domainanalysesrevealthat chlamydiatrachomatis … ... causes blinding trachoma, ... (tok.a.f.). s...
TRANSCRIPT

Domain Analyses Reveal That Chlamydia trachomatis CT694Protein Belongs to the Membrane-localized Family of Type IIIEffector Proteins*□S
Received for publication, May 31, 2012, and in revised form June 15, 2012 Published, JBC Papers in Press, June 18, 2012, DOI 10.1074/jbc.M112.386904
Holly D. Bullock1, Suzanne Hower1, and Kenneth A. Fields2
From the Department of Microbiology and Immunology, The Miller School of Medicine, Miami, Florida 33136
Background: The Chlamydia trachomatis secreted effector CT694 is deployed during invasion and exerts multiple effectson host cells.Results: Residues 40–80 of CT694 contain a domain necessary and sufficient for peripheral localization to eukaryoticmembranes.Conclusion: CT694 employs membrane association to manifest effects on host cells.Significance: Elucidating functional protein domains is essential to understand molecular mechanisms of infection employedby the pathogen C. trachomatis.
TheChlamydia trachomatis type three-secreted effector pro-tein CT694 is expressed during late-cycle development yet issecreted by infectious particles during the invasion process.Wehave previously described the presence of at least two functionaldomains within CT694. CT694 was found to interact with thehuman protein Ahnak through a C-terminal domain and affectformation of host-cell actin stress fibers. Immunolocalizationanalyses of ectopically expressed pEGFP-CT694 also revealedplasmamembrane localization for CT694 that was independentof Ahnak binding. Here we provide evidence that CT694 con-tains multiple functional domains. Plasma membrane localiza-tion andCT694-induced alterations in host cell morphology aredependent on an N-terminal domain. We demonstrate thatmembrane association of CT694 is dependent on a domainresembling a membrane localization domain (MLD) found inanti-host proteins fromYersinia,Pseudomonas, and Salmonellaspp. This domain is necessary and sufficient for localization andmorphology changes but is not required forAhnakbinding. Fur-ther, the CT694MLD is able to complement ExoS �MLDwhenectopically expressed. Taken together, our data indicate thatCT694 is a multidomain protein with the potential to modulatemultiple host cell processes.
Chlamydia trachomatis infectionhas been themost reportedsexually transmitted disease in the United States because1994,with over 1.2 million cases reported in 2009 (1). However, it isbelieved that the true number of cases is much higher becauseof the potential for asymptomatic infections, particularly inmales (1). Sequelae resulting fromuntreated or repeatedC. tra-chomatis serovar D-K infections can include infertility, pelvicinflammatory disease, ectopic pregnancy, or pelvic pain (2).
Additionally, ocular infectionwithC. trachomatis serovarsA-Ccauses blinding trachoma, the leading cause of preventableblindness worldwide, particularly in developing countries (3).An obligate intracellular bacterium, C. trachomatis exhibits
a biphasic developmental cycle consisting of an extracellular,non-metabolic elementary body (EB)3 and an intracellular, rep-licative reticulate body (RB) (4). Both particle types posses afunctional type-III secretion system (T3SS), which is essentialfor bacterial development (5). Contact with the host cell surfacetriggers secretion of effectors through the T3SS into the hostcell (6). In Chlamydia, these immediate early effectors are pre-packaged during the reticulate body-to-EB transition to bereadily available immediately upon host cell contact (7, 8). Onesuch effector is the conserved translocated actin recruitingphosphoprotein (TarP), which is translocated into host cellswithin minutes of host cell contact and induces actin cytoskel-etal reorganization that is important for invasion (9). The abil-ity to dynamically reorganize the host cytoskeleton is a com-mon general theme among bacteria expressing T3SS, includingYersinia spp., Salmonella, Shigella spp., and Pseudomonasaeruginosa (10–12).To achieve efficient anti-host function, some T3SS effec-
tors must be targeted to the correct subcellular compart-ment (12). The presence of a membrane localization domain(MLD) is one mechanism employed to accomplish this goal.For example, discrete MLD domains within YopE of Yersiniaspp. or ExoS of P. aeruginosa mediate association with hostmembranes (12). These MLDs lack the characteristic pre-dicted hydrophobic � helix of a transmembrane domain (13)but contain a leucine-rich region that is essential for mem-brane association (12, 14). It has been proposed that inter-actions with membranes could be direct (15). Alternatively,there is evidence that these membrane-localized effectors do
* This work was supported by Public Health Service Grant AI065530 fromNIAID, National Institutes of Health (to K. A. F.).
□S This article contains supplemental Fig. S1 and Table S1.1 Both authors contributed equally to this work.2 To whom correspondence should be addressed: Department of Microbiol-
ogy and Immunology, University of Miami, 1600 NW 10th Ave., Miami, FL.Tel.: 305-243-6711; E-mail: [email protected].
3 The abbreviations used are: EB, elementary body; T3SS, type-III secretionsystem; MLD, membrane localization domain; hpi, hours post-infection;BD, binding domain; AD, activating domain; MOI, multiplicity of infection;MOMP, major outer membrane protein; GAP, GTPase-activating protein;EGFP, enhanced green fluorescent protein.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 287, NO. 33, pp. 28078 –28086, August 10, 2012© 2012 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
28078 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 33 • AUGUST 10, 2012
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

so through interactions with membrane-associated proteinsrather than direct interactions with the host cell membrane(14, 16, 17). Regardless of the mechanism, localization ofeffectors to cellular membranes allows a targeted response inwhich respective effector proteins manifest activities in aconstrained microenvironment.CT694 is a recently described C. trachomatis-specific T3SS
effector (18). Aside from a predicted coiled-coil domain fromresides 285–305, the primary sequence of CT694 does not con-tain additional domains identifiable via similarity searches ordomain predictions (18). Like TarP (19), this protein is translo-cated during host cell invasion (18). CT694 protein levelsdecrease slightly around 1 h post-infection (hpi) but are thenmaintained until a robust increase during de novo CT694 syn-thesis at 18–24 hpi during late-cycle development (7, 18). Pre-vious work (18) demonstrated that a GFP-CT694 chimeralocalizes to the plasma membrane, where it interacts withAhnak, a large human protein involved in cytoskeleton main-tenance and cell signaling (20, 21). Ectopic expression studiesalso revealed that deletion of the C terminus of CT694 pre-cludes the interaction with Ahnak but does not affect mem-brane localization (18). Herein, we test the possibility that theNterminus of CT694 expresses an MLD that is necessary forlocalization of CT694 to host membranes.
