dormancy, germination and the cell … germination and the cell cycle in developing and imbibing...
TRANSCRIPT

DORMANCY, GERMINATION AND THE CELL CYCLE INDEVELOPING AND IMBIBING TOMATO SEEDS1
RENATO D. DE CASTRO2 AND HENK W.M. HILHORST3
2Universidade Federal de Lavras - Lavras - MG, 3Wageningen University - Wageningen - The Netherlands.
ABSTRACT - Seed development and germination are characterized by an arrest and a resumption ofgrowth, respectively. Evidently, the cell cycle plays an important role in these events. Tomato(Lycopersicon esculentum) is one of the best studied dicot species with respect to the role of the cell cyclein seed development, germination and dormancy. Developmental arrest after the completion of histo-differentiation and morphogenesis is paralelled by an arrest of the cell cycle in the G1 phase anddissapearance of the microtubular cytoskeleton. Resumption of growth occurs during germination withDNA synthesis and replication, accumulation of tubulins, reassembly of microtubular networks and,finally, mitosis. In dormant seeds cell cycle activity is suppressed whereas in non-dormant abscisic-acid-deficient sitiens (sitw) mutant cell cycle activities are reactivated still during seed development, leading tosubsequent viviparous germination.
ADDITIONAL INDEX TERMS: Bromodeoxyuridine, cell cycle, DNA, dormancy, embryogenesis,germination, Lycopersicon esculentum, microtubules, seeds, tubulin.
DORMÊNCIA, GERMINAÇÃO E O CICLO CELULARNO DESENVOLVIMENTO E EMBEBIÇÃO DE SEMENTES DE TOMATE
RESUMO - O desenvolvimento e a germinação de sementes são respectivamente caracterizados pelainativação e reativação do crescimento. Evidentemente, o ciclo celular tem função relevante no contextodestes eventos. O tomate é uma das espécies de dicotiledôneas mais bem estudadas no que diz respeito aopapel do ciclo celular no desenvolvimento, germinação e dormência de sementes. A interrupção dodesenvolvimento ao completar a histo-diferenciação e morfogênese é acompanhada do bloqueio dociclo celular na fase G1 e completo desaparecimento do citoesqueleto microtubular durante a fase dematuração. O crescimento é complementado durante a germinação, através do reinicio da síntese e replicaçãode DNA, acúmulo de tubulinas, restabelecimento do citoesqueleto, levando a ocorrência de mitoses e divisõescelulares. Em sementes dormentes, as atividades do ciclo celular são suprimidas, enquanto que nas sementessem dormência do mutante sitiens (sitw), deficiente em ácido abscísico, o ciclo celular é reativado aindadurante o desenvolvimento da semente, resultando na subseqüente germinação vivípara.
TERMOS ADICIONAIS PARA INDEXAÇÃO: Bromodeoxiuridina, ciclo celular, DNA, dormência,embriogênese, germinação, Lycopersicon esculentum, microtubulos, sementes, tubulina.
1. Based on an invited lecture presented at the VII Brazilian Plant Physiology Congress, Brasília, July, 1999, in the CoordinatedSession on Seed Physiology
2. Corresponding author. Universidade Federal de Lavras, Departamento de Agricultura, Laboratório de Sementes, Caixa Postal37, Lavras-MG, 37200-000, Brasil. (present address). Fax: +35 3829-1301; E-mail: [email protected]
3. Wageningen University, Laboratory of Plant Physiology, Arboretumlaan 4, 6703 BD, Wageningen, The Netherlands.(permanent address)
Footnotes: BrdU – bromodeoxyuridine, DAP – days after pollination

106 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
INTRODUCTION
Seeds are the primary dispersal units ofhigher plants containing the complete genetic makeup of the species. They are complex biologicalstructures, which, in millions of years, haveadapted to diverse and often austere environments.Seeds are generally able to withstand drought andextreme temperatures and may remain viable forprolonged periods of time, which can extend tohundreds of years. These features make seeds themain contributor to the preservation of the plantworld’s genetic diversity, and are a fascinating systemfor biological research. Furthermore, seeds are ofeminent importance as a source of food and nutritionfor more than two thirds of the world population.
The seed consists of nutrient reservestorage tissue(s), an embryo and encapsulatingstructures which aim protection and that may alsoregulate germination. Seeds are unique, as theirformation requires the involvement of twoconsecutive phases in the plant’s life cycle (thesporophyte and the gametophyte), from whichtissues with differing ploidy levels develop, i.e.,the embryo and the endosperm. They may retainalso the unique characteristic of withstandingdesiccation while retaining the capacity forimmediate metabolic reactivation uponrehydration. Likewise, seeds may possessmechanisms to sense the environment, thusensuring germination under favourable conditions.
Seeds are objects of many different kindsof research. Over the past decades a number ofplant species have developed into models to studydifferent aspects of seed biology. Cereal grainshave long been used to unravel the pathways andregulation of endosperm mobilization by thealeurone layer (Fincher, 1989; Jones and Jacobsen,1991). Seeds of pea (Pisum sativum) have beenused extensively for the study of seed developmentand assimilate partitioning (Wang and Hedley,1991). Arabidopsis thaliana seeds are used forgenetic and molecular studies, employing largecollections of mutants (Feldman et al., 1994). In
several cereals, such as maize (Zea mays) andwheat (Triticum aestivum), extensive studies havebeen undertaken to improve seed quality, both forstand improvement and nutritional value. However,the tomato seed has been used most extensively tostudy the physiology and biochemistry of seeddevelopment, germination and dormancy (Hilhorstet al., 1998).
The morphological and physiologicalprocesses that occur during seed development andgermination have been studied and describedextensively (Figure 1A and C). However,information about the regulatory mechanismscontrolling these processes has begun to emergeonly after the introduction of genetic andmolecular-biological technology to this field(reviewed by Bewley and Black, 1994; Galau etal., 1991; Goldberg et al., 1994; Harada, 1997;Hilhorst, 1995; Kermode, 1995; Koornneef andKarssen, 1994; Raghavan, 1997). Analysis ofchanges in gene expression patterns that occurduring seed development and postgerminativegrowth (Figure 1B) has provided clues about theregulatory programs governing both periods (Chlanand Dure, 1983; Dure et al., 1981; Dure, 1985).
These modern molecular-geneticapproaches have often been based on studies usingseeds and have immensely contributed to ourunderstanding of plant developmental biology.However, seeds have been used in these studiesmostly because they are convenient objects ofstudy. Yet, the present knowledge is by far notenough to explain and understand the functioningand the behavior of seeds.
There is no doubt that ourunderstanding of the processes involved in seeddevelopment and germination has expanded overthe last decades. Notwithstanding theseadvances, very little is known about dormancy,as well as many other subjects of primaryimportance in seed science. As discussed byKarssen (1993), a great deal of integrated,interdisciplinary study is still required for seedscience sensu strictu, in order to betterunderstand seed function and behavior.
Dormancy, Germination and the ... 107
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000

108 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
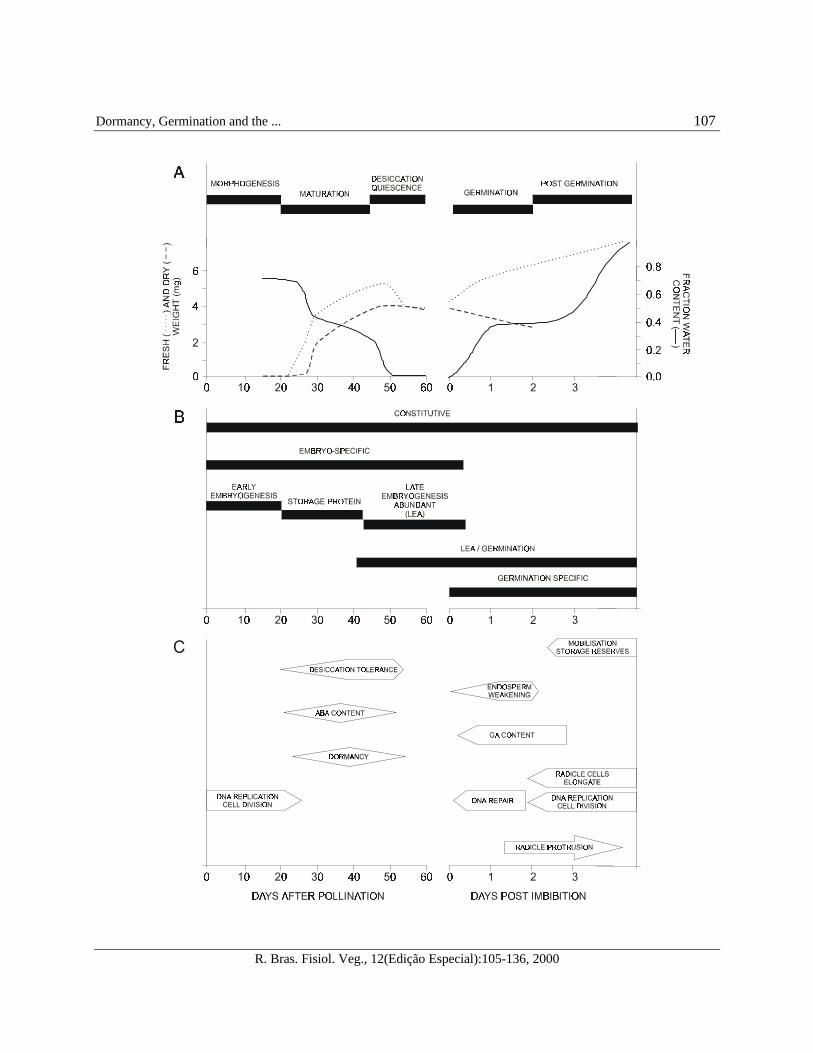
FIGURE 1 - Seed development and germination.
(A) Physiological parameters divide seed development into distinct phases. Time course of changes in fresh and dryweights and in fraction water content (g H2O/g dry weight) of developing and germinating embryos, and post-germinative seedlings are shown. Bars indicate stages of seed development and germination. Data are from oilseedrape embryos and seedlings, taken from Comai and Harada (1990), reviewed by Harada (1997).(B) Gene expression patterns divide seed development into specific stages. In a conceptual representation of theaccumulation of seven mRNA sets that are present during seed development, germination and post-germinativegrowth, bars indicate the period during which the indicated mRNA set is detected. For points of reference, mRNAaccumulation patterns are fit to the time scale in (a). Adapted from Dure (1985) and Goldberg et al. (1989).(C) Time course of major events associated with seed development, germination and subsequent post-germinativegrowth. For points of reference, the physiological events are fit to the time scale in (a) and (b). Adapted fromBewley (1997) and Hilhorst et al. (1998).
Here we attempt to give an integratedoverview and functional analysis of cell cycleevents, such as DNA synthesis and configurationsof the microtubular cytoskeleton during seeddevelopment, imbibition and germination, usingthe tomato seed as a model system. Specialemphasis is given to the progression and arrestof embryo growth through cell division andexpansion, in relation to dormancy andquiescence.
Phases of Seed Development and Germination
Seed development usually is divided intwo conceptually distinct phases (Figure 1A). Thefirst phase is considered a period of morphogenesisduring which the embryo’s body plan is establishedthrough intensive cell divisions and the embryonicorgans and tissues are formed (reviewed byGoldberg et al., 1994; Meinke, 1995; West andHarada, 1993). The second phase is considered asubsequent period of seed maturation, whichincludes the arrest of tissue and organ formation,the accumulation of nutrient reserves, changes inembryo size and in fresh and dry weights, thesuppression of precocious germination, theacquisition of desiccation tolerance, dehydrationand quiescence, and, in many species, theinduction of dormancy (Koornneef and Karssen,1994). Germination and postgerminative growth(Figure 1A) represent the phases during whichmetabolic and morphogenetic reactivation of thequiescent seed occurs (reviewed by Bewley, 1997;Harada et al., 1988, Harada, 1997).
Developmental Arrest
It has been argued that developmentalarrest is an imposed condition during seedmaturation that is not required for the formationof a viable embryo (Walbot, 1978). Contrary tohigher plants, seeds from lower plants, forinstance, do not undergo developmental arrestand metabolic quiescence. The implication isthat seed plants have incorporated processesrelated to seed maturation, dormancy andgermination into the continuous mode ofmorphogenetic developmental characteristics ofmany lower plants. The regulatory programscontrolling the arrest of growth and metabolismduring development, which leads to quiescenceand in some cases dormancy, and thereactivation of growth and metabolism duringgermination are, therefore, of relevance. Bothphases will certainly involve the arrest andreactivation of cell cycle related events.
THE CELL CYCLE
Growth depends on cell division andelongation. The sequence of processes that occurduring cell division is referred to as the cell cycle,which is dependent on DNA synthesis andreplication, and which leads to specific patterns oforganogenesis and morphogenesis, in respect tocellular differentiation (Figure 2). At the ‘deeper’,cytological and molecular levels, the cell cycleinvolves a chromosome cycle in which DNAsynthesis towards replication occurs duringinterphase, and a mitotic cycle which leads to celldivision (Figure 3). The patterning of the cell

