers
were not affected (Eom et al., 2004).
Bcl-2 may be a direct effector of neuronal differentiation by influencing MAPK
signaling pathways. It was recently shown that Bcl-2 gene expression is directly
correlated with p38α activation, and triggers the formation of neuronal networks when
overexpressed in a global p38α-repressed environment (Trouillas et al., 2008).
Furthermore, specific activation of c-Jun N-terminal kinase (JNK) was involved during
Bcl-2 mediated neurite extention of MN9D dopaminergic neuronal cells overexpressing
Bcl-2 (Eom et al., 2004).
Taken together, these data indicate that Bcl-2 may act as an intercellular factor that
control neurite extension and axonal regeneration. Importantly, transplantation of ES
cells overexpressing Bcl-2 further increased the survival of transplanted ES cells, as well
as neuronal differentiation, and functional outcome (Wei et al., 2005). Thus,
manipulation of Bcl-2 should be valuable for improving transplantation efficacy of NS
cells.
1.2.3.2 Bcl-xL
Bcl-xL is the most potent antiapoptotic protein among Bcl-2 family members, both
in vitro and in vivo (Gonzalez-Garcia et al., 1995; Krajewska et al., 2002). More
specifically, Bcl-xL is essential for neuronal survival during brain development and in the
adult CNS (Motoyama et al., 1995; Parsadanian et al., 1998). Similar to Bcl-2, Bcl-xL
expression increases through the early developmental phase of the brain and peaks during
the neurogenesis period (Krajewska et al., 2002). Bcl-xL facilitates the in vitro
differentiation of both mouse and human ES cells into the neuronal lineage (Liste et al.,
2004; Shim et al., 2004). Also, recent data from conditional Bcl-xL mutant mice add

Chapter 1
26
support to these observations (Chang et al., 2007a; Savitt et al., 2005). Bcl-xL directs the
fate of both immortalized and naïve human NS cells and was found to enhance neuron
generation by increasing proliferation of neuronal progenitors, while decreasing glia
generation (Liste et al., 2007). These findings support the possibility that the
determination of neuronal versus astroglial lineages may be actively regulated by
balanced levels of anti- and pro-apoptotic Bcl-2 family proteins in neural precursors
(Chang et al., 2007a). In fact, Chang and coworkers found that Bcl-xL instructively
directs the fate of embryonic cortical precursors toward neurons (Chang et al., 2007a).
Finally, the expression of Bcl-xL has been shown to be down-regulated during
differentiation of human hematopoietic progenitor cells along the granulocyte lineage, but
increased during differentiation into monocytes/macrophages (Sanz et al., 1997). In
addition, Bcl-xL increased along the erythroid differentiation of progenitor cells (Gregoli
and Bondurant, 1997) and in K562 cell differentiation into megakaryocyte (Terui et al.,
1998).
1.2.3.3 Bax and Bim
Scarce data is available on the role of Bax during neural differentiation. In fact,
during neurodevelopment, Bax expression is coincident with that of astrocyte formation
(Sauvageot and Stiles, 2002), and declines during postnatal development (Krajewska et
al., 2002). In accordance, Bax was shown to instruct embryonic cortical precursors to
undergo astrocytic differentiation (Chang et al., 2007a). Bax has no effect on neuronal
differentiation, which is supported by the observation that Bax levels decreased or
remained unchanged in differentiating PC12 cells (Vekrellis et al., 1997) and retinoic
acid-mediated differentiation of NTera-2/DI cells (Guillemain et al., 2000), respectively.
The BH3-only protein Bim was recently added to the list of pro-apoptotic proteins
with a function in cell differentiation. Using gene-deficiency mice, Bunk and coworkers
reported a possible role of pro-apoptotic Bim in neuronal differentiation of adult-born
neural precursor cells (Bunk et al., 2010). Despite bim and puma deficiency did not
change early markers of neuronal differentiation in dentate gyrus, BrdU/NeuN double-
labeling revealed that the deficiency of bim, but not puma, accelerated the differentiation
of newly generated cells into a neuronal phenotype. In apoptotic context, Bim binds to
and “neutralizes” Bcl-2. It is therefore possible that endogenous Bim levels determine the

General Introduction
27
amount of sequestered Bcl-2, and by doing so influence neuronal differentiation or axonal
outgrowth.
1.2.4 Others apoptotic players in cell differentiation
Death receptors are cell-surface receptors belonging to the tumor necrosis factor
(TNF) superfamily that includes CD95 (FAS/APO-1), TNF-receptor type I (TNFR1) and
tumor necrosis factor-related apoptosis inducing ligand (TRAIL) receptors. TNF is a
major mediator of apoptosis as well as inflammation and immunity, and it has been
implicated in the pathogenesis of a wide spectrum of human diseases, including cancer
(Wang et al., 2009) and autoimmune diseases (Zapata et al., 2009). TNF binds to two
specific receptors, TNFR1 and TNFR2 which differ in structure and function. A death
domain is present only in TNFR1, and thus, TNFR2 activation does not directly lead to
caspase activation. TNFR2 is believed to initiate primarily pro-inflammatory and pro-
survial signaling. On the other hand, TNFR1 activation leads to recruitment of
intracellular adaptor proteins that activate multiple signal transduction pathways (Tracey
et al., 2008). Recently, members of the TNF family have emerged as apoptosis-
associated proteins that operate in differentiation-related processes.
Current experiments suggest that lower concentrations of TNF can elicit myoblast
differentiation and engage behavior associated activated muscle stem cells, such as
enhanced chemotaxis (Chen et al., 2005; Torrente et al., 2003). Furthermore, recent
evidence also suggests that components of the TNF/TRAIL dependent pathways stimulate
differentiation in a variety of cell types, including dendritic and human intestinal cells
(Chomarat et al., 2003; Rimondi et al., 2006). In addition, TNF-α was recently shown to
have a pro-neurogenic effect in neonatal SVZ cell cultures of mice, which also involved
TNFR1 activation (Bernardino et al., 2008).
Stimulation of the death receptor, CD95 in NS cell does not trigger apoptosis but
unexpectedly leads to stem cell survival and neuronal specification (Corsini et al., 2009).
These effects are mediated via activation of the Src and phosphatidylinositol 3-kinase
(PI3K)-Akt-mTOR signaling pathway, ultimately leading to a global increase in protein
translation.

Chapter 1
28
1.3 microRNAs
The recent discovery of microRNAs (miRNAs or miRs) introduces a novel
regulatory control over gene expression in several processes, including neuronal
differentiation. miRNAs comprise a large family of single stranded non-coding RNAs, ~
21-nucleotide-long that have emerged as key post-transcriptional regulators of gene
expression in metazoans and plants (Bartel, 2009). miRNAs undergo several processing
steps until the mature miRNA is produced and exerts its biological role. Many miRNAs
tend to be in close genomic proximity, accounting for about 37% of miRNAs transcribed
as polycistronic transcripts (Altuvia et al., 2005). Transcription of miRNA genes is
regulated in a similar manner to that of protein-coding genes, and is a major level of
control responsible for tissue-specific or development-specific expression of miRNAs
(Krol et al., 2010).
In mammals, miRNAs are predicted to control the activity of ~ 50% of all protein-
coding genes (for review, see Krol et al., 2010). Functional studies indicate that miRNAs
participate in the regulation of almost every cellular process investigated so far, and that
changes in miRNA expression are associated with many human pathologies. Indeed,
stem cell and miRNA fields have converged with the identification of stem cell-specific
and lineage-specific miRNAs (Gangaraju and Lin, 2009). Owing to their functions in the
regulation of translation, miRNAs can regulate stem cell fate and behavior by fine-tuning
the protein levels of various factors required for stem cell proliferation and
differentiation.
1.3.1 Overview of miRNA biogenesis and function
The majority of miRNAs are transcribed from independent genes by RNA
polymerase II, giving rise to a 5’ 7-methyl-guanosine capped and 3’ polyadenilated
primary miRNA (pri-miRNAs) (Cai et al., 2004) (Fig. 1.6). Pri-miRNA transcripts can
originate from intergenic regions, where the miRNA can reside within an exon, or an
intron of a non-coding transcript. However, many mammalian miRNAs reside in introns
of protein coding genes, and display a similar transcription pattern to that of protein-
coding genes (Baskerville and Bartel, 2005; Rodriguez et al., 2004).
pri-miRNAs present a typical hairpin structure, and act as substrates for two
members of the RNase III family of enzymes, Drosha and Dicer. The product of Drosha
cleavage, an ~ 70-nucleotide precursor miRNA (pre-miRNA), is exported to the

General Introduction
29
cytoplasm, where Dicer processes it to an ~ 20-bp miRNA duplex. One strand of this
duplex, the guide strand, is then incorporated into the miRNA-induced silencing complex
(RISC). As part of the RISC, miRNAs basepair to target mRNAs and induce their
translational repression of deadenylation and degradation. The Argonaute (AGO) protein,
which directly interact with miRNAs, and glycine-tryptophan protein of 182 kDa (GW
182) proteins, which act as downstream effectors in the repression, are key factors in the
assembly and function of RISC. Furthermore, the role of both Drosha and Dicer in
miRNA maturation is assisted by a number of cofactors or accessory proteins, with some
playing an important regulatory function (Krol et al., 2010).
Figure 1.6. Schematic overview of miRNA biogenesis. miRNAs are transcribed as primary
transcripts (pri-miRNA) by RNA polymerase II. Each miRNA is cleaved in two steps, catalyzed

Chapter 1
30
by two members of the RNase III family of enzymes, Drosha and Dicer, operating together with
specific cofactors, such as DiGeorge syndrome critical region gene 8 (DGCR8) and
transactivation-responsive (TAR) RNA-binding protein (TRBP) in mammals. In the first nuclear
step, the Drosha-DGCR8 complex processes pri-miRNA into a ~ 70-nucleotide stem loop known
as the precursor miRNA (pre-miRNA), which is exported to the cytoplasm by exportin 5.
Cleavage of pre-miRNA by Dicer, assisted by TRBP, yields an ~ 20-bp mature miRNA duplex in
the cytoplasm. Following processing, the guide strand of the duplex is preferentially incorporated
into a miRNA-induced silencing complex (RISC), whereas the other strand (passenger) is
released and degraded (not shown). Mature miRNA allows the RISC to recognize target mRNA
through partial sequence complementarity with its target. The target mRNA can be either cleaved
by AGO2, or deadelylated, decapped and exonucleolyticaly degraded. Adapted from Inui et al.
2010.
Recent discoveries have revealed a model in which complex miRNA regulatory
events are woven into the known transcription factor and signaling networks that control
cell fate and differentiation, modulating their activity through positive and negative
feedback loops to reinforce cellular decisions. The important regulatory role of miRNAs
in development and differentiation clearly emerged from the study of ES cells null for the
Dicer gene. Ablation of Dicer leads to defects in embryonic stem cell division and
proliferation (Murchison et al., 2005), causing death in mice and complete loss of
pluripotent stem cells (Bernstein et al., 2003). In addition, Dicer ablated embryonic stem
cells also fail to differentiate. Therefore, miRNAs can affect stem cell behavior at both
proliferation and differentiation levels.
1.3.2 Apoptosis-associated miRNAs as regulators of differentiation and cell fate
decision
Based on the capacity of miRNAs to regulate pro- and antiapoptotic genes,
Garofalo et al. recently classified miR-15 and miR-16, let-7 family, members of miR-34
family, miR-122, miR-101 and miR-29 family members as pro-apoptotic miRNAs
(Garofalo et al., 2010). Not surprisingly, some of these miRNAs are reported to be
involved in cell differentiation.

General Introduction
31
1.3.2.1 miR15 and miR-16
miR-15a and miR-16-1 are deleted or downregulated in the majority of chronic
lymphocytic leukemias, and they are known to negatively regulate Bcl-2 expression
levels (Cimmino et al., 2005). Interestingly, miR-15a and miR-16 were reported to
decrease considerably during human monocyte-macrophage differentiation (Li et al.,
2010b). In addition, miR-15a was shown to play a role during hematopoietic
differentiation (Zhao et al., 2009). Furthermore, miRNA profile in mouse inner ear
tissues identified miR-15a-1 as having a differential expression profile during
development of mouse inner ear tissue (Friedman et al., 2009), which was also observed
during zebrafish inner ear development and morphogenesis. Interestingly, both miR-15a
and miR-16-1 were upregulated in retinoic acid-mediated granulocytic differentiation of
NB4 cells (Garzon et al., 2008). These results are correlated with downregulation of Bcl-
2 mRNA and protein levels during retinoic acid treatment. Importantly, the miR-15a/16-
1 cluster also downregulates expression of cyclin dependent kinase 6 (CDK6), caspase
recruitment domain-containing protein 10 (CARD10), and cell division cycle protein 27
(CDC27), preventing cells from entering S phase, which can also contribute to a role in
cell differentiation (Ivey and Srivastava, 2010).
1.3.2.2 Let-7 family
The lethal-7 (let-7) gene was initially discovered in C. elegans (Reinhart et al.,
2000). let-7 and its family members are highly conserved across species, from worms to
humans, in sequence and function (Pasquinelli et al., 2000). Misregulation of let-7 leads
to a less differentiated cellular state and the development of cell-based diseases, such as
cancer. The size of the let-7-family can differ between organisms; in invertebrates is
smaller, only nine related family members in nematodes and one let-7 in fly (Lagos-
Quintana et al., 2001), compared with 13 closely related let-7 family genes in humans
(Roush and Slack, 2008).
The major role of let-7 is to promote differentiation of cells. It was shown to
regulate stem cell differentiation in C. elegans (Reinhart et al., 2000), neuromusculature
development and adult behavior in flies (Caygill and Johnston, 2008; Sokol et al., 2008),
limb development in chicken and mouse (Lancman et al., 2005; Schulman et al., 2005),
and cell proliferation and differentiation (Wulczyn et al., 2007; Yu et al., 2007a; Yu et al.,

Chapter 1
32
2007b). Still, let-7 was reported to be strongly induced during neural differentiation of
ES cells and embryocarcinoma cells (Rybak et al., 2008; Wulczyn et al., 2007).
In mammals, let-7 levels increase during embryogenesis and during brain
development (Schulman et al., 2005; Wulczyn et al., 2007). Let-7 is undetectable in
human and mouse embryonic stem cells, and increases following differentiation
(Wulczyn et al., 2007). This high expression of let-7 is then maintained in various adult
tissues (Sempere et al., 2002). However, the direct contribution of let-7 in development
has not been demonstrated. It was recently shown that the introduction of let-7 miRNAs
can suppress the self-renewal in Dgrc8-/- mouse ES cells (Melton et al., 2010).
Furthermore, inhibition of the let-7 family promotes de-differentiation of somatic cells
into iPS cells. Together, these findings suggest that let-7 miRNAs act in order to stabilize
the self-renewing versus differentiated cell fates.
Differentiation is tightly linked with exit from cell cycle and numerous genes whose
products promote G1/S or G2/M transitions, including CDK6, CDC25A, and CCND2, are
direct let-7 targets (Ivey and Srivastava, 2010). In addition, let-7 indirectly influences
cell cycle by negatively regulating oncogenes such as MYC (Sampson et al., 2007), RAS
(Johnson et al., 2005) and high-mobility group AT-hook 2 HGMA2 (Park et al., 2007),
which would otherwise promote proliferation. Caspase-3 was also reported to be a direct
let-7 target (Tsang and Kwok, 2008). However, the relative importance of these targeting
in a differentiation context is unknown.
1.3.2.3 miR-34 family
The miRNA-34 family comprises three members in mammalians; miR-34a, which
is generated from a larger transcriptional unit on chromosome 1p36, and miR-34b and
miR-34c, both of which are generated by processing of a bicistronic transcript from
chromosome 11q23 (for a review, see Hermeking, 2010). In mice, miR-34a is
ubiquitously expressed with the highest expression in the brain (Lodygin et al., 2008),
whereas miR-34b/c is mainly expressed in lung tissues (Bommer et al., 2007).
Importantly, several reports showed that members of the miR-34 family are direct p53
targets, and their upregulation induces apoptosis and cell cycle arrest (Bommer et al.,
2007; Chang et al., 2007b; Corney et al., 2007; He et al., 2007a; Raver-Shapira et al.,
2007; Tarasov et al., 2007) (Fig. 1.7).

General Introduction
33
Figure 1.7. Involvement of members of miR-34 family in p53 network. Under genotoxic
stress, p53 is activated by upstream kinases and transactivates several target genes, including
miRNA-34 family members. After processing, mature miR-34a, -b, and -c mediates inhibition of
translation or RNA degradation of the indicated validated, and presumably many others not yet
confirmed targets, leading to apoptosis, cell cycle arrest, senescence or inhibition of migration.
Adapted from Hermeking, 2010.
Despite the involvement of miR-34 family members in mediating p53 effect on
apoptosis and cell cycle, emerging evidence suggests a role for miR-34a during cell
differentiation. miR-34a was shown to be involved in monocyte-derived dendritic cell
differentiation (Hashimi et al., 2009), megakaryocytic differentiation of K562 cells
(Ichimura et al., ; Navarro et al., 2009), and differentiation of mouse embryonic stem cells
(Tarantino et al.). In addition, targets of miR-34a were distinct in each differentiation
context and include JAG1, CDK4 and CDK6, MYB, mitogen-activated protein kinase
kinase 1 (MEK1) and silent information regulator 1 (SIRT1). Interestingly, the induction
of miR-34a during megakaryocytic differentiation of K562 cells was shown to be
independent of p53 (Navarro et al., 2009). However, the same attempt to analyze a
possible link between p53 and miR-34a expression in a differentiation context was not
performed in the others studies and therefore the hypothesis that miR-34a mediates p53
effect in a differentiation context deserves further investigation.
1.3.2.4 miR-122, miR-101, and miR-29 family
miR-122 is a highly abundant, liver-specific miRNA whose expression is
specifically turned on in the mouse liver during embryogenesis (Chang et al., 2004).

Chapter 1
34
Therefore, it is expected to be involved in liver development. In addition, Lin et al.
demonstrated that the anti-apoptotic Bcl-w is a direct target of miR-122 (Lin et al., 2008).
Recently, miR-122 was shown to contribute to liver development by regulating the
balance between proliferation and differentiation of hepatocytes, at least in part by
targeting CUTL1, a transcriptional repressor of genes specifying terminal differentiation
(Xu et al., 2010).
miR-101 was suggested to have pro-apoptotic functions by targeting Mcl-1 (Su et
al., 2009). The only link between miR-101 expression and cell differentiation came from
the study of Garzon and coworkers that showed an upregulation of miR-101 during
megakaryocyte differentiation (Garzon et al., 2006).
The miR-29 family of miRNAs consists of miR-29a, -29b, and -29c. In addition,
Mcl-1 was also confirmed to be a target of miR-29b (Mott et al., 2007). Interestingly,
miR-29b was suggested to be a key regulator of development of the osteoclast phenotype
by targeting anti-osteogenic factors and modulating bone extracellular matrix proteins (Li
et al., 2009b). The other members of miR-29 family, miR-29a and miR-29c, are also
upregulated during osteoblastic differentiation in vitro, possibly induced by Wnt signaling
(Kapinas et al., 2009). In addition, miR-29 was reported to be more expressed in
astrocytes than neurons (Smirnova et al., 2005), and was also shown to accelerate
myogenesis (Wang et al., 2008a).
Although the above data suggest a possible role for apoptosis-associated miRNAs
in cell differentiation, it is important to further explore the mechanism(s) by which
apoptosis-associated miRNAs regulate cell differentiation, adding to our understanding
the potential of miRNAs to define the fate of developing cells.

35
Objectives
The studies presented in this thesis were driven by the hypothesis that specific
apoptosis-associated signaling pathways play a critical role in NS differentiation. Three
main objectives were designed. First, our goal was to determine the involvement of
specific apoptosis-associated proteins in mouse NS cell differentiation, and investigate
their interaction with proliferation to commit cells to differentiation. Second, we sought
to evaluate the potential involvement of apoptosis-associated miRNAs in different models
of neural differentiation. Finally, we aimed to further explore the role of miR-34a, an
apoptosis-associated miRNA whose expression is induced by p53, during NS cell
differentiation. Specifically, we wanted to identify putative targets of miR-34a, and
further clarify the mechanism(s) by which miR-34a regulates cell differentiation.
The specific questions addressed in this thesis are:
1. Are apoptosis-associated proteins integral components of the differentiation
process of NS cells? Do they interfere with proliferation pathways, such as
the PI3K/Akt signaling pathway?
2. Are apoptosis-associated miRNAs involved in the differentiation process of
neural cells?
3. What is the role of miR-34a expression during NS cell differentiation?
Which molecular pathways does it modulate?
The overarching goal of the research presented in this thesis is to increase our
understanding regarding the basic molecular mechanisms of NS cells differentiation that
require specific factors of the apoptosis machinery. The results may have broad
implications in advancing our understanding of differentiation and its modulation.
Further, our findings may prove useful in the development of novel therapeutic strategies
to improve long term-survival and differentiation of transplanted stem cells.


2 Caspases and p53 modulate FOXO3A/Id1 signaling during mouse neural stem cell
differentiation
Márcia M. Aranha1, Susana Solá1, Walter C. Low2,3, Clifford J. Steer4,
Cecília M. P. Rodrigues1*
1iMed.UL, Faculty of Pharmacy, University of Lisbon, Lisbon, Portugal; 2,3Department of
Neurosurgery, and Stem Cell Institute, University of Minnesota Medical School, Minneapolis,
Minnesota, USA; 4Department of Medicine, and Genetics, Cell Biology, and Development,
University of Minnesota Medical School, Minneapolis, Minnesota, USA
Journal of Cellular Biochemistry 2009; 107 (4): 748-758

Reprinted from Journal of Cellular Biochemistry, vol 107, Issue 4, Márcia M. Aranha,
Susana Solá, Walter C. Low, Clifford J. Steer, Cecília M. P. Rodrigues. Caspases and
p53 modulate FOXO3A/Id1 signaling during mouse neural stem cell differentiation,
pages 748-758, Copyright 2009, with permission from John Wiley and Sons. All rights
reserved.

Apoptotic machinery in differentiation
39
2.1 Abstract
Neural stem cells (NSCs) differentiate into neurons and glia, and a large percentage
undergoes apoptosis. The engagement and activity of apoptotic pathways may favor
either cell death or differentiation. In addition, Akt represses differentiation by up-
regulating the inhibitor of differentiation 1 (Id1), through phosphorylation of its repressor
FOXO3A. The aim of this study was to investigate the potential cross-talk between
apoptosis and proliferation during mouse NSC differentiation. We determined the time of
neurogenesis and gliogenesis using neuronal β-III tubulin and astroglial GFAP to confirm
that neurogenesis and gliogenesis occur at ~ 3 and 8 days, respectively. p-Akt, p-
FOXO3A, and Id1 were significantly reduced throughout differentiation. Caspase-3
processing, p53 phosphorylation, and p53 transcriptional activation increased at three
days of differentiation, with no evidence of apoptosis. Importantly, in cells exposed to
the pancaspase inhibitor z-VAD.fmk, p-FOXO3A and Id1 were no longer down-
regulated, p53 phosphorylation and transcriptional activation were reduced, while
neurogenesis and gliogenesis were significantly delayed. The effect of siRNA-mediated
silencing of p53 on FOXO3A/Id1 was similar to that of z-VAD.fmk only at 3 days of
differentiation. Interestingly, caspase inhibition further increased the effect of p53
knockdown during neurogenesis. In conclusion, apoptosis-associated factors such as
caspases and p53 temporally modulate FOXO3A/Id1 signaling and differentiation of
mouse NSCs.

Chapter2
40
2.2 Introduction
Neural stem cells (NSCs) are defined by their capacity for self-renewal and ability
to generate major cell types of the mammalian central nervous system (Doe, 2008).
These features make them a therapeutic tool to treat a broad spectrum of human
neurological disorders, including Parkinson’s, Alzheimer’s, and Huntington’s diseases as
well as acute stroke (Schwarz et al., 2006). Despite the therapeutic potential of NSCs,
major hurdles must be overcome to render these cells a reliable and efficient system to
produce specific cell types for neuro-replacement therapies (Hsu et al., 2007). Tight
regulation of the differentiation process remains a critical issue. In fact, the mechanisms
controlling differentiation and lineage specification of NSCs are still poorly understood,
and many molecules and their specific functions are yet unknown.
Apoptosis is often thought to result from a failure of cells to correctly exit the cell
cycle and differentiate. In this respect, it has been postulated that apoptotic pathways
may also regulate other basic cell functions that do not involve cell death per se (Garrido
and Kroemer, 2004). It was recently suggested that several conserved elements of
apoptosis are also integral components of terminal differentiation. Therefore, it is not
surprising that during differentiation, specific cellular changes occur in a similar manner
to those observed during apoptosis (Chasis and Schrier, 1989). Terminal erythroid
differentiation, for example, is characterized by major morphological changes that include
chromatin condensation and complete removal of the nucleus (Garrido and Kroemer,
2004). In addition, it has been demonstrated that the fate of erythroid precursors is
determined downstream of caspase activation by the pattern of cleaved targets (Ribeil et
al., 2005).
Caspases have recently been implicated in the differentiation process for a variety
of model systems (Schwerk and Schulze-Osthoff, 2003). Specifically, caspase-8 was
shown to be required for T cell activation (Misra et al., 2007), while bone morphogenetic
protein-induced differentiation of osteoblastic cells has been associated with transient and
potent activation of caspase-8, -2, and -3 (Mogi and Togari, 2003). In addition, it has
been demonstrated that NSC differentiation depends on endogenous caspase-3 activity
that may influence the kinase activity associated with changes in phenotype (Fernando et
al., 2005). The time of engagement and activity level of these pathways is thought to
ultimately determine the choice between cell death or maturation (Fernando and

Apoptotic machinery in differentiation
41
Megeney, 2007). Nevertheless, the precise role of caspase-3 in NSC differentiation is
still largely unknown.
Curiously, widespread neurogenesis and gliogenesis occur in the adult mammalian
nervous system in response to certain stress conditions, including hypoxia (Magavi et al.,
2000). The tumor suppressor p53 is known to regulate key cellular functions involved in
differentiation. p53 is activated by cellular stress, hyperproliferative signals, and
developmental stimuli. One well characterized outcome of p53 activation is irreversible
cell cycle exit required for either apoptosis, senescence, or differentiation (Stiewe, 2007).
In fact, p53 was shown to contribute to differentiation of mouse embryonic stem cells by
suppressing expression of Nanog, which is known to maintain self-renewal and the
undifferentiated state of stem cells (Lin et al., 2005). In addition, reduced p53 expression
in human embryonic stem cells has been shown to decrease spontaneous differentiation
and slow differentiation (Qin et al., 2007). Other studies have shown that p53 is required
for differentiation of PC12 cells (Zhang et al., 2006), myoblasts (Porrello et al., 2000),
and epithelial cells (Saifudeen et al., 2002).
Interestingly, p53 may contribute to differentiation by limiting self-renewal and the
undifferentiated state of cells through inactivation of the Akt-mediated phosphoinositide
3-kinase (PI3K) survival pathway. In fact, p53 may inhibit the Akt proliferation pathway
by modulating important downstream targets, such as the lipid phosphatase and tensin
homolog deleted from chromosome 10 (PTEN) (Stambolic et al., 2001). Moreover,
differentiation of PC12 cells has been associated with enhanced expression of
proapoptotic molecules and reduced activation of Akt survival signaling (Mielke and
Herdegen, 2002). Finally, Akt represses cellular differentiation by up-regulating the
inhibitor of differentiation 1 (Id1), through phosphorylation of its repressor FOXO3A.
Akt regulates the sub-cellular localization of FOXO3A by phosphorylation, thereby
preventing its nuclear translocation and inhibition of Id1 (Birkenkamp et al., 2007).
Having provided a rationale for similarities between apoptosis and differentiation
cell machineries, the next challenge will be to further define and manipulate an operative
pathway common to each process. Half of all neurons produced during neurogenesis die
through apoptosis before maturation of the nervous system. This could be explained, at
least in part, by the high rate of cell death that occurs during the G1/S cell cycle
checkpoint (Liu and Greene, 2001). In this regard, it is crucial to understand which
apoptosis mechanism(s) overlaps with neuronal differentiation pathways that reduce cell
death and improve lineage specification for potential neuro-replacement therapies.

Chapter2
42
Here, we further investigate the role of caspases and p53 during mouse NSC
differentiation, and elucidate the cross-talk between apoptosis-associated elements and
the Akt survival pathway. Our data strongly indicate that caspases and p53 act in a
synergistic manner in suppressing p-FOXO3A/Id1 signaling and, thereby inducing
neurogenesis of NSCs.
2.3 Methods
2.3.1 Mouse NSC Culture and Differentiation
Mouse NSCs containing a constitutively expressed marker for green fluorescence
protein (GFP) were used to investigate the process of neuronal differentiation. Cells were
obtained from E14 mouse embryo central nervous system and cultured as described by
Rietze et al (Rietze and Reynolds, 2006) and Reynold and Weiss (Reynolds and Weiss,
1992). Mouse NSCs were maintained as neurospheres in undifferentiation medium,
serum-free, 1:1 mix of DMEM/F12 (Invitrogen Corp., Grand Island, NY) with 1× N-2
supplement (Invitrogen Corp.), 20 ng/ml EGF, 20 ng/ml b-FGF (R & D Systems Inc.,
Minneapolis, MN), and 1% penicillin-streptomycin (Invitrogen Corp.), at 37ºC in
humidified atmosphere of 5% CO2. Subculture occurred at day 7 with mechanical
dissociation of neurospheres. Cellular plating was performed at 5 × 104 cell/ml density
on T75 flasks and half of the culture medium was changed after 3 days. The
differentiation of mouse NSCs in vitro was induced by culturing dissociated cells in
differentiation medium containing DMEM/F12 with 1× N-2 supplement, 100 ng/ml b-
FGF, 10% FBS (Invitrogen Corp.), 500 nM all-trans retinoic acid (Sigma Chemical Co.,
St. Louis, MO), 50 µM taurine (Sigma Chemical Co.), 10 ng/ml TGF-β2 (R & D Systems
Inc.) and 1% penicillin-streptomycin in tissue culture plates pre-coated with poly-D-
lysine (Sigma Chemical Co.). The culture medium was changed every 3 days.
Differentiated cells at 5 × 104 cells/ml were fixed at days 3, 6, 8, 10, 12, 14 and 17 and
processed for immunostaining and evaluation of differentiation. Neurons and astrocytes
were obtained at 3 and 8 days, respectively, in differentiation medium. For western blot
analysis, cultures at 5 × 105 cells/ml were processed at days 1, 3, and 8 to assess the role
of apoptosis during both neuron and astrocyte differentiation. All experiments were
performed using adherent cells only to exclude detached apoptotic cells.