EXPERIMENTAL PROCEDURES
Strains and Culture Conditions—HeLa 229 epithelial cells(CCL 2.1, ATCC) were maintained in RPMI 1640 (Invitrogen)supplemented with 10% (v/v) FBS (Sigma-Aldrich, St. Louis,MO) at 37 °C in the presence of 5% CO2/95% humidified air.C. trachomatis serovar L2 (LGV 434, ATCC)was propagated inHeLa cells and purified through MD-76R (Mallinckrodt, St.Louis, MO) density gradients as described previously (22). Forinfections, HeLa monolayers were inoculated with C. tracho-matis in Hank’s balanced salt solution (Invitrogen) and incu-bated at 37 °C for 1 h as described (22, 23). Inocula werereplaced with RPMI 1640 � 10% FBS (v/v) and incubated for24 h, unless otherwise indicated.DNA Methods—C. trachomatis open reading frames were
amplified from C. trachomatis serovar L2 genomic DNA usingEconoTaq PLUS Green Master Mix (Lucigen, Middleton, WI)according to the guidelines of themanufacturer and using customoligonucleotide primers containing engineered restriction sites,synthesized by Integrated DNA Technologies (Coralville, IA).Cloningwasperformedaccording to standardprotocols (24). PCRproductswere ligated intovectorsutilizing theappropriate restric-tion enzymes, unless otherwise noted. Primer sequences withrestriction sites are listed in supplemental Table S1. Transforma-tions with plasmid DNAs were performed using chemically com-petent Escherichia coliDH5� strains (Invitrogen). CT694 trunca-tions were cloned into pEGFP-C3 (Clontech, Mountain View,CA). Primers SHCT694-1A and SHCT694-6B were usedfor CT694-�39, SHCT694-1A and SHCT694–8B for CT694-�192, SHCT694–1A and SHCT694-9B for CT694-�243, andSHCT694-10A and SHCT694-4B2 for �42-CT694. CT694�40-80 was generated using primers CT694 MLD F and CT694MLD R. The resulting PCR products were digested with theappropriate restriction enzymes and ligated into pEGFP-C3.
CT694-�40-80 was generated by Genewiz, Inc. (South Plain-field, NJ) using the GFP-CT694 backbone. GFP-ExoS/694-MLD was generated by insertion of restriction enzymesites flanking the ExoS MLD on full-length GFP-ExoS byQuikchange Lightning (Agilent Technologies, Inc., SantaClara,CA) according to the directions of the manufacturer and usingprimers ExoS-EcoRI-QC F/R and ExoS-KpnI-QC. The CT694MLD insert was amplified using primers CT694 MLD EcoRIand CT694 MLD KpnI. Resulting PCR products were digestedwith the appropriate restriction enzymes and ligated to createGFP-ExoS/694 MLD. Primers CT694 MLD EcoRI 2 forwardand reverse were used to correct out-of-frame ligation. Allexpression constructs were verified by DNA sequencing(Oncogenomics Core Facility, Sylvester Comprehensive Can-cer Center, University of Miami Miller School of Medicine,Miami, FL).P. aeruginosa open reading frames were amplified from
either P. aeruginosa PA01 genomic DNA (Greg Plano, MillerSchool ofMedicine, University ofMiamiMiller School ofMed-icine, Miami, FL) or P. aeruginosa PA103�UT expressing plas-mid-encoded pUCP-ExoSmutants (JoanOlson, Robert C. ByrdHealth Sciences Center, West Virginia University, Morgan-town,WV) using EconoTaq PLUSGreenMasterMix (Lucigen)and primers ExoS F and ExoS R. Full-length ExoS and ExoSmutants were digested with the appropriate restrictionenzymes and ligated into pEGFP-C3 (Clontech).Immunodetection—For immunoblot analysis, samples were
precipitated with either 10% trichloroacetic acid (sucrose gra-dients) or 50% acetone (Triton X-114 extractions), solubilizedin 3� Laemmli buffer and resolved by SDS-PAGE 12% (v/v)homogenous polyacrylamide gels or SDS-PAGE 4–20% gradi-ent polyacrylamide gels (Bio-Rad Laboratories, Hercules, CA),followed by transfer to Immobilon-P membranes (Millipore,Billerica, MA) in Tris-glycine buffer. Detection of specific pro-teins was accomplished using �-CT694 (18), �-MOMP (18),�-Tarp C terminus (19), �-Caveolin-1 (BD Biosciences),�-GAPDH (Millipore), or �-GFP (Sigma). After incubationwith specific primary antibodies, immunoblots were probedwith the appropriate horseradish peroxidase-conjugated sec-ondary antibodies (Sigma) followed by development withAmersham Biosciences ECL Plus (GE Healthcare UK Limited,Buckinghamshire, UK).Transfection and Microscopy—Semiconfluent HeLa mono-
layers were grown on 12-mm-diameter glass coverslips forimmunofluorescence analysis. Monolayers were directlyinfected with C. trachomatis or transfected with endotoxin-free plasmid DNAs. Transfections were accomplished usingLipofectamine 2000 according to the directions of the manu-facturer (Invitrogen). For indirect immunofluorescence, sam-ples were fixed with methanol, blocked in 5% BSA (Sigma) inTris-buffered saline plus Tween 20 (TBST) (137mMNaCl, 2.68mM KCl, 2.48 mM Tris base (pH 7.4), supplemented with 0.5%(v/v) Tween 20), and appropriate antibodies were diluted in 5%BSA inTBST. Proteinswere visualized by direct fluorescence ofGFP-containing proteins or, where indicated, with �-GFP(Sigma), and the appropriate secondary antibody conjugated toAlexa Fluor 488 (Invitrogen). Nuclear staining was achieved bystaining with DAPI (Invitrogen). Images were acquired by epi-
Membrane Localization Domain of CT694
AUGUST 10, 2012 • VOLUME 287 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 28079
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

fluorescence microscopy using a �60 apochromat objectiveplus �1.5 intermediate magnification on a TE2000U invertedphotomicroscope (Nikon,Melville, NY) equipped with a RetigaEXi 1394, 12-bit monochrome CCD camera (QImaging, Sur-rey, BC, Canada) and MetaMorph imaging software version6.3r2 (Molecular Devices, Downington, PA). Images were pro-cessed using Adobe Photoshop CS2 version 9.0 (Adobe Sys-tems, San Jose, CA).Yeast Two-hybrid Assay—Yeast two-hybrid assays were per-
