do ruffed lemurs form a hybrid zone? distribution and
TRANSCRIPT

Copyright q American Museum of Natural History 2002 ISSN 0003-0082
P U B L I S H E D B Y T H E A M E R I C A N M U S E U M O F N AT U R A L H I S T O RY
CENTRAL PARK WEST AT 79TH STREET, NEW YORK, NY 10024
Number 3376, 26 pp., 4 figures, 2 tables Month 00, 2002
Do Ruffed Lemurs Form a Hybrid Zone?Distribution and Discovery of Varecia, withSystematic and Conservation Implications
NATALIE VASEY1 AND IAN TATTERSALL2
ABSTRACT
Since their discovery by Western explorers traveling to Madagascar in the 17th century, theruffed lemurs have undergone numerous taxonomic revisions. During the 19th and 20th cen-turies, it was intermittently suggested that black-and-white and red ruffed lemurs hybridize innature. Despite the fact that a natural hybrid zone has never been documented, this suggestionhas played a large role in designating the two forms as subspecies of the single species Vareciavariegata. Through a review and synthesis of historical documents, taxonomic literature, mu-seum collections, menagerie and zoo records, recent survey work, genetic data, and vocali-zation data, we examine the evidence for a natural hybrid zone and suggest taxonomic revi-sions. Our work indicates a more extensive hybrid zone than previously suggested—but onein which hybridization is the exception rather than the rule. Furthermore, our findings warrantupgrading the black-and-white ruffed lemur and the red ruffed lemur from subspecies to fullspecies, Varecia variegata (Kerr, 1792) and Varecia rubra (E. Geoffroy, 1812). Our resultssupport the current captive breeding practices of U.S. and European zoos participating in theruffed lemur ‘‘Species Survival Plan’’ and the ‘‘European Endangered Species Programme’’.Lastly, and possibly most importantly, we can now set specific geographic priorities for con-serving the habitat of these highly endangered lemurs in northern Madagascar.
1 Department of Anatomy, College of Medicine, Howard University, 520 W Street NW, Washington, D.C. 20059;now at Anthropology Department, Portland State University, P.O. Box 751, Portland, Oregon 97207. e-mail:[email protected].
2 Curator, Department of Anthropology, American Museum of Natural History. e-mail: [email protected].

2 NO. 3376AMERICAN MUSEUM NOVITATES
INTRODUCTION
The French explorers Francois Cauche(1651) and Etienne de Flacourt (1658) pro-vided Westerners with the first descriptionsof the Malagasy primate now commonlyknown as the ‘‘variegated’’ or ‘‘black-and-white ruffed’’ lemur. More than a centurylater, these animals were formally namedLemur macaco variegatus by Kerr (1792).A related red-and-black form, Lemur ruber,was named shortly afterward by E. Geoffroy(1812) and now goes by the common nameof ‘‘red ruffed lemur’’. Tremendous vari-ability in coat color and pattern (see fig. 1),the ability of the black-and-white form tohybridize with the red form in captivity, andthe relatively recent discovery of the unusu-al life history traits of both ruffed lemurshave subsequently spurred repeated taxo-nomic revisions. Currently the two colorforms are generally classified as subspeciesof the same species and are united withinthe genus Varecia Gray, 1863 in the strep-sirhine primate family Lemuridae. Theblack-and-white form is usually known asVarecia variegata variegata and the redform as Varecia variegata rubra.
In his pathbreaking review of lemur sys-tematics, Schwarz (1931) alluded to the ex-istence of a natural hybrid zone betweenthe two color forms north of the Bay ofAntongil (northeastern Madagascar; seefigs. 2 and 3), but he provided no directevidence for such a zone. Documentationof such a natural hybrid zone would be ofgreat biological interest in designating geo-graphic priorities for habitat conservation,for captive management, and for lemur sys-tematics generally. However, no recent sur-veys have located such a zone. The balanceof evidence on published record so far restson the somewhat slim foundation of a sin-gle hybrid specimen housed at the Ameri-can Museum of Natural History which wascollected 40 km NW of Maroantsetra bythe Archbold expedition in 1930 (Rand,1936; Tattersall, 1982; Buettner-Januschand Tattersall, 1985), although various sim-ilarly puzzling specimens also exist else-where. In writing a review of the naturalhistory of Varecia, Vasey (in press) wasdisinclined to broach the biological mys-
tery concerning a natural hybrid zone, andinstead we try to solve this problem here.To assess whether a natural hybrid zoneever existed, we undertook a review andsynthesis of historical documents, taxo-nomic literature, museum collections, me-nagerie and zoo records, and recent surveywork. We also summarize recent geneticwork on ruffed lemurs and previously pub-lished work on their vocalizations. Thesereviews suggest that hybridization in thewild probably occurred over a much largerarea than previously thought, but that it isthe exception rather than the rule. Onlyremnants of the hybrid zone habitat as itexisted prior to human interference may re-main, given the subsequent extensive hab-itat loss north of the Bay of Antongil. Al-ternatively, it is possible that a poorly doc-umented variety or subspecies of ruffed le-mur has virtually been lost, and that theso-called hybrids are remnants of this pop-ulation; however, there is little substanti-ating evidence for this. In conservationterms, forest corridors are needed to con-nect Varecia populations across the water-sheds north of the Bay of Antongil. Finally,the results of our review support upgradingboth the black-and-white ruffed lemur andthe red ruffed lemur to full species, Vareciavariegata (Kerr, 1792) and Varecia rubra(E. Geoffroy 1812), as has also been sug-gested by Groves (2001). However, we areunable to follow Groves in promoting thepelage variants of the black-and-white spe-cies to subspecies status.
ABBREVIATIONS
AMNH American Museum of Natural History,New York
BMNH British Museum of Natural History,London
DUPC Duke University Primate Center, Dur-ham, North Carolina
MCZ Museum of Comparative Zoology,Cambridge, Massachusetts
MNHN Museum National d’Histoire Naturelle,Paris
RMNH Rijksmuseum van Natuurlijke Historie,Leiden
USNM United States National Museum(Smithsonian), Washington, D.C.

2002 3VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
NORMATIVE DESCRIPTIONS OFBLACK-AND-WHITE AND RED
RUFFED LEMURS
Varecia is the largest-bodied genus of theprimate family Lemuridae, possessing an es-pecially long, luxuriant coat relative to otherlemurs. Within this genus the sexes are moreor less identical in body size and pelage col-oration. Adult wild Varecia range in bodymass from 2.6 to 4.1 kg (Vasey, in press).Both black-and-white and red forms possessblack fur on the ventrum, tail, inner aspectof limbs, manus, pes, crown, forehead, andface. Surrounding the face and ears is a ruffof long fur: white in black-and-white ruffedlemurs and red or honey-blond in red ruffedlemurs. But consistency in coat color andpattern ends there, particularly in the black-and-white variant: Varecia was aptly namedthe ‘‘variegated’’ lemur. In a review pub-lished two decades ago, one of us (Tattersall,1982) recognized four principal variantsamong the black-and-white ruffed lemurs(shown in schematic form in fig. 1). Threeof these had already been established in themid-nineteenth-century account of I. Geof-froy (1851), who elaborated three varieties ofhis Lemur varius (‘‘varietes’’ a, b, and c).These varieties correspond, respectively, tothe forms represented by Kerr’s Lemur ma-caco variegatus (Kerr, 1792), Osman Hill’sLemur variegatus editorum (Hill, 1953), andSmith’s Prosimia subcincta (Smith, 1833).For other equivalences, see Tattersall (1982),who also divided variety b (editorum) intotwo subvariants (b1 and b2). Figure 1 dis-plays the basic pelage coloration differenceson which the four resulting varieties arebased (see also below).
A thorough examination of almost allavailable museum black-and-white speci-mens indicates the presence of more pelage-color variation than is reflected in these foursimple descriptions: an unpublished reviewby one of us (I.T.) several years ago revealedthat as many as 10 repeated variants couldbe recognized in museum collections. How-ever, especially in view of the regrettably lit-tle that we know of geographic distributions(see fig. 2), an infinite multiplication of for-malized variants hardly seems desirable oreven possible. Examination of the available
evidence of coat pattern among black-and-white ruffed lemurs contradicts the notion ofa simple north–south cline, although themodest trend noted by Petter et al. (1977)and Tattersall (1982) toward a reduction inthe overall amount of black in the pelage infavor of white toward the south does seemto hold in a general way. Nonetheless, thereis some notable geographic randomization ofpelage pattern distribution (see fig. 2 and be-low). Analysis of the matter is complicatedby the fact that while ruffed lemur skins arequite well represented in museum collec-tions, locality records are not. Moreover, re-liable field reports are sparse. However, thereare clearly broad areas within which morethan one variant is to be found (see fig. 2 andbelow). We are thus unable to concur withGroves (2001) that at least three distinct sub-species of Varecia variegata are to be rec-ognized.
To summarize the data presented in figure1, in lighter forms (e.g., the variegata vari-ety), the dorsal fur is white with black re-stricted to the shoulder, the upper arm to theelbow, and front of the thigh (color plate);whereas in darker forms known from thenorth (e.g., the subcincta variety), the dor-sum is mainly black with two variably thickbands of white fur—one band encircles thetorso and sometimes extends longitudinallytoward the nape of the neck, and the otherband extends across the rump, down the pos-terior aspect of the thigh, and onto the lateralsurface of the lower leg. In another variety(editorum), the black shoulder patches ex-tend posteriorly onto the flanks and meetmidsagittally, forming a black mantle acrossthe back and shoulders. These various coatpatterns have been illustrated frequently (seeHistorical Documents, Taxonomy, and Illus-trations). Indistinct white rings on the tail,said to resemble those of the ring-tailed le-mur, have also been noted in museum spec-imens collected by J.-P. Audebert (Handfest,1968, cited in Ceska et al., 1992) and in liv-ing wild animals (Evans et al., 1993–1994).Dark regions are not always pure black, butmay grade into silver, light brown, or darkbrown.
While the variegated coat of the black-and-white ruffed lemur has long beenknown, the pied pelage of the red ruffed le-

4 NO. 3376AMERICAN MUSEUM NOVITATES
Fig. 1. Schematic dorsal view of the four pelage variants recognized here among black-and-whiteruffed lemurs. Demarcation between black and white pelage areas is generally but not invariably sharp;black noses are usually frosted.

2002 5VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
Fig. 2. Map showing the general areas of distribution of black-and-white and red ruffed lemurs. Seefigure 3 for detail of the contact area in the region of Maroantsetra, with field sightings located; referto the key at upper left for symbols (filled symbols are museum records, open ones are field sightings).Hatched areas indicate approximate distribution limits and do not imply continuous distribution withinthe regions indicated. Field identifications indicated here in the black-and-white ruffed lemur range southof the Bay of Antongil are by Andrea Katz (Betampona), Elizabeth Balko (Ranomafana/Kianjavatoregion), and I.T. (extreme south).