Apoptotic machinery in differentiation
43
2.3.2 Immunocytochemistry
Mouse NSCs were fixed with 4% paraformaldehyde in phosphate-buffered saline
(PBS) for 30 minutes during differentiation. Cells were then blocked for 1 hour at room
temperature in PBS containing 0.1% Triton-X-100, 1% FBS, and 10% normal donkey
serum (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Subsequently,
cells were incubated with either monoclonal antibodies to glial fibrillary acidic protein
(GFAP) (1:1000), neuronal nuclei (NeuN) (1:100) and oligodendrocyte marker O4
(1:100) (Chemicon Int., Temecula, CA) or the polyclonal antibody to β-III tubulin at a
dilution of 1:1000 (Covance, Princeton, NJ) and to active caspase-3 (1:50) (R&D
Systems, Inc.) in blocking solution, overnight at 4ºC. After three washes with PBS, cells
were incubated with the AMCA-anti-mouse (Invitrogen Corp.) or the Alexa Fluor 594-
anti-rabbit conjugated secondary antibodies (Invitrogen Corp.) for 2 hour at room
temperature. The localization of GFAP, NeuN, O4, β-III tubulin, and caspase-3 were
visualized using an Axioskop fluorescence microscope (Carl Zeiss, Jena, Germany).
Total GFP-positive cells were counted on a computer screen grid from at least four
random fields (x400). Results were expressed as the average percentage of GFP-positive
cells, which co-labeled for β-III tubulin or GFAP.
2.3.3 Evaluation of Apoptosis
Hoechst labeling and the TUNEL staining of mouse NSCs were used to detect
apoptotic nuclei. In brief, for morphologic evaluation of apoptosis, the medium was
gently removed at the indicated times with minimal detachment of the cells. Attached
cells were fixed with 4% paraformaldehyde in PBS, pH 7.4, for 10 minutes at room
temperature, incubated with Hoechst dye 33258 (Sigma Chemical Co.) at 5 µg/ml in PBS
for 5 minutes, washed with PBS and mounted using PBS:glycerol (3:1, v/v). Fluorescent
nuclei were scored blindly and categorized according to the condensation and staining
characteristics of chromatin. Normal nuclei showed non-condensed chromatin dispersed
over the entire nucleus. Apoptotic nuclei were identified by condensed chromatin,
contiguous to the nuclear membrane, as well as nuclear fragmentation of condensed
chromatin. Three random microscopic fields per sample were counted and mean values
expressed as the percentage of apoptotic nuclei. For TUNEL staining, apoptotic cells
were quantitated using the deoxynucleotidyltransferase-mediated dUTP nick end labeling
(TUNEL) assay. Cells were fixed with 4% formaldehyde and processed using an

Chapter2
44
Apoptag in situ apoptosis detection kit (Chemicon Int., Temecula, CA). The number of
TUNEL-positive cells was counted on a computer screen grid from at least three random
fields (400x).
2.3.4 Total, Cytosolic, and Nuclear Protein Extraction
For total protein extracts, adherent mouse NSCs were lysed in ice-cold buffer (10
mM Tris-HCl, pH 7.6, 5 mM MgCl2, 1.5 mM KAc, 1% Nonidet P-40, 2 mM DTT, and
protease inhibitor cocktail tablets (Complete; Roche Applied Science, Mannheim,
Germany)) for 30 minutes, and then homogenized with 20 strokes in a loose fitting
Dounce. The lysate was centrifuged at 3200g for 10 minutes at 4ºC and the supernatant
recovered. For nuclear and cytosolic extracts, cells were lysed with hypotonic buffer (10
mM Tris-HCl, pH 7.6, 5 mM MgCl2, 1.5 mM KAc, 2 mM DTT, and protease inhibitors),
homogenized with 20 strokes in a loose fitting Dounce, and centrifuged at 500g for 10
minutes at 4ºC. The cytosolic proteins were recovered in the supernatant while the
nuclear pellet was washed in buffer containing 10 mM Tris-HCl, pH 7.6, 5 mM MgCl2,
0.25 M sucrose, 0.5% Triton X-100, and protease inhibitors, then resuspended and
sonicated in buffer containing 10 mM Tris-HCl, pH 7.6, 0.25 M sucrose with protease
inhibitors. Finally, the suspension was centrifuged through 0.88 M sucrose at 2000g for
20 minutes at 4ºC, and nuclear proteins were recovered in the supernatant.
2.3.5 Immunoblotting
Steady-state levels of p53, p-p53 (Ser15), p-Akt (Ser473), Akt1/2, p-FOXO3A
(Thr32), FOXO3A, and Id1 as well as caspase-3 processing were determined by Western
blot, using primary rabbit polyclonal antibodies reactive to p-p53 (Ser15) (Calbiochem,
Darmstadt, Germany), p-Akt (Ser473), total Akt, Id1 and caspase-3 (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), FOXO3A and p-FOXO3A (Thr32) (Chemicon
Int.) or primary mouse monoclonal antibody to p53 (Santa Cruz Biotechnology); as well
as secondary antibodies conjugated with horseradish peroxidase (Bio-Rad Laboratories,
Hercules, CA, USA). Membranes were processed for protein detection using Super
SignalTM substrate (Pierce, Rockford, IL, USA). β-actin was used to control for lane
loading while acetyl-histone H3 was used as a marker for nuclear protein extraction.
Protein concentrations were determined using the Bio-Rad protein assay kit according to
the manufacture’s specifications.

Apoptotic machinery in differentiation
45
2.3.6 p53-DNA Binding ELISA Assay
The Universal EZ-TFA p53 Transcription Factor Assay Chemiluminescent Kit
(Upstate-Millipore, Billerica, MA) was used according to the manufacturer’s protocol.
Nuclear extracts of mouse NSC cells were applied to plates containing biotinylated
oligonucleotides with a p53 consensus binding site (5`-
GGACATGCCCGGGCATGTCC-3`). After 1 hour incubation at room temperature,
plates were washed and incubated with diluted p53 antibody (1:1000) for an additional 1
hour. Diluted anti-rabbit horseradish peroxidase-conjugated antibody (1:500) was then
added to previously washed plates. The chemiluminescence substract solution was added
and incubated for 5-8 minutes to allow luminescence development. The reaction was
stopped and samples read at 1-second integration time in a microplate luminometer. A
p53 specific competitor oligonucleotide having the same consensus sequence as the p53
capture probe, but not biotinylated was used as negative control.
2.3.7 Inhibition of Caspase Activity
Mouse NSC were exposed to 50 µM of the pan-caspase inhibitor z-VAD.fmk
(Calbiochem, La Jolla, CA) 23 h after platting in differentiation medium, and then
collected after 1, 3, and 8 days. The culture medium supplemented with z-VAD.fmk was
changed after 3 days in culture. Total proteins were extracted for immunobloting.
Attached cells were fixed for Hoechst staining and immunocytochemistry analysis.
2.3.8 Short Interference RNA and Transfection
A pool of 4 short interference RNA (siRNA) nucleotides designed to knockdown
p53 gene expression in mouse was purchased from Dharmacon (Waltham, MA). A
control siRNA containing a scrambled sequence that does not result in degradation of any
known cellular mRNA was used as control. Twenty-four hours after platting, the culture
differentiation medium was changed to medium without 1% penicillin-streptomycin.
Mouse NSCs were transfected using Lipofectamine™ Transfection Reagent for siRNA
(Polyplus Transfections, Illkirch, France), according to the manufacturer’s instructions for
an additional 24 h. The final concentration of siRNAs was 100 nM. To assess gene
silencing, transcript and protein levels of p53 were determined by RT-PCR and Western

Chapter2
46
blot, respectively. Our results indicated that the silencing efficiency of the siRNAs for
p53 was ~ 90% in mouse NSC. To investigate the effect of p53 siRNA throughout the
differentiation process, attached cells treated or non-treated with z-VAD.fmk were either
harvested for immunoblot or used for immunocytochemistry assays after 3 and 8 days of
differentiation. Attached cells were also fixed for Hoechst staining.
2.3.9 RNA Isolation and RT-PCR
Transcript expression of p53 was determined by RT-PCR. Total RNA was
extracted from mouse NSCs using the TRIZOL reagent (Invitrogen Corp.). For RT-PCR,
5 µg of total RNA was reverse-transcribed using oligo(dT) (Integrated DNA
Technologies Inc., Coralville, IA) and SuperScript II reverse transcriptase (Invitrogen
Corp.). Specific oligonucleotide primer pairs were incubated with cDNA template for
PCR amplification using the Expand High FidelityPLUS PCR System from Roche Applied
Science. The following sequences were used as primers: p53 sense
5’-GTGAAGCCCTCCGAGTGTCAGGAGC-3’; p53 antisense 5’-GGTGGGCAG
CGCTCTCTTTGCGC-3’; β-actin sense 5’-GTGGGCCGCTCTAGGCACCAA-3’; and
β-actin antisense 5’-CTCTTTGATGTCACGCACGATTTC-3’. The product of the
β-actin RNA was used as control.
2.3.10 Densitometry and Statistical Analysis
The relative intensities of protein and nucleic acid bands were analyzed using the
Quantity One Version 4.6 densitometric analysis program (Bio-Rad Laboratories).
Results from different groups were compared using the Student’s t test, or one-way
ANOVA. Kruskal-Wallis or the Mann-Whitney U tests were also used whenever the
assumptions of the parametric test were not satisfied. Values of p < 0.05 were considered
statistically significant. All statistical analysis was performed with GraphPad InStat
software (GraphPad Software, Inc, San Diego, CA).
2.4 Results
2.4.1 The Akt/p-FOXO3A/Id1 Pathway is Down-Regulated during Differentiation
To investigate the potential cross-talk between survival and differentiation
pathways during mouse NSC differentiation, we first determined the time of neurogenesis
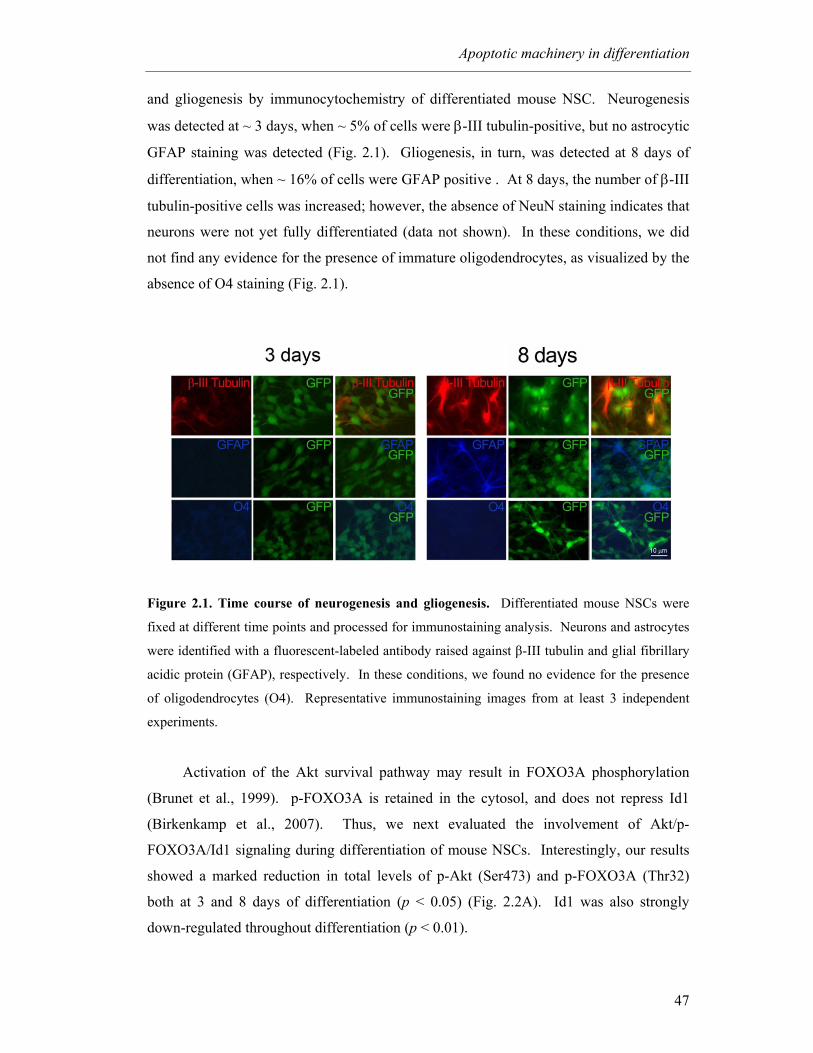
Apoptotic machinery in differentiation
47
and gliogenesis by immunocytochemistry of differentiated mouse NSC. Neurogenesis
was detected at ~ 3 days, when ~ 5% of cells were β-III tubulin-positive, but no astrocytic
GFAP staining was detected (Fig. 2.1). Gliogenesis, in turn, was detected at 8 days of
differentiation, when ~ 16% of cells were GFAP positive . At 8 days, the number of β-III
tubulin-positive cells was increased; however, the absence of NeuN staining indicates that
neurons were not yet fully differentiated (data not shown). In these conditions, we did
not find any evidence for the presence of immature oligodendrocytes, as visualized by the
absence of O4 staining (Fig. 2.1).
Figure 2.1. Time course of neurogenesis and gliogenesis. Differentiated mouse NSCs were
fixed at different time points and processed for immunostaining analysis. Neurons and astrocytes
were identified with a fluorescent-labeled antibody raised against β-III tubulin and glial fibrillary
acidic protein (GFAP), respectively. In these conditions, we found no evidence for the presence
of oligodendrocytes (O4). Representative immunostaining images from at least 3 independent
experiments.
Activation of the Akt survival pathway may result in FOXO3A phosphorylation
(Brunet et al., 1999). p-FOXO3A is retained in the cytosol, and does not repress Id1
(Birkenkamp et al., 2007). Thus, we next evaluated the involvement of Akt/p-
FOXO3A/Id1 signaling during differentiation of mouse NSCs. Interestingly, our results
showed a marked reduction in total levels of p-Akt (Ser473) and p-FOXO3A (Thr32)
both at 3 and 8 days of differentiation (p < 0.05) (Fig. 2.2A). Id1 was also strongly
down-regulated throughout differentiation (p < 0.01).

Chapter2
48
Figure 2.2. The Akt/p-FOXO3A/Id1 signaling pathway is downregulated during mouse NSC
differentiation. NSCs were induced to differentiate for 1, 3 and 8 days. Total, nuclear, and
cytosolic proteins were extracted for immunoblot analysis. A) Representative immunoblots of p-
Akt (Ser473), Akt1/2, p-FOXO3A (Thr32), FOXO3A, and Id1 protein expression. B)
Representative immunoblots (top) and corresponding histograms (bottom) of cytosolic levels of p-
FOXO3A and nuclear levels of FOXO3A in mouse NSCs through differentiation. The results are
expressed as mean ± SEM arbitrary units of at least 6 independent experiments. *p < 0.05 and †p
< 0.01 from 1 day of differentiation. β-actin was used as loading control. Cytosolic and nuclear
fractions were also immunobloted for acetyl-histone H3.
The regulation of FOXO3A was further characterized by evaluating cytosolic p-
FOXO3A (Thr32) and nuclear FOXO3A (Fig. 2.2B). Curiously, the inactive form of
FOXO3A, p-FOXO3A (Thr32), decreased by ~ 60% in the cytosol at 3 days, and was
almost undetectable at 8 days of differentiation (p < 0.01). In contrast, nuclear levels of

Apoptotic machinery in differentiation
49
active, non-phosphorylated FOXO3A, increased by ~ 2- and 3-fold at 3 and 8 days,
respectively. Thus, modulation of the Akt/p-FOXO3A/Id1 survival pathway during
differentiation of mouse NSCs appears to be an important regulatory event. Specifically,
downregulation of Id1, possibly through FOXO3A, appears to be required for the
induction of mouse NSC differentiation.
2.4.2 Apoptosis Factors are Increased during Differentiation
Components of the cell death machinery have been implicated in the differentiation
program (Fernando and Megeney, 2007; Lin et al., 2005). We next evaluated activation
of caspase-3 and total levels of p53 during mouse NSC differentiation using only
adherent cells to exclude dead detached cells. Interestingly, the results demonstrated that
caspase-3 processing increased 3- and 2-fold at 3 and 8 days of differentiation,
respectively (p < 0.05) (Fig. 2.3A). This was confirmed through detection of active
caspase-3 by immunocytochemistry (Fig. 2.3B). Caspase-3 activity increased 20% from
day 3 to day 8 of differentiation (data not shown). Total levels of p53 were not
significantly modulated during differentiation (Fig. 2.3A). Nevertheless, phosphorylation
of p53 at serine 15, which regulates p53 stability and activity (Xu, 2003) and is typically
required for transactivation was up-regulated in early stages of differentiation. In fact, p-
p53 (Ser15) increased by > 2-fold at 3 days, returning to control levels at 8 days of
differentiation. In addition, p53 DNA binding activity was increased by ~ 3- and 2-fold
at 3 and 8 days of differentiation (p < 0.05) (Fig. 2.3C), respectively, further indicating
that regulation of p53 transcriptional activity is an important event for differentiation of
mouse NSCs. Finally, both nuclear morphology analysis and TUNEL assays showed
only ~ 2% of apoptotic cells during differentiation (Fig. 2.3D). This indicates that
changes in pro-apoptotic molecules were not associated with increased cell death.

Chapter2
50
Figure 2.3. p53 activity and caspase-3 cleavage are modulated during mouse NSC
differentiation. NSCs were induced to differentiate for 1, 3 and 8 days. Cells were fixed and
processed for morphological evaluation of apoptosis. Total proteins were extracted for
immunoblot analysis, while nuclear extracts were used for p53-DNA binding assays. A)
Representative immunoblots of caspase-3 processing, and total p53 and phosphorylated p53
(Ser15) expression. β-actin was used as loading control. B) Detection of active caspase-3.
Representative immunostaining images from at least 3 independent experiments. C) p53-DNA
binding activity. Nuclear protein extracts were prepared as described in “Material and Methods”.
The level of p53 present in nuclear lysates that can bind to its DNA consensus recognition
sequence was determined by the Universal EZ-TFA p53 assay and expressed as fold change
relative to day 1 of differentiation. *p < 0.05 from day 1. D) Evaluation of apoptosis by Hoechst
staining and TUNEL assay. Representative images from at least 3 independent experiments.
GFP, green fluorescent protein.
2.4.3 Caspases Inhibit p-FOXO3A/Id1 Signaling during Differentiation
We next characterized possible interactions between apoptosis executioners and
regulation of the Akt/p-FOXO3A/Id1 pathway during differentiation of mouse NSCs.
For this purpose, we suppressed caspase activity with a pan-caspase inhibitor, z-
VAD.fmk, and re-evaluated changes in Akt/p-FOXO3A/Id1 expression, p53

Apoptotic machinery in differentiation
51
phosphorylation (Ser 15), and p53-DNA binding activity. As expected, cells treated with
z-VAD.fmk showed reduced caspase-3 activation at both 3 and 8 days of differentiation
(Fig. 2.4A). Notably, caspase suppression markedly decreased p-p53 (Ser15) expression
(Fig. 2.4A) and p53 transcriptional activation (Fig 2.4B) (p < 0.05). More importantly, in
the absence of caspase activity, the p-FOXO3A/Id1 pathway was not down-regulated
during differentiation. Indeed, p-FOXO3A (Thr32) increased ~ 2-fold at 3 (p < 0.01) and
8 (p < 0.05) days. Id1, in turn, increased 40% at 3 days (p < 0.05) and remained
unchanged at 8 days of differentiation. p-Akt (Ser473) was maintained throughout
differentiation (data not shown). Finally, the absence of significant nuclear fragmentation
and TUNEL-positive cells after caspase inhibition (Fig. 2.4C) suggests that z-VAD.fmk-
induced changes were not associated with increased cell death. Thus, our results indicate
that caspases are required, at least in part, to down-regulate p-FOXO3A and Id1,
facilitating differentiation of mouse NSCs.
Figure 2.4. Pan-caspase inhibition modulates p53 activity and interferes with the Akt/p-
FOXO3A/Id1 pathway during mouse NSC differentiation. NSCs were incubated with a pan-
caspase inhibitor, z-VAD.fmk, and collected after 3 and 8 days in differentiation medium. Cells
were fixed and stained for morphological evaluation of apoptosis. Total proteins were extracted
for immunoblot analysis, while nuclear extracts were used for p53-DNA binding assays. A)
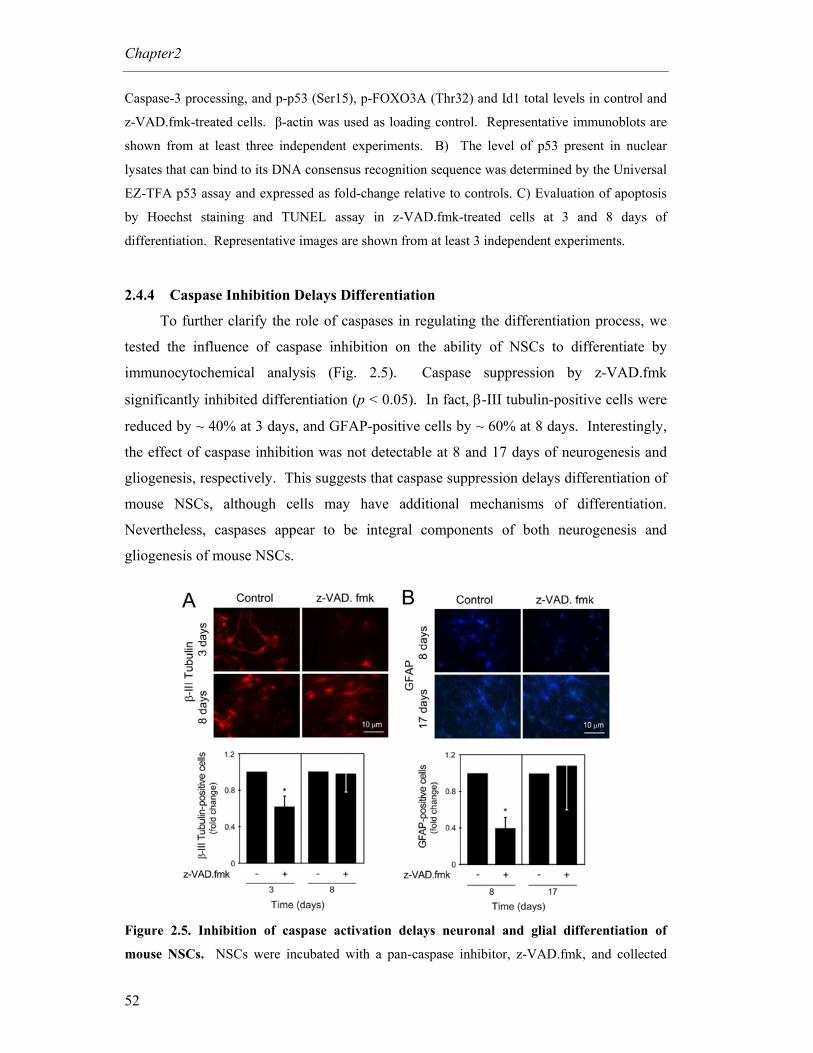
Chapter2
52
Caspase-3 processing, and p-p53 (Ser15), p-FOXO3A (Thr32) and Id1 total levels in control and
z-VAD.fmk-treated cells. β-actin was used as loading control. Representative immunoblots are
shown from at least three independent experiments. B) The level of p53 present in nuclear
lysates that can bind to its DNA consensus recognition sequence was determined by the Universal
EZ-TFA p53 assay and expressed as fold-change relative to controls. C) Evaluation of apoptosis
by Hoechst staining and TUNEL assay in z-VAD.fmk-treated cells at 3 and 8 days of
differentiation. Representative images are shown from at least 3 independent experiments.
2.4.4 Caspase Inhibition Delays Differentiation
To further clarify the role of caspases in regulating the differentiation process, we
tested the influence of caspase inhibition on the ability of NSCs to differentiate by
immunocytochemical analysis (Fig. 2.5). Caspase suppression by z-VAD.fmk
significantly inhibited differentiation (p < 0.05). In fact, β-III tubulin-positive cells were
reduced by ~ 40% at 3 days, and GFAP-positive cells by ~ 60% at 8 days. Interestingly,
the effect of caspase inhibition was not detectable at 8 and 17 days of neurogenesis and
gliogenesis, respectively. This suggests that caspase suppression delays differentiation of
mouse NSCs, although cells may have additional mechanisms of differentiation.
Nevertheless, caspases appear to be integral components of both neurogenesis and
gliogenesis of mouse NSCs.
Figure 2.5. Inhibition of caspase activation delays neuronal and glial differentiation of
mouse NSCs. NSCs were incubated with a pan-caspase inhibitor, z-VAD.fmk, and collected

Apoptotic machinery in differentiation
53
after 3, 8, and 17 days in differentiation medium. Differentiated cells were fixed at indicated
times and processed for immunostaining assays. Neurons and astrocytes were identified with a
fluorescent-labeled antibody raised against β-III tubulin and glial fibrillary acidic protein (GFAP),
respectively. A) Representative immunocytochemistry images from at least 3 independent
experiments. B) Corresponding histograms of β-III tubulin and GFAP positive cells throughout
differentiation. The results are expressed as mean ± SEM arbitrary units of at least 3 independent
experiments. *p < 0.05 from controls.
2.4.5 p53 Represses p-FOXO3A/Id1 Signaling in Early Stages of Differentiation
We next investigated whether caspases require p53 to regulate differentiation of
NSCs. First, we determined the effect of siRNA-mediated silencing of p53 on p-
FOXO3A and Id1. p53 mRNA and protein expression decreased by ~ 90 and 70%,
respectively, in mouse NSCs after transfection with siRNAs for p53 (p < 0.01) (Fig.
2.6A). More importantly, siRNA-mediated silencing of p53 reversed the down-regulation
of p-FOXO3A (Thr32) and Id1 at 3 days of differentiation, but not at 8 days (Fig. 2.6B).
In fact, at 3 days of differentiation, p-FOXO3A (Thr32) and Id1 increased by ~ 50 (p <
0.01) and 30% (p < 0.05) after p53 suppression, respectively. Inhibition of p53 resulted
in increased caspase-3 cleavage (data not shown), suggesting that caspases may act
upstream of p53 to decrease p-FOXO3A and Id1 during early stages of differentiation. In
contrast, the absence of p53 does not influence p-FOXO3A/Id1 signaling at 8 days of
differentiation. We then treated p53 siRNA-transfected cells with z-VAD.fmk and
evaluated changes at 3 days of differentiation. Notably, the suppression of both caspases
and p53 induced a ~ 4-fold increase in p-FOXO3A compared with 2-fold in cells only
transfected with p53 siRNA (p < 0.05). Id1, in turn, was ~ 4-fold greater after z-
VAD.fmk incubation in p53 siRNA-transfected cells compared with control siRNA-
transfected cells (p < 0.05). Thus, at 3 days of differentiation, caspase inhibition further
enhanced p53 silencing effects, indicating that caspases do not act simply in a p53-
dependent manner. Rather, caspases might work with p53 in regulating p-FOXO3A and
Id1.

Chapter2
54
Figure 2.6. Silencing of p53 decreases p-FOXO3A and Id1 expression levels during mouse
NSC differentiation, without potentiating z-VAD.fmk effects. NSCs were incubated with
either control or p53 siRNA and collected after 3 and 8 days in differentiation medium. Cells
were fixed and stained to assess apoptosis. Total proteins were extracted for immunoblot
analysis. A) Representative RT-PCR (left) and Western blot (right) of p53 and β-actin in cells
transfected with either control or p53 siRNA. B) Representative immunoblots of p-FOXO3A
(Thr32) and Id1 in cells transfected with either control siRNA, p53 siRNA, or p53 siRNA plus z-
VAD.fmk. β-actin was used as loading control. C) Evaluation of apoptosis by Hoechst staining
and TUNEL assay in p53 siRNA transfected cells at 3 days (top) and 8 days (bottom) of
differentiation in the presence or absence of z-VAD.fmk. Representative images from at least 3
independent experiments are shown
Finally, morphology and TUNEL assays revealed that p53 siRNA-mediated effects
were not associated with increased cell death throughout differentiation of mouse NSCs
(Fig. 2.6C top and bottom). Therefore, our results demonstrated that p53 might be
required for early differentiation, and that p53 effects in repressing p-FOXO3A and Id1 is
further enhanced by caspase activation.
2.4.6 Caspases and p53 Act in a Synergistic Manner to Modulate Neurogenesis
To confirm the role of p53 in regulating the differentiation process, we tested the
influence of p53 knockdown on the ability of NSCs to differentiate into neurons and
astrocytes through immunocytochemical analysis of cells transfected with either control
or p53 siRNA. Our results showed that p53 suppression significantly reduced the number

Apoptotic machinery in differentiation
55
of positive cells for β-III tubulin compared with control siRNA-transfected cells (Fig.
2.7A). In addition, inhibition of neurogenesis by p53 siRNA increased after caspase
suppression at 3 days, again implying that caspases and p53 might act in a synergistic
manner. In contrast, siRNA-mediated silencing of p53 did not change the number of
positive cells for GFAP (Fig. 2.7B), suggesting that p53 is not crucial for astrocyte
maturation. Curiously, the silencing effect of p53, in the presence or absence of caspase
inhibition, was not present at 8 days of neurogenesis. This indicates that downregulation
of p53 only delays neurogenesis in mouse NSCs. Nevertheless, caspases and p53 are
crucial elements of differentiation in mouse NSCs, acting in a synergistic manner for
regulating neurogenesis. In contrast, the mechanism that regulates gliogenesis in mouse
NSCs appears to involve caspases, which in turn bypass p53 to promote glial cell
differentiation.
Figure 2.7. Downregulation of p53 delays neuronal and glial differentiation of mouse NSCs.
NSCs transfected with control siRNA, p53 siRNA, or p53 siRNA plus z-VAD.fmk, and collected
after 3, 8, and 17 days in differentiation medium. Cells were fixed at indicated times and
processed for immunostaining assays. Neurons and astrocytes were identified with a
fluorescent-labeled antibody raised against β-III tubulin and glial fibrillary acidic protein (GFAP),
respectively. A) Representative immunocytochemistry from at least 3 independent experiments.

