distinct genetic influences on locomotor senescence in drosophila revealed by a series of metrical...
TRANSCRIPT

www.elsevier.com/locate/expgero
Experimental Gerontology 41 (2006) 877–881
Short Report
Distinct genetic influences on locomotor senescence inDrosophila revealed by a series of metrical analyses
Ian Martin, Michael S. Grotewiel *
Department of Human Genetics and Neuroscience Program, Virginia Commonwealth University School of Medicine, Richmond, VA 23298, USA
Received 9 May 2006; received in revised form 20 June 2006; accepted 27 June 2006Available online 7 August 2006
Abstract
Decline of locomotor function is a prominent feature of aging in most species. We previously described a series of metrical analysesfor quantifying functional declines associated with aging. Here, we have applied these metrics to elucidate differences in the nature oflocomotor senescence between Drosophila cohorts from four different genetic backgrounds (Canton-S, Oregon-R, Samarkand and Lau-sanne-S) and two long-lived mutant strains, Indy206 and chico1. These analyses demonstrate that senescence of locomotor function inOregon-R and Canton-S proceeds more rapidly than in Samarkand and Lausanne-S flies. Our analyses also show that while chico1
and Indy mutants both exhibit attenuated locomotor declines, the impact of chico loss of function on locomotor senescence is substan-tially greater. Our studies illustrate how important differences in functional senescence characteristics can be unveiled by in-depth anal-ysis on functional senescence data sets.� 2006 Elsevier Inc. All rights reserved.
Keywords: Aging; Behavior; Negative geotaxis; Method; Metrics
1. Introduction
Senescence of physiological function (functional senes-cence) is believed to drive the decrease in quality of lifeand the increase in mortality associated with aging(Grotewiel et al., 2005). Accordingly, understanding func-tional senescence is crucial to understanding the biologyof aging. To facilitate the investigation of functional senes-cence, we previously designed a series of metrical analysesfor quantifying and comparing age-associated functionalimpairments between cohorts (Martin et al., 2005). Thesemetrics describe the rate of functional decline, the age atwhich function declines to 75%, 50% and 25% of initial per-formance and total function across the assessment period.In the current study, we used these metrics to rigorouslyevaluate senescence of negative geotaxis, a locomotorbehavior in Drosophila. Age-related decline in this behavior
0531-5565/$ - see front matter � 2006 Elsevier Inc. All rights reserved.
doi:10.1016/j.exger.2006.06.052
* Corresponding author. Tel.: +1 804 828 9632x160; fax: +1 804 8283760.
E-mail address: [email protected] (M.S. Grotewiel).
is sensitive to genetic background and alleles of chico andIndy that extend life span (Gargano et al., 2005). Here,using a series of detailed metrical analyses, we show (1) thatnegative geotaxis declines more rapidly in Canton-S andOregon-R than in Samarkand and Lausanne-S flies and(2) that mutation of chico has significantly more pro-nounced beneficial effects on locomotor senescence thandoes mutation of Indy.
2. Materials and methods
2.1. Fly stocks, husbandry and aging
Canton-S, Oregon-R, Samarkand and Lausanne-Sstrains flies were used as representative genetic back-grounds. The Canton-S strain was provided by Ron Davis(Baylor College of Medicine, Houston, TX, USA). TheOregon-R, Samarkand and Lausanne-S strains wereobtained from the Bloomington Drosophila Stock Centerat Indiana University (Bloomington, IN, USA). TheIndy206 and chico1 mutants were provided by Stephen

878 I. Martin, M.S. Grotewiel / Experimental Gerontology 41 (2006) 877–881
Helfand and Marc Tatar (Brown University, Providence,RI, USA). Flies were housed, collected and tested as previ-ously described (Gargano et al., 2005).
2.2. Negative geotaxis assays
Rapid iterative negative geotaxis (RING) assays wereperformed as previously described (Gargano et al., 2005).Ten vials initially containing 25 adults each were assessedfor each genotype and genetic background in longitudinalstudies at weekly intervals. The performance of each vialwas treated as a single datum at each age.