6 NO. 3376AMERICAN MUSEUM NOVITATES
mur has not been fully appreciated (Vasey,1997, in press). In this form, dorsal fur variesfrom blood red, to red-orange, to honey-blond. Bands of white, honey-blond, or lightred fur may encircle the base of the tail, tar-sus, metatarsus, or digits. White fur may ap-pear on the muzzle and lateral aspect of theankle, extending up to the knee or thigh; andthe nape of the neck almost always bears alarge patch of white fur (see color plate). Apale line across the nuchal region, tippedwith white laterally, has also been observed.Some of these characteristics were noted byE. Geoffroy (1824a), I. Geoffroy (1851), El-iot (1913), Hill (1953), Petter et al. (1977),Tattersall (1982), Hekkala and Rakonton-dratsima (1999), and were also illustratedearly on (see Historical Documents, Taxon-omy, and Illustrations). Tattersall (1982)found several museum specimens ‘‘muchlighter in color than are typical V. v. rubra’’and likened them to captive-bred hybrids. Itis now clear, however, that lighter colorationby itself (i.e., honey blond vs. dark red) fallswell within the normative description, anddeep within the geographic range, of redruffed lemurs: animals from within a singlecommunity in the Andranobe Forest, for ex-ample, show all of the variation in coat colorand patterning described above, and in par-ticular the honey-blond coloration (colorplate).
Hybrids and putative hybrids (see colorplate) are black-capped and ventrally black,but vary in the proportions of darker fur onthe dorsum. The spectrum generally runsfrom a predominantly rufous hue to a whitishpelage lightly washed with raspberry tones,especially near the shoulders and rump.None, however, possesses the precise pat-terning of coloration typical of either of theparental populations, although in the occa-sional hybrid the principal effect is of blackvs. white, with the addition of some reddishfur.
HISTORICAL DOCUMENTS,TAXONOMY, AND ILLUSTRATIONS
Buffon (1765) provided the first scientificdescription of the black-and-white ruffed le-mur, referring to it simply by the vernacularnames ‘‘vari’’ and ‘‘maki-pie’’. His descrip-
tion and accompanying illustration (Buffon,1765: pl. XXVII) represent what is currentlyreferred to as the variegata variety (seeabove). Almost thirty years later, Kerr (1792)assigned this form the trinomen Lemur ma-caco variegatus,3 referencing the animal de-scribed by Buffon and providing the vernac-ular English names ‘‘pied vari’’ and ‘‘ruffedmaucauco’’. Shortly afterward, E. Geoffroy(1796) introduced le ‘‘vari a ceinture’’ (the‘‘vari’’ with a belt), currently identified as thesubcincta variety after Smith (1833). JeanBaptiste Audebert illustrated both of theseblack-and-white forms in his monograph,Histoire Naturelle des Singes et Makis (Au-debert, 1797, pls. V and VI). Buffon, Kerr,J. B. Audebert, and later E. Geoffroy and(1824b) and Pollen (1868) all recounted thefew details known of these animals in theirwild state that had been given by Flacourt inhis Histoire de la grande Isle Madagascar(1658: 153):
Il y a de diuerses sortes de singes, il y en a de grands,qui sont blancs, & ont des taches noires sur les costez,& sur la teste, ils ont le museau long comme vn re-nard, ils les nomment a Manghabei varicossy4: ceux-cy sont furieux comme des tigres, ils font tel bruitdans les bois, que s’il y en a deux, il semble qu’il yen a vn cent. I’en ay eu deux que ie fis porter dansnostre barque; mais ils se ietterent dans la mer, ilssont tres difficilles a apprivoiser; si on ne les a deieunesse.5
3 Kerr built on a binomen and description publishedearlier by Gmelin—Lemur macaco: corpore nigro andalbo mixto (Systema Naturæ, 13th ed.1788). However,Linnaeus had previously assigned the binomen Lemurmacaco to the black lemur (Systema Naturæ, 12th ed.1766). The latter has long been considered a separatespecies, but in early days it was considered one and thesame species as the black-and-white ruffed lemur untilsufficient numbers of captive animals had passedthrough European menageries so that coat colors ofmales and females could be distinguished and habits re-vealed.
4 Today the Malagasy vernacular ‘‘varicossy’’(5 ‘‘varikosy’’) refers to the white-fronted brown lemur,Eulemur fulvus albifrons, which is found sympatricallywith the ruffed lemur in the northeast of Madagascar.The Malagasy vernaculars for black-and-white and redruffed lemurs are ‘‘varikandana’’ (or ‘‘varikandra’’) and‘‘varinena’’ (or ‘‘varikamena’’), respectively.
5 ‘‘There are several kinds of monkey, there are bigones, which are white, and have black patches on theirribs, and on their heads, they have a long muzzle like afox, at Mangabey they call them varicossy: they are asfierce as tigers, they make so much noise in the trees,that if there are two of them it seems that there are a

2002 7VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
Early in the 19th century, the red form wasnamed Lemur ruber by E.Geoffroy (1812):in the vernacular, ‘‘maki rouge’’. A morethorough description of the red form and il-lustrations of both the red and black-andwhite forms were later published (E. Geof-froy 1824a, 1824b, Livraisons XV andXLIII). Although the botanist Commerson il-lustrated the red form in 1763, his drawingsremained unpublished when they were sentback to Paris after his death shortly thereaf-ter. Only after the naturalist Peron returnedto France from Madagascar with skins (ca.1802), and a merchant boat had brought alive animal back to France from Madagascar,was Commerson’s drawing of the red formunearthed from among his papers and a newspecies named (E. Geoffroy, 1824a, 1824b).The animal illustrated by Geoffroy and Cu-vier (1824a) depicted the seaborne animal,which was then housed at the menagerie inthe Jardin des Plantes at the Museumd’Histoire Naturelle in Paris (now known asthe MNHN). This female possessed a chest-nut-red coat dorsally, with white bands of furaround each tarsus. Strangely, this animalnever vocalized. Presumably this is why herclose relationship to the black-and-whiteform was not yet obvious. Geoffroy and Cu-vier (1824a, 1824b) assumed that, like otherknown lemurs, females and males of the‘‘maki rouge’’ and the ‘‘maki vari’’ wouldhave different coat colors and patterns.
Not surprisingly in light of ongoing broadefforts to name and precisely classify the le-murs during the 19th century, several mile-stones in ruffed lemur systematics would bepassed before 1900. Importantly, however,that scientists in different parts of Europewere sometimes not aware of each other’swork for many years after each had pub-lished. The French savant I. Geoffroy (1851)disentangled the ruffed lemur from the blacklemur (Lemur macaco Linnaeus, 1766, also
hundred. I had two of them brought on our ship; butthey threw themselves in the water, they are very diffi-cult to tame unless you get them when they are young’’.Note that in the original French text, the letter ‘‘j’’ wasprinted as ‘‘i’’, the letter ‘‘v’’ as ‘‘u’’ when in the middleof a word, and the letter ‘‘u’’ as ‘‘v’’ when beginning aword. Two editions of Flacourt were consulted for thistranslation (see references), in addition to Tattersall(1982).
described as Lemur niger by E. Geoffroy,1812) by advancing the species name Lemurvarius to distinguish it; and he also retainedthe species Lemur ruber. He further brieflydescribed varieties a, b, and c of the black-and-white form (which, as noted above, cor-respond to the varieties variegata, editorum,and subcincta, respectively). The English-man Gray (1863) introduced the genus Va-recia for the ruffed lemurs, thereby unitingthe black-and-white form (Varecia varia)and the red form (Varecia rubra) at the ge-neric level using as key characteristics theruff surrounding the head and the tufted ears.This generic distinction was not immediatelyaccepted. Gray continued to include theblack lemurs with the ruffed lemurs (Gray1863, 1870, 1871), yet later questioned hisown taxonomic revisions (Gray, 1872), basedon the observations of several other scientists(Bartlett, 1862; Schlegel, 1866; Sclater,1871). Sclater (1871) correctly observed thatamong black lemurs the sexes were dichro-matic, whereas in black-and-white ruffed le-murs the sexes had similar coat patterns. Itwas also recognized that Lemur varius andLemur niger differed in their ‘‘voices’’ (vo-calizations; Bartlett, 1862; Sclater, 1871).Despite these caveats, Gray’s examination ofspecimens in the BMNH, especially thosecollected by Alfred Crossley in the 1860s,led him to think that Lemur niger, Lemurvarius, and Lemur ruber were ‘‘all one spe-cies, extremely variable in colour, some be-ing black, others red, and others white, andall the intermediate shades and variations’’(Gray, 1872). Schlegel, a museum naturalistin Leiden, and Pollen, a Dutch explorer, drewsimilar conclusions. Noting excessively var-iable coloration Schlegel (1866, 1876) con-sidered Lemur ruber to be nothing more thana variety of Lemur varius, but was also thefirst to remark that Lemur macaco was sex-ually dichromatic, and that compared to Le-mur varius, it was smaller in size, had amuch weaker voice, and a far less thick, tuft-ed, and woolly coat. Having traveled onlythrough the northwest of Madagascar, Pollenrelied on information provided by nativesand by other explorers, and on observationshe made on captive ruffed lemurs at the Mu-see de Saint-Denis (Reunion) where he sawa tricolored animal. Without providing any

8 NO. 3376AMERICAN MUSEUM NOVITATES
sources he drew a rather extreme conclusion,lumping all black, white, red, and black-and-white forms into Lemur varius. He claimed,moreover, that there were no differences incolor by sex or age, and stated that Lemurruber had the same habits as the vari, ‘‘withwhich [presumably in captivity] it lives andmates’’ (Pollen, 1868: 21). So, while somein this period came to understand that ruffedlemurs were not sexually dichromatic, werespecifically distinct from black lemurs, andconsisted of two distinct species (e.g., I.Geoffroy, 1851)—and perhaps even of aunique genus (Gray, 1863)—a polar viewhad also emerged: that animals of all colorsand coat patterns belonged to one single spe-cies.
Schwarz (1931) attempted to sort I. Geof-froy’s (1851) three varieties geographically,but acknowledged that many original muse-um labels had disappeared and that he hadencountered much difficulty in identifyinggeographical names on labels, in literature,and on older maps (Schwarz, 1931: 399–400). Schwarz unequivocally stated, howev-er, that the red form was a color mutation ofI. Geoffroy’s variety a, ‘‘with which it occursand interbreeds’’. The only evidence cited insupport of this claim was that crosses bornat the Berlin Zoo resembled some of A.Milne-Edwards and A. Grandidier’s (1890)illustrations (see color plate), which were,unfortunately, published without descriptionsor subspecific designations.
Hill (1953) drew directly upon Schwarz(1931) with regard to taxonomy and distri-bution. He upgraded I. Geoffroy’s varietiesa, b, and c to the subspecies Lemur varie-gatus variegatus (Kerr, 1792), Lemur varie-gatus editorum Hill, 1953, and Lemur var-iegatus subcincta (Smith, 1833), respective-ly. He produced a distribution map for thesubspecies, taking considerable license inbuilding on the information provided bySchwarz (1931). This map contains gross er-rors relative to distributional informationknown both at that time and currently (cf.Pollen, 1868; Jentink, 1892; Elliot, 1913;Kaudern, 19156). Hill (1953: 400) further
6 Pollen, collating information from indirect sources,stated that Lemur varius (both forms) occurs in the re-gion between Tintingue, Tamatave, and Antananarivo.
embellished on Schwarz (1931) by statingthat the red form and variety a (L. v. varie-gatus) occur together geographically and in-terbreed in nature. Unfortunately, Hill pro-vided no documentation for this claim otherthan to cite Schwarz’s observations that hy-brids were produced at the Berlin Zoo, andthat the hybrids in turn resembled A. Milne-Edwards and A. Grandidier’s (1890) plates.Consequently, the idea (but no documenta-tion) of a natural hybrid zone emerged in tan-dem with a notion (albeit misconstrued) ofthe geographical distribution of the red andblack-and-white forms.
Thus, by the mid-20th century, notions re-ported by Pollen (1868) as unverified claimshad become highly speculative ‘‘facts’’. Butone last other clue from the 19th century wasyet to be seized upon. Schwarz (1931) notedthat ‘‘black-and-white and red specimenshave been collected in the same place byJ.[-P.] Audebert in the coast region north ofthe Bay of Antongil’’, citing Jentink of Lei-den (Catalogue Systematique des Mammifer-es, 1892). Schwarz was alluding to the col-lections made by the German-born Josef-Pe-ter Audebert, who worked in Madagascar be-tween 1876 and 1879. His collection was
Jentink, using J.-P. Audebert’s collection records, dem-onstrated that L. varius occurred north of the Bay ofAntongil and south as far as Mananare, Vidoutra, andMahambo. Elliot indicated that the black-and-whiteform occurs in northeastern Madagacar from Adenponeto Cape Masoala at the entrance of Antongil Bay, andinto the interior to Bengoa; and that the red form occursin eastern Madagascar, from the Bay of Antongil in thenorth to Masindrano in the south. Many of the placenames cited by Elliot cannot be located (presumably hedrew these localities from museum labels). His distri-bution for the red form more accurately reflects the dis-tribution for the black-and-white form, assuming Mas-indrano refers to the town bearing this name in south-eastern Madagascar, not the village bearing this namejust south of Mahalevona on the Bay of Antongil. Kau-dern described a subcincta-like animal from one daywest of Fenerive (‘‘has only a white band around itsbody’’, Kaudern, 1915, pl. 3, fig. 1), and an editorum-like animal from south of Tamatave (‘‘almost all whitein the rear’’). Despite the availability of this distribu-tional information, Hill (1953: 401, Fig. 130) illustratedthe range of L. variegatus as extending only as far southas Tamatave, but well into the northern high plateau andall along the northern coast as far as Diego Suarez! Hedid not take the records from south of Tamatave, Anta-nanarivo, and possibly Masindrano into account, where-as the distribution he gave for ruffed lemurs in northernMadagascar is hugely exaggerated.

