
Introduction to
Genetic AnalysisTENTH EDITION
Introduction to
Genetic AnalysisTENTH EDITION
Griffiths • Wessler • Carroll • Doebley
© 2012 W. H. Freeman and Company
CHAPTER 12Regulation of Gene Expression
in Eukaryotes

CHAPTER OUTLINE12.1 Transcriptional regulation in eukaryotes: an overview
12.2 Lessons from yeast: the GAL system
12.3 Dynamic chromatin
12.4 Short-term activation of genes in a chromatin environment
12.5 Long-term inactivation of genes in a chromatin environment
12.6 Gender-specific silencing of genes and whole chromosomes
12.7 Post-transcriptional gene repression by miRNAs

The first cloned mammal
Dolly, the Finn Dorset lamb in 1996 and her surrogate Scottish Blackface mother
Dolly and Bonnie

Dolly in Royal Museum of Scotland
Ian Wilmut

Dolly
Dolly Parton

globinmyosin
erythrocytemuscle cell
All genes
housekeeping
All cells have the same genome, but each cell expresses only a subset of all genes
Cells differ in gene expression

Overview of transcriptional regulation
nucleus(membrane)
chromatin

Gene regulation at multiple levels
TransportLocalizationModificationComplex formationDegradation
Many regulatory proteins have to import into nucleus

Promoter-proximal elements precede the promoter of a eukaryotic gene

Promoter-proximal elements are necessary for efficient transcription
Point mutations throughout the promoter region were analyzed for their effects on transcription rates. The height of each line represents the transcription level relative to a wild-type promoter or promoter-proximal element (1.0).

Transcription factors need multiple functional domains
1. A domain that recognizes a DNA regulatory sequence (the protein’s DNA-binding site)
2. A domain that interacts with one or more proteins of the transcriptional apparatus (RNA polymerase or a protein associated with RNA polymerase)
3. A domain that interacts with proteins bound to nearby regulatory sequences on DNA such that they can act cooperatively to regulate transcription
4. A domain that influences chromatin condensation either directly or indirectly
5. A domain that acts as a sensor of physiological conditions within the cell
Direct or indirect (by interacting with other proteins)

Model Organism Yeast
Brewer’s yeast Baker’s yeast
Saccharomyces cerevisiaeBudding yeast

The Gal pathway
Expressed at low level
Induced by galactose
Regulated by Gal4

Transcriptional activator proteins bind to UAS elements in yeast
UAS: Upstream Activation Sequences
Binding site for Gal4
Can be far from promoter

Transcriptional activator proteins are modular
Reporter gene
domain swap

Transcriptional activator proteins may be activated by an inducer
Galactose: inducing signal

Transcriptional activator proteins recruit the transcriptional machinery
Co-activator:Does not directly bind DNA
Specific recognition of target sequenceEnhancer can function
far away from promoter
Enhancer can function upstream or downstream, even far away

Transcriptional complexes

Combinations of regulatory proteins control cell types
Mating type
Combinations of binding partners => different binding specificities

The structure of chromatin
~150 bp linker DNA

A nucleosome is composed of DNA wrapped around eight histones
Histone octamer (H2A2H2B2H32H42)
DNA exposed on the outside

The structure of chromatin

The structure of chromatin
Euchromatin(loose)
Heterochromatin
•Condensed
•Repetitive sequences
•Late replicating
•Genes silenced

Chromatin remodeling exposes regulatory sequences
Shifting of nucleosome position
Exposes regulatory sequences
Linker DNA: sensitive to nuclease
Nucleosomal DNA: protected from nuclease digestion
Use nuclease sensitivity to determine chromatin state (open/closed) or nucleosome position
+ SWI-SNF + ATP

The SWI-SNF complex for chromatin remodeling
Yeast mutant screen
sugar nonfermenting (snf)
Mating type switch (swi)
swi2=snf2
swi2/snf2 (“switch-sniff”) locus
SWI-SNF complex

Modifications of histone tails results in chromatin remodeling
Histone tails are exposed, can be modified
Modifies Lysine (K) and Arginine (R) (basic aa)
Acetylation: negative charges => repulsion

Histone modifications

Histone modifications

Acetylation of histones
Histone acetyltransferase (HAT)
Histone de-acetylase (HDAC)

Histone modifications
Alternative modifications on the same residue
Histone code

Regulation of gene expression by histone acetylation

Histone deacetylation can turn off gene transcription
HDAC(corepressor)

Inheritance of chromatin states
Epigenetic memory: heritable traits (over rounds of cell division and sometimes transgenerationally) that do not involve changes to the underlying DNA sequence. (e.g. chromatin state)

