distribution of ?-conotoxin gvia binding sites in teleost cerebellar and electrosensory neurons
TRANSCRIPT

THE ,JOURNAL OF COMPARATIVE NEUROLOGY 364:456472 (1996)
Distribution of w-Conotoxin GVIA Binding Sites in Teleost Cerebellar and
Electrosensory Neurons
Y. THARANI, G.A. THURLOW, ANI) R.W. TURNER Neuroscience Research Group, University of Calgary, Calgary, Alberta, Canada, T2N 4N1
ABSTRACT The distribution of w-Conotoxin GVIA (CgTx) binding sites was used to localize putative
N-type Ca2+ channels in an electrosensory cerebellar lobule, the eminentia granularis pars posterior, and in the electrosensory lateral line lobe of a gymnotiform teleost (Apteronotus leptorhynchus). The binding sites for CgTx revealed by an anti-CgTx antibody had a consistent distribution on somatic and dendritic membranes of specific cell types in both structures. The distribution of CgTx binding was unaffected by co-incubation with nifedipine or AgaToxin IVA, blocking agents for L- and P-type Ca2+ channels, respectively. Incubation with CgTx in the presence of varying levels of extracellular Ca2& altered the number but not the cell types exhibiting immunolabel. A punctate immunolabel was detected on somatic membranes of granule and stellate cell interneurons in both the eminentia granularis pars posterior and the electrosensory lateral line lobe. Punctate CgTx binding sites were also present on spherical cell somata and on the large presynaptic terminals of primary afferents that terminate on spherical cells in the electrosensory lateral line lobe. No label was detected in association with distal dendritic membranes of any cell class, or with parallel fibers in the respective molecular layers. Binding sites for CgTx in the eminentia granularis are consistent with the established role for N-type Ca2+ channels in cell migrations, an activity which is known to persist in this layer in adult Apteronotus. The distribution of labeled stellate cells with respect to topographic maps in the electrosensory lateral line lobe further suggest that N-type Ca2+ channels are expressed in relation to functional activity across these sensory maps. ~ ~ f i ~ i l e y - ~ i s s . Inc.
Indexing terms: electric fish, calcium channel, calcium channel blockers, cerebellum
The specific distribution of Ca2+ channels over neuronal membranes is recognized as an important determinant of cell activity. Several Ca2+ channel subtypes have been identified in neural tissue, with the traditional classifica- tion including a low voltage-activated T-type and high voltage-activated L-, N-, P-, Q-, and R-types (Llinas et al., 1992; Olivera et al., 1994; Dunlap et al., 1995). The neurotoxin o-conotoxin GVIA (CgTx) from the marine snail Conus geographus (Olivera et al., 1994) has been shown to bind selectively and with high affinity to neuronal N-type Ca2+ channels (alB; Birnbaumer et al., 1994) with a dissocia- tion constant slow enough to be considered essentially irreversible (Wagner et al., 1988). Our understanding of the distribution and function of N-type Ca2+ channels has thus been advanced through electrophysiological, immunocyto- chemical, and binding studies employing CgTx.
Although CgTx evolved as a paralytic agent directed against the teleost nervous system (Olivera et al., 19941, Ca2+ channels with N-type biophysical properties and/or CgTx sensitivity have been reported in neural tissues of animals that span the vertebrate classes, including elasmo-
branchs (Koyano et al., 1987; Farinas et al., 19931, amphib- ians (Lipscombe et al., 1988; Robitaille et al., 1990; Cohen et al., 1991), birds (Fox et al., 1987; Stanley and Goping, 19911, and mammals (Albensi et al., 1993; Witcher et al., 1993; Wheeler et al., 1994; Boland et al., 1994). Several recent studies have taken advantage of the binding proper- ties of CgTx to localize putative N-type Ca2+ channels at the cellular level by using labeled CgTx or antisera directed against CgTx (Jones et al., 1989; Robitaille et al., 1990; Cohen et al., 1991; Fortier et al., 1991; Haydon et al., 1994; Mills et al., 1994). These and other studies have established that CgTx binding sites are distributed across the CNS in a manner that is distinct from L- or P-type Ca2+ channels (Takemura et al., 1987; Wagner et al., 1988; Llinas et al., 1992; Westenbroek et al., 1992). Thus, biotin- or fluoro- phore-conjugated CgTx binding sites have been localized to
Accepted July 30, 1995. Address reprint requests to Ray W. Turner, Department of Anatomy,
University of Calgary, 3330 Hospital Dr N.W.,CalgaIy, Alberta. Canada T2N 4N1.
7 1996 WILEY-LISS. INC.

N-TYPE CALCIUM CHANNEL DISTRIBUTION
the frog neuromuscular junction (Robitaille et al., 1990; Cohen et al., 19911, bird ciliary ganglion (Haydon et al., 1994), neurites of cultured rat hippocampal pyramidal cells (Jones et al., 19891, and somata and dendrites of rat hippo- campal pyramidal cells (Mills et al., 1994). Putative N-type Ca2+ channels have also been localized through immunocy- tochemical or binding studies to mammalian cerebellar granule cell and molecular layers (Takemura et al., 1989; Komuro and Rakic, 1992; Westenbroek et al., 1992; Albensi et al., 1993) and Purkinje cell somata and dendrites (Fortier et al., 1991; Westenbroek et al., 1992). Surprisingly, very little attention has been paid to the cellular targets for CgTx binding in the teleost central nervous system.
Studying membrane ion channels in the electrosensory system of weakly electric fish allows one to interpret ion channel and single cell activity with respect to neural circuits that have a known physiological function. Periph- eral electroreceptors in these animals monitor an emitted electric organ discharge (EOD) and project primary afferent inputs encoding EOD distortions to the medullary electro- sensory lateral line lobe (ELL; Carr and Maler, 1986). The ELL is a well defined laminar structure comprised of four separate topographic maps (segments) of electroreceptor distribution (Maler, 1979; Carr and Maler, 1986). An overlying cerebellar lobule, the eminentia granularis pars posterior (EGp), also receives electrosensory input and gives rise to a major descending parallel fiber projection to the ELL (Sas and Maler, 1987).
457
Maler, 1988). Calcium conductances have long been known to play a central role in somatic and dendritic activity of cerebellar Purkinje cells (Llinas et al., 1992), and recent studies on N-type Ca2+ channels in mammalian cerebellum suggest an important role for this Ca2+ channel subtype in cerebellar activity (Fortier et al., 1991; Komuro and Rakic, 1992; Westenbroek et al., 1992; Haws et al., 1993). Deter- mining the cellular distribution of N-type Ca2+ channels in the ELL and EGp would therefore facilitate electrophysi- ological studies of the functional role for Ca2+ conductances in cells of the electrosensory system.
The present study examines the distribution of CgTx binding sites as a marker of putative N-type Ca2+ channels in electrosensory cells of the ELL and EGp of the weakly electric fish Apteronotus leptorhynchus. A punctate distribu- tion of CgTx binding sites consistent with N-type Ca2+ channels was found on pre- and postsynaptic membranes of specific cell classes in these two electrosensory structures. Furthermore, CgTx binding sites are distributed in the ELL with respect to major boundaries between separate topographic maps, and in the eminentia granularis in a manner resembling the developing mammalian cerebellum during granule cell migration. An abstract on this work has been presented elsewhere (Turner and Hawkes, 1992).
The activity of cells within the ELL has been well characterized, with pyramidal cells in each segment respond- ing differentially to the frequency of EOD amplitude modu- lations in vivo (Shumway, 1989). Recordings in an in vitro slice preparation reveal both a slow oscillatory activity in ELL circuitry (Mathieson and Maler, 1988; Turner et al., 1991) and an intrinsic fast oscillatory spike discharge in ELL pyramidal cells (Turner et al., 1994). Previous work provides further evidence of oscillatory activity in pyrami- dal cells during the presentation of EOD field modulations in vivo (Bastian, 1981; Metzner and Heiligenberg, 1991). Descending parallel fiber input from EGp granule cells acts as an important gain control mechanism in the ELL that can modify pyramidal cell receptive field properties (Bas- tian, 1986). The importance of determining the distribution of ion channels when examining the basis for electrosen- sory processing was emphasized in a recent demonstration that fast oscillatory spike discharge in ELL pyramidal cells results from a reciprocal depolarization of somatic and dendritic membranes by voltage-dependent Na+ channels (Turner et al., 1994). A role for Ca2+ channels in determin- ing ELL circuit output has also been suggested by the recording of Ca2 '-spikes in pyramidal cells (Mathieson and
C CCb CLS CMS DNL EGm EGP ELL GCL LS MS mol PCL
Abbreviations
cerebello-medullary cistern corpus cerebelli centrolateral segment centromedial segment deep neuropil layer eminentia granularis pars medialis eminentia granularis pars posterior electrosensory lateral line lobe granule cell layer lateral segment medial segment molecular layer pyramidal cell layer
MATERIALS AND METHODS Slice preparation
Weakly electric gymnotiform fish (Apteronotus leptorhyn- chus; Brown Ghost Knife; n > 50) were obtained from local importers and maintained at 26-28°C in aerated fresh water aquaria. Transverse brain slices were prepared accord- ing to Turner et al. (1994). Briefly, fish of 11-15 cm length were anaesthetized by immersion in aquarium water con- taining 0.05% 2-phenoxy-ethanol and then transferred to a custom-made stereotaxic apparatus for superfusion of the gills with oxygenated aquarium water (95% 02/5% COz) containing 0.05% 2-phenoxy-ethanol. The skin and cra- nium were removed under microscopic observation and the brain superfused with cold (4°C) oxygenated artificial cere- brospinal fluid (ACSF) consisting of: 124 mM NaC1,2.0 mM KCl, 1.25 mM KH2P04, 1.5 mM CaC12, 1.5 mM MgS04, 24 mM NaHC03, and 10 mM D-glucose, pH 7.4 (all chemicals were obtained from SIGMA unless otherwise noted). A mini-scalpel was used to transect the brain at the mid-tectal level at approximately 90" to the longitudinal axis. The rostra1 surface of the blocked brain was attached to a pre-cooled block with cyano-acrylate glue, and the brain surrounded by a 20% solution of gelatinidistilled water (40°C). Transverse 300 pm slices of the medulla were cut on a Vibratome containing cold oxygenated ACSF and the gelatin cut away from each slice with forceps and iridec- tomy scissors. Slices were then transferred to a Petri dish containing cold ACSF bubbled with 95% 0215% COa gas.
CgTx treatment A stock solution of CgTx (20 pM; Molecular Probes) was
made up in a medium consisting of 148 mM NaCl, 3.25 mM KCl, 1.5 mM CaC12, 1.5 mM MgCIZ, 10 mM N-2-hydroxy- ethanepiperizine-Nf-2-ethane sulphonic acid (HEPES), and 10 mM D-glucose, pH 7.4 (osmolarity = 310 mmolikg. A final dilution of CgTx (range of 10 nM-1 PM) was made up in standard ACSF or HEPES-buffered medium (above).
Slices were placed in 30 p1 aliquots of 0.5 pM CgTx contained within 8 mm diameter frosted circular wells on a

458 Y. THARANI ET AL.
ground peroxidase activity was eliminated in sections to be reacted with streptavidin-HRP by using 0.3% H202/PBS for 10 minutes, and HRP revealed using diaminobenzidine (0.2 mgiml) in 0.1 M sodium acetate buffer (pH 6.0), re- acted with 0.2% glucose and 0.075% glucose-oxidase. Sec- tions were then dehydrated through an ethanol series and xylene prior to coverslipping in Permount (Fisher Scientific).