Chapter2
56
B) Corresponding histograms of β-III tubulin and GFAP positive cells throughout the
differentiation period. The results are expressed as mean ± SEM arbitrary units of at least 3
independent experiments. *p < 0.05 from controls.
2.5 Discussion
Neural stem cell differentiation and commitment to either neuronal or glial lineage
have important implications in developmental, pathological, and regenerative processes.
It is, therefore, crucial to thoroughly understand the molecular mechanisms involved in
NSC processing. It has been previously reported that cell death and differentiation
processes may use similar molecular pathways (Fernando and Megeney, 2007). Our
results provide an extended mechanism of action for the apoptosis executioners
throughout the differentiation process. Specifically, they show that caspases are integral
components of differentiation of mouse NSCs that together with p53 can induce
neurogenesis or gliogenesis. Moreover, these specific apoptotic factors interfere with
elements of the Akt/p-FOXO3A/Id1 intracellular signaling pathway to promote
differentiation of NSCs.
It has been demonstrated that Akt/p-FOXO3A/Id1 signaling is an important
regulator of cell differentiation. In fact, the phosphorylation of the forkhead transcription
factor FOXO3A was shown to play a pivotal role in maintaining the hematopoietic stem
cell pool (Miyamoto et al., 2007); and the down-regulation of active FOXO3A accelerates
cellular senescence in human dermal fibroblasts (Kyoung Kim et al., 2005). In addition,
it was recently demonstrated that active FOXO3A is a negative target of Notch signalling
(Mandinova et al., 2008), thus supporting its role of maintaining cells in an
undifferentiated state. Our results indicate that this downstream target of Akt is indeed
altered during differentiation of mouse NSCs. In fact, Akt phosphorylation was
decreased throughout differentiation and this was associated with reduced levels of
cytosolic p-FOXO3A. Akt expression was also attenuated in the early stages of
differentiation in human endothelial progenitor cells and gradually upregulated during
cell maturation (Mogi et al., 2008). Subsequently, we showed that a reduction in Akt-
induced FOXO3A phosphorylation lead to increased activity of FOXO3A, and
subsequently decreased Id1 in the nucleus. The link between FOXO3A and Id1 in
triggering differentiation has already been established in other cell types (Birkenkamp et
al., 2007). The decrease in Id1 expression in mouse NSCs may, in turn, allow basic helix

Apoptotic machinery in differentiation
57
loop helix (bHLH) transcription factors to induce expression of differentiation-associated
genes (Norton, 2000).
Importantly, the present study reveals that apoptosis executioners interfere with
FOXO3A/Id1 signalling to promote both neurogenesis and gliogenesis in mouse NSCs.
Several studies have already elucidated the importance of programmed cell death in
establishing cell lineage for model systems of neural differentiation. However, a precise
role for apoptosis-associated elements in the generation of both neurons and astrocytes is
still largely unknown. Curiously, it was recently reported that members of the Bcl-2
family such as Bcl-XL and Bax play an instructive role in determining the fate of
embryonic cortical precussor cells (Chang et al., 2007a). Bcl-XL and Bax mediate
differentiation of neurons and astrocytes, respectively, independently from their roles in
cell survival and apoptosis.
Herein, we investigated whether caspases and p53 were upregulated during mouse
NSC differentiation. Our data indicated that cleavage of caspase-3 was significantly
increased at both 3 and 8 days of differentiation. In fact, the non-proteolytic functions of
caspases in the regulation of cell survival, differentiation, and inflammation had already
suggested that caspases may become activated without inducing an apoptotic cascade
(Lamkanfi et al., 2007). For example, in erythroblasts, keratinocytes, and lens epithelial
cells undergoing differentiation, enucleation has been regarded as a caspase-mediated
incomplete apoptotic process. In addition, others have previously shown that caspase
activation specifically contributes to the differentiation of monocytes into macrophages,
in the absence of cell death (Sordet et al., 2002). Differentiation-associated caspase
activation involved the release of cytochome c from the mitochondria and led to cleavage
of the protein acinus, while poly(ADP-ribose)polymerase remained uncleaved. In our
study, the role of caspases during differentiation of mouse NSCs was further
characterized. We evaluated the potential cross-talk of caspases with the proliferation
pathway Akt/p-FOXO3A/Id1 in mouse NSCs. Our results revealed that both p-FOXO3A
(Thr 32) and Id1 increased after caspase inhibition, which in turn resulted in delayed
neuronal and astrocyte formation at 3 and 8 days of differentiation, respectively. In
addition, although total levels of p53 were unchanged, its DNA-binding activity
significantly increased throughout differentiation, suggesting that p53 might be also
important for both neuronal and astrocyte formation.
The role of p53 on NSC differentiation was further investigated by using siRNA-
mediated p53 silencing. In fact, the results showed that p53 is pivotal for inducing

Chapter2
58
neurogenesis, and its function is not associated with cell death. Although, we did not
observe an increase of p-p53 (Ser15) at 8 days of differentiation, this does not exclude the
involvement of p53, since this transcription factor may be activated by phosphorylation in
many other residues. Nevertheless, p53 silencing did not compromise both p-FOXO3A
and Id1 at 8 days of differentiation, and even astrocyte generation. The absence of p53
silencing effect at 8 days of differentiation could be explained by several potential
mechanisms. First, silencing of p53 at day 1 of differentiation may not be sufficient to
inhibit p53 expression throughout the course of differentiation. Indeed, mouse NSCs may
re-establish normal levels of p53 mRNA at 8 days of differentiation. Nevertheless, this
hypothesis was excluded when we further transfected cells with p53 siRNA at 4 days of
differentiation, and both p-FOXO3A and Id1 total levels remained unchanged (data not
shown). An alternate explanation is the potential substitution of p53 function by other
family members at day 8 of differentiation (Stiewe, 2007). In fact, it has been
demonstrated that knockouts for p53, p63 or p73 show only slight delays on development.
However, viability is compromised when double or triple knockouts for these genes are
generated, indicating that other members of this family are able to substitute the lack of
one transcription factor (Stiewe, 2007).
Importantly, we have demonstrated that at 3 days of differentiation, caspases and
p53 act in a synergistic manner to induce p-FOXO3A/Id1 repression. In fact, caspase
suppression not only decreased p53-DNA binding activity but it also further increased
siRNA p53 effects during early differentiation of mouse NSCs. Interestingly, in contrast
with the apoptosis cascade, where p53 acts upstream of caspases, our data strongly
suggest that caspases may act upstream of p53 during neurogenesis. Indeed, caspase
inhibition significantly reduced both p53-DNA binding activity and p-p53 (Ser15)
expression levels, and p53 silencing did not decrease caspase-3 cleavage (data not
shown). However, caspases might act in a p53-independent manner, since the inhibition
of both caspases and p53 did not promote the same level of inhibition in neurogenesis, but
rather a cumulative effect. Therefore, caspases and p53 may act, at least in part,
independently to induce neuronal differentiation of mouse NSC.
Our results coupled with previous studies showing that overexpression of Bcl-2, a
negative target of p53, significantly reduced the number of neurons (Esdar et al., 2001),
support the pivotal role of p53 activation during neurogenesis. Further, the critical
involvement of p53 for the fate of neuronal cells was indeed demonstrated by others,

Apoptotic machinery in differentiation
59
showing that p53 is required for neurite outgrowth in cultured cells including primary
neurons as well as for axonal regeneration in mice (Di Giovanni et al., 2006).
It remains to be investigated whether p53 downregulates Akt activity or whether
Akt repression induces p53 activity during neurogenesis of NSC. Interestingly, it has
been demonstrated that Akt downregulation may induce p53 transcriptional activation
through Sirt2. In fact, 14-3-3 β/γ augment deacetylation and down-regulate p53
transcriptional activity by Sirt2 in an Akt-dependent manner (Jin et al., 2008). However,
p53 may also repress the Akt pathway through PTEN (Stambolic et al., 2001).
In addition, the transcription factor FOXO3A might be phosphorylated by either
serum- and glucocorticoid-inducible kinase (SGK1) (You et al., 2004) or IκB kinase
(IKK) (Hu et al., 2004). In fact, SGK1 expression was markedly increased by p53 (You
et al., 2004). In this respect, p53 silencing would lead to decreased p-FOXO3A. On the
other hand, corroborating our results, the β subunit of IKK may be negatively regulated
by p53 (Gu et al., 2004). Indeed, p53 might have acted on differentiation via the IKK
kinase, after levels of p-FOXO3A were markedly increased by p53 knockdown in mouse
NSCs. Importantly, it has been shown that Id1 may be downregulated in a p53-dependent
manner (Qian and Chen, 2008).
Finally, it will be important to investigate which cofactors and/or molecular
pathways interfere with caspases and p53 to influence differentiation versus apoptosis in
mouse NSCs. One explanation for this commitment derives largely from caspase
targeting and activation of substrates or cofactors that are unique to each event. Most
likely, levels of activation of apoptosis executioners are different between apoptosis and
differentiation. In addition, our results do not exclude the possibility that suphydryl-
dependent enzymes other than caspases might also be involved. Thus, the precise role of
caspases and the identification of specific pertinent caspases during differentiation should
be further elucidated.
This study provides an extended mechanism of action for apoptosis-associated
elements, linking major components of cell death with the differentiation process of
mouse NSCs. Caspases modulate differentiation, further enhancing p53 effects at 3 days
of differentiation. Nevertheless, both caspases and p53 were shown to signal the p-
FOXO3A pathway and inhibit repression of differentiation by Id1. Collectively, our
results may contribute to the identification of novel targets for NSC differentiation
procedures that may translate ultimately into improved neuro-replacement therapies.

Chapter2
60
Acknowledgements
This work was supported by grant PTDC/SAU-FCF/67912/2006 from Fundação
para a Ciência e a Tecnologia (FCT) and FEDER. S.S. and M.M.A. were recipients of
postdoctoral SFRH/BPD/20834/2004 and Ph.D. SFRH/BD/28429/2006 fellowships,
respectively, from FCT.

3 Apoptosis-associated microRNAs are modulated in mouse, rat and human
neural differentiation
Márcia M. Aranha1, Daniela M. Santos1, Joana M. Xavier1, Walter C. Low2,3,
Clifford J. Steer4,5, Susana Solá1, Cecília M.P. Rodrigues1
1Research Institute for Medicines and Pharmaceutical Sciences, Faculty of Pharmacy,
University of Lisbon, Lisbon 1649-003, Portugal; 2Department of Neurosurgery,
University of Minnesota Medical School, Minneapolis, MN 55455, USA; 3Stem Cell
Institute, University of Minnesota Medical School, Minneapolis, MN 55455, USA;
4Department of Medicine, University of Minnesota Medical School, Minneapolis, MN
55455, USA; 5Department of Genetics, Cell Biology, and Development, University of
Minnesota Medical School, Minneapolis, MN 55455, USA
BMC Genomics 2010; 11: 514

Reprinted from BMC Genomics, vol. 11: 514, Márcia M. Aranha, Daniela M.
Santos, Joana M. Xavier, Walter C. Low, Clifford J. Steer, Susana Solá, Cecília M.P.
Rodrigues. Apoptosis-associated microRNAs are modulated in mouse, rat and human
neural differentiation, Copyright 2010, with permission from BioMed Central. All rights
reserved.

Apoptosis-associated miRNAs and differentiation
63
3.1 Abstract
MicroRNAs (miRs or miRNAs) regulate several biological processes in the cell.
However, evidence for miRNAs that control the differentiation program of specific neural
cell types has been elusive. Recently, we have shown that apoptosis-associated factors,
such as p53 and caspases participate in the differentiation process of mouse neural stem
(NS) cells. To identify apoptosis-associated miRNAs that might play a role in neuronal
development, we performed global miRNA expression profiling experiments in NS cells.
Next, we characterized the expression of proapoptotic miRNAs, including miR-16, let-7a
and miR-34a in distinct models of neural differentiation, including mouse embryonic
stem cells, PC12 and NT2N cells. In addition, the expression of antiapoptotic miR-19a
and 20a was also evaluated.
The expression of miR-16, let-7a and miR-34a was consistently upregulated in
neural differentiation models. In contrast, expression of miR-19a and miR-20a was
downregulated in mouse NS cell differentiation. Importantly, differential expression of
specific apoptosis-related miRNAs was not associated with increased cell death.
Overexpression of miR-34a increased the proportion of postmitotic neurons of mouse NS
cells.
In conclusion, the identification of miR-16, let-7a and miR-34a, whose expression
patterns are conserved in mouse, rat and human neural differentiation, implicates these
specific miRNAs in mammalian neuronal development. The results provide new insights
into the regulation of neuronal differentiation by apoptosis-associated miRNAs.

Chapter 3
64
3.2 Introduction
Adult neural stem (NS) cells and embryonic stem (ES) cells are capable of
differentiating into multiple cell types of the adult nervous system (Doetsch, 2003;
Temple, 2001b). This unique property promises future medical applications, ranging
from regenerative medicine to drug screening. However, the therapeutic use of NS cells
and ES cells will require the identification of cellular conditions that promote the efficient
commitment of these progenitors to particular cell phenotypes.
Much progress has been made toward the elucidation of specific differentiation
signaling pathways. Surprisingly, recent evidence suggests a functional role for
apoptosis-associated proteins such as p53 (Maimets et al., 2008; Porrello et al., 2000),
caspases (Fernando et al., 2005) and Bcl-2 (Zhang et al., 1996) in differentiation and
development. In addition, the recent discovery of microRNAs (miRs or miRNAs)
introduced a novel type of regulatory control of gene expression during developmental
processes (Ambros, 2004; Bartel, 2004). miRNAs are a class of endogenous noncoding,
highly conserved RNAs of ~ 22 nucleotides in length (Du and Zamore, 2005). They
regulate mRNA expression either by translation repression or cleavage of targeted mRNA
(Bartel, 2004). In the past few years, our understanding of miRNAs has expanded, and a
role for apoptosis-associated miRNAs in cell differentiation is emerging.
Let-7a, a member of the let-7 family, is associated with apoptosis by directly
targeting caspase-3 (Tsang and Kwok, 2008). Let-7 was first identified in Caenorhabditis
elegans and reported to control the timing of fate specification during larval development
(Reinhart et al., 2000). In addition, let-7a was also implicated in neuronal differentiation
(Liu et al., 2009; Sempere et al., 2004). However, the mechanism by which let-7a
regulates cell differentiation is unknown.
Members of the miR-34 family are direct p53 targets, and their upregulation
induces apoptosis and cell cycle arrest (Bommer et al., 2007; Chang et al., 2007b; Corney
et al., 2007; He et al., 2007a; Raver-Shapira et al., 2007; Tarasov et al., 2007). In this
regard, miR-34a has been shown to regulate genes involved in cell cycle control and
apoptosis, including cyclin-dependent kinase 4 (CDK4), CDK6, cyclin D1, E2F3 and
SIRT1 (He et al., 2007a; Sun et al., 2008; Welch et al., 2007). A role for miR-34a in
megakaryocytic differentiation was recently reported, where the miRNA regulates the
expression of MYB, and CDK4 and CDK6, thus promoting cell cycle arrest (Navarro et
al., 2009). Furthermore, miR-34a is involved in dendritic cell differentiation (Hashimi et

Apoptosis-associated miRNAs and differentiation
65
al., 2009), and is required for proper differentiation of mouse ES cells by targeting Sirt1
(Tarantino et al., 2010).
miR-16 is implicated in induction of apoptosis by targeting Bcl-2 (Cimmino et al.,
2005), and is involved in cell cycle regulation by targeting CDK6, cell division cycle
protein 27 (CDC27), the caspase recruitment domain-containing protein 10 (CARD10),
cyclin D1 and cyclin E (Chen et al., 2008; Linsley et al., 2007; Liu et al., 2008).
Nevertheless, an involvement of miR-16 in cell differentiation is virtually unknown.
To gain further insight into the role of apoptosis-associated factors during cell
differentiation, we monitored specific apoptosis-associated miRNAs during NS cell
differentiation. Our results showed that the differential expression of miR-16, let-7a and
miR-34a during mouse NS cell differentiation was not associated with cell death. In
addition, upregulation of these miRNAs observed in distinct models of neural
differentiation strongly suggests that apoptosis-associated miRNAs may play a role in
differentiation.
3.3 Methods
3.3.1 Cell Lines
Mouse NS Cells. Mouse NS cells containing a constitutively expressed marker for
green fluorescent protein (GFP) were used to investigate the process of neural
differentiation. Primary cells were obtained from central nervous system tissue of
embryonic mice and cultured primarily as previously described (Reynolds and Weiss,
1992, 1996; Rietze and Reynolds, 2006). Mouse NS cells were maintained as
neurospheres in undifferentiation medium, serum-free, 1:1 mix of DMEM/F12
(Invitrogen Corp., Grand Island, NY) with 1× N-2 supplement (Invitrogen Corp.), 20
ng/ml epidermal growth factor (EGF) (R & D Systems Inc., Minneapolis, MN), 20 ng/ml
basic fibroblast growth factor (bFGF) (PeproTech EC, London, UK), and 1% penicillin-
streptomycin (Invitrogen Corp.), at 37ºC in humidified atmosphere of 5% CO2.
Subculture occurred at day 7 with mechanical dissociation of neurospheres. The
differentiation of mouse NS cells in vitro was induced by culturing dissociated cells in
differentiation medium containing DMEM/F12 with 1× N-2 supplement, 100 ng/ml
bFGF, 10% fetal bovine serum (FBS) (Invitrogen Corp.), 500 nM all-trans retinoic acid
(Sigma Chemical Co., St. Louis, MO), 50 µM taurine (Sigma Chemical Co.), 10 ng/ml

Chapter 3
66
transforming growth factor-β2 (TGF-β2) (R & D Systems Inc.) and 1% penicillin-
streptomycin in tissue culture plates pre-coated with poly-D-lysine (Sigma Chemical
Co.). The culture medium was changed every 3 days. Differentiated cells at 5 × 104
cells/ml were fixed at different time points, and processed for immunostaining and
evaluation of differentiation. Cultures at 5 × 105 cells/ml were processed for western blot
and PCR analysis.
In parallel experiments, cell death was prevented by adding tauroursodeoxycholic
acid (TUDCA; Sigma Chemical Co.) to the culture medium. Briefly, cells with 3 days of
differentiation were treated with 50 µM of TUDCA for 72 hours. At 6 days, cells were
collected and processed for evaluation of apoptosis and miRNA expression.
miR-34a expression was modulated using 100 nM of precursor control and pre-
miR-34a (Applied Biosystems, Foster City, CA). Mouse NS cells (2.5 × 105 cells/ml)
were transfected at 3 or 5 days of differentiation using Lipofectamine 2000 (Invitrogen
Corp) following manufacturer’s instructions.
Mouse ES Cells. The ES cell line 46C (Sox1-GFP) (Ying et al., 2003) was grown
and maintained as previously reported (Abranches et al., 2009) and induced to
differentiate using a monolayer protocol. Specifically, ES cells were plated in serum-
free medium ESGRO Complete Clonal Grade medium (Millipor Inc.) at high density.
After 24 hours, ES cells were gently dissociated and plated onto 0.1% (v/v) gelatin-coated
tissue culture plastic at 1 × 104 cells/cm2 in RHB-A media (StemCell Science Inc.). For
replating on day 4, cells were dissociated and plated at 2 × 104 cells/cm2 in laminin-
coated tissue culture plastic, using RHB-A medium supplemented with 5 ng/ml murine
bFGF (PeproTech EC, London, UK). Cells at days 4 and 8 of differentiation were
collected to evaluate expression levels of differentiation markers and apoptosis-associated
miRNAs. A positive control for neural differentiation was also included at day 8 using
LY411575, a γ-secretase inhibitor known to inhibit the Notch pathway and thus enhance
neuronal differentiation. Cells were supplemented with either 0.01% DMSO (control) or
10 nM LY411575 (in 0.01% DMSO) for 12 hours. LY411575-treated cells and
respective controls were photographed under an inverted microscope Leica DMIL using a
DC200 camera (Leica Wetzlar, Germany)

Apoptosis-associated miRNAs and differentiation
67
PC12 Cells. Rat adrenal pheochromocytoma (PC12) cells were propagated in
RPMI 1640 medium (Invitrogen Corp.) supplemented with 10% horse serum (Invitrogen
Corp.), 5% FBS (Invitrogen Corp.) and appropriate antibiotics in an humidified incubator
maintained at 37ºC with 5% CO2. PC12 cells were differentiated in RPMI 1640 medium
containing 1% horse serum and antibiotics with either none, 5 or 50 ng/ml mouse nerve
growth factor, (NGF 2.5S) (Chemicon, Temecula, CA) at 24 hours. Differentiation
medium was replaced and fresh NGF 2.5D was added every third day to differentiating
cells. Cells were collected at 1, 2, 4 and 7 days prior to differentiation to evaluate
expression levels of apoptosis-associated miRNAs.
NT2N Cells. The human teratocarcinoma-derived Ntera2/D1 neuron-like (NT2N)
cells were obtained based on standard protocols with minor alterations (Cheung et al.,
1999; Megiorni et al., 2005). Briefly, cells were differentiated by treating adherent
dividing cells with 10 µM all-trans retinoic acid (Sigma Chemical Co.) for up to 3 weeks.
Cells were collected at various time points (0, 14 and 21 days); and for some a replate
onto Matrigel coated culture dishes after 14 days of retinoic acid was performed.
Replated cells were incubated with mitosis inhibitors (1 µM cytosine D-
arabinofuranoside, 10 µM fluorodeoxyuridine, 10 µM uridine) (Sigma Chemical Co.), in
the absence of retinoic acid, and collected 7 days after treatment to evaluate expression
levels of apoptosis-associated miRNAs.
3.3.2 RNA isolation and semiquantitative reverse-transcriptase-polymerase chain
reaction (RT-PCR)
Total RNA was isolated from mouse NS cells, PC12 and NT2N cells by TRIzol
reagent (Invitrogen Corp.) according to manufacturer’s protocol. ES cell RNAs were
extracted using High Pure RNA Isolation kit (Roche Diagnostics), with the inclusion of
DNAse I treatment according to manufacturer’s instructions. For semiquantitative RT-
PCR, 0.5-1 µg of total RNA from NS and ES cells were reverse-transcribed using
oligo(dT) (Integrated DNA Technologies Inc., Coralville, IA) and SuperScript II reverse
transcriptase (Invitrogen Corp.). Specific oligonucleotide primer pairs were incubated
with cDNA template for PCR amplification using the Expand High FidelityPLUS PCR
System from Roche Applied Science. The following sequences were used as primers:
Nestin sense 5’-CTGGAACAGAGATTGGAAGGCCGCT-3’; Nestin antisense

Chapter 3
68
5’-GGATCCTGTGTCTTCAGAAAGGCTGTCAC-3’; Mash1 sense 5’-AGATGAG
CAAGGTGGAGACG-3’; Mash1 antisense 5’-TGGAGTAGTTGGGGGAGATG-3’;
Ngn1 sense 5’-ATGCCTGCCCCTTTGGAGAC-3’; Ngn1 antisense 5’-TGCA
TGCGGTTGCGCTCGC-3’; Ngn2 sense 5’-ACCGCATGCACAACCTAAAC-3’; Ngn2
antisense 5’-AGCGCCCAGATGTAATTGTG-3’; β-III tubulin sense 5’-AATGAG
GCCTCCTCTCACAA-3’; β-III tubulin antisense 5’-CTTGCTGATGAG
CAGTGTGC-3’; GFAP sense 5’-CCAAACTGGCTGATGTCTACC-3’; and GFAP
antisense 5’-GCTTCATGTGCCTCCTGTCTA-3’¸ GAPDH sense 5’-ATTCAA
CGGCACAGTCAAGG-3’; and GAPDH antisense 5’-TGGATGCAGGGATGAT
GTTC-3’. The ribosomal RNA subunit 28S was used as control when using mouse NS
cells, while GAPDH was used as control in mouse ES cells.
3.3.3 Preparation of labeled RNA and array hybridization
Microarray analysis was performed using a custom microarray platform, highly
sensitive, allowing the profiling of even weakly expressed miRNAs (Kalscheuer et al.,
2008). Briefly, a miRNA probe set containing ~1140 oligonucleotides as probes,
complementary to Caenorhabditis elegans, Drosophila, zebra fish, mouse, rat and human
miRNAs was purchased from Invitrogen. The set also included a number of internal and
negative control probes. Oligonucleotides were printed in quadruplicates on Corning
GAPSII-coated slides in the Microarray Facility at the University of Minnesota. For
RNA labeling, 25 µg of total RNA was ligated to 0.5 µg of a synthetic linker, pCU-
DY547 (Dharmacon, Lafayette, CO, USA). To control the hybridization process,
reference DNA oligonucleotides complementary to a subset of mammalian miRNAs were
combined and labeled with a ULYSIS Alexa Fluor 647 Kit (Invitrogen Corp.). Labeled
RNAs and DNAs were then mixed and hybridized to microarray slides (Thomson et al.,
2004). Finally, slides were scanned with a ScanArray 5000 machine (Perkin Elmer,
Waltham, MA, USA). Microarray images were processed with Bluefuse software
(BlueGnome, Cambridge, UK) to quantify pixel intensities. Individual spots on the slides
were further inspected to exclude abnormal spots from subsequent calculations.

Apoptosis-associated miRNAs and differentiation
69
3.3.4 Evaluation of miRNAs expression levels by quantitative Real Time-PCR
Real-time PCR was performed in an Applied Biosystems 7300 Sequence Detection
System (Applied Biosystems, Foster City, CA). Ten nanograms of total RNA were
reverse transcribed using a TaqMan® MicroRNA Reverse Transcription (RT) kit from
Applied Biosystems. Each RT reaction contained 1x stem-loop RT specific primer, 1x
reaction buffer, 0.25 mM each of dNTPs, 3.33 U/µl Multiscribe RT enzyme and 0.25
U/µl RNase inhibitor. The 15-µl reactions were incubated for 30 min at 16ºC, 30 min at
42ºC, and 5 min at 85ºC and then held at 4ºC. The PCR reaction was performed using a
standard TaqMan® PCR kit protocol (Applied Biosystems). Briefly, following the RT
step, 1.33 µl of the RT reaction were combined with 1 µl of a TaqMAn MicroRNA Assay
(20x; forward primer, reverse primer and probe) and 17.67 µl of TaqMan® Universal
PCR Master Mix, No AmpErase® UNG in a 20 µl final volume. The reactions were
incubated at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min;
and were run in triplicate. miRNA expression levels relative to GAPDH (mouse NS cells,
ES cells, and PC12) or RNU6B (NT2N) were calculated on the basis of ∆∆Ct method.
GeNorm and Norm-Finder were used to identify the best stable combination of reference
genes/single gene for normalization in real-time PCR assays. Calibration was set using
undifferentiated mouse NS cells, mouse ES cells with 4 days of differentiation or
untreated with LY411575, PC12 with 1 day of differentiation and undifferentiated NT2N
cells or non-replated NT2N cells. The n-fold change in miRNAs expression was
determined according to the method of 2-∆∆CT.
3.3.5 Immunoblotting
Steady-state levels of β-III tubulin, neuronal nuclei (NeuN) and glial fibrillary
acidic protein (GFAP) were determined by Western blot. Total protein extracts of mouse
NS cells were prepared in lysis buffer, following standard established protocols. Protein
content was measured by the Bio-Rad protein assay kit according to the manufacture’s
specification, using bovine serum albumin as standard. 50-80 µg of total protein extracts
were separated on 12% sodium dodecyl sulphate-polyacrylamide electrophoresis gel and
then subjected to immunoblots using primary mouse monoclonal antibody reactive to β-
III tubulin (Tuj1, Covance, Princeton, NJ), NeuN (MAB377, Chemicon International),
and GFAP (MAB360, Chemicon International). Blots were subsequently incubated with
secondary antibodies conjugated with horseradish peroxidase (Bio-Rad Laboratories,

Chapter 3
70
Hercules, CA, USA). Finally, membranes were processed for protein detection using
Immobilon (Millipore Corporation, Billerica, MA) or SuperSignal reagent (Pierce,
Rockford, IL). Ponceau S staining was used to assess equal gel loading (Romero-Calvo
et al.).
3.3.6 Immunocytochemistry
Mouse NS cells were fixed with paraformaldehyde (4% w/v) in phosphate-buffered
saline (PBS) and blocked for 1 hour at room temperature in PBS containing 0.1% Triton-
X-100, 1% FBS, and 10% normal donkey serum (Jackson ImmunoResearch Laboratories,
Inc., West Grove, PA). Subsequently, cells were incubated with either anti-β-III tubulin
(rabbit, polyclonal; Chemicon International) or anti-GFAP antibodies (mouse,
monoclonal; Chemicon International) at a dilution of 1:1000 in blocking solution,
overnight at 4ºC. Cells were then incubated with either anti-mouse or anti-rabbit IgG
conjugated to AMCA (Jackson ImmunoResearch Laboratories, Inc.) and Alexa 594
(Molecular Probes-Invitrogen), respectively, for 2 hour at room temperature. Samples
were mounted using Fluoromount-G™ (Beckman Coulter, Inc.). Fluorescence
microscopy assessments were performed with a Zeizz AX10 microscope (Carl Zeiss,
Jena, Germany) equipped with a Leica DFC490 camera (Leica Weitzlar, Germany).
Living undifferentiated cells were photographed under an inverted microscope Leica
DMIL with a DC200 camera (Leica Weitzlar, Germany).
3.3.7 Assessment of apoptosis
Apoptosis was assessed by evaluating DNA fragmentation, nuclear morphology and
phosphatidylserine exposure.
Evaluation of DNA fragmentation by TUNEL assay. Apoptosis-induced DNA
fragmentation was determined using the transferase mediated deoxyuridine triphosphate
(dUTP)-digoxigenin nick-end labeling (TUNEL) assay. Cells were fixed with 4% (w/v)
paraformaldehyde and processed using an Apoptag in situ apoptosis detection kit
according to the manufacturer’s protocol (Chemicon Int., Temecula, CA). The samples
were examined using a Leica DM 2500 bright-field microscope (Leica Weitzlar,
Germany).

Apoptosis-associated miRNAs and differentiation
71
Morphologic evaluation of apoptosis. Hoechst labeling of mouse NS cells was used
to detect apoptotic nuclei. In brief, the medium was gently removed at the indicated
times with minimal detachment of the cells. Attached cells were fixed with 4% (w/v)
paraformaldehyde in PBS, pH 7.4, for 10 minutes at room temperature, incubated with
Hoechst dye 33258 (Sigma Chemical Co.) at 5 µg/ml in PBS for 5 minutes, washed with
PBS and mounted using PBS:glycerol (3:1, v/v). Microscopy assessments were
performed with a Zeizz AX10 microscope (Carl Zeiss, Jena, Germany) equipped with a
Leica DFC490 camera (Leica Weitzlar, Germany). Normal nuclei showed non-
condensed chromatin dispersed over the entire nucleus. Apoptotic nuclei were identified
by condensed chromatin, contiguous to the nuclear membrane, as well as nuclear
fragmentation of condensed chromatin.
Cytofluorometric analysis of apoptosis. Apoptotic cells were quantified by
cytofluorometric analysis using a FACSCalibur (Becton Dickinson, Mountain View, CA)
as described previously (Schutte et al., 1998). Cells were stained with the vital dye
propidium iodide (PI; 5µg/mL; Sigma, Steinheim, Germany) and concomitantly with
Annexin-V-APC (eBioscience, Inc.) according to manufacturer’s instructions to
determine the phosphatidylserine exposure. Data were statistically evaluated using
FlowJo software (Tree Star, Inc, Ashland, OR).
3.3.8 Flow Cytometry
Cells were trypsinized (0.025% trypsin/EDTA; Invitrogen Corp.) and harvested in
Ca2+-free and Mg2+-free PBS and 2% FBS. After washing, 0.5 - 1 × 106 cells were fixed
with paraformaldehyde (4% w/v) in PBS for 20 min on wet ice, and blocked for 20 min in
PBS, containing 0.25% saponin (Fluka, Biochemika, Switzerland) and 5% normal donkey
serum (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Subsequently,
cells were incubated with anti-NeuN (MAB377; Chemicon International) at a dilution of
1:20 in PBS, containing 0.1% saponin and 5% normal donkey serum, for 30 min. Cells
were then incubated with anti-mouse antibody conjugated to Cy5 (Jackson
ImmunoResearch Laboratories, Inc.), for 20 min. Cells were analyzed using a
FACSCalibur (Becton Dickinson, Mountain View, CA).

Chapter 3
72
3.3.9 Statistical Analysis
Results from different groups were compared using the Student’s t test, or one-way
ANOVA. Kruskal-Wallis or the Mann-Whitney U tests were also used whenever the
assumptions of the parametric test were not satisfied. Values of p < 0.05 were considered
statistically significant. All statistical analysis was performed with GraphPad InStat
software (GraphPad Software, Inc, San Diego, CA).
3.4 Results
3.4.1 NS cells have both neurogenic and gliogenic potential in vitro
It is known that both embryonic neural tissue and certain regions of the adult
vertebrate central nervous system contain resident populations of progenitor/precursor
stem cells (Alvarez-Buylla and Garcia-Verdugo, 2002). NS cells can be isolated and
expanded as undifferentiated floating clusters called neurospheres in the presence of
bFGF and/or EGF (Tropepe et al., 1999). In addition, NS cells have the ability to
differentiate into neurons, astrocytes and oligodendrocytes in a time controlled fashion
(Qian et al., 2000). Thus, NS cells represent a powerful in vitro model system for
studying and elucidating the molecular mechanisms that underlie neural differentiation.
Using mouse NS cells constitutively expressing GFP (Fig. 3.1A), we confirmed the
differentiation potential toward neuronal and glial fates by monitoring the appearance of
various cell-specific markers.
The proneural basic helix–loop–helix (bHLH) transcription factor Neurogenin1
(Ngn1) was not expressed in neurospheres, while Mash1 was readily detected (Fig. 3.1B).
Following incubation in differentiation medium, cells displayed a sharp increase in
markers for neurons and neural precursors. Ngn1 was readily detected at 6 hours and
tended to decrease during cell differentiation. Mash1 was slightly increased but not
significantly modulated. Upregulation of proneural bHLH transcription factors was
associated with a subsequent increase in β-III tubulin at both the mRNA and protein
levels, indicating that neurogenesis is induced early after cell incubation in differentiation
medium. At the protein level, β-III tubulin expression peaked at 1 day of differentiation
and gradually decreased thereafter (Fig. 3.1C). These results suggested that the
neurogenic potential of mouse NS cells rapidly decreased with differentiation.

