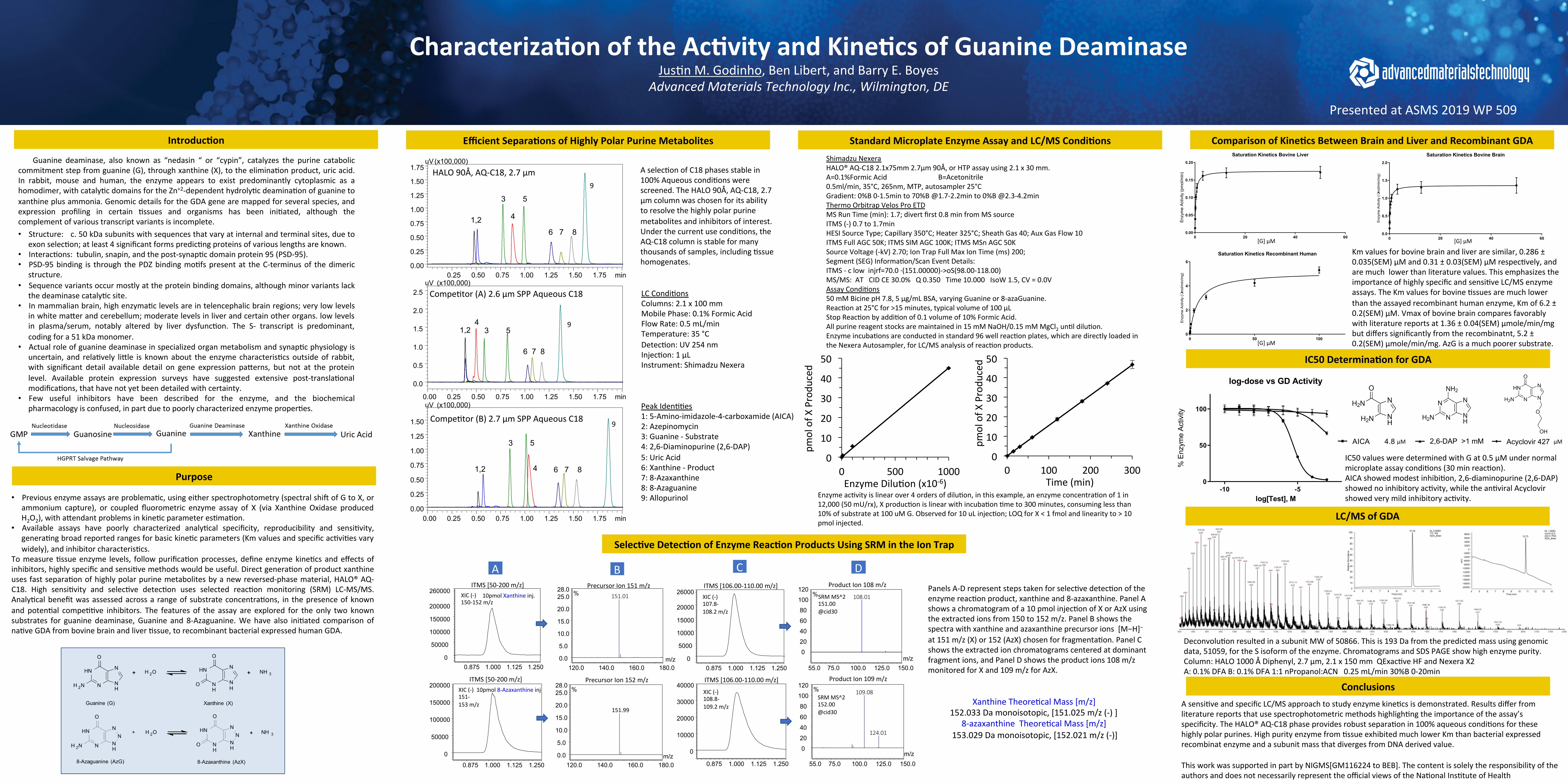
IC50Determina.onforGDA
IC50valuesweredeterminedwithGat0.5µMundernormalmicroplateassaycondi<ons(30minreac<on).AICAshowedmodestinhibi<on,2,6-diaminopurine(2,6-DAP)showednoinhibitoryac<vity,whilethean<viralAcyclovirshowedverymildinhibitoryac<vity.