The density of cells exhibiting CgTx immunolabel was determined by counting labeled cells on photographic mon- tages of tissue sections (1-6 per animal) and expressed as cells per 100 pm2 area. The dorsal boundary of the EGp granule cell layer was estimated by drawing a line from an inflection that is apparent on the lateral edge of a section between the corpus cerebelli (CCb) and the transitional zone (Sas and Maler, 1987) along a ventromedial course to the cerebello-medullary cistern perpendicular to the gran- ule and molecular layer boundary. In some cases, the EGp granule cell layer was further subdivided into three areas of equal width along the dorso-ventral axis to quantify CgTx- labeled cells across the granule cell layer. The boundaries between segments in the ELL were estimated by identifying characteristic irregularities in the deep neuropil and pyra- midal cell layers. The corresponding molecular layer bound- aries were then estimated by drawing lines from the pyramidal cell layer boundaries to the cerebella-medullary cistern following the dorso-medial curvature of dendritic or glial structures that extend from the pyramidal cell layer. The large surface area of the EGp granule cell layer allowed us to obtain counts of cell density over a minimum of ten 100 pmz sample areas per section. Measurements of tissue area within the more irregular boundaries of the ELL cell or molecular layers were obtained by digitizing an overlay of these boundaries through image capture (Image, NIH), and measuring each area using a drawing tool calibrated to the corresponding scale bar on a Macintosh-based imaging program (Image, NIH). Average values were expressed as mean 5 standard deviation and statistical significance was calculated for two samples using an unpaired Student’s t-test, and for multiple samples by a one-way ANOVA followed by a post hoc pairwise comparison using Tukey’s protected t-test. The number of cases used for each calcula- tion refers to the number of animals unless otherwise noted.
Cell structure was identified in some cases by backload- ing cells with Neurobiotin in 400 pm tissue slices main- tained in an in vitro recording chamber (Turner et al., 1994). Neurobiotin in crystal form was placed over the ELL and/or EGp molecular layer and pushed into the tissue using a glass electrode of 20-50 pm tip diameter. After 4-8 hours the slices were removed from the chamber, fixed in 4% PARA overnight, and cryostat-cut in 10 pm sections. Sections were treated with 0.3% Triton-X 100 for 30 minutes, washed in PBS (3 x 20 min), and reacted with streptavidin-CY-3 (1:1,500) for 45 minutes. Cells were photographed using reflected light fluorescence and a CY-3 filter set, brightfield optics, or differential interference contrast optics (Olympus BH-2).
microscope slide (Cel-line) for 30 minutes at room tempera- ture, and during this time were superfused with 95% OJ5% C02 gas through polyethylene tubing under a covering dish. Except where otherwise noted, this and all other incuba- tions were performed at room temperature. The slices were then transferred to 24 mm diameter multiwell culture dishes (Falcon) and washed with 7-10 changes of oxygen- ated medium for 30 minutes while superfusing 95% 02/5c/c C02 gas under the Petri dish lid. The viability of slices subjected to the full test and wash procedures was verified in some cases by subsequent electrophysiological record- ings in an in vitro slice chamber. Slices were fixed flat on the surface of filter paper presoaked in 4% paraformaldehyde (PARA) in 0.1 M phosphate buffer supplemented with 0.9% NaCl (PBS) for 5 minutes, covered with PARA/PBS, and postfixed overnight at 4°C.
For studies examining competition between Caz+ and CgTx, all ACSF used during slice preparation consisted of Ca”--free HEPES-buffered medium (as above). Slices were preincubated for 30 minutes in cold, oxygenated HEPES- buffered ACSF containing either 0 mM, 1.5 mM, or 30 mM Ca2*. Slices were maintained in the corresponding Ca2+ concentration in HEPES-buffered ACSF during subse- quent exposure to 0.5 pM CgTx (30 min) and post incuba- tion washes (30 min). For studies examining the effects of nifedipine (5 pM; ICN Biochemicals) and AgaToxin-IVA ( 100 nM; Research Biochemicals) on CgTx binding, slices were preincubated in oxygenated ACSF containing nifedi- pine and AgaToxin-IVA for 15-30 minutes. Slices were then co-treated in 0.5 pM CgTx for 30 minutes and subsequently washed in oxygenated ACSF containing nifedipine and AgaToxin-IVA for 10-15 minutes, followed by 20-30 min- utes in ACSF. Nifedipine was initially made as a 1 mM stock solution in dimethyl sulfoxide and then diluted to the appropriate concentration in ACSF.
Immunocytochemistry Binding sites for CgTx were revealed using a monoclonal
antibody directed against CgTx (Fortier et al., 1991; gift of R.B. Hawkes) and standard immunocytochemical proce- dures. Following overnight fixation, slices were rinsed in PBS for 1-3 hours and cryoprotected in 10% sucrose/PBS before rapid freezing on the cut surface of a frozen block of OCT compound ( - -“24C). Tissue sections of 8-12 pm thickness were cut by cryostat and thaw-mounted on poly-D-lysine coated microscope slides, air-dried for 1 hour and washed in PBS (3 x 10 min) and PBS containing 3% normal goat serum (NGS, 10-30 min, Amersham). Tissue was exposed to anti-CgTx (2:l; Fortier et al., 1991) in NGSiPBS overnight (4°C) and subsequently washed in NGSIPBS (3 x 10 min; all subsequent reactions carried out at room temperature). A biotin-conjugated sheep-anti- mouse IgG (1:200; Amersham) was applied in NGSiPBS (45-60 min) and sections washed in NGSiPBS (3 x 10 min). The tertiary label consisted of streptavidin-conju- gated CY-3 ( 1: 1500) or streptavidin-conjugated horseradish peroxidase (HRP; 1 : l O O ; Amersham) in PBS (30-45 rnin), with subsequent washes in PBS (3 x 10 rnin). Control experiments consisted of omission of CgTx, primary anti- body, secondary antibody, or substitution of the primary antibody with the medium used to propagate the monoclo- nal antibody.
Sections stained with CY-3 were coverslipped in antifade medium (90% glycerol/O.l% p-phenylenediamine in PBS; PH 10). sealed with nail polish. and stored at -20°C. Back-
RESULTS Electrosensory cells in Apteronotus medulla The major laminae and cell types of the ELL and EGp
have been previously described (Finger, 1978, 1983; Maler, 1979; Maler et al., 1981; Mathieson et al., 1987; Sas and Maler. 1987: Maler and Mumaini. 1994). but will be brieflv summarized here. The ELL contains 4 ’topographic ma,”,
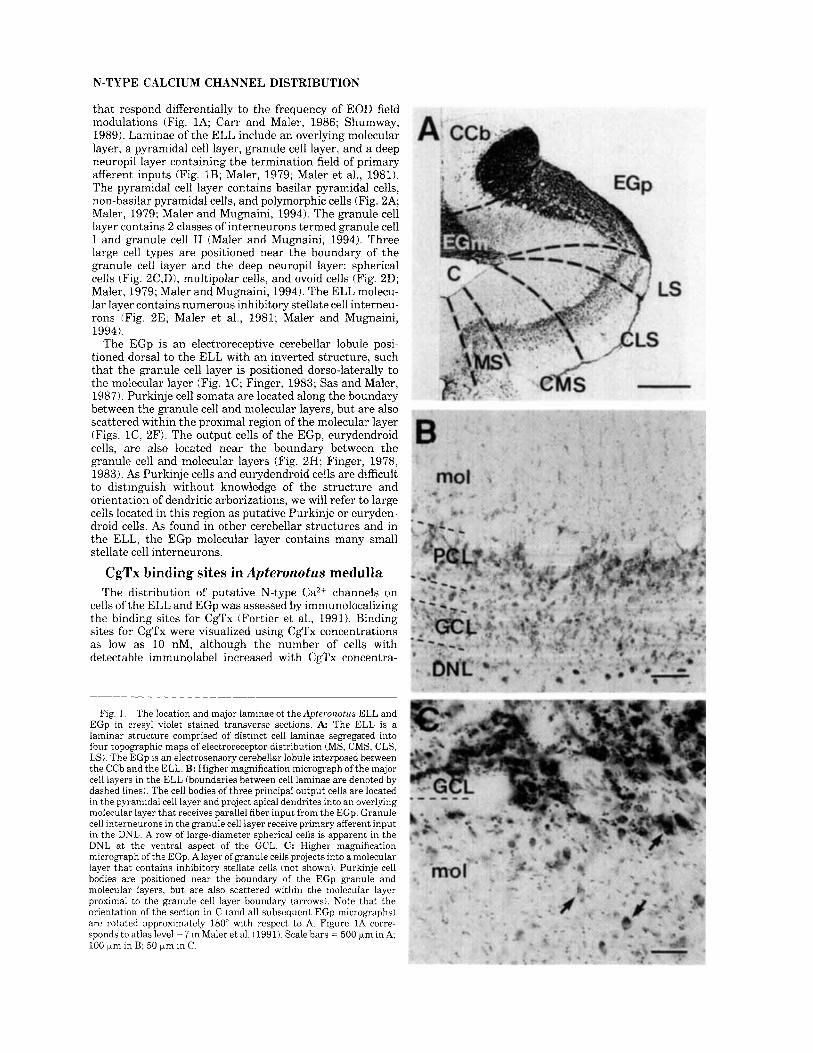
N-TYPE CALCIUM CHANNEL DISTRIBUTION
that respond differentially to the frequency of EOD field modulations (Fig. 1A; Carr and Maler, 1986; Shumway, 1989). Laminae of the ELL include an overlying molecular layer, a pyramidal cell layer, granule cell layer, and a deep neuropil layer containing the termination field of primary afferent inputs (Fig. 1B; Maler, 1979; Maler et al., 1981). The pyramidal cell layer contains basilar pyramidal cells, non-basilar pyramidal cells, and polymorphic cells (Fig. 2A; Maler, 1979; Maler and Mugnaini, 1994). The granule cell layer contains 2 classes of interneurons termed granule cell I and granule cell I1 (Maler and Mugnaini, 1994). Three large cell types are positioned near the boundary of the granule cell layer and the deep neuropil layer: spherical cells (Fig. 2C,D), multipolar cells, and ovoid cells (Fig. 2D; Maler, 1979; Maler and Mugnaini, 1994). The ELL molecu- lar layer contains numerous inhibitory stellate cell interneu- rons (Fig. 2E; Maler et al., 1981; Maler and Mugnaini, 1994).
The EGp is an electroreceptive cerebellar lobule posi- tioned dorsal to the ELL with an inverted structure, such that the granule cell layer is positioned dorso-laterally to the molecular layer (Fig. 1C; Finger, 1983; Sas and Maler, 1987). Purkinje cell somata are located along the boundary between the granule cell and molecular layers, but are also scattered within the proximal region of the molecular layer (Figs. lC, 2F). The output cells of the EGp, eurydendroid cells, are also located near the boundary between the granule cell and molecular layers (Fig. 2H; Finger, 1978, 1983). As Purkinje cells and eurydendroid cells are difficult to distinguish without knowledge of the structure and orientation of dendritic arborizations, we will refer to large cells located in this region as putative Purkinje or euryden- droid cells. As found in other cerebellar structures and in the ELL, the EGp molecular layer contains many small stellate cell interneurons.
CgTx binding sites in Apteronotus medulla The distribution of putative N-type Ca2+ channels on
cells of the ELL and EGp was assessed by immunolocalizing the binding sites for CgTx (Fortier et al., 1991). Binding sites for CgTx were visualized using CgTx concentrations as low as 10 nM, although the number of cells with detectable immunolabel increased with CgTx concentra-
Fig. 1. The location and major laminae of the Apteronotus ELL and EGp in cresyl violet stained transverse sections. A The ELL is a laminar structure comprised of distinct cell laminae segregated into four topographic maps of electroreceptor distribution (MS, CMS, CLS, LS). The EGp is an electrosensory cerebellar lobule interposed between the CCb and the ELL. B: Higher magnification micrograph of the major cell layers in the ELL (boundaries between cell laminae are denoted by dashed lines). The cell bodies of three principal output cells are located in the pyramidal cell layer and project apical dendrites into an overlying molecular layer that receives parallel fiber input from the EGp. Granule cell interneurons in the granule cell layer receive primary afferent input in the DNL. A row of large-diameter spherical cells is apparent in the DNL a t the ventral aspect of the GCL. C: Higher magnification micrograph of the EGp. A layer of granule cells projects into a molecular layer that contains inhibitory stellate cells (not shown). Purkinje cell bodies are positioned near the boundary of the EGp granule and molecular layers, but are also scattered within the molecular layer proximal to the granule cell layer boundary (arrows). Note that the orientation of the section in C (and all subsequent EGp micrographs) are rotated approximately 180" with respect to A. Figure 1A corre- sponds to atlas level - 7 in Maler et al. (1991). Scale bars = 500 Fm in A 100 pm in B; 50 pm in C.
