
American Journal of Medical Genetics Supplement 2: 183-190 (1986) Developmental Field Concept:393-400
Genetic Analysis of Cleft Lip With or Without Cleft Palate in Chinese Kindreds
Michael Melnick, Mary L. Marazita, and Dan-Ning Hu
Craniofacial Biology Program, University of Southern California, Los Angeles (M. M.), and Mental Retardation Research Center, UCLA Medical Center, Los Angeles (M. L. M.), California; Population Genetics Research Group, Shanghai Society of Genetics, Shanghai, China (D.-N. H.)
Cleft lip with or without cleft palate (CL P) affects 1 in 500-1,000 newborns worldwide; the cause remains unclear. For CL & P, Asians are at higher risk than Caucasians or Blacks. This report presents the results of a genetic analysis of 163 CL f P proband families ascertained in Shanghai, China. Based on statistical tests of several predictions from the classical multifactorial/threshold model, no evidence could be found to support it. Further, goodness-of-fit (PGOODFIT) tests of this model were also unsupportive, there being more families with two or more affected sibs than expected. Classical segregation analysis for Mendelian inheri- tance showed a maximum likelihood estimate of the segregation ratio of 0.187 0.068. This value was not significantly less than 0.25 (recessive inheri- tance) but was significantly less than 0.50 (dominant inheritance). Sex ratio data from these Chinese families suggest that manifestation of the putative single major gene is sex influenced. The apparent reduced penetrance is likewise to be expected if manifestation of CL -t P also depends on in utero exposure to deleterious environmental agents, as demonstrated in a variety of human and animal model studies.
Key words: cleft lip and palate, genetics, sex ratio
INTRODUCTION
Cleft lip with or without cleft palate (CL f P) is a major public health problem worldwide. Approximately 1 in 500-1,OOO newborns is affected with this malforma- tion, the incidence varying by race and nationality [Melnick et al, 19801. Despite nearly half a century of intensive investigation, the cause of CL f P remains elusive. Unquestionably, both genetic and environmental factors play a role in all human
Received for publication June 21, 1985; revision received November 4, 1985.
Address reprint requests to Michael Melnick, D.D.S., Ph.D., Craniofacial Biology Program, DEN 4242,University of Southern California, Los Angeles, CA 90089-0641.
0 1986 Alan R. Liss, Inc.

184 Melnick, Marazita, and Hu
development including malformation. However, what remains unclear for CL * P is: 1) the nature of the genetic component; 2) the proportion of persons with the relevant genotype that expresses the phenotype; 3) the relative importance of environ- mental effects. Although numerous studies have been done worldwide, real progress in understanding the cause of CL & P (and ultimately its prevention) awaits unequi- vocal answers to all three of these critical questions. Though empiric risk figures have value, they are inadequate substitutes for genetic risk estimates based on clear knowledge of causation.
The genetic model most often proposed for nonsyndromic CL k P is multifac- torial threshold inheritance (MF/T) [Carter, 1976; Fraser, 1970; 19761, although it has never been satisfactorily demonstrated for well-ascertained pedigree data [Mel- nick et al, 19801. Recently, Carter et a1 [ 19821 reported that their large British CL P data set was suggestive of MF/T, but they tested neither that hypothesis nor the one of genetic heterogeneity.
However, other studies, do provide some important causal clues, primarily because they have utilized well-designed mathematical analyses of population and family data. Chung et a1 [1974] performed a large study involving various racial crosses in Hawaii. Although the complex segregation analysis offered no clear-cut discrimination between the single major-gene and MF/T models, the best-fitting single major-gene hypothesis appeared to be autosomal recessive inheritance with reduced penetrance (0.195), no phenocopies, and an estimated gene frequency of 0.0811. According to Chung et a1 [ 19741, several other investigators had demonstrated that very high heritability can be considered an indicator of the presence of major genes. They concluded that the heritability for CL & P in their study population of 0.99 (after adjusting for ascertainment probability and affected parents) was consistent with the single major-gene hypothesis, as was the observed concordance rate in published monozygotic twin data.