Dormancy, Germination and the ... 109
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
FIGURE 2 - The relation between DNA replication, cell division and organogenesis in plants.An autopoietic system (Varela et al., 1975) showing the sequential outcome of processes (horizontal arrows) inrelation to the complexity of the system (upward-directed arrows). Adapted from Barlow (1993).
division cycle resolves into recursive patterns ofconfigurations of the cytoskeleton components, asthe ”sub cellular machinery” required for celldivision and growth.
Microtubules are one main component ofthe cytoskeleton. They are unbranched cylinders ofabout 25 nm in external diameter, with an opencentral channel of about 15 nm, and are assembledfrom heterodimers containing one α-tubulin andone β-tubulin polypeptide, each with a molecularweight around 50 to 55 kD conprising 450 aminoacids (Figure 4). Microtubules are related to thecell cycle through distinct reorganizational orconfigurational arrays, i.e., the interphase corticalarrays, the preprophase bands, spindles andphragmoplast mitotic arrays (Derksen et al., 1990;Goddard et al., 1994) (Figure 3).
Synthesis and replication of DNA occursduring interphase and may last several hours.Therefore, interphase is a phase which, within thecell cycle, lasts longer than mitosis. Withininterphase there is a phase of quiescence (G0)coupled to a phase of growth and pre-synthesis(G1) during which the nuclei of diploid cellscontain an arbitrary 2C DNA value, referring tothe amount of DNA per nucleus, and duringwhich DNA repair may occur. Subsequently,synthesis towards replication of DNA occursduring the synthetic or S-phase. Finally, the G2
phase, comprising nuclei with 4C DNA values,resolves interphase in preparation for mitosis.The mitotic phase comprises several distinctstages based on the microtubular arrays, i.e.,(pre) prophase, metaphase, anaphase andtelophase (Figure 3).
Level inHierarchy Process Outcome
Whole PlantRoot and Shoot Systems
Organ-meristem
Molecule
Morphogenesis
Organogenesis
Cell Cell division
DNA replication
Biomass,productivity
Cell production
Cell cycle
Chromosome cycle

110 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
FIGURE 3 - Diagram of microtubular cytoskeleton arrays in relation to cell cyle phases.DNA replication (2C to 4C DNA) occurs in a period that lasts several hours, known as “S-phase” of the cell cycle.Mitosis (“M-phase”) commences after a “gap” (“G2-phase”). The “G1-phase” follows mitosis, to complete thecycle. Changes in the microtubular cytoskeleton system are geared to these events. Cortical arrays are found duringG1- and S-phases. The preprophase band begins to form at the end of the S-phase and is fully condensed just beforethe nuclear envelope breaks at the end of prophase of mitosis. It is then supplanted by the mitotic microtubularcytoskeleton arrays of the spindle, which in turn, give way to the phragmoplast array. Finally cytokinesis occurs andcortical arrays are reinstated. Adapted from Gunning and Steer (1996).
Both DNA synthesis and themicrotubular organization have been extensivelystudied in seeds, taking into consideration subjectssuch as cellular events in embryogenesis andendosperm formation during seed development,and in organogenesis during postgerminativegrowth (reviewed by Bershadsky and Vasiliev,
1988; Barlow, 1993; Clayton, 1985; Francis andHerbert, 1993; Raghavan, 1997; XuHan, 1995).Yet, a great deal of knowledge is still required inorder to understand the relationships among cellcycle events in seeds, and their contribution to seedfunctioning during development and germination,including dormancy.

Dormancy, Germination and the ... 111
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
DORMANCY
Dormancy is defined as the inability ofseeds to germinate under favourable environmentalconditions. The mechanisms of dormancy are stillto be elucidated and, therefore, the classification ofthe different types of dormancy is entirely based onits expression under various conditions (see Eiraand Caldas, this volume). For example, dormancyinduced by elevated temperatures is usuallyindicated as thermodormancy. However, it is notknown whether this type of dormancy differsfrom other types, e.g. photodormancy, in amechanistical sense. Thus, it was proposed todistinguish only two types of dormancy: primaryand secondary dormancy (Karssen, 1982; Amen,1986; Hilhorst, 1995). This classification isbased on timing. Primary dormancy is associated
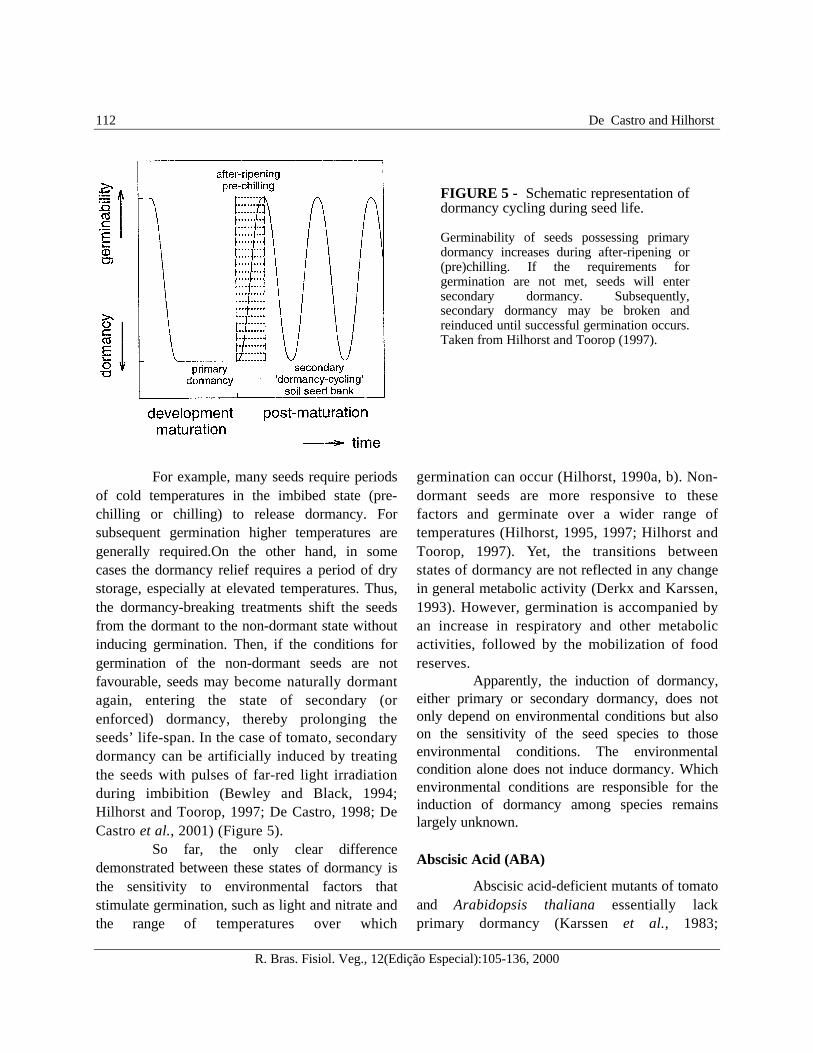
with seed development, whereas secondarydormancy occurs essentially after dispersal andis basic to seasonal dormancy cycling in soilseed banks (Figure 5).
The basic characteristic of dormancy issimply the absence of germination. Unfortunately,this is as yet the only way to quantify dormancy. Anon-germinating seed may be non-dormant ordormant. A non-dormant seed will germinate underthe right set of conditions, whereas a dormant seedwill usually display much greater restrictions in theconditions under which it will germinate. Duringmaturation seeds may enter a state of true primarydormancy, which may or may not be sustainedafter maturity, and which requires another set ofconditions to break their dormancy. Therequirements for dormancy relief may be differentfrom those for germination.
FIGURE 4 - Schematic model of the microtubule.The microtubule is a hollow cylinder whose wall is composed of 13 protofilaments. Each protofilament isconstructed from alternating α-tubulin and ß-tubulin molecules. Modified from Kleinsmith and Kish (1995).
112 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
For example, many seeds require periodsof cold temperatures in the imbibed state (pre-chilling or chilling) to release dormancy. Forsubsequent germination higher temperatures aregenerally required.On the other hand, in somecases the dormancy relief requires a period of drystorage, especially at elevated temperatures. Thus,the dormancy-breaking treatments shift the seedsfrom the dormant to the non-dormant state withoutinducing germination. Then, if the conditions forgermination of the non-dormant seeds are notfavourable, seeds may become naturally dormantagain, entering the state of secondary (orenforced) dormancy, thereby prolonging theseeds’ life-span. In the case of tomato, secondarydormancy can be artificially induced by treatingthe seeds with pulses of far-red light irradiationduring imbibition (Bewley and Black, 1994;Hilhorst and Toorop, 1997; De Castro, 1998; DeCastro et al., 2001) (Figure 5).
So far, the only clear differencedemonstrated between these states of dormancy isthe sensitivity to environmental factors thatstimulate germination, such as light and nitrate andthe range of temperatures over which
germination can occur (Hilhorst, 1990a, b). Non-dormant seeds are more responsive to thesefactors and germinate over a wider range oftemperatures (Hilhorst, 1995, 1997; Hilhorst andToorop, 1997). Yet, the transitions betweenstates of dormancy are not reflected in any changein general metabolic activity (Derkx and Karssen,1993). However, germination is accompanied byan increase in respiratory and other metabolicactivities, followed by the mobilization of foodreserves.
Apparently, the induction of dormancy,either primary or secondary dormancy, does notonly depend on environmental conditions but alsoon the sensitivity of the seed species to thoseenvironmental conditions. The environmentalcondition alone does not induce dormancy. Whichenvironmental conditions are responsible for theinduction of dormancy among species remainslargely unknown.
Abscisic Acid (ABA)
Abscisic acid-deficient mutants of tomatoand Arabidopsis thaliana essentially lackprimary dormancy (Karssen et al., 1983;
FIGURE 5 - Schematic representation ofdormancy cycling during seed life.
Germinability of seeds possessing primarydormancy increases during after-ripening or(pre)chilling. If the requirements forgermination are not met, seeds will entersecondary dormancy. Subsequently,secondary dormancy may be broken andreinduced until successful germination occurs.Taken from Hilhorst and Toorop (1997).

Dormancy, Germination and the ... 113
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
Koornneef et al., 1985). This has led to theconclusion that the transient rise in ABA contentduring mid-development is responsible for theinduction of primary dormancy. However,evidence for this is at best circumstantial(Hilhorst, 1995). First, there is often a lack ofcorrelation between ABA content and depth ofdormancy. For instance, the tomato cultivarCaruso has more than 10 times higher levels ofABA than the cultivar Moneymaker, yet thelatter shows a much deeper dormancy (Berry andBewley, 1992, Hilhorst, 1995). Secondly, it hasbeen shown that complementation of the ABAmutation in Arabidopsis thaliana by theadmission of ABA reversed all mutant traits tothe wild type phenotype, except for thedormancy (Koornneef et al., 1989). Thirdly, inArabidopsis thaliana, mutants have been isolatedthat lack dormancy but possesses wild type ABAlevels (Léon-Kloosterziel et al., 1996). Apparently,the sensitivity to the hormone plays an importantrole. ABA – insensitive mutants of Arabidopsisthaliana, containing wild type ABA contents, alsolack primary dormancy (Koornneef et al., 1984).Vivipary or pre-harvest sprouting in wheat hasbeen demonstrated to be parallel to a reducedsensitivity to ABA (Walker-Simmons, 1987).During the second half of seed developmentsensitivity to ABA decreased in seeds ofmuskmelon (Welbaum et al., 1990) and alfafa (Xuet al., 1990; Xu and Bewley, 1991). It is likely thatthis is also the case in tomato, but data are notavailable. Only when the sensitivity to ABAamong seed batches is similar, can ABA content becorrelated with germination, as was shown fortomato and Arabidopsis thaliana seeds (Hilhorst,1995). Apparently, the very low remainingamounts of ABA in the mature seeds inhibited ordelayed germination. This would also explainwhy either dry storage or cold imbibition couldbreak dormancy. It has been shown in tomato(Groot and Karssen, 1992) and lettuce (Dulson et
al., 1988) that both treatments, may lead to adecrease in endogenous ABA and an increase ingermination.
THE TOMATO SEED
Tomato (Lycopersicon esculentum Mill.)is one of the most important annual vegetables inthe world. Many new varieties are created eachyear by intensive breeding efforts to meet differentrequirements all over the world. Common tomatoplants are diploid and reproduced by seeds. Newlybred varieties are almost all hybrids, normallyproduced by artificial crossing. Therefore, high-quality seeds are required to match these high-costhybrids to ensure their authentic value. Moreprofound knowledge of the physiological andbiochemical changes and their interactions duringdevelopment, pre-treatments and germination willcertainly contribute to the improvement of seedquality.
Compared with seed of Arabidopsisthaliana, a model plant species for genetic andmolecular research, the seed of tomato isconsidered a convenient research object because ofits relatively simple structure and large size. Themature seed consists of a full-grown embryoembedded in a thick-walled endosperm and issurrounded by a thin seed coat (Smith, 1935; Vargaand Bruinsma, 1986). Its size, in the order of 2-5mm, allows for easy manipulation and dissection.Therefore, tomato seeds have been used mostextensively as a “model system” to studyphysiology and biochemistry, including thefunctional analysis of cell cycle events duringdevelopment, pre-treatments, germination anddormancy (Bino et al., 1992; De Castro, 1998; DeCastro et al., 1995, 1998, 2000, 2001; Groot et al.,1987; Hilhorst et al., 1998; Liu et al., 1994, 1996,1997).
Tomato seeds develop and mature in themoist environment of a developing fruit. Thedevelopment of the fruit is governed by the