2002 9VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
sold to various museums, but most of hisruffed lemurs went to the RMNH in Leidenand were well documented by Jentink(1892). It is important to remember that nei-ther Schwarz nor Hill had examined the Lei-den material firsthand, and more will be saidof this in the section below on Museum Col-lections and Records.
Fieldwork and careful study of museumskins and their provenances by Petter et al.(1977) and Tattersall (1982) improved ourunderstanding of the taxonomy and distri-bution of ruffed lemurs. However, these stud-ies by no means resulted in a tidy, discretedistribution of the black-and-white forms.Considering cranial anatomy, social behav-ior, and the unusual reproductive traits ofruffed lemurs, Petter (1962) reintroduced thegenus Varecia Gray, 1863 to distinguishruffed lemurs from other members of thefamily Lemuridae. Petter et al. (1977) de-scribed and provided locality data, to the ex-tent possible, for seven black-and-whiteforms. These authors retained the subspeciesadvanced by Hill in 1953 (V. v. variegata, V.v. editorum, and V. v. subcincta) and furtherdescribed, but did not name, another four.The southernmost population then known,inhabiting the remaining coastal forest southof Farafangana, was said to be almost entire-ly white dorsally. Like various earlier work-ers, Petter et al. (1977) viewed the red formas simply another subspecies of ruffed le-mur—V. variegata rubra. However, unlikeearlier workers, these authors indicated thatthe Antainambalana River (fig. 3) clearlyseparates the red form from the black-and-white form, specifically V. v. subcincta7. Thisis the first reference to a geographic dividebetween the two color forms.8 Tattersall(1982) recognized just two subspecies, cor-responding to the red form (V. v. rubra) onthe one hand, and to all black-and-whiteforms combined (V. v. variegata) on the oth-er. Building on Geoffroy (1851), he de-scribed four black-and-white types (a, b1, b2,
7 Black-and-white ruffed lemurs (subcincta type) wereobserved at 400 m in the hills to the west of the An-tainambalana in 1969 by Petter et al. (1977: 225).
8 Hill (1953) produced a distribution map showing thisdivide as well, but he provided neither provenance datanor details of the specimens upon which the map wasbased.
c) and indicated that better knowledge of thedistribution of these varieties might ultimate-ly warrant their recognition as distinct sub-species. Unfortunately, the key for this dis-tribution map contained drafting errors; wereproduce it correctly in the footnote below9.Groves (2001) raised the variants of theblack-and-white forms to subspecies of thespecies Varecia variegata.
The new distribution data presented byPetter et al. (1977) and Tattersall (1982) con-flict in one particularly important way withthose published earlier by Schwarz (1931)and Hill (1953). The latter authors placed thelightest form (type a, variegata) farthestnorth, interbreeding with the red form northof the Bay of Antongil, whereas the formerdetermined that it was the darkest form (typec, subcincta) that occurred farthest north,largely separated from the red form by theAntainambalana River. Despite the discoveryof a geographic divide, however, the idea ofa natural hybrid zone reemerged. Based onreports of a rufous brown variant west of theBay of Antongil, Tattersall (1977) suggestedsuch animals might be hybrids or a localform of V. v. variegata. Later it was notedthat several museum specimens are muchlighter in color than are typical V. v. rubraand resemble 50/50 V. v. rubra 3 V. v. var-iegata hybrids bred in captivity at the DUPC(Tattersall, 1982; Buettner-Janusch and Tat-tersall, 1985). These authors thought at thattime that only one such specimen was bothwild-caught and documented to locality. Thiswas collected by the Archbold expedition in1930, 40 km NW of Maroantsetra up theVohimaro River (Rand, 1936). On this basis,Tattersall (1982) suggested that V. v. varie-gata and V. v. rubra might be in secondarycontact between the confluent Vohimaro andAntainambalana Rivers. This suggestionprompted several surveys, the findings ofwhich are reviewed below (see Recent Sur-veys).
In concluding this section it is appropriateto summarize the basis upon which a naturalhybrid zone was proposed. The black-and-
9 The correct correspondence between V. variegatavariegata types and symbols in figure 3.15 of Tattersall(1982: 72) is as follows: type a (variegata) m; type b1(editorum) ,; type b2 (editorum) V; type c (subcincta)M.

10 NO. 3376AMERICAN MUSEUM NOVITATES
Fig. 3. Masoala Peninsula and region north of the Bay of Antongil and its major watersheds. Arrows(→) indicate the Vohimaro, Antainambalana, Andranofotsy, and Mahalevona rivers. Triangles (m) showlocalities where hybrid ruffed lemurs were collected (see table 1). Large dots (v) show localities wheremore than one form of ruffed lemur was collected and/or sighted (see table 2). Stippling represents areaof overlap between black-and-white and red ruffed lemurs where hybridization has occasionally oc-curred. Small dots (●) show other localities mentioned in the text. Andaparaty: black-and-white ruffedlemurs sighted; Ankorongana: red ruffed lemurs reported to the north (Ambodibilahy Mtn.) and to theeast (Anjanaharibe Mtn.). At nearby Sahantaha village to the south, black-and-white ruffed lemurs keptas pets were apparently captured on the west side of the Antainambalana River. Andranobe Forest: redruffed lemurs at this site show all the pelage variations described under the normative description.Samples used in genetic analyses were collected from this population (Vasey, 1997). Dashed linesindicate boundaries of the Masoala National Park. Note several narrow corridors bridging forestedblocks. Coordinates for important localities during recent surveys: Andaparaty village 158129S, 498379E;

2002 11VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
←
Ankorongana village 1581097860S, 4983594580E; Sahantaha village 1581193000S, 4983499000E; Sahavaryvillage 158199S, 498509E; Belampona River 1581898480S, 4985495080E (see text for references). Amboh-imarahavavy is an Archbold Expedition collecting locality (‘‘2 days NE of Maroantsetra’’) wherered ruffed lemurs were collected. Other important coordinates: Andranobe field site 1584099060S,4985793990E; Ambohimarahavavy 1581690200S, 5080390000E; Bevato 1581097400S, 4982891400E.
white and red forms were considered differ-ent species throughout most of the 19th cen-tury (E. Geoffroy, 1812; E. Geoffroy, 1824a,1824b; Lesson, 1840, 1842; I. Geoffroy,1851; Gray 1863, 1870), but by the end ofthe 19th century and throughout the 20th vir-tually all systematists classified them as sub-species or varieties of a single highly vari-able species (Schlegel, 1866, 1876; Gray,1872; Forbes, 1894; Elliot, 191310; Schwarz,1931; G. Grandidier and Petit, 1932; Hill,1953; Petter et al., 1977; Tattersall, 1982).While Schwarz (1931) and Hill (1953) bothalluded to the existence of a hybrid zone, nei-ther author documented any wild-caught hy-brids in museum holdings. Rather, their worksuggests that the gradual taxonomic down-grading of black-and-white and red ruffed le-murs, from species to subspecies, had beeninfluenced largely by the existence of hybridsthat had been produced in captivity. For, asevidence for a hybrid zone, they were ableonly to offer captive-bred crosses and theplates of A. Milne-Edwards and A. Grandi-dier (1890; see also color plate).
This observation prompts three questions.What did the oldest illustrations of ruffed le-murs look like? Were there any hybridsamong them? And when were hybrids firstproduced in menageries and zoos? In answerto the first two questions, until we arrive atthe plates published by A. Milne-Edwardsand A. Grandidier (1890), all early illustra-tions of ruffed lemurs are true forms, eitherred or black-and-white (Commerson, 1763unpubl.; Buffon, 1765; Schreber, 1775; J. B.Audebert, 1797; E. Geoffroy 1824a, 1824b;Gervais, 1854). A. Milne-Edwards and A.Grandidier (1890) illustrated three black-and-white forms corresponding to Geoffroy’s
10 Elliot (1913) was the first author in 121 years prop-erly to attribute the species name Lemur variegatus toKerr (1792). Along with Forbes (1894) he was first toapply the now commonly used English vernaculars,‘‘ruffed lemur’’ and ‘‘red ruffed lemur’’.
varieties a, b and c (pls. 123, 124, 125), twored forms falling within the normative de-scription (pls. 12711 and 128), and one pos-sible hybrid (pl. 126; see color plate herein).Unfortunately these plates were publishedwithout descriptions, taxonomic assignments,or references as to where the illustrated ani-mals originated. It is possible that these il-lustrations were based on wild (or wild-caught) animals, as A. Grandidier undertookdetailed surveys in Madagascar and crossedthe island in several directions between 1865and 1870. However, it cannot be ruled outthat the hybrid depicted was produced in cap-tivity. And documentation of a hybrid zonerequires more than zoo-bred hybrids and un-annotated illustrations. It requires either theidentification of such a zone in the wild, orprecise locality documentation of wild-caught hybrids in existing museum holdings.We next turn our attention to museum col-lections before reviewing menagerie and zoorecords, recent survey work, genetics, andvocalizations.
MUSEUM COLLECTIONS ANDRECORDS
The above review suggests that collectionsmade in northeastern Madagascar by J.-P.Audebert between 1876 and 1879, and by theMission Zoologique Franco-Anglo-Ameri-caine a Madagascar 1929–1931 (commonlyreferred to by Anglophones as the Archboldexpedition), hold promise in documentingwild-caught hybrids and/or regions of over-lap between the black-and-white and redforms of Varecia. We have studied these col-lections, in addition to many others, and werefer largely to published work of one of us
11 Schwarz (1931: 418) considered plates 126 and 127of Milne-Edwards and A. Grandidier (1890) to representhybrids. However, upon examining these plates, one ofus (N.V.) has determined that plate 127 lies within thenormative description of the red ruffed lemur.