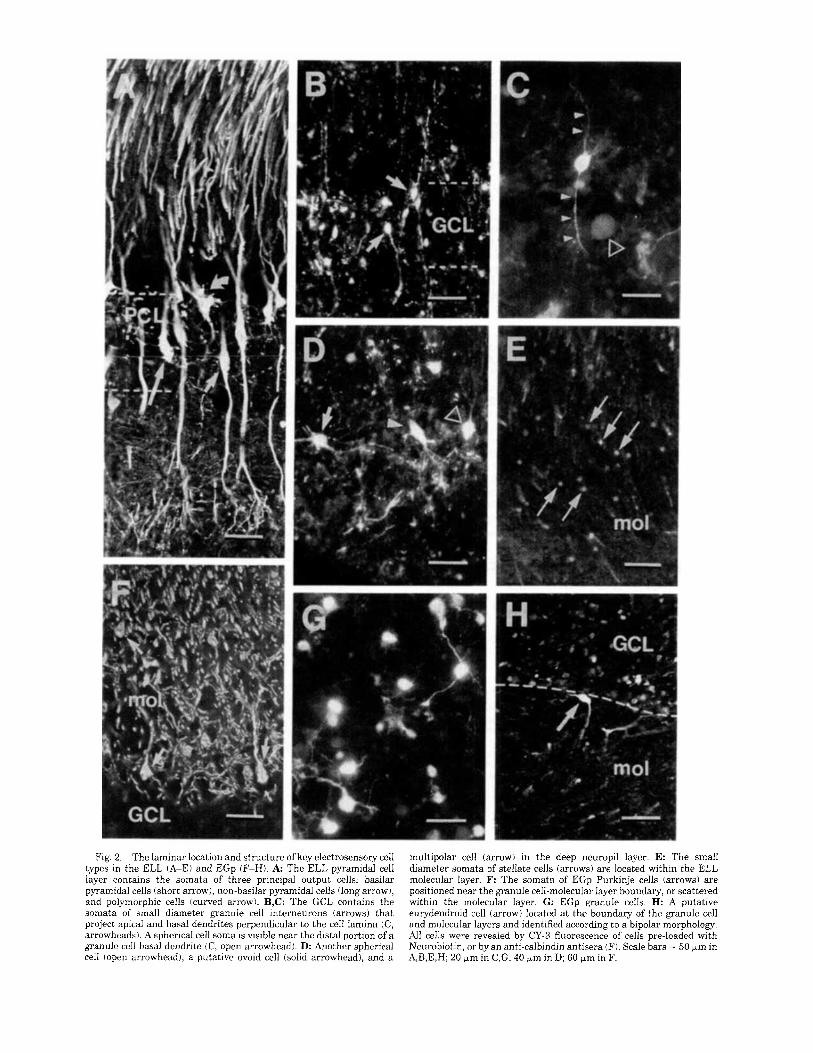
Fig. 2. The laminar location and structure of key electrosensory cell types in the ELL (A-E) and EGp (F-H). A The ELL pyramidal cell layer contains the somata of three principal output cells: basilar pyramidal cells (short arrow), non-basilar pyramidal cells (long arrow), and polymorphic cells (curved arrow). B,C: The GCL contains the somata of small diameter granule cell interneurons (arrows) that project apical and basal dendrites perpendicular to the cell lamina (C, arrowheads). A spherical cell soma is visible near the distal portion of a granule cell basal dendrite (C, open arrowhead). D: Another spherical cell (open arrowhead), a putative ovoid cell (solid arrowhead), and a
multipolar cell (arrow) in the deep neuropil layer. E: The small diameter somata of stellate cells (arrows) are located within the ELL molecular layer. F: The somata of EGp Purkinje cells (arrows) are positioned near the granule cell-molecuiar layer boundary, or scattered within the molecular layer. G: EGp granule cells. H: A putative eurydendroid cell (arrow) located a t the boundary of the granule cell and molecular layers and identified according to a bipolar morphology. All cells were revealed by CY-3 fluorescence of cells pre-loaded with Neurobiotin, or by an anti-calbindin antisera (F). Scale bars = 50 )*m in A,B,E,H; 20 wm in C,G; 40 Fm in D; 60 p,m in F.
N-TYPE CALCIUM CHANNEL DISTRIBUTION 461
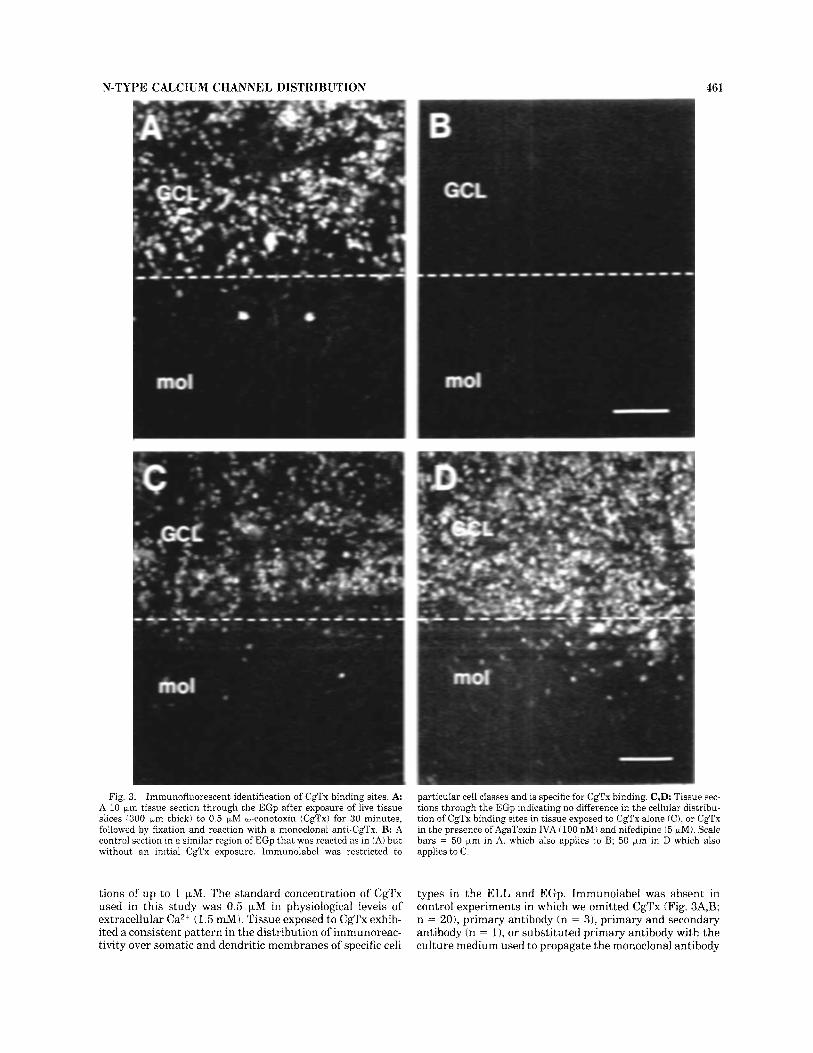
Fig. 3. Immunofluorescent identification of CgTx binding sites. A A 10 pm tissue section through the EGp after exposure of live tissue slices 1300 pm thick) to 0.5 FM w-conotoxin (Cgl’x) for 30 minutes, followed by fixation and reaction with a monoclonal anti-CgTx. B: A control section in a similar region of EGp that was reacted as in (A) but without an initial CgTx cxposure. Immunolabel was restricted to
particular cell classes and is specific for CgTx binding. C,D: Tissue sec- tions through the EGp indicating no difference in the cellular distribu- tion of CgTx binding sites in tissue exposed to CgTx alone (C), or CgTx in the presence ofAgaToxin IVA (100 nM) and nifedipine (5 pM). Scale bars = 50 pm in A, which also applies to B; 50 Fm in D which also applies to C.
tions of up to 1 pM. The standard concentration of CgTx used in this study was 0.5 FM in physiological levels of extracellular Ca2+ (1.5 mM). Tissue exposed to CgTx exhib- ited a consistent pattern in the distribution of immunoreac- tivity over somatic and dendritic membranes of specific cell
types in the ELL and EGp. Immunolabel was absent in control experiments in which we omitted CgTx (Fig. 3A,B; n = ZO), primary antibody (n = 31, primary and secondary antibody (n = 11, or substituted primary antibody with the culture medium used to propagate the monoclonal antibody

462 Y. THARANI ET AL.
(n = 1). The distribution and average number of cells exhibiting CgTx binding was also unaffected by co-incuba- tion with AgaToxin IVA (100 nM) or nifedipine (5 pM; Fig. 3C,D), antagonists of P- and L-type Ca2+ channels, respec- tively (Olivera et al., 1994). Specifically, we found no significant difference in the density of immunolabeled EGp granule cells per 100 pm2 area between tissue sections exposed to CgTx alone (76.9 t_ 16.53; n = 2) or CgTx in the presence of AgaToxin and nifedipine (79.2 t 12.63; n = 2; P < 0.25). A similar distribution of immunolabel was detected with biotin-conjugated CgTx but with a lowered sensitivity, as predicted by previous binding studies (Jones et al., 1989).
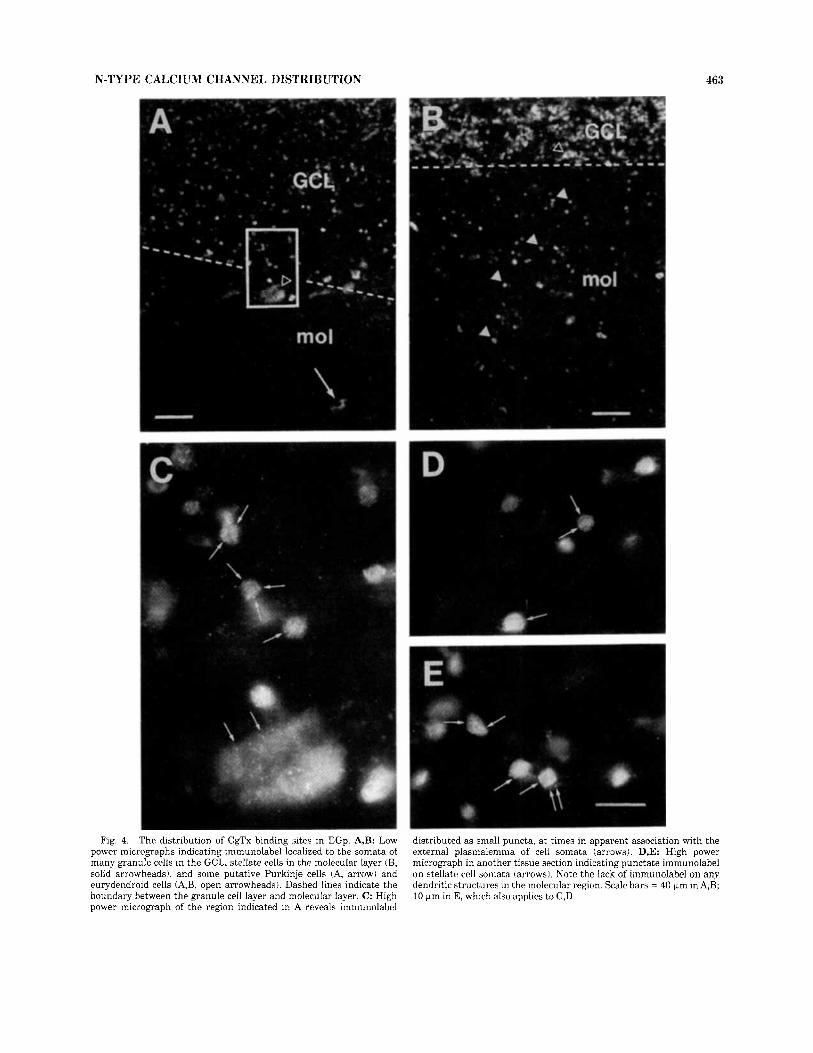
In the EGp, CgTx immunola- be1 was most often associated with the somata of granule cells, and with stellate cells in the molecular layer (Fig. 4A,B). In some cases, the cell bodies of Purkinje cells and eurydendroid cells were also labeled (Fig. 4A). No immuno- label was found in association with dendritic membranes of any labeled cell type, with synaptic glomeruli in the granule cell layer, or with parallel fibers in the molecular layer. At 100 x magnification, punctate CgTx immunolabel was de- tected on both EGp granule cells (Fig. 4C) and stellate cells (Fig. 4D,E). This label was often positioned near the plasmalemmal surface in a manner suggestive of membrane- associated Ca2+ channels (Fig. 4C-E). In most putative Purkinje and eurydendroid cells, CgTx immunolabel was distributed uniformly across the somata, although punc- tate labeling could be detected in some cases (Fig. 4C). Similar CgTx binding sites were observed on granule and stellate cells of the eminentia granularis pars medialis (EGm) and the CCb (not shown).