2.3. Metrical analyses and statistical tests
Metrical analyses described previously (Martin et al.,2005) were used on previously published senescence of neg-ative geotaxis data sets (Gargano et al., 2005). The absoluterate of decline (aROD) was calculated as the change in neg-ative geotaxis scores between each assessment interval. Theproportional rate of decline (pROD) was calculated as theproportion of a function lost during each interval. Declinetime75 (DT75), DT50 and DT25, representing the timerequired for function to decline to 75%, 50% and 25% ofinitial values, respectively, were interpolated or extrapolat-
Fig. 1. Negative geotaxis senescence in different genetic backgrounds. (A) NeSamarkand (Sam) males. There was a significant effect of age and genetic backgand significant interactions between age and genetic background (p < 0.0001). Dexperiments with 10 vials of up to 25 flies each. (B) There was a significant effecage and genetic background on aROD (two-way ANOVA, p < 0.0001 for all fac75%, 50% and 25% of initial negative geotaxis (individual ANOVAs, p < 0.0Lausanne-S were significantly different than all other backgrounds (p < 0.001Oregon-R and Canton-S (p < 0.001). (D) There was a significant effect ofBonferroni’s post hoc tests revealed that total negative geotaxis was different
ed from second-order curve fits. Total negative geotaxisduring each experiment was determined as the area underthe curve. Metrics were calculated from each vial individu-ally. Data are reported as mean ± SEM for the 10 vialsconstituting each genotype or genetic background. Sincethe effects of genetic background on senescence of negativegeotaxis were similar in males and females and the effects ofthe Indy206 and Indy302 alleles on senescence of negativegeotaxis were similar, data from only males in the geneticbackgrounds and only the Indy206 allele are reported herefor brevity. Data from females for chico1 mutants and con-trols were analyzed since loss of function in chico has morepronounced effects on aging in this sex (Tu et al., 2002).Parametric tests in Prism (GraphPad Software, San Diego,CA, USA) were used to assess statistical significance(p < 0.05).
3. Results
3.1. Effect of genetic background on senescence of locomotor
function
Age-dependent loss of negative geotaxis varies withgenetic background (Fig. 1A, (Gargano et al., 2005)).Toward a better understanding of how genetic background
gative geotaxis in Canton-S (CS), Oregon-R (OR) Lausanne-S (LS) andround on negative geotaxis (two-way ANOVA, p < 0.0001 for both factors)ata (mean ± SEM) are taken from Gargano et al. (2005) and are from twot of age and genetic background as well as significant interactions betweentors). (C) There was a significant effect of genetic background on decline to
001). Bonferroni’s post hoc tests revealed that DT75, DT50 and DT25 for) and that DT25 for Samarkand was significantly different from that ofgenetic background on total negative geotaxis (ANOVA, P < 0.0001).
in all genetic backgrounds (p < 0.001).

I. Martin, M.S. Grotewiel / Experimental Gerontology 41 (2006) 877–881 879
can impact senescence of negative geotaxis, we performed aseries of metrical analyses as previously described (Martinet al., 2005) on these data. To determine whether the paceof senescence in negative geotaxis was influenced by geneticbackground, we calculated the absolute rate of decline(aROD) and proportional rate of decline (pROD) (Martinet al., 2005) from the data in Fig. 1A. Genetic backgroundand age had significant effects on the aROD of negativegeotaxis (Fig. 1B). aROD was higher overall in Canton-Sand Oregon-R than in Samarkand and Lausanne-S.Statistical interactions between age and genetic back-ground indicate that the effects of age on aROD were notuniform across the different genetic backgrounds tested.Interestingly, Canton-S, Oregon-R and Lausanne-S allexhibited a significant increase in aROD with age whereasSamarkand displayed an age-associated decrease in thismeasure (Fig. 1B). The age-related changes in aROD inLausanne-S and Samarkand, however, were quite small.Similarly, the pROD of negative geotaxis was affected byage and genetic background, and was higher overall in Can-ton-S and Oregon-R than in Samarkand and Lausanne-S(data not shown). These analyses establish that the rateof negative geotaxis senescence was influenced by geneticbackground.