114 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
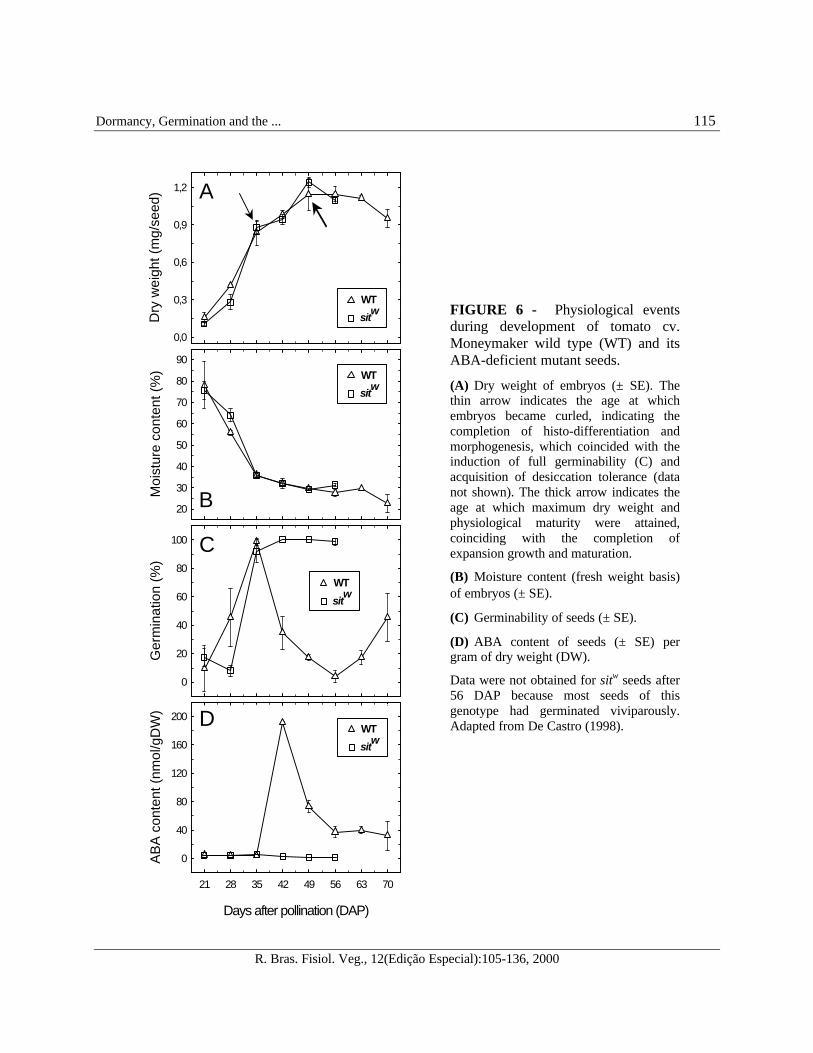
developing seeds. The number of seeds and fruitgrowth are positively correlated as the final fruitsize and weight are largely determined by the totalnumber of seeds (Varga and Bruinsma, 1986).Studies of the hormonal control of seeddevelopment have shown that the fruit fresh weightof ABA-deficient tomato mutants was less thanhalf of the control wild type fruits during thesecond half of development. However, there wereno significant differences between seedwater/moisture content and dry weight (Groot etal., 1987; Liu et al., 1996, De Castro, 1998). Thus,tomato seed development, in terms of dry mass orwater content, appears to be independent from theaction of ABA (Figure 6A and B).
The tomato seed attains maximum dryweight and its final curled shape between 35 and50 days after pollination (DAP). Fruits are ripeand red after approximately 60 DAP. Severalcrucial events take place simultaneously duringthe period of 35-50 DAP, i.e., seeds become fullygerminable, desiccation tolerance is induced andwater content decreases to approximately 50%(fresh weight basis) (as seen in Figure 6).Concurrently, the endosperm solidifies,abscission of the funiculus occurs, and the testaturns brown (Berry and Bewley, 1991; Liu et al.,1996; De Castro, 1998). Primary dormancy maybe induced in wild type seeds once fullgeminability is attained whereas sitw seeds donot become dormant. This correlates with theABA content in seeds of both genotypes duringdevelopment (Figure 6C and D).
DORMANCY AND THE CELL CYCLEDURING TOMATO SEED DEVELOPMENT
The functional analysis of cell cycleevents in developing tomato seeds (cv.Moneymaker) shows that the developmentalarrest of the embryo is characterized not only bycessation of DNA synthesis but also by a
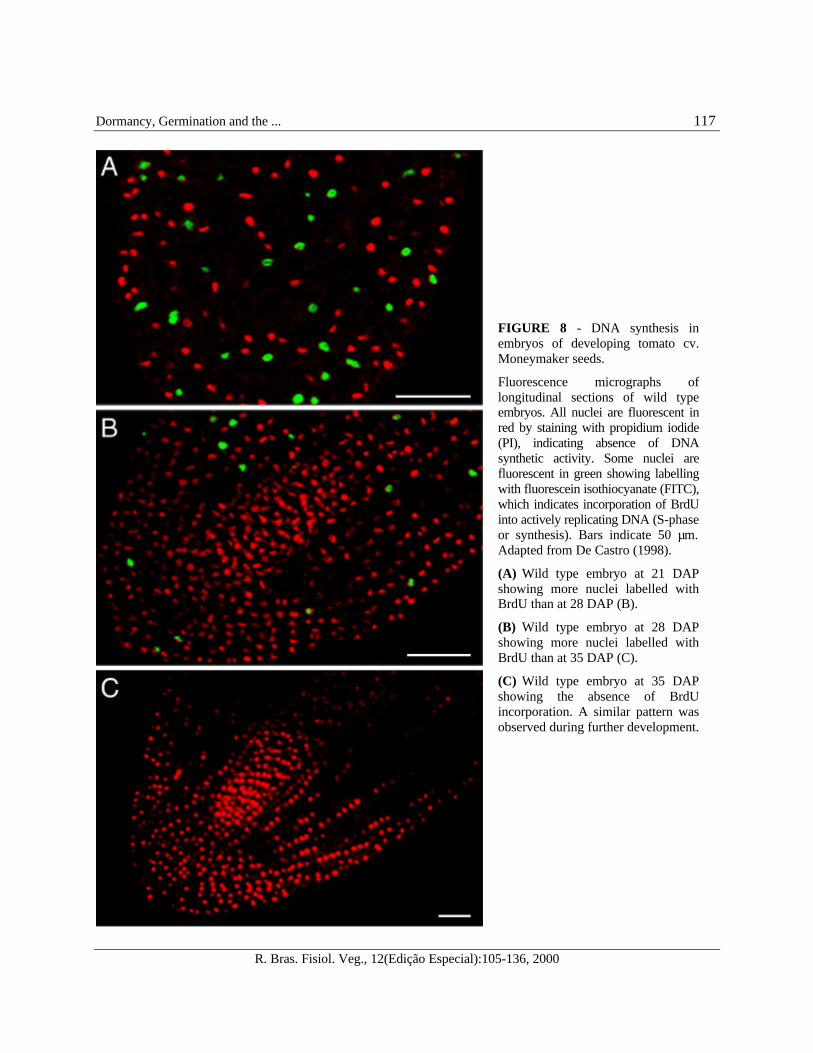
complete breakdown of the microtubularcytoskeleton (De Castro, 1998). DNA contentdata obtained by means of flow cytometry showthat the number of embryonic 4C DNA nuclei isrelatively high during early stages of embryodevelopment in wild type and sitw seeds, i.e., at 21DAP. This is an indication of high cell cycleactivity, but which appears to decrease as thenumber of 4C DNA nuclei decreases and becomesconstant at low levels from 35 DAP onwards (Figure7), with the majority of the cells containing 2C DNAnuclei. This pattern has been confirmed by a decreasein the number of embryonic nuclei labelled withbromodeoxyuridine (BrdU), which is a labelingreagent used for the detection of DNA synthesis andcell proliferation analysis (Gratzner, 1982).Bromodeoxyuridine labelled nuclear DNA isobserved in developing tomato embryos only until35 DAP, showing that DNA synthesis is arrestedat 35 DAP prior to maturation (Figure 8).
The initial high number of 4C DNAnuclei is paralleled by high amounts of ß-tubulindetected between 21 and 35 DAP (Figure 9, lanes 5to 7), which corresponds to an elaboratemicrotubular cytoskeleton network composed ofmitotic and cortical microtubular arrays throughoutthe embryonic tissue, indicating histo-differentiation (Figure 10A to C). Thereafter,mitotic events and DNA synthesis are not detectedanymore in wild type embryos, indicating that thecompletion of embryo histo-differentiation andmorphogenesis occurs between 28 and 35 DAP.
The decrease in β-tubulin content in wildtype embryos from 35 DAP onwards (Figure 9,lanes 7 to 12) is the result of the degradation of thecortical microtubular cytoskeleton network (Figure10 C to F). Apparently, cortical microtubules arerequired during this period of the maturation phase
in order to provide the embryo with thestructural machinery for further expansion growthafter histo-differentiation and morphogenesis havebeen completed. This occurs concomitantly withthe increase in dry weight during this same periodDAP (Figure 6A).
Dormancy, Germination and the ... 115
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
21 28 35 42 49 56 63 70
Dry
wei
ght (
mg/
seed
)
0,0
0,3
0,6
0,9
1,2
WT
sitw
21 28 35 42 49 56 63 70
Moi
stur
e co
nten
t (%
)
20
30
40
50
60
70
80
90WT
sitw
Days after pollination (DAP)
21 28 35 42 49 56 63 70
AB
A c
onte
nt (
nmol
/gD
W)
0
40
80
120
160
200WT
sitw
21 28 35 42 49 56 63 70
Ger
min
atio
n (%
)
0
20
40
60
80
100
WT
sitw
A
C
D
B
FIGURE 6 - Physiological eventsduring development of tomato cv.Moneymaker wild type (WT) and itsABA-deficient mutant seeds.
(A) Dry weight of embryos (± SE). Thethin arrow indicates the age at whichembryos became curled, indicating thecompletion of histo-differentiation andmorphogenesis, which coincided with theinduction of full germinability (C) andacquisition of desiccation tolerance (datanot shown). The thick arrow indicates theage at which maximum dry weight andphysiological maturity were attained,coinciding with the completion ofexpansion growth and maturation.
(B) Moisture content (fresh weight basis)of embryos (± SE).
(C) Germinability of seeds (± SE).
(D) ABA content of seeds (± SE) pergram of dry weight (DW).
Data were not obtained for sitw seeds after56 DAP because most seeds of thisgenotype had germinated viviparously.Adapted from De Castro (1998).