-10 -50
50
100
log-dose vs GD Activity
log[Test], M
% E
nzym
e A
ctiv
ity AICA 4.8 mM
2,6-DAP >1 mM
Acyclovir 427 mM
-10 -50
50
100
log-dose vs GD Activity
log[Test], M
% E
nzym
e A
ctiv
ity AICA 4.8 mM
2,6-DAP >1 mM
Acyclovir 427 mM
-10 -50
50
100
log-dose vs GD Activity
log[Test], M
% E
nzym
e A
ctiv
ity AICA 4.8 mM
2,6-DAP >1 mM
Acyclovir 427 mM
-10 -50
50
100
log-dose vs GD Activity
log[Test], M
% E
nzym
e A
ctiv
ity AICA 4.8 mM
2,6-DAP >1 mM
Acyclovir 427 mM
Guanine deaminase, also known as “nedasin “ or “cypin”, catalyzes the purine cataboliccommitmentstepfromguanine(G),throughxanthine(X),totheelimina<onproduct,uricacid.In rabbit, mouse and human, the enzyme appears to exist predominantly cytoplasmic as ahomodimer,withcataly<cdomainsfortheZn+2-dependenthydroly<cdeamina<onofguaninetoxanthineplusammonia.GenomicdetailsfortheGDAgenearemappedforseveralspecies,andexpression profiling in certain <ssues and organisms has been ini<ated, although thecomplementofvarioustranscriptvariantsisincomplete.
• Structure:c.50kDasubunitswithsequencesthatvaryatinternalandterminalsites,duetoexonselec<on;atleast4significantformspredic<ngproteinsofvariouslengthsareknown.
• Interac<ons:tubulin,snapin,andthepost-synap<cdomainprotein95(PSD-95).• PSD-95binding is through thePDZbindingmo<fspresentat theC-terminusof thedimeric
structure.• Sequencevariantsoccurmostlyattheproteinbindingdomains,althoughminorvariantslack
thedeaminasecataly<csite.• Inmammalianbrain,highenzyma<clevelsareintelencephalicbrainregions;verylowlevels
inwhitemaYerandcerebellum;moderatelevelsinliverandcertainotherorgans.lowlevelsin plasma/serum, notably altered by liver dysfunc<on. The S- transcript is predominant,codingfora51kDamonomer.
• Actualroleofguaninedeaminaseinspecializedorganmetabolismandsynap<cphysiologyisuncertain, and rela<vely liYle is knownabout theenzyme characteris<csoutsideof rabbit,with significant detail available detail on gene expression paYerns, but not at the proteinlevel. Available protein expression surveys have suggested extensive post-transla<onalmodifica<ons,thathavenotyetbeendetailedwithcertainty.
• Few useful inhibitors have been described for the enzyme, and the biochemicalpharmacologyisconfused,inpartduetopoorlycharacterizedenzymeproper<es.
PresentedatASMS2019WP509
Characteriza.onoftheAc.vityandKine.csofGuanineDeaminaseJus<nM.Godinho,BenLibert,andBarryE.Boyes
AdvancedMaterialsTechnologyInc.,Wilmington,DE
Introduc.on
Purpose
EfficientSepara.onsofHighlyPolarPurineMetabolites StandardMicroplateEnzymeAssayandLC/MSCondi.ons ComparisonofKine.csBetweenBrainandLiverandRecombinantGDA
INNOVATION YOU CAN TRUST – PERFORMANCE YOU CAN RELY ON fused-core.com
• Previousenzymeassaysareproblema<c,usingeitherspectrophotometry(spectralshicofGtoX,orammonium capture), or coupled fluorometric enzyme assay of X (via Xanthine Oxidase producedH2O2),withaYendantproblemsinkine<cparameteres<ma<on.
• Available assays have poorly characterized analy<cal specificity, reproducibility and sensi<vity,genera<ngbroadreportedrangesforbasickine<cparameters(Kmvaluesandspecificac<vi<esvarywidely),andinhibitorcharacteris<cs.
Tomeasure<ssueenzyme levels, followpurifica<onprocesses,defineenzymekine<csandeffectsofinhibitors,highlyspecificandsensi<vemethodswouldbeuseful.Directgenera<onofproductxanthineusesfastsepara<onofhighlypolarpurinemetabolitesbyanewreversed-phasematerial,HALO®AQ-C18. High sensi<vity and selec<ve detec<on uses selected reac<on monitoring (SRM) LC-MS/MS.Analy<calbenefitwasassessedacrossa rangeof substrateconcentra<ons, in thepresenceofknownand poten<al compe<<ve inhibitors. The features of the assay are explored for the only two knownsubstrates for guanine deaminase, Guanine and 8-Azaguanine.We have also ini<ated comparison ofna<veGDAfrombovinebrainandliver<ssue,torecombinantbacterialexpressedhumanGDA.