12 NO. 3376AMERICAN MUSEUM NOVITATES

2002 13VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
←
COLOR PLATE. A, Black-and-white ruffed lemur, variegata variety (by D. Haring, DUPC). B, Redruffed lemur, honey-blonde variety from Andranobe River, Madagascar (by N. Vasey). C, Putative hybridin Plate 126 of Milne-Edwards and Grandidier (1890); courtesy of AMNH Library. D, Red ruffed lemur(by D. Haring, DUPC). See Normative Description section for further details.
(Tattersall, 1982; Buettner-Janusch and Tat-tersall, 1985) and to his museum notes in oursearch for undocumented hybrids. In addi-tion, Jentink’s Catalogue Systematique desMammiferes from Leiden (1892), Rand’s de-tailed notes on collection localities of theArchbold Expedition (Rand, 1932, 1936),and the Catalogue of Primates in the BritishMuseum (Natural History) Part IV (Jenkins,1987) have been consulted, the last of theseproviding detailed information on Archboldmaterial accessioned into the BMNH in ad-dition to the field listings held by the De-partment of Mammalogy at the AMNH. Wehave also studied the holdings of the USNM,which include the excellent Abbott Collec-tion, to improve upon the normative descrip-tions provided above and to search for ruffedlemur specimens collected in northern Mad-agascar. Topographic maps issued by the In-stitut Cartographique de Madagascar (FTM)at 1:100,000 scale, and several geo-referenc-ing databases have been consulted to verifylocalities, place names, and topographic fea-tures. The latter resources include the UnitedStates Board on Geographic Names Gazet-teer (USGS), the Madagascar Gazetteer(Missouri Botanical Garden), and, especially,the Platform d’Analyse Project for the Ma-soala Region, Madagascar (data courtesy ofDavid Lees).
Three possible wild-caught hybrids withprovenance data were located for this study,one each in museum holdings at the RMNH(Lemur varius w, aka ‘‘Lemur ruber Cr. g duCat. Ost.’’ in Jentink’s 1892 catalog), theBMNH (ZD.1935.1.8.29), and the AMNH(100510). Of these, only the last had beenpublished as a hybrid (Buettner-Janusch andTattersall, 1985). Table 1 provides detailedinformation on these specimens and on twomore for which there are no provenance data.Although Plate 126 (see color plate) of A.Milne-Edwards and A. Grandidier (1890)suggests that hybrids might have been de-posited in the MNHN, we could not find any
there. The three wild-caught hybrids origi-nate from 40 km NW of Maroantsetra alongthe Vohimaro River (Bevato village) andfrom the environs of Mahalevona village (ta-ble 1, fig. 3). These provenance records sug-gest that ruffed lemurs may have hybridizedover a much larger area than previously sug-gested: one extending across three majordrainage systems that empty south into theBay of Antongil. From east to west, these arethe valleys of the Mahalevona, the Andran-ofotsy, and the Antainambalana Rivers, theVohimaro River being a major tributary ofthe last of these.
Currently, the RMNH hybrid is catalogedas Lemur ruber (Jentink, 1892). It comesfrom ‘‘Malewo’’ (old orthography for Ma-halevona) and falls outside the range of var-iation seen in either red ruffed lemurs gen-erally, or in comparison to other specimenscollected in the same location. It is a light-colored specimen with many white hairs inthe dorsal fur, giving it a ‘‘washed-out’’ look.The AMNH hybrid (100510) and the BMNHhybrid (ZD.1935.1.8.29) are cataloged underV. v. rubra (Buettner-Janusch and Tattersall,1985; Jenkins, 1987) and both come from40 km NW of Maroantsetra. BMNH1935.1.8.29 is described as ‘‘light coloured’’(Jenkins, 1987), with the top of the head andcheeks black; the ventrum, extremities, andtail black; and the rest of the dorsum whitishwith reddish elements, especially posteriorly.According to our notes, BMNH 1935.1.8.29resembles AMNH 100510. This AMNHspecimen is whitish, with black headcap,ventrum and extremities. Its tail is very darkexcept at the proximal base, where it lightensup for about one-fourth of its length. Thereare red-brown patches bilaterally at the topof the forelimbs, and there are hints of red-dish fur in the center of the dorsum and atthe sides posteriorly. AMNH 100510 haspreviously been described as similar to V. v.variegata 3 V. v. rubra crosses from theDUPC (Buettner-Janusch and Tattersall,

14 NO. 3376AMERICAN MUSEUM NOVITATES
TABLE 1Wild-Caught Hybrids of Varecia in Museum Collectionsa
1985). These wild hybrids resemble captivehybrids (color plate). Two specimens of un-known provenance from Ward’s Natural His-tory Establishment, AMNH 267 and 268,generally resemble the BMNH and AMNHhybrids just described. They have light fur,dark headcaps, white necks and most of thedorsum white, with faded reddish patches onthe upper forelimbs and extending across theback. There are also darker patches on theinsides of the upper legs and lighter reddishpatches on the lower back, at the sides. Theirtails are very dark.
Schwarz’s indication that black-and-whiteand red forms were collected in the sameplace (Maroantsetra) by J.-P. Audebert fueledthe provocative suggestion that a natural hy-brid zone existed (e.g., Simons and Lindsay,1987). Our review has revealed three, andpossibly four, localities where more than oneform was found, demonstrating that overlapoccurred over a considerably larger area thanpreviously recognized (table 2, fig. 3). In the19th century, Alfred Crossley and J.-P. Au-debert collected black-and-white and redforms in the environs of Maroantsetra andJ.-P. Audebert collected red forms and onehybrid in the environs of Mahalevona vil-lage. In May 1930, the Archbold Expeditioncollected black-and-white forms and two hy-brids 40 km NW of Maraoantsetra near Be-
vato village. In the Andranofotsy valley, nearthe village of Savary, J.-P. Audebert collect-ed black-and-white forms in 1878; in a recentsurvey, red ruffed lemurs were sighted nearthis village (see Recent Surveys). However,another village along the AntainambalanaRiver (north of the confluence with the Voh-imaro) bears a similar place name (Saha-fary), so this co-occurrence must remainspeculative. These regions of overlap, espe-cially those where hybrids occur, buttress theclaim for a natural hybrid zone. However, de-spite documenting a more expansive hybridzone and several regions of overlap, museumholdings turned up only three documentedhybrids. This being the case, hybridizationappears to be the exception rather than therule in the overlap zone of red and black-and-white ruffed lemurs.
MENAGERIE AND ZOO RECORDS
Those managing captive ruffed lemurbreeding programs are well aware that black-and-white and red ruffed lemurs form hy-brids readily in captivity, as do many othercongeneric lemur taxa (Petter, 1969). Be-cause the red ruffed lemur is rare in zooholdings, it was commonly crossed withblack-and-white ruffed lemurs, notably inEuropean zoos, and these crosses have been
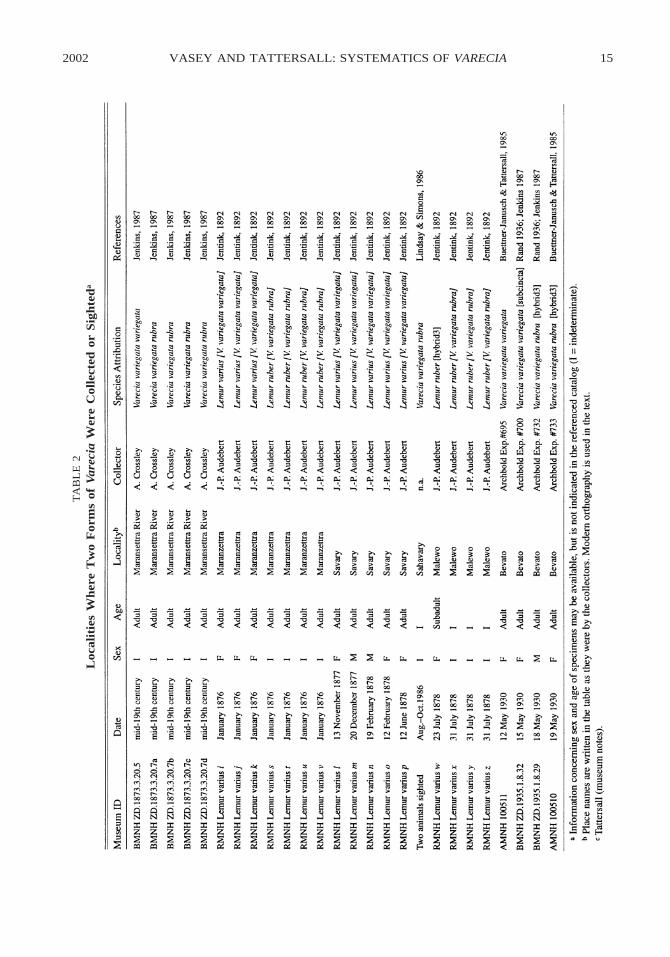
2002 15VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
TA
BL
E2
Loc
alit
ies
Wh
ere
Tw
oF
orm
sof
Var
ecia
Wer
eC
olle
cted
orS
igh
ted
a