116 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
The depolymerization of microtubulesafter 35 DAP is a gradual process, first apparent inthe cotyledons, to proceed in time towards theshoot meristem, hypocotyl and radicle regions ofthe mature embryo (De Castro, 1998). Theappearance of fluorescent granules between 42 and49 DAP (Figure 10D and E) indicate clusters ofdepolymerised tubulin prior to its completedegradation (Figure 10F), and coincided with theattainment of minimal moisture content, maximumdry weight and achievement of physiologicalmaturity (Figure 6A, B).
Apparently, the cessation ofdevelopmental growth starts in the embryo`scotyledonary and shoot apical region, precedingthat of the hypocotyl and radicle basal region (DeCastro, 1998). Once physiological maturity hasbeen achieved, expansion growth is arrested
leading to a cessation of the structuralrearrangements (Clayton, 1985). Therefore, fromthis stage onwards the microtubular cytoskeleton isno longer required (Figure 10F) and cell cycleactivities are apparently fully arrested, indicating aphase of quiescence, despite further changes ingerminability.
The physiologically mature tomato seedshave a water potential between -0.50 MPa and -1.00MPa. Viviparous germination is prevented by thelocular tissue, which has a water potential ofapproximately -1.00 MPa. Remarkably, the waterpotential of the embryo is much lower, approximately-4.00 MPa. Thus, it is clear that endosperm and testaplay a decisive role in preventing viviparousgermination and maintaining seeds in a quiescentstate as mentioned above.
21 DAP 28 DAP 63 DAP35 DAP A
Days after pollination (DAP)
21 28 35 42 49 56 63
4C D
NA
con
tent
(%
)
0
5
10
15
20
sitw
WT B
21 DAP 28 DAP 35 DAP 63 DAP A
FIGURE 7 - Total 4C DNA contents inembryos of developing tomato cv.Moneymaker seeds.
(A) Flow cytometry histogram profiles of wildtype (WT) diploid embryo nuclei showing therelative proportions of unreplicated 2C DNAnuclei and replicated 4C DNA nuclei.
(B) Number of nuclei with 4C DNA content(± SE), expressed as percentage of the totalnumber of WT and sitw nuclei (2C + 4C), asdetermined by flow cytometry (A).
A minimum of 10.000 gated nuclei countswere considered for analysis of all samples.This represented a minimum of 70% of thetotal embryonic nuclei counts. Adapted fromDe Castro (1998).
B
Dormancy, Germination and the ... 117
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
FIGURE 8 - DNA synthesis inembryos of developing tomato cv.Moneymaker seeds.
Fluorescence micrographs oflongitudinal sections of wild typeembryos. All nuclei are fluorescent inred by staining with propidium iodide(PI), indicating absence of DNAsynthetic activity. Some nuclei arefluorescent in green showing labellingwith fluorescein isothiocyanate (FITC),which indicates incorporation of BrdUinto actively replicating DNA (S-phaseor synthesis). Bars indicate 50 µm.Adapted from De Castro (1998).
(A) Wild type embryo at 21 DAPshowing more nuclei labelled withBrdU than at 28 DAP (B).
(B) Wild type embryo at 28 DAPshowing more nuclei labelled withBrdU than at 35 DAP (C).
(C) Wild type embryo at 35 DAPshowing the absence of BrdUincorporation. A similar pattern wasobserved during further development.
118 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
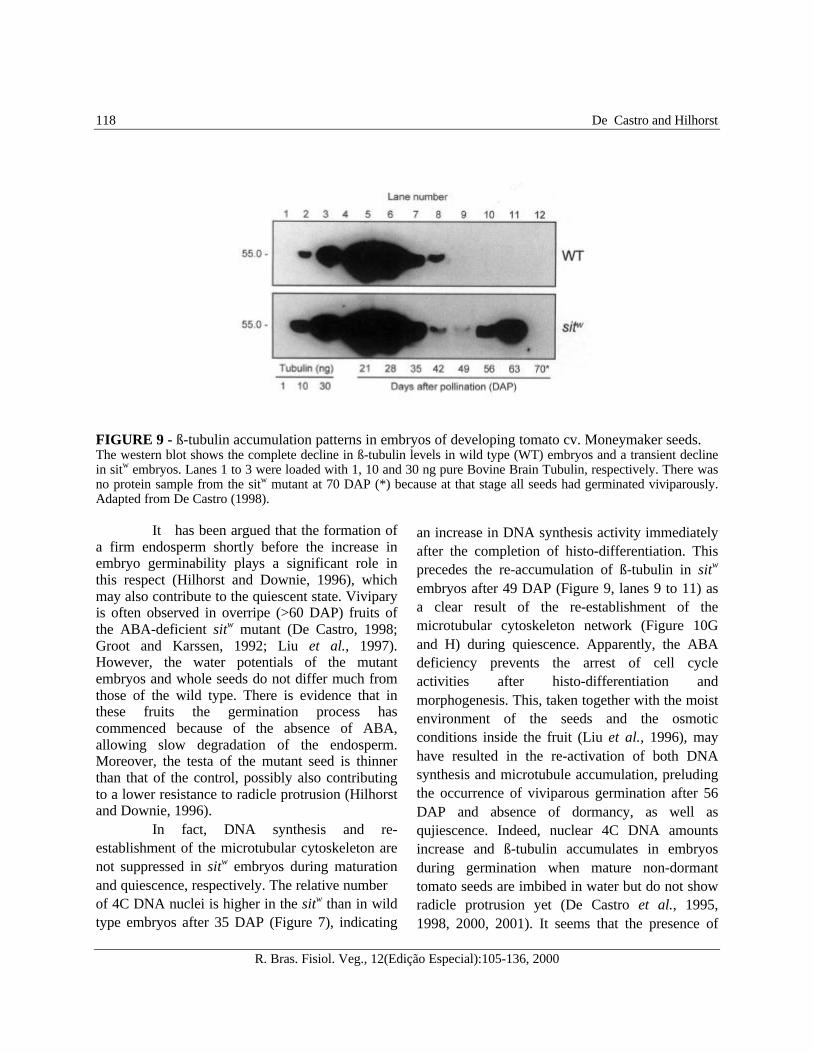
FIGURE 9 - ß-tubulin accumulation patterns in embryos of developing tomato cv. Moneymaker seeds.The western blot shows the complete decline in ß-tubulin levels in wild type (WT) embryos and a transient declinein sitw embryos. Lanes 1 to 3 were loaded with 1, 10 and 30 ng pure Bovine Brain Tubulin, respectively. There wasno protein sample from the sitw mutant at 70 DAP (*) because at that stage all seeds had germinated viviparously.Adapted from De Castro (1998).
It has been argued that the formation ofa firm endosperm shortly before the increase inembryo germinability plays a significant role inthis respect (Hilhorst and Downie, 1996), whichmay also contribute to the quiescent state. Viviparyis often observed in overripe (>60 DAP) fruits ofthe ABA-deficient sitw mutant (De Castro, 1998;Groot and Karssen, 1992; Liu et al., 1997).However, the water potentials of the mutantembryos and whole seeds do not differ much fromthose of the wild type. There is evidence that inthese fruits the germination process hascommenced because of the absence of ABA,allowing slow degradation of the endosperm.Moreover, the testa of the mutant seed is thinnerthan that of the control, possibly also contributingto a lower resistance to radicle protrusion (Hilhorstand Downie, 1996).
In fact, DNA synthesis and re-establishment of the microtubular cytoskeleton arenot suppressed in sitw embryos during maturationand quiescence, respectively. The relative numberof 4C DNA nuclei is higher in the sitw than in wildtype embryos after 35 DAP (Figure 7), indicating
an increase in DNA synthesis activity immediatelyafter the completion of histo-differentiation. Thisprecedes the re-accumulation of ß-tubulin in sitw
embryos after 49 DAP (Figure 9, lanes 9 to 11) asa clear result of the re-establishment of themicrotubular cytoskeleton network (Figure 10Gand H) during quiescence. Apparently, the ABAdeficiency prevents the arrest of cell cycleactivities after histo-differentiation andmorphogenesis. This, taken together with the moistenvironment of the seeds and the osmoticconditions inside the fruit (Liu et al., 1996), mayhave resulted in the re-activation of both DNAsynthesis and microtubule accumulation, preludingthe occurrence of viviparous germination after 56DAP and absence of dormancy, as well asqujiescence. Indeed, nuclear 4C DNA amountsincrease and ß-tubulin accumulates in embryosduring germination when mature non-dormanttomato seeds are imbibed in water but do not showradicle protrusion yet (De Castro et al., 1995,1998, 2000, 2001). It seems that the presence of
Dormancy, Germination and the ... 119
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
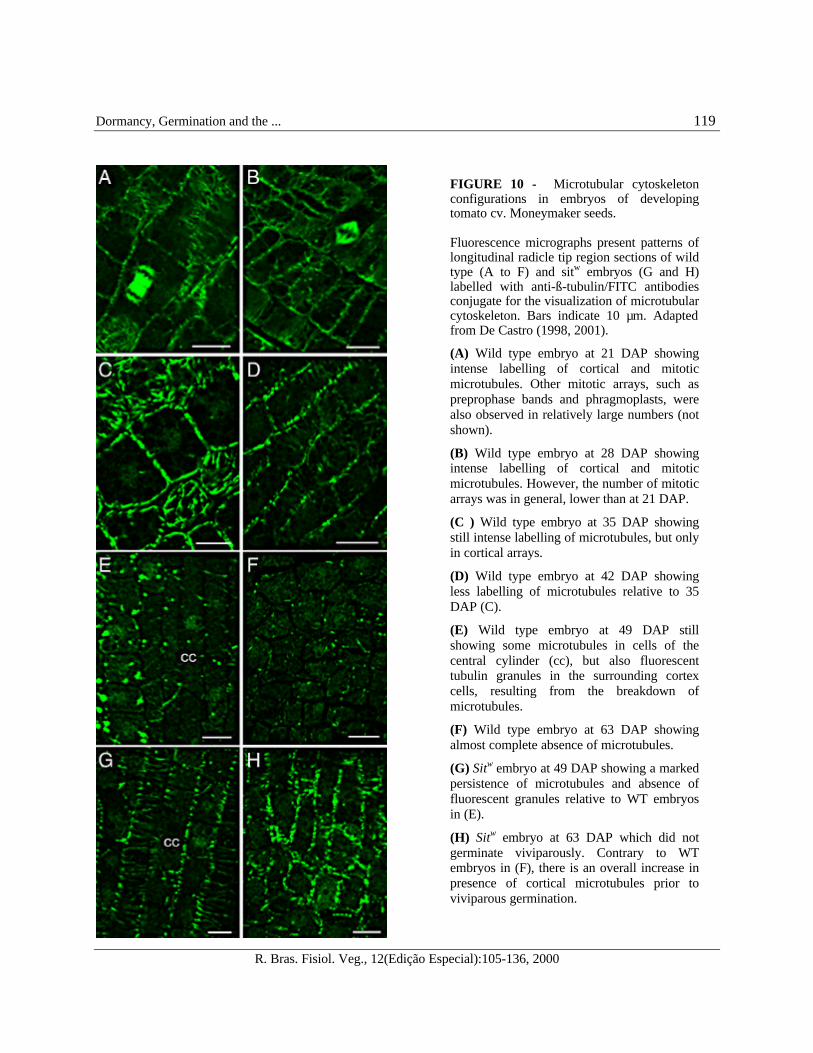
FIGURE 10 - Microtubular cytoskeletonconfigurations in embryos of developingtomato cv. Moneymaker seeds.
Fluorescence micrographs present patterns oflongitudinal radicle tip region sections of wildtype (A to F) and sitw embryos (G and H)labelled with anti-ß-tubulin/FITC antibodiesconjugate for the visualization of microtubularcytoskeleton. Bars indicate 10 µm. Adaptedfrom De Castro (1998, 2001).
(A) Wild type embryo at 21 DAP showingintense labelling of cortical and mitoticmicrotubules. Other mitotic arrays, such aspreprophase bands and phragmoplasts, werealso observed in relatively large numbers (notshown).
(B) Wild type embryo at 28 DAP showingintense labelling of cortical and mitoticmicrotubules. However, the number of mitoticarrays was in general, lower than at 21 DAP.
(C ) Wild type embryo at 35 DAP showingstill intense labelling of microtubules, but onlyin cortical arrays.
(D) Wild type embryo at 42 DAP showingless labelling of microtubules relative to 35DAP (C).
(E) Wild type embryo at 49 DAP stillshowing some microtubules in cells of thecentral cylinder (cc), but also fluorescenttubulin granules in the surrounding cortexcells, resulting from the breakdown ofmicrotubules.
(F) Wild type embryo at 63 DAP showingalmost complete absence of microtubules.
(G) Sitw embryo at 49 DAP showing a markedpersistence of microtubules and absence offluorescent granules relative to WT embryosin (E).
(H) Sitw embryo at 63 DAP which did notgerminate viviparously. Contrary to WTembryos in (F), there is an overall increase inpresence of cortical microtubules prior toviviparous germination.