Studies of CL + P in Denmark have provided similar results. Melnick et a1 [1980], after statistically testing and rejecting the MF/T model, took a fresh look at one of the older and more simple multifactorial models, that of a single major-gene and heterogeneous environmental triggers that are present during embryogenesis. Using the mathematical relationships Penrose [ 19531 derived for this purpose, and the frequency of CL f P in the general Danish population and in sibs of CL k P probands, they were able to derive estimates under the hypothesis of recessive inheritance: the estimated gene frequency being 0.0864 and penetrance 0.174. Thus, the frequency of genetically susceptible embryos would be about 1 in 135, a common condition; manifestation would depend on in utero exposure to appropriate environ- mental agents during the critical period of facial development, and this would lower the frequency of CL & P to about 1 in 800. These results were remarkably similar to those of Chung et a1 [1974] and are supported by a variety of animal model data [Goldman, 1984; Gupta et al, 1985; Melnick et al, 1981al. Additional analyses of the Danish kindreds [Marazita et al, 19841 again rejected the MF/T model, and found evidence of genetic heterogeneity for CL k P with the strong possibility of a major autosomal recessive gene in a substantial portion of the kmdreds.
Asians (Chinese, Japanese, Koreans, and Filipinos) are clearly at higher risk for CL -t P than Caucasians or Blacks [Chung et al, 1974; Hu et a1 1982; Koguchi, 1975; Tanaka et al, 19691. These racial differences still persist in Hawaii, where the environment is relatively uniform among different races and after removal of ascer-
394:DFC

Genetics of CL+P in Shanghai 185
TABLE I. Sex Ratios in CL f P Proband Sibships
Male Female Ratio X:. P
CL k Pprobands 102 61 1.67 8.872 <0.005 CL k P sibs 6 6 1 .OO 0.005 >0.90 Unaffected sibs 108 135 0.80 4.179 <0.05 Total 216 202 1.07 0.086 >0.50
aCalculated from an expected M:F ratio of 1.04.
tainment biases [Chung et al, 19741. To date, we have looked to limited and inconclu- sive Japanese studies [Koguchi, 1975; 1980; Tanaka et al, 19691 for family data on Asians in their country of origin. The need to perform genetic hypothesis testing with families who reside in various Asian countries remains.
The present study is a genetic analysis of 163 CL 5 P proband families ascer- tained in Shanghai, China, by Hu et a1 [1982]. Predictions of the MF/T model are tested; goodness-of-fit tests of the MF/T model and segregation analysis are also utilized to clarify the genetic component of the cause of CL f P in this study population.
MATERIALS AND METHODS
Medical records on file at the Shanghai International Peace Maternity and Infant Hospital were used to determine the incidence of CL L- P among births occurring in this hospital for the years 1971-1980 [Hu et al, 19821. Every newborn infant was examined by a physician, and any apparent congenital anomaly was recorded. Over the 10-year period, 60 nonsyndromic cases of CL f P were recorded among 45,072 newborn infants; this is an incidence of 1.33 per 1,OOO births. This incidence is midway between that of the Japanese (1.70) and Caucasian (- 1.10) populations. For family studies, nonsyndromic CL P probands were ascertained through the afore- mentioned hospital and Department of Plastic Surgery, Ninth People Hospital (Shang- hai Second Medical College). A detailed family history was obtained from each case, including parental consanguinity. A total of 163 probands, 583 first-degree relatives, 1,032 second-degree relatives, and 1,727 third-degree relatives were reviewed. Re- garding the 163 nonsyndromic CL * P probands, 17 had cleft lip alone and 146 had cleft lip and cleft palate.
The predictions of the MF/T model [Carter, 19761 were tested in the usual manner [Melnick et al, 19801. The MF/T model was also investigated using the goodness-of-fit test (PGOODFIT) described by Gladstien et a1 [ 19781. Simple Men- delian inheritance was tested by the classical segregation analysis of Morton [ 19621.