16 NO. 3376AMERICAN MUSEUM NOVITATES
referred to as ‘‘tri-colored ruffed lemurs’’(Bernirschke and Miller, 1981). This breed-ing practice, however, has been discouragedfor over two decades (e.g., Lindsay, 1977),and in particular by the Species SurvivalPlan (SSP, a branch of the American Zoo andAquarium Association), by the European En-dangered Species Programme (EEP), and bythe Prosimian Taxon Advisory Group (abranch of the World Conservation Union,formerly the IUCN). This returns us to thequestion posed at the end of the first section(Historical Documents, Taxonomy, and Illus-trations) regarding when hybrids were firstproduced in menageries and zoos. Since ourreview of museum collections and recordsplaces discussion of a natural hybrid zone onmuch firmer footing, why should one botherto determine the earliest occurrence of cap-tive hybrids? This effort remains importantbecause it affects ruffed lemur taxonomyand, in turn, conservation initiatives. The de-cision to lump the black-and-white and redforms into one species was initially made onthe basis of their excessively variable coatcoloration, without recognizing that some ofthe variation in the available sample mighthave been induced through captive hybrid-ization (e.g., Pollen, 1868). Lumping wascompleted before J.-P. Audebert and theArchbold Expedition brought back naturalhybrids in the late 19th and early 20th cen-turies, and long before the distinctive geo-graphic distributions of the two color formswere known (Petter et al., 1977; Tattersall,1977; see Survey Work). These facts suggestthat the down-ranking of black-and-whiteand red ruffed lemurs to subspecies warrantsreconsideration, especially if it can be dem-onstrated that their down-ranking was largelyinfluenced by captive hybridization duringthe 19th century. This section is not intendedto be an exhaustive review of captive breed-ing in ruffed lemurs, but rather to reveal theearliest records of captive breeding practicesand to substantiate whether captive-bred hy-brids had undue influence in collapsing thespecific distinction between black-and-whiteand red ruffed lemurs.
Although lemurs were already kept in Eu-ropean menageries by the 18th century, turn-over was high and captive lives short: mates
were rarely available12 and illness went un-diagnosed or untreated. Lemurs hardly faredbetter during the first part of the 19th cen-tury. One of the first references to a captiveblack-and-white ruffed lemur is found inKerr (1792: 87): ‘‘The vari is fond of sun-shine, yet always chooses to sleep in a darkplace, and preserves its nest very clean. Inconfinement, it refuses to feed on eggs, flesh,or fish; and, when eating, growls, with avoice like that of a lion’’. The Empress Jo-sephine Bonaparte kept a pair of black-and-white ruffed lemurs at Malmaison that werepresented to the Museum d’Histoire Naturel-le in Paris in 1809, presumably upon theirdeath. This pair mated and produced triplets,which did not survive (I. Geoffroy, 1851).The first red ruffed lemur brought live to Eu-rope was mute and lived alone at the menag-erie in the Jardin des Plantes in Paris ca.1820, as did a second animal circa 1839. Fewprimates reproduced in the Paris menageriebetween 1793 and 1851, and none that didwere lemurs, despite the reception of 87strepsirhines during that period (I. Geoffroy,1851).
In the second half of the 19th century andduring the early 20th century, life in menag-eries took something of a turn for the better.In an attack on Darwin’s Origin of Species,Gray inadvertently provided the most tellinginformation regarding 19th century menag-eries and hybridization:
All the specimens that have come under my obser-vation have been living in menageries; and all theskins in the Museum are obtained from specimenswhich have been so confined; and some of them havebeen even born in confinement, and are probably thehybrid offspring of two species, arising from the in-termixture of different kinds in the same cages. Undersuch circumstances, it is very natural that thereshould be difficulties in separating them, and thatthere may be intermediate forms. Yet I may state that,when the specimens which have come under my ex-amination have been carefully compared, I have hadno difficulty in distinguishing them, and I have notfound a single specimen which I have had the slight-est reason to believe is a passage from one species tothe other (Gray, 1863: 134, cf. Gray, 1872).
Pollen’s (1868) somewhat oblique reference
12 Regarding his pet ‘‘mongous’’ (brown lemur), Buf-fon (1765: 177) wrote: ‘‘il cherchoit les chattes, & memese satisfaisoit avec elles, mais sans accouplement intime& sans production’’.

2002 17VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
to captive breeding and hybridization ofruffed lemurs at the Musee de St. Denis wasfollowed by much more substantial records.Between 1845 and 1932, 66 lemur birthswere recorded in the London Zoo: eight werehybrids (two from before 1867), althoughnone was a ruffed lemur (Zuckerman, 1932).Between 1899 and 1922, 120 lemurs wereborn at the Giza Zoological Gardens inEgypt, of which seven were black-and-whiteruffed lemurs (Flower, 1933). By 1935, De-chambre of the Paris menagerie had achievedmany successes in captive care and breedingof lemurs. He had produced three-generationlineages of five lemur species (including Le-mur variegatus) and had defeated severaltypes of illness in the lemur colony throughimprovements in diet and housing. Decham-bre’s success (Dechambre, 1935) was re-warded in that he was able to give the firstdetailed descriptions of Varecia‘s unique re-productive pattern.
The documents at hand indicate that cap-tive hybridization of lemurs occurred before1870. Thus, it is highly likely that the taxo-nomic revisions of Varecia in the later 19thcentury were influenced by such practices,whether intentional or not. More recently, le-mur subspecies, and even separate speciesand genera, have intentionally been hybrid-ized as a means of clarifying their taxonomy(e.g., A. P. Gray, 1954; Petter, 1969). In thisregard, Petter et al. (1977: 257) obtained ahybrid between a female V. v. rubra and amale V. v. subcincta. Although the ruffed le-mur captive breeding program (established inthe late 1960s) discourages interbreedingamong the two color forms, hybrids are stilloccasionally produced (I. Porton, personalcommun.), and crossings have been mostcommon in European zoos (Bernirschke andMiller, 1981). Illustrations of pelage colorand pattern based in any part on captive-bredanimals with histories of hybridization (aliveor in museum holdings) may complicaterather than clarify ruffed lemur taxonomy be-cause some of the variation illustrated maynot represent natural variation or natural hy-brids (see, e.g., Ceska et al., 1992, pls. 1–4).It is also true that some unusual wild-col-lected forms have not yet been illustrated(e.g., USNM 84381, which combines edito-rum and variegata features, see also Tatter-
sall, 1982: 70). Clearly, it will be importantto document the pelage and color patterns ofwild-caught animals and especially of naturalhybrids whenever such opportunities arise.One such opportunity existed at Zoo Ivoloinain Tamatave, Madagascar, where an apparentwild hybrid resided from 16 July 1990 untilher death on 2 July 1996. Zoo Ivoloina staffwere told that she was one of a pair of twinsthat had fallen from a tree along a river ‘‘nearMaroantsetra’’. She was kept for some timeas a pet in Maroantsetra and was only latertransferred to Zoo Ivoloina. She appearedsimilar to the 50–50 V. v. rubra 3 V. v. var-iegata hybrids that were at one time pro-duced at the DUPC (A. Katz, personal com-mun.). Unfortunately, photographs were nottaken of this hybrid, nor was the skin pre-served.
SURVEY WORK
The largely separate geographic distribu-tions of black-and-white and red ruffed le-murs throughout most of their range were notknown until long after their reduction to sub-species of the same species had become en-trenched. The red ruffed lemur occurs prin-cipally on the Masoala Peninsula, while theblack-and-white ruffed lemur has a muchlarger range extending from south of theMananara River (at least to the region northof Midongy du Sud) to the AntainambalanaRiver in the north (near Maroantsetra) (fig.2; Petter et al., 1977; Tattersall, 1977). Petteret al. (1977) and Tattersall (1982) stated thatthe Antainambalana River currently appearsto divide the two subspecies; however, thenorthern and western reaches of the redruffed lemur’s range are incompletelyknown. Petter et al. (1977) illustrated thenorthernmost distribution of the red ruffedlemur as Andapa, whereas Tattersall’s infor-mants, in particular the late Georges Randri-anasolo, a highly experienced observer, in-dicated that this form occurred only as farnorth as Cap Est—although it might have ex-tended as far north as Antalaha until recently(Tattersall, 1977). Perhaps significantly, noruffed lemur bony remains are known fromany of the subfossil sites in northern Mada-gascar (L. Godfrey, personal commun.).Based on local reports of a rufous brown var-

18 NO. 3376AMERICAN MUSEUM NOVITATES
iant NW of Maroantsetra (Tattersall 1977),and a study of museum holdings and collec-tion records (Tattersall 1982; Buettner-Jan-usch and Tattersall, 1985), it was suggestedthat V. v. variegata and V. v. rubra are insecondary contact (i.e., hybridize) betweenthe confluent Vohimara and AntainambalanaRivers. These suggestions prompted surveysin the environs of Maroantsetra, north of theBay of Antongil, in tandem with conserva-tion initiatives.
Two surveys completed since the mid-1980s seem to confirm the AntainambalanaRiver as the current geographic divide be-tween the black-and-white form and the redform, although neither survey was successfulin actually locating hybrids. While V. v. var-iegata (subcincta type) was confirmed justwest of the Antainambalana River near An-daparaty village (Lindsay and Simons, 1986;Simons and Lindsay, 1987), local villagers atAndaparaty, Ankorongana, and Sahantahaclaim that ‘‘varinena’’ (vernacular for the redform) occur east of the Antainambalana Riv-er (fig. 3; Lindsay and Simons, 1986; Simonsand Lindsay, 1987; Hekkala and Rakoton-dratsima, 1999). Unfortunately, lemur trap-ping and extensive swidden agriculture occurbetween the Antainambalana and Andrano-fotsy River valleys, but according to localreports, V. v. rubra may still be abundant inat least one locality there–on Anjanaharibe,a mountain rising to 1150 m (E. Hekkala,personal commun.). Although red ruffed le-murs were not actually sighted by surveyteams between the Antainambalana and An-dranofotsy River valleys, they were seen intwo locations further east between the An-dranofotsy and Mahalevona Rivers: near Sa-havary village along the east bank of the An-dranofotsy River (Lindsay and Simons,1986; Simons and Lindsay, 1987), and alongthe Belampona, a small river between theAndranofotsy and Mahalevona watershedssouth of the Masoala National Park border(Hekkala and Rakotondratsima, 1999). Le-mur trapping and swidden agriculture are,unfortunately, common in this region as well(Hekkala and Rakotondratsima, 1999).Black-and-white ruffed lemurs may havebeen collected at Sahavary more than 100years prior to 1986 (table 2).
These two surveys prompt further ques-
tions and suggest several conservation initia-tives worthy of pursuit. First, the northerndistribution of Varecia still remains unveri-fied up to Andapa. Second, the survey teamspaid special attention to the region approxi-mately 40 km NW of Maroantsetra along theAntainambalana River (fig. 3), given theprovenance of the one wild-caught hybridknown at the time. As this review indicatesthat the hybrid zone extended across two ad-ditional watersheds to the east, future sur-veys should investigate these regions in moredepth, the Mahalevona watershed in partic-ular. The survey by Lindsay and Simons didnot include the Mahalevona watershed at all,and that by Hekkala and Rakotondratsimadid not penetrate very far up it. There areverified reports of red ruffed lemurs in theupper reaches of the Mahalevona watershedfrom individuals involved in forest corridorwork for the Masoala National Park (seeConservation Implications; Holloway, 1997;and see also Rand, 1932). Third, locatingpopulations of ruffed lemurs between theAntainambalana and Andranofotsy riversshould be a priority. Local villagers refer tothe lemurs in this region as ‘‘varinena’’ andindicate that they possess an abdominal beltof white fur, as in the black-and-white ruffedlemur (subcincta type) (Hekkala and Rako-tondratsima, 1999). The suffix nena comesfrom mena, which in Malagasy refers notonly to red, but also to light brown (as inbrown sugar). Perhaps the ‘‘rufous brownvariant’’ reported in this region by Malagasyinformants, Georges Randrianasolo in partic-ular (Tattersall, 1977), and hybrids are oneand the same. Alternatively, if the ‘‘hybridzone’’ is indeed much larger than previouslysuspected, it is worth pondering whether anentirely distinct variety or subspecies occursor occurred within this region. If so, its num-bers have been perilously reduced by humanactivity over the last century or more. Thismight explain why only ‘‘true’’ forms, black-and-white or red, have been witnessed in thisregion during recent surveys. Such popula-tions might then be filtering in from areasfarther east and farther west, where they havenot been ravaged as extensively by habitatloss and hunting. It is well known that de-graded habitat (and ecotones) are major fac-tors that bring about hybridization as a result