0.25 0.50 0.75 1.00 1.25 1.50 1.75 min 0.00 0.25 0.50 0.75 1.00 1.25 1.50 1.75 uV (x100,000)
0.00 0.25 0.50 0.75 1.00 1.25 1.50 1.75 min 0.0 0.5 1.0 1.5 2.0 2.5
uV (x100,000)
0.00 0.25 0.50 0.75 1.00 1.25 1.50 1.75 min 0.00 0.25 0.50 0.75 1.00 1.25 1.50
uV (x100,000)
1,2
3 5
6 7 8
4
9
1,2
3 5
6 7 84
9
1,2 3 5
6 7 8
4 9
PeakIden<<es1:5-Amino-imidazole-4-carboxamide(AICA)2:Azepinomycin3:Guanine-Substrate4:2,6-Diaminopurine(2,6-DAP)5:UricAcid6:Xanthine-Product7:8-Azaxanthine8:8-Azaguanine9:Allopurinol
LCCondi<onsColumns:2.1x100mmMobilePhase:0.1%FormicAcidFlowRate:0.5mL/minTemperature:35°CDetec<on:UV254nmInjec<on:1µLInstrument:ShimadzuNexera
HALO90Å,AQ-C18,2.7µm
Compe<tor(B)2.7µmSPPAqueousC18
Compe<tor(A)2.6µmSPPAqueousC18
Aselec<onofC18phasesstablein100%Aqueouscondi<onswerescreened.TheHALO90Å,AQ-C18,2.7µmcolumnwaschosenforitsabilitytoresolvethehighlypolarpurinemetabolitesandinhibitorsofinterest.Underthecurrentusecondi<ons,theAQ-C18columnisstableformanythousandsofsamples,including<ssuehomogenates.
PanelsA-Drepresentstepstakenforselec<vedetec<onoftheenzymereac<onproduct,xanthineand8-azaxanthine.PanelAshowsachromatogramofa10pmolinjec<onofXorAzXusingtheextractedionsfrom150to152m/z.PanelBshowsthespectrawithxanthineandazaxanthineprecursorions[M−H]−at151m/z(X)or152(AzX)chosenforfragmenta<on.PanelCshowstheextractedionchromatogramscenteredatdominantfragmentions,andPanelDshowstheproductions108m/zmonitoredforXand109m/zforAzX.
XanthineTheore<calMass[m/z]152.033Damonoisotopic,[151.025m/z(-)]
8-azaxanthineTheore<calMass[m/z]153.029Damonoisotopic,[152.021m/z(-)]
ITMS[50-200m/z]
0.875 1.000 1.125 1.250 0
50000
100000
150000
200000
260000 10pmolXanthineinj.XIC(-)
150-152m/z
PrecursorIon151m/z ITMS[106.00-110.00m/z]
0.875 1.000 1.125 1.250 0
5000
10000
15000
20000
26000 XIC(-)107.8-108.2m/z
ProductIon108m/z
55.0 75.0 100.0 125.0 150.0
0
20
40
60
80
100
120 108.01
m/z
%
120.0 140.0 160.0 180.0 0.0
5.0
10.0
15.0
20.0
25.0 28.0
151.01
m/z
%
0.875 1.000 1.125 1.250
0
10000
20000
30000
40000 ITMS[106.00-110.00m/z]
XIC(-)108.8-109.2m/z
ProductIon109m/z
55.0 75.0 100.0 125.0 150.0
0
20
40
60
80
100
120 109.08
124.01
m/z
%
0.875 1.000 1.125 1.250
0
50000
100000
150000
200000 ITMS[50-200m/z]
XIC(-)10pmol8-Azaxanthineinj151-153m/z
PrecursorIon152m/z
120.0 140.0 160.0 180.0 0.0
5.0
10.0
15.0
20.0
25.0 28.0
151.99
m/z
%
A B C D
SRMMS^2151.00@cid30
SRMMS^2152.00@cid30
Selec.veDetec.onofEnzymeReac.onProductsUsingSRMintheIonTrap
ShimadzuNexeraHALO®AQ-C182.1x75mm2.7µm90Å,orHTPassayusing2.1x30mm.A=0.1%FormicAcid B=Acetonitrile0.5ml/min,35°C,265nm,MTP,autosampler25°CGradient:0%B0-1.5minto70%[email protected]%[email protected](min):1.7;divertfirst0.8minfromMSsourceITMS(-)0.7to1.7minHESISourceType;Capillary350°C;Heater325°C;SheathGas40;AuxGasFlow10ITMSFullAGC50K;ITMSSIMAGC100K;ITMSMSnAGC50KSourceVoltage(-kV)2.70;IonTrapFullMaxIonTime(ms)200;Segment(SEG)Informa<on/ScanEventDetails:ITMS-clowinjrf=70.0·(151.00000)->oS(98.00-118.00)MS/MS:ATCIDCE30.0%Q0.350Time10.000IsoW1.5,CV=0.0VAssayCondi<ons50mMBicinepH7.8,5µg/mLBSA,varyingGuanineor8-azaGuanine.Reac<onat25°Cfor>15minutes,typicalvolumeof100µLStopReac<onbyaddi<onof0.1volumeof10%FormicAcid.Allpurinereagentstocksaremaintainedin15mMNaOH/0.15mMMgCl2un<ldilu<on.Enzymeincuba<onsareconductedinstandard96wellreac<onplates,whicharedirectlyloadedintheNexeraAutosampler,forLC/MSanalysisofreac<onproducts.