RESULTS
As noted in Table I, the M/F proband sex ratio of 1.67 was significantly greater than expected. Altered sex ratios of the kind observed here may have several explanations, including sex-influenced inheritance, biased gametic selection, and sex- biased, natural parental selection. Given Nishimura’s [ 19751 observation that the frequency of CL f P was far greater in spontaneous abortions than in live births, it
DFC:395
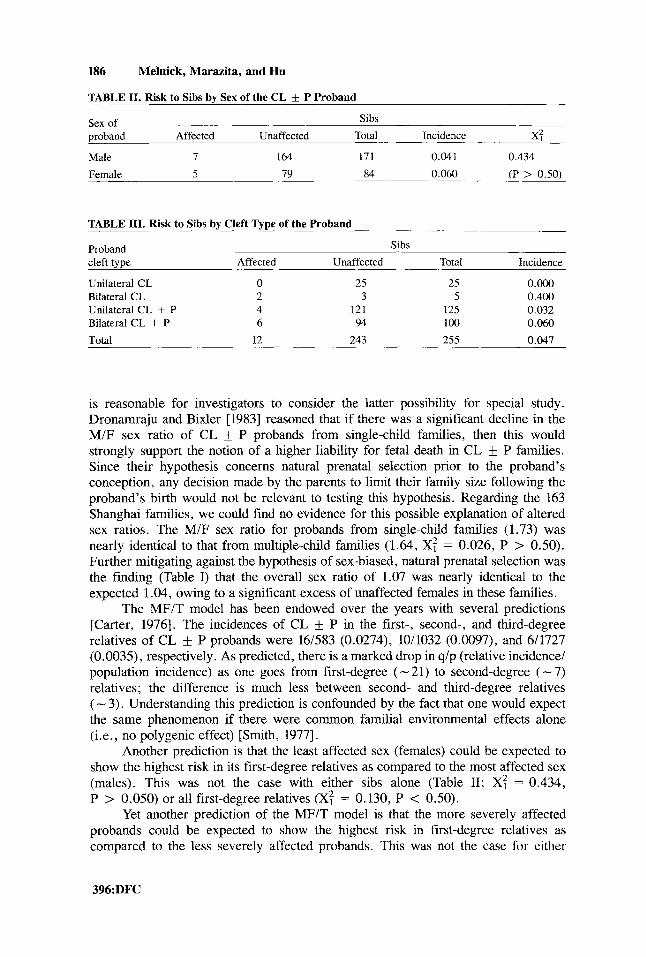
186 Melnick, Marazita, and Hu
TABLE 11. Risk to Sibs by Sex of the CL f P Proband
Sex of Sibs proband Affected Unaffected Total Incidence x: Male 7 164 171 0.041 0.434
Female 5 79 84 0.060 (P > 0.50)
TABLE 111. Risk to Sibs by Cleft Type of the Proband
Proband cleft type Affected Unaffected Total Incidence
Unilateral CL 0 25 25 O.Oo0 Bilateral CL 2 3 5 0.400 Unilateral CL + P 4 121 125 0.032 Bilateral CL + P 6 94 100 0.060
Total 12 243 255 0.047
Sibs
is reasonable for investigators to consider the latter possibility for special study. Dronamraju and Bixler [1983] reasoned that if there was a significant decline in the M/F sex ratio of CL f P probands from single-child families, then this would strongly support the notion of a higher liability for fetal death in CL f P families. Since their hypothesis concerns natural prenatal selection prior to the proband’s conception, any decision made by the parents to limit their family size following the proband’s birth would not be relevant to testing this hypothesis. Regarding the 163 Shanghai families, we could find no evidence for this possible explanation of altered sex ratios. The M/F sex ratio for probands from single-child families (1.73) was nearly identical to that from multiple-child families (1.64, X: = 0.026, P > 0.50). Further mitigating against the hypothesis of sex-biased, natural prenatal selection was the finding (Table I) that the overall sex ratio of 1.07 was nearly identical to the expected 1.04, owing to a significant excess of unaffected females in these families.