120 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
ABA is essential for the suppression of DNAsynthesis activity and subsequent re-establishmentof the microtubular cytoskeleton during thetransition to the maturation and quiescent phases.Consequently, sitw seeds do not acquire dormancy(Figure 6C). The absence of the ABA peak in sitw
seeds at 42 DAP correlates with the increase in thenumber of nuclei with replicated 4C DNAamounts, preluding viviparous germination(Figures 6D and 7). It is tempting to speculate thatABA action is required to block resumption DNAsynthesis. In this way seeds may become dormant.However, this also depends on specificenvironmental conditions.
Some wild type and sitw seeds are able togerminate during embryo histo-differentiation, butare unable to produce viable seedlings. At thisstage, torpedo shaped embryos appear not to be yetfully differentiated and still undergo intense cellcycle activity, at low dry weight and high moisturecontent. Only after the completion of histo-differentiation and accumulation of reserves, i.e.,from 35 DAP onwards, germination results innormal seedlings (De Castro, 1998). Apparently,the switch from a developmental mode to agerminative mode does not require the completionof histo-differentiation. Berry and Bewley (1991)concluded that the inability of tomato seeds toproduce viable seedlings at these early stages ofdevelopment is due to the lack of sufficientreserves in the still liquid endosperm. Theyobserved that young embryos isolated from theseseeds grew into viable seedlings when placed on anutrient medium.
DORMANCY AND THE CELL CYCLEDURING TOMATO SEED GERMINATION
Embryos of dry mature non-dormantwild type tomato seeds contain low levels of
nuclear 4C DNA and ß-tubulin (Bino et al., 1992,1993; De Castro et al., 1995, 1998, 2000; Liu etal., 1994, 1997) (Figure 12 and Figure 14, lane 5).This correlates with the absence of BrdUincorporation into embryonic nuclear DNA fromdry control seeds, showing the lack of DNAsynthesis activity (Figure 13A), whereas theabsence of a microtubular cytoskeleton network(Figure 15A) reflects the absence of ß-tubulin(Figure 14, lane 5). Seeds in a dry state have mostof their metabolic activities suppressed (Vertucci,1989), which may contribute to maintaining thecell cycle of most embryo cells in the G1 phase, asobserved during the process of seed maturation anddehydration.
The reactivation of cell cycle activitiesand growth is observed during re-hydration of thenon-dormant seeds. Non-dormant tomato seedsinitiate germination, i.e. radicle protrusion, after 48h of imbibition (Figure 11). DNA synthesis, asdetected through BrdU incorporation, initiatesaround 12 h of imbibition in embryo radicles(Figure 13B and C), concomitantly with the initialaccumulation of β-tubulin (Figure 14, lane 6) andassembly of cortical microtubules (Figure 15B andC). This indicates that initial DNA synthesis isparalleled by the initiation of embryo growththrough cell expansion, prior to the occurrence ofmitotic events. However, the amount of tubulinincreases further (Figure 14, lanes 7 to 9) andmitosis can be observed in tomato embryos alreadyat 24 h of imbibition (Figure 15C), prior to radicleprotrusion and completion of germination, which isinitiated after 48 h of imbibition (Figure 11). It isconcluded that the completion of germination intomato seeds occurs normally by means of cellexpansion as well as cell division (De Castro et al.,2000). Although embryonic DNA replication intomato can be blocked by hydroxyurea withoutaffecting the accumulation of ß-tubulin and radicleprotrusion, subsequent seedling development does
Dormancy, Germination and the ... 121
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
Incubation time (d)
0 1 2 3 4 5 6 7 8 9 10
4C D
NA
con
tent
(%
)
0
10
20
30
40
50
Ch
1D2D
sitsitwND
Incubation time (d)
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Ger
min
ated
see
ds (
%)
0
10
20
30
40
50
60
70
80
90
100
+
+
++ + + + +
FR
Ch
NDsitw+1D+Ch1D+L1D+GA2D+Ch2D+L2D+GA
Max SD
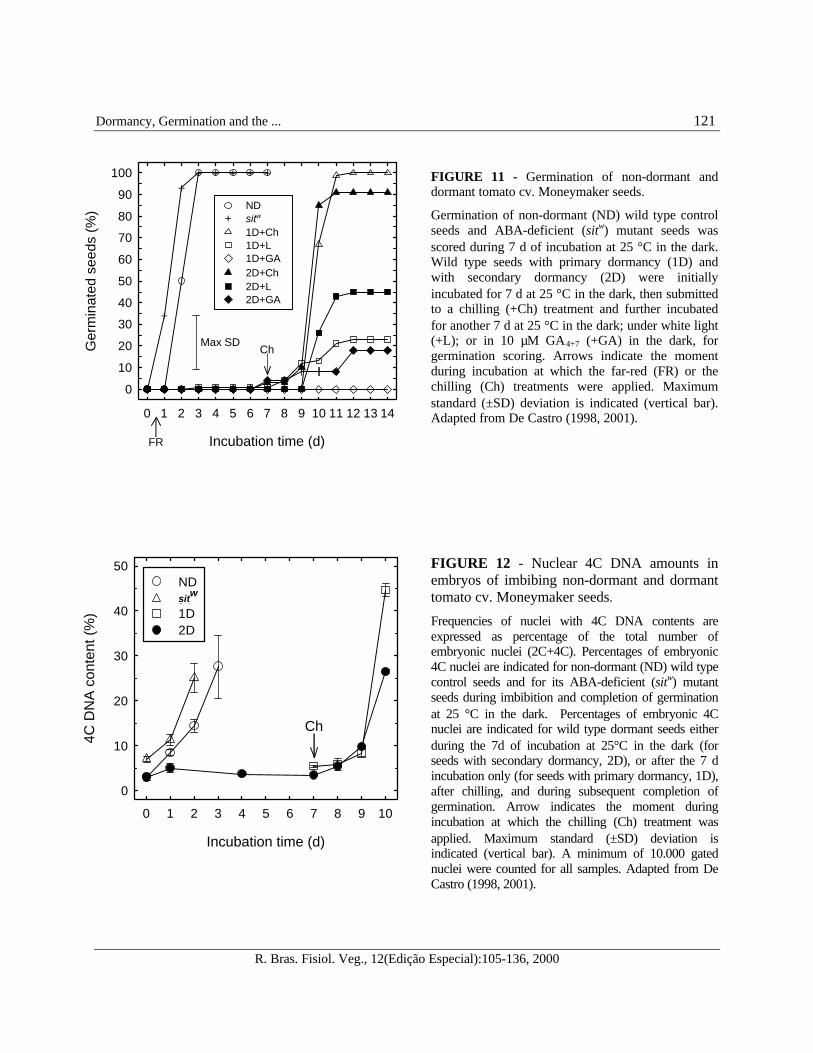
FIGURE 11 - Germination of non-dormant anddormant tomato cv. Moneymaker seeds.
Germination of non-dormant (ND) wild type controlseeds and ABA-deficient (sitw) mutant seeds wasscored during 7 d of incubation at 25 °C in the dark.Wild type seeds with primary dormancy (1D) andwith secondary dormancy (2D) were initiallyincubated for 7 d at 25 °C in the dark, then submittedto a chilling (+Ch) treatment and further incubatedfor another 7 d at 25 °C in the dark; under white light(+L); or in 10 µM GA4+7 (+GA) in the dark, forgermination scoring. Arrows indicate the momentduring incubation at which the far-red (FR) or thechilling (Ch) treatments were applied. Maximumstandard (±SD) deviation is indicated (vertical bar).Adapted from De Castro (1998, 2001).
FIGURE 12 - Nuclear 4C DNA amounts inembryos of imbibing non-dormant and dormanttomato cv. Moneymaker seeds.
Frequencies of nuclei with 4C DNA contents areexpressed as percentage of the total number ofembryonic nuclei (2C+4C). Percentages of embryonic4C nuclei are indicated for non-dormant (ND) wild typecontrol seeds and for its ABA-deficient (sitw) mutantseeds during imbibition and completion of germinationat 25 °C in the dark. Percentages of embryonic 4Cnuclei are indicated for wild type dormant seeds eitherduring the 7d of incubation at 25°C in the dark (forseeds with secondary dormancy, 2D), or after the 7 dincubation only (for seeds with primary dormancy, 1D),after chilling, and during subsequent completion ofgermination. Arrow indicates the moment duringincubation at which the chilling (Ch) treatment wasapplied. Maximum standard (±SD) deviation isindicated (vertical bar). A minimum of 10.000 gatednuclei were counted for all samples. Adapted from DeCastro (1998, 2001).

122 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
not proceed (Liu and Groot, personalcommunication). Similar observations were madein cabbage seeds (Górnik et al., 1997). Thisimplies that cell division is not a prerequisite forradicle protrusion in tomato. However, the retardedcompletion of germination in the presence ofhydroxyurea suggests that, simultaneously withcell expansion, mitotic divisions are required fornormal tomato seed germination and seedlinggrowth (De Castro et al., 2000). So far, celldivision was considered to occur in tomatoembryos only after completion of germination(Coolbear and Grierson, 1979; Haigh, 1988).However, this was based on quantitative analysisof nucleic acids only. Evidently, the substantiallymore sensitive immunocytological approachconfirms that this does not proceed and thatnormal radicle protrusion in tomato does requirecell division (De Castro et al., 2000). Apparently,the mechanisms of germination is dependent oneach species. Radicle protrusion in cucumber(Cucumis sativus L.) seeds does occur through cellexpansion only, whereas cell division is onlyobserved after radicle protrusion, at 21 h ofimbibition (Jing et al., 1999).
Tomato seeds with primary dormancy donot show any detectable cell cycle activity in theembryos when incubated in the dark, not even afterprolonged incubation of over one month (DeCastro et al,. 2001), i.e. the number of 4C DNAnuclei does not increase and DNA synthesis is notdetected through BrdU incorporation, neither doestubulin and microtubules accumulate (Figures 12to 15). Similar results were obtained with seedsfrom dormant lines of Avena fatua, in which only aslow replacement of DNA was observed whichwas related to DNA replair (Elder and Osborne,1993). Gibberellins (GAs) and white light appearsonly slightly effective, at best, in inducinggermination (Figure 11). This insensitivity is alsoreflected in an overall absence of cell cycleactivities. A small amount of β-tubulin can bedetected seeds with primary as well as in seeds
with secondary dormancy when incubated inpresence of GAs or white light (Figure 14, lanes 14and 15, 21 and 22), but there is no significantlevels of microtubules assembly, neither any DNAsynthetic activity (De Castro et al,. 2001).However, chilling does increase the sensitivity tolight and GAs, resulting in almost full germinationafter transferring the seeds back to 25°C (Figure 11).This supports the hypothesis that dormancy can bedefined by a lack of sensitivity to germinationstimulants (Hilhorst and Karssen, 1992; Derkx andKarssen, 1993, Derkx et al., 1994). It is presumedthat cell cycle activities may have also beeninduced as the levels of β-tubulin increases (Figure14, lane 11) and cortical microtubular cytoskeleton(Figure 15E) after the chilling treatment, but priorto transfer to 25 °C. However, it has not beenobserved a significant increase in the number of 4CDNA nuclei (Figure 12),nor any detectable nuclearDNA synthesis activity through BrdUincorporation. The initial accumulation of tubulinsand subsequent formation of microtubularcytoskeleton is a prerequisite for the initiation ofcell cycle events in embryonic tissue during seedimbibition and germination (De Castro et al., 1995,1998, 2000, 2001; Jing, et al., 1999), and rootgrowth (Gunning and Sammut, 1990; Baluška andBarlow, 1993). This suggests that the initialformation of the microtubular cytoskeleton isassociated with the dormancy-breaking processsensu stricto. Transferring the seeds to 25 °C afterchilling results in a sequential increase in theamount of β-tubulin (Figure 14, lanes 12 and 13),number of microtubules (Figure 15E and F), andinitiation of DNA synthesis which is resumed uponfull DNA replication prior to mitosis (Figure 12and Figure 13E, F), evidently associated with thegermination process (De Castro et al., 1995, 1998,2000; Jing, et al., 1999).The situation in seeds inwhich secondary dormancy is artificially inducedby treating the seeds with pulses of far-red lightirradiation during imbibition appears to be
Dormancy, Germination and the ... 123
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000