formed as described (18). Briefly, specific plasmid constructswere transformed into Saccharomyces cerevisiae AH109 usingthe S. cerevisiae Easycomp transformation kit (Invitrogen) fol-lowed by selection on S.D. agar plates deficient in leucine (Leu)and tryptophan (Trp) (Clontech). CT694 (full-length and trun-cations) were cloned in-frame downstream of the gal4 bindingdomain (BD) and used as bait in the MATCHMAKER TwoHybrid System 3 (Clontech). Primers used are listed in supple-mental Table S1. Interaction studies with Ahnak were per-formed with engineered Ahnak constructs cloned downstreamof the gal4 activating domain (AD) of pGADT7 (Clontech).Colonies expressing interacting proteins were selected for onQDOx-�-gal. To rule out growth because of auto-activation onselective media, BD/CT694 was cotransformed with AD/T orpGADT7 (AD/Empty). In the same way, Ahnak constructswere cotransformed with BD/53 or pGBKT7 (BD/Empty).Yeast protein expression was evaluated by immunoblot with�-AD, �-BD, �-HA (Santa Cruz Biotechnology, Santa Cruz,CA), �-c-myc (BD Biosciences), or �-CT694.Membrane Fractionation—Semiconfluent HeLa monolayers
were mock-infected or inoculated with C. trachomatis at anMOI of 100 for 2 or 24 h, as indicated. The cultures were thenscraped into 10ml of ice-cold 0.25 M sucrose buffer (10mMTris(pH 7.5), 1 mM EDTA, 0.25 M sucrose) supplemented with pro-tease inhibitors (complete mixture, Roche). Whole-culturematerial was homogenized for 20–25 strokes in a Douncehomogenizer followed by centrifugation to remove cell debris.The resulting supernatant was layered over a 24–44% sucrosegradient, centrifuged, and processed as described (25).Triton X-114 extractions were performed as described pre-
viously (26). HeLa monolayers were transfected or infectedwithC. trachomatis serovar L2. Cultures were lysed in 1.5ml ofice-cold 1% Triton lysis buffer (1% Triton X-114 (Sigma), 100mM KCl, 50 mM Tris-HCl (pH 7.4)), rotated for 30 min at 4 °C,and clarified by centrifugation for 35 min (14,000 rpm at 4 °C)using an Eppendorf 5810 R centrifuge (Eppendorf AG, Ham-burg, Germany). The resulting supernatant was incubated at37 °C for 10 min, followed by centrifugation for 12 min (14,000rpmat room temperature) to separate the detergent phase (bot-tom fraction) from the aqueous phase (top fraction). The aque-ous phase was removed to a fresh tube containing 200�l of 10%Triton X-114 buffer (10% Triton X-114, 100 mM KCl, 50 mM
Tris-HCL (pH 7.4)), and the detergent phase was mixed with 1ml of buffer 1 (50 mM Tris (pH 7.4), 100 mM KCl). Both phaseswere incubated on ice for 10 min, followed by 10 min of incu-bation at 37 °C, and finally centrifugation for 12 min (14,000rpm at room temperature). The top fraction of the detergentphase was removed, and the detergent-rich phase was mixedwith 1 ml of buffer 1. The detergent-depleted aqueous phase
was again mixed with 200 �l of 10% Triton X-114 buffer, andeach tube was subjected to the same incubations on ice, fol-lowed by 37 °C, followed by centrifugation, repeated a total offour times. The final detergent and aqueous phases were pre-cipitated in 50% acetone (v/v) at �20 °C overnight.Statistical Analyses—All experiments were repeated a mini-
mum of three times, and representative data are shown. Statis-tical analysis was evaluated using a two-tailed Student’s t testwhere appropriate. In all figures, p values were labeled as fol-lows: *, p � 0.01 and **, p � 0.001. Statistical analysis was per-formed using GraphPad Prism software.
RESULTS
Endogenous CT694 Is Membrane-associated During EarlyCycle Development—We have demonstrated previously thatectopically expressed CT694 is capable of colocalization withthe plasma membrane of transfected HeLa cells (18). Thesedata raised the possibility that CT694 membrane localizationcorrelates with effector function(s). We first sought to investi-gate the subcellular partitioning of endogenous CT694 to con-firm the relevance of interaction with host cell membranes.HeLa cells were infected with C. trachomatis serovar L2 at anMOI of 100, disrupted 2 hpi, and lysates were subjected to asucrose gradient fractionation. Mock-infected parallel cultureswere similarly processed as an immunoblot control for anti-body specificity (data not shown). Proteins in subsequent gra-dient fractions were probed in immunoblot analyses with anti-bodies specific for CT694 or TarP. Immunoblot analyses wereprobed with C. trachomatis major outer membrane protein(MOMP) antibodies or humanCaveolin-1 as controls for intactbacteria and hostmembranes, respectively (Fig. 1). BothCT694and TarP were detected in fractions 1–5 as well as fraction 14.Conversely, MOMP was detected only in the lower columnfractions, indicating migration of intact chlamydiae. Asexpected, the host transmembrane protein Caveolin-1 waspresent in the top fractions, indicating that these fractions con-tained the majority of host cell membrane material (Fig. 1).These results are consistentwithmembrane association of bothCT694 and TarP during C. trachomatis infections. This local-ization was specific for early infection time points becauseCT694 was detected solely in fractions containing intact bacte-ria when cultures were harvested during late-cycle develop-ment (supplemental Fig. S1).We examined phase separation of CT694 in detergent
extracts to further assess the subcellular localization of endog-
FIGURE 1. CT694 partitions with membranes and intact bacteria in whole-culture sucrose floatation gradients. HeLa cell monolayers were infected atan MOI of 100 with C. trachomatis (L2). Cultures were harvested at 2 hpi, andmaterial was separated via centrifugation in a sucrose density gradient. Mate-rial from 14 equivalent column fractions (top panel, fraction 1; bottom panel,fraction 14) was concentrated via TCA precipitation, and proteins wereresolved via SDS-PAGE. Fractions were probed in immunoblot analyses forCT694, TarP, and the fractionation controls MOMP and Caveolin-1.