To quantify the locomotor declines in Fig. 1A further,we derived the DT75, DT50 and DT25, which representthe time required for function to decline to 75%, 50%and 25% of initial values, respectively (Martin et al.,2005). Genetic background had significant effects on thesemeasures (Fig. 1C). Specifically, decline times in Lausanne-S were consistently longer than in the other three back-grounds. Interestingly, DT25 in Samarkand was longerthan in Canton-S and Oregon-R, although DT75 andDT50 were indistinguishable in these three genetic back-grounds. This suggests that Samarkand males exhibited apattern of decline in negative geotaxis distinct from Can-ton-S and Oregon-R. These data demonstrate that the timeat which negative geotaxis declined by quartile percentamounts was sensitive to genetic background.
We calculated area under the curve to assess the effect ofgenetic background on total negative geotaxis throughoutour experiments. Total negative geotaxis was different inall four genetic backgrounds (Fig. 1D). Lausanne-S hadthe highest total negative geotaxis followed in descendingorder by Samarkand, Oregon-R and Canton-S. These datareveal that total negative geotaxis can vary between differ-ent genetic backgrounds by more than twofold. This metricillustrates a marked difference in the total functionality ofthese four genetic backgrounds.
3.2. Effects of chico and Indy mutations on senescence of
locomotor function
Mutations in chico (Clancy et al., 2001; Tu et al., 2002)and Indy (Rogina et al., 2000) extend life span in Drosoph-
ila. We previously reported that these mutations also altersenescence of negative geotaxis (Figs. 2A and B, (Gargano
et al., 2005)). To quantify and compare age-related loss ofnegative geotaxis behavior in chico1 homozygous andIndy206 heterozygous mutants, we assessed rate of decline,decline times and total negative geotaxis from the data inFigs. 2A and B.
There was a significant effect of age on aROD for bothchico1 and Indy206 mutants and their respective controls(Figs. 2C and D). Interestingly, all strains tested in thesestudies exhibited a trend towards an increase in aROD inthe second half of the assessment period. Overall, chico1
mutants had lower aROD values than control flies(Fig. 2C). In contrast, overall aROD values in Indy206
mutants were not statistically distinguishable from controls(Fig. 2D). chico1 mutants appeared to have reducedaRODs at most assessments whereas Indy206 flies did not.This suggests that while the beneficial effects of chico1 onfunctional senescence manifested throughout the experi-ment, for Indy206 mutants these effects were restricted tothe first few weeks of life. The pROD in negative geotaxisin chico1 and Indy206 mutants followed a pattern similar tothat of aROD; there were significant effects of age andgenotype on pROD in chico1 flies, but in Indy206 mutantsthere was a significant effect of age only (data not shown).
All three DTs for chico1 mutants were significantlyincreased (Fig. 2E). Hence, a lower rate of functionaldecline resulted in a delay in the decline time to 75%,50% and 25% of initial function. This further supportsthe idea that chico1 delayed senescence of negative geotaxisthroughout the function span. DTs for the Indy206 alleleindicate a significant delay in the DT75 and DT50
(Fig. 2F). No difference was found, however, for theDT25. This is consistent with the positive effects of Indy
mutations on senescence of negative geotaxis occurringmainly during the first few weeks of life.
Total negative geotaxis during the experiments inchico1 mutants was increased by 62% relative to controls(Fig. 2G). Although chico1 did not bestow any increasein peak locomotor function in young flies (Fig. 2A), theattenuated pattern of senescence in chico1 animals con-ferred a substantial increase in their total locomotor func-tionality. Indy mutants also exhibited an increase in totalnegative geotaxis (Fig. 2H), although the effect was not aslarge as in chico1 flies. The larger increase in total negativegeotaxis in chico1 mutants is consistent with the idea thatchico1 loss of function had a greater impact on aging ofnegative geotaxis throughout the life span than did muta-tions in Indy.