The MF/T model has been endowed over the years with several predictions [Carter, 19761. The incidences of CL f P in the first-, second-, and third-degree relatives of CL f P probands were 16/583 (0.0274), 10/1032 (0.0097), and 6/1727 (0.0035), respectively. As predicted, there is a marked drop in q/p (relative incidence/ population incidence) as one goes from first-degree ( - 21) to second-degree ( - 7) relatives; the difference is much less between second- and third-degree relatives ( - 3). Understanding this prediction is confounded by the fact that one would expect the same phenomenon if there were common familial environmental effects alone (i.e., no polygenic effect) [Smith, 19771.
Another prediction is that the least affected sex (females) could be expected to show the highest risk in its first-degree relatives as compared to the most affected sex (males). This was not the case with either sibs alone (Table 11; X: = 0.434, P > 0.050) or all first-degree relatives (XT = 0.130, P < 0.50).
Yet another prediction of the MF/T model is that the more severely affected probands could be expected to show the highest risk in first-degree relatives as cornparcd to the less severely affected probands. This was not the case for either
396:DFC

Genetics of CL P in Shanghai 187
TABLE IV. PGOODFIT Test of the MF/T Model
Ascertainment probability ( K ) ~ Heritability (H2) 0.1 0.3 0.5 0.7 0.9
0.3 3.1 x lo-'' 9.0 x lo-" 2.2 x lo-" 4.1 x 5.3 x 10-" 0.1 9.1 x 1 0 - ~ ~ 4.0 x 10- l~ 2.5 x 1 0 - l ~ 2.0 x 1 0 - l ~ 1 . 8 x 1 0 - l ~
0.5 1.3 x 10-6 4.1 x 10-7 1.1 x 10-7 2.2 x 10-8 3.0 x 10-9 0.7 5.5 x lop4 1.9 x 1 0 - ~ 6.0 x 1.4 x 2.3 x 1 0 - ~ 0.9 2.6 x 1.2 x lo-* 4.4 x 1.3 x 2.6 x lop4
aEntries are the probabilities (P-values) of obtaining the observed number (11) or more of nuclear families with two or more affected children under the MFlT model for various heritability values.
comparisons of CL and CL f P (0.067 and 0.044, respectively) or comparisons of unilateral CL t- P and bilateral (X: = 1.03, P > 0.10); the numbers were too small for comparisons of unilateral and bilateral CL (Table 111).
Finally, it is predicted that there will be a higher consanguinity rate in these families than in the general population. The frequency of first-cousin marriages was 1.84% in these families as compared to 0.77% in the general population; these rates were not significantly different (X: = 2.42, P > 0.10). In summary, then, based on tests of the stated predictions of the MF/T model [Carter, 19761, we found essentially no evidence for it in this study population. In this regard, this Shanghai data set is like all others worldwide [Melnick et al, 19801.
We next performed two other genetic analyses on this group of CL f P families: goodness-of-fit tests of the MF/T model and classical segregation analysis. These analyses were done for 109 proband nuclear families, excluding consanguinous and single-child families. The trait analyzed was "cleft (CL k P) or not." The goodness-of-fit test of the MF/T model has been described by Gladstien et a1 [ 19781. The method incorporates the necessary corrections for ascertainment bias. The test is applicable to disorders showing different incidence rates in males and females, and involves a comparison of the observed number of nuclear families having at least two affected offspring with the expected distribution of such families under the MF/T model. The computation of the expected distribution is conditioned on several family- dependent variables, including parental affected status, numbers of sons and daugh- ters, and whether each family was ascertained through only affected males or through at least one affected female. The probabilities of observing the actual study data are derived for a range of ascertainment probabilities (r) and heritabilities (h2).
In 11 of the 109 nuclear families there were 2 or more affected sibs. For a range of P (0.1-0.9) and h2 (0.1-0.9) two probabilities (P-values) were calculated: (a) the probability of observing 11/109 or more families with two or more affected children under the MF/T model, and (b) the probability of observing fewer than 11/109. Regarding the P-values for (a), for every combination of T and h2 it was less than 0.03 (Table IV); regarding the P-values for (b), for each combination of P and h2 it was equal to 1.0. Thus, the MF/T model for these data can be rejected because there are more families with two or more affected sibs than would be expected under this model.