Apoptosis-associated miRNAs and differentiation
73
Figure 3.1. Mouse NS cells have both neurogenic and gliogenic potential in vitro. Mouse NS
cells were expanded in the presence of EGF and bFGF, and induced to differentiate as described
in Materials and Methods. Cells were harvested for total RNA and protein extraction, or fixed at
different time points and processed for immunostaining analysis. Neuronal differentiation was
readily detected after incubation in differentiation medium, while glial production was only
observed from day 3. A) Mouse NS cells constitutively expressing GFP were grown in
suspension as neurospheres that increased in number and size with cell proliferation (phase
contrast and GFP fluorescence images). B) Semi-quantitative RT-PCR analysis for selected
markers of lineage commitment at successive time-points of mouse NS cell differentiation.
Neurospheres (undifferentiated NS cells) collected before platting in differentiation medium were
considered 0 time differentiation. Data was normalized to 28S. C) Representative immunoblots
showing protein expression of neuronal (β-III tubulin and NeuN) and glial (GFAP) markers
during mouse NS cell differentiation. Ponceau S was used as loading control. D) Mouse NS cells
labeled with anti-β-III tubulin and anti-GFAP antibodies, to visualize neurons and glial cells,
respectively. Representative immunostaining images were from at least 3 independent
experiments. Scale bar: 50 µm. Ngn1, neurogenin1; GFAP, glial fibrillary acidic protein; NeuN,
neuronal nuclei; h, hours; d, days.

Chapter 3
74
In cell culture, there is an inverse relationship between Ngn expression and the
propensity of the neural progenitors to undergo differentiation into astrocytes, suggesting
that proneural bHLH factors might act to suppress the formation of astrocytes (Sun et al.,
2001). In agreement with these observations, the glial marker GFAP was only detected at
day 3, in association with decreased levels of Ngn1 and concomitant with a reduction in
the number of neurons. Importantly, analysis of NeuN protein expression revealed
postmitotic neurons after 6 days of differentiation. Finally, analysis by
immunocytochemistry using β-III tubulin and GFAP markers further confirmed the
decreased neurogenic potential of mouse NS cells throughout differentiation, and the
generation of glial cells after the peak of neuronal production (Fig. 3.1D).
3.4.2 Apoptosis-associate miRNAs are differentially modulated during mouse
neural stem cell differentiation
miRNAs are thought to play a role in cancer pathogenesis. In fact, a specific subset
of miRNAs is upregulated in most cancer profiles, and are, therefore, considered to be
antiapoptotic miRNAs (Garofalo et al., 2010). The best characterized of that subgroup
are miR-21, miR-222, miR-221, miR-17-92 cluster, miR-106 and miR-155. In contrast,
proapoptotic miRNAs are usually downregulated in cancer, and include miR-15, miR-16,
the let-7 family and members of the miR-34 family. Importantly, we have recently shown
that apoptosis-associated factors temporally modulate mouse NS cell differentiation
(Aranha et al., 2009). As miRNAs have recently been implicated in neural differentiation
(Li and Jin, 2010), we sought to investigate whether apoptosis-specific miRNAs are
particularly relevant during mouse NS cell differentiation.
Global miRNA expression profiling studies revealed differential expression patterns
of apoptosis-related miRNAs throughout mouse NS cell differentiation (Tables 3.1 and
3.2). In general, let-7 family members were upregulated at 3 days, and decreased at 8
days of differentiation. Interestingly, and similar to miR-34a/b/c, miR-16 was
upregulated at both 3 and 8 days of neural differentiation. In contrast, antiapoptotic
miRNAs were in general downregulated at 8 days. Further analysis of the differentially
expressed miRNAs involved clustering analysis into different groups, according to
variations in the apoptosis-associated miRNAs expression. Less differentiated cells (1
and 3 days) were grouped separately from those that were more differentiated (8 days)

Apoptosis-associated miRNAs and differentiation
75
(data not shown). Importantly, similar grouping was not achieved in clustering analysis
using all detected miRNAs.
Table 3.1. Proapoptotic miRs expression during mouse NS cell differentiation Time (days)
miR 1 3 8
miR-15a 1.00 0.75 0.71
miR-15b 1.00 1.11 0.61
miR-16 1.00 1.91 4.09
Let-7a 1.00 1.72 0.91
Let-7b 1.00 3.68 0.72
Let-7c 1.00 1.84 0.60
Let-7d 1.00 1.31 0.40
Let-7e 1.00 0.83 0.38
Let-7f 1.00 2.20 0.75
Let-7g 1.00 1.40 1.13
Let-7i 1.00 1.20 0.77
miR-34a 1.00 3.01 3.39
miR-34b 1.00 2.36 2.56
miR-34c 1.00 1.18 1.67
miR-122a 1.00 0.85 0.60
miR-29a 1.00 0.66 0.52
miR-29b 1.00 0.98 0.21
miR-29c 1.00 1.09 0.78
miR-101 1.00 4.33 4.70
Results are presented as fold change variation versus day 1 of differentiation. Data are
mean values from two independent experiments.

Chapter 3
76
Table 3.2. Antiapoptotic miRs expression during mouse NS cell differentiation
Time (days)
miR 1 3 8
miR-21 1.00 0.68 1.03
miR-221 1.00 0.51 0.61
miR-222 1.00 0.72 1.68
miR-106b 1.00 0.44 0.29
miR-93 1.00 0.88 0.82
miR-25 1.00 1.21 2.58
miR-17-5p 1.00 0.64 0.47
miR-17-3p 1.00 2.98 1.48
miR-18a 1.00 1.16 0.46
miR-19a 1.00 1.78 0.57
miR-19b 1.00 0.21 0.18
miR-20a 1.00 0.35 0.22
miR-92b 1.00 1.03 1.04
miR-155 1.00 1.62 0.41
Results are presented as fold change variation versus day 1 of differentiation. Data are
mean values from two independent experiments.
Based on a possible link between miR-16, let-7a and miR-34a with known
apoptotic molecules that have already been associated with differentiation, we decided to
validate microarray data for the three proapoptotic miRNAs throughout mouse NS cell
differentiation by quantitative real time-PCR (Fig. 3.2). Antiapoptotic miR-19a and miR-
20a, two miR-17-92 cluster members were also investigated. Proapoptotic miRNAs were
confirmed to be upregulated during mouse NS cell differentiation. Notably, miR-16
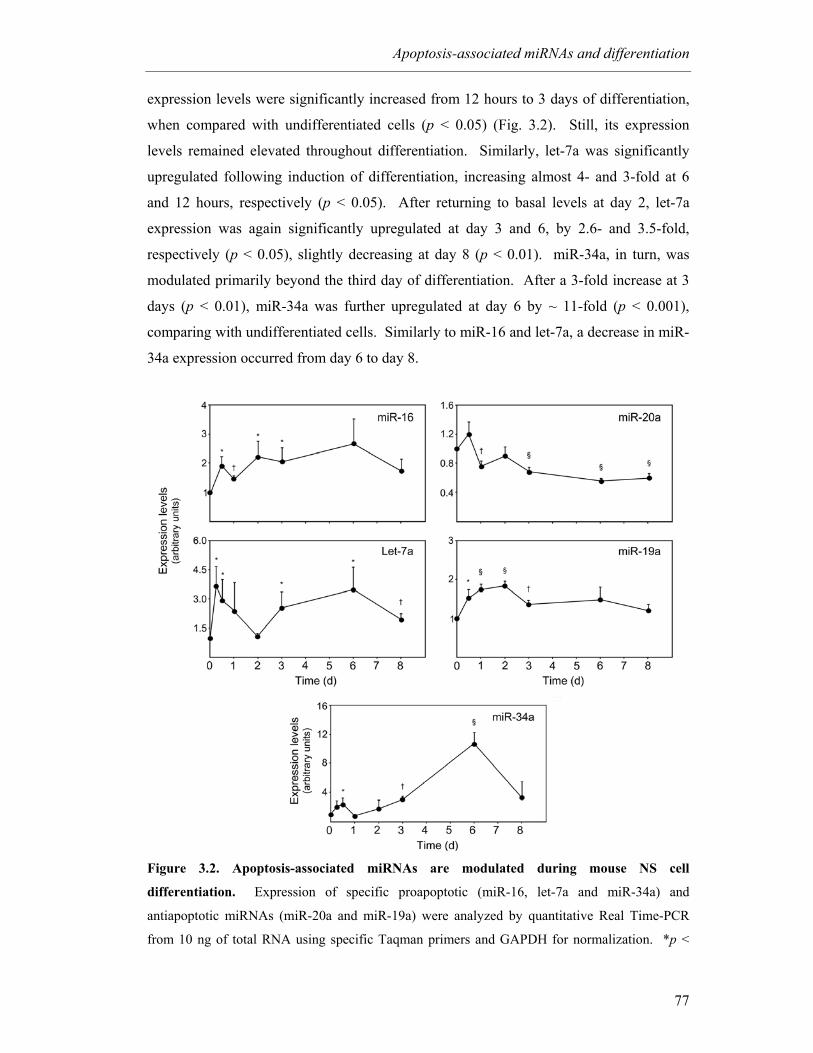
Apoptosis-associated miRNAs and differentiation
77
expression levels were significantly increased from 12 hours to 3 days of differentiation,
when compared with undifferentiated cells (p < 0.05) (Fig. 3.2). Still, its expression
levels remained elevated throughout differentiation. Similarly, let-7a was significantly
upregulated following induction of differentiation, increasing almost 4- and 3-fold at 6
and 12 hours, respectively (p < 0.05). After returning to basal levels at day 2, let-7a
expression was again significantly upregulated at day 3 and 6, by 2.6- and 3.5-fold,
respectively (p < 0.05), slightly decreasing at day 8 (p < 0.01). miR-34a, in turn, was
modulated primarily beyond the third day of differentiation. After a 3-fold increase at 3
days (p < 0.01), miR-34a was further upregulated at day 6 by ~ 11-fold (p < 0.001),
comparing with undifferentiated cells. Similarly to miR-16 and let-7a, a decrease in miR-
34a expression occurred from day 6 to day 8.
Figure 3.2. Apoptosis-associated miRNAs are modulated during mouse NS cell
differentiation. Expression of specific proapoptotic (miR-16, let-7a and miR-34a) and
antiapoptotic miRNAs (miR-20a and miR-19a) were analyzed by quantitative Real Time-PCR
from 10 ng of total RNA using specific Taqman primers and GAPDH for normalization. *p <

Chapter 3
78
0.05, †p < 0.01 and §p < 0.001 compared to day 0 (undifferentiated cells). Expression levels were
calculated by the ∆∆Ct method using undifferentiated cells as calibrator. Data represent mean ±
SEM of four independent experiments.
The synchronous induction of proapoptotic miRNAs suggested that they may play a
role during mouse NS cell differentiation. In contrast, antiapoptotic miR-20a was mainly
downregulated throughout neural differentiation. In fact, its expression levels were ~ 2-
fold downregulated from 6 days and beyond (p < 0.001), compared to undifferentiated
cells. Curiously, miR-19a expression was mostly increased during the first 3 days of
differentiation, gradually decreasing toward control levels. The onset of miR-19a
expression correlates with induction of proliferation during the first 3 days of
differentiation (data not shown).
3.4.3 Expression of apoptosis-related miRNAs is not associated with increased cell
death during mouse NS cell differentiation
To exclude the possibility that expression of proapoptotic miRNAs is associated
with increased cell death, we evaluated cellular apoptosis during mouse NS cell
differentiation using Hoechst staining, TUNEL assay and cytofluorometric analysis.
Undifferentiated cells exhibited major apoptosis-associated alterations, including
chromatin condensation and nuclear fragmentation (Fig. 3.3A, Hoechst), DNA
fragmentation (Fig. 3.3A, TUNEL), exposure of phosphatidylserine on the outer leaflet of
the plasma membrane (detected with annexin V-APC conjugates) (Fig. 3.3B and C), as
well as membrane permeabilization, as indicated by the staining with the vital dye PI.
Importantly, induction of cell differentiation resulted in minor alterations in nuclear
morphology, while DNA fragmentation was slightly detected during the first two days of
differentiation. In addition, phosphatidylserine exposure was only slightly increased from
day 3 to day 6, but always less than 6% during differentiation. Furthermore, no
significant loss of cell viability was observed. Taken together, our results indicated that
differentiation of mouse NSC was not associated with a significant increase of cell death.

Apoptosis-associated miRNAs and differentiation
79
Figure 3.3. Differentiation of mouse NS cells is not associated with increased cell death.
Mouse NS cells at different days of differentiation were fixed and processed for either
morphological evaluation of apoptosis or TUNEL assay, or were stained with Annexin-V-
APC/PI. A) Evaluation of apoptosis by Hoechst staining and TUNEL assay. Representative
images from at least three independent experiments. Arrows indicate apoptotic cells. B)
Representative FACS diagrams depicting the percentages of either dying (Annexin+/PI-) or dead
(Annexin+/PI+) cells. C) Quantitation of the data depicted in FACS diagrams. Results are mean
± SEM. This experiment was repeated at least three times, yielding comparable results. Scale
bar: 10 µm. d, days.
All three proapoptotic miRNAs increased from day 3 to day 6 of differentiation. In
addition, a concomitant increase in cell death was also observed. Thus, to exclude the
role of these specific miRNAs in apoptosis, we inhibited apoptosis at 6 days and
reevaluated changes on miRNA expression (Fig. 3.4). Incubation with the antiapoptotic
TUDCA for 72 hours, reduced Annexin+/PI- cells from 6.4% to 5.3% (Fig. 3.4A and B).
In addition, loss of cell viability (Annexin+/PI+ cells) was significantly reduced from
14.2% to 10.6% (p < 0.05). More importantly, the decrease in cell death after TUDCA
treatment did not decrease the expression of proapoptotic miRNAs (Fig. 3.4C). These
results strongly suggested that modulation of miR-16, let-7a and miR-34a was most likely
due to cell differentiation rather than cell death.

Chapter 3
80
Figure 3.4. Inhibition of apoptosis by TUDCA was not associated with a decrease in
proapoptotic miRNAs expression. Mouse NS cells with 3 days of differentiation were either
untreated or treated with 50 µM of TUDCA for 72 hours. Collected cells were stained with
Annexin-V-APC/PI to evaluate cell death, or processed for total RNA extraction and miRNAs
expression evaluation by quantitative Real Time-PCR. A) Representative Annexin V-APC/PI
stainings showing decreased cell death after TUDCA incubation. B) Quantitation of either dying
(Annexin+/PI-) or dead (Annexin+/PI+) cells depicted in FACS diagrams. Results are mean ±
SEM of triplicates. C) Expression of proapoptotic miRNAs at 3 and 6 days, with or without
TUDCA treatment. miR-16, let-7a and miR-34a expression were evaluated from 10 ng of total
RNA, using specific primers for each miRNA, and GAPDH for normalization. Expression levels
were calculated by the ∆∆Ct method using differentiated cells at 3 days as calibrator. Data
represent mean ± SEM of three independent experiments. *p < 0.05 compared to respective
nontreated cells at 6 days.
3.4.4 Apoptosis-associated miRNAs are upregulated in different models of neural
differentiation
To further validate the role of the proapoptotic miRNAs, miR-16, let-7a and miR-
34a in neural cell differentiation, we investigated whether they were upregulated in other
neural differentiation models, including mouse ES cells, PC12 and NT2N cell lines. The
generation of neurons from ES cells has reproducibly been characterized in vitro

Apoptosis-associated miRNAs and differentiation
81
(Abranches et al., 2009; Ying et al., 2003). Neural progenitors derived from ES cells
organize themselves into rosette-like structures that can subsequently differentiate into
neurons, astrocytes and oligodendrocytes. Using the same differentiation protocol,
neurogenesis was evidenced by increased Ngn2 and β-III tubulin mRNA expression
levels at 8 days of differentiation (Fig. 3.5A).
Figure 3.5. miR-16, let-7a and miR-34a are increased during mouse ES cell differentiation.
Mouse ES cells (Sox1-GFP 46C) were differentiated using an adherent monolayer protocol. Cells
at 4 and 8 days were collected for total RNA extraction and subsequently processed for evaluation
of specific differentiation markers, as well as proapoptotic miRNA expression by quantitative
Real Time-PCR. A positive control for neural differentiation was also performed at day 8 by
treating cells with either 10 nM LY411575 or 0.01%DMSO (control) for 12 hours. A) Semi-
quantitative RT-PCR analysis for selected markers of lineage commitment in day 4 and 8, as well

Chapter 3
82
as in LY411575-treated and untreated cells. B) Representative bright-field, phase contrast images
showing increased neuronal differentiation after LY411575 incubation compared with control
(DMSO-treated) cells. C) Expression of miR-16, Let-7a and miR-34a at 4 and 8 days of ES cell
differentiation and in control (DMSO-treated) and LY411575-treated rosette cultures at 8 days.
miRNAs expression were evaluated from 10 ng of total RNA, using specific primers for each
miRNA, and GAPDH for normalization. Expression levels were calculated by the ∆∆Ct method
using either differentiated cells at 4 days or LY411575-untreated cells as calibrator. Data
represent mean ± SEM of three independent experiments. *p < 0.05 compared to respective
nontreated cells. Scale bar: 50 µm. d, days.
The neural progenitor marker Nestin was also increased, suggesting a concomitant
increase in neural progenitors. In fact, at this stage of differentiation, expression levels of
miR-16, let-7a and miR-34a were increased when compared with day 4 (Fig. 3.5B). The
inhibition of Notch pathway at 8 days of differentiation by LY411575 treatment resulted
in increased neuronal production. In fact, neuronal markers, particularly Ngn2, were
increased, and morphological evaluation was consistent with the presence of
differentiated cells (Fig. 3.5A and C). Notably, LY411575-induced neurogenesis resulted
in a significant increased of miR-16 and miR-34a, by 3- and 2-fold (Fig. 3.5B),
respectively (p < 0.05), supporting the potential involvement of both miRNAs in neuronal
differentiation. Glial differentiation was not detected in the time points analyzed (data
not shown).
It has been previously described that differentiation of PC12 cells can be induced by
NGF (Greene and Tischler, 1976). Cell differentiation increases during the first 7 days of
treatment with NGF, as measured by neuronal outgrowth and mRNA levels of specific
neurofilaments (Schimmelpfeng et al., 2004). Moreover, this correlation decreases with
lower NGF concentrations. Our results demonstrated that treatment of PC12 cells with 50
ng/ml of NGF markedly induced miR-16 expression at 2 and 4 days, compared with
NGF-untreated cells (Fig. 3.6A). Indeed, miR-16 expression increased by 1.8- (p < 0.01)
and 1.4-fold (p < 0.05) at 2 and 4 days, respectively. Nevertheless, no difference in miR-
16 expression levels was detected at 7 days, under different NGF treatments.
Interestingly, 24 hours after treatment with 50 ng/ml of NGF, let-7a expression was ~ 5-
fold upregulated in differentiated PC12 cells (p < 0.05). Under identical conditions, let-
7a expression remained elevated throughout the time course (p < 0.05). Notably, miR-
34a expression was also gradually upregulated after NGF treatment. Although to a lesser

Apoptosis-associated miRNAs and differentiation
83
extent, similar results were obtained for all three miRNAs under suboptimal NGF 5 ng/ml
concentrations. None of the selected miRs were modulated in NGF-untreated cells.
Human teratocarcinoma NT2 cells generate postmitotic neurons (NT2N) in
response to retinoic acid and mitotic inhibitors (Hara et al., 2008). Retinoic acid
treatment of NT2 cells induced neuronal differentiation as evidenced by the expression of
the neuronal marker NeuroD at 14 and 21 days, as well as increased expression of β-III
tubulin (data not shown). In NT2N, miR-16 expression was increased by 3.6-fold at day
14 (p < 0.001) and almost 4-fold at day 21 (p < 0.05) of differentiation (Fig. 3.6B).
Similarly, let-7a was highly upregulated, particularly at 21 days of retinoic acid treatment
(p < 0.05). In contrast, miR-34a was not modulated during cell differentiation. Replated
cells showed increased differentiation, as evidenced by the increase in β-III tubulin-
positive cells and detection of NeuN-positive cells (data not shown). Surprisingly,
increased differentiation after replating was associated with a significant decrease in both
miR-16 and let-7a expression by ~ 2 (p < 0.05) and 5-fold (p < 0.001), while miR-34a
increased by 4.5-fold (p < 0.05), compared with non-replated cells.
Figure 3.6. Differentiation of PC12 and NT2N cells were associated with modulated levels of
miR-16, let-7a and miR-34a expression. PC12 cells were differentiated upon NGF treatment at

Chapter 3
84
1 day with either 5 or 50 ng/ml. NGF-untreated cells were also prepared. Cells were collected at
1, 2 4 and 7 days. NT2N cells were induced to differentiate with 10 µM all-trans retinoic acid and
collected at 0, 14 and 21 days. In parallel experiments, cells were replated at 14 days in Matrigel,
deprived of retinoic acid and incubated with mitotic inhibitors and further cultured for 7 days.
Control cells were maintained in initial retinoic acid treatment. Both PC12 and NT2N-collected
cells were processed for total RNA extraction and evaluation of miRNAs expression levels using
quantitative Real Time-PCR. A and B) miR-16, let-7a and miR-34a expression during PC12 and
NT2N differentiation, respectively. miRNAs expression were evaluated from 10 ng of total RNA,
using specific primers for each miRNA. GAPDH and RNU6B were used for normalization in
PC12 and NT2N cells, respectively. Expression levels were calculated by the ∆∆Ct method.
NGF-untreated cells at 1 day was used as calibrator in PC12 cells, while undifferentiated (day 0)
or unreplated cells were used as calibrator in NT2N cells. Data represent mean ± SEM of three
independent experiments. A) *p < 0.05 and †p < 0.01 compared to 1 day. miR-16 and let-7a
expressions in cells treated with 5 and 50 ng/ml NGF were also significantly different from NGF
untreated cells at day 4 (p < 0.05) and day 2 (p < 0.01), respectively. B) *p < 0.05 and §p < 0.001
compared to day 0; †p < 0.05 and ‡p < 0.001 compared to unreplated cells. d, days.
miR-34a was the most significantly upregulated miRNA during mouse NS cell
differentiation (~ 11-fold between days 3 and 6; p < 0.001) compared to other apoptosis-
associated miRNAs. Because miR-34a upregulation coincides with the onset of
postmitotic neurons, we investigated the effect of miR-34a upregulation in this population
by evaluating NeuN expression. We used flow cytometry to determine the proportion of
the postmitotic neuronal population. The proportion of NeuN-positive cells was
significantly increased when compared to cells transfected with unrelated RNA duplexes
(Fig. 3.7A). In fact, NeuN-positive cells increased from 29.7 to 38.9% 24 h after
transfection. This increase was further enhanced from 27.3 to 66.6% at 72 h post-
transfection. In addition, these results were confirmed by Western blot analysis (Fig.
3.7B). No changes were detected in the proportion of astrocytes in the pre-miR-34a
transfected cells (data not shown). These results indicated that miR-34a contributes to
neuronal differentiation in mouse NS cells.

Apoptosis-associated miRNAs and differentiation
85
Figure 3.7. miR-34a overexpression increases the ratio of postmitotic neurons in mouse NS
cells. Mouse NS cells were transfected using 100 nM of either control or pre-miR-34a, and
collected 24 and 72 h after transfection. Collected cells were labeled for NeuN detection by flow
cytometry, or processed for total protein extraction and NeuN expression evaluation by Westen
blot. A) Percentage of NeuN positive cells in negative control and pre-miR-34a-transfected cells,
24 and 72 h after transfection. In independent repetitions of these experiments, similar increases
in the relative number of NeuN-positive cells were observed in cultures overexpressing miR-34a
when compared to control pre-miR-transfected cells. B) Representative blot showing increased
NeuN expression in pre-miR-34a transfected cells.
3.5 Discussion
Differentiation of NS and ES cells in vitro has attracted wide interest as an
experimental system for investigating specific molecular pathways of cell development.
Here, we have demonstrated neuronal and glial conversion of mouse NS cells. In this
model, the neuronal population is formed mainly during the first 2 days, as indicated by
the upregulation of β-III tubulin. Nevertheless, commitment to mature neurons was
confirmed by the expression of NeuN at 6 days, which indicates the presence of immature
postmitotic neurons. Astrocytic population peaked at 6 days, and this was maintained
throughout differentiation.
Functional studies of apoptosis-associated miRNAs have focused intensely on
cancer research. However, our results demonstrate that specific miRNAs regulating pro-
and antiapoptotic genes may also participate in differentiation processes. The
involvement of miR-16 in cell differentiation has not been previously reported. Indeed,

Chapter 3
86
miR-16 expression was markedly increased throughout mouse, rat and human neural
differentiation. Nevertheless, the onset of miR-16 expression during mouse NS cell
differentiation was not associated with the appearance of any specific cell type. miR-16
was shown to be implicated in cell cycle regulation, as well as apoptosis induction by
targeting Bcl-2 (Chen et al., 2008; Cimmino et al., 2005; Linsley et al., 2007; Liu et al.,
2008). Apoptosis is not markedly regulated during mouse NS cell differentiation, and
Bcl-2 expression is increased (data not shown). Therefore, a possible role for miR-16 in
the context of differentiation may be associated with cell cycle control in downregulating
the proliferation potential of differentiating cells. During differentiation of neural
precursors, a tightly coordinated regulation of cell cycle exit is crucial for the generation
of appropriate number of neurons and proper wiring of neuronal circuits (Politis et al.,
2008). Nevertheless, here-to-fore unrecognized miR-16 targets may exert distinct, yet
crucial functions during cell differentiation.
The let-7 family consists of eleven very closely related genes (Pasquinelli et al.,
2000). They are highly conserved in animals from worms to humans, and their
expression increases after differentiation and maturation of tissues (Lee et al., 2005). In
addition to regulating apoptosis by targeting caspase-3 (Tsang and Kwok, 2008), it was
also demonstrated that let-7 family members regulate RAS and HMGA2 oncogene
through the 3’UTR (Akao et al., 2006; Johnson et al., 2005; Lee and Dutta, 2007). In the
present study, let-7a expression was shown to be cyclic during mouse NS cells
differentiation, and corresponded to the onset of neurogenesis and gliogenesis. This
suggests that let-7a expression is associated with a general mechanism of differentiation
rather than differentiation of specific cell types. Our results are supported by the previous
report showing that let-7a is a critical regulator of neuronal differentiation (Liu et al.,
2009; Sempere et al., 2004). In fact, increased let-7a expression during ES cells, PC12
and NT2N differentiation also underscores the important role of let-7a during general
differentiation. Additional studies are warranted to evaluate the mechanism(s) by which
let-7a regulates cell differentiation. Importantly, although highly modulated during cell
differentiation, both let-7a and miR-16 were significantly expressed in neurospheres (data
not shown). Therefore, it is possible that additional mechanisms exist to antagonize let-
7a and miR-16 expression during NS cell differentiation.
A specific role for miR-34a during neuronal differentiation has not been reported.
Interestingly, herein we showed a significant upregulation of miR-34a during mouse NS
cell differentiation, and this paralleled the appearance of postmitotic neurons. In addition,

Apoptosis-associated miRNAs and differentiation
87
in vitro transient overexpression of miR-34a increased the proportion of NeuN-positive
cells. Given that miR-34a has been shown to regulate genes involved in cell cycle arrest,
it is possible that miR-34a upregulation is related to cell cycle exit and the subsequent
appearance of immature postmitotic neurons. In fact, miR-34a upregulation in ES cells
after LY treatment, as well as in NT2N cells after incubation with the mitosis inhibitor,
supports this hypothesis. In addition, it has been previously shown that miR-34a can
suppress cell-cycle genes and induce neural phenotype in neuroblastoma tumors (Wei et
al., 2008). Importantly, it has been previously reported that miR-34a overexpression had
no effect on cell cycle arrest and survival/apoptosis of astrocytes (Guessous et al., 2010;
Li et al., 2009a). However, additional studies are required to confirm the influence of
miR-34a in neurogenesis and evaluate whether it mediates p53 effects on cell
differentiation. In fact, several potential miR-34a targets might be involved in the
transition towards postmitotic neurons. It was previously demonstrated that miR-34a
targets Sirt1 and is regulated by p53, which in turn, is activated by Sirt1 suppression. In
addition, miR-34a may also regulate the differentiation process by influencing Notch
signaling pathway (Guessous et al., 2010; Hermeking, 2010). In contrast to let-7a and
miR-16, miR-34a was barely detected in undifferentiated cells, supporting its specific
involvement in cell differentiation.
miR-19a and miR-20a are members of the miR-17-92 cluster (Bonauer and
Dimmeler, 2009), which consists of seven mature miRNAs, previously linked to
tumorigenesis. Recently, additional functions have been assigned to this cluster,
particularly to miR-20a and miR-19a. Specifically, miR-20a was shown to control
monocyte differentiation (Fontana et al., 2007). In fact, transfection of hematopoietic
progenitors with miR-20a increased the proliferation of monocytes and blocked
differentiation, whereas inhibition of miR-20a caused a decrease in proliferation and more
rapid differentiation and maturation. Our results, showing that miR-20a decreases during
mouse NS cell differentiation, are in agreement with this previous report.
Only limited information is available regarding the physiological role of miR-19a.
It has been demonstrated that it acts as a positive regulator of antiapoptotic Stat3/IL-6
receptor signaling by directly suppressing SOCS-1, thereby facilitating malignant growth
of multiple myeloma (Pichiorri et al., 2008). Further, miR-19 appears to affect the level
of proapoptotic protein Bim, thereby preventing apoptosis and promoting cell survival.
The possible role of miR-19a on cell survival, may explain why this miRNA was
upregulated at early stages after the induction of mouse NS cell differentiation. However,

Chapter 3
88
additional studies are required to determine the specific role of both miR-20a and miR-
19a during cell differentiation, and also evaluate if their expression is restricted to a
specific cell type.
In conclusion, our results demonstrate that apoptosis-associated miRNAs are
differentially expressed during neural differentiation, in the absence of cell death, and
identify miR-16, let-7a and miR-34a as important players.
Acknowledgments
We would like to acknowledge Yan Zeng (University of Minnesota Medical
School, Minneapolis, Minnesota, USA) for miRNA expression profiling, and Rui Castro
(iMed.UL, Lisbon, Portugal) for microarray data analysis. The authors thank Ricardo
Viana and Inês Milagre (iMed.UL, Lisbon, Portugal) for providing PC12 and NT2N cell
cultures, and Domingos Henrique (IMM, Lisbon, Portugal) for ES cell lines. This work
was supported by grant PTDC/BIA-BCM/67922/2006 from Fundação para a Ciência e a
Tecnologia (FCT), Lisbon, Portugal. MM Aranha and DM Santos are recipients of PhD
fellowships from FCT, Portugal (SFRH/BD/28429/2006 and SFRH/BD/42008/2007,
respectively).

4 miR-34a regulates mouse neural stem cell differentiation
Márcia M. Aranha1, Susana Solá1, Daniela M. Santos1, Walter C. Low2,3, Clifford J.
Steer4,5, Cecília M.P. Rodrigues1
1Research Institute for Medicines and Pharmaceutical Sciences, Faculty of Pharmacy,
University of Lisbon, Lisbon, Portugal; 2Department of Neurosurgery, 3Stem Cell
Institute, 4Department of Medicine, and 5Department of Genetics, Cell Biology, and
Development, University of Minnesota Medical School, Minneapolis, Minnesota, USA
Manuscript in preparation.


miR-34a regulates neural differentiation
91
4.1 Abstract
MicroRNAs (miRNAs or miRs) participate in the regulation of several biological
processes, including cell differentiation. Recently, miR-34a has been implicated in
differentiation of monocyte-derived dendritic cells, human erythroleukemia cells, and
mouse embryonic stem cells. In addition, members of the miR-34 family have been
described as direct p53 targets. However, the function of miR-34a in the control of the
differentiation program of specific neural cell types remains largely unknown. Here, we
investigated the role of miR-34a in regulating mouse neural stem (NS) cell
differentiation. miR-34a overexpression increased the proportion of postmitotic neurons
of mouse NS cells, whereas miR-34a downregulation had the opposite effect. SIRT1 was
identified as a target of miR-34a and shown to mediate miR-34a effects on neuronal
differentiation. In addition, p53-DNA binding activity was increased and cell death
unchanged after miR-34a overexpression, thus reinforcing the role of p53 during neural
differentiation. Interestingly, in conditions where SIRT1 was activated by pharmacologic
treatment with resveratrol, miR-34a overexpression increased the proportion of astrocytes
through a SIRT1-independent mechanism. In conclusion, our results extend the
molecular mechanism by which miR-34a modulates neural differentiation, suggesting that
miR-34a is required for proper neuronal differentiation, in part, by targeting SIRT1 and
affecting p53 activity.