4. Discussion
Although most multicellular species experience someform of functional senescence during the post-reproductivephase of life, the nature of this senescence varies betweendifferent species and between individuals of the same spe-cies (Arking, 1998). Variability among individuals of thesame species depends both on external factors related tothe environment and internal factors including genetic
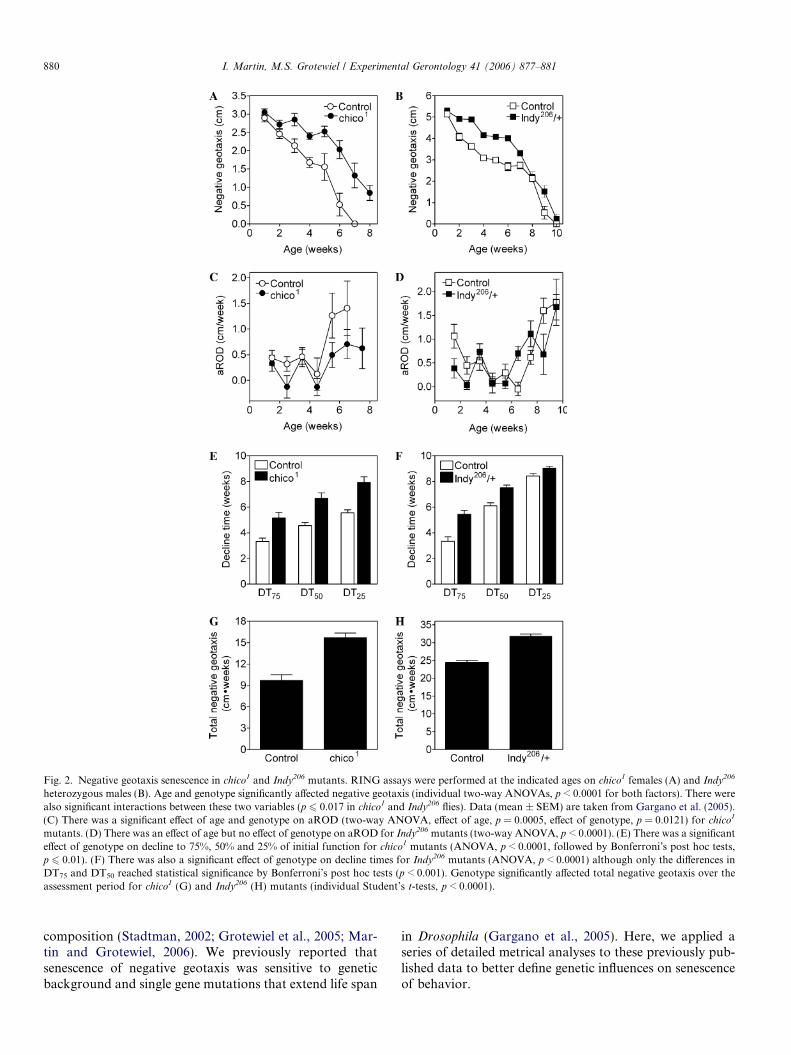
Fig. 2. Negative geotaxis senescence in chico1 and Indy206 mutants. RING assays were performed at the indicated ages on chico1 females (A) and Indy206
heterozygous males (B). Age and genotype significantly affected negative geotaxis (individual two-way ANOVAs, p < 0.0001 for both factors). There werealso significant interactions between these two variables (p 6 0.017 in chico1 and Indy206 flies). Data (mean ± SEM) are taken from Gargano et al. (2005).(C) There was a significant effect of age and genotype on aROD (two-way ANOVA, effect of age, p = 0.0005, effect of genotype, p = 0.0121) for chico1
mutants. (D) There was an effect of age but no effect of genotype on aROD for Indy206 mutants (two-way ANOVA, p < 0.0001). (E) There was a significanteffect of genotype on decline to 75%, 50% and 25% of initial function for chico1 mutants (ANOVA, p < 0.0001, followed by Bonferroni’s post hoc tests,p 6 0.01). (F) There was also a significant effect of genotype on decline times for Indy206 mutants (ANOVA, p < 0.0001) although only the differences inDT75 and DT50 reached statistical significance by Bonferroni’s post hoc tests (p < 0.001). Genotype significantly affected total negative geotaxis over theassessment period for chico1 (G) and Indy206 (H) mutants (individual Student’s t-tests, p < 0.0001).