Methylation of DNA
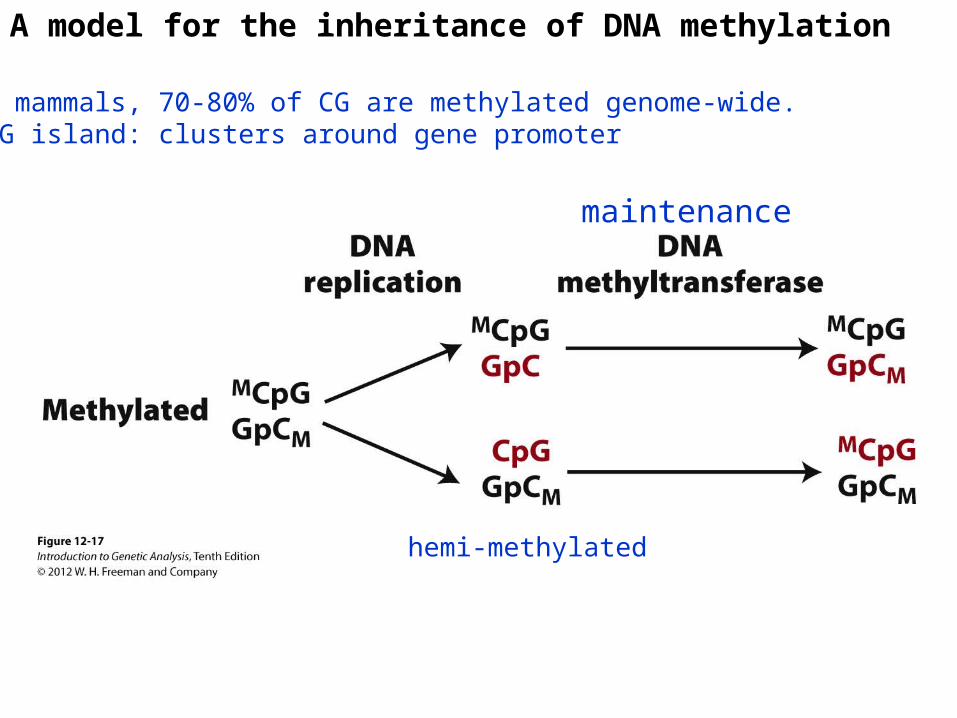
A model for the inheritance of DNA methylation
In mammals, 70-80% of CG are methylated genome-wide.CpG island: clusters around gene promoter
hemi-methylated
maintenance

Enhanceosomes help recruit the transcriptional machinery

Enhanceosomes recruit chromatin remodelers
Enhancers contain binding sites for many transcription factors, which bind and interact cooperatively.

Enhancer-blocking insulators prevent enhancer activation

Model for how enhancer-blocking insulators might work

Mating-type switching is controlled by recombination of DNA cassettes
ds break in MAT made by HO endonuclease
=> gene conversion
silent information regulators (SIR)Sir2 (HDAC)

Gene silencing is caused by the spread of heterochromatin
w+ is expressed in some cells => not a mutation in w gene
Clonal => epigenetic memory
Position-effect variegation (PEV)

Heterochromatin in Drosophila chromosomes
~30% of genome
H3K4me2, enriched in euchromatinH3K9me2, enriched in heterochromatin

Some genes enhance or suppress the spread of heterochromatin
Enhancer
Suppresor
Su(var)2-5 = HP1 (heterochromatic protein 1)Su(var)3-9 = histone methylatransferase

Multiple states of Lysine methylation

Heterochromatin may spread farther in some cells than in others

Barrier insulators stop the spread of heterochromatin

Genomic imprinting

Genomic imprinting
Phenotype depends on the parental origin of the genes

Inactivation of genes and chrosomomes

Genomic imprinting
No changes in DNA sequence
mouse/human ~100 imprinted genes

Genomic imprinting requires insulators
DNA methylation
imprinting control region

Unusual inheritance of imprinted genes

Steps required for imprinting
Igf2: maternal imprinting (inactive)H19: paternal imprinting (inactive)
H19Igf2

X inactivation
Dosage compensation for X chromosome female: XX male: XY

Barr body and Lyon Hypothesis of X inactivation
Murray Barr: discoverer
Mary Lyon
Epigenetic memory
Xi: H3K9me, histone hypoacetylation, DNA hypermethylation~ heterochromatin

Xist
non-coding

Xist RNA covers one of the two copies of the X chromosome
RNA fluorescent in situ hybridization (FISH)
metaphase chromosomes
female fibroblast cell line
Xist expression => cis-inactivation

A model for X-chromosome inactivation

Possible models for the repression of translation by miRNA