Membrane Localization Domain of CT694
28080 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 33 • AUGUST 10, 2012
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

enous CT694 during C. trachomatis infection. HeLa cells wereinfected with C. trachomatis at anMOI of 100 and subjected toTriton X-114 extraction at 1, 3, and 6 hpi. At each time point,CT694 was detected in both the detergent and aqueous phasesof infected cells (Fig. 2A). GAPDH, Caveolin-1, and MOMPwere used as controls and fractionated as expected. UnlikeCT694, TarPwas detected only in aqueous fractions, indicatingthat anymembrane association of TarP is fundamentally differ-ent from and perhaps less direct than that of CT694. TritonX-114 extraction was also performed on equivalent amounts ofdensity gradient-purified EBs. In these experiments, CT694was detected solely in aqueous material, indicating that Chla-mydia-localized CT694 is not associated with membranes (Fig.2B). In aggregate, these data are consistentwith secretedCT694stably associating with eukaryotic membranes during invasionand early-cycle development.Membrane Localization Domain of CT694—Transmem-
brane prediction tools such as TMPred (27) and Kyte Doolittle(13) analysis do not indicate the presence of typical transmem-brane domainswithin the primary sequence of CT694 (data notshown). Therefore, a systematic series of GFP-tagged CT694truncations were engineered and expressed in HeLa cells todelineate the minimal domain necessary for membrane local-ization (Fig. 3A). Equivalent truncations were generated in theyeast two-hybrid expression system to assess the impact onCT694 interaction with human Ahnak. As reported previously(18), concentrations of full-length CT694 (694-FL, Fig. 3C)could be detected in both cytosolic aggregates and plasmamembranes. The cytosolic aggregates are not present in allcells, andwe cannot rule out the possibility that they are a prod-uct of CT694 overexpression. Curiously, the accumulation ofaggregates seems positively correlated with membrane local-ization. Deletion of the C-terminal 39 (694-�39), 101 (694-�101), or 192 residues (694-�192) did not affect CT694 local-ization but did disrupt CT694 association with Ahnak in yeasttwo-hybrid studies. Expression of CT694 residues 43–322(�42–694) did not interfere with Ahnak interaction but didresult in less overt localization of CT694 to plasmamembranes.Finally, in-frame deletion of residues 40–80 (694-�40–80)completely abolishedmembrane localization of CT694, but didnot affect the interaction with Ahnak. These data indicate that
a domain within residues 40–80 is necessary for membranelocalization but not for interaction with Ahnak. Consistentwith previous studies, CT694-transfected HeLa cells also rou-tinely displayed aberrant cell morphology in which cellularedges had a feathered appearance (Fig. 3B). This phenotypecorrelated with membrane localization because versions ofCT694 (694-�243, �42–694) that showed decreased abun-dance in membrane localization also mediated less overt alter-ations in HeLa morphology. In addition, HeLa cells transfectedwith 694-�40–80 were comparable with the GFP control withrespect to gross morphology (Fig. 3C).Residues 40–80 of CT694 were expressed as a GFP-fusion
protein to further assess the ability of this segment to act as amembrane localization domain (Fig. 4A). Similar to full-length694, the signal for GFP�40–80 was detectible in the plasmamembrane of transfected HeLa cells. Interestingly, the cellularmorphology appeared comparable with the GFP-only control.Triton X-114 extraction was also performed on HeLa cellsexpressing GFP, 694-FL, 694-�40–80, or GFP�40–80 to con-firm and quantify membrane localization of chimeric proteins.Immunoblot analyses confirmed that ectopically expressedCT694, like endogenous CT694, partitioned into both thedetergent and aqueous fractions (Fig. 4B). Densitometry analy-sis of immunoblots revealed nearly equal distribution of protein(Fig. 4C). Partitioning of 694-�40–80 appeared similar to theGFP-only control and was detected predominantly in the aque-ous fraction. Conversely, GFP�40–80 was detected mostabundantly in the detergent phase, indicating that these resi-dues were both necessary and sufficient formembrane localiza-tion of CT694.Residues 40–80 Constitute a Functional Membrane Local-
ization Domain—The mechanism for membrane associationwas unclear given the lack of a predicted transmembranedomain inCT694. Interestingly, T3SS effector proteins in othersystems have been shown to possess a functional MLD that isessential for these proteins to express respective anti-host func-tions. The best characterized effectors of this family include theYersinia T3SS effector YopE and ExoS of P. aeruginosa (12).Both YopE and ExoS are targeted to the host cell plasma mem-brane upon translocation via a discreteMLD domain, and suchlocalization is necessary to exert RhoGAP activity (12, 28–30).Consistent with the divergent nature of amino acids among theMLD family of proteins, there was no significant similaritybetween CT694 and theMLD domains of either YopE or ExoS.Likewise, there is no significant overall sequence similaritybetween full-length CT694 and either YopE or ExoS. This is incontrast to direct comparisons of YopE and ExoS, which sharesignificant N-terminal homology (12, 31). However, all threedomains are predicted to contain an�helix of approximately 20residues (residues 51–72 of ExoS and 54–75 of YopE) (data notshown, 12). Because the YopE and ExoS MLD domains can befunctionally exchanged (12), we tested whether replacing theMLDof ExoSwith amino acids 40–80 ofCT694would result infunctional complementation.Transient expression of GFP-ExoS in mammalian cells
results in cell rounding, and mutation of the MLD is sufficientto abrogate this phenotype because maintained membranelocalization is required for optimal RhoGAP activity of ExoS
FIGURE 2. Endogenous CT694 associates with host cell membranes invivo. HeLa cell monolayers infected with C. trachomatis (L2) at an MOI of 100for 1, 3, or 6 h (A) or density gradient-purified EBs (B) were subjected to TritonX-114 detergent extraction. Proteins in detergent (D) and aqueous (A) phaseswere concentrated via acetone precipitation and resolved by SDS-PAGE.A, fractions were probed in immunoblot for CT694 as well as TarP and MOMPas bacterial aqueous and detergent controls, respectively, or GAPDH andCaveolin-1 as respective host cell aqueous and detergent controls. B, deter-gent phase and aqueous phase proteins from EBs were probed in immuno-blot for CT694, TarP, or MOMP.