Chapter 4
92
4.2 Introduction
MicroRNAs (miRNAs or miRs) are small, ~ 21-nucleotide-long regulatory RNA
molecules encoded in plant and animal genomes. miRNAs regulate the expression of
target genes by binding to the 3’-untranslated regions of specific mRNAs and triggering
mRNA destabilization or suppression of translation (Bartel, 2004). They fine-tune gene
expression by effecting more subtle and rapid changes than global transcriptional control
mechanisms (Shivdasani, 2006). Each miRNA have the capacity to regulate multiple
genes; in mammals, miRNAs are predicted to control the activity of ~ 50% of all protein-
coding genes (Krol et al., 2010).
Functional studies indicate that miRNAs participate in the regulation of almost
every cellular processes investigated so far, including cell differentiation. The important
regulatory role of miRNAs in development and differentiation clearly emerged from the
study of embryonic stem cells null for the Dicer gene, which encodes an RNase III
required for miRNA biogenesis. Ablation of Dicer leads to defects in embryonic stem
cell division and proliferation (Murchison et al., 2005), causing death in mice and
complete loss of pluripotent stem cells (Bernstein et al., 2003). In addition, Dicer ablated
embryonic stem cells also fail to differentiate. Therefore, miRNAs can affect stem cell
fate at the levels of both proliferation and differentiation.
miR-34a is a member of the miR-34 family which comprises three processed
miRNAs in mammals: miR-34a, -34b and -34c (Hermeking, 2010). miR-34a is encoded
by its own transcript, whereas miR-34b and miR-34c share a common primary transcript.
In mice, miR-34a is ubiquitously expressed with the highest expression in the brain
(Lodygin et al., 2008), whereas miR-34b and c are mainly expressed in lung tissue
(Bommer et al., 2007). Importantly, several reports showed that members of the miR-34
family are direct p53 targets, and their upregulation induces apoptosis and cell cycle
arrest (Bommer et al., 2007; Chang et al., 2007b; Corney et al., 2007; He et al., 2007a;
Raver-Shapira et al., 2007; Tarasov et al., 2007). However, emerging evidence suggests a
role for miR-34a during cell differentiation. miR-34a was shown to be involved in
monocyte-derived dendritic cell differentiation by targeting JAG1 (Hashimi et al., 2009).
In addition, miR-34a regulates phorbol ester-induced megakaryocytic differentiation of
K562 cells by inhibiting cell proliferation and inducing cell cycle arrest, through direct
regulation of its known targets cyclin-dependent kinase 4 and 6 (CDK4 and CDK6),
MYB and mitogen-activated protein kinase kinase 1 (MEK1) (Ichimura et al., 2010;
Navarro et al., 2009). Recently, miR-34a was shown to modulate mouse embryonic stem

miR-34a regulates neural differentiation
93
cell differentiation by regulating silent information regulator 1 (SIRT1) expression levels
(Tarantino et al., 2010). Nevertheless, the precise role of miR-34a during neuronal
differentiation has never been reported.
SIRT1 is a NAD-dependent deacetylase involved in cellular resistance to stress,
metabolism, aging, and tumor suppression (Blander and Guarente, 2004; Finkel et al.,
2009; Haigis and Guarente, 2006; Mantel and Broxmeyer, 2008). In mammals, SIRT1
function is mediated by its deacetylating activity not only on histones, but also on key
transcription factors, such as p53 (p53), forkhead transcription factors (FOXO), p300
histone acetyltransferase, tumor suppressor protein p73 (p73), E2F transcription factor 1
(E2F1), DNA repair factor Ku antigen 70-kDa subunit (Ku70), nuclear factor κ-B (NF-
κB), and androgen receptor (AR) (for review, see (Guarente and Picard, 2005). In
addition, SIRT1 knockout mice demonstrated embryonic and postnatal developmental
defects (Wang et al., 2008b), suggesting that SIRT1 is important in embryonic
development. Indeed, SIRT1 has been identified as a redox sensor in mouse embryonic
fibroblasts and undifferentiated muscle cells, in which it controls proliferation, cell cycle
arrest and differentiation (Brunet et al., 2004; Fulco et al., 2003). SIRT1 was also
reported to play a role in cellular differentiation of human keratinocytes (Blander et al.,
2009), in mouse embryonic stem cell differentiation and hematopoiesis (Ou et al., 2010),
and in differentiation of neural progenitors (Hisahara et al., 2008; Prozorovski et al.,
2008).
Here, we identified the role of miR-34a in regulating mouse neural stem (NS) cell
differentiation by a mechanism that involves SIRT1 downregulation and increased p53-
DNA binding activity.
4.3 Methods
4.3.1 Cell Culture
Mouse NS cells containing a constitutively expressed marker for green fluorescent
protein (GFP) were used to investigate the process of neural differentiation. Primary cells
were obtained from central nervous system tissue of embryonic mice, and cultured
primarily as previously described (Reynolds and Weiss, 1992, 1996; Rietze and
Reynolds, 2006). Mouse NS cells were maintained as neurospheres in undifferentiation
medium, serum-free, 1:1 mix of DMEM/F12 (Invitrogen Corp., Grand Island, NY) with
1× N-2 supplement (Invitrogen Corp.), 20 ng/ml epidermal growth factor (EGF) (R & D

Chapter 4
94
Systems Inc., Minneapolis, MN), 20 ng/ml basic fibroblast growth factor (bFGF)
(PeproTech EC, London, UK), and 1% penicillin-streptomycin (Invitrogen Corp.), at
37ºC in humidified atmosphere of 5% CO2. Subculture occurred at day 7 with
mechanical dissociation of neurospheres. The differentiation of mouse NS cells in vitro
was induced by culturing dissociated cells in differentiation medium containing
DMEM/F12 with 1× N-2 supplement, 100 ng/ml bFGF, 10% fetal bovine serum (FBS)
(Invitrogen Corp.), 500 nM all-trans retinoic acid (Sigma Chemical Co., St. Louis, MO),
50 µM taurine (Sigma Chemical Co.), 10 ng/ml transforming growth factor-β2 (TGF-β2)
(R & D Systems Inc.) and 1% penicillin-streptomycin in tissue culture plates pre-coated
with poly-D-lysine (Sigma Chemical Co.). The culture medium was changed every 3
days. Differentiated cells at 5 × 105 cells/ml were processed for immunobloting assays.
Transfections and cell treatments were carried out at 2.5 × 105 cells/ml cellular density.
4.3.2 Small interfering RNA transfections, overexpression/antisense experiments
and cell treatments
miR-34a expression was modulated using 100 nM of pre-anti-miR negative control
and pre-anti-miR-34a (Applied Biosystems, Foster City, CA). The sequences of small
interfering RNA (siRNA) duplex for SIRT1 were sense
5’-GUGAGAAAAUGCUGGCCUA-3’ and antisense 5’-UAGGCCAGCAUUU
UCUCAC-3’ Cells were transfected using 100 nM of SIRT1 siRNA or treated with 5 µM
resveratrol. Mouse NS cells were transfected at 2 or 3 days of differentiation using
Lipofectamine 2000 (Invitrogen Corp) following manufacturer’s instructions.
Efficiencies of miR-34a modulation and SIRT1 silencing were assessed by quantitative
real-time RT-PCR and immunoblotting, respectively.
4.3.3 Evaluation of miR-34a expression levels by quantitative Real Time-PCR
Real-time PCR was performed in an Applied Biosystems 7300 Sequence Detection
System (Applied Biosystems, Foster City, CA). Ten ng of total RNA isolated by TRIzol
reagent (Invitrogen Corp.) were reverse transcribed using a TaqMan® MicroRNA
Reverse Transcription (RT) kit from Applied Biosystems. Each RT reaction contained 1x
stem-loop RT specific primer, 1x reaction buffer, 0.25 mM each of dNTPs, 3.33 U/µl
Multiscribe RT enzyme and 0.25 U/µl RNase inhibitor. The 15-µl reactions were
incubated for 30 min at 16ºC, 30 min at 42ºC, and 5 min at 85ºC and then held at 4ºC.
The PCR reaction was performed using a standard TaqMan® PCR kit protocol (Applied

miR-34a regulates neural differentiation
95
Biosystems). Briefly, following the RT step, 1.33 µl of the RT reaction were combined
with 1 µl of a TaqMAn MicroRNA Assay (20x; forward primer, reverse primer and
probe) and 17.67 µl of TaqMan® Universal PCR Master Mix, No AmpErase® UNG in a
20 µl final volume. The reactions were incubated at 95°C for 10 min, followed by 40
cycles of 95°C for 15 s and 60°C for 1 min; and were run in triplicate. miRNA
expression levels relative to GAPDH were calculated on the basis of ΔΔCt method. The
n-fold change in miRNAs expression was determined according to the method of 2-ΔΔCT.
4.3.4 Flow cytometry analysis of Nestin, β-III Tubulin, NeuN and GFAP
Cells were trypsinized (0.025% trypsin/EDTA; Invitrogen Corp.) and harvested in
Ca2+-free and Mg2+-free PBS and 2% FBS. After washing, 0.5 - 1 × 106 cells were fixed
with paraformaldehyde (4% w/v) in PBS for 20 min on wet ice, and blocked for 20 min in
PBS, containing 0.25% saponin (Fluka, Biochemika, Switzerland) and 5% normal donkey
serum (Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Subsequently,
cells were incubated with antibodies reactive to Nestin (MAB 353), neuronal nuclei
(NeuN) (MAB377), glial fibrillary acidic protein (GFAP) (MAB360; Chemicon
International, Temecula, CA ) and β-III Tubulin (Tuj1) at a dilution of 1:100, 1:20,
1:2000 and 1:500, respectively, in PBS containing 0.1% saponin and 5% normal donkey
serum, for 30 min. Cells were then incubated with anti-mouse antibody conjugated to
Cy5 (Jackson ImmunoResearch Laboratories, Inc.), for 30 min. Cells were analyzed
using a FACSCalibur (Becton Dickinson, Mountain View, CA).
4.3.5 Immunoblotting
Steady-state levels of SIRT1 were determined by immunoblotting. Total protein
extracts of mouse NS cells were prepared in lysis buffer, following standard established
protocols. Protein content was measured by the Bio-Rad protein assay kit according to
the manufacture’s specification, using bovine serum albumin as standard. 50-80 µg of
total protein extracts were separated on 12% sodium dodecyl sulphate-polyacrylamide
electrophoresis gel and then subjected to immunoblots using primary rabbit polyclonal
antibody reactive to SIRT1 (07-131, Chemicon International). Blots were subsequently
incubated with secondary antibodies conjugated with horseradish peroxidase (Bio-Rad
Laboratories, Hercules, CA, USA). Finally, membranes were processed for protein

Chapter 4
96
detection using Immobilon (Millipore Corporation, Billerica, MA) or SuperSignal reagent
(Pierce, Rockford, IL). Ponceau S staining was used to assess equal gel loading
(Romero-Calvo et al., 2010).
4.3.6 Immunocytochemistry
Mouse NS cells were fixed with paraformaldehyde (4% w/v) in phosphate-buffered
saline (PBS) and blocked for 1 hour at room temperature in PBS containing 0.1% Triton-
X-100, 1% FBS, and 10% normal donkey serum (Jackson ImmunoResearch Laboratories,
Inc., West Grove, PA). Subsequently, cells were incubated with either anti-SIRT1
(rabbit, polyclonal), anti-Nestin, anti-β-III Tubulin or anti-GFAP (mouse, monoclonal)
antibodies (Chemicon International) at a dilution of 1:200, 1:100, 1:500 and 1:1000,
respectively, in blocking solution, overnight at 4ºC. Cells were then incubated with either
anti-mouse or anti-rabbit IgG conjugated to AMCA (Jackson ImmunoResearch
Laboratories, Inc.) and Alexa 594 (Molecular Probes-Invitrogen), respectively, for 2 h at
room temperature. For GFAP detection, an Alexa 568-conjugated anti-mouse antibody
(Molecular Probes-Invitrogen) was used. Samples were mounted using Fluoromount-
G™ (Beckman Coulter, Inc.). Fluorescence microscopy assessments were performed
with a Zeizz AX10 microscope (Carl Zeiss, Jena, Germany) equipped with a Leica
DFC490 camera (Leica Weitzlar, Germany).
4.3.7 Nuclear protein extraction
Nuclear extracts from p53- and miR-34a-overexpressing cells were prepared as
previously described (Schreiber et al., 1989). p53-overexpressing cells were obtained by
transfecting 8 ug of wild-type p53 (pCMV-p53wt).
4.3.8 Electrophoretic mobility shift assay
Electrophoretic mobility shift assay (EMSA) was performed as previously
described (Milagre et al., 2008). DNA probe and competitors were generated by
hybridization with complementary single-stranded oligonucleotides and used in double-
stranded form (supplementary Table 1). Briefly, 8 µg nuclear extracts were incubated
with γ32P-labeled oligonucleotide probe (p53-cons) containing a p53 consensus binding
site for 20 min. In competition assays, excess unlabeled oligonucleotide was pre-

miR-34a regulates neural differentiation
97
incubated for 30 min, prior to incubation with the probe p53-cons for additional 20 min.
Super-shift reactions were performed with 1 µl of anti-p53 (DO-1 X, Santa Cruz
Biotechnology), which was added to the reaction media after incubation of the probe.
Protein-DNA complexes were resolved on 5% non-denaturing PAGE gels (29:1
acrylamide:bisacrylamide) in 0.5 × Tris-borate buffer (45 mM Tris-borate, 1 mM EDTA).
The gels were eletrophoresed for 2 h and 30 m, at 20 mA.
4.3.9 Cytofluorometric analysis of apoptosis
Apoptotic cells were quantified by cytofluorometric analysis using a FACSCalibur
(Becton Dickinson, Mountain View, CA) as described previously (Schutte et al., 1998).
Cells were stained with the vital dye propidium iodide (PI; 5µg/mL; Sigma, Steinheim,
Germany) and concomitantly with Annexin-V-APC (eBioscience, Inc.) according to
manufacturer’s instructions to determine phosphatidylserine exposure. Data were
statistically evaluated using FlowJo software (Tree Star, Inc, Ashland, OR).
4.3.10 Statistical Analysis
Results from different groups were compared using the Student’s t test, or one-way
ANOVA. Values of p < 0.05 were considered statistically significant. All statistical
analysis was performed with GraphPad InStat software (GraphPad Software, Inc, San
Diego, CA).
4.4 Results
4.4.1 miR-34a modulates the proportion of postmitotic neurons
We have previously reported that induction of mouse NS cell differentiation results
in a mixed cell population composed by both neuronal and glial cells, which in turn
differentiate in a time-controlled fashion within 8 days (Aranha et al., 2009). In addition,
we have shown that miR-34a expression is increased during mouse NS cell differentiation
concomitantly with the appearance of postmitotic neurons and astrocytes (Aranha et al.,
2010). To determine the effect of miR-34a on neural differentiation, we explored the
effect of miR-34a modulation on the percentage of neural progenitors (Nestin+), neuronal
precursors (β-III Tubulin+), postmitotic neurons (NeuN+) and astrocytes (GFAP+). Pre-
and anti-miR-34a, as well as respective controls, were transfected into differentiating

Chapter 4
98
mouse NS cells at 3 days. miR-34a expression was analyzed by real-time RT-PCR 24 h
later, to confirm its functional modulation (data not shown). Flow cytometry analysis
revealed substantial alterations in NeuN expression, when cells were transfected with
either anti- or pre-miR-34a (Fig. 4.1A).
Figure 4.1. miR-34a modulates the proportion of NeuN-positive cells. Mouse NS cells were
transfected using 100 nM of either anti- or pre-miR-34a at 3 days, and collected after 24 and 72 h,
respectively. Cells were subsequently labeled for Nestin, β-III Tubulin, NeuN and GFAP
detection by flow cytometry. A) Representative histograms of Nestin, β-III Tubulin, NeuN and
GFAP detection in anti-miR-control (red line) and anti-miR-34a-transfected cultures (blue line)
(top), or in pre-miR-control (red line) and pre-miR-34a-transfected cultures (blue line) (bottom).

miR-34a regulates neural differentiation
99
In four independent repetitions of these experiments, similar alterations in the relative number of
positive cells for each marker were observed. B) Fold change increase in NeuN-positive cells
after miR-34a modulation. Results are shown as mean ± SEM. *p < 0.05 from cells transfected
with respective control.
Notably, transfections with anti-miR-34a for 24 h reduced the proportion of NeuN+
cells by 26%, compared with cells transfected with an anti-miR negative control (p <
0.05). Further, the percentage of NeuN+ cells significantly increased by 93% after pre-
miR-34a transfection for 72 h (p < 0.05) (Fig. 4.1B). Modulation of miR-34a had no
effect on the number of Nestin+ and β-III Tubulin+, while GFAP+ cells were only slightly
affected. Importantly, cell counting and apoptosis assessment suggested that alterations
in the number of NeuN+ cells are not related with altered cell number or increased cell
death (data not shown).
4.4.2 SIRT1 expression is modulated by miR-34a during neural differentiation
Several signaling pathways can be potentially affected by miR-34a, and implicated
in the transition of dividing neuronal precursors to immature postmitotic neurons. SIRT1
has recently been suggested to be a target of miR-34a during embryonic stem cell
differentiation (Tarantino et al., 2010). In addition, SIRT1 expression was shown to be
downregulated during differentiation of human embryonic stem cells (Calvanese et al.,
2010). To investigate the role of SIRT1 during miR-34a-mediated neural differentiation,
we analyzed SIRT1 expression throughout the differentiation period. Immunoblot
analysis revealed that SIRT1 was highly expressed in undifferentiated neurospheres and
also in differentiating mouse NS cells during the first 2 days. Notably, SIRT1 expression
markedly decreased from the third day of differentiation onwards (Fig. 4.2A). Curiously,
decreased SIRT1 expression at day 3 is concomitant with previously described miR-34a
upregulation (Aranha et al., 2010). Immunofluorescence staining of SIRT1 at 2 days of
differentiation revealed that SIRT1 staining was mostly nuclear and present in distinct
cell populations, including Nestin+ and β-III Tubulin+ cells (Fig. 4.2B). Next, we
evaluated whether overexpression of miR-34a resulted in decreased SIRT1 at the protein
level. SIRT1 protein was significantly decreased after miR-34a overexpression (p < 0.05)
(Fig. 4.2C), suggesting that the effect of miR-34a on the number of postmitotic neurons is
mediated by SIRT1.

Chapter 4
100
Figure 4.2. SIRT1 expression levels decrease after miR-34a overexpression. SIRT1
expression in mouse NS cells was detected by immunoblotting at different times of differentiation
or by immunocytochemistry at 2 days of differentiation. Cells transfected with either control or
pre-miR-34a at 3 days of differentiation were also processed for SIRT1 detection by
immunoblotting. A) Representative immunoblot showing a marked decrease in SIRT1 expression
levels from day 3 of differentiation onwards. Ponceau S was used as loading control. B)
Immunofluorescence detection of cells labeled with anti-SIRT1, anti-Nestin and anti-β-III tubulin
antibodies to visualize SIRT1 expression in neural progenitors and neuronal precursors,
respectively. Scale bar: 10 µm. C) Representative immunoblot (top) and corresponding
densitometry analysis (bottom) showing decreased SIRT1 expression in pre-miR-34a transfected
cells. Data represent mean ± SEM of three independent experiments. *p < 0.05 from cells
transfected with control.
4.4.3 miR-34a regulates neuronal differentiation through a SIRT1-mediated
mechanism
To further confirm if SIRT1 participates in miR-34a-mediated neural
differentiation, we determined whether SIRT1 modulation affects the percentage of

miR-34a regulates neural differentiation
101
postmitotic neurons. To address this question, cells were either incubated with
resveratrol, a pharmacologic SIRT1 activator, or transfected with SIRT1 siRNA, at 12 h
after incubation in differentiation medium. Changes in percentage of NeuN+ cells were
evaluated by flow cytometry at 3 days of differentiation. SIRT1 silencing efficiency was
confirmed by immunoblotting at 24 h after transfection (Fig. 4.3A).
Figure 4.3. miR-34a promotes the appearance of post-mitotic neurons by downregulating
SIRT1 expression. SIRT1 expression was modulated in mouse NS cells at 12 h after induction
of differentiation. Inhibition of SIRT1 was achieved by transfecting cells with either scrambled
control or 100 nM of SIRT1 siRNA, while activation of SIRT1 was performed by treating cells
with either DMSO (control) or 5 µM of resveratrol. In parallel experiments, miR-34a expression
was modulated using anti- or pre-miR-34a and respective controls at 100 nM. Cells were

Chapter 4
102
collected at 3 days, fixed and processed for NeuN labeling and detection by flow cytometry. The
blue line corresponds to SIRT1 and/or miR-34a modulation, while the red line corresponds to the
respective control. A) Representative immunoblot showing decreased SIRT1 expression after
transfection with SIRT1 siRNA. Ponceau S was used as loading control. B) Silencing of SIRT1
increased the percentage of NeuN+ cells. C) Representative fluorescence microscopy image of
mouse NS cells expressing GFP after SIRT1 downregulation. D) Increased percentage of NeuN+
cells after simultaneous downregulation of miR-34a and SIRT1. E) Decreased proportion of
NeuN+ cells after resveratrol treatment (left) and increased percentage of NeuN+ cells with
simultaneous SIRT1 activation and miR-34a overexpression (right). Data represent mean ± SEM
of at least three independent experiments. *p < 0.05 from cells transfected with respective
control. Scale bar: 40 µm.
Interestingly, downregulation of SIRT1 increased the proportion of NeuN+ cells
(Fig. 4.3B), confirmed by morphological evaluation (Fig. 4.3C). Simultaneous silencing
of miR-34a and SIRT1 resulted in a similar increase by ~ 25% in the percentage of
NeuN+ cells when compared with control transfected cells ( p < 0.05) (Fig. 4.3D). The
effect of SIRT1 activation on NeuN expression was less pronounced, although still
significant (p < 0.05) (Fig. 4.3E, left). Importantly, NeuN+ cells increased by almost 40%
when cells were simultaneously treated with resveratrol and transfected with pre-miR-34a
(p < 0.05) (Fig. 4.3E, right). These results suggest that miR-34a regulates the appearance
of postmitotic neurons by targeting SIRT1 mRNA.
4.4.4 Astrogliogenesis can be modulated by miR-34a through a SIRT1-
independent mechanism
Transient transfection of miR-34a into mouse NS cells had a minor effect on
astroglial subpopulation (Fig. 4.1A). Nevertheless, SIRT1 knockdown consistently
decreased the percentage of GFAP+ cells by ~ 12% (p < 0.001), while activation of
SIRT1 increased GFAP+ cells by ~ 45% (p < 0.05) (Fig. 4.4A). After simultaneous
silencing of SIRT1 and miR-34a, the proportion of GFAP+ cells was no longer down-
regulated (Fig. 4.4B) Interestingly, overexpression of miR-34a and simultaneous
treatment with resveratrol resulted in a significant increase in GFAP+ cells by ~ 3.4 fold
(p < 0.05) (Fig. 4.4C). Immunofluorescence analysis of GFAP corroborated this
observation (Fig. 4.4D). Therefore, resveratrol treatment further enhanced the effect of
miR-34a modulation. As miR-34a negatively regulates SIRT1 expression, we would

miR-34a regulates neural differentiation
103
expect to have an opposite effect of miR-34a on GFAP+ cells. Thus, our results indicate
that SIRT1 positively regulates the astrocytic subpopulation. Moreover, under certain
conditions, miR-34a regulates astrocytic differentiation through a SIRT1-independent
mechanism.
Figure 4.4. miR-34a positively regulates the astrocytic population under resveratrol
treatment. SIRT1 expression was modulated in mouse NS cells after 12 h of induction of
differentiation. Inhibition of SIRT1 was achieved by transfecting cells with either scrambled
control or 100 nM of SIRT1 siRNA. SIRT1 was activated by treating cells with either DMSO
(control) or 5 µM of resveratrol. In parallel experiments, miR-34a expression was also
modulated. Cells were collected at 3 days, fixed and processed for GFAP labeling and detection
by flow cytometry and immunofluorescence. The blue line corresponds to SIRT1 and/or miR-34a
modulation, while the red line represents the respective control. A) Decreased percentage of

Chapter 4
104
GFAP+ cells after downregulation of SIRT1 (left), and increased percentage of GFAP+ after
SIRT1 activation (right). B) Simultaneous inhibition of SIRT1 and miR-34a reversed the
phenotype observed after SIRT1 silencing alone. C) Under resveratrol treatment, overexpression
of miR-34a resulted in increased proportion of GFAP+ cells. D) Immunofluorescence showing
increased number of GFAP+ cells under resveratrol treatment and miR-34a overexpression
(bottom) when compared with controls (top). Scale bar: 40 µm. Images taken with a
magnification of 400 × are shown in detail. *p < 0.05 and §p < 0.001 from cells transfected with
respective control.
4.4.5 miR-34a upregulation increases p53-DNA binding activity
SIRT1 has been shown to interact with and deacetylate the p53 protein, leading to a
suppression in its activity (Langley et al., 2002). The induction of miR-34a during mouse
NS cell differentiation and the concomitant decrease of SIRT1 suggest that p53 may be
involved in this regulation. To test this hypothesis, we upregulated miR-34a expression
levels and evaluated p53-DNA binding activity by EMSA. To verify the specificity of
the complex(es) formed when a p53 consensus oligonucleotide (p53-cons) was used as a
probe, we performed a supershift assay using an anti-p53 antibody, as well as competition
assays using oligonucleotides with different affinities for p53 binding, in mouse NS cells
overexpressing p53. As shown in Fig. 4.5A, nuclear extracts from mouse NS cells
overexpressing p53 exhibited very strong DNA-binding activity. In the presence of the
p53 antibody, there was a shift of the specific complex, resulting in slower-migrating
bands (Fig. 4.5A, arrowheads). Competition assays using 10 and 100-fold molar excess
of each competitor resulted in reduced efficiency of DNA binding, according to their
affinity for p53. In fact, competitor p53-A contains two quarter-sites known to be
consensus sites for p53 (Wang et al., 1995), while p53-B contains only one. Therefore,
the efficiency of competition was proportional to the number of repeats. As expected,
competition with unlabeled p53-cons resulted in a significant decrease in the efficiency of
DNA binding, and this effect was completely abolished when using p53-cons bearing a
mutation in the consensus site. Indeed, the formation of p53/p53-cons complex was not
competed by an unrelated DNA sequence. Importantly, the EMSA results show that total
nuclear proteins capable of binding to the p53-cons probe markedly increase under miR-
34a overexpression (Fig. 4.5B). These results suggest that miR-34a overexpression
increases p53-DNA binding activity. Notably, this was not associated with increased cell
death, as evaluated by PI/Annexin staining (Fig. 4.5C).

miR-34a regulates neural differentiation
105
Figure 4.5. miR-34a overexpression increases p53-DNA binding activity. Mouse NS cells
with 3 days of differentiation were transfected with either pre-miR-control or pre-miR-34a for 72
h. Cells were then collected for nuclear protein extraction or stained with Annexin-V-APC/PI to
evaluate cell death. p53-overexpressing cells were used for supershift and competition
experiments. Radio-labeled double-stranded oligonucleotide corresponding to the p53 consensus
(p53 -cons) was used as a probe. A) Representative EMSA showing the specificity of the
complex formed with p53-cons probe. Supershift experiments were performed using an anti-p53
antibody (DO-1, Santa Cruz Biotechnology). Competition experiments were performed by
adding 10- or 100-fold excess of unlabeled double-stranded oligonucleotides bearing a mutation
in the consensus site (p53-cons-mut), or containing either two or one of quarter-sites known to be

Chapter 4
106
consensus sites for p53 (p53-A and p53-B, respectively) or a non-specific sequence (NS). B)
EMSA showing increased p53-DNA binding activity in pre-miR-34a transfected cells for 48 and
74 h. C) Representative Annexin V-APC/PI staining showing absence of cell death after miR-34a
modulation.
4.5 Discussion
miR-34a is a well-known miRNA involved in various cellular functions that acts as
a tumor suppressor (Chang et al., 2007b; He et al., 2007a). In this study, we demonstrate
a novel function for miR-34a during neuronal differentiation, implicating miR-34a
expression on the appearance of postmitotic neurons by a mechanism that involves SIRT1
downregulation. This mechanism implies also increased p53-DNA binding activity, in
common with neural differentiation.
We have previously found that miR-34a expression was concomitant with the
appearance of postmitotic neurons and astrocytes during mouse NS cell differentiation
(Aranha et al., 2010). Here we show that miR-34a promotes the appearance of
postmitotic neurons. This is in agreement with previous observations that miR-34a can
suppress cell-cycle genes and induce a neural phenotype (Wei et al., 2008). miR-34a may
have a wide range of molecular targets during cell differentiation. Here, we confirmed
and extended other results implicating SIRT1 in the differentiation activity of miR-34a
(Tarantino et al., 2010a). In addition, miR-34a can regulate Notch-1 and Notch-2 protein
expression (Li et al., 2009a), and decrease CDK4 and CDK6 (Navarro et al., 2009), as
well as MEK1 (Ichimura et al., 2010), which regulate cell cycle progression and inhibit
cell proliferation, respectively, as a prerequisite for cells to enter differentiation.
Although constituting an intriguing topic, the role of SIRT1 during neuronal
differentiation is not entirely new. The results presented in this study are in accordance
with recent findings showing that SIRT1 expression is regulated during human embryonic
stem cell differentiation (Calvanese et al., 2010). Specifically, SIRT1 down-regulation
was shown to be necessary for reactivation of key developmental genes, such as the
neuroretinal morphogenesis effector delta-like protein 4 (DLL4), the T-box transcription
factor (TBX3), and paired box gene 6 (PAX6). Similarly, downregulation of SIRT1
during mouse NS cell differentiation might be necessary for the accomplishment of the
transition toward postmitotic neurons and completion of a correct neuronal differentiation
program. However, the observation that SIRT1 is expressed in β-III Tubulin+ cells at two

miR-34a regulates neural differentiation
107
days of differentiation indicates that SIRT1 is necessary in the initial stages of the
differentiation process. In agreement, SIRT1 has been shown to positively regulate
neuronal differentiation by inhibiting the transactivation of Hes1, a transcription factor
that negatively regulates cell differentiation (Hisahara et al., 2008). In addition, loss of
SIRT1 in embryonic stem cells was shown to delay the switch off of Oct4 and Nanog
expression, affecting the capacity to exit the pluripotent stem cell program (Ou et al.,
2010). This suggests that a tight regulation of SIRT1 expression might exist and that
timing and levels of expression may determine commitment toward a certain neural
phenotype.
Our results also demonstrate that SIRT1 expression positively regulates
astrogliogenesis. In fact, it has been previously shown that increased SIRT1 activity
causes differentiation of neural progenitor cells into astrocytes at the expense of neurons
(Prozorovski et al., 2008). SIRT1 represses the pro-neuronal basic helix-loop-helix
(bHLH) transcription factor Mash1 and influences cell-fate decision. By regulating
SIRT1 expression in the neuronal subpopulation, miR-34a may impair the developmental
program toward the astroglial lineage and favor neurogenesis. Nevertheless, we do not
exclude the possibility that miR-34a may also contribute to neuronal differentiation by
additional mechanisms. This indicates that SIRT1 might play a distinct role on
differentiation under specific conditions, according to substrate and co-factor availability,
as well as redox state of the cell.
miR-34a expression was found not to be involved in SIRT1 downregulation during
human embryonic stem cell differentiation (Calvanese et al., 2010). In fact, miR-34a
expression is not increased after induction of differentiation into embryonic bodies.
Differences associated with specific properties of each biological model may explain this
discrepancy. In addition, it also shows that different mechanisms might exist that
regulate SIRT1 expression levels.
We found that transient transfection of miR-34a into mouse NS cells had almost no
effect on the astrocytic subpopulation, which is in accordance with previous studies
showing that forced miR-34a expression did not affect human astrocytes (Li et al., 2009).
Curiously, a positive regulation of miR-34a on GFAP+ cells became notorious after
increased SIRT1 activity. Given that miR-34a negatively regulates SIRT1 expression and
SIRT1 induces the astroglial lineage, we would expect to have an opposite effect of miR-
34a in GFAP+ cells. This suggests that under certain conditions, miR-34a may have the

Chapter 4
108
capacity to influence commitment toward astrogliogenesis by a SIRT1-independent
mechanism.
miR-34a is part of the p53 regulatory network (He et al., 2007b). miR-34a
transcription is directly activated by p53, and in turn miR-34a regulates the expression of
some p53 target genes (Chang et al., 2007b; He et al., 2007a; Raver-Shapira et al., 2007).
Nevertheless, transfection with p53 siRNA did not affect miR-34a expression, suggesting
that induction of miR-34a during mouse NS cell differentiation is p53 independent (data
not shown). These results are in accordance with previous data showing a p53-
independent role for miR-34a during megakaryocytic differentiation of K562 cells
(Navarro et al., 2009). Instead, p53 seems to act downstream of miR-34a in this cellular
context. Our results suggest that miR-34a indirectly regulates p53, possible through a
SIRT1-dependent mechanism. Further, SIRT1 inhibits p53-mediated suppression of
Nanog expression, which is required to maintain cells in an undifferentiated status (Han
et al., 2008). Increased p53-DNA binding activity resulting from miR-34a
overexpression may mediate miR-34a induction of postmitotic neurons by controlling cell
cycle progression of neuronal precursors. Additional experiments are required to confirm
whether SIRT1 mediates miR-34a activation of p53. Nevertheless, the present study
provides a foundation for the role of miR-34a in differentiation of neural stem cells.
In conclusion, here we report the role of miR-34a in regulating neural
differentiation. In addition, we demonstrate that miR-34a-mediated silencing of SIRT1
may be necessary for correct establishment of specific differentiation programs during
neural stem cell differentiation.
Acknowledgments
We would like to acknowledge Inês Milagre and Dr. Elsa Rodrigues (iMed.UL,
Lisbon, Portugal) for help with EMSA. This work was supported by grant PTDC/BIA-
BCM/67922/2006 and PTDC/SAU-GMG/099162/2008 from Fundação para a Ciência e a
Tecnologia (FCT), Lisbon, Portugal. MM Aranha and DM Santos are recipients of PhD
fellowships from FCT, Portugal (SFRH/BD/28429/2006 and SFRH/BD/42008/2007,
respectively).