An alternative to the MF/T model is simple Mendelian inheritance. This was tested in the same 109 families by the classical segregation analysis of Morton [ 19621.
DFC:397

188 Melnick, Marazita, and Hu
Maximum likelihood estimates of the segregation ratio (p) and proportion of sporadic cases (x) were obtained under a model of segregation with incomplete ascertainment (a < l), admixture of sporadic cases and a uniform segregation frequency. The sample size was too small to simultaneously estimate T. Thus, we set a = 0.832 for all analyses (?r = R/A, where R = no of probands and A = total no of affected persons). The maximum likelihood estimate of p was 0.187 f 0.068 and for x 0.206 0.228. The value of p is not significantly less than 0.25 (the expected value for recessive inheritance) but is significantly less than 0.50 (the expected value for dominant inheritance). The value of x is not significantly greater than 0. In summary, these results are consistent with autosomal recessive inheritance of CL k P in these 109 Chinese families.
DISCUSSION
To summarize our analyses of 163 CL k P proband families from Shanghai, there is no evidence to support the MF/T model but there is evidence for autosomal recessive inheritance of CL & P. It should be noted that regarding our segregation analysis, the likelihood surface was rather flat (probably due to sample size) and the above estimates may thus represent local maxima. Future analyses will include many additional families and should solve any convergence problems. Further, we recog- nize that the best statistical approach is to test the MF/T model and Mendelian hypothesis simultaneously under the mixed model [Morton and MacLean, 19741. Again, this requires larger sample sizes, like those used with the Danish population [Marazita et al, 19841. Plans are under way to ascertain similarly large sample sizes in Shanghai for the mixed-model approach. With these caveats duly considered, we conclude that the outcome of this study provides an important piece to the causal puzzle.
Beginning with the extensive study by Chung et a1 [1974], and through the studies of Melnick et a1 [1980], Marazita et a1 [1984], and the present one, the knowledge of CL f P etiology appears to be coming more clear. This process has been slow, but surely more effective owing to real hypothesis testing.
Firstly, these aforementioned studies have found little or no evidence to support the MF/T model of Carter [1976] and Fraser [1976]. Indeed, this is evident from statistical analysis of data sets worldwide [Melnick et al, 19801. In this regard, the Shanghai data set does not differ from the rest.
Secondly, there appears to be growing evidence that a major autosomal recessive gene is an important causal element of a substantial portion of the CL f P cases. This was initially suggested by Chung et a1 [1974], then Melnick et a1 [1980], and most recently by Mazarita et a1 119841. With the present findings on 109 Shanghai families consistent with autosomal recessive inheritance, it would appear that this inheritance pattern cuts across Asian and Caucasian racial lines. The data from these studies, including the present one, also suggest a reduced penetrance of approximately 20-25 % . This is entirely expected if manifestation of CL k P also depends upon in utero exposure to deleterious environmental agents during the critical period of facial development, as suggested by Melnick et a1 [1980] and demonstrated in a variety of animal model studies of CL f P and isolated cleft palate [Melnick et al, 1981a,b; Goldman, 19841.
398:DFC

Genetics of CL+P in Shanghai 189
Finally, the sex ratio data from these Chinese families (Table I) is most intrigu- ing. There is the usual excess of males among the CL f P probands, but the overall sibship sex ratio is not at all deviant, owing to the excess of unaffected females. This differs substantially from the Caucasian data [Melnick et al, 19801, where it is found that the overall family sex ratio also deviates significantly from normal expectations, and there appears to be an overall deficiency of females in the proband families. Considered together with the evidence for an autosomal recessive gene, this rather striking finding in the Chinese families would suggest that manifestation of the putative single major gene is sex influenced. At least in these Shanghai families, the altered proband sex-ratio does not appear to be a spurious finding based on liveborn ascertainment bias. Rather, it appears to be biologically significant.