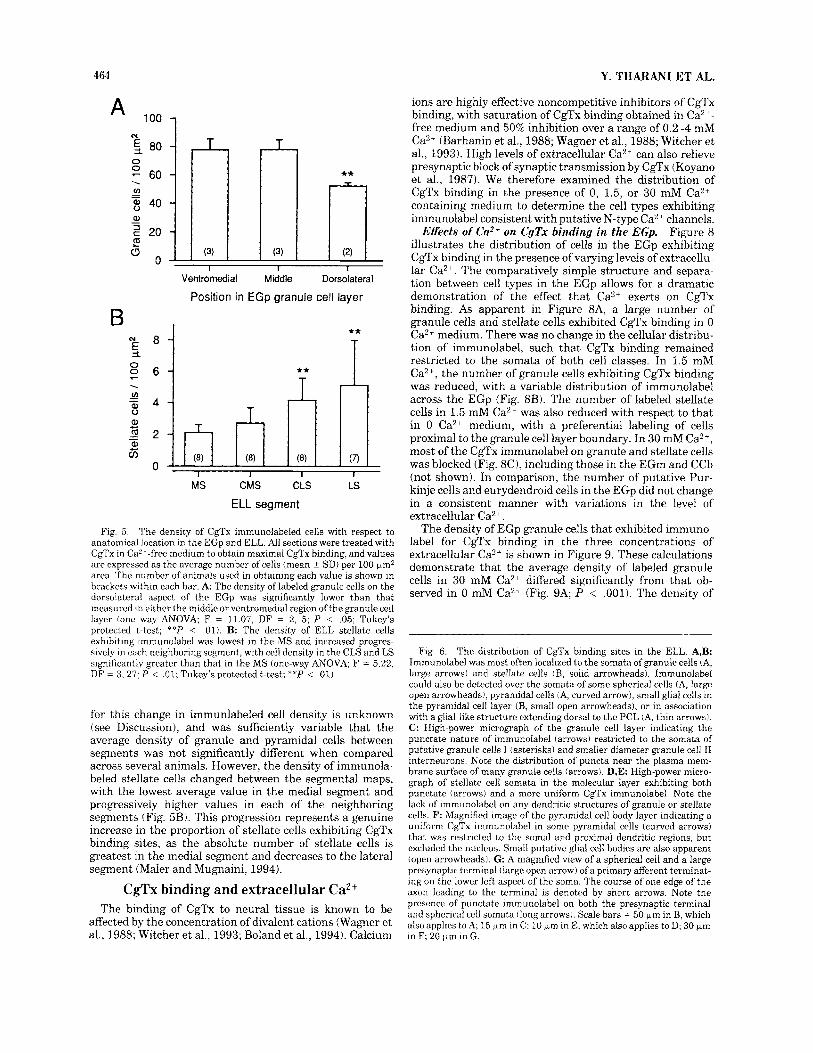
Although there are no established anatomical or func- tional subdivisions within the EGp granule cell layer, we often detected an apparent difference in the number of CgTx immunolabeled cells across this structure (Fig. 8B). We therefore compared the density of granule cells across the cell layer by separately examining three regions of equal width running parallel to the granule cell-molecular layer boundary. This revealed a gradient in the density of cells exhibiting CgTx binding sites across the EGp granule cell layer, such that the ventromedial area bordering the molecu- lar layer and the adjacent region had a significantly greater density of labeled granule cells than the dorsolateral region (Fig. 5A). In comparison, there was no significant difference in the density of immunolabeled granule cells between rostra1 and caudal sections (not shown; n = 8 sections, 1 animal). Counts of the number of stellate cells through the EGp molecular layer could not be obtained, as this region also contains cells migrating from the EGm that compli- cates cell identification in this region (Zupanc and Horschke, 1995; Zupanc et al., 1995). Putative Purkinje and euryden- droid cells were labeled too infrequently to obtain reliable counts.
In the ELL, CgTx immunola- be1 was most often associated with the somata of granule cells and stellate cells (Figs. 6A-E, 7) and less frequently with the somata of polymorphic and pyramidal cells (Figs. 6A,F; 7). Immunolabel could also be detected in two classes of glial cells. In one case the label was restricted to the somata of a small glial cell class localized to the pyramidal cell body layer (Figs. 6B, 7; Maler and Mugnaini, 1994; Turner and Borg, 1995). In the second, immunolabel was found over fenestrated processes that extended 50 pm above the pyramidal cell body layer (Figs. 6A, 7); structures
CqTx binding in the EGp.
CgTx binding in the ELL.
that we have previously shown to be GFAP immunoreactive (Turner and Borg, 1995).
At lOOx magnification, CgTx binding was observed on ELL granule and stellate cell somata primarily as a punc- tate immunolabel (Fig. 6C-E). Two cell types consistent with granule cells I and I1 were distinguished. The granule cell I had a larger cell body (11 pm diameter) and a well defined nucleus and cytoplasm, while granule cell I1 somata were more numerous and smaller in diameter (6 pm diameter) with relatively larger nuclei (Fig. 6C; Maler et al., 1981; Maler and Mugnaini, 1994). A punctate CgTx immu- nolabel was distributed over the region of granule cell I and I1 somata, with immunolabel near the plasmalemmal sur- face most clearly distinguished on the smaller granule cell I1 subtype (Fig. 6 0 . We did not detect label extending over the apical or basal dendrites of either granule cell class. Punctate immunolabel could also be detected near the plasma membrane surface of stellate cell somata (Fig. 6D,E), but not on dendritic membranes of stellate cells or the larger diameter apical dendrites of pyramidal cells in the molecular layer. As found for some cells in the EGp, CgTx immunolabel on ELL pyramidal, spherical, polymor- phic, glial, and some stellate cells appeared more uniform in the soma1 region (Fig. 6F,G). However, we were also able to visualize punctate CgTx immunolabel near the surface of some ELL spherical cells (Fig. 6G), and in association with the large presynaptic terminals of primary afferents (T-type electroreceptors) that synapse on spherical cell somata (Fig. 6G; Maler et al., 1981).
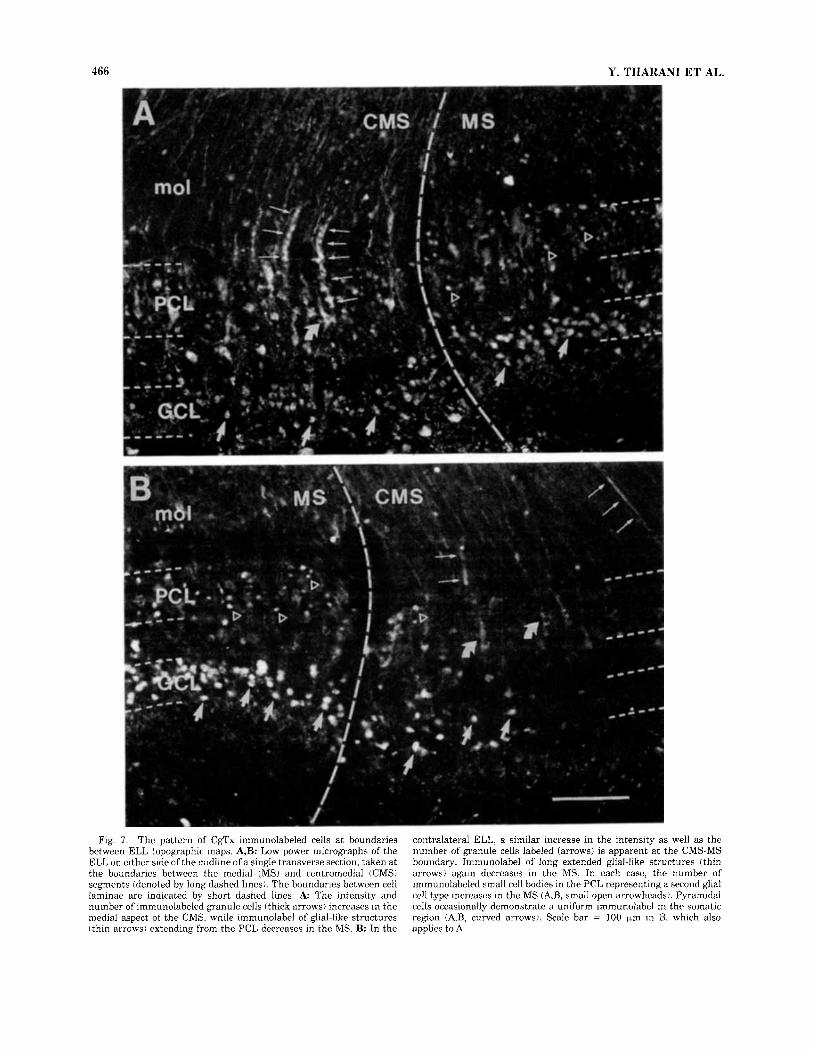
C9Tx binding across ELL segmental maps. As stated above, the cell classes exhibiting CgTx immunolabel were consistent in both the EGp and ELL. However, the inten- sity and number of cells that labeled could vary, a result that was more obvious in the ELL than the EGp. This was detected both within a single tissue section and between experimental animals. Local variations in CgTx binding affinity likely contribute to this result, in part through competition with extracellular Ca2+ (see below). Differences between experimental animals may also be related to age, as the distribution of CgTx binding sites in mammals changes during postnatal development (Komuro and Rakic, 1992; Filloux et al., 19941, and the fraction of N-type Ca2+ current blocked by CgTx is greater in younger as compared to adult rats (Ikeda, 1991; Regan et al., 1991; Zhu and Ikeda, 1993; Boland et al., 1994). In this regard, the age of animals used in our study was not determined and could differ consider- ably, as the rate of teleost growth will vary according to environmental factors. However, close examination indi- cates that at least some of this variability was attributable to changes in the density of CgTx immunolabeled cells near the boundaries of ELL topographic maps. Thus, the inten- sity and number of CgTx labeled cells could change near segment boundaries; a pattern that was often consistent through several sections cut from the same animal. On the other hand, this “segment-associated” label could differ between the ELLs on either side of one tissue section.