5 Concluding Remarks


Concluding Remarks
111
The studies presented in this thesis contribute to a better understanding of the
molecular mechanisms controlling neural stem (NS) cell differentiation. More
specifically, these studies confirm and extend our knowledge in considering apoptosis-
associated molecules as integral components of the differentiation process.
In brief, we demonstrated a role for p53 and caspase-3 in the differentiation process
of NS cells, and suggested that these molecules temporally modulate FOXO3A/Id1
signaling. In addition, we implicated apoptosis-associated microRNAs (miRNAs or
miRs), such as miR-16, let-7a and miR-34a, in neural differentiation of mouse, rat and
human cells, independently of their role in apoptosis. Finally, we further explored the
function of miR-34a in mediating the transition between neural progenitors and post-
mitotic neurons, and implicated SIRT1 and p53 activity in this regulatory network. The
data presented here provide new insights into the regulation of neural differentiation by
apoptosis-associated molecules, and represents a step forward in understanding the
molecular mechanisms of NS cell differentiation. In this last concluding chapter, we
integrate our findings, and address future perspectives arising from these studies.
The tumor suppressor p53 and caspase-3 have been implicated in the differentiation
process of subventricular zone (SVZ)- and striatum-derived NS cells, respectively
(Fernando et al., 2005; Gil-Perotin et al., 2006). To extend these studies, we investigated
whether p53 and caspase-3 molecules would also be implicated in the differentiation
process of NS cells, extracted from E14 mouse embryos. Our results indicated that NS
cell differentiation is associated with an increase in active caspase-3, as well as
phosphorylated p53 (Ser 15), concomitant with increased p53 DNA-binding activity. We
also performed functional studies and demonstrated that caspase-3 inhibition decreases
both neuronal and astroglial subpopulations, while p53 down-regulation affects only the
neuronal lineage. It would be interesting to address in the future, whether this strict
subpopulation effect is due to specific localized expression of p53 in neuronal precursors.
In addition, most of the relevant p53 transcriptional targets and caspase-3 substrates have
yet to be determined in a differentiation context. Experiments aimed at addressing these
questions should include high-throughput proteomics analysis and genome-wide
chromatin immunoprecipitation assays.
Id1 is a bHLH transcription factor that positively contributes to cell cycle
progression (Hamajima et al., 2010), and is responsible for maintaining the proliferative
state of cortical progenitors (Ross et al., 2003). In addition, it has been demonstrated that
non-phosphorylated FOXO3A transcriptionally represses Id1 expression (Birkenkamp et

Chapter 5
112
al., 2007). Given the fact that differentiation results in progressive loss of self-renewal
and unrestricted development potential, we dissected the Akt/p-FOXO3A/Id1 pathway
during NS cell differentiation, and confirmed the downregulation of this signaling
pathway. We next addressed whether the involvement of p53 and caspase-3 in NS cell
differentiation cross-talks with this pathway, thus influencing Id1 expression. We found
that in the absence of caspase-3 activity or after siRNA mediated-silencing of p53, the p-
FOXO3A/Id1 pathway is no longer down-regulated, suggesting that the role of p53 and
caspase-3 during NS cell differentiation relies on a cross-talk with proliferation pathways,
ultimately leading to down-regulation of molecules responsible for maintaining the
proliferative state of stem cells, such as Id1.
The involvement of p53 in the differentiation process may also provide important
information for future research in the field of induced pluripotent stem (iPS) cells. It was
recently shown that inhibition of p53 greatly increases the reprogramming efficiency
process (Hong et al., 2009). Nevertheless, if the inactivation of p53 is crucial for
successful reprogramming, this approach raises concerns about the genomic stability and
tumorigenecity of iPS cells, and may compromise the differentiation capacity of iPS cells
into specific lineages. In addition, efforts to block apoptosis via caspase inhibition for
therapeutic purposes may have much broader implications than initially thought,
especially for stem cells.
Recently, noncoding RNAs belonging to the miRNA family have emerged as a new
class of cell lineage determinants. As repressors, miRNAs may promote differentiation
by limiting the expression of genes that support the self-renewing progenitor state.
Hence, we were interested in determining whether miRNAs linked to apoptosis are
involved in the differentiation of NS cells. To address this issue, we performed
microarray miRNA profiling in NS cells at different times of differentiation to detect
miRNA expression. Our results revealed differential expression of several apoptosis-
associated miRNAs, including miR-16, let-7a and miR-34a, and this was also validated
by quantitative real time-PCR. Importantly, we found that expression levels of miR-16,
let-7a and miR-34a are also modulated in mouse ES cell, PC12 and NT2N differentiation,
implicating these miRNAs in mammalian neuronal development. It would be interesting
to determine the expression patterns of apoptosis-associated miRNAs specifically in
neuronal and glial lineages, using a system with β-III tubulin-GFP and GFAP-GFP
expressing cells, respectively. Dissecting the mechanisms associated with the expression

Concluding Remarks
113
of apoptosis-associated miRNAs is crucial for a better understanding of their role and
relevance in the differentiation process.
Contrary to transcription factors, miRNAs usually function as a fine-tuning
mechanism, and not as an on/off switch. This may be important for future applications
involving the modulation of apoptosis-associated pathways by miRNAs, because minor
changes in activity levels may ultimately dictate the cell choice between cell death and
differentiation. By fine-tuning protein expression, miRNA effects are not as dramatic,
which makes miRNAs potential ideal therapeutic targets. Further, the ability of a single
miRNA to control the expression of several mRNAs might be crucial, especially if
targeted mRNAs belong to a common signaling pathway. Finally, miRNA processing is
faster than protein translation, allowing miRNAs to rapidly affect gene expression (Tsang
et al., 2007). Modulation of miRNAs for therapeutic purpose is now under investigation
and represents a promising area for future clinical application.
It has been reported that miR-34a expression increases during retinoic acid-induced
differentiation of neuroblastoma cell lines (Welch et al., 2007). Based on the
identification of miR-34a as a p53 target (Bommer et al., 2007), we became interested in
confirming the role of miR-34a in cell differentiation. We found that miR-34a positively
regulates the appearance of post-mitotic neurons. Nevertheless, ectopic expression of
miR-34a in embryonic mouse central nervous system through in vivo electroporation, and
the use of miR-34a knockout mice would both add valuable information regarding the
role of miR-34a in neural differentiation. In addition, in situ hybridization during mouse
embryonic development would add evidence for the involvement of miR-34a during
neuronal development.
We have also addressed the question of whether p53 is responsible for miR-34a
induction during NS cell differentiation. We found that p53 down-regulation using
siRNA is not associated with decreased miR-34a expression, suggesting that miR-34a
acts in a p53-independent manner. Nevertheless it is tempting to speculate that the other
members of the p53 family, p63 and especially p73 can participate in the regulation of
miR-34a expression. For instance, p73 knockout mice display varying degrees of
abnormal neurological development (Yang et al., 2000), and p73 has been shown to
induce differentiation in neuroblastoma cells in vitro (De Laurenzi et al., 2000).
Modulation of p73 expression levels and the use of reporter plasmids carrying wild-type
and mutated miR-34a sequence for p73 putative binding should be explored in the future.

Chapter 5
114
SIRT1 is a validated miR-34a target, recently reported to be involved in miR-34a-
mediated ES cell differentiation (Tarantino et al., 2010b). Therefore, we became
interested in assessing whether SIRT1 is involved in miR-34a-mediated NS cell
differentiation. Our results showed that miR-34a modulation significantly affects SIRT1
expression. Nevertheless, modulation of SIRT1 only slightly influences the proportion of
post-mitotic neurons, indicating that other miR-34a targets may also contribute to this
phenotype. In the future, an attempt should be taken to further identify others miR-34a
targets. Differentiation is tightly linked with cell cycle exit, and the capacity of miR-34a
to modulate cell cycle regulators may suggest their intervention on the transition between
mitotic neuronal precursors to immature post-mitotic neurons.
SIRT1 is a NAD-dependent deacetylase, which has been shown to inhibit several
pro-apoptotic proteins, including p53. Not surprisingly, miR-34a-mediated
downregulation of SIRT1 resulted in increased p53 activity (Yamakuchi et al., 2008). To
investigate if a similar mechanism is involved in NS cell differentiation, we determined
p53-DNA binding activity after miR-34a overexpression. Our results revealed that p53-
DNA binding activity is increased after upregulation of miR-34a. Given the data
suggesting a positive regulation of differentiation by p53, miR-34a-mediated induction of
p53 might result in an amplification of the signal dictating cell differentiation. Thus, our
results suggest that miR-34a might be a suitable target for therapeutic intervention.
Surprisingly, our results showed that miR-34a positively regulates astrocytes.
However, this was only revealed when cells were pre-treated with the SIRT1-activator,
resveratrol. It is thought that miRNAs may act as a buffering system, modulating
molecules whenever the cell balance is lost. Therefore, simple inactivation of a miRNA
may not suffice to collapse the signaling networks in which miRNAs are involved.
Previous studies showed that for most miRNA genes of C. elegans, single-gene
knockouts did not result in detectable mutant phenotypes (Miska et al., 2007).
Alternatively, miRNA functions may be revealed under more sophisticated experimental
assays that take into account the cell complexity. It was recently suggested that the use of
sensitized genetic backgrounds provides an efficient approach for miRNA functional
analysis (Brenner et al., 2010). Therefore, especial particular environment might be
generated after treatment with resveratrol, which in turn may reveal other miR-34a
functions, such as the capacity to regulate the glial lineage. Nevertheless, additional
studies, possibly using genetic backgrounds with reduced processing and activity of all

Concluding Remarks
115
miRNAs or with reduced activity of a wide array of regulatory pathways would be
necessary to confirm the capacity of miR-34a to regulate astrogliogenesis.
Accumulating evidence demonstrating a role for apoptosis-associated molecules in
differentiation has advanced our understanding of the mechanisms regulating cell
differentiation. The challenge for the future will be to identify mechanisms and nuances
that ultimately guide this machinery to either cell death or differentiation, which may
reveal novel mechanisms regulating neurogenesis as well as new targets for therapeutic
intervention.


117
References
Abe-Dohmae, S., Harada, N., Yamada, K., and Tanaka, R. (1993). Bcl-2 gene is highly
expressed during neurogenesis in the central nervous system. Biochemical and
Biophysical Research Communications 191, 915-921.
Abranches, E., Silva, M., Pradier, L., Schulz, H., Hummel, O., Henrique, D., and
Bekman, E. (2009). Neural differentiation of embryonic stem cells in vitro: a road map to
neurogenesis in the embryo. PloS One 4, e6286.
Akao, Y., Nakagawa, Y., and Naoe, T. (2006). let-7 microRNA functions as a
potential growth suppressor in human colon cancer cells. Biological & Pharmaceutical
Bulletin 29, 903-906.
Aladjem, M.I., Spike, B.T., Rodewald, L.W., Hope, T.J., Klemm, M., Jaenisch, R., and
Wahl, G.M. (1998). ES cells do not activate p53-dependent stress responses and undergo
p53-independent apoptosis in response to DNA damage. Current Biology 8, 145-155.
Altman, J., and Das, G.D. (1965). Autoradiographic and histological evidence of
postnatal hippocampal neurogenesis in rats. The Journal of Comparative Neurology 124,
319-335.
Altuvia, Y., Landgraf, P., Lithwick, G., Elefant, N., Pfeffer, S., Aravin, A.,
Brownstein, M.J., Tuschl, T., and Margalit, H. (2005). Clustering and conservation
patterns of human microRNAs. Nucleic Acids Research 33, 2697-2706.
Alvarez-Buylla, A., and Garcia-Verdugo, J.M. (2002). Neurogenesis in adult
subventricular zone. Journal of Neuroscience 22, 629-634.
Alvarez-Buylla, A., Garcia-Verdugo, J.M., and Tramontin, A.D. (2001). A unified
hypothesis on the lineage of neural stem cells. Nature Review in Neuroscience 2, 287-
293.
Ambros, V. (2004). The functions of animal microRNAs. Nature 431, 350-355.

References
118
Anthony, T.E., and Heintz, N. (2008). Genetic lineage tracing defines distinct
neurogenic and gliogenic stages of ventral telencephalic radial glial development. Neural
Development 3, 30.
Arama, E., Agapite, J., and Steller, H. (2003). Caspase activity and a specific
cytochrome C are required for sperm differentiation in Drosophila. Developmental Cell 4,
687-697.
Aranha, M.M., Santos, D.M., Xavier, J.M., Low, W.C., Steer, C.J., Sola, S., and
Rodrigues, C.M. (2010). Apoptosis-associated microRNAs are modulated in mouse, rat
and human neural differentiation. BMC Genomics 11, 514.
Aranha, M.M., Sola, S., Low, W.C., Steer, C.J., and Rodrigues, C.M. (2009). Caspases
and p53 modulate FOXO3A/Id1 signaling during mouse neural stem cell differentiation.
Journal of Cellular Biochemistry 107, 748-758.
Armesilla-Diaz, A., Bragado, P., Del Valle, I., Cuevas, E., Lazaro, I., Martin, C.,
Cigudosa, J.C., and Silva, A. (2009). p53 regulates the self-renewal and differentiation of
neural precursors. Neuroscience 158, 1378-1389.
Armstrong, J.F., Kaufman, M.H., Harrison, D.J., and Clarke, A.R. (1995). High-
frequency developmental abnormalities in p53-deficient mice. Current Biology 5, 931-
936.
Bartel, D.P. (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell
116, 281-297.
Bartel, D.P. (2009). MicroRNAs: target recognition and regulatory functions. Cell 136,
215-233.
Baskerville, S., and Bartel, D.P. (2005). Microarray profiling of microRNAs reveals
frequent coexpression with neighboring miRNAs and host genes. RNA 11, 241-247.
Bernardi, P., and Azzone, G.F. (1981). Cytochrome c as an electron shuttle between
the outer and inner mitochondrial membranes. The Journal of Biological Chemistry 256,
7187-7192.
Bernardino, L., Agasse, F., Silva, B., Ferreira, R., Grade, S., and Malva, J.O. (2008).
Tumor necrosis factor-alpha modulates survival, proliferation, and neuronal
differentiation in neonatal subventricular zone cell cultures. Stem Cells 26, 2361-2371.

References
119
Bernstein, E., Kim, S.Y., Carmell, M.A., Murchison, E.P., Alcorn, H., Li, M.Z., Mills,
A.A., Elledge, S.J., Anderson, K.V., and Hannon, G.J. (2003). Dicer is essential for
mouse development. Nature Genetics 35, 215-217.
Birkenkamp, K.U., Essafi, A., van der Vos, K.E., da Costa, M., Hui, R.C., Holstege,
F., Koenderman, L., Lam, E.W., and Coffer, P.J. (2007). FOXO3a induces differentiation
of Bcr-Abl-transformed cells through transcriptional down-regulation of Id1. The Journal
of Biological Chemistry 282, 2211-2220.
Black, S., Kadyrov, M., Kaufmann, P., Ugele, B., Emans, N., and Huppertz, B. (2004).
Syncytial fusion of human trophoblast depends on caspase 8. Cell Death and
Differentiation 11, 90-98.
Blander, G., Bhimavarapu, A., Mammone, T., Maes, D., Elliston, K., Reich, C.,
Matsui, M.S., Guarente, L., and Loureiro, J.J. (2009). SIRT1 promotes differentiation of
normal human keratinocytes. The Journal of Investigative Dermatology 129, 41-49.
Blander, G., and Guarente, L. (2004). The Sir2 family of protein deacetylases. Annual
Review of Biochemistry 73, 417-435.
Bode, A.M., and Dong, Z. (2004). Post-translational modification of p53 in
tumorigenesis. Nature Review in Cancer 4, 793-805.
Bommer, G.T., Gerin, I., Feng, Y., Kaczorowski, A.J., Kuick, R., Love, R.E., Zhai, Y.,
Giordano, T.J., Qin, Z.S., Moore, B.B., et al. (2007). p53-mediated activation of
miRNA34 candidate tumor-suppressor genes. Current Biology 17, 1298-1307.
Bonauer, A., and Dimmeler, S. (2009). The microRNA-17-92 cluster: still a miRacle?
Cell Cycle 8, 3866-3873.
Brenner, J.L., Jasiewicz, K.L., Fahley, A.F., Kemp, B.J., and Abbott, A.L. (2010).
Loss of individual microRNAs causes mutant phenotypes in sensitized genetic
backgrounds in C. elegans. Current Biology 20, 1321-1325.
Brooks, C.L., and Gu, W. (2003). Ubiquitination, phosphorylation and acetylation: the
molecular basis for p53 regulation. Current Opinion in Cell Biology 15, 164-171.
Browes, C., Rowe, J., Brown, A., and Montano, X. (2001). Analysis of trk A and p53
association. FEBS Letters 497, 20-25.

References
120
Brunet, A., Bonni, A., Zigmond, M.J., Lin, M.Z., Juo, P., Hu, L.S., Anderson, M.J.,
Arden, K.C., Blenis, J., and Greenberg, M.E. (1999). Akt promotes cell survival by
phosphorylating and inhibiting a Forkhead transcription factor. Cell 96, 857-868.
Brunet, A., Sweeney, L.B., Sturgill, J.F., Chua, K.F., Greer, P.L., Lin, Y., Tran, H.,
Ross, S.E., Mostoslavsky, R., Cohen, H.Y., et al. (2004). Stress-dependent regulation of
FOXO transcription factors by the SIRT1 deacetylase. Science 303, 2011-2015.
Brynczka, C., Labhart, P., and Merrick, B.A. (2007). NGF-mediated transcriptional
targets of p53 in PC12 neuronal differentiation. BMC Genomics 8, 139.
Bunk, E., König, H.-G., Bernas, T., Engel, T., Henshall, D., Kirby, B., and Prehn, J.
(2010). BH3-only proteins BIM and PUMA in the regulation of survival and neuronal
differentiation of newly generated cells in the adult mouse hippocampus. Cell Death and
Disease 1, 1-9.
Cai, X., Hagedorn, C.H., and Cullen, B.R. (2004). Human microRNAs are processed
from capped, polyadenylated transcripts that can also function as mRNAs. RNA 10,
1957-1966.
Cain, K., Bratton, S.B., and Cohen, G.M. (2002). The Apaf-1 apoptosome: a large
caspase-activating complex. Biochimie 84, 203-214.
Calvanese, V., Lara, E., Suarez-Alvarez, B., Abu Dawud, R., Vazquez-Chantada, M.,
Martinez-Chantar, M.L., Embade, N., Lopez-Nieva, P., Horrillo, A., Hmadcha, A., et al.
(2010). Sirtuin 1 regulation of developmental genes during differentiation of stem cells.
Proceedings of the National Academy of Sciences of the United States of America 107,
13736-13741.
Carlile, G.W., Smith, D.H., and Wiedmann, M. (2004). Caspase-3 has a nonapoptotic
function in erythroid maturation. Blood 103, 4310-4316.
Cathelin, S., Rebe, C., Haddaoui, L., Simioni, N., Verdier, F., Fontenay, M., Launay,
S., Mayeux, P., and Solary, E. (2006). Identification of proteins cleaved downstream of
caspase activation in monocytes undergoing macrophage differentiation. The Journal of
Biological Chemistry 281, 17779-17788.
Caygill, E.E., and Johnston, L.A. (2008). Temporal regulation of metamorphic
processes in Drosophila by the let-7 and miR-125 heterochronic microRNAs. Current
Biology 18, 943-950.

References
121
Chang, J., Nicolas, E., Marks, D., Sander, C., Lerro, A., Buendia, M.A., Xu, C.,
Mason, W.S., Moloshok, T., Bort, R., et al. (2004). miR-122, a mammalian liver-specific
microRNA, is processed from hcr mRNA and may downregulate the high affinity
cationic amino acid transporter CAT-1. RNA Biology 1, 106-113.
Chang, M.Y., Sun, W., Ochiai, W., Nakashima, K., Kim, S.Y., Park, C.H., Kang, J.S.,
Shim, J.W., Jo, A.Y., Kang, C.S., et al. (2007a). Bcl-XL/Bax proteins direct the fate of
embryonic cortical precursor cells. Molecular and Cellular Biology 27, 4293-4305.
Chang, T.C., Wentzel, E.A., Kent, O.A., Ramachandran, K., Mullendore, M., Lee,
K.H., Feldmann, G., Yamakuchi, M., Ferlito, M., Lowenstein, C.J., et al. (2007b).
Transactivation of miR-34a by p53 broadly influences gene expression and promotes
apoptosis. Molecular Cell 26, 745-752.
Chao, C., Saito, S., Anderson, C.W., Appella, E., and Xu, Y. (2000). Phosphorylation
of murine p53 at ser-18 regulates the p53 responses to DNA damage. Proceedings of the
National Academy of Sciences of the United States of America 97, 11936-11941.
Chasis, J.A., and Schrier, S.L. (1989). Membrane deformability and the capacity for
shape change in the erythrocyte. Blood 74, 2562-2568.
Chen, R.W., Bemis, L.T., Amato, C.M., Myint, H., Tran, H., Birks, D.K., Eckhardt,
S.G., and Robinson, W.A. (2008). Truncation in CCND1 mRNA alters miR-16-1
regulation in mantle cell lymphoma. Blood 112, 822-829.
Chen, S.E., Gerken, E., Zhang, Y., Zhan, M., Mohan, R.K., Li, A.S., Reid, M.B., and
Li, Y.P. (2005). Role of TNF-α signaling in regeneration of cardiotoxin-injured muscle.
American Journal of Physiology 289, 1179-1187.
Cheung, W.M., Fu, W.Y., Hui, W.S., and Ip, N.Y. (1999). Production of human CNS
neurons from embryonal carcinoma cells using a cell aggregation Methods in
BioTechniques 26, 946-948, 950-942, 954.
Chipuk, J.E., and Green, D.R. (2005). Do inducers of apoptosis trigger caspase-
independent cell death? Nature Reviews 6, 268-275.
Choi, B.H., and Lapham, L.W. (1978). Radial glia in the human fetal cerebrum: a
combined Golgi, immunofluorescent and electron microscopic study. Brain Research 148,
295-311.

References
122
Chojnacki, A., and Weiss, S. (2008). Production of neurons, astrocytes and
oligodendrocytes from mammalian CNS stem cells. Nature Protocols 3, 935-940.
Chomarat, P., Dantin, C., Bennett, L., Banchereau, J., and Palucka, A.K. (2003). TNF
skews monocyte differentiation from macrophages to dendritic cells. Journal of
Immunology 171, 2262-2269.
Cimmino, A., Calin, G.A., Fabbri, M., Iorio, M.V., Ferracin, M., Shimizu, M., Wojcik,
S.E., Aqeilan, R.I., Zupo, S., Dono, M., et al. (2005). miR-15 and miR-16 induce
apoptosis by targeting BCL2. Proceedings of the National Academy of Sciences of the
United States of America 102, 13944-13949.
Conti, L., and Cattaneo, E. (2010). Neural stem cell systems: physiological players or
in vitro entities? Nature Review in Neuroscience 11, 176-187.
Corney, D.C., Flesken-Nikitin, A., Godwin, A.K., Wang, W., and Nikitin, A.Y.
(2007). MicroRNA-34b and MicroRNA-34c are targets of p53 and cooperate in control of
cell proliferation and adhesion-independent growth. Cancer Research 67, 8433-8438.
Corsini, N.S., Sancho-Martinez, I., Laudenklos, S., Glagow, D., Kumar, S., Letellier,
E., Koch, P., Teodorczyk, M., Kleber, S., Klussmann, S., et al. (2009). The death receptor
CD95 activates adult neural stem cells for working memory formation and brain repair.
Cell Stem Cell 5, 178-190.
Curtis, M.A., Penney, E.B., Pearson, A.G., van Roon-Mom, W.M., Butterworth, N.J.,
Dragunow, M., Connor, B., and Faull, R.L. (2003). Increased cell proliferation and
neurogenesis in the adult human Huntington's disease brain. Proceedings of the National
Academy of Sciences of the United States of America 100, 9023-9027.
De Botton, S., Sabri, S., Daugas, E., Zermati, Y., Guidotti, J.E., Hermine, O.,
Kroemer, G., Vainchenker, W., and Debili, N. (2002). Platelet formation is the
consequence of caspase activation within megakaryocytes. Blood 100, 1310-1317.
De Laurenzi, V., Raschella, G., Barcaroli, D., Annicchiarico-Petruzzelli, M., Ranalli,
M., Catani, M.V., Tanno, B., Costanzo, A., Levrero, M., and Melino, G. (2000).
Induction of neuronal differentiation by p73 in a neuroblastoma cell line. The Journal of
Biological Chemistry 275, 15226-15231.
Debatin, K.M., and Krammer, P.H. (2004). Death receptors in chemotherapy and
cancer. Oncogene 23, 2950-2966.

References
123
Denecker, G., Hoste, E., Gilbert, B., Hochepied, T., Ovaere, P., Lippens, S., Van den
Broecke, C., Van Damme, P., D'Herde, K., Hachem, J.P., et al. (2007). Caspase-14
protects against epidermal UVB photodamage and water loss. Nature Cell Biology 9,
666-674.
Di Giovanni, S., Faden, A.I., Yakovlev, A., Duke-Cohan, J.S., Finn, T., Thouin, M.,
Knoblach, S., De Biase, A., Bregman, B.S., and Hoffman, E.P. (2005). Neuronal
plasticity after spinal cord injury: identification of a gene cluster driving neurite
outgrowth. FASEB Journal 19, 153-154.
Di Giovanni, S., Knights, C.D., Rao, M., Yakovlev, A., Beers, J., Catania, J.,
Avantaggiati, M.L., and Faden, A.I. (2006). The tumor suppressor protein p53 is required
for neurite outgrowth and axon regeneration. The EMBO Journal 25, 4084-4096.
Dix, M.M., Simon, G.M., and Cravatt, B.F. (2008). Global mapping of the topography
and magnitude of proteolytic events in apoptosis. Cell 134, 679-691.
Doe, C.Q. (2008). Neural stem cells: balancing self-renewal with differentiation.
Development 135, 1575-1587.
Doetsch, F. (2003). The glial identity of neural stem cells. Nature Neuroscience 6,
1127-1134.
Doetsch, F., and Alvarez-Buylla, A. (1996). Network of tangential pathways for
neuronal migration in adult mammalian brain. Proceedings of the National Academy of
Sciences of the United States of America 93, 14895-14900.
Doetsch, F., Caille, I., Lim, D.A., Garcia-Verdugo, J.M., and Alvarez-Buylla, A.
(1999). Subventricular zone astrocytes are neural stem cells in the adult mammalian
brain. Cell 97, 703-716.
Doetsch, F., and Scharff, C. (2001). Challenges for brain repair: insights from adult
neurogenesis in birds and mammals. Brain, Behavior and Evolution 58, 306-322.
Donehower, L.A., Harvey, M., Slagle, B.L., McArthur, M.J., Montgomery, C.A., Jr.,
Butel, J.S., and Bradley, A. (1992). Mice deficient for p53 are developmentally normal
but susceptible to spontaneous tumours. Nature 356, 215-221.
Du, T., and Zamore, P.D. (2005). microPrimer: the biogenesis and function of
microRNA. Development 132, 4645-4652.

References
124
Ellis, R.E., Yuan, J.Y., and Horvitz, H.R. (1991). Mechanisms and functions of cell
death. Annual Review of Cell Biology 7, 663-698.
Eom, D.S., Choi, W.S., and Oh, Y.J. (2004). Bcl-2 enhances neurite extension via
activation of c-Jun N-terminal kinase. Biochemical and Biophysical Research
Communications 314, 377-381.
Esdar, C., Milasta, S., Maelicke, A., and Herget, T. (2001). Differentiation-associated
apoptosis of neural stem cells is effected by Bcl-2 overexpression: impact on cell lineage
determination. European Journal of Cell Biology 80, 539-553.
Feng, L., Hatten, M.E., and Heintz, N. (1994). Brain lipid-binding protein (BLBP): a
novel signaling system in the developing mammalian CNS. Neuron 12, 895-908.
Fernando, P., Brunette, S., and Megeney, L.A. (2005). Neural stem cell differentiation
is dependent upon endogenous caspase 3 activity. FASEB Journal 19, 1671-1673.
Fernando, P., Kelly, J.F., Balazsi, K., Slack, R.S., and Megeney, L.A. (2002). Caspase
3 activity is required for skeletal muscle differentiation. Proceedings of the National
Academy of Sciences of the United States of America 99, 11025-11030.
Fernando, P., and Megeney, L.A. (2007). Is caspase-dependent apoptosis only cell
differentiation taken to the extreme? FASEB Journal 21, 8-17.
Finkel, T., Deng, C.X., and Mostoslavsky, R. (2009). Recent progress in the biology
and physiology of sirtuins. Nature 460, 587-591.
Fontana, L., Pelosi, E., Greco, P., Racanicchi, S., Testa, U., Liuzzi, F., Croce, C.M.,
Brunetti, E., Grignani, F., and Peschle, C. (2007). MicroRNAs 17-5p-20a-106a control
monocytopoiesis through AML1 targeting and M-CSF receptor upregulation. Nature Cell
Biology 9, 775-787.
Friedman, L.M., Dror, A.A., Mor, E., Tenne, T., Toren, G., Satoh, T., Biesemeier,
D.J., Shomron, N., Fekete, D.M., Hornstein, E., et al. (2009). MicroRNAs are essential
for development and function of inner ear hair cells in vertebrates. Proceedings of the
National Academy of Sciences of the United States of America 106, 7915-7920.
Fujita, J., Crane, A.M., Souza, M.K., Dejosez, M., Kyba, M., Flavell, R.A., Thomson,
J.A., and Zwaka, T.P. (2008). Caspase activity mediates the differentiation of embryonic
stem cells. Cell Stem Cell 2, 595-601.