Membrane Localization Domain of CT694
AUGUST 10, 2012 • VOLUME 287 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 28081
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE 3. CT694 truncation analyses. A, CT694 truncations were engineered and expressed either in HeLa cells as GFP fusion proteins to examine membranelocalization or in S. cerevisiae for yeast-two hybrid (Y2H) screens to assess interaction with Ahnak. A “�” for Ahnak-binding indicates growth on stringentselection medium and is indicative of interaction between Ahnak and the respective version of CT694. A “�” for membrane localization indicates fluorescencemicroscopy detection of the recombinant protein in a pattern consistent with plasma membrane localization. B, representative images demonstratingaberrant cell morphology of HeLa cells expressing GFP-694-FL compared with cells expressing GFP empty vector. Insets represent digitally zoomed areasindicated by arrows. C, representative images for membrane localization and morphology changes for the indicated GFP-CT694 mutants. Scale bar � 10 �m.
Membrane Localization Domain of CT694
28082 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 33 • AUGUST 10, 2012
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

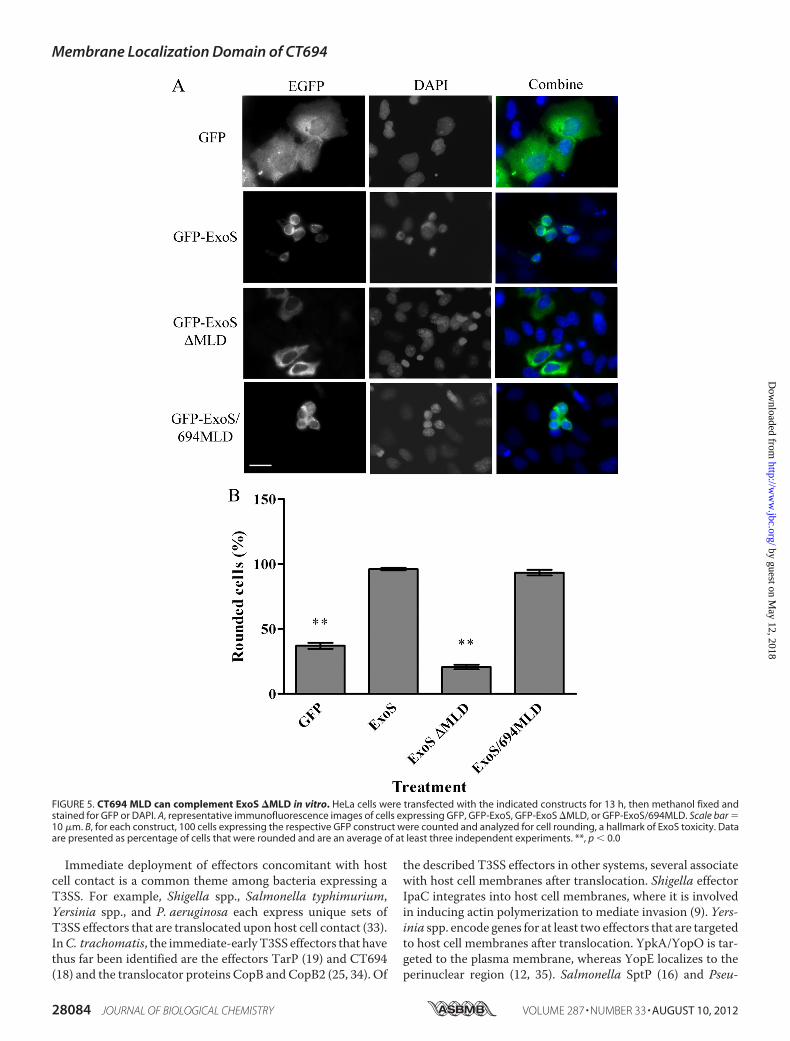
(14, 28, 32). We tested whether complementation of ExoS�MLD with the CT694 MLD (ExoS/694MLD) would restoreExoS-mediated HeLa cell rounding. HeLa cells were transfectedwith 400ngof the indicatedDNA, andGFPorDAPI stainingweresubsequently used to assess subcellular localization and cellrounding (Fig.5).Aspublishedpreviously,GFP-ExoSwasdetectedin perinuclear concentrations, whereas GFP-ExoS �MLD waslocalized diffusely in the cytosol (15). CT694 MLD was able tocomplement both the perinuclear localization and the roundedcell phenotype incells expressingGFP-ExoS/694MLD(Fig. 5A). Inaddition, rounded cells were quantified for each construct, withGFP-ExoS/694MLD displaying a similar percentage of roundedcells compared with GFP-ExoS WT, whereas vector-only andGFP-ExoS �MLD had significantly fewer rounded cells (Fig. 5B).These data are therefore consistent with residues 40–80 contain-ing a functional MLD that targets the chlamydial effector to theplasmamembraneof infectedhost cells, similar to theMLDfamilyof T3S effector proteins.