Figure 7 compares the distribution of CgTx immunola- beled cells near the boundary between the medial and centromedial segments of the ELL on either side of one tissue section. In this case, granule cells exhibiting CgTx binding sites were prominent in the medial segment of both ELLs, while their numbers decreased sharply in the medial aspect of the neighboring centromedial segment. In con- trast, the number of labeled glial or pyramidal cells in- creased in the centromedial segment (Fig. 7). The reason
N-TYPE CALCIUM CHANNEL DISTRIBUTION 463
Fig. 4. The distribution of CgTx binding sites in EGp. A,B: Low power micrographs indicating immunolabel localized to the somata of many granule cells in the GCL, stellate cells in the molecular layer (B, solid arrowheads), and some putative Purkinje cells (A, arrow) and eurydendroid cells (A,B, open arrowheads). Dashed lines indicate the boundary between the granule cell layer and molecular layer. C: High power micrograph of the region indicated in A reveals immunolabel
distributed as small puncta, a t times in apparent association with the external plasmalemma of cell somata (arrows). D,E: High power micrograph in another tissue section indicating punctate immunolabel on stellate cell somata (arrows). Note the lack of immunolabel on any dendritic structures in the molecular region. Scale bars = 40 +m in A,B; 10 pm in E, which also applies to C,D.
464 Y. THARANI ET AL.
loo 1 A
N 5 80 - 60 8 40
0 0 . v) - -
" I I I
Ventrornedial Middle Dorsolateral
Position in EGp granule cell layer
B I
.
**
I ** T
MS CMS CLS LS
ELL segment
Fig. j. The density of CgTx immunolabeled cells with respect to anatomical location in the EGp and ELL. All sections were treated with CgTx in CaL--free medium to obtain maximal CgTx binding, and values are expressed as the average number of cells (mean ? SDI per 100 pm2 area. The number of animals used in obtaining each value is shown in brackets within each bar. A: The density of labeled granule cells on the dorsolateral aspect of the EGp was significantly lower than that measured in either the middle or ventromedial region of the granule cell layer (one way ANOVA; F = 11.07, DF = 2, 5; P < .05; Tukey's protected t-test; **P < .01). B: The density of ELL stellate cells exhibiting immunolabel was lowest in the MS and increased progres- sively in each neighhoringsegment, with cell density in the CLS and LS significantly greater than that in the MS (one-way ANOVA; F = 5.22, DF = 3, 27; P < .01; Tukey's protected t-test; **P < .01j.
for this change in immunlabeled cell density is unknown (see Discussion), and was sufficiently variable that the average density of granule and pyramidal cells between segments was not significantly different when compared across several animals. However, the density of immunola- beled stellate cells changed between the segmental maps, with the lowest average value in the medial segment and progressively higher values in each of the neighboring segments (Fig. 5B). This progression represents a genuine increase in the proportion of stellate cells exhibiting CgTx binding sites, as the absolute number of stellate cells is greatest in the medial segment and decreases to the lateral segment (Maler and Mugnaini, 1994).
CgTx binding and extracellular Ca2+ The binding of CgTx to neural tissue is known to be
affected by the concentration of divalent cations (Wagner et al., 1988; Witcher et al., 1993; Boland et al., 1994). Calcium
ions are highly effective noncompetitive inhibitors of CgTx binding, with saturation of CgTx binding obtained in Ca2+- free medium and 50% inhibition over a range of 0.2-4 mM Ca2+ (Barhanin et al., 1988; Wagner et al., 1988; Witcher et al., 1993). High levels of extracellular Ca2+ can also relieve presynaptic block of synaptic transmission by CgTx (Koyano et al., 1987). We therefore examined the distribution of CgTx binding in the presence of 0, 1.5, or 30 mM Ca2+- containing medium to determine the cell types exhibiting immunolabel consistent with putative N-type Ca2+ channels.
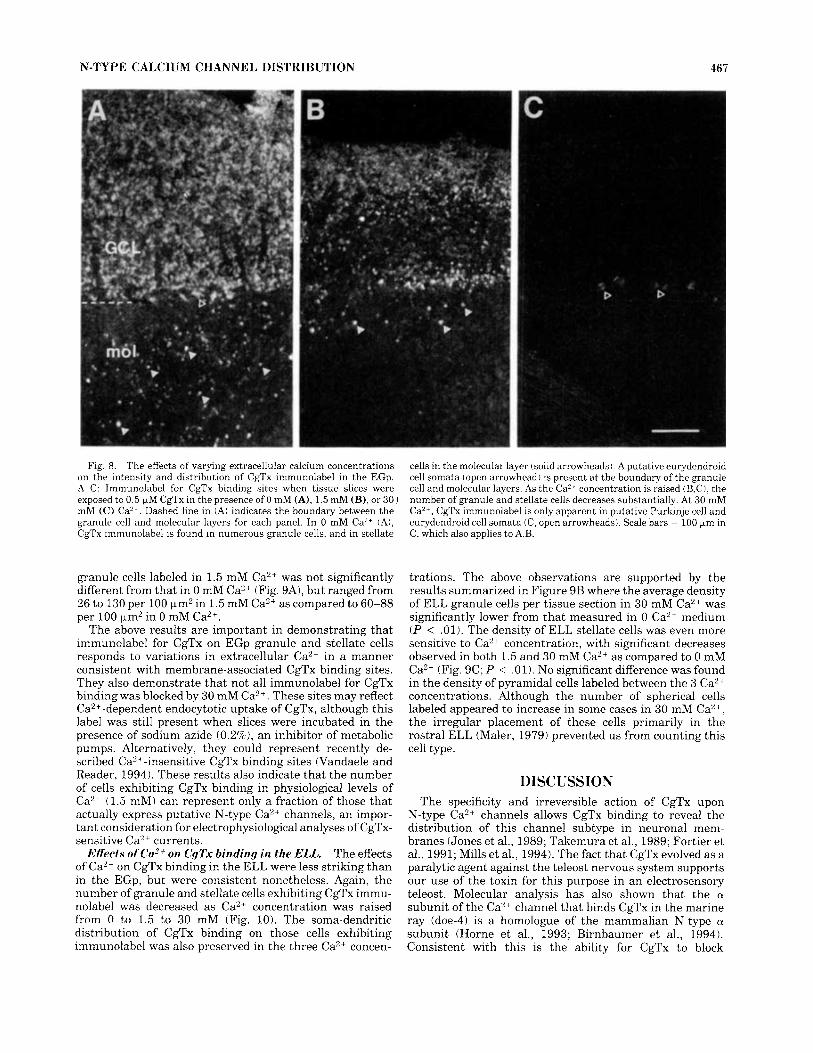
Figure 8 illustrates the distribution of cells in the EGp exhibiting CgTx binding in the presence of varying levels of extracellu- lar Ca2+. The comparatively simple structure and separa- tion between cell types in the EGp allows for a dramatic demonstration of the effect that Ca2+ exerts on CgTx binding. As apparent in Figure 8A, a large number of granule cells and stellate cells exhibited CgTx binding in 0 Ca2+ medium. There was no change in the cellular distribu- tion of immunolabel, such that CgTx binding remained restricted to the somata of both cell classes. In 1.5 mM Ca2+, the number of granule cells exhibiting CgTx binding was reduced, with a variable distribution of immunolabel across the EGp (Fig. 8B). The number of labeled stellate cells in 1.5 mM Ca2' was also reduced with respect to that in 0 Ca2+ medium, with a preferential labeling of cells proximal to the granule cell layer boundary. In 30 mM Ca2+, most of the CgTx immunolabel on granule and stellate cells was blocked (Fig. 8C), including those in the EGm and CCb (not shown). In comparison, the number of putative Pur- kinje cells and eurydendroid cells in the EGp did not change in a consistent manner with variations in the level of extracellular Ca2+.
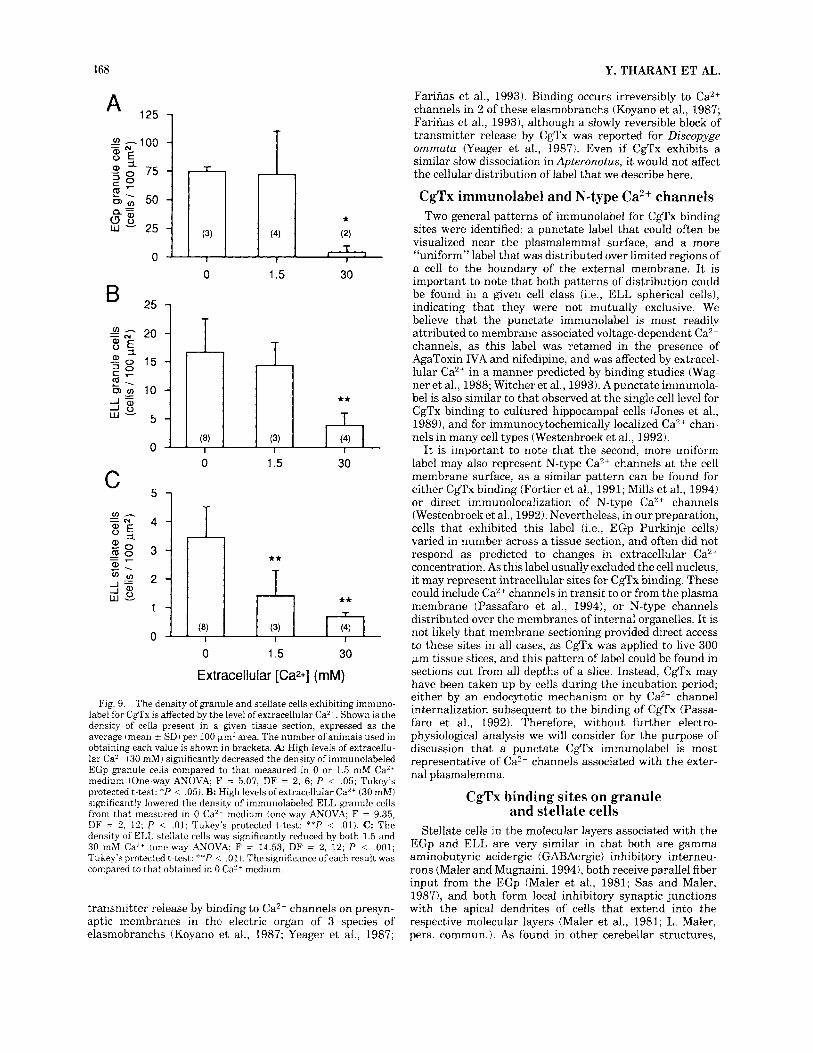
The density of EGp granule cells that exhibited immuno- label for CgTx binding in the three concentrations of extracellular Ca2+ is shown in Figure 9. These calculations demonstrate that the average density of labeled granule cells in 30 mM Ca2+ differed significantly from that ob- served in 0 mM Ca2+ (Fig. 9A; P < .001). The density of
Effects of Ca" on CgTx binding in the EGp.
Fig. 6. The distribution of CgTx binding sites in the ELL. A,B: Immunolabel was most often localized to the somata of granule cells (A, large arrows) and stellate cells (B, solid arrowheads). Immunolabel could also be detected over the somata of some spherical cells (A, large open arrowheads), pyramidal cells (A, curved arrow), small glial cells in the pyramidal cell layer (B, small open arrowheads), or in association with a glial-like structure extending dorsal to the PCL (A, thin arrows). C: High-power micrograph of the granule cell layer indicating the punctate nature of immunolabel (arrows) restricted to the somata of putative granule cells I (asterisks) and smaller diameter granule cell I1 interneurons. Note the distribution of puncta near the plasma mem- brane surface of many granule cells (arrows). D , E High-power micro- graph of stellate cell somata in the molecular layer exhibiting both punctate (arrows) and a more uniform CgTx immunolabel. Note the lack of immunolabel on any dendritic structures of granule or stellate cells. F: Magnified image of the pyramidal cell body layer indicating a uniform CgTx immunolabel in some pyramidal cells (curved arrows) that was restricted to the soma1 and proximal dendritic regions, but excluded the nucleus. Small putative glial cell bodies are also apparent (open arrowheads). G A magnified view of a spherical cell and a large presynaptic terminal (large open arrow) of a primary afferent terminat- ing on the lower left aspect of the soma. The course of one edge of the axon leading to the terminal is denoted by short arrows. Note the presence of punctate immunolabel on both the presynaptic terminal and spherical cell somata (long arrows). Scale bars = 50 pm in B, which also applies to A; 15 pm in C; 10 pm in E, which also applies to D; 30 pm in F; 20 pm in G.