124 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
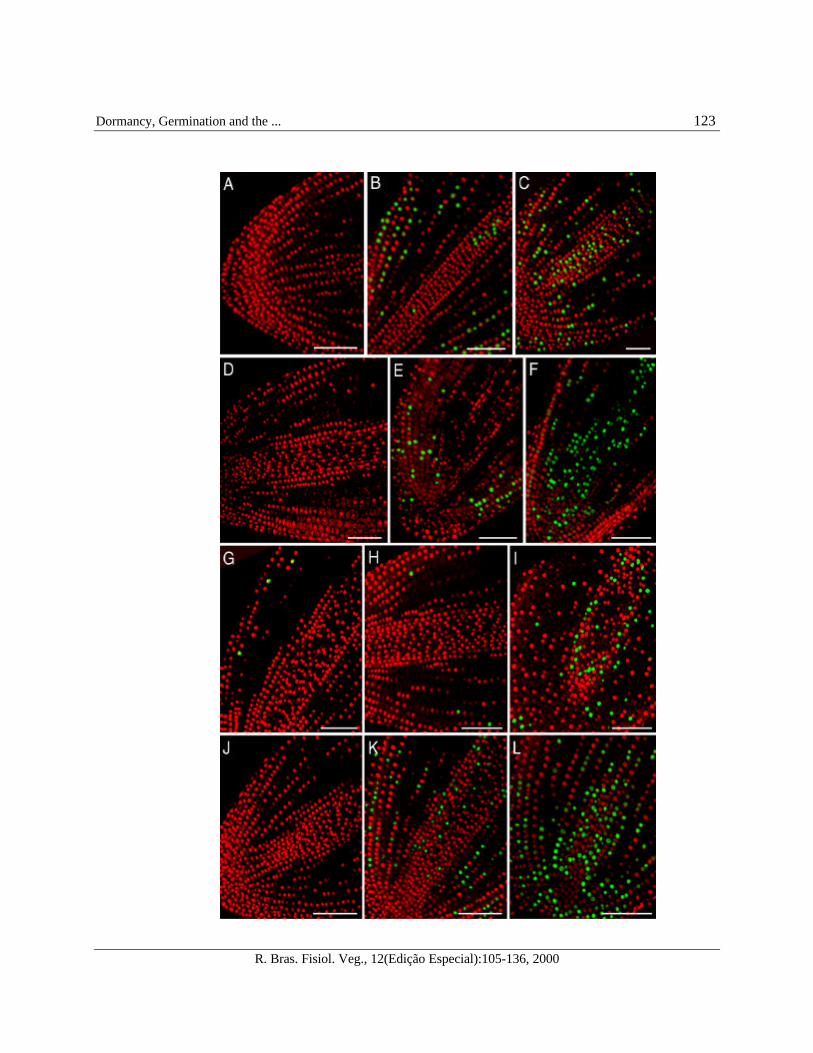
FIGURE 13 - Activation of DNA synthesis in embryos of imbibing tomato cv. Moneymaker seeds.
Fluorescence micrographs of longitudinal sections of the radicle tip region of embryos from non-dormant wild typecontrol and its ABA-deficient sitw mutant seeds, and from wild type seeds with primary and secondary dormancy.Most nuclei show red fluorescence as a result of staining with propidium iodide (PI), indicating absence of DNAsynthesis activity. Nuclei which show green fluorescence are labelled with anti-BrdU/FITC antibodies conjugate,indicative of BrdU incorporation into actively replicating DNA (S-phase). Bars indicate 100 µm. Adapted from DeCastro (1998, 2001).
(A-C) Embryos from non-dormant WT control seeds: (A) embryo of dry seeds showing absence of BrdUincorporation, which indicates absence of nuclear DNA synthesis; (B) embryo showing incorporation of BrdU andactivation of nuclear DNA synthesis after 12 h of seed imbibition; (C) embryo showing a larger number of nucleiwith actively replicating DNA after 24 h of imbibition.
(D-F) Embryos from WT seeds with primary dormancy: (D) embryo from imbibed primarily dormant seedsshowing absence of BrdU incorporation and DNA synthesis after 1 month plus 7 d of incubation at 25 °C in dark;(E) embryo showing activation of DNA synthesis immediately after release of primary dormancy by a chillingtreatment; (F) embryo of ungerminated seeds, 1 d after the chilling treatment, showing a larger number of nucleilabelled with active replicating nuclear DNA.
(G-I) Embryos from WT seeds with secondary dormancy: (G) embryos upon induction of secondary dormancy by afar-red irradiation showing some nuclei labelled with BrdU, indicating the initiation of DNA synthesis during theperiod of far-red irradiation and induction of dormancy; (H) detection of DNA synthesis immediately after thechilling treatment in levels comparable to that after the far-red treatment and subsequent 7 d of incubation at 25 ºCin the dark (not shown); (I) higher levels of DNA synthesis 1d after the chilling treatment indicating release ofsecondary dormancy.
(J-L) Embryos from the fully non-dormant sitw seeds: (J) dry seeds showing absence of DNA synthesis; (K) embryoof ungerminated sitw seeds after 1 d of imbibition showing activation of DNA synthesis; (L) embryo of sitw seedsthat had germinated after 1 d of imbibition showing a larger number of nuclei with actively synthesizing DNA.
different from what is observed in relation toprimary dormancy (De Castro et al., 2001).Although germination is completely prevented bythe far-red irradiation, some ß-tubulin accumulates(Figure 14, lanes 16 and 17) and results in theassembly of microtubules during the far-redirradiation treatment (Figure 15G), as after thechilling treatment in seeds with primary dormancy.But, again, it has not been detected any increase inthe number of 4C nuclei after the far-redirradiation treatment (Figure 12), indicating thatDNA replication appears to be suppressed also intomato seeds with secondary dormancy (Groot etal., 1997). However, some BrdU labelled nucleicould be detected after the far-red irradiation, asafter chilling (Figure 13G and H). As mentioned
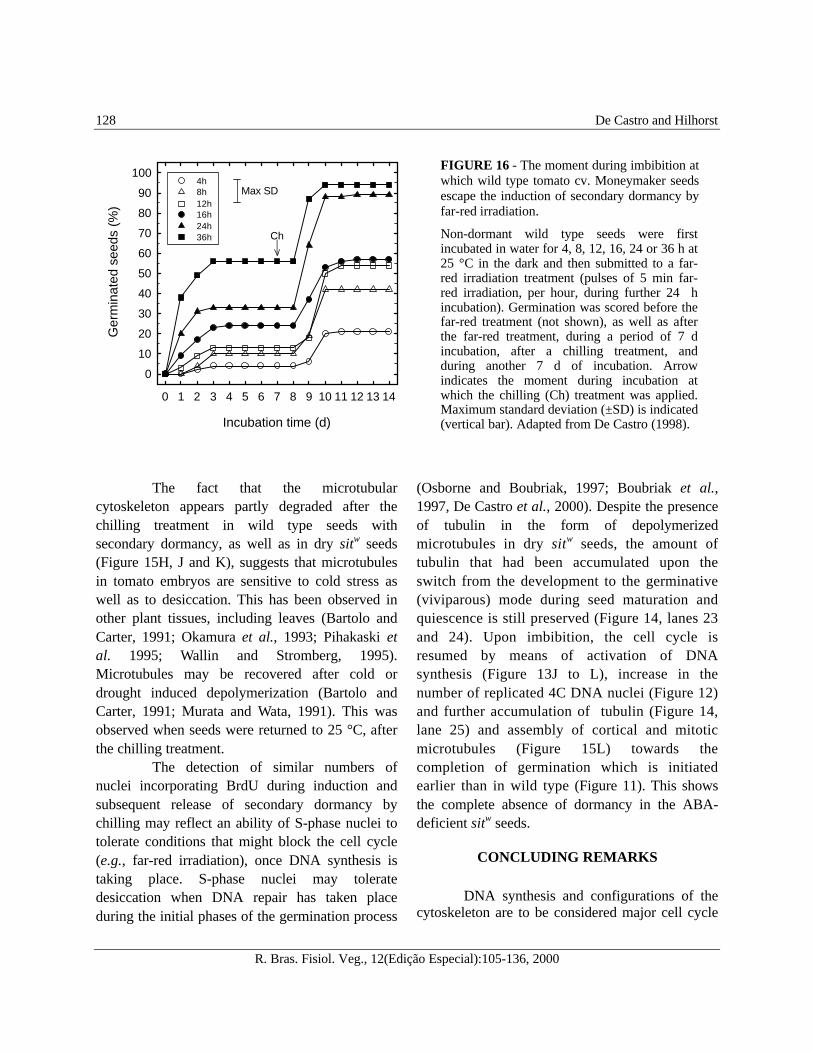
before, the initiation of DNA synthesis towards fullDNA replication occurs in the radicle tip of non-dormant seeds within the first 12 h of imbibition.This occurs concomitantly with the accumulationof β-tubulin and the establishment of corticalmicrotubular cytoskeleton. Apparently, the far-red irradiation induces secondary dormancywhen the cell cycle had been already activatedduring the first hours of seed imbibition. Thissuggests that secondary dormancy may beinduced, and block the cell cycle, while thegermination process is in progress. Indeed,results of an escape experiment with imbibingtomato seeds indicate that the far-red irradiationmay induce secondary dormancy when appliedbefore 36 h of imbibition (Figure 16) (De Castro,

Dormancy, Germination and the ... 125
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
1998). At that phase of the germinationprocess DNA synthesis and the formation ofmicrotubular cytoskeleton have progressed to sucha degree that mitosis has started in a large numberof cells (Figure 15C). Necrotic regions can beobserved in the root tips of abnormal seedlingsfrom those seeds which do escape the inhibition byfar-red irradiation after 36h of imbibition. Thismay be related to the initiation of mitotic events incells of these specific regions (De Castro et al.,2000). However, further investigations are neededto address this aspect. Furthermore, as shown
before in the ABA-deficient sitw mutant,germination follows the completion of seedmorphogenesis within the fruit without interveningdevelopmental arrest (Figures 6, 8 and 9) (Liu etal, 1997; De Castro, 1998). Consequently, embryosfrom dry mature sitw seeds contain amounts ofreplicated 4C DNA nuclei, tubulin andmicrotubular cytoskeleton that are comparablewith those of germinating wild type seeds at animbibition interval of 24 hours (Figure 12; Figure14, lanes 7 and 23; Figure 15B and J) (De Castro etal., 2001).
FIGURE 14 - ß-Tubulin accumulation in embryos of imbibing non-dormant and dormant tomato cv.Moneymaker seeds.
The western blot shows the accumulation of â-tubulin in embryos of non-dormant wild type (WT) and ABA-deficient (sitw) mutant seeds and in dormant wild type seeds. Lanes 1-3 were loaded with 1, 10 and 30 ng purebovine brain tubulin, respectively. Molecular weight (kDa) is indicated on the left hand side. (g) indicate embryos ofseeds that had germinated. Adapted from De Castro (1998, 2001).
(Lanes 5 to 9) ß-tubulin levels are shown for embryos from dry non-dormant wild type control seeds, and after 24 to48 h of imbibition.
(Lanes 10 to 15) ß-tubulin levels in embryos of wild type seeds with primary dormancy; (10) after an incubationperiod of 1month plus 7 d at 25 ºC in dark; (11) at the end of the chilling (Ch) treatment; (12 and 13) 24 and 48 hafter the chilling treatment, respectively; (14 and 15) seeds incubated for 7 d at 25 ºC in the dark and thensubsequently incubated for another 7 d in 10 µM GA4+7 or under white light, respectively.
(Lanes 16 to 22) ß-tubulin levels in embryos of seeds with secondary dormancy; (16) after the far-red lightirradiation treatment; (17) after a subsequent 7 d incubation at 25 ºC in dark; (18) immediately after the chillingtreatment; (19 and 20) 24 and 48 h after the chilling, respectively; (21 and 22) seeds incubated for 7 d at 25 ºC in thedark and then subsequently incubated for another 7 d in 10 µM GA4+7 or under white light, respectively.
(Lanes 23 to 25) ß-tubulin levels in embryos of dry sitw seeds, and after 24 and 48 h of imbibition, respectively.