2002 19VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
of disrupted premating isolating mechanisms(i.e., ecological separation and normal be-havioral incompatibility between congeners),especially under conditions where hybridsmight be better adapted to disturbed habitatthan are parental taxa (Moore, 1977).
GENETIC DATA
With modern methods that require nomore than plucked hairs to assess relation-ships, museum hybrids and black-and-whiteand red forms of known provenance that co-occur comprise potential material for geneticstudy of the apparent ruffed lemur hybridzone (tables 1 and 2). To date, however, ge-netic studies of ruffed lemurs have drawnprimarily on captive animals whose lineagescan in most cases be traced back to specificwild-caught individuals, but whose ancestralprovenance is unclear or unknown. There area few important exceptions. Tissue samplesof wild red ruffed lemurs from AndranobeForest in the Masoala National Park (Vasey,1997) and of wild black-and-white ruffed le-murs from the Betampona Strict Nature Re-serve (Britt et al., 1998), the Manombo Spe-cial Reserve (Ratsimbazafy, in prep.), andRanomafana National Park (Balko, 1998)were recently collected. In combination withsamples from captive ruffed lemurs, the sam-ples from wild ruffed lemurs of known prov-enance have been used in recent genetic stud-ies to assess variation within and amongblack-and-white and red ruffed lemurs. Twoseparate studies of variation between black-and-white and red ruffed lemurs, using dif-ferent genes and different methods, are sum-marized here.
One study (Pastorini, 2000) includes wildred ruffed lemurs from Andranobe Forest (n5 3), captive red ruffed lemurs (n 5 3), andcaptive black-and-white ruffed lemurs (n 53). The captives originate from zoos in Eu-rope and Madagascar, and all can be tracedto wild-caught, true forms (I. Porton, person-al commun.). The sample of red ruffed le-murs shows absolute pairwise genetic dis-tances consistent with within-subspecies var-iation, whereas the sample of black-and-white ruffed lemurs shows genetic distancessufficient to designate two subspecies. Threeseparate tree-building methods (maximum
parsimony, neighbor-joining, maximum like-lihood) show the red ruffed lemur samplesclustering between two discrete sets of theblack-and-white ruffed lemur samples.mtDNA genes used in this analysis consistof the 39 end of the COIII gene (53 bp), thecomplete NADH-dehydrogenase subunitsND3 (348 bp), ND4L (297 bp) and ND4(1378 bp), along with the glycine (72 bp),arginine (68 bp), histidine (70 bp), serine (64bp), and part of the leucine (47 bp) tRNAgenes (Pastorini, 2000).
A second study includes wild red ruffedlemurs from Andranobe Forest (n 5 3); wildblack-and-white ruffed lemurs from Betam-pona (n 5 3), Manombo (n 5 6), and Ran-omafana (n 5 14); wild-caught animals re-cently held at Zoo Ivoloina, Tamatave (n 56); captive ruffed lemurs from the SpeciesSurvival Plan (SSP) population (n 5 12); ared ruffed lemur traceable to wild-caughtparents (n 5 1); and black-and-white ruffedlemurs traceable to wild-caught individuals(n 5 11). The black-and-white ruffed lemursamples form two diagnosable units consist-ing of a southern group from Manombo andRanomafana, and a northern group from Be-tampona (Wyner et al., 1999). Red ruffed le-murs comprise a distinct clade and form thesister group to all of the black-and-whiteruffed lemurs sampled (Wyner, 2000; G.Amato, personal commun.). The highly var-iable control region (D-loop) mtDNA frag-ment (548 bp) was used to study variationamong and between black-and-white ruffedlemur populations. The more conservedmtDNA ribosomal 12s rRNA and 16s rRNAregions were also sequenced, to examine therelationships between the black-and-whiteand red ruffed lemur samples. These phylo-genetically informative ribosomal genes in-dicate that these two lineages had separatedearlier than had the various black-and-whiteforms. We recognize that genetic distances asmeasured here are not absolutely correlatedwith speciation, and also that speciation isnot an immediate or inevitable consequenceof allopatry. Most if not all speciation eventsamong primates probably occur only in al-lopatry, but morphological differentiationand speciation are not the same thing. For-mally named subspecies may similarly be re-garded as geographically subdivided popu-

20 NO. 3376AMERICAN MUSEUM NOVITATES
lations (although, unlike species, they arevariants that may intergrade phenotypicallywithin a species’ range, and in practicalterms are essentially a taxonomist’s conve-nience); and other processes besides allopa-try can also result in speciation. These mayeven include hybridization (Arnold andEmms, 1998; Grant and Grant, 1998). Thus,the mtDNA ribosomal genetic data do not bythemselves provide a definitive argument fora speciation event, although we regard thedata given above as highly suggestive of this.Other approaches can thus usefully bebrought to bear on recognizing species pop-ulations, and among them are the vocaliza-tion data we examine below.
VOCALIZATION DATA
Among vertebrates, species (and admitted-ly sometimes subspecies) commonly differ intheir vocalizations. Vocalizations are espe-cially good indicators of species statusamong nocturnal strepsirhines whose com-munication systems rely heavily on olfactoryand auditory cues rather than on visual ones.Loud calls (advertisement calls in particular)give a strong indication that the communi-cation systems of females and males havebeen finely tuned to one another, and, in ef-fect, that a particular specific-mate recogni-tion system has emerged (Patterson, 1985).Species definitions vary, but a stable specific-mate recognition system is generally agreedto be a critical feature in recognizing the re-productive inclusiveness of a species, wheth-er in sympatry or allopatry, and is a strongindicator that divergence in the fertilizationsystem inherited from the parental specieshas occurred. Specific-mate recognition sys-tems have been especially well demonstratedin closely related galagos, whose advertise-ment calls show distinct, species-specificacoustic structures, which seem to be in-volved both in mate recognition and matechoice (Zimmerman et al., 1988; Zimmer-man, 1995a, 1995b). Vocalization data areparticularly useful for distinguishing strepsir-hine species in the field, often providing thefirst clues that groups of animals may differin other features of their fertilization system,anatomy, life history, genetics, ecological
niche, and social behavior (Masters, 1998;Bearder, 1999).
One loud call emitted by ruffed lemurs isreferred to as the ‘‘pulsed squawk’’ (aka ‘‘uh-uh-uh . . . ’’ call). It functions as a terrestrialpredator alarm call but is also used in otherhigh arousal contexts (Macedonia and Taylor,1985; Morland, 1991; pers. obs.). This calldiffers between black-and-white and redruffed lemurs in median high frequency,pulse rate, and pulse duration; and becauseof its relatively narrow frequency band pulse,it has been considered a good indicator ofphyletic divergence (Macedonia and Taylor,1985). Nonetheless, in the study in whichthis was revealed, Macedonia and Taylor(1985) did not consider upgrading black-and-white and red ruffed lemurs to full speciesbased on their acoustic study of this call,since they simply retained the subspecific no-menclature that was available when theypublished. Divergence in the ‘‘pulsedsquawk’’ call is clearly not just a matter ofdialects, such as those recently discovered inneighboring populations of the grey mouselemur, Microcebus murinus (Hafen et al.,1998); black-and-white and red ruffed lemursare separated geographically, and do not nor-mally form overlapping demes.
In conjunction with the evidence noted forphylogenetic splitting from mtDNA ribosom-al genes, the two forms’ largely distinct geo-graphic distributions, their differences in pel-age color and pattern, and other historicalfactors detailed in previous sections, suchquantified differences in the ruffed lemurs’terrestrial antipredator calls constitute per-suasive evidence that black-and-white andred ruffed lemurs warrant separate speciesstatus and should appropriately be referred toby the binomina Varecia variegata (Kerr,1792) and Varecia rubra (E. Geoffroy,1812), respectively. We feel reasonably con-fident that our case for upgrading black-and-white and red ruffed lemurs to separate spe-cies will be strengthened by field and captivestudies of sexually dimorphic calls. Suchstudies would provide especially useful ad-ditional evidence to evaluate whether diver-gent specific-mate recognition systems haveevolved in the two forms. The ruffed lemurvocal repertoire includes various sex-specificcalls and a loud call to which females and

2002 21VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
males contribute differently (e.g. the ‘‘roar-shriek’’; Morland, 1991; personal obs.).Demonstration of specificity in such callswould form a solid basis for an acousticstudy of specific-mate recognition systemsamong ruffed lemurs.
SYSTEMATIC SUMMARY
The fact that the two forms of Vareciamay occasionally form hybrids, and readilydo so in zoos, is not necessarily significantin determining their species status. Specia-tion, after all, is not a unitary mechanism, butis instead an outcome, with numerous poten-tial underlying causes. Isolating mechanismsmay operate at any of various levels, whetherbehavioral, chromosomal, anatomic, or de-velopmental. The essential question is notwhether hybrid zygotes may develop nor-mally, or even reproduce successfully asadults, but is rather whether under normalcircumstances there is any biologicallymeaningful gene exchange between the twopopulations. Among the two kinds of ruffedlemur this does not appear to be the case, foralthough there appears plausibly to be, or tohave been, a geographically quite extensivezone of occasional hybridization between theblack-and-white and red ruffed lemurs in theregion shown in figure 3, hybridization with-in that zone has apparently been a rare oc-currence at best. There is certainly nothingknown that could be described as a zone ofintergradation among the two populations.Habitat deterioration caused by human activ-ity in the region might conceivably have re-sulted in the disappearance of a distinctivepopulation bearing hybridlike characteristics.However, it is very much more likely thatsuch habitat disruption has artificially facili-tated hybridization within the zone. This, ofcourse, would make the apparent rarity ofhybridization even more puzzling if we weredealing with but one species. Our conclusion,then, is ineluctible. The two major forms ofruffed lemur warrant recognition as two dis-tinct species (two effectively autonomous re-productive units): Varecia variegata and V.rubra.
As actual or potential evolutionary ephem-era, subspecies may be recognized if they areuseful to the taxonomist. And while, as not-
ed, there is more variation in pelage colora-tion amongst the red ruffed lemurs than hasusually been appreciated, such variation ap-pears to be relatively evenly distributed with-in the red ruffed lemur population and showsno geographical patterning. Varecia rubracan thus be regarded as variable but mono-morphic. On the other hand, the black-and-white form does show at least four relativelydistinctive variants, of which three have beengiven their own taxonomic designations: var-iegata, editorum, and subcincta. Distribu-tional data on museum specimens represent-ing these variants has been woefully lacking.But combining what little is certainly knownfrom museum specimens with reliable recentfield observations (kindly contributed by An-drea Katz [for Betampona] and ElizabethBalko [for Ranomafana and Kianjavato], inaddition to unpublished observations by I.T.both to the north and the south of the Man-anara River in the far south) leads to a com-plex picture.
Although there appears to be a very vaguetendency toward lightness in the pelage ofVarecia variegata from north to south, thefour distinctive variants recognized by Tat-tersall (1982) seem to be relatively randomlydistributed across space (fig. 2). Individualsof the variegata type are found mainly southof Mananjary, but have also been collectedas far north as Mananara; subcincta is foundin the region to the west of Toamasina (Ta-matave), but then so are variegata and edi-torum; subcincta is found as far north as theMaroantsetra region, but also as far south asFenoarivo, but with variegata in between;and so on. In the absence of any clear geo-graphic patterning within the huge area ofdistribution of Varecia variegata, it seemsmore realistic to regard this species as a pop-ulation embracing several possibly largelydiscrete pelage coloration patterns, which areprobably quite simply genetically deter-mined, than as one that is usefully divisibleinto discrete geographic subspecies. The con-clusion as regards the black-and-white ruffedlemurs thus appears to be variable (in a fash-ion different from the red ruffed lemur), butmonotypic. We cannot concur with Groves(2001) in raising variants of Varecia varie-gata to subspecies status.