N-TYPE CALCIUM CHANNEL DISTRIBUTION 465
Figure 6
466 Y. THARANI ET AL.
Fig. 7 . The pattern of CgTx immunolabeled cells at boundaries between ELL topographic maps. A,B: Low power micrographs of the ELL on either side of the midline of a single transverse section, taken at the boundaries between the medial (MS) and centromedial (CMS) segments (denoted by long dashed lines). The boundaries between cell laminae are indicated by short dashed lines. A The intensity and number of immunolabeled granule cells (thick arrows) increases in the medial aspect of the CMS, while immunolabel of glial-like structures (thin arrows) extending from the PCL decreases in the MS. B: In the
contralateral ELL, a similar increase in the intensity as well as the number of granule cells labeled (arrows) is apparent a t the CMS-MS boundary. Immunolabel of long extended glial-like structures (thin arrows) again decreases in the MS. In each case, the number of immunolabeled small cell bodies in the PCL representing a second glial cell type increases in the MS (A,B, small open arrowheads). Pyramidal cells occasionally demonstrate a uniform immunolabel in the somatic region (A,B, curved arrows). Scale bar = 100 pm in B, which also applies to A.
N-TYPE CALCIUM CHANNEL DISTRIBUTION 467
Fig. 8. The effects of varying extracellular calcium concentrations on the intensity and distribution of CgTx immunolabel in the EGp. A-C: Immunolabel for CgTx binding sites when tissue slices were exposed to 0.5 FM CgTx in the presence of 0 mM (A), 1.5 mM (B), or 30 J mM (C) Ca’*. Dashed line in (A) indicates the boundary between the granule cell and molecular layers for each panel. In 0 mM Ca2+ (A), CgTx immunolabel is found in numerous granule cells, and in stellate
cells in the molecular layer (solid arrowheads). A putative eurydendroid cell somata (open arrowhead) is present a t the boundary of the granule cell and molecular layers. As the Ca’* concentration is raised iB,C), the number of granule and stellate cells decreases substantially. At 30 mM Ca”’, CgTx immunolabel is only apparent in putative Purkinje cell and eurydendroid cell somata iC, open arrowheads). Scale bars = 100 pm in C, which also applies to A.B.
granule cells labeled in 1.5 mM Ca2+ was not significantly different from that in 0 mM Ca2+ (Fig. 9A), but ranged from 26 to 130 per 100 pm2 in 1.5 mM Ca2+ as compared to 60-88 per 100 km2 in 0 mM Ca”.
The above results are important in demonstrating that immunolabel for CgTx on EGp granule and stellate cells responds to variations in extracellular Ca2+ in a manner consistent with membrane-associated CgTx binding sites. They also demonstrate that not all immunolabel for CgTx binding was blocked by 30 mM Ca2+. These sites may reflect Ca”+-dependent endocytotic uptake of CgTx, although this label was still present when slices were incubated in the presence of sodium azide (0.2%), an inhibitor of metabolic pumps. Alternatively, they could represent recently de- scribed Ca”-insensitive CgTx binding sites (Vandaele and Reader, 1994). These results also indicate that the number of cells exhibiting CgTx binding in physiological levels of Ca2+ (1.5 mM) can represent only a fraction of those that actually express putative N-type Ca2+ channels, an impor- tant consideration for electrophysiological analyses of CgTx- sensitive Ca2+ currents.
The effects of Ca2+ on CgTx binding in the ELL were less striking than in the EGp, but were consistent nonetheless. Again, the number of granule and stellate cells exhibiting CgTx immu- nolabel was decreased as Ca2+ concentration was raised from 0 to 1.5 to 30 mM (Fig. 10). The soma-dendritic distribution of CgTx binding on those cells exhibiting immunolabel was also preserved in the three Ca2+ concen-
Effects of Ca2+ on CgTx binding in the ELL.
trations. The above observations are supported by the results summarized in Figure 9B where the average density of ELL granule cells per tissue section in 30 mM Ca2+ was significantly lower from that measured in 0 Ca2+ medium ( P < .01). The density of ELL stellate cells was even more sensitive to Ca2+ concentration, with significant decreases observed in both 1.5 and 30 mM Ca2+ as compared to 0 mM Ca2+ (Fig. 9C; P < .01). No significant difference was found in the density of pyramidal cells labeled between the 3 Ca2+ concentrations. Although the number of spherical cells labeled appeared to increase in some cases in 30 mM Ca2++, the irregular placement of these cells primarily in the rostra1 ELL (Maler, 1979) prevented us from counting this cell type.
DISCUSSION The specificity and irreversible action of CgTx upon
N-type Ca2+ channels allows CgTx binding to reveal the distribution of this channel subtype in neuronal mem- branes (Jones et al., 1989; Takemura et al., 1989; Fortier et al., 1991; Mills et al., 1994). The fact that CgTx evolved as a paralytic agent against the teleost nervous system supports our use of the toxin for this purpose in an electrosensory teleost. Molecular analysis has also shown that the OL
subunit of the Ca2+ channel that binds CgTx in the marine ray (doe-4) is a homologue of the mammalian N-type a subunit (Horne et al., 1993; Birnbaumer et al., 1994). Consistent with this is the ability for CgTx to block
168 Y. THARANI ET AL.
125 A
0 0 1.5 30
25 1 B
I -F
0 1.5 30
Extracellular [Ca2+] (mM)
Fig. 9. The density of granule and stellate cells exhibiting immuno- label for CgTx is affected by the level of extracellular Ca2 +. Shown is the density of cells present in a given tissue section, expressed as the average (mean 2 SD) per 100 pn2 area. The number of animals used in obtaining each value is shown in brackets. A High levels of extracellu- lar CaY’ 130 mM) significantly decreased the density of immunolabeled EGp granule cells compared to that measured in 0 or 1.5 mM Ca” medium (One-way ANOVA; F = 5.07, DF = 2 , 6; P < .05; Tukey’s protected t-test: *P < .05). B: High levels ofextracellular Ca2+ (30 mM) significantly lowered the density of immunolabeled ELL granule cells from that measured in 0 Ca2- medium (one-way ANOVA; F = 9.35, DF = 2, 12; P < .01; Tukey’s protected t-test: **P < .01). C: The density of ELL stellate cells was significantly reduced by both 1.5 and 30 mM Ca2+ (one-way ANOVA; F = 14.53, DF = 2, 12; P < ,001; Tukey’s protected t-test: **P < .01). The significance ofeach result was compared to that obtained in 0 Ca2+ medium.
transmitter release by binding to Ca2+ channels on presyn- aptic membranes in the electric organ of 3 species of elasmobranchs (Koyano et al., 1987; Yeager et al., 1987;
Farinas et al., 1993). Binding occurs irreversibly to Ca2+ channels in 2 of these elasmobranchs (Koyano et al., 1987; Farinas et al., 19931, although a slowly reversible block of transmitter release by CgTx was reported for Discopyge ommata (Yeager et al., 1987). Even if CgTx exhibits a similar slow dissociation in Aptemnotus, it would not affect the cellular distribution of label that we describe here.
CgTx immunolabel and N-type Ca2+ channels Two general patterns of immunolabel for CgTx binding
sites were identified: a punctate label that could often be visualized near the plasmalemmal surface, and a more “uniform” label that was distributed over limited regions of a cell to the boundary of the external membrane. It is important to note that both patterns of distribution could be found in a given cell class (i.e., ELL spherical cells), indicating that they were not mutually exclusive. We believe that the punctate immunolabel is most readily attributed to membrane-associated voltage-dependent Ca2+ channels, as this label was retained in the presence of AgaToxin IVA and nifedipine, and was affected by extracel- lular Ca2+ in a manner predicted by binding studies (Wag- ner et al., 1988; Witcher et al., 1993). A punctate immunola- be1 is also similar to that observed at the single cell level for CgTx binding to cultured hippocampal cells (Jones et al., 1989), and for immunocytochemically localized Ca2+ chan- nels in many cell types (Westenbroek et al., 1992).
It is important to note that the second, more uniform label may also represent N-type Ca2+ channels at the cell membrane surface, as a similar pattern can be found for either CgTx binding (Fortier et al., 1991; Mills et al., 1994) or direct immunolocalization of N-type Ca2+ channels (Westenbroek et al., 19921. Nevertheless, in our preparation, cells that exhibited this label (i.e., EGp Purkinje cells) varied in number across a tissue section, and often did not respond as predicted to changes in extracellular Ca2+ concentration. As this label usually excluded the cell nucleus, it may represent intracellular sites for CgTx binding. These could include Ca2+ channels in transit to or from the plasma membrane (Passafaro et al., 19941, or N-type channels distributed over the membranes of internal organelles. It is not likely that membrane sectioning provided direct access to these sites in all cases, as CgTx was applied to live 300 pm tissue slices, and this pattern of label could be found in sections cut from all depths of a slice. Instead, CgTx may have been taken up by cells during the incubation period; either by an endocytotic mechanism or by Ca2+ channel internalization subsequent to the binding of CgTx (Passa- far0 et al., 1992). Therefore, without further electro- physiological analysis we will consider for the purpose of discussion that a punctate CgTx immunolabel is most representative of CaZ+ channels associated with the exter- nal plasmalemma.
CgTx binding sites on granule and stellate cells
Stellate cells in the molecular layers associated with the EGp and ELL are very similar in that both are gamma aminobutyric acidergic (GABAergc) inhibitory interneu- rons (Maler and Mugnaini, 19941, both receive parallel fiber input from the EGp (Maler et al., 1981; Sas and Maler, 19871, and both form local inhibitory synaptic junctions with the apical dendrites of cells that extend into the respective molecular layers (Maler et al., 1981; L. Maler, pers. commun.). As found in other cerebellar structures,

N-TYPE CALCIUM CHANNEL DISTRIBUTION 469
Fig. 10. The effects of varying extracellular Ca” concentrations on the intensity and distribution of Cg”x immunolabel in the centromedial segment of the ELL. A-C: CgTx immunolabel when tissue slices were exposed to 0.5 GM CgTx in the presence of 0 mM (A), 1.5 mM (B), or 30 mM (C) Ca” . Dashed lines indicate the boundaries of the PCL and GCL. In 0 mM and 1.5 mM Ca2’ (A,B), CgTx immunolabel is found on granule cells (arrows), stellate cells in the molecular layer (solid arrowheads,, and a small diameter glial cell type in the PCL (small open
EGp granule cells receive mossy fiber input and project an axon into the molecular layer to form parallel fiber output (Finger, 1983; Sas and Maler, 1987). Granule cells of the ELL also act as interneurons that receive primary afferent input in the DNL and make synaptic and gap junction contact with pyramidal cell somata (Maler et al., 1981). An important consideration at the functional level is whether the punctate CgTx immunolabel observed on these cells is distributed over pre- or postsynaptic membranes. Virtually all presynaptic input in the EGp and ELL molecular layers is derived from a massive parallel fiber projection frdm the EGp (Maler et al., 1981; Sas and Maler, 1987). This input forms synaptic contact with dendritic membranes of stel- late cells, while very few synaptic contacts can be distin- guished on stellate cell somata at the ultrastructural level (Maler et al., 1981). These factors suggest a postsynaptic localization for the CgTx binding detected on EGp and ELL stellate cell somata.