126 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000

Dormancy, Germination and the ... 127
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
FIGURE 15 - Microtubular cytoskeleton configurations in embryos of imbibing non-dormant anddormant tomato cv. Moneymaker seeds.
Fluorescence micrographs of longitudinal sections of the radicle tip region of embryos from non-dormant wild typecontrol and sitw seeds, and from wild type seeds with primary and secondary dormancy labelled with anti-ß-tubulin/FITC antibodies conjugate. Bars indicate 25 µm. Adapted from De Castro (1998, 2001).
(A-C) Embryos from non-dormant wild type control seeds: (A) embryo of dry seeds showng the absence ofmicrotubules; (B) embryo showing the initial assembly and accumulation of cortical microtubules after 12 h ofimbibiton; (C) embryo showing larger amounts of cortical microtubular arrays and appearance of mitotic arrays after24 h of imbibition, indicating the initiation of cell divisions prior to radicle protrusion and completion ofgermination.
(D-F) Embryos from wild type seeds with primary dormancy: (D) embryo from imbibed primarily dormant seedsshowing absence of microtubules after 1 month plus 7 d of incubation at 25 °C in dark; (E) embryo showingappearance of cortical microtubules after the chilling treatment, indicating dormancy release; (F) appearance of anelaborate cortical and mitotic microtubular cytoskeleton network 1d after the chilling treatment, e.g., the arrowheadsindicate 2 mitotic phragmoplast microtubular arrays of cells in early cytokinesis, whereas the arrow indicates amitotic microtubular spindle array of a cell in metaphase.
(G-I) Embryos from wild type seeds with secondary dormancy: (G) embryos upon induction of secondary dormancyshowing appearance of cortical microtubules (arrowheads) after the far-red light irradiation treatment; (H) embryoshowing presence of cortical microtubules and fluorescent tubulin granules, apparently indicating some breakdownof microtubules after a subsequent period of 7 d incubation at 25 ºC in dark; (I) embryo showing the recovery of themicrotubular network after the chilling treatment, showing cortical and mitotic arrays after a 1 d subsequentimbibition period, indicating dormancy release.
(J-L) Embryos from non-dormant sitw seeds: (J and K) radicle and cotyledonary regions showing presence ofcortical microtubules and tubulin granules in embryos of dry sitw seeds; (L) embryo of ungerminated sitw seeds after1 d of imbibition, showing the recovery of cortical and mitotic microtubular arrays, e.g., arrowheads indicate 2mitotic phragmoplast microtubular arrays of cells in late cytokinesis.
Seeds with far-red induced secondarydormancy also do not to respond to GA but dorespond moderately to light (Figure 11). Thechilling treatment alone is effective enough toinduce almost full germination withoutrequirement for light (Figure 11). However,contrary to the seeds with primary dormancy, inthe seeds with secondary dormancy the chillingtreatment appears to induce partial degradation oftubulin (Figure 14, lanes 11 and 18) andbreakdown of microtubular cytoskeleton (Figure15H) while BrdU incorporation can still bedetected (Figure 13H). It has been shown thatduring development primary dormancy is inducedat a moment when embryo histo-diferentiation hasbeen completed and there is no DNA synthesis, i.e.after 35 DAP. However, at that stage there is stillan elaborate cortical microtubular cytoskeletonpresent, which appears to be active during the final
morphological expansion growth, i.e., between 35and 49 DAP (De Castro, 1998). Secondarydormancy can be induced under conditions ofactive DNA synthesis and the presence of themicrotubular cytoskeleton. Therefore, it isapparent that the cell cycle is blocked by the far-red irradiation treatment and is resumed after thechilling. The fact that the germination responseto GA and light by seeds with secondarydormancy is greater than in seeds with primarydormancy indicates that the difference betweenprimary and secondary dormancy in tomato islargely quantitative. Apparently, the initiation ofthe cell cycle contributes to this response. Theescape experiment shows that the longer theseeds are imbibed before far-red irradiation, theless dormant they become. This is expressed inthe effectiveness of the chilling treatment(Figure 16).
128 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
The fact that the microtubularcytoskeleton appears partly degraded after thechilling treatment in wild type seeds withsecondary dormancy, as well as in dry sitw seeds(Figure 15H, J and K), suggests that microtubulesin tomato embryos are sensitive to cold stress aswell as to desiccation. This has been observed inother plant tissues, including leaves (Bartolo andCarter, 1991; Okamura et al., 1993; Pihakaski etal. 1995; Wallin and Stromberg, 1995).Microtubules may be recovered after cold ordrought induced depolymerization (Bartolo andCarter, 1991; Murata and Wata, 1991). This wasobserved when seeds were returned to 25 °C, afterthe chilling treatment.
The detection of similar numbers ofnuclei incorporating BrdU during induction andsubsequent release of secondary dormancy bychilling may reflect an ability of S-phase nuclei totolerate conditions that might block the cell cycle(e.g., far-red irradiation), once DNA synthesis istaking place. S-phase nuclei may toleratedesiccation when DNA repair has taken placeduring the initial phases of the germination process
(Osborne and Boubriak, 1997; Boubriak et al.,1997, De Castro et al., 2000). Despite the presenceof tubulin in the form of depolymerizedmicrotubules in dry sitw seeds, the amount oftubulin that had been accumulated upon theswitch from the development to the germinative(viviparous) mode during seed maturation andquiescence is still preserved (Figure 14, lanes 23and 24). Upon imbibition, the cell cycle isresumed by means of activation of DNAsynthesis (Figure 13J to L), increase in thenumber of replicated 4C DNA nuclei (Figure 12)and further accumulation of tubulin (Figure 14,lane 25) and assembly of cortical and mitoticmicrotubules (Figure 15L) towards thecompletion of germination which is initiatedearlier than in wild type (Figure 11). This showsthe complete absence of dormancy in the ABA-deficient sitw seeds.
CONCLUDING REMARKS
DNA synthesis and configurations of thecytoskeleton are to be considered major cell cycle
Incubation time (d)
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Ger
min
ated
see
ds (
%)
0
10
20
30
40
50
60
70
80
90
1004h8h12h16h24h36h Ch
Max SD
FIGURE 16 - The moment during imbibition atwhich wild type tomato cv. Moneymaker seedsescape the induction of secondary dormancy byfar-red irradiation.
Non-dormant wild type seeds were firstincubated in water for 4, 8, 12, 16, 24 or 36 h at25 °C in the dark and then submitted to a far-red irradiation treatment (pulses of 5 min far-red irradiation, per hour, during further 24 hincubation). Germination was scored before thefar-red treatment (not shown), as well as afterthe far-red treatment, during a period of 7 dincubation, after a chilling treatment, andduring another 7 d of incubation. Arrowindicates the moment during incubation atwhich the chilling (Ch) treatment was applied.Maximum standard deviation (±SD) is indicated(vertical bar). Adapted from De Castro (1998).

Dormancy, Germination and the ... 129
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
FIGURE 17 - The cell cycle in developing and germinating tomato cv. Moneymaker seeds.
A model describing the development and germination of wild type and its ABA-deficient sitw mutant tomato cv.Moneymaker seeds in relation with activities of cell cycle events and with dormancy. Diverging lines indicateincreasing and converging lines decreasing activities. The ‘gray fill’ indicate presence of the activity, whereasthe ‘white fill’ indicate absence of the activity.

130 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
events which allow a better understanding of theinactivation of growth during seed developmentand reactivation of growth during seedgermination. It is shown during tomato seeddevelopment (Figure 17) that the arrest of DNAsynthesis and replication, coupled with the arrest ofmitotic events defines the completion of embryohisto-differentiation and morphogenesis as theinitial phase of seed development. Further embryogrowth occurs only through cell expansion, as a resultof water uptake and subsequent reserveaccumulation, and is arrested with the fulldegradation of the cortical microtubular cytoskeleton.This defines maturation, as the second phase of seeddevelopment. Thereafter, seed quiescence takes placein the absence of cell cycle events as the final phaseof seed development. Resumption of growth occursduring seed imbibition when cell cycle events arereactivated (Figure 17). New cortical microtubulesare assembled and DNA synthesis is reinitiatedtowards replication leading to mitotic events andcompletion of germination and radicle protrusion. Inthe ABA-deficient sitw mutant developmental arrestand suppression of the cell cycle are absent (Figure17), resulting in a continuum between morphogenesisand germination, which is typically expressed asvivipary. The sitw mutant is considered adevelopmental mutant in which dormancy is neverexpressed and in which there is no quiescence.Dormancy is characterized by a suppression of cellcycle events. Quatitative differences have beenobserved when comparing primary and secondarydormancy.
REFERENCES
AMEN, R.D. A model of seed dormancy.Botanical Review, 34: 1-31, 1986.
BERSHADSKY, A.D. & VASILIEV, J.M.Cytoskeleton. New York, Plenum Press, 1988.
BALUŠKA, F. & BARLOW, P.W. The role of themicrotubular cytoskeleton in determiningnuclear chromatin structure and passage ofmaize root cells through the cell cycle.European Journal of Cell Biology, 61: 160-167, 1993.
BARLOW, P.W. The cell division cycle in relationto root organogenesis. In: ORMROD, J.C. &FRANCIS, D. (Ed.) Molecular and CellularBiology of the Plant Cell Cycle. Dordrecht,Kluwer Academic Publishers, 1993. p. 179-199.
BARTOLO, M.E. & CARTER, J.V. Microtubulesin the mesophyll cells of nonacclimated andcold-acclimated spinach. Plant Physiology,97: 175-181, 1991.
BERRY, T. & BEWLEY, J. D. Seeds of tomato(Lycopersicon esculentum Mill.) whichdevelop in a fully hydrated environment in thefruit switch from a developmental to agerminative mode without a requirement fordesiccation. Planta, 186: 27-34, 1991.
BERRY, T. & BEWLEY, J.D. A role for thesurrounding fruit tissue in preventing thegermination of tomato (Lycopersiconesculentum) seeds. Plant Physiology, 100:951-957, 1992.
BEWLEY, J.D. & BLACK, M. Seeds: Physiologyof Development and Germination. NewYork, Plenum Press, 1994. 445p
BEWLEY, J.D. Seed germination and dormancy.Plant Cell, 9: 1055-1066, 1997.
BINO, R.J.; DE VRIES, J.N.; KRAAK, H.L. &PIJLEN, J.G. Flow cytometric determinationof nuclear replication stages in tomato seedsduring priming and germination. Annals ofBotany, 69: 231-236, 1992.
BINO, R.J.; LANTERI, S.; VERHOEVEN, H.A. &KRAAK, H.L. Flow cytometric determinationof nuclear replication stages in seed tissues.Annals of Botany, 72: 181-187, 1993.
BOUBRIAK, I.; KARGIOLAKI, H.; LYNE, L. &OSBORNE, D.J. The requirement for DNArepair in desiccation tolerance of germinatingembryos. Seed Science Research, 7: 97-105,1997.

Dormancy, Germination and the ... 131
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
CHLAN, C.A. & DURE, L.I. Plant seedembryogenesis as a tool for molecularbiology. Molecular Cell Biochemestry, 55:5-15, 1983.
CLAYTON, L. The cytoskeleton and the plant cellcycle. In: BRYANT, J.A. & FRANCIS, D.(Ed.) The Cell Division Cycle in Plants.Cambridge, Cambridge University Press,1985. p. 113-131.
COOLBEAR, P. & GRIERSON, D. Studies on thechanges in the major nucleic acid componentsof tomato seeds (Lycopersicon esculentumMill.) resulting from presowing treatments.Journal of Experimental Botany, 30: 1153-1162, 1979.
COMAI, L. & HARADA J.J. Transcriptionalactivities in dry seed nuclei indicate the timingof the transition from embryogeny togermination. Proceedings of the NationalAcademy of Scieces, USA. 87: 2671-2674,1990.
DE CASTRO, R.D.; ZHENG, X.;BERGERVOET, J.H.W.; DE VOS, R.C.H. &BINO, R.J. ß-tubulin accumulation and DNAreplication in imbibing tomato seeds. PlantPhysiology, 109: 499-504, 1995.
DE CASTRO, R.D. A functional analysis of cellcycle events in developing and germinatingtomato seeds. Wageningen, WageningenAgricultural University, 1998. 110p. PhDThesis.
DE CASTRO, R.D.; HILHORST, H.W.M.;BERGERVOET, J.H.W.; GROOT, S.P.C. &BINO, R.J. Detection of ß-tubulin in tomatoseeds: optimization of extraction andimmunodetection. Phytochemistry, 47: 689-694, 1998.
DE CASTRO, R.D.; VAN LAMMEREN, A.A.M.;GROOT, S.P.C.; BINO, R.J. & HILHORST,H.W.M. Cell division and subsequent radicleprotrusion in tomato seeds are inhibited byosmotic stress, but DNA synthesis andformation of microtubular cytoskeleton arenot. Plant Physiology, 114: 1-9, 2000.
DE CASTRO, R.D.; BINO, R.J.; JING, H-C.;KIEFT, H. & HILHORST, H.W.M. Depth ofdormancy in tomato (Lycopersiconesculentum Mill.) seeds is related to theprogression of the cell cycle prior to theinduction of dormancy. Seed ScienceResearch, in press, 2001.
DERKSEN, J.; WILMS, F.H.A. & PIERSON, E.S.The plant cytoskeleton: its significance in plantdevelopment. Acta Botanica Neerlandica. 39:1-18, 1990.
DERKX, M.P.M. & KARSSEN, C.M. Changingsensitivity to light and nitrate but not togibberellins regulates seasonal dormancypatterns in Sysimbrium officinale seeds.Plant, Cell and Environment, 16: 469-479,1993.
DERKX, M.P.M.; VERMEER, E. & KARSSEN,C.M. Gibberellins in seeds of Arabidopsisthaliana: Biological activities, identificationand effects of light and chilling onendogenous levels. Plant GrowthRegulators, 15: 223-234, 1994.
DULSON, J.; BEWLEY, J.D. & JOHNSTON,R.N. Abscisic acid is an endogenous inhibitorin the regulation of mannanase production byisolated lettuce (Lactuca sativa cv. GrandRapids) endosperms. Plant Physiology, 87:660-666, 1988.