22 NO. 3376AMERICAN MUSEUM NOVITATES
CONSERVATION IMPLICATIONS OFVARECIA SYSTEMATICS
Earlier we indicated that documentation ofa natural hybrid zone would be of great sig-nificance in designating geographic prioritiesfor habitat conservation, for captive manage-ment, and for lemur systematics generally.Our review of historical documents and tax-onomy suggests that the down-ranking of theblack-and-white and red ruffed lemurs tosubspecies was largely influenced by the sub-stantial number of hybrids produced in cap-tivity. Captive hybridization of lemurs oc-curred before 1870, and it is highly likelythat the taxonomic revisions of Varecia inthe 19th century were influenced by suchbreeding practices, whether intentional ornot. We now know that black-and-white andred ruffed lemurs have distinct geographicdistributions, genetic profiles, and loud calls,and that in sum these distinctions make astrong case for separate species recognitionfor black-and-white and red ruffed lemurs.
A review of museum holdings and collec-tion records suggests that a natural hybridzone between black-and-white and red ruffedlemurs occurred north of the Bay of Anton-gil, extending as far east as the MahalevonaRiver and as far west as the Vohimara River,a tributary of the Antainambalana (fig. 3).Although hybridization appears to be rare,this hybrid zone is considerably larger thanpreviously thought. Recent surveys have lo-cated populations of both the red form andthe black-and-white form in this region; de-graded habitat may currently prevent the twoforms from coming into frequent contact, butit may equally also bring about hybridizationat times when contact is made. As with theAntainambalana and Andranofotsy Rivervalleys (Lindsay and Simons, 1986; Hekkalaand Rakotondratsima, 1999), the Mahale-vona River valley is well populated by hu-mans, especially in its southern reaches(CARE/WCS/PF, 1995). A subcincta-likeform with reddish brown fur in place ofblack may exist between the Antainambalanaand Andranofotsy watersheds, but this reportrequires verification. Future surveys shouldalso investigate the Mahalevona watershed,where we have documented an additional hy-brid and regions of overlap.
The recently established Masoala NationalPark was designed to shelter several patchesof littoral forest, a considerable, contiguousportion of the Masoala Peninsula, a band offorest running northeast by southwest be-tween the Ambanizana and Mahalevona wa-tersheds, and a small band of forest runningeast to west between the Mahalevona andAndranofotsy River valleys (fig. 3). Thesebands of forest in the northernmost reachesof the Masoala National Park are connectedto the main body of the Park via three forestcorridors which are under active restoration,but are each less than 1 km wide (Holloway,1997, 2000). The Masoala Peninsula, includ-ing the corridors, suffered devastating dam-age during Cyclone Huddah, which struck on2 April 2000. Cyclones leave a poignantmessage behind when they rupture such ten-uous links between forest patches. Given ourcurrent understanding of the ruffed lemur hy-brid zone, forest corridors between intact for-est patches will be more critical than ever inmaintaining population networks for ruffedlemurs (and other forest-bound taxa) north ofthe Bay of Antongil. However, to bridge theentire hybrid zone, a protected network ofintact forest patches and actively managedforest corridors must extend farther east be-tween the Andranofotsy and Antainambalanawatersheds. If we are successful in establish-ing a network of forest patches and corridors,perhaps we will know again what naturalistsmay have known long ago; that two distinctvariegated lemurs meet in a restricted hybridzone in northern Madagascar, but that hy-bridization between them is and was rare. Inthe longer term, biodiversity in northernMadagascar must be held intact by forestcorridors connecting protected areas furthernorth (the Parc National de Marojejy and theReserve Speciale d’Anjanaharibe-Sud), far-ther south (the Parc National de Verezanant-soro), and farther east (the Reserve Specialede Manongarivo and the Reserve Naturellede Tsaratanana) (see also Ganzhorn et al.,1996/1997; Goodman, 1999).
ACKNOWLEDGMENTS
We thank many individuals for kindly ar-ranging access to museum holdings and doc-uments, including Richard Thorington and

2002 23VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
Linda Gordon (Washington, D.C.), PaulaJenkins and the late Prue Napier (London),Chris Smeenk and A.M. Husson (Leiden),Francis Petter, Michel Tranier, and Roger Sa-ban (Paris), Edi Rutzmoser (Cambridge,Mass.), and Guy Musser, Sydney Anderson,and Chris Norris (New York). Ian Tattersallparticularly expresses his appreciation to thelate Georges Randrianasolo for so generouslysharing his expertise on lemur distributions.We thank George Amato (WCS) and YaelWyner (AMNH) who made a valuable con-tribution to the genetics section of this paper.Ken Mowbray of the AMNH helped in manyways, and particularly in preparing the colorplate with images furnished by David Haring(DUPC) and Natalie Vasey. Ken Cameron,D.V.M. (Cincinnati Zoo) and Stephen Nash(SUNY, Stony Brook) also helped with vi-sual documentation. Bridget Thomas kindlydrafted figures 1 and 2. We would also liketo thank the staff librarians at the UnitedStates National Museum, the National Zoo-logical Park, the Library of Congress (Wash-ington, D.C.), and the American Museum ofNatural History for assistance in locatingmany documents, including rare illustrationsof ruffed lemurs. Thanks also go to StefanFalke for help with German translation, toEvon Hekkala and Marius Rakontondratsimafor access to their unpublished survey report,to David Lees and Russell Thorstrom forhelp with geo-referencing sites, to IngridPorton for providing pedigrees for captive-born ruffed lemurs, and to Andrea Katz andElizabeth Balko for field identifications fromBetampona and Ranomafana/Kianjavato, re-spectively. We thank Laurie Godfrey, ErikGunther, Judith Masters, Elwyn Simons,Anne Yoder, and Bob Sussman for their com-ments on an earlier draft of this paper.
REFERENCES
Arnold, M.L., and S.K. Emms. 1998. Paradigmlost: Natural hybridization and evolutionary in-novations. In D. J. Howard and S. H. Belocher(editors), Endless forms: species and specia-tion: 379–389. New York and Oxford: OxfordUniversity Press.
Audebert, J.B. 1797. Histoire Naturelle des Singeset des Makis. Pls. V and VI. Paris: Desray.
Balko, E.A. 1998. A behaviorally plastic responseto forest composition and logging disturbance
by Varecia variegata variegata in RanomafanaNational Park, Madagascar. Ph.D. dissertation,State University of New York, College of En-vironmental Science and Forestry, Syracuse,NY.
Bartlett, A.D. 1862. Description of a new speciesof lemur. Proceedings of the Zoological Societyof London, 1862: 347.
Bearder, S.K. 1999. Physical and social diversityamong nocturnal primates: a new view basedon long term research. Primates 40: 267–282.
Benirschke, K., and C. Miller. 1981. Weights andneonatal growth rings of ring-tailed lemurs andruffed lemurs. Journal of Zoo and Animal Med-icine 12: 107–111.
Britt, A., C. Welch, and A. Katz. 1998. The firstrelease of captive-bred lemurs into their naturalhabitat. Lemur News 3: 8–11.
Buettner-Janusch, J., and I. Tattersall. 1985. Anannotated catalogue of Malagasy primates(families Lemuridae, Indriidae, Daubentoni-idae, Megaladapidae, Cheirogaleidae) in thecollections of the American Museum of NaturalHistory. American Museum Novitates 2834: 1–45.
Buffon, G.L.L. 1765. Histoire naturelle, generaleet particuliere. Tome XIII, pl. XXVII. Paris:Imprimerie Royale.
CARE/WCS/TPF 1995. Proposition des limitesdu Parc National Masoala. Unpublished reportby CARE International Madagascar, WildlifeConservation Society, and the Peregrine Fund.
Cauche, F. 1651 [Translated in 1711 by J. Ste-vens]. A Voyage to Madagascar, the AdjacentIslands, and the Coast of Afrik. In: A new col-lection of voyages and travels, vol. 7. London.
Ceska, V., H.U. Hoffman, and K.-H. Winkelstra-ter. 1992. Lemuren im Zoo. Berlin: Verlag PaulParey.
Dechambre, E. 1935. Observations sur les makisde la menagerie du Jardin des Plantes. Bulletindu Museum National d’Histoire Naturelle,2eme serie, 7(6): 315–319.
Elliot, D.G. 1913. A review of the primates.Monographs of the American Museum of Nat-ural History, vol. 1.
Evans, M.I., P.M. Thompson, and A. Wilson.1993–1994. A survey of the lemurs of Amba-tovaky Special Reserve. Primate Conservation(14–15): 13–21.
Flacourt, E. de. 1658. Histoire de la grande isleMadagascar. Paris: G. de Lvyne.
Flacourt, E. de. 1658. Histoire de la grande isleMadagascar. Reprinted 1991, Sainte Clotilde,La Reunion: A.R.S. Terres Creoles (CollectionMascarin).
Flower, S.S. 1933. Breeding season of lemurs.