A pre- or postsynaptic location for CgTx binding to EGp granule cells on this basis can not be discriminated at this time, although a postsynaptic N-type Ca2+ conductance has been recorded in mouse cerebellar granule cells (Haws et al., 1993). The punctate label for CgTx binding on the ELL granule cell I1 may be postsynaptic, as this cell receives excitatory synaptic contact on basilar dendrites (Maler et al., 1981; Mathieson et al., 1987), while virtually no axoso-
arrowheads). Immunolabel on pyramidal cell somata is also present a t 1.5 mM Ca’+ (B, curved arrow). As the Ca2+ concentration is raised to 30 mM (C), the number of granule cells (arrows) and stellate cells (solid arrowheads) decreases substantially, with immunolabel detected more frequently in spherical cells (C, open arrowheads), a polymorphic cell (C, curved arrow), and radially extending glial cell processes within and dorsal to the PCL (C, thin arrows). Scale bars = 100 bm in C, which also applies to A,B.
matic synaptic boutons have been detected (Maler et al., 1981; Maler and Mugnaini, 1994). As the ELL granule cell I subclass receives GABAergic synaptic contact on the soma (Maler and Mugnaini, 1994) the possible association of CgTx binding with synaptic terminals on this cell can not be ruled out.
Regional differences in CgTx immunolabel in the ELL
An interesting result in the present study was the observed increase in the proportion of Cox-labeled stellate cells from the medial to the lateral segmental map (Fig. 5B). Although we could often visualize a change in the density of labeled granule cells near the boundaries of segmental maps (Fig. 7), these differences were not consistent enough to indicate statistical significance. Yet both results are interesting in that pyramidal cells, the output cells of the ELL, exhibit a map-specific frequency sensitivity to EOD amplitude modulations in vivo (Shumway, 1989; Metzner and Heiligenberg, 199 l), and different frequencies of oscilla- tory discharge across segmental maps in vitro (Turner et al., 1991, 1994). Regional variations in CgTx binding may then correspond to functional differences in the activity within each topographic map. Indeed, there is evidence that the level of expression of Ca2+ channels can be modified by

170 Y. THARANI ET AL.
somata, and in a patch-like manner over Purkinje cell dendrites (Fortier et al., 1991; Westenbroek et al., 1992). In contrast to these general findings in mammals, punctate CgTx binding sites were localized almost exclusively to somatic membranes in our preparation; no CgTx immuno- label was found on the dendrites of EGp or ELL granule or stellate cells, or EGp Purkinje cells. Although pyramidal cells of the ELL have large diameter dendrites, CgTx binding sites were detected on these structures only in the proximal dendritic region as a uniform label. We obviously can not rule out the possibility that CgTx binding sites exist on dendritic membranes, but the present data indicates primarily a somatic distribution for the N-type Ca2+ chan- nel in these electrosensory cell types.
Binding sites for CgTx have been localized anatomically to presynaptic membranes in Torpedo electric organ (Farinas et al., 1993), the nerve terminal calyx on chick ciliary ganglion cells (Haydon et al., 1994 ), and mossy fiber terminals in hippocampus (Westen- broek et al., 1992). We have now detected punctate label for CgTx binding on the large presynaptic terminal of primary afferents that contact ELL spherical cells (Fig. 6G). As these terminals are known to exclusively form gap junction connections in Apteronotus (Maler et al., 1981), these channels must participate in functions other than Ca2+ entry for the purpose of transmitter release; perhaps the transmission of spike discharge or the regulation of gap junction conductance. The possible distribution of putative N-type CaZt channels on presynaptic terminals of other afferents in the ELL or EGp could not be resolved at the light microscopic level.
Functional significance to electrosensory processing
Presynaptic distribution.
Some important functional considerations for electrosen- sory processing are apparent, even if we assume that N-type Ca2+ channels are localized only to EGp and ELL granule and stellate cells. The placement of N-type Ca2+ channels on these cells is optimal for modifying the transmission of electrosensory input at the level of primary afferents (via ELL granule cells) or descending feedback pathways to the ELL (via EGp granule and stellate cells, and ELL stellate cells). For instance, GABAergic, noradrenergic, and gluta- matergic afferents capable of modifying N-type Ca2+ chan- nels (Schwartz and Bean, 1992; Diverse-Pierluissi and Dunlap, 1993; Mintz and Bean, 1993) terminate in the ELL granule cell layer (Sas et al., 1990; Maler and Monaghan, 1991; Maler and Mugnaini, 1994). Granule cells are known to shape both local and surround inhibition in ELL pyrami- dal cells (Shumway and Maler, 1989) and are suspected to underlie a slow oscillatory discharge in the ELL (Turner et al., 1991). Therefore, any modification ofN-type Ca2+ chan- nel conductance by transmitter actions could dramatically alter pyramidal cell receptive field properties through the activity of granule cells. The ability of N-type Ca2+ conduc- tance to undergo facilitation with repetitive activation (Ikeda, 1991) may also be particularly relevant to electrosen- sory input, as primary afferents repetitively depolarize ELL cells in the course of encoding beat frequencies of EOD amplitude modulations (Metzner and Heiligenberg, 1991).
several factors, some of which are activity-dependent. Thus, L-type Ca2+ channels can be internalized in response to K+-dependent membrane depolarizations (Liu et al., 19941, and N-type Ca2+ channel expression is increased by nerve growth factor or protein kinase C (Plummer et al., 1989; Knox et al., 1992). Passafaro et al. (1994) also reported an intracellular pool of CgTx-binding sites that generated functional N-type Ca2+ channels when translocated to the plasma membrane in response to a 30 minute exposure to CgTx or Cd+. Notably, this should not affect the distribu- tion of CgTx binding if such a mechanism is present in our tissue, as CgTx was immediately washed from the sections several times following CgTx exposure. Future studies will be required to determine whether activity during electrosen- sory processing is sufficient to modulate the segmental distribution of CgTx binding sites in the ELL.
Comparison of N-type Ca”channe1 distribution in other vertebrate cell types
Cerebellar distribution. The distribution of CgTx bind- ing sites in the EGp contrasts sharply with radioligand studies in adult mammalian cerebellum that demonstrate a predominance of CgTx label in the cerebellar molecular layer and a lower density in the granule cell layer (Take- mura et al., 1989; Albensi et al., 1993; Filloux et al., 1994). Rather, the distribution of CgTx binding sites in Apterono- tus is strikingly similar to that observed in developing mouse cerebellum, where the primary binding site is on cerebellar granule cells, with little label in the molecular layer (Komuro and Rakic, 1992). Expression of this label was shown by Komuro and Rakic (1992) to coincide with the migration of granule cells from the external to the internal granule cell layer. Indeed, the rate of granule cell migration was markedly reduced by CgTx application, implicating N-type Ca2+ channels in the process of cell movement (Komuro and Rakic, 1992). In this regard, Zupanc and Horschke ( 1995) recently demonstrated that in Apteronotus the EGm is a proliferative zone for cell division throughout the life of the animal. Once formed in the EGm, cells migrate through the molecular layer towards the EGp granule cell layer (Zupanc et al., 1995). The binding sites for CgTx in the EGm and EGp are then likely related to the ongoing migration of cells between these granule cell layers in the adult Apteronotus. Interestingly, the distribution of nicotinamide adenine dinucleotide phosphate (NADPH)- diaphorase in EGp Purkinje cells and granule cells is also similar to that in the developing, but not the adult, mammalian cerebellum (Turner and Moroz, 1995). The above results are thus of special interest in that they suggest the adult apteronotid eminentia granularis most closely resembles a developing mammalian cerebellum. The wealth of information on mammalian cerebellar develop- ment should provide several predictions by which to test this hypothesis (Altman, 1982; Hawkes and Mascher, 1994).
Soma-dendritic distribution. Both immunocytochemi- cal and CgTx binding studies report that putative N-type Ca2+ channels are distributed over somatic or dendritic membranes in a cell-specific manner in mammalian neural tissue (Fortier et a]., 1991; Westenbroek et al., 1992; Mills et al., 1994). In general, previous studies have emphasized the localization of this Caz+ channel subtype to dendritic membranes (Westenbroek et al., 1992; Mills et al., 1994). In particular, putative N-type Ca2+ channels were reported over the cell body and proximal dendritic region of cerebel- lar stellate cells (Fortier et al., 1991), over Purkinje cell
ACKNOWLEDGMENTS This work was supported by the Medical Research Coun-
cil of Canada and by the Alberta Heritage Foundation for

N-TYPE CALCIUM CHANNEL DISTRIBUTION 471
Medical Research. We thank R.B. Hawkes and L. Maler for helpful discussions and comments on the manuscript, and L. Borg for expert technical assistance. R.W.T. is a Canada MRC and AHFMR Scholar.
LITERATURE CITED Albensi, 13.C., K.T. Ryujin, J.M. McIntosh, S.R. Naisbitt, B.M. Olivera, and
F. Filloux 11993) Localization ofl l2..'I Iw-conotoxin GVIA binding in human hippocampus and cerebellum. Neuroreport 4: 1331-1334.
Altman. J. t 1982) Morphological development of the rat cerebellum and some of its mechanisms. In S.L. Palay and V. Chan-Palay (eds): The Cerebellum. New Vistas. Berlin: Springer-Verlag, pp. 8 4 9 .
Barhanin, ,J.. A. Schmid, and M. Lazdunski (1988) Properties of structure and interaction of the receptor for w-conotoxin, a polypeptide active on Ca" channels. Biochem. Biophys. Res. Commun. 150:1051-1062.
Bastian, J. i 19811 Electrolocation. 11. The effects of movingobjects andother electrical stimuli on the activities of two categories of posterior lateral line lobe cells in Apteronotus alhrfrons. J . Comp. Physiol. ( A ) 144.481- 494.
Bastian, J . 11986) Gain control in the electrosensory system mediated by descending inputs to the electrosensory lateral line lobe. J. Neurosci. 6:553-562.
Birnbaumer, I,., K.P. Campbell, W.A. Catterall, M.M. Harpold, F. Hofmann, W.A. Home. Y. Mori, A. Schwartz, T.P. Snutch, T. Tanabe, and R.W. Tsien i 19941 The naming of voltage-gated calcium channels. Neuron 13:50.5--506.
Boland, L.M., Morrill, J .A., and B.P. Bean (19941 w-conotoxin block of N-type calcium channels in frog and rat sympathetic neurons. J. Neurosci. 14,501 1-5027.
Carr, C.E.. and L. Maler (1986) Electroreception in gymnotiform fish. In T.H. Bullock and W. Heiligenberg (eds): Electroreception. New York: John Wiley & Sons, pp. 319-373.
Cohen, M.W., O.T. Jones, and K.J. Angelides 11991) Distribution of Ca2+ channels on frog motor nerve terminals revealed by fluorescent o-cono- toxin. J. Neurosci. I1:1032-1039.
DiversL;-Picduissi, M., and K. Dunlap (1993) Distinct, convergent second messenger pathways modulate neuronal calcium currents. Neuron 10:753-760.
Dunlap, K.. Luebke, J.I., and Turner, T.J. (1995) Exocytotic Ca2+ channels in mammalian central neurons. TINS 18:89-98.
Fariiias, I., G. Egea, J. Blasi, C. Cases, and J. Marsal(1993) Calcium channel antagonist omega-conotoxin binds to intramembrane particles of iso- lated nerve terminals. Neuroscience 54:745-752.
Filloux, F.. A. Schapper, S.R. Naisbitt, B.M. Olivera, and J.M McIntosh I 1994 I Complex patterns of 1 '"'Ilornega-conotoxin GVIA binding site expression during postnatal rat brain development. Dev. Brain Res. 78: 131-136.
Finger, T.E. i 1978) Efferent neurons of the teleost cerebellum. Brain Kes. I 53:60X-6 15.