References
125
Fukuda, S., Kato, F., Tozuka, Y., Yamaguchi, M., Miyamoto, Y., and Hisatsune, T.
(2003). Two distinct subpopulations of nestin-positive cells in adult mouse dentate gyrus.
Journal of Neuroscience 23, 9357-9366.
Fulco, M., Schiltz, R.L., Iezzi, S., King, M.T., Zhao, P., Kashiwaya, Y., Hoffman, E.,
Veech, R.L., and Sartorelli, V. (2003). Sir2 regulates skeletal muscle differentiation as a
potential sensor of the redox state. Molecular Cell 12, 51-62.
Gage, F.H. (2000). Mammalian neural stem cells. Science 287, 1433-1438.
Gangaraju, V.K., and Lin, H. (2009). MicroRNAs: key regulators of stem cells. Nature
Reviews 10, 116-125.
Garcia, A.D., Doan, N.B., Imura, T., Bush, T.G., and Sofroniew, M.V. (2004). GFAP-
expressing progenitors are the principal source of constitutive neurogenesis in adult
mouse forebrain. Nature Neuroscience 7, 1233-1241.
Garofalo, M., Condorelli, G.L., Croce, C.M., and Condorelli, G. (2010). MicroRNAs
as regulators of death receptors signaling. Cell Death and Differentiation 17, 200-208.
Garrido, C., and Kroemer, G. (2004). Life's smile, death's grin: vital functions of
apoptosis-executing proteins. Current Opinion in Cell Biology 16, 639-646.
Garzon, R., Pichiorri, F., Palumbo, T., Iuliano, R., Cimmino, A., Aqeilan, R., Volinia,
S., Bhatt, D., Alder, H., Marcucci, G., et al. (2006). MicroRNA fingerprints during
human megakaryocytopoiesis. Proceedings of the National Academy of Sciences of the
United States of America 103, 5078-5083.
Garzon, R., Volinia, S., Liu, C.G., Fernandez-Cymering, C., Palumbo, T., Pichiorri, F.,
Fabbri, M., Coombes, K., Alder, H., Nakamura, T., et al. (2008). MicroRNA signatures
associated with cytogenetics and prognosis in acute myeloid leukemia. Blood 111, 3183-
3189.
Gil-Perotin, S., Marin-Husstege, M., Li, J., Soriano-Navarro, M., Zindy, F., Roussel,
M.F., Garcia-Verdugo, J.M., and Casaccia-Bonnefil, P. (2006). Loss of p53 induces
changes in the behavior of subventricular zone cells: implication for the genesis of glial
tumors. Journal of Neuroscience 26, 1107-1116.
Gonzalez-Garcia, M., Garcia, I., Ding, L., O'Shea, S., Boise, L.H., Thompson, C.B.,
and Nunez, G. (1995). bcl-x is expressed in embryonic and postnatal neural tissues and

References
126
functions to prevent neuronal cell death. Proceedings of the National Academy of
Sciences of the United States of America 92, 4304-4308.
Gottlieb, E., Haffner, R., King, A., Asher, G., Gruss, P., Lonai, P., and Oren, M.
(1997). Transgenic mouse model for studying the transcriptional activity of the p53
protein: age- and tissue-dependent changes in radiation-induced activation during
embryogenesis. The EMBO journal 16, 1381-1390.
Gotz, M., and Huttner, W.B. (2005). The cell biology of neurogenesis. Nature Reviews
6, 777-788.
Green, D.R., and Evan, G.I. (2002). A matter of life and death. Cancer Cell 1, 19-30.
Green, D.R., and Kroemer, G. (2004). The pathophysiology of mitochondrial cell
death. Science 305, 626-629.
Greene, L.A., and Tischler, A.S. (1976). Establishment of a noradrenergic clonal line
of rat adrenal pheochromocytoma cells which respond to nerve growth factor.
Proceedings of the National Academy of Sciences of the United States of America 73,
2424-2428.
Gregoli, P.A., and Bondurant, M.C. (1997). The roles of Bcl-X(L) and apopain in the
control of erythropoiesis by erythropoietin. Blood 90, 630-640.
Gu, L., Zhu, N., Findley, H.W., Woods, W.G., and Zhou, M. (2004). Identification and
characterization of the IKKalpha promoter: positive and negative regulation by ETS-1
and p53, respectively. The Journal of Biological Chemistry 279, 52141-52149.
Guarente, L., and Picard, F. (2005). Calorie restriction--the SIR2 connection. Cell 120,
473-482.
Guessous, F., Zhang, Y., Kofman, A., Catania, A., Li, Y., Schiff, D., Purow, B., and
Abounader, R. (2010). microRNA-34a is tumor suppressive in brain tumors and glioma
stem cells. Cell Cycle.
Guillemain, I., Gaboyard, S., Fontes, G., Saunier, M., Privat, A., and Patey, G. (2000).
Differential expression of Bcl-2-related proteins in differentiating NT2 cells. Neuroreport
11, 1421-1425.
Haigis, M.C., and Guarente, L.P. (2006). Mammalian sirtuins--emerging roles in
physiology, aging, and calorie restriction. Genes & Development 20, 2913-2921.

References
127
Hamajima, Y., Komori, M., Preciado, D.A., Choo, D.I., Moribe, K., Murakami, S.,
Ondrey, F.G., and Lin, J. (2010). The role of inhibitor of DNA-binding (Id1) in
hyperproliferation of keratinocytes: the pathological basis for middle ear cholesteatoma
from chronic otitis media. Cell Proliferation 43, 457-463.
Han, M.K., Song, E.K., Guo, Y., Ou, X., Mantel, C., and Broxmeyer, H.E. (2008).
SIRT1 regulates apoptosis and Nanog expression in mouse embryonic stem cells by
controlling p53 subcellular localization. Cell Stem Cell 2, 241-251.
Hanna, J., Saha, K., Pando, B., van Zon, J., Lengner, C.J., Creyghton, M.P., van
Oudenaarden, A., and Jaenisch, R. (2009). Direct cell reprogramming is a stochastic
process amenable to acceleration. Nature 462, 595-601.
Hara, K., Yasuhara, T., Maki, M., Matsukawa, N., Masuda, T., Yu, S.J., Ali, M., Yu,
G., Xu, L., Kim, S.U., et al. (2008). Neural progenitor NT2N cell lines from
teratocarcinoma for transplantation therapy in stroke. Progress in Neurobiology 85, 318-
334.
Harris, S.L., and Levine, A.J. (2005). The p53 pathway: positive and negative
feedback loops. Oncogene 24, 2899-2908.
Hashimi, S.T., Fulcher, J.A., Chang, M.H., Gov, L., Wang, S., and Lee, B. (2009).
MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and
WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 114, 404-414.
Haubensak, W., Attardo, A., Denk, W., and Huttner, W.B. (2004). Neurons arise in the
basal neuroepithelium of the early mammalian telencephalon: a major site of
neurogenesis. Proceedings of the National Academy of Sciences of the United States of
America 101, 3196-3201.
He, L., He, X., Lim, L.P., de Stanchina, E., Xuan, Z., Liang, Y., Xue, W., Zender, L.,
Magnus, J., Ridzon, D., et al. (2007a). A microRNA component of the p53 tumour
suppressor network. Nature 447, 1130-1134.
He, X., He, L., and Hannon, G.J. (2007b). The guardian's little helper: microRNAs in
the p53 tumor suppressor network. Cancer Research 67, 11099-11101.
Hermeking, H. (2010). The miR-34 family in cancer and apoptosis. Cell Death and
Differentiation 17, 193-199.

References
128
Hisahara, S., Chiba, S., Matsumoto, H., Tanno, M., Yagi, H., Shimohama, S., Sato,
M., and Horio, Y. (2008). Histone deacetylase SIRT1 modulates neuronal differentiation
by its nuclear translocation. Proceedings of the National Academy of Sciences of the
United States of America 105, 15599-15604.
Hong, H., Takahashi, K., Ichisaka, T., Aoi, T., Kanagawa, O., Nakagawa, M., Okita,
K., and Yamanaka, S. (2009). Suppression of induced pluripotent stem cell generation by
the p53-p21 pathway. Nature 460, 1132-1135.
Hsu, Y.C., Lee, D.C., and Chiu, I.M. (2007). Neural stem cells, neural progenitors, and
neurotrophic factors. Cell Transplantation 16, 133-150.
Hu, M.C., Lee, D.F., Xia, W., Golfman, L.S., Ou-Yang, F., Yang, J.Y., Zou, Y., Bao,
S., Hanada, N., Saso, H., et al. (2004). IkappaB kinase promotes tumorigenesis through
inhibition of forkhead FOXO3a. Cell 117, 225-237.
Hudson, C.D., Morris, P.J., Latchman, D.S., and Budhram-Mahadeo, V.S. (2005).
Brn-3a transcription factor blocks p53-mediated activation of proapoptotic target genes
Noxa and Bax in vitro and in vivo to determine cell fate. The Journal of Biological
Chemistry 280, 11851-11858.
Huesmann, G.R., and Clayton, D.F. (2006). Dynamic role of postsynaptic caspase-3
and BIRC4 in zebra finch song-response habituation. Neuron 52, 1061-1072.
Hughes, A.L., Gollapudi, L., Sladek, T.L., and Neet, K.E. (2000). Mediation of nerve
growth factor-driven cell cycle arrest in PC12 cells by p53. Simultaneous differentiation
and proliferation subsequent to p53 functional inactivation. The Journal of Biological
Chemistry 275, 37829-37837.
Ichimura, A., Ruike, Y., Terasawa, K., Shimizu, K., and Tsujimoto, G. (2010).
MicroRNA-34a inhibits cell proliferation by repressing mitogen-activated protein kinase
kinase 1 during megakaryocytic differentiation of K562 cells. Molecular Pharmacology
77, 1016-1024.
Imura, T., Kornblum, H.I., and Sofroniew, M.V. (2003). The predominant neural stem
cell isolated from postnatal and adult forebrain but not early embryonic forebrain
expresses GFAP. Journal of Neuroscience 23, 2824-2832.
Ishizaki, Y., Jacobson, M.D., and Raff, M.C. (1998). A role for caspases in lens fiber
differentiation. The Journal of Cell Biology 140, 153-158.

References
129
Ivey, K.N., and Srivastava, D. (2010). MicroRNAs as regulators of differentiation and
cell fate decisions. Cell Stem Cell 7, 36-41.
Janzen, V., Fleming, H.E., Riedt, T., Karlsson, G., Riese, M.J., Lo Celso, C.,
Reynolds, G., Milne, C.D., Paige, C.J., Karlsson, S., et al. (2008). Hematopoietic stem
cell responsiveness to exogenous signals is limited by caspase-3. Cell Stem Cell 2, 584-
594.
Jessberger, S., and Kempermann, G. (2003). Adult-born hippocampal neurons mature
into activity-dependent responsiveness. The European Journal of Neuroscience 18, 2707-
2712.
Jin, K., Galvan, V., Xie, L., Mao, X.O., Gorostiza, O.F., Bredesen, D.E., and
Greenberg, D.A. (2004). Enhanced neurogenesis in Alzheimer's disease transgenic
(PDGF-APPSw,Ind) mice. Proceedings of the National Academy of Sciences of the
United States of America 101, 13363-13367.
Jin, Y.H., Kim, Y.J., Kim, D.W., Baek, K.H., Kang, B.Y., Yeo, C.Y., and Lee, K.Y.
(2008). Sirt2 interacts with 14-3-3 beta/gamma and down-regulates the activity of p53.
Biochemical and Biophysical Research Communication 368, 690-695.
Johnson, S.M., Grosshans, H., Shingara, J., Byrom, M., Jarvis, R., Cheng, A.,
Labourier, E., Reinert, K.L., Brown, D., and Slack, F.J. (2005). RAS is regulated by the
let-7 microRNA family. Cell 120, 635-647.
Kalscheuer, S., Zhang, X., Zeng, Y., and Upadhyaya, P. (2008). Differential
expression of microRNAs in early-stage neoplastic transformation in the lungs of F344
rats chronically treated with the tobacco carcinogen 4-(methylnitrosamino)-1-(3-pyridyl)-
1-butanone. Carcinogenesis 29, 2394-2399.
Kamada, S., Shimono, A., Shinto, Y., Tsujimura, T., Takahashi, T., Noda, T.,
Kitamura, Y., Kondoh, H., and Tsujimoto, Y. (1995). bcl-2 deficiency in mice leads to
pleiotropic abnormalities: accelerated lymphoid cell death in thymus and spleen,
polycystic kidney, hair hypopigmentation, and distorted small intestine. Cancer Research
55, 354-359.
Kang, T.B., Ben-Moshe, T., Varfolomeev, E.E., Pewzner-Jung, Y., Yogev, N.,
Jurewicz, A., Waisman, A., Brenner, O., Haffner, R., Gustafsson, E., et al. (2004).

References
130
Caspase-8 serves both apoptotic and nonapoptotic roles. Journal of Immunology 173,
2976-2984.
Kapinas, K., Kessler, C.B., and Delany, A.M. (2009). miR-29 suppression of
osteonectin in osteoblasts: regulation during differentiation and by canonical Wnt
signaling. Journal of Cellular Biochemistry 108, 216-224.
Kawamura, T., Suzuki, J., Wang, Y.V., Menendez, S., Morera, L.B., Raya, A., Wahl,
G.M., and Belmonte, J.C. (2009). Linking the p53 tumour suppressor pathway to somatic
cell reprogramming. Nature 460, 1140-1144.
Kempermann, G., and Gage, F.H. (2000). Neurogenesis in the adult hippocampus.
Novartis Foundation symposium 231, 220-235; discussion 235-241, 302-226.
Kempermann, G., Gast, D., Kronenberg, G., Yamaguchi, M., and Gage, F.H. (2003).
Early determination and long-term persistence of adult-generated new neurons in the
hippocampus of mice. Development 130, 391-399.
Kokovay, E., Shen, Q., and Temple, S. (2008). The incredible elastic brain: how neural
stem cells expand our minds. Neuron 60, 420-429.
Komarova, E.A., Chernov, M.V., Franks, R., Wang, K., Armin, G., Zelnick, C.R.,
Chin, D.M., Bacus, S.S., Stark, G.R., and Gudkov, A.V. (1997). Transgenic mice with
p53-responsive lacZ: p53 activity varies dramatically during normal development and
determines radiation and drug sensitivity in vivo. The EMBO Journal 16, 1391-1400.
Korsmeyer, S.J., Wei, M.C., Saito, M., Weiler, S., Oh, K.J., and Schlesinger, P.H.
(2000). Pro-apoptotic cascade activates BID, which oligomerizes BAK or BAX into pores
that result in the release of cytochrome c. Cell Death and Differentiation 7, 1166-1173.
Krajewska, M., Mai, J.K., Zapata, J.M., Ashwell, K.W., Schendel, S.L., Reed, J.C.,
and Krajewski, S. (2002). Dynamics of expression of apoptosis-regulatory proteins Bid,
Bcl-2, Bcl-X, Bax and Bak during development of murine nervous system. Cell Death
and Differentiation 9, 145-157.
Kroemer, G., Galluzzi, L., and Brenner, C. (2007). Mitochondrial membrane
permeabilization in cell death. Physiological Reviews 87, 99-163.
Kroemer, G., and Martin, S.J. (2005). Caspase-independent cell death. Nature
Medicine 11, 725-730.

References
131
Krol, J., Loedige, I., and Filipowicz, W. (2010). The widespread regulation of
microRNA biogenesis, function and decay. Nature Review in Genetics 11, 597-610.
Kuhn, H.G., Dickinson-Anson, H., and Gage, F.H. (1996). Neurogenesis in the dentate
gyrus of the adult rat: age-related decrease of neuronal progenitor proliferation. Journal of
Neuroscience 16, 2027-2033.
Kuo, C.T., Zhu, S., Younger, S., Jan, L.Y., and Jan, Y.N. (2006). Identification of
E2/E3 ubiquitinating enzymes and caspase activity regulating Drosophila sensory neuron
dendrite pruning. Neuron 51, 283-290.
Kyoung Kim, H., Kyoung Kim, Y., Song, I.H., Baek, S.H., Lee, S.R., Hye Kim, J., and
Kim, J.R. (2005). Down-regulation of a forkhead transcription factor, FOXO3a,
accelerates cellular senescence in human dermal fibroblasts. The Journal of Gerontology.
Series A, Biological Sciences and Medical Sciences 60, 4-9.
Lagos-Quintana, M., Rauhut, R., Lendeckel, W., and Tuschl, T. (2001). Identification
of novel genes coding for small expressed RNAs. Science 294, 853-858.
Lamkanfi, M., Festjens, N., Declercq, W., Vanden Berghe, T., and Vandenabeele, P.
(2007). Caspases in cell survival, proliferation and differentiation. Cell Death and
Differerentiation 14, 44-55.
Lancman, J.J., Caruccio, N.C., Harfe, B.D., Pasquinelli, A.E., Schageman, J.J.,
Pertsemlidis, A., and Fallon, J.F. (2005). Analysis of the regulation of lin-41 during chick
and mouse limb development. Developmental Dynamics 234, 948-960.
Langley, E., Pearson, M., Faretta, M., Bauer, U.M., Frye, R.A., Minucci, S., Pelicci,
P.G., and Kouzarides, T. (2002). Human SIR2 deacetylates p53 and antagonizes
PML/p53-induced cellular senescence. The EMBO Journal 21, 2383-2396.
Lavin, M.F., and Gueven, N. (2006). The complexity of p53 stabilization and
activation. Cell Death and Differentiation 13, 941-950.
LeBrun, D.P., Warnke, R.A., and Cleary, M.L. (1993). Expression of bcl-2 in fetal
tissues suggests a role in morphogenesis. The American Journal of Pathology 142, 743-
753.
Lee, Y.S., and Dutta, A. (2007). The tumor suppressor microRNA let-7 represses the
HMGA2 oncogene. Genes & Development 21, 1025-1030.

References
132
Lee, Y.S., Kim, H.K., Chung, S., Kim, K.S., and Dutta, A. (2005). Depletion of human
micro-RNA miR-125b reveals that it is critical for the proliferation of differentiated cells
but not for the down-regulation of putative targets during differentiation. The Journal of
Biological Chemistry 280, 16635-16641.
Lengner, C.J., Steinman, H.A., Gagnon, J., Smith, T.W., Henderson, J.E., Kream,
B.E., Stein, G.S., Lian, J.B., and Jones, S.N. (2006). Osteoblast differentiation and
skeletal development are regulated by Mdm2-p53 signaling. The Journal of Cell Biology
172, 909-921.
Levitt, P., and Rakic, P. (1980). Immunoperoxidase localization of glial fibrillary
acidic protein in radial glial cells and astrocytes of the developing rhesus monkey brain.
The Journal of Comparative Neurology 193, 815-840.
Li, F., He, Z., Shen, J., Huang, Q., Li, W., Liu, X., He, Y., Wolf, F., and Li, C.Y.
(2010a). Apoptotic Caspases Regulate Induction of iPSCs from Human Fibroblasts. Cell
Stem Cell 7, 508-520.
Li, H., Zhu, H., Xu, C.J., and Yuan, J. (1998). Cleavage of BID by caspase 8 mediates
the mitochondrial damage in the Fas pathway of apoptosis. Cell 94, 491-501.
Li, J., and Yuan, J. (2008). Caspases in apoptosis and beyond. Oncogene 27, 6194-
6206.
Li, L.Y., Luo, X., and Wang, X. (2001). Endonuclease G is an apoptotic DNase when
released from mitochondria. Nature 412, 95-99.
Li, T., Morgan, M.J., Choksi, S., Zhang, Y., Kim, Y.S., and Liu, Z.G. (2010b).
MicroRNAs modulate the noncanonical transcription factor NF-kappaB pathway by
regulating expression of the kinase IKKalpha during macrophage differentiation. Nature
Immunology 11, 799-805.
Li, X., and Jin, P. Roles of small regulatory RNAs in determining neuronal identity.
Nature Reviews 11, 329-338.
Li, Y., Guessous, F., Zhang, Y., Dipierro, C., Kefas, B., Johnson, E., Marcinkiewicz,
L., Jiang, J., Yang, Y., Schmittgen, T.D., et al. (2009a). MicroRNA-34a inhibits
glioblastoma growth by targeting multiple oncogenes. Cancer Research 69, 7569-7576.
Li, Z., Hassan, M.Q., Jafferji, M., Aqeilan, R.I., Garzon, R., Croce, C.M., van Wijnen,
A.J., Stein, J.L., Stein, G.S., and Lian, J.B. (2009b). Biological functions of miR-29b

References
133
contribute to positive regulation of osteoblast differentiation. The Journal of Biological
Chemistry 284, 15676-15684.
Lin, C.J., Gong, H.Y., Tseng, H.C., Wang, W.L., and Wu, J.L. (2008). miR-122 targets
an anti-apoptotic gene, Bcl-w, in human hepatocellular carcinoma cell lines. Biochemical
and Biophysical Research Communications 375, 315-320.
Lin, T., Chao, C., Saito, S., Mazur, S.J., Murphy, M.E., Appella, E., and Xu, Y.
(2005). p53 induces differentiation of mouse embryonic stem cells by suppressing Nanog
expression. Nature Cell Biology 7, 165-171.
Linsley, P.S., Schelter, J., Burchard, J., Kibukawa, M., Martin, M.M., Bartz, S.R.,
Johnson, J.M., Cummins, J.M., Raymond, C.K., Dai, H., et al. (2007). Transcripts
targeted by the microRNA-16 family cooperatively regulate cell cycle progression.
Molecular and Cellular Biology 27, 2240-2252.
Lippens, S., Kockx, M., Knaapen, M., Mortier, L., Polakowska, R., Verheyen, A.,
Garmyn, M., Zwijsen, A., Formstecher, P., Huylebroeck, D., et al. (2000). Epidermal
differentiation does not involve the pro-apoptotic executioner caspases, but is associated
with caspase-14 induction and processing. Cell Death and Differentiation 7, 1218-1224.
Liste, I., Garcia-Garcia, E., Bueno, C., and Martinez-Serrano, A. (2007). Bcl-XL
modulates the differentiation of immortalized human neural stem cells. Cell Death and
Differentiation 14, 1880-1892.
Liste, I., Garcia-Garcia, E., and Martinez-Serrano, A. (2004). The generation of
dopaminergic neurons by human neural stem cells is enhanced by Bcl-XL, both in vitro
and in vivo. Journal of Neuroscience 24, 10786-10795.
Liu, D.X., and Greene, L.A. (2001). Neuronal apoptosis at the G1/S cell cycle
checkpoint. Cell Tissue Research 305, 217-228.
Liu, Q., Fu, H., Sun, F., Zhang, H., Tie, Y., Zhu, J., Xing, R., Sun, Z., and Zheng, X.
(2008). miR-16 family induces cell cycle arrest by regulating multiple cell cycle genes.
Nucleic Acids Research 36, 5391-5404.
Liu, S.P., Fu, R.H., Yu, H.H., Li, K.W., Tsai, C.H., Shyu, W.C., and Lin, S.Z. (2009).
MicroRNAs regulation modulated self-renewal and lineage differentiation of stem cells.
Cell Transplantation 18, 1039-1045.

References
134
Lledo, P.M., Alonso, M., and Grubb, M.S. (2006). Adult neurogenesis and functional
plasticity in neuronal circuits. Nature Review in Neuroscience 7, 179-193.
Lodygin, D., Tarasov, V., Epanchintsev, A., Berking, C., Knyazeva, T., Korner, H.,
Knyazev, P., Diebold, J., and Hermeking, H. (2008). Inactivation of miR-34a by aberrant
CpG methylation in multiple types of cancer. Cell Cycle 7, 2591-2600.
Lomonosova, E., and Chinnadurai, G. (2008). BH3-only proteins in apoptosis and
beyond: an overview. Oncogene 27 Suppl 1, S2-19.
Louis, J.M., McFarland, V.W., May, P., and Mora, P.T. (1988). The phosphoprotein
p53 is down-regulated post-transcriptionally during embryogenesis in vertebrates.
Biochimica et Biophysica Acta 950, 395-402.
Lu, Q.L., Poulsom, R., Wong, L., and Hanby, A.M. (1993). Bcl-2 expression in adult
and embryonic non-haematopoietic tissues. The Journal of Pathology 169, 431-437.
Magavi, S.S., Leavitt, B.R., and Macklis, J.D. (2000). Induction of neurogenesis in the
neocortex of adult mice. Nature 405, 951-955.
Mahrus, S., Trinidad, J.C., Barkan, D.T., Sali, A., Burlingame, A.L., and Wells, J.A.
(2008). Global sequencing of proteolytic cleavage sites in apoptosis by specific labeling
of protein N termini. Cell 134, 866-876.
Maimets, T., Neganova, I., Armstrong, L., and Lako, M. (2008). Activation of p53 by
nutlin leads to rapid differentiation of human embryonic stem cells. Oncogene 27, 5277-
5287.
Mandinova, A., Lefort, K., Tommasi di Vignano, A., Stonely, W., Ostano, P.,
Chiorino, G., Iwaki, H., Nakanishi, J., and Dotto, G.P. (2008). The FoxO3a gene is a key
negative target of canonical Notch signalling in the keratinocyte UVB response. The
EMBO Journal 27, 1243-1254.
Mantel, C., and Broxmeyer, H.E. (2008). Sirtuin 1, stem cells, aging, and stem cell
aging. Current Opinion in Hematology 15, 326-331.
Matas, D., Milyavsky, M., Shats, I., Nissim, L., Goldfinger, N., and Rotter, V. (2004).
p53 is a regulator of macrophage differentiation. Cell Death and Differentiation 11, 458-
467.

References
135
Megiorni, F., Mora, B., Indovina, P., and Mazzilli, M.C. (2005). Expression of
neuronal markers during NTera2/cloneD1 differentiation by cell aggregation method.
Neuroscience Letters 373, 105-109.
Meletis, K., Wirta, V., Hede, S.M., Nister, M., Lundeberg, J., and Frisen, J. (2006).
p53 suppresses the self-renewal of adult neural stem cells. Development 133, 363-369.
Melton, C., Judson, R.L., and Blelloch, R. (2010). Opposing microRNA families
regulate self-renewal in mouse embryonic stem cells. Nature 463, 621-626.
Merkle, F.T., and Alvarez-Buylla, A. (2006). Neural stem cells in mammalian
development. Current Opinion in Cell Biology 18, 704-709.
Merkle, F.T., Mirzadeh, Z., and Alvarez-Buylla, A. (2007). Mosaic organization of
neural stem cells in the adult brain. Science 317, 381-384.
Merry, D.E., Veis, D.J., Hickey, W.F., and Korsmeyer, S.J. (1994). bcl-2 protein
expression is widespread in the developing nervous system and retained in the adult PNS.
Development 120, 301-311.
Middleton, G., Pinon, L.G., Wyatt, S., and Davies, A.M. (1998). Bcl-2 accelerates the
maturation of early sensory neurons. Journal of Neuroscience 18, 3344-3350.
Mielke, K., and Herdegen, T. (2002). Fatal shift of signal transduction is an integral
part of neuronal differentiation: JNKs realize TNFalpha-mediated apoptosis in
neuronlike, but not naive, PC12 cells. Molecular and Cellular Neuroscience 20, 211-224.
Milagre, I., Nunes, M.J., Gama, M.J., Silva, R.F., Pascussi, J.M., Lechner, M.C., and
Rodrigues, E. (2008). Transcriptional regulation of the human CYP46A1 brain-specific
expression by Sp transcription factors. Journal of Neurochemistry 106, 835-849.
Miska, E.A., Alvarez-Saavedra, E., Abbott, A.L., Lau, N.C., Hellman, A.B.,
McGonagle, S.M., Bartel, D.P., Ambros, V.R., and Horvitz, H.R. (2007). Most
Caenorhabditis elegans microRNAs are individually not essential for development or
viability. PLoS Genetics 3, e215.
Misra, R.S., Russell, J.Q., Koenig, A., Hinshaw-Makepeace, J.A., Wen, R., Wang, D.,
Huo, H., Littman, D.R., Ferch, U., Ruland, J., et al. (2007). Caspase-8 and c-FLIPL
associate in lipid rafts with NF-kappaB adaptors during T cell activation. The Journal
Biological Chemistry 282, 19365-19374.

References
136
Miura, M., Chen, X.D., Allen, M.R., Bi, Y., Gronthos, S., Seo, B.M., Lakhani, S.,
Flavell, R.A., Feng, X.H., Robey, P.G., et al. (2004). A crucial role of caspase-3 in
osteogenic differentiation of bone marrow stromal stem cells. The Journal of Clinical
Investigation 114, 1704-1713.
Miyamoto, K., Araki, K.Y., Naka, K., Arai, F., Takubo, K., Yamazaki, S., Matsuoka,
S., Miyamoto, T., Ito, K., Ohmura, M., et al. (2007). Foxo3a is essential for maintenance
of the hematopoietic stem cell pool. Cell Stem Cell 1, 101-112.
Mogi, M., and Togari, A. (2003). Activation of caspases is required for osteoblastic
differentiation. The Journal of Biological Chemistry 278, 47477-47482.
Mogi, M., Walsh, K., Iwai, M., and Horiuchi, M. (2008). Akt-FOXO3a signaling
affects human endothelial progenitor cell differentiation. Hypertension Research 31, 153-
159.
Montano, X. (1997). P53 associates with trk tyrosine kinase. Oncogene 15, 245-256.
Mori, S., Matsuyama, K., Mori, F., and Nakajima, K. (2001). Supraspinal sites that
induce locomotion in the vertebrate central nervous system. Advances in Neurology 87,
25-40.
Motoyama, N., Wang, F., Roth, K.A., Sawa, H., Nakayama, K., Nakayama, K.,
Negishi, I., Senju, S., Zhang, Q., Fujii, S., et al. (1995). Massive cell death of immature
hematopoietic cells and neurons in Bcl-x-deficient mice. Science 267, 1506-1510.
Mott, J.L., Kobayashi, S., Bronk, S.F., and Gores, G.J. (2007). mir-29 regulates Mcl-1
protein expression and apoptosis. Oncogene 26, 6133-6140.
Mu, Y., Lee, S.W., and Gage, F.H. (2010). Signaling in adult neurogenesis. Current
Opinion in Neurobiology 20, 416-423.
Murchison, E.P., Partridge, J.F., Tam, O.H., Cheloufi, S., and Hannon, G.J. (2005).
Characterization of Dicer-deficient murine embryonic stem cells. Proceedings of the
National Academy of Sciences of the United States of America 102, 12135-12140.
Murray-Zmijewski, F., Slee, E.A., and Lu, X. (2008). A complex barcode underlies the
heterogeneous response of p53 to stress. Nature Reviews 9, 702-712.

References
137
Navarro, F., Gutman, D., Meire, E., Caceres, M., Rigoutsos, I., Bentwich, Z., and
Lieberman, J. (2009). miR-34a contributes to megakaryocytic differentiation of K562
cells independently of p53. Blood 114, 2181-2192.
Netea, M.G., Lewis, E.C., Azam, T., Joosten, L.A., Jaekal, J., Bae, S.Y., Dinarello,
C.A., and Kim, S.H. (2008). Interleukin-32 induces the differentiation of monocytes into
macrophage-like cells. Proceedings of the National Academy of Sciences of the United
States of America 105, 3515-3520.
Nicholson, D.W. (1999). Caspase structure, proteolytic substrates, and function during
apoptotic cell death. Cell Death and Differentiation 6, 1028-1042.
Niizuma, H., Nakamura, Y., Ozaki, T., Nakanishi, H., Ohira, M., Isogai, E.,
Kageyama, H., Imaizumi, M., and Nakagawara, A. (2006). Bcl-2 is a key regulator for the
retinoic acid-induced apoptotic cell death in neuroblastoma. Oncogene 25, 5046-5055.
Nishikawa, S., Jakt, L.M., and Era, T. (2007). Embryonic stem-cell culture as a tool
for developmental cell biology. Nature Reviews 8, 502-507.
Norton, J.D. (2000). ID helix-loop-helix proteins in cell growth, differentiation and
tumorigenesis. Journal of Cell Science 113, 3897-3905.
Ogawa, Y., Takebayashi, H., Takahashi, M., Osumi, N., Iwasaki, Y., and Ikenaka, K.
(2005). Gliogenic radial glial cells show heterogeneity in the developing mouse spinal
cord. Developmental Neuroscience 27, 364-377.
Oomman, S., Strahlendorf, H., Dertien, J., and Strahlendorf, J. (2006). Bergmann glia
utilize active caspase-3 for differentiation. Brain Research 1078, 19-34.
Ou, X., Chae, H.D., Wang, R.H., Shelley, W.C., Cooper, S., Taylor, T., Kim, Y.J.,
Deng, C.X., Yoder, M.C., and Broxmeyer, H.E. (2010). SIRT1 deficiency compromises
mouse embryonic stem cell hematopoietic differentiation, and embryonic and adult
hematopoiesis in the mouse. Blood.
Pankratz, M.T., Li, X.J., Lavaute, T.M., Lyons, E.A., Chen, X., and Zhang, S.C.
(2007). Directed neural differentiation of human embryonic stem cells via an obligated
primitive anterior stage. Stem Cells 25, 1511-1520.
Park, S.M., Shell, S., Radjabi, A.R., Schickel, R., Feig, C., Boyerinas, B., Dinulescu,
D.M., Lengyel, E., and Peter, M.E. (2007). Let-7 prevents early cancer progression by
suppressing expression of the embryonic gene HMGA2. Cell Cycle 6, 2585-2590.