24 NO. 3376AMERICAN MUSEUM NOVITATES
Proceedings of the Zoological Society of Lon-don, 1933: 317.
Forbes, H.O. 1894. A hand-book to the primates.Pl. VII. London: Edward Lloyd.
Ganzhorn, J.U., O. Langrand, P.C. Wright, S.O’Connor, B. Rakotosamimanana, A. T.C.Feistner, and Y. Rumpler. 1996/1997. The stateof lemur conservation in Madagascar. PrimateConservation 17: 70–86.
Geoffroy Saint-Hilaire, E. 1796. Memoire sur lesrapports naturels des makis Lemur L. et de-scription d’une espece nouvelle de Mammifere,par le citoyen Geoffroy. Magasin Encyclope-dique, 2eme annee, 1: 20–50.
Geoffroy Saint-Hilaire, E. 1812. Suite au tableaudes quadrumanes. Seconde famille. Lemuriens.Strepsirrhini. Paris: Annales du Museumd’Histoire naturelle 19: 156–170.
Geoffroy Saint-Hilaire, E. 1824a. Le maki rouxfemelle (Livraison 15, April 1820). HistoireNaturelle des Mammiferes. Tome Premier. Par-is: A. Belin.
Geoffroy Saint-Hilaire, E. 1824b. Vari (Livraison43, August 1824). Histoire naturelle des mam-miferes. Tome Cinquieme. Paris: A. Belin.
Geoffroy Saint-Hilaire, I. 1851. Premiere partie.Mammiferes. Catalogue des primates. In Cata-logue methodique de la collection des mam-miferes de la collection des oiseaux et des col-lections annexes. Museum d’Histoire Naturellede Paris. Paris: Gide et Baudry.
Gervais, P. 1854. Histoire naturelle des mammi-feres. Pl. X. Paris: L. Curmer.
Gmelin, J.F. 1788. Systema Naturæ. 13th ed. Vol.1. Lipsice: G.E. Beer.
Goodman, S.M. 1999. Biological inventories offorest fragments and forest corridors in easternand central Madagascar. Lemur News 4: 28–30.
Grandidier, G., and G. Petit. 1932. Zoologie deMadagascar. Paris: Societe d’Editions Geograp-hiques, Maritimes et Coloniales.
Grant, B.R., and P.R. Grant. 1998. Hybridizationand speciation in Darwin’s finches: The role ofsexual imprinting on a culturally transmittedtrait. In D.J. Howard and S.H. Belocher (edi-tors), Endless forms: species and speciation:404–422. New York and Oxford: Oxford Uni-versity Press.
Gray, A.P. 1954. Mammalian Hybrids. (A checklist with bibliography). London: C.A.B.
Gray, J.E. 1863. Revision of the species of le-muroid animals, with the description of somenew species. Proceedings of the Zoological So-ciety of London 1863: 129–153.
Gray, J.E. 1870. Catalogue of monkeys, lemurs,and fruit-eating bats in the collection of theBritish Museum. London: Taylor and Francis.
Gray, J.E. 1871. On a new species of lemur from
Madagascar, and on the changes of Lemur ma-caco, Linnean Annual Magazine of NaturalHistory, 4th series, 7: 339–340.
Gray, J.E. 1872. Notes on Propithecus, Indris, andother lemurs (Lemurina) in the British Muse-um. Proceedings of the Zoological Society ofLondon 1872: 846–860.
Groves, C. 2001. Primate taxonomy. Washington,D.C.: Smithsonian Institution Press.
Hafen, T., H. Neveu, Y. Rumpler, I. Wilden, andE. Zimmerman. 1998. Acoustically dimorphicadvertisement calls separate morphologicallyand genetically homogeneous populations ofthe grey mouse lemur (Microcebus murinus).Folia Primatologica 69 (suppl. 1): 342–356.
Handfest, K. 1967. Josef-Peter Audebert, derMadagaskarforscher aus Dillingen-Saar. KarlHandfest, 6623 Luisenthal-Saar-Postfach 33.[Independently published by the author]
Hekkala, E.R., and M. Rakotondratsima. 1999.Report on preliminary surveys for distributionand population genetic structure of the red-ruffed lemur (Varecia variegata rubra) in theAntainambalana/Andranofotsy River Basin ofMadagascar: an assessment of forest remnantsas corridors for conservation. [Unpublished re-port presented to Wildlife Conservation Socie-ty, Madagascar]
Hill, W.C.O. 1953. Primates: comparative anato-my and taxonomy. I. Strepsirrhini. New York:Interscience Publishers, Inc.; Edinburgh: TheUniversity Press.
Holloway, L. 1997. Catalysing natural regenera-tion of rainforest: Masoala corridors. [Unpub-lished report presented to Wildlife Conserva-tion Society, Madagascar]
Holloway, L. 2000. Catalysing rainforest restora-tion in Madagascar. In W. R. Lourenco and S.M. Goodman (editors), Diversity and ende-mism in Madagascar: 1–10. Paris: Memoires dela Societe de Biogeographie.
Jenkins, P.D. 1987. Catalogue of primates in theBritish Museum (Natural History) and else-where in the British Isles. Part IV: SuborderStrepsirrhini, including the subfossil Madagas-can lemurs and family Tarsiidae. London: Brit-ish Museum of Natural History.
Jentink, F.A. 1892. Catalogue systematique desmammiferes (singes, carnivores, ruminants,pachyderms, sirens, et cetaces), Tome XI. Mu-seum d’Histoire Naturelle des Pays-Bas. Lei-den: E.J. Brill.
Kaudern W. 1915. Saugetiere aus Madagaskar.Arkiv for Zoologi 9(18): 1–101 (1 4 plates).
Kerr, R. 1792. Animal kingdom. Class I. Mam-malia. Edinburgh: W. Creech.
Lesson, R.P. 1840. Species des mammiferes bi-manes et quadrumanes. Paris: J.-B. Bailliere.

2002 25VASEY AND TATTERSALL: SYSTEMATICS OF VARECIA
Lesson, R.P. 1842. Nouveau tableau du regne an-imal. Mammiferes. Paris: Arthus Bertrand.
Lindsay, N.B.D. 1977. Notes on the taxonomicstatus and breeding of the ruffed lemur. Dodo14: 65–69.
Lindsay, N.B.D., and H.J. Simons. 1986. Noteson Varecia in the northern limits of its range.Dodo 23: 29–24.
Linnaeus, C. 1766. Systema Naturæ. 12th ed., to-mus 1. Stockholm: Laurentii Salvii.
Macedonia, J.M., and L.M. Taylor. 1985. Subspe-cific divergence in a loud call of the ruffed le-mur (Varecia variegata). American Journal ofPrimatology 9: 295–304.
Masters, J.C. 1998. Speciation in the lesser gala-gos. Folia Primatologica 69(suppl.1): 357–370.
Milne-Edwards, A., and A. Grandidier. 1890. His-toire physique, naturelle et politique de Mada-gascar. Atlas II, vol. X, tome V, Pls. CXXIII–CXXIX.
Moore, W.S. 1977. An evaluation of narrow hy-brid zones in vertebrates. Quarterly Review ofBiology 52: 263–277.
Morland, H.S. 1991. Social organization and ecol-ogy of black and white ruffed lemurs (Vareciavariegata variegata) in lowland rain forest,Nosy Mangabe, Madagascar. Ph.D. dissertation,Yale University, New Haven, CT.
Pastorini, J. 2000. Molecular systematics of le-murs. Ph.D. dissertation, University of Zurich,Switzerland.
Patterson, H.E.H. 1985. The recognition conceptof species. In E.S. Vrba (editor), Species andspeciation: 21–29. Pretoria: Transvaal Museum.
Petter, J.J. 1962. Recherches sur l’ecologie etl’ethologie des lemuriens Malgaches. Memo-ires du Museum National d’Histoire Naturelleserie A, Zoologie, 27(1): 1–146.
Petter, J.J. 1969. Speciation in Madagascan le-murs. Biological Journal of the Linnaean So-ciety 1: 77–84.
Petter, J.J., R. Albignac, and Y. Rumpler. 1977.Faune de Madagascar 44: mammiferes lemu-riens (Primates, prosimiens). Paris: ORSTOM/CNRS.
Pollen, F.P.L. 1868. Notices sur quelques autresmammiferes habitant Madagascar et les ılesvoisines. In F.P.L. Pollen and D.C. Van Dam(editors), Recherches sur la faune de Madagas-car et de ses dependences. Pt. 2. H. Schlegeland F.P.L. Pollen. Mammiferes et oiseaux: 19–29. Leiden: J. K. Steenhoff.
Rand, A.L. 1932. Mission Franco-Anglo-Ameri-caine a Madagascar. Notes du voyage. L’Oiseauet la Revue Francaise d’Ornithologie (new ser.)2: 227–282.
Rand, A.L. 1936. On the distribution and habitsof Madagascar birds. Summary of the field
notes of the Mission Zoologique Franco-Anglo-Americaine a Madagascar. Bulletin of theAmerican Museum of Natural History 72(5):143–499.
Ratsimbazafy J. In prep. Ph.D. dissertation, StateUniversity of New York, Stony Brook.
Schlegel, H. 1866. Contributions a la faune deMadagascar et des ıles avoisinantes, d’apres lesdecouvertes et observations de Mm. FrancoisPollen et M.D.-C. van Dam. NederlandschTijdschrift voor de Dierkunde 3: 73–89.
Schlegel, H. 1876. Revue methodique et critiquedes collection deposees dans cet etablissement.Tome 7, monographie 40: Simiae. Leiden: Mu-seum d’Histoire Naturelle des Pays-Bas.
Schreber, J.C.D.von. 1775. Die Saugethiere inAbildungen nach der Natur mit Beschreibun-gen. Vol. 1 (text), and vol. 1 (plates). Erlangen:in der Expedition des schreber’schen saugthier-und des Esper’schen Schmetterlingswerkes. Pl.XLB (after Buffon).
Schwarz, E. 1931. A revision of the genera andspecies of Madagascar Lemuridae. Proceedingsof the Zoological. Society of London, 1931:399–428.
Sclater, P.L. 1871. Notes on rare or little-knownanimals now or lately living in the Society’sgardens. Part I. Mammalia. Proceedings of theZoological Society of London, 1871: 221–240.
Simons, H.J., and N. B.D. Lindsay. 1987. Surveywork on ruffed lemurs (Varecia variegata) andother primates in the northeastern rain forestsof Madagascar. Primate Conservation 8: 88–91.
Smith A. 1833. An epitome of African zoology,pt. 1, Mammalia. South African Quarterly Jour-nal (Capetown) II, 17–32, pt. 2: 49–64, 81–96.
Tattersall, I. 1977. Distribution of lemurs. Part 1:The lemurs of northern Madagascar. Annals ofthe New York Academy of Sciences 293: 160–169.
Tattersall, I. 1982. The primates of Madagascar.New York: Columbia University Press.
Vasey, N. 1997. Community ecology and behav-ior of Varecia variegata rubra and Lemur ful-vus albifrons on the Masoala Peninsula, Mad-agascar. Ph.D. dissertation, Washington Univer-sity, St. Louis, MO.
Vasey, N. In press. Varecia. In S. M. Goodmanand J. Benstead (editors), Natural History ofMadagascar: [page numbers]. Chicago: Univer-sity of Chicago Press.
Wyner, Y.M. 2000. Species boundaries and con-servation units in two species in the family Le-muridae. Ph.D. dissertation, Yale University,New Haven, CT.
Wyner, Y.M., G. Amato, and R. DeSalle. 1999.Captive breeding, reintroduction, and the con-servation genetics of black and white ruffed le-

26 NO. 3376AMERICAN MUSEUM NOVITATES
murs, Varecia variegata variegata. MolecularEcology 8: S107–S115.
Zimmerman, E. 1995a. Acoustic communicationin nocturnal prosimians. In L. Alterman, M.K.Izard, and G.A. Doyle (editors), Creatures ofthe dark: the nocturnal prosimians: 311–330.New York: Plenum Press.
Zimmerman, E. 1995b. Loud calls in nocturnalprosimians: structure, evolution and ontogeny.In E. Zimmerman, J.D. Newman, and U. Jur-gens (editors), Current topics in primate vocalcommunication: 47–72. New York: PlenumPress.
Zimmerman, E., S.K. Bearder, G.A. Doyle, andA.B. Andersson. 1998. Variations in vocal pat-terns of Lesser Bushbabies (Galago senegalen-sis and Galago moholi) and their implicationsfor taxonomic relationships. Folia Primatologi-ca 51: 87–105.
Zuckerman, M.A. 1932. The menstrual cycle ofthe primates. Part VI. Further observations onthe breeding of primates, with special referenceto the suborders Lemuroidea and Tarsioidea.Proceedings of the Zoological Society of Lon-don, 1932: 1059–1075.

Recent issues of the Novitates may be purchased from the Museum. Lists of back issues of theNovitates and Bulletin published during the last five years are available at World Wide Web sitehttp://library.amnh.org. Or address mail orders to: American Museum of Natural History Library,Central Park West at 79th St., New York, NY 10024. TEL: (212) 769-5545. FAX: (212) 769-5009. E-MAIL: [email protected]
a This paper meets the requirements of ANSI/NISO Z39.48-1992 (Permanence of Paper).