Finger. T.E. 11983) Organization of the teleost cerebellum. In R.G. North- cutt and R.E. Davis (eds): Fish Neurobiology Ann Arbor: University of Michigan. pp. 261-284.
Fortier, L.-P.. J.-P. Tremblay, J . Rafrafi, and R. Hawkes (19911 A monoclo- nal antibody to conotoxin reveals the distribution of a subset of calcium channels in the rat cerebellar cortex. Mol. Brain Res. 9:209-215.
Fox, A.P., M.C. Nowycky, and R.W. Tsien (1987) Kinetic and pharmacologic properties distinguishing three types of calcium currents in chick sensory neurons. J. Physiol. (Lond.) 94.149-172.
Hawkes, R.. and C. Mascher I 1994) The development of molecular compart- mentation in the cerebellar cortex. Acta. Anat. 151r139-149.
Haws, C.M.. P.A. Slesinger, and J.B. Lansman (1993) Dihydropyridine-and u,-conotoxin-sensitive CaJ- currents in cerebellar neurons: persistent block of L-type channels by a pertussis toxin-sensitive G-protein. J. Neurosci. 13:1148-1156.
Haydon. P.G.. E. Henderson, and E.F. Stanley (1994) Localization of individual calcium channels at the release face of a presynaptic nerve terminal. Neuron 1,71275-1280.
Horne, W.A.. P.T. Ellinor, I . Inman, M. Zhou, R.W. Tsien,andT.L. Schwartz (19931 Molecular diversity of Ca" channel u l subunits from the marine ray D~scopyge ommata. Proc. Natl. Acad. Sci. U.S.A. 90:3787-3791
Ikeda, S.R. (19911 Double-pulse calcium channel facilitation in adult rat sympathetic neurons. J. Physiol. (Land.) 43.9181-214.
Jones. 0.T.. D.1,. Kunze, and K.J. Angelides I19891 Localization and mobility
of w-conotoxin-sensitive Ca" channels in hippocampal CAI neurons. Science 244:1189-1193.
Knox, R.J., E.A. Quattrocki, J .A. Connor, and L.K. Kaczmarek (1992) Recruitment of Cai' channels by protein kinase C during rapid forma- tion of putative neuropeptide release sites in isolated Aplysia neurons. Neuron 8:883-889.
Komuro, H., and P. Rakic 11992) Selective role of N-type calcium channels in neuronal migration. Science 257:806-809.
Koyano, K., T. Abe, Y. Nishiuchi, and S. Sakakibara (1987) Effects of synthetic w-conotoxin on synaptic transmission. Eur. J. Pharmacol. 135:337-343.
Lipscombe, D., D.V. Madison, M. Poenie, H. Keuter, R.Y. Tsien, and R.W. Tsien (1988) Spatial distribution of calcium channels and cytosolic calcium transients in LTowth cones and cell bodies of sympathetic neurons. Proc. Natl. Acad. Sci. U.S.A. 85:2398-2402.
Liu, J., R. Bangalore, A. Rutledge, and D.J. Triggle (1994) Modulation of L-type Ca" channels in clonal rat pituitary cells by membrane depolar- ization. Mol. Pharmacol. 45:1198-1206.
Llinas, R., M. Sugimori, D.E. Hillman, and B. Cherksey (19921 Distribution and functional significance of the P-type voltage-dependent CaL- chan- nels in the mammalian central nervous system. TINS 15:351-355.
Maler, L. (1979) The posterior lateral line lobe of certain gymnotoid fish: Quantitative light microscopy. J. Comp. Neurol. 18.3:323-363.
Maler, L., and D. Monaghan (1991 1 The distribution of excitatory amino acid binding sites in the brain of an electric fish, Aptrronotus lrptorhynchus. J. Chem. Neuroanat. 4.39-61.
Maler, L., and E. Mugnaini (1994) Correlating GABAergc circuits and sensory function in the Electrosensory Lateral Line Lobe (ELL) of a Gymnotiform fish. J. Comp. Neurol. 345,2244252.
Maler, L., E. Sas, S.A. Johnston, and W.G. Ellis (1991). An atlas of the brain oftheelectric fish Apteronotus leptorhynchus. J. Chem. Neuroanat. 4:l-38.
Maler, L., E. Sas, and J. Rogers 11981) The cytology of the posterior lateral line lobe of high frequency weakly electric fish (Gymnotidae): Dendritic differentiation and synaptic specificity in a simple cortex. J. Comp. Neurol. 195.87-139.
Mathieson, W.B., W. Heiligenberg, and L. Maler (1987) Ultrastructural studies of physiologically identified electrosensory afferent synapses in the Gymnotiform fish, Eigenrnannra. J. Comp. Neurol. 255,526-537.
Mathieson, W.B., and L. Maler ( 19881 Morphological and electrophysiolog- cal properties of a novel in uitro preparation: the electrosensory lateral line lobe brain slice. J . Comp. Physiol. A 16.7:489-506.
Metzner, W., and W. Hciligenberg 11991) The coding of signals in the electric communication of the gymnotiform fish Eigenrnannia: From electrore- ceptors to neurons in the torus semicircularis of the midbrain. J . Comp. Physiol. 169:135-150.
Mills, L.R., C.E. Niesen, A.P. So. P.L. Carlen, I. Spigelman, and O.T. Jones (19941 N-Type Ca2+ channels are located on somata, dendrites, and a subpopulation of dendritic spines on live hippocampal pyramidal neu- rons. J. Neurosci. 14:6815-6824.
Mintz, I.M., and B.P. Bean (1993) GABAll receptor inhibition of P-type Ca2+ channels in central neurons. Neuron 10:889-898.
Olivera, B.M., G.P. Miljanich, J . Ramachandran, and M.E. Adams (1994) Calcium channel diversity and neurotransmitter release: The w-conotox- ins and w-agatoxins. Annu. Rev. Biochem. 63823-867.
Passafaro, M., F. Clementi, and I.:. Sher (1992) Metabolism of w-conotoxin- sensitive voltage-operated calcium channels in human neuroblastoma cells: Modulation by cell differentiation and anti-channel antibodies. J . Neurosci. 12:3372-3379.
Passafaro, M., F. Clementi, A. Pollo, E. Carbone, and E. Sher (1994) w-conotoxin and CdY' stimulate the recruitment to the plasmamem- brane of an intracellular pool ofvoltage-operated Ca" channels. Neuron 12:317-326.
Plummer, M.R., D.E. Logothetis, and P . Hess (19891 Elementary properties and pharmacological sensitivities of calcium channels in mammalian peripheral neurons. Neuron 2: 1453-1463.
Regan, L.J., D.W.Y. Sah, and B.P. Bean (1991) Ca" channels in rat central and peripheral neurons: high-threshold current resistant to dihydropyri- dine blockers and w-conotoxin. Neuron 6269-280.
Robitaille, R., E.M. Adler, and M.P. Charlton (1990) Strategc location of calcium channels at transmitter release sites of frog neuromuscular synapses. Neuron ,51773-779.
Sas, E. , and L. Maler (1987) The organization of afferent input to the caudal lobe of the cerebellum of the gymnotid fish Aptrronotus lrptorhynchus. Anat. Embryol. 177.65-79.

Y. THARANI ET AL.
Sas. E., L. Maler, and B. Tinner (1990) Catecholaminergic systems in the brain of a gymnotiform teleost fish: an immunohistochemical study. J. Comp. Neurol. 292.127-162.
Schwartz, K.J., and B.P. Bean (1992) Inhibition of calcium channels in rat CA3 pyramidal neurons by a metabotropic glutamate receptor. J. Neurosci. 12:4358-4371.
Shumway, C.A. (1989) Multiple electrosensory maps in the medulla of weakly electric gymnotiform fish. I. Physiological differences. J . Neuro- sci. 9:4388-4399.
Shumway, C.A., and L. Maler (1989) GABAergic inhibition shapes temporal and spatial response properties of pyramidal cells in the electrosensory lateral line lobe of gymnotiform fish. J. Comp. Neurol. 164:391-407.
Stanley. E.F., and G. Goping (1991) Characterization o fa calcium current in a vertebrate cholinergic presynaptic nerve terminal. J. Neurosci. 11:985- 993.
Takemura, M., H. Fukui, and H. Wada (1987) Different localization of receptors for w-conotoxin and nitrendipine in rat brain. Biochem. Biophys. Res. Commun. I49:982-988.
Takemura, M., H. Kiyama, H. Fukui, M. Tohyama, and H. Wada (1989) Distribution of the w-conotoxin receptor in rat brain. An autoradio- graphic mapping. Neuroscience 32:405-4 16.
Turner, R.W., and L. Borg (1995) Acutely isolated and cultured cells from the electrosensory lateral line lobe of a gymnotiform teleost. J . Comp. Neurol. 358:305-323.
Turner. R.W., and R.B. Hawkes ( 19921 The immunocytochemical distribu- tion of w-conotoxin-binding sites in the electrosensory Lateral line lobe IELL) ofilpteronotus leptorhynchus. XXII Proc. Sac. Neurosci. 18:970.
Turner, R.W., and L.L. Moroz (1995) Localization of nicotinamide adenine dinucleotide phosphate-diaphorase in electrosensory and electromotor systems of a gymnotiform teleost, Apteronotus leptorhynchus. J . Comp. Neurol. ,356r261-274.
Turner, R.W.. J . Plant, and L. Maler (1991) Conditional oscillatory discharge in topographic maps of the electrosensory lateral line lobe. XXI Proc. Soc. Neurosci. 171405.
Turner, R.W., L. Maler, T. Deerinck, R.S. Levinson, and M.H. Ellisman (1994) TTX-sensitive dendritic sodium channels underlie oscillatory discharge in a vertebrate sensory neuron. J. Neurosci. 1416453-6471.
Vandaele, S.F., and T.A. Reader (1994) Ca('')-sensitive and insensitive omega-conotoxin GVIA binding sites in rat brain. Neuroreport 5:1121- 1124.
Wagner, J.A., A.M. Snowman, A. Biswas, B.M. Olivera, and S.H. Snyder (1988) w-conotoxin GVIA binding to a high-affinity receptor in brain: Characterization, calcium sensitivity, and solubilization. J . Neurosci. 8:3354-3359.
Westenbroek, R.E., J.W. Hell, C. Warner. S.J. Dubel, T.P. Snutch, and W.A. Catterall ( 1992) Biochemical properties and subcellular distribution of an N-type calcium channel a1 subunit. Neuron 9:1099-1115.
Wheeler, D.B., A. Randall, and R.W. Tsien (1994) Holes of N-type and Q-type Ca2+ channels in supporting hippocampal synaptic transmission. Science 264:107-111.
Witcher, D.R., M. DeWaard, and K.P. Campbell (1993) Characterization of the purified N-type Ca'. channel and the cation sensitivity of w-cono- toxin GVIA binding. Neuropharmacology 32,1127-1139,
Yeager, R.E., D. Yoshikami, J . Rivier, L.J. Cruz, and G.P Miljanich (1987) Transmitter release from presynaptic terminals of electric organ: Inhibi- tion by the calcium channel antagonist omega Conus toxin. J . Neurosci. 7.2390-2396.
Zhu, Y., and S.R. Ikeda (1993) Adenosine modulates voltage-gated Ca2* channels in adult rat sympathetic neurons. J. Neurophysiol. 70:610-620.
Zupanc, G.K.H., and I. Horschke (1995) Proliferation zones in the brain of adult gymnotiform fish: a quantitative mapping study. J . Comp. Neurol. 3 5 3 2 13-233.
Zupanc, G.K.H., I. Horschke, R. Ott, and G.B. Rascher (1995) The postem- bryonic development of the cerebellum in a teleost fish, Apteronotus leptorhynchus. In N. Elsner and R. Menzel (edsi: Gottingen Neurobiol- ogy Report 1995. Volume 11. StuttgartiNew York: Georg Thieme Verlag, p. 549.