Enzymeac<vityislinearover4ordersofdilu<on,inthisexample,anenzymeconcentra<onof1in12,000(50mU/rx),Xproduc<onislinearwithincuba<on<meto300minutes,consuminglessthan10%ofsubstrateat100uMG.Observedfor10uLinjec<on;LOQforX<1fmolandlinearityto>10pmolinjected.
0
10
20
30
40
50
0 100 200 300
pmolofX
Produ
ced
Time(min)
0
10
20
30
40
50
0 500 1000
pmolofX
Produ
ced
EnzymeDilu<on(x10-6)
Conclusions
Deconvolu<onresultedinasubunitMWof50866.Thisis193Dafromthepredictedmassusinggenomicdata,51059,fortheSisoformoftheenzyme.ChromatogramsandSDSPAGEshowhighenzymepurity.Column:HALO1000ÅDiphenyl,2.7µm,2.1x150mmQExac<veHFandNexeraX2A:0.1%DFAB:0.1%DFA1:1nPropanol:ACN0.25mL/min30%B0-20min
0 50 1000
2
4
6
Saturation Kinetics Recombinant Human
[G] mM
Enz
yme
Act
ivity
(mm
ol/m
in/m
g)
0 20 40 600.00
0.05
0.10
0.15
0.20
Saturation Kinetics Bovine Liver
[G] mM
Enz
yme
Act
ivity
(pm
ol/m
in)
0 20 40 600.0
0.5
1.0
1.5
2.0
Saturation Kinetics Bovine Brain
[G] mM
Enz
yme
Act
ivity
(mm
ol/m
in/m
g)
Kmvaluesforbovinebrainandliveraresimilar,0.286±0.035(SEM)µMand0.31±0.03(SEM)µMrespec<vely,andaremuchlowerthanliteraturevalues.Thisemphasizestheimportanceofhighlyspecificandsensi<veLC/MSenzymeassays.TheKmvaluesforbovine<ssuesaremuchlowerthantheassayedrecombinanthumanenzyme,Kmof6.2±0.2(SEM)µM.Vmaxofbovinebraincomparesfavorablywithliteraturereportsat1.36±0.04(SEM)µmole/min/mgbutdifferssignificantlyfromtherecombinatnt,5.2±0.2(SEM)µmole/min/mg.AzGisamuchpoorersubstrate.
Asensi<veandspecificLC/MSapproachtostudyenzymekine<csisdemonstrated.Resultsdifferfromliteraturereportsthatusespectrophotometricmethodshighligh<ngtheimportanceoftheassay’sspecificity.TheHALO®AQ-C18phaseprovidesrobustsepara<onin100%aqueouscondi<onsforthesehighlypolarpurines.Highpurityenzymefrom<ssueexhibitedmuchlowerKmthanbacterialexpressedrecombinatenzymeandasubunitmassthatdivergesfromDNAderivedvalue.ThisworkwassupportedinpartbyNIGMS[GM116224toBEB].ThecontentissolelytheresponsibilityoftheauthorsanddoesnotnecessarilyrepresenttheofficialviewsoftheNa<onalIns<tuteofHealth
LC/MSofGDA
[G]µM
[G]µM [G]µM
µM µM