DISCUSSION
The data presented here support amodel in which theC. tra-chomatis T3SS effector CT694 contains multiple functional
domains. These domains include the previously describedAhnak binding domain (18), an undefined toxicity domainthat manifests as altered cell morphology in transfectedHeLa cells, and a membrane localization domain residingwithin amino acids 40 to 80. The CT694MLD is necessary totarget CT694 to host cell membranes. Our transfection stud-ies show that this localization is independent of the Ahnakbinding domain, and the MLD alone is sufficient to localizeGFP to the plasma membrane. However, the MLD alone isnot sufficient to induce host cell morphology changes, sug-gesting that although the Ahnak-binding activity is inde-pendent of localization, toxicity is not. In the absence of atractable genetic system, we cannot directly establish therole of this domain during C. trachomatis infection. How-ever, endogenous CT694 also localizes to cellular mem-branes as early as immediately post-infection and stays asso-ciated with membranes for up to 6 h post-infection. Thesedata provide evidence that secreted CT694 does associatewith host membranes and is consistent with our model inwhich CT694 plays a role in host cell invasion or establish-ment of a subcellular niche for C. trachomatis development.
FIGURE 4. Amino acids 40 – 80 are sufficient for membrane localization. A, HeLa cell monolayers were transfected for 18 h with vector only or GFP�40 – 80,and images were captured by direct immunofluorescence. Scale bar � 10 �m. B and C, HeLa cell monolayers were transfected for 18 h with the indicatedplasmid DNAs and then subjected to Triton X-114 detergent extraction. Proteins in detergent (D) and aqueous (A) phases were concentrated via acetoneprecipitation and resolved by SDS-PAGE. Immunoblot analyses were probed for GFP or GAPDH. B, a representative image. C, total protein in each fraction wasquantified by densitometry, and statistics were generated by Student’s t test.
Membrane Localization Domain of CT694
AUGUST 10, 2012 • VOLUME 287 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 28083
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from
Immediate deployment of effectors concomitant with hostcell contact is a common theme among bacteria expressing aT3SS. For example, Shigella spp., Salmonella typhimurium,Yersinia spp., and P. aeruginosa each express unique sets ofT3SS effectors that are translocated upon host cell contact (33).InC. trachomatis, the immediate-earlyT3SS effectors that havethus far been identified are the effectors TarP (19) and CT694(18) and the translocator proteins CopB and CopB2 (25, 34). Of
the described T3SS effectors in other systems, several associatewith host cell membranes after translocation. Shigella effectorIpaC integrates into host cell membranes, where it is involvedin inducing actin polymerization to mediate invasion (9). Yers-inia spp. encode genes for at least two effectors that are targetedto host cell membranes after translocation. YpkA/YopO is tar-geted to the plasma membrane, whereas YopE localizes to theperinuclear region (12, 35). Salmonella SptP (16) and Pseu-
FIGURE 5. CT694 MLD can complement ExoS �MLD in vitro. HeLa cells were transfected with the indicated constructs for 13 h, then methanol fixed andstained for GFP or DAPI. A, representative immunofluorescence images of cells expressing GFP, GFP-ExoS, GFP-ExoS �MLD, or GFP-ExoS/694MLD. Scale bar �10 �m. B, for each construct, 100 cells expressing the respective GFP construct were counted and analyzed for cell rounding, a hallmark of ExoS toxicity. Dataare presented as percentage of cells that were rounded and are an average of at least three independent experiments. **, p � 0.0
Membrane Localization Domain of CT694
28084 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 33 • AUGUST 10, 2012
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

domonas ExoS (14) also have established roles that requireassociation with host membranes. AexT and AexU of Aeromo-nas spp. likely represent additional familymembers on the basisof sequence similaritywith theMLDofYopE (29). Although theMLD of YopE can functionally complement loss of the ExoSMLD (33), the precise nature of the membrane associationremains unclear. Barbieri and colleagues (12) and Zhang andBarbieri (14) proposed that a leucine-rich motif arranged onone side of a predicted� helix within theMLDof these proteinsmediates membrane association. More hydrophilic residueswithin the domain are proposed to have a membrane-stabiliz-ing role (28).The data presented here demonstrate that CT694 may asso-
ciate with host membranes in a similar manner as ExoS andYopE. CT694 is translocated through the T3SS upon host cellcontact and is then targeted to the host cell membrane. Thismembrane localization is dependent on a discrete domain, sim-ilar to YopE and ExoS, that also lacks a predicted transmem-brane structure (13).However, CT694 appears to lack an appar-ent leucine-rich domain. Therefore, we cannot rule out thepossibility of distinct molecular mechanisms manifesting asstable interaction with host membranes. Interestingly, ectopi-cally expressed ExoS also partitioned to the TritonX114 phaseof fractionated HeLa cells similar to CT694 (data not shown).Coupled with our complementation data, we believe that,regardless of mechanism, the MLD of CT694 accomplishes afunctional role similar to that of other MLD-containingeffectors.Although this class of membrane-localized effector proteins
displays some similarities, the role each effector plays variesdepending on the system. YopE and ExoS each contain GTPaseactivating protein (GAP) domains targeting Rho familyGTPases to prevent host cell phagocytosis of bacteria (30, 36).On the opposite end of the spectrum, the Shigella effector IpaCplays an important role in cytoskeleton rearrangement to pro-mote bacterial invasion and uptake by host cells (37). SptP, aSalmonella T3SS effector, also localizes to host cell plasmamembranes, where it is hypothesized to participate in repairingthe host cell membrane after bacterial invasion through theactivity of the SptPN-terminalGAPdomain (11, 16, 38).On thebasis of our collective observations, we believe that CT694functions at the host cell membrane duringC. trachomatis hostcell invasion and early cycle development. Later functions havenot been ruled out but would presumably not require mem-brane association because endogenous CT694 did not fraction-ate with the membrane during late-cycle development. Intransfected cells, CT694 is able to disrupt actin stress fibers(18). Moreover, our domain analyses indicate an activity resid-ing between the MLD and Ahnak binding domain that medi-ates altered cellular morphology in transfected HeLa cells. It isformally possible that CT694 expresses RhoGAP activity, yetthe morphological phenotype induced by CT694 differs signif-icantly from those induced by YopE (39), ExoS (40), and SptP(11). TarP-mediated arrangement of actin and recruitment tothe invasion site is a necessary step to C. trachomatis invasion(9), andRac1, but not RhoAorCdc42, is recruited and activatedduring C. trachomatis invasion (41). We have been unable todetect Rac1-specific GAP activity for CT694 (data not shown)
but cannot exclude the possibility of GAP activity for CT694directed toward alternative host targets. Interestingly, Salmo-nella utilize a careful balance of effectors to promote actinnucleation through SopEGEF activity and revert and repair thehost cell cytoskeleton and membrane through SptP GAP activ-ity (42). It is possible that CT694may have a similar general roleduringC. trachomatis infection and that one function ofCT694is to repair cytoskeletal insult because of TarP activity. We arecurrently pursuing work to address this intriguing possibility.