REFERENCES
Carter CO (1976): Genetics of common single malformations. Br Med Bull 32:21-26. Carter CO, Evans K, Coffey R, Roberts JAF, Buck A, Roberts MF (1982): A three generation family
study of cleft lip with or without cleft palate. J Med Genet 19:246-261. Chung CS, Ching GHS, Morton NE (1974): A genetic study of cleft lip and palate in Hawaii. 11.
Complex segregation analysis and genetic risks. Am J Hum Genet 26:177-188. Dronamraju KR, Bixler D (1983): Fetal mortality in oral cleft families (VI): A search for early
embryonic and zygotic mortality. Clin Genet 24:346-349. Fraser FC (1970): Genetics of cleft lip and cleft palate. Am J Hum Genet 22:336-352. Fraser FC (1976): The multifactorial threshold concept-uses and misuses. Teratology 14:267-280. Gladstien K, Lange K, Spence MA (1978): A goodness-of-fit test for the polygenic threshold model:
Application to pyloric stenosis. Am J Med Genet 2:7-13. Goldman AS (1984): Biochemical mechanics of glucocorticoid- and phenytoin-induced cleft palate. In
Zimmerman EF (ed): “Current Topics in Developmental Biology.” New York: Academic Press, Inc., pp 217-239.
Gupta C, Katsumata M, Goldman AS, (1985): H-2 histocompatability region influences the inhibition of arachidonic acid cascade by dexamethasone and phenytoin in mouse embryonic palates. J Cranio- facial Genet Devel Biol 5:277-285.
Hu DN, Li JH, Chen HY, Chang HS, Wu BX, Lu ZK, Wang DZ, Liu XG (1982): Genetics of cleft lip and cleft palate in China. Am J Hum Genet 34
Koguchi H (1975): Recurrence rate in offspring and s atients with cleft lip and/or cleft palate. Jpn J Hum Genet 20:207-221.
Koguchi H (1980): Population data on cleft lip and cleft palate in the Japanese. In Melnick M, Bixler D, Shields , ED (eds): “Etiology of Cleft Lip and Cleft Palate.” New York: Alan R. Liss, Inc., pp
Marazita ML, Spence MA, Melnick M (1984): Genetic analysis of cleft lip with or without cleft palate in Danish kindreds. Am J Med Genet 19:9-18.
Melnick M, Bixler D, Fogh-Andersen P, Conneally PM (1980): Cleft lip f cleft palate: An overview of the literature and an analysis of Danish cases born between 1941 and 1968. Am J Med Genet
Melnick M, Jaskoll T, Slavkin HC (1981a): Corticosteroid-induced cleft palate in mice and H-2 haplotype: Maternal and embryonic effects. Immunogenetics 13:443-450, 1981.
Melnick M, Jaskoll T, Slavkin HC (1981b): Corticosteroid-induced cleft lip in mice: A teratologic, topographic, and histologic investigation. Am J Med Genet 10:333-350.
Morton NE (1962): Segregation and linkage. In Burdette WJ (ed): “Methodolgy in Human Genetics.” San Francisco: Holden-Day, pp 17-52.
Morton NE, MacLean CJ (1974): Analysis of family resemblance. 111. Complex segregation of quanti- tative traits. Am J Hum Genet 26:489-503.
Nishimura H (1975): Prenatal versus postnatal malformations based on the Japanese experience on induced abortions in the human being. In Blandau RJ (ed): “Ageing Gametes.” Basel: S.Karger,
297-323.
6~83-97.
DFC:399

190 Melnick, Marazita, and Hu
pp 349-368. Penrose LS (1953): The genetical background of common diseases. Acta Genet 4:257-265. Smith C (1977): The multifactorial model of liability in familial disease: Methods, applications, and
limitations. In Japan Medical Research Foundation (ed): “Gene-Environment Interaction in Common Diseases.” Tokyo: University of Tokyo Press, pp 127-138.
Tanaka K, Fujino H, Fujita Y, Tashiro H, Sanui Y (1969): Cleft lip and palate: Some evidences for the multifactorial trait and estimation of heritability based upon Japanese data. Jpn J Hum Genet 14: 1-9.
Edited by John M. Opitz and James F. Reynolds
400:DFC