132 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
DURE, L.I.; GREENWAY, S.C. & GALAU, G.A.Developmetal biochemistry of cotton-seedembryogenesis and germination: changingmessenger ribonucleic acid populations asshown by in vitro protein synthesis.Biochemistry, 20: 4162-4168, 1981.
DURE, L.I. Embryogenesis and gene expressionduring seed formation. Oxford Survey onPlant Molecular and Cell Biology, 2: 179-197, 1985.
ELDER, R.H. & OSBORNE, D.J. Function ofDNA synthesis and DNA repair in thesurvival of embryos during early germinationand in dormancy. Seed Science Research, 3:43-53, 1993.
FELDMAN, K.A.; MALMBERG, R.L. & DEAN,C. Mutagenesis in Arabidopsis. In:SOMMERVILLE, C.R. & MEYEROWITZ,E.M. (Ed.) Arabidopsis. New York, CSHLPress, 1994. p. 137-172.
FINCHER, G.B. Molecular and cellular biologyassociated with endosperm mobilization ingerminating cereal grains. Annual Review ofPlant Physiology and Plant MololecularBiology, 40: 305-346, 1989.
FRANCIS, D. & HERBERT, R.J. Regulation ofcell division in the shoot apex. In: ORMROD,J.C. & FRANCIS, D. (Ed.) Molecular andCellular Biology of the Plant Cell Cycle.Dordrecht, Kluwer Academic Publishers,1993. p. 201-210.
GALAU, G.A.; JAKOBSEN, K.S. & HUGHES,D.W. The controls of late dicotembryogenesis and early germination.Physiologia Plantarum, 81: 280-288, 1991.
GODDARD, R.H.; WICK, S.M.; SILFLOW, C.D.& SNUSTAD, D.P. Microtubule componentsof the plant cell cytoskeleton. PlantPhysiology, 104: 1-6, 1994.
GOLDBERG, R.B.; BARKER, S.J. & PEREZ-GRAU, L. Regulation of gene expressionduring plant embryogenesis. Cell, 56: 149-160, 1989.
GOLDBERG, R.B.; DE PAIVA, G. &YEDEGARI ,R. Plant embryogenesis: zygoteto seed. Science, 266: 605-614, 1994.
GÓRNIK, K.; DE CASTRO, R.D.; LIU, Y.;BINO, R.J. & GROOT, S.P.C. Inhibition ofcell division during cabbage (Brassicaoleracea L.) seed germination. Seed ScienceResearch, 7: 333-340, 1997.
GRATZNER, H.G. Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: a new reagentfor detection of DNA replication. Science, 218:474-475, 1982.
GROOT, S.P.C.; BRUINSMA, J. & KARSSEN,C.M. The role of endogenous gibberellinsin seed and fruit development of tomato:studies with a gibberellin-deficientmutant. Physiologia Plantarum, 71: 184-190, 1987.
GROOT, S. P. C. & KARSSEN, C.M. Dormancyand germination of abscisic acid-deficienttomato seeds: studies with the sitiens mutant.Plant Physiology, 99: 952 - 958, 1992.
GROOT, S.P.C.; DE CASTRO, R.D.; LIU, Y. &BINO, R.J. Cell cycle analysis in dormant andgerminating tomato seeds. In: ELLIS, R.H.;BLACK, M.; MURDOCH, A.J. & HONG,T.D. (Ed.) Basic and Applied Aspects ofSeed Biology. Dordrecht, Kluwer AcademicPublishers, 1997. p. 395-402.
GUNNING, B.E.S. & STEER, M.W. Plant cellbiology, structure and function. Massachusets,Jones & Bartlett Publishers, 1996.

Dormancy, Germination and the ... 133
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
GUNNING, B.E.S. & SAMMUT, M.Rearrangement of microtubules involved inestablishing cell division planes startimmediately after DNA synthesis and arecompleted just before mitosis. Plant Cell, 2:1273-1282, 1990.
HAIGH, A.M. Why do tomato seed prime? Sidney,Macquarie University, 1988. PhD thesis.
HARADA, J.J.; DIETRICH, R.; COMAI, L., &BADEN, C.S. Regulation of gene expressionduring seed germination and postgerminativedevelopment. In: VERMA, D.P.S. &GOLDBERG, R.B. (Ed.) Plant GeneResearch, Volume 5, Temporal and SpatialRegulation of Plant Genes. New York,Springer-Verlag. 1988. p. 26-39.
HARADA, J.J. Seed maturation and control ofgermination. In: LARKINS, B.A. & VASIL,I.K. (Ed.) Cellular and Molecular Biology ofPlant Seed Development. Dordrecht, KluwerAcademic Publishers. 1997. p. 545-592.
HILHORST, H.W.M. Dose-response analysis offactors involved in germination and secondarydormancy of seeds of Sisymbrium officinale. I.Phytochrome. Plant Physiology, 94: 1090-1095, 1990a.
HILHORST, H.W.M. Dose-response analysis offactors involved in germination and secondarydormancy of seeds od Sisymbrium officinale.II. Nitrate. Plant Physiology, 94: 1096-1102,1990b.
HILHORST, H.W.M. & KARSSEN, C.M. Seeddormancy and germination: The role ofabscisic acid and gibberellins and theimportance of hormone mutants. PlantGrowth Regulators, 11: 225-238, 1992.
HILHORST, H.W.M. A critical update on seeddormancy. I. Primary dormancy. SeedScience Research, 5: 61-73, 1995.
HILHORST, H.W.M. & DOWNIE, B. Primarydormancy in tomato (Lycopersiconesculentum cv. Moneymaker) seeds: Studieswith the sitiens mutant. Journal ofExperimental Botany, 47: 89-97, 1996.
HILHORST, H.W.M. Seed dormancy. SeedScience Research, 7: 221-223, 1997.
HILHORST, H.W.M. & TOOROP, P.E. Reviewon dormancy, germinability, and germinationin crop and weed seeds. Advances inAgronomy, 61: 112-165, 1997.
HILHORST, H.W.M.; GROOT, S.P.C. & BINO,R.J. The tomato seed as a model system to studyseed development and germination. ActaBotanica Neerlandica, 47: 169-183, 1998.
JING, H-C.; VAN LAMMEREN, A.M.; DECASTRO, R.D.; BINO, R.J.; HILHORST,H.W.M. & GROOT, S.P.C. β-tubulinaccumulation and DNA synthesis aresequentially resumed in embryo organs ofcucumber (Cucumis sativus L.) seeds duringgermination. Protoplasma, 208: 230-239,1999.
JONES, R.L. & JACOBSEN, J.V. Regulation ofsynthesis and transport of secreted proteins incereal aleurone. International Review onCytology, 126: 49-88, 1991.
KARSSEN, C.M. Seasonal patterns of dormancyin weed seeds. In: KHAN, A.A. (Ed.) ThePhysiology and Biochemistry of SeedDevelopment, Dormancy and Germination.Amsterdam, Elsevier. 1982. p. 243-270.
KARSSEN, C.M.; BRINKHORST-VAN, D.E.R.;SWAN, D.L.C.; BREEKLAND, A.E. &KOORNNEEF, M. Induction of dormancyduring seed development by endogenousabscisic acid: studies on abscisic aciddeficient genotypes of Arabidopsis thaliana.Planta, 157: 58-165, 1983.

134 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
KARSSEN, C.M. Seed science: characteristics andobjectives. In: CÔME, D. & CORBINEAU,F. (Ed.) Fourth International Workshop onSeeds: Basic and Applied Aspects of SeedBiology. Angers, Université Pierre et MarieCurie. 1993. p. 3-9.
KERMODE, A.R. Regulatory mechanisms in thetransition from seed development togermination: interactions between the embryoand the seed environment. In: KIGEL, J. &GALILI, G. (Ed.) Seed Development andGermination. New York, MarcelDekker.1995. p. 273-332.
KLEINSMITH, L.J. & KISH, V.M. Principles ofCell and Molecular Biology. New York,Harper Collins College Publishers, 1995.
KOORNNEEF, M.; REULING, G. & KARSSEN,C.M. The isolation and characterisation ofabscisic acid insensitive mutants ofArabidopsis thaliana. Plant Physiology, 61:377-383, 1984.
KOORNNEEF, M.; CONE, J.W.; KARSSEN,C.M.; KENDRICK, R.E.; VAN DER VEEN,J.H. & ZEEVAART, J.A.D. Plant hormoneand photoreceptor mutants in Arabidopsis andtomato. In: FREELING, M. (Ed.) PlantGenetics, UCLA Symposia on Molecularand Cellular Biology, New Series Vol. 35.New York, Alan Liss Inc., 1985. p. 1-12.
KOORNNEEF, M.; HANHART, C.J.; HILHORST,H.W.M. & KARSSEN, C.M. In vivo inhibitionof seed development and reserve proteinaccumulation in recombinants of abscisic acidbiosynthesis and responsiveness mutants inArabidopsis thaliana. Plant Physiology, 90:463-469, 1989.
KOORNNEEF, M. & KARSSEN, C.M. Seeddormancy and germination. In:MEYEROWITZ, E.M. & SOMMERVILLE,C.R. (Ed.) Arabdopsis. Cold Spring Harbor,Cold Spring Harbor Laboratory Press, 1994. p.313-334.
LÉON-KLOOSTERZIEL, K.M.; VAN DE BUNT,G.A.; ZEEVAART, J.A.D. & KOORNNEEF,M. Arabidopsis mutants with reduced seeddormancy. Plant Physiology, 110: 233-240,1996.
LIU, Y.; BERGERVOET, J.H.W.; RIC DE VOS,C.H.; HILHORST, H.W.M.; KRAAK, H.L.;KARSSEN, C.M. & BINO, R.J. Nuclearreplication activities during imbibition ofgibberellin- and abscisic acid-deficient tomato(Lycopersicon esculentum Mill.) mutant seeds.Planta, 194: 378-373, 1994.
LIU, Y.; BINO, R.J.; KARSSEN, C.M. &HILHORST, H.W.M. Water relations of GA-and ABA-deficient tomato mutants duringseed and fruit development and their influenceon germination. Physiologia Plantarum, 96:425-432, 1996.
LIU, Y.; HILHORST, H.W.M.; GROOT, S.P.C. &BINO, R.J. Amounts of nuclear DNA andinternal morphology of gibberellin andabscisic acid-deficient tomato (Lycopersiconesculentum Mill.) seeds during maturation,imbibition and germination. Annals ofBotany, 79: 161-168, 1997.
MEINKE, D.W. Molecular genetics of plantembryogenesis. Annual Review of PlantPhysiology and Plant Mololecular Biology,46: 369-394, 1995.

Dormancy, Germination and the ... 135
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
MURATA, T. & WATA, M. Re-formation ofpreprophase band after cold-induceddepolymerization of microtubules inAdiantum protonemata. Plant CellPhysiology, 32: 1145-1151, 1991.
OKAMURA, S.; KAKIUSHI, M.; SANO, A. &KAWAJIRI, M. Loss of tubulin during coldtreatment of cultured carrot cells. PhysiologyPlantarum, 88: 93-98, 1993.
OSBORNE, D.J. & BOUBRIAK, I.I. DNA status,replication and repair in desiccation toleranceand germination. In: ELLIS, R.H.; BLACK,M.; MURDOCH, A.J. & HONG, T.D. (Ed.)Basic and applied aspects of seed biology.Dordrecht, Kluwer Academic Publishers,1997. p. 23-32.
PIHAKASKI, M.K. & PUHAKAINEN, T. Effectof cold exposure on cortical microtubules ofrye (Secale cereale) as observed byimmunocytochemistry. PhysiologiaPlantarum, 93: 563-571, 1995.
RAGHAVAN, V. Molecular Embryology ofFlowering Plants. New York, CambridgeUniversity Press, 1997. 690p.
SMITH, O. Pollination and life-history studies oftomato (Lycopersicon esculentum Mill.).Cornell University Agricutural Stn. Memo,184: 3-16, 1935.
VARELA, F.G.; MATURANA, H.R. & URIBE,R. Autopoiesis: the organization of livingsystems, its characterisation and a model.Biosystems, 5: 187-196, 1975.
VARGA, A. & BRUINSMA, J. Tomato. In:MONSELISE, S.P. (Ed.) CRC Handbook ofFruit Set and Development. Boca Raton,CRC Press, 1986. p. 461-481.
VERTUCCI, C.W. The effects of cooling rate onseeds exposed to liquid nitrogentemperatures. Plant Physiology, 90: 1478-1485, 1989.
XUHAN, X. Seed development in Phaseolusvulgaris L., Populus nigra L., andRanunculus sceleratus L. with specialreference to the microtubular cytoskeleton.Wageningen, Wageningen University, 1995.150p. PhD. Thesis.
XU, N.; COULTER, K.M. & BEWLEY, J.D.Abscisic acid and osmoticum preventgermination of alfafa embryos, but onlyosmoticum maintains the synthesis ofdevelopmental proteins. Planta, 182: 382-390,1990.
XU, N. & BEWLEY, J.D. Sensitivity to abscisicacid and osmoticum changes duringembryogenesis of alfafa (Medicago sativa).Journal of Experimental Botany, 42: 821-826, 1991.
WALBOT, V. Control mechanisms for plantembryogeny. In: CLUTTER, M.E. (Ed.)Dormancy and Developmental Arrest. NewYork, Academic Press, Inc., 1978. p. 113-116.
WALKER-SIMMONS, M. ABA levels andsensitivity in developing wheat embryos ofsprouting resistant and susceptible cultivars.Plant Physiology, 84: 61-66, 1987.
WALLIN, M. & STROMBERG, E. Cold-stableand cold adapted microtubules. InternationalReview of Cytology, 157: 1-31, 1995.
WANG, T.L. & HEDLEY, C.L. Seed developmentin peas: knowing your three 'r's' (or four,or five). Seed Science Research, 1: 3-14,1991.

136 De Castro and Hilhorst
R. Bras. Fisiol. Veg., 12(Edição Especial):105-136, 2000
WELBAUM, G.E.; TISSAOUI, T. & BRADFORD,K.J. Water relations of seed development andgermination in muskmelon (Cucumis melo L.).III Sensitivity of germination to water potentialand abscisic acid during development. PlantPhysiology, 92: 1029-1037, 1990.
WEST, M.A. & HARADA, J.J. Embryogenesis inhigher plants: An overview. Plant Cell, 5:1361-1369, 1993.