Acknowledgments—We thank Drs. K. Wolf, H. Betts-Hampikian, K.Mueller, M. McKuen, and B. Chellas-Géry for critical reading of themanuscript. We are also grateful to B. Chellas-Géry for technicalassistance and Drs. Greg Plano and Joan Olson for their kind gifts ofP. aeruginosa strains and genomic DNA.
REFERENCES1. Sexually Transmitted Disease Surveillance 2009 (2010) United States De-
partment of Health andHuman Services, Centers for Disease Control andPrevention
2. Haggerty, C. L., Gottlieb, S. L., Taylor, B. D., Low,N., Xu, F., andNess, R. B.(2010) Risk of sequelae after Chlamydia trachomatis genital infection inwomen. J. Infect. Dis. 201, S134-S155
3. Burton, M. J., and Mabey, D. C. (2009) The global burden of trachoma. Areview. Travel Med. Infect. 3, e460
4. Moulder, J. W. (1991) Interaction of chlamydiae and host cells in vitro.Microbiol. Rev. 55, 143–190
5. Wolf, K., Betts, H. J., Chellas-Géry, B., Hower, S., Linton, C. N., and Fields,K. A. (2006) Treatment of Chlamydia trachomatis with a small moleculeinhibitor of the Yersinia type III secretion system disrupts progression ofthe chlamydial developmental cycle.Mol. Microbiol. 61, 1543–1555
6. Betts-Hampikian, H. J., and Fields, K. A. (2010) The chlamydial type IIIsecretion mechanism: revealing cracks in a tough nut. Front. Microbiol. 1,1
7. Belland, R. J., Zhong, G., Crane, D. D., Hogan, D., Sturdevant, D., Sharma,J., Beatty, W. L., and Caldwell, H. D. (2003) Genomic transcriptional pro-filing of the developmental cycle of Chlamydia trachomatis. (2011) Proc.Natl. Acad. Sci. U.S.A. 100, 8478–8483
8. Fields, K. A., Mead, D. J., Dooley, C. A., and Hackstadt, T. (2003) Chla-mydia trachomatis type III secretion. Evidence for a functional apparatusduring early cycle development.Mol. Microbiol. 48, 671–683
9. Jewett, T. J., Miller, N. J., Dooley, C. A., and Hackstadt, T. (2010) Theconserved Tarp actin binding domain is important for chlamydial inva-sion. PLoS Pathog. 6, e1000997
10. Carabeo, R. A., Grieshaber, S. S., Fischer, E., and Hackstadt, T. (2002)Chlamydia trachomatis induces remodeling of the actin cytoskeleton dur-ing attachment and entry into HeLa cells. Infect. Immun. 70, 3793–3803
11. Fu, Y., andGalán, J. E. (1998) The Salmonella typhimurium tyrosine phos-phatase SptP is translocated into host cells and disrupts the actin cytoskel-eton.Mol. Microbiol. 27, 359–368
12. Krall, R., Zhang, Y., and Barbieri, J. T. (2004) Intracellular membranelocalization of Pseudomonas ExoS and Yersinia YopE inmammalian cells.J. Biol. Chem. 279, 2747–2753
13. Kyte, J., and Doolittle, R. F. (1982) A simple method for displaying thehydropathic character of a protein. J. Mol. Biol. 157, 105–132
14. Zhang, Y., and Barbieri, J. T. (2005) A leucine-rich motif targets Pseu-domonas aeruginosa ExoS within mammalian cells. Infect. Immun. 73,7938–7945
15. Deng, Q., Zhang, Y., and Barbieri, J. T. (2007) Intracellular trafficking ofPseudomonas ExoS, a type III cytotoxin. Traffic 8, 1331–1345
16. Cain, R. J., Hayward, R. D., andKoronakis, V. (2004) The target cell plasmamembrane is a critical interface for Salmonella cell entry effector-hostinterplay.Mol. Microbiol. 54, 887–904
17. Marcus, S. L., Knodler, L. A., and Finlay, B. B. (2002) Salmonella enterica
Membrane Localization Domain of CT694
AUGUST 10, 2012 • VOLUME 287 • NUMBER 33 JOURNAL OF BIOLOGICAL CHEMISTRY 28085
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

serovar Typhimurium effector SigD/SopB is membrane-associated andubiquitinated inside host cells. Cell. Microbiol. 4, 435–446
18. Hower, S.,Wolf, K., and Fields, K. A. (2009) Evidence that CT694 is a novelChlamydia trachomatis T3S substrate capable of functioning during in-vasion or early cycle development.Mol. Microbiol. 72, 1423–1437
19. Clifton, D. R., Fields, K. A., Grieshaber, S. S., Dooley, C. A., Fischer, E. R.,Mead, D. J., Carabeo, R. A., and Hackstadt, T. (2004) A chlamydial type IIItranslocated protein is tyrosinephosphorylated at the site of entry andassociated with recruitment of actin. Proc. Natl. Acad. Sci. U.S.A. 101,10166–10171
20. Benaud, C., Gentil, B. J., Assard, N., Court, M., Garin, J., Delphin, C., andBaudier, J. (2004) AHNAK interaction with the annexin 2/S100A10 com-plex regulates cell membrane cytoarchitecture. J. Cell Biol. 164, 133–144
21. Lee, I. H., You, J. O., Ha, K. S., Bae, D. S., Suh, P. G., Rhee, S. G., and Bae,Y. S. (2004) AHNAK-mediated activation of phospholipase C-�1 throughprotein kinase C. J. Biol. Chem. 279, 26645–26653
22. Caldwell, H. D., Kromhout, J., and Schachter, J. (1981) Purification andpartial characterization of the major outer membrane protein of Chla-mydia trachomatis. (2011) Infect. Immun. 31, 1161–1176