References
138
Parsadanian, A.S., Cheng, Y., Keller-Peck, C.R., Holtzman, D.M., and Snider, W.D.
(1998). Bcl-xL is an antiapoptotic regulator for postnatal CNS neurons. The Journal of
Neuroscience 18, 1009-1019.
Pasquinelli, A.E., Reinhart, B.J., Slack, F., Martindale, M.Q., Kuroda, M.I., Maller, B.,
Hayward, D.C., Ball, E.E., Degnan, B., Muller, P., et al. (2000). Conservation of the
sequence and temporal expression of let-7 heterochronic regulatory RNA. Nature 408,
86-89.
Pichiorri, F., Suh, S.S., Ladetto, M., Kuehl, M., Palumbo, T., Drandi, D., Taccioli, C.,
Zanesi, N., Alder, H., Hagan, J.P., et al. (2008). MicroRNAs regulate critical genes
associated with multiple myeloma pathogenesis. Proceedings of the National Academy of
Sciences of the United States of America 105, 12885-12890.
Plenchette, S., Cathelin, S., Rebe, C., Launay, S., Ladoire, S., Sordet, O., Ponnelle, T.,
Debili, N., Phan, T.H., Padua, R.A., et al. (2004). Translocation of the inhibitor of
apoptosis protein c-IAP1 from the nucleus to the Golgi in hematopoietic cells undergoing
differentiation: a nuclear export signal-mediated event. Blood 104, 2035-2043.
Politis, P.K., Thomaidou, D., and Matsas, R. (2008). Coordination of cell cycle exit
and differentiation of neuronal progenitors. Cell Cycle 7, 691-697.
Poluha, W., Schonhoff, C.M., Harrington, K.S., Lachyankar, M.B., Crosbie, N.E.,
Bulseco, D.A., and Ross, A.H. (1997). A novel, nerve growth factor-activated pathway
involving nitric oxide, p53, and p21WAF1 regulates neuronal differentiation of PC12
cells. The Journal of Biological Chemistry 272, 24002-24007.
Porrello, A., Cerone, M.A., Coen, S., Gurtner, A., Fontemaggi, G., Cimino, L.,
Piaggio, G., Sacchi, A., and Soddu, S. (2000). p53 regulates myogenesis by triggering the
differentiation activity of pRb. The Journal of Cellular Biology 151, 1295-1304.
Prozorovski, T., Schulze-Topphoff, U., Glumm, R., Baumgart, J., Schroter, F.,
Ninnemann, O., Siegert, E., Bendix, I., Brustle, O., Nitsch, R., et al. (2008). Sirt1
contributes critically to the redox-dependent fate of neural progenitors. Nature Cell
Biology 10, 385-394.
Qian, X., Shen, Q., Goderie, S.K., He, W., Capela, A., Davis, A.A., and Temple, S.
(2000). Timing of CNS cell generation: a programmed sequence of neuron and glial cell
production from isolated murine cortical stem cells. Neuron 28, 69-80.

References
139
Qian, Y., and Chen, X. (2008). ID1, Inhibitor of Differentiation/DNA Binding, Is an
Effector of the p53-dependent DNA Damage Response Pathway. The Journal of
Biological Chemistry 283, 22410-22416.
Qin, H., Yu, T., Qing, T., Liu, Y., Zhao, Y., Cai, J., Li, J., Song, Z., Qu, X., Zhou, P.,
et al. (2007). Regulation of apoptosis and differentiation by p53 in human embryonic
stem cells. The Journal of Biological Chemistry 282, 5842-5852.
Radley, J.M., and Scurfield, G. (1980). The mechanism of platelet release. Blood 56,
996-999.
Ramon y Cajal, S. (1913). Degeneration and regeneration of the nervous system
(London, Oxford University Press).
Raver-Shapira, N., Marciano, E., Meiri, E., Spector, Y., Rosenfeld, N., Moskovits, N.,
Bentwich, Z., and Oren, M. (2007). Transcriptional activation of miR-34a contributes to
p53-mediated apoptosis. Molecular Cell 26, 731-743.
Reichel, M.B., Ali, R.R., D'Esposito, F., Clarke, A.R., Luthert, P.J., Bhattacharya,
S.S., and Hunt, D.M. (1998). High frequency of persistent hyperplastic primary vitreous
and cataracts in p53-deficient mice. Cell Death and Differentiation 5, 156-162.
Reinhart, B.J., Slack, F.J., Basson, M., Pasquinelli, A.E., Bettinger, J.C., Rougvie,
A.E., Horvitz, H.R., and Ruvkun, G. (2000). The 21-nucleotide let-7 RNA regulates
developmental timing in Caenorhabditis elegans. Nature 403, 901-906.
Reynolds, B.A., Tetzlaff, W., and Weiss, S. (1992). A multipotent EGF-responsive
striatal embryonic progenitor cell produces neurons and astrocytes. The Journal of
Neuroscience 12, 4565-4574.
Reynolds, B.A., and Weiss, S. (1992). Generation of neurons and astrocytes from
isolated cells of the adult mammalian central nervous system. Science 255, 1707-1710.
Reynolds, B.A., and Weiss, S. (1996). Clonal and population analyses demonstrate
that an EGF-responsive mammalian embryonic CNS precursor is a stem cell.
Developmental Biology 175, 1-13.
Ribeil, J.A., Zermati, Y., Vandekerckhove, J., Cathelin, S., Kersual, J., Dussiot, M.,
Coulon, S., Moura, I.C., Zeuner, A., Kirkegaard-Sorensen, T., et al. (2007). Hsp70
regulates erythropoiesis by preventing caspase-3-mediated cleavage of GATA-1. Nature
445, 102-105.

References
140
Ribeil, J.A., Zermati, Y., Vandekerckhove, J., Dussiot, M., Kersual, J., and Hermine,
O. (2005). Erythropoiesis: a paradigm for the role of caspases in cell death and
differentiation. The Journal Social Biology 199, 219-231.
Rietze, R., and Reynolds, B. (2006). Neural stem cell isolation and characterization.
Methods in Enzymology 419, 3-23.
Rimondi, E., Secchiero, P., Quaroni, A., Zerbinati, C., Capitani, S., and Zauli, G.
(2006). Involvement of TRAIL/TRAIL-receptors in human intestinal cell differentiation.
Journal of Cellular Physiology 206, 647-654.
Rodriguez, A., Griffiths-Jones, S., Ashurst, J.L., and Bradley, A. (2004). Identification
of mammalian microRNA host genes and transcription units. Genome Research 14, 1902-
1910.
Rogel, A., Popliker, M., Webb, C.G., and Oren, M. (1985). p53 cellular tumor antigen:
analysis of mRNA levels in normal adult tissues, embryos, and tumors. Molecular and
Cellular Biology 5, 2851-2855.
Rohn, T.T., Cusack, S.M., Kessinger, S.R., and Oxford, J.T. (2004). Caspase
activation independent of cell death is required for proper cell dispersal and correct
morphology in PC12 cells. Experimental Cell Research 295, 215-225.
Romero-Calvo, I., Ocon, B., Martinez-Moya, P., Suarez, M.D., Zarzuelo, A.,
Martinez-Augustin, O., and de Medina, F.S. Reversible Ponceau staining as a loading
control alternative to actin in Western blots. Analytical Biochemistry 401, 318-320.
Romero-Calvo, I., Ocon, B., Martinez-Moya, P., Suarez, M.D., Zarzuelo, A.,
Martinez-Augustin, O., and de Medina, F.S. (2010). Reversible Ponceau staining as a
loading control alternative to actin in Western blots. Analytical Biochemistry 401, 318-
320.
Ross, S.E., Greenberg, M.E., and Stiles, C.D. (2003). Basic helix-loop-helix factors in
cortical development. Neuron 39, 13-25.
Rotter, V., Schwartz, D., Almon, E., Goldfinger, N., Kapon, A., Meshorer, A.,
Donehower, L.A., and Levine, A.J. (1993). Mice with reduced levels of p53 protein
exhibit the testicular giant-cell degenerative syndrome. Proceedings of the National
Academy of Sciences of the United States of America 90, 9075-9079.

References
141
Roush, S., and Slack, F.J. (2008). The let-7 family of microRNAs. Trends in Cell
Biology 18, 505-516.
Rybak, A., Fuchs, H., Smirnova, L., Brandt, C., Pohl, E.E., Nitsch, R., and Wulczyn,
F.G. (2008). A feedback loop comprising lin-28 and let-7 controls pre-let-7 maturation
during neural stem-cell commitment. Nature Cell Biology 10, 987-993.
Sah, V.P., Attardi, L.D., Mulligan, G.J., Williams, B.O., Bronson, R.T., and Jacks, T.
(1995). A subset of p53-deficient embryos exhibit exencephaly. Nature Genetics 10, 175-
180.
Saifudeen, Z., Dipp, S., and El-Dahr, S.S. (2002). A role for p53 in terminal epithelial
cell differentiation. The Journal of Clinical Investigation 109, 1021-1030.
Saifudeen, Z., Dipp, S., Stefkova, J., Yao, X., Lookabaugh, S., and El-Dahr, S.S.
(2009). p53 regulates metanephric development. The Journal of American Society in
Nephrology 20, 2328-2337.
Sampson, V.B., Rong, N.H., Han, J., Yang, Q., Aris, V., Soteropoulos, P., Petrelli,
N.J., Dunn, S.P., and Krueger, L.J. (2007). MicroRNA let-7a down-regulates MYC and
reverts MYC-induced growth in Burkitt lymphoma cells. Cancer Research 67, 9762-
9770.
Sanz, C., Benito, A., Silva, M., Albella, B., Richard, C., Segovia, J.C., Insunza, A.,
Bueren, J.A., and Fernandez-Luna, J.L. (1997). The expression of Bcl-x is downregulated
during differentiation of human hematopoietic progenitor cells along the granulocyte but
not the monocyte/macrophage lineage. Blood 89, 3199-3204.
Sato, N., Hotta, K., Waguri, S., Nitatori, T., Tohyama, K., Tsujimoto, Y., and
Uchiyama, Y. (1994). Neuronal differentiation of PC12 cells as a result of prevention of
cell death by bcl-2. Journal of Neurobiology 25, 1227-1234.
Sauvageot, C.M., and Stiles, C.D. (2002). Molecular mechanisms controlling cortical
gliogenesis. Current Opinion in Neurobiology 12, 244-249.
Savitt, J.M., Jang, S.S., Mu, W., Dawson, V.L., and Dawson, T.M. (2005). Bcl-x is
required for proper development of the mouse substantia nigra. The Journal of
Neuroscience 25, 6721-6728.
Sawamoto, K., Wichterle, H., Gonzalez-Perez, O., Cholfin, J.A., Yamada, M.,
Spassky, N., Murcia, N.S., Garcia-Verdugo, J.M., Marin, O., Rubenstein, J.L., et al.

References
142
(2006). New neurons follow the flow of cerebrospinal fluid in the adult brain. Science
311, 629-632.
Scaffidi, C., Fulda, S., Srinivasan, A., Friesen, C., Li, F., Tomaselli, K.J., Debatin,
K.M., Krammer, P.H., and Peter, M.E. (1998). Two CD95 (APO-1/Fas) signaling
pathways. The EMBO Journal 17, 1675-1687.
Schimmelpfeng, J., Weibezahn, K.F., and Dertinger, H. (2004). Quantification of
NGF-dependent neuronal differentiation of PC-12 cells by means of neurofilament-L
mRNA expression and neuronal outgrowth. Journal of Neuroscience Methods 139, 299-
306.
Schmid, P., Lorenz, A., Hameister, H., and Montenarh, M. (1991). Expression of p53
during mouse embryogenesis. Development 113, 857-865.
Schreiber, E., Matthias, P., Muller, M.M., and Schaffner, W. (1989). Rapid detection
of octamer binding proteins with 'mini-extracts', prepared from a small number of cells.
Nucleic Acids Research 17, 6419.
Schulman, B.R., Esquela-Kerscher, A., and Slack, F.J. (2005). Reciprocal expression
of lin-41 and the microRNAs let-7 and mir-125 during mouse embryogenesis.
Developmental Dynamics 234, 1046-1054.
Schutte, B., Nuydens, R., Geerts, H., and Ramaekers, F. (1998). Annexin V binding
assay as a tool to measure apoptosis in differentiated neuronal cells. Journal of
Neuroscience Methods 86, 63-69.
Schwarz, J., Schwarz, S.C., and Storch, A. (2006). Developmental perspectives on
human midbrain-derived neural stem cells. Neurodegenerative Diseases 3, 45-49.
Schwerk, C., and Schulze-Osthoff, K. (2003). Non-apoptotic functions of caspases in
cellular proliferation and differentiation. Biochemical Pharmacology 66, 1453-1458.
Sempere, L.F., Dubrovsky, E.B., Dubrovskaya, V.A., Berger, E.M., and Ambros, V.
(2002). The expression of the let-7 small regulatory RNA is controlled by ecdysone
during metamorphosis in Drosophila melanogaster. Developmental Biology 244, 170-
179.
Sempere, L.F., Freemantle, S., Pitha-Rowe, I., Moss, E., Dmitrovsky, E., and Ambros,
V. (2004). Expression profiling of mammalian microRNAs uncovers a subset of brain-

References
143
expressed microRNAs with possible roles in murine and human neuronal differentiation.
Genome Biology 5, R13.
Shi, Y. (2004). Caspase activation, inhibition, and reactivation: a mechanistic view.
Protein Science 13, 1979-1987.
Shibata, T., Yamada, K., Watanabe, M., Ikenaka, K., Wada, K., Tanaka, K., and Inoue,
Y. (1997). Glutamate transporter GLAST is expressed in the radial glia-astrocyte lineage
of developing mouse spinal cord. The Journal of Neuroscience 17, 9212-9219.
Shim, J.W., Koh, H.C., Chang, M.Y., Roh, E., Choi, C.Y., Oh, Y.J., Son, H., Lee,
Y.S., Studer, L., and Lee, S.H. (2004). Enhanced in vitro midbrain dopamine neuron
differentiation, dopaminergic function, neurite outgrowth, and 1-methyl-4-
phenylpyridium resistance in mouse embryonic stem cells overexpressing Bcl-XL. The
Journal of Neuroscience 24, 843-852.
Shivdasani, R.A. (2006). MicroRNAs: regulators of gene expression and cell
differentiation. Blood 108, 3646-3653.
Sims, R.J., 3rd, and Reinberg, D. (2008). Is there a code embedded in proteins that is
based on post-translational modifications? Nature Reviews 9, 815-820.
Smirnova, L., Grafe, A., Seiler, A., Schumacher, S., Nitsch, R., and Wulczyn, F.G.
(2005). Regulation of miRNA expression during neural cell specification. The European
Journal of Neuroscience 21, 1469-1477.
Sokol, N.S., Xu, P., Jan, Y.N., and Ambros, V. (2008). Drosophila let-7 microRNA is
required for remodeling of the neuromusculature during metamorphosis. Genes &
Development 22, 1591-1596.
Sordet, O., Rebe, C., Plenchette, S., Zermati, Y., Hermine, O., Vainchenker, W.,
Garrido, C., Solary, E., and Dubrez-Daloz, L. (2002). Specific involvement of caspases in
the differentiation of monocytes into macrophages. Blood 100, 4446-4453.
Stambolic, V., MacPherson, D., Sas, D., Lin, Y., Snow, B., Jang, Y., Benchimol, S.,
and Mak, T.W. (2001). Regulation of PTEN transcription by p53. Molecular Cell 8, 317-
325.
Stanfield, B.B., and Trice, J.E. (1988). Evidence that granule cells generated in the
dentate gyrus of adult rats extend axonal projections. Experimental Brain Research 72,
399-406.

References
144
Stiewe, T. (2007). The p53 family in differentiation and tumorigenesis. Nature Review
in Cancer 7, 165-168.
Su, H., Yang, J.R., Xu, T., Huang, J., Xu, L., Yuan, Y., and Zhuang, S.M. (2009).
MicroRNA-101, down-regulated in hepatocellular carcinoma, promotes apoptosis and
suppresses tumorigenicity. Cancer Research 69, 1135-1142.
Suh, H., Consiglio, A., Ray, J., Sawai, T., D'Amour, K.A., and Gage, F.H. (2007). In
vivo fate analysis reveals the multipotent and self-renewal capacities of Sox2+ neural
stem cells in the adult hippocampus. Cell Stem Cell 1, 515-528.
Sun, F., Fu, H., Liu, Q., Tie, Y., Zhu, J., Xing, R., Sun, Z., and Zheng, X. (2008).
Downregulation of CCND1 and CDK6 by miR-34a induces cell cycle arrest. FEBS
Letters 582, 1564-1568.
Sun, Y., Nadal-Vicens, M., Misono, S., Lin, M.Z., Zubiaga, A., Hua, X., Fan, G., and
Greenberg, M.E. (2001). Neurogenin promotes neurogenesis and inhibits glial
differentiation by independent mechanisms. Cell 104, 365-376.
Susin, S.A., Lorenzo, H.K., Zamzami, N., Marzo, I., Snow, B.E., Brothers, G.M.,
Mangion, J., Jacotot, E., Costantini, P., Loeffler, M., et al. (1999). Molecular
characterization of mitochondrial apoptosis-inducing factor. Nature 397, 441-446.
Suzuki, A., and Tsutomi, Y. (1998). Bcl-2 accelerates the neuronal differentiation: new
evidence approaching to the biofunction of bcl-2 in the neuronal system. Brain Research
801, 59-66.
Takahashi, K., and Yamanaka, S. (2006). Induction of pluripotent stem cells from
mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676.
Tarantino, C., Paolella, G., Cozzuto, L., Minopoli, G., Pastore, L., Parisi, S., and
Russo, T. (2010). miRNA 34a, 100, and 137 modulate differentiation of mouse
embryonic stem cells. FASEB Journal 24, 3255-3263.
Tarasov, V., Jung, P., Verdoodt, B., Lodygin, D., Epanchintsev, A., Menssen, A.,
Meister, G., and Hermeking, H. (2007). Differential regulation of microRNAs by p53
revealed by massively parallel sequencing: miR-34a is a p53 target that induces apoptosis
and G1-arrest. Cell Cycle 6, 1586-1593.

References
145
Tebar, M., Boex, J.J., and Ten Have-Opbroek, A.A. (2001). Functional overexpression
of wild-type p53 correlates with alveolar cell differentiation in the developing human
lung. The Anatomical Record 263, 25-34.
Tedeschi, A., and Di Giovanni, S. (2009). The non-apoptotic role of p53 in neuronal
biology: enlightening the dark side of the moon. EMBO Reports 10, 576-583.
Tedeschi, A., Nguyen, T., Puttagunta, R., Gaub, P., and Di Giovanni, S. (2009). A
p53-CBP/p300 transcription module is required for GAP-43 expression, axon outgrowth,
and regeneration. Cell Death and Differentiation 16, 543-554.
Temple, S. (2001a). The development of neural stem cells. Nature 414, 112-117.
Temple, S. (2001b). Stem cell plasticity--building the brain of our dreams. Nature
Reviews 2, 513-520.
Terui, Y., Furukawa, Y., Kikuchi, J., Iwase, S., Hatake, K., and Miura, Y. (1998). Bcl-
x is a regulatory factor of apoptosis and differentiation in megakaryocytic lineage cells.
Experimental Hematology 26, 236-244.
Thomson, J.M., Parker, J., Perou, C.M., and Hammond, S.M. (2004). A custom
microarray platform for analysis of microRNA gene expression. Nature Methods 1, 47-
53.
Torrente, Y., El Fahime, E., Caron, N.J., Del Bo, R., Belicchi, M., Pisati, F., Tremblay,
J.P., and Bresolin, N. (2003). Tumor necrosis factor-alpha (TNF-alpha) stimulates
chemotactic response in mouse myogenic cells. Cell Transplantation 12, 91-100.
Tracey, D., Klareskog, L., Sasso, E.H., Salfeld, J.G., and Tak, P.P. (2008). Tumor
necrosis factor antagonist mechanisms of action: a comprehensive review. Pharmacology
& Therapeutics 117, 244-279.
Tramontin, A.D., Garcia-Verdugo, J.M., Lim, D.A., and Alvarez-Buylla, A. (2003).
Postnatal development of radial glia and the ventricular zone (VZ): a continuum of the
neural stem cell compartment. Cerebral Cortex 13, 580-587.
Tropepe, V., Sibilia, M., Ciruna, B.G., Rossant, J., Wagner, E.F., and van der Kooy, D.
(1999). Distinct neural stem cells proliferate in response to EGF and FGF in the
developing mouse telencephalon. Developmental Biology 208, 166-188.

References
146
Trouillas, M., Saucourt, C., Duval, D., Gauthereau, X., Thibault, C., Dembele, D.,
Feraud, O., Menager, J., Rallu, M., Pradier, L., et al. (2008). Bcl2, a transcriptional target
of p38alpha, is critical for neuronal commitment of mouse embryonic stem cells. Cell
Death and Differentiation 15, 1450-1459.
Tsang, J., Zhu, J., and van Oudenaarden, A. (2007). MicroRNA-mediated feedback
and feedforward loops are recurrent network motifs in mammals. Molecular Cell 26, 753-
767.
Tsang, W.P., and Kwok, T.T. (2008). Let-7a microRNA suppresses therapeutics-
induced cancer cell death by targeting caspase-3. Apoptosis 13, 1215-1222.
Van de Craen, M., Van Loo, G., Pype, S., Van Criekinge, W., Van den brande, I.,
Molemans, F., Fiers, W., Declercq, W., and Vandenabeele, P. (1998). Identification of a
new caspase homologue: caspase-14. Cell Death and Differentiation 5, 838-846.
Veis, D.J., Sorenson, C.M., Shutter, J.R., and Korsmeyer, S.J. (1993). Bcl-2-deficient
mice demonstrate fulminant lymphoid apoptosis, polycystic kidneys, and hypopigmented
hair. Cell 75, 229-240.
Vekrellis, K., McCarthy, M.J., Watson, A., Whitfield, J., Rubin, L.L., and Ham, J.
(1997). Bax promotes neuronal cell death and is downregulated during the development
of the nervous system. Development 124, 1239-1249.
Vousden, K.H., and Prives, C. (2009). Blinded by the Light: The Growing Complexity
of p53. Cell 137, 413-431.
Wang, H., Garzon, R., Sun, H., Ladner, K.J., Singh, R., Dahlman, J., Cheng, A., Hall,
B.M., Qualman, S.J., Chandler, D.S., et al. (2008a). NF-kappaB-YY1-miR-29 regulatory
circuitry in skeletal myogenesis and rhabdomyosarcoma. Cancer Cell 14, 369-381.
Wang, P., Qiu, W., Dudgeon, C., Liu, H., Huang, C., Zambetti, G.P., Yu, J., and
Zhang, L. (2009). PUMA is directly activated by NF-kappaB and contributes to TNF-
alpha-induced apoptosis. Cell Death and Differentiation 16, 1192-1202.
Wang, R.H., Sengupta, K., Li, C., Kim, H.S., Cao, L., Xiao, C., Kim, S., Xu, X.,
Zheng, Y., Chilton, B., et al. (2008b). Impaired DNA damage response, genome
instability, and tumorigenesis in SIRT1 mutant mice. Cancer Cell 14, 312-323.

References
147
Wang, S., Rosengren, L., Hamberger, A., and Haglid, K. (2001). Antisense inhibition
of BCL-2 expression induces retinoic acid-mediated cell death during differentiation of
human NT2N neurons. Journal of Neurochemistry 76, 1089-1098.
Wang, Y., Schwedes, J.F., Parks, D., Mann, K., and Tegtmeyer, P. (1995). Interaction
of p53 with its consensus DNA-binding site. Molecular and Cellular Biology 15, 2157-
2165.
Weber, G.F., and Menko, A.S. (2005). The canonical intrinsic mitochondrial death
pathway has a non-apoptotic role in signaling lens cell differentiation. The Journal of
Biological Chemistry 280, 22135-22145.
Wei, J.S., Song, Y.K., Durinck, S., Chen, Q.R., Cheuk, A.T., Tsang, P., Zhang, Q.,
Thiele, C.J., Slack, A., Shohet, J., et al. (2008). The MYCN oncogene is a direct target of
miR-34a. Oncogene 27, 5204-5213.
Wei, L., Cui, L., Snider, B.J., Rivkin, M., Yu, S.S., Lee, C.S., Adams, L.D., Gottlieb,
D.I., Johnson, E.M., Jr., Yu, S.P., et al. (2005). Transplantation of embryonic stem cells
overexpressing Bcl-2 promotes functional recovery after transient cerebral ischemia.
Neurobiology of Disease 19, 183-193.
Welch, C., Chen, Y., and Stallings, R.L. (2007). MicroRNA-34a functions as a
potential tumor suppressor by inducing apoptosis in neuroblastoma cells. Oncogene 26,
5017-5022.
Williams, D.W., Kondo, S., Krzyzanowska, A., Hiromi, Y., and Truman, J.W. (2006).
Local caspase activity directs engulfment of dendrites during pruning. Nature
Neuroscience 9, 1234-1236.
Wong, K., Zhang, J., Awasthi, S., Sharma, A., Rogers, L., Matlock, E.F., Van Lint, C.,
Karpova, T., McNally, J., and Harrod, R. (2004). Nerve growth factor receptor signaling
induces histone acetyltransferase domain-dependent nuclear translocation of p300/CREB-
binding protein-associated factor and hGCN5 acetyltransferases. The Journal of
Biological Chemistry 279, 55667-55674.
Wulczyn, F.G., Smirnova, L., Rybak, A., Brandt, C., Kwidzinski, E., Ninnemann, O.,
Strehle, M., Seiler, A., Schumacher, S., and Nitsch, R. (2007). Post-transcriptional
regulation of the let-7 microRNA during neural cell specification. FASEB Journal 21,
415-426.

References
148
Xu, H., He, J.H., Xiao, Z.D., Zhang, Q.Q., Chen, Y.Q., Zhou, H., and Qu, L.H. (2010).
Liver-enriched transcription factors regulate microRNA-122 that targets CUTL1 during
liver development. Hepatology 52, 1431-1442.
Xu, Y. (2003). Regulation of p53 responses by post-translational modifications. Cell
Death and Differentiation 10, 400-403.
Yamakuchi, M., Ferlito, M., and Lowenstein, C.J. (2008). miR-34a repression of
SIRT1 regulates apoptosis. Proceedings of the National Academy of Sciences of the
United States of America 105, 13421-13426.
Yang, A., Walker, N., Bronson, R., Kaghad, M., Oosterwegel, M., Bonnin, J., Vagner,
C., Bonnet, H., Dikkes, P., Sharpe, A., et al. (2000). p73-deficient mice have
neurological, pheromonal and inflammatory defects but lack spontaneous tumours.
Nature 404, 99-103.
Yi, C.H., and Yuan, J. (2009). The Jekyll and Hyde functions of caspases.
Developmental Cell 16, 21-34.
Ying, Q.L., Stavridis, M., Griffiths, D., Li, M., and Smith, A. (2003). Conversion of
embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nature
Biotechnology 21, 183-186.
You, H., Jang, Y., You-Ten, A.I., Okada, H., Liepa, J., Wakeham, A., Zaugg, K., and
Mak, T.W. (2004). p53-dependent inhibition of FKHRL1 in response to DNA damage
through protein kinase SGK1. Proc Natl Acad Sci U S A 101, 14057-14062.
Yu, F., Yao, H., Zhu, P., Zhang, X., Pan, Q., Gong, C., Huang, Y., Hu, X., Su, F.,
Lieberman, J., et al. (2007a). let-7 regulates self renewal and tumorigenicity of breast
cancer cells. Cell 131, 1109-1123.
Yu, J., Vodyanik, M.A., Smuga-Otto, K., Antosiewicz-Bourget, J., Frane, J.L., Tian,
S., Nie, J., Jonsdottir, G.A., Ruotti, V., Stewart, R., et al. (2007b). Induced pluripotent
stem cell lines derived from human somatic cells. Science 318, 1917-1920.
Yuan, J., Shaham, S., Ledoux, S., Ellis, H.M., and Horvitz, H.R. (1993). The C.
elegans cell death gene ced-3 encodes a protein similar to mammalian interleukin-1 beta-
converting enzyme. Cell 75, 641-652.
Yuasa, S. (2001). Development of astrocytes in the mouse hippocampus as tracked by
tenascin-C gene expression. Archives of Histology and Cytology 64, 149-158.

References
149
Zapata, J.M., Llobet, D., Krajewska, M., Lefebvre, S., Kress, C.L., and Reed, J.C.
(2009). Lymphocyte-specific TRAF3 transgenic mice have enhanced humoral responses
and develop plasmacytosis, autoimmunity, inflammation, and cancer. Blood 113, 4595-
4603.
Zermati, Y., Garrido, C., Amsellem, S., Fishelson, S., Bouscary, D., Valensi, F., Varet,
B., Solary, E., and Hermine, O. (2001). Caspase activation is required for terminal
erythroid differentiation. The Journal of Experimental Medicine 193, 247-254.
Zhang, J., Yan, W., and Chen, X. (2006). p53 is required for nerve growth factor-
mediated differentiation of PC12 cells via regulation of TrkA levels. Cell Death and
Differentiation 13, 2118-2128.
Zhang, K.Z., Westberg, J.A., Holtta, E., and Andersson, L.C. (1996). BCL2 regulates
neural differentiation. Proceedings of the National Academy of Sciences of the United
States of America 93, 4504-4508.
Zhao, C., Deng, W., and Gage, F.H. (2008a). Mechanisms and functional implications
of adult neurogenesis. Cell 132, 645-660.
Zhao, H., Kalota, A., Jin, S., and Gewirtz, A.M. (2009). The c-myb proto-oncogene
and microRNA-15a comprise an active autoregulatory feedback loop in human
hematopoietic cells. Blood 113, 505-516.
Zhao, T., and Xu, Y. (2010). p53 and stem cells: new developments and new concerns.
Trends in Cell Biology 20, 170-175.
Zhao, Y., Yin, X., Qin, H., Zhu, F., Liu, H., Yang, W., Zhang, Q., Xiang, C., Hou, P.,
Song, Z., et al. (2008b). Two supporting factors greatly improve the efficiency of human
iPSC generation. Cell Stem Cell 3, 475-479.
Zheng, H., Ying, H., Yan, H., Kimmelman, A.C., Hiller, D.J., Chen, A.J., Perry, S.R.,
Tonon, G., Chu, G.C., Ding, Z., et al. (2008). p53 and Pten control neural and glioma
stem/progenitor cell renewal and differentiation. Nature 455, 1129-1133.





![No limiar do universo aranha 01 [de 05]](https://cdn.vdocuments.us/doc/165x107/579054ea1a28ab900c92a259/no-limiar-do-universo-aranha-01-de-05.jpg)