23. Hackstadt, T., Rockey, D. D., Heinzen, R. A., and Scidmore, M. A. (1996)Chlamydia trachomatis interrupts an exocytic pathway to acquire (2011)EMBO J. 15, 964–977
24. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1982)Molecular Cloning. ALaboratory Manual, Cold Spring Harbor, NY
25. Chellas-Géry, B., Wolf, K., Tisoncik, J., Hackstadt, T., and Fields, K. A.(2011) Biochemical and localization analyses of putative type III secretiontranslocator proteins CopB and CopB2 of Chlamydia trachomatis revealsignificant distinctions. Infect. Immun. 79, 3036–3045
26. Bordier, C. (1981) Phase separation of integral membrane proteins in Tri-ton X-114 solution. J. Biol. Chem. 256, 1604–1607
27. Hoffman, K., and Stoffel, W. (1993) TMbase-A database of membranespanning protein segments. (2011) J. Biol. Chem. 374
28. Zhang, Y., Deng, Q., Porath, J. A., Williams, C. L., Pederson-Gulrud, K. J.,and Barbieri, J. T. (2007) Plasma membrane localization affects theRhoGAP specificity of Pseudomonas ExoS. Cell. Microbiol. 9, 2192–2201
29. Isaksson, E. L., Aili, M., Fahlgren, A., Carlsson, S. E., Rosqvist, R., andWolf-Watz, H. (2009) The membrane localization domain is required forintracellular localization and autoregulation of YopE in Yersinia tubercu-losis. (2011) Infect. Immun. 77, 4740–4749
30. Black, D. S., and Bliska, J. B. (2000) The RhoGAP activity of the Yersiniapseudotuberculosis cytotoxinYopE is required for antiphagocytic function
and virulence.Mol. Microbiol. 37, 515–52731. Evdokimov, A. G., Tropea, J. E., Routzahn, K. M., andWaugh, D. S. (2002)
Crystal structure of the Yersinia pestis GTPase activator YopE. ProteinScience 11, 401–408
32. Pederson, K. J., Krall, R., Riese, M. J., and Barbieri, J. T. (2002) Intracellularlocalization modulates targeting of ExoS, a type III cytotoxin, to eukary-otic signaling proteins.Mol. Microbiol. 46, 1381–1390
33. Hueck, C. J. (1998) Type III protein secretion systems in bacterial patho-gens of animals and plants.MMBR 62, 379–433
34. Fields, K. A., Fischer, E. R., Mead, D. J., and Hackstadt, T. (2005) Analysisof putative Chlamydia trachomatis chaperones Scc2 and Scc3 and theiruse in the identification of type III secretion substrates. J. Bacteriol. 187,6466–6478
35. Håkansson, S., Galyov, E. E., Rosqvist, R., and Wolf-Watz, H. (1996) TheYersiniaYpkASer/Thr kinase is translocated and subsequently targeted tothe inner surface of the HeLa cell plasma membrane.Mol. Microbiol. 20,593–603
36. Frithz-Lindsten, E., Du, Y., Rosqvist, R., and Forsberg, A. (1997) Intracel-lular targeting of exoenzyme S of Pseudomonas aeruginosa via type III-de-pendent translocation induces phagocytosis resistance, cytotoxicity anddisruption of actin microfilaments.Mol. Microbiol. 25, 1125–1139
37. Mounier, J., Popoff, M. R., Enninga, J., Frame, M. C., Sansonetti, P. J., andVan Nhieu, G. T. (2009) The IpaC carboxyterminal effector domain me-diates Src-dependent actin polymerization during Shigella invasion of ep-ithelial cells. PLoS Pathog. 5, e1000271
38. Fu, Y., and Galán, J. E. (1999) A Salmonella protein antagonizes Rac-1 andCdc42 to mediate host cell recovery after bacterial invasion. Nature 401,293–297
39. Rosqvist, R., Forsberg, A., and Wolf-Watz, H. (1991) Intracellular target-ing of the Yersinia YopE cytotoxin in mammalian cells induces actin mi-crofilament disruption. Infect. Immun. 59, 4562–4569
40. McGuffie, E. M., Fraylick, J. E., Hazen-Martin, D. J., Vincent, T. S., andOlson, J. C. (1999) Differential sensitivity of human epithelial cells to Pseu-domonas aeruginosa exoenzyme S. Infect. Immun. 67, 3494–3503
41. Carabeo, R. A., Grieshaber, S. S., Hasenkrug, A., Dooley, C., and Hack-stadt, T. (2004) Requirement for the RacGTPase inChlamydia trachoma-tis invasion of non-phagocytic cells. Traffic 5, 418–425
42. Zhou,D., andGalán, J. (2001) Salmonella entry into host cells. Thework inconcert of type III secreted effector proteins. Microbes. Infect. 3,1293–1298
Membrane Localization Domain of CT694
28086 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 287 • NUMBER 33 • AUGUST 10, 2012
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Holly D. Bullock, Suzanne Hower and Kenneth A. Fieldsthe Membrane-localized Family of Type III Effector Proteins
CT694 Protein Belongs toChlamydia trachomatisDomain Analyses Reveal That
doi: 10.1074/jbc.M112.386904 originally published online June 18, 20122012, 287:28078-28086.J. Biol. Chem.
10.1074/jbc.M112.386904Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
Supplemental material:
http://www.jbc.org/content/suppl/2012/06/18/M112.386904.DC1
http://www.jbc.org/content/287/33/28078.full.html#ref-list-1
This article cites 39 references, 15 of which can be accessed free at
by guest on May 12, 2018
http://ww
w.jbc.org/
Dow
nloaded from