880 I. Martin, M.S. Grotewiel / Experimental Gerontology 41 (2006) 877–881
composition (Stadtman, 2002; Grotewiel et al., 2005; Mar-tin and Grotewiel, 2006). We previously reported thatsenescence of negative geotaxis was sensitive to geneticbackground and single gene mutations that extend life span
in Drosophila (Gargano et al., 2005). Here, we applied aseries of detailed metrical analyses to these previously pub-lished data to better define genetic influences on senescenceof behavior.

I. Martin, M.S. Grotewiel / Experimental Gerontology 41 (2006) 877–881 881
Our studies demonstrate that many aspects of senes-cence of negative geotaxis are dependent on genetic back-ground. The rates of decline were different in the fourgenetic backgrounds as were the age-dependent changesin this metric. Both aROD and pROD increased with agein Canton-S, Oregon-R and Lausanne-S during the first 8weeks of life. In contrast, Samarkand showed a decreasein aROD with age and no significant change in pRODfor the same period. This indicates that altered senescenceof negative geotaxis can result from distinct changes in theage-dependent dynamics of aROD and pROD. All three ofthe DT metrics were longer in Lausanne-S than the othergenetic backgrounds, reflecting the lower rate of locomotordecline in Lausanne-S. DT25 for Samarkand was longerthan that of both Canton-S and Oregon-R due to theincrease in locomotor decline with age in these latter twobackgrounds. Finally, total negative geotaxis was differentin all four genetic backgrounds and was lowest in Canton-Sand Oregon-R, the two strains that exhibited the fastestpace of locomotor decline. Together, these data demon-strate that senescence of negative geotaxis in Canton-Sand Oregon-R flies proceeds more rapidly than it does inLausanne-S and Samarkand flies. These studies illustratethat senescence of negative geotaxis proceeds via patternsof decline that are characteristic of each genetic back-ground, highlighting the need to control for genetic back-ground in behavioral aging studies.
Both chico1 and Indy206 mutants have altered senescenceof locomotor function (Gargano et al., 2005). Detailedanalysis of these age-related declines using our series ofmetrics revealed subtle, but important, differences in theway functional senescence is modified in these twomutants. The rates of decline for chico1 mutants werereduced throughout most of the assessment period. Thisresulted in a significant increase in all decline time measuresin addition to a large increase in total negative geotaxisfunction in chico1 flies. For Indy206 mutants, reductions inthe rate of decline were rather marginal overall and restrict-ed to the early stages of senescence. Interestingly, earlyreductions in decline rates appeared to be followed byincreases at greater ages. Nevertheless a small initial decel-eration in the rate of decline in Indy206 flies manifested asextensions in DT75 and DT50 in addition to increased totalnegative geotaxis activity. Since mutation of chico hadgreater effects than mutation of Indy on the rates of func-tional decline, the time required for function to declineand total function, our analyses indicate that the chico1
mutation exerts a larger benefit on the biological systemsthat support locomotor function throughout the Drosoph-
ila life span.The aROD, decline time and total function descriptors
we applied here to analyze senescence of negative geotaxis
should be useful for characterizing age-related decline inmany other functions across age (Martin et al., 2005).When function is highest in the youngest groups anddeclines with age (as we typically find for negative geotax-is), the use of the descriptors is fairly straightforward.Although, we have not extensively investigated the use ofthese descriptors on data sets in which peak function ini-tially increases with age and then declines (e.g., femalefecundity in various species (Novoseltsev et al., 2003)),we predict that they will provide important insights evenin these somewhat more complicated scenarios when usedin an integrated fashion. It seems likely, however, thatadditional descriptors will be required to provide a compre-hensive analysis of more complex data sets. For example, ameasure of time to peak function might be informative insuch cases.
Acknowledgements
The authors thank Ron Davis, Marc Tatar and StephenHelfand for providing fly stocks. Work in the author’s lab-oratory is supported by grants from the National Institutesof Health and the American Federation for AgingResearch.
References
Arking, R., 1998. The Biology of Aging: Observations and Principles.Sinauer Associates, Sunderland, MA.
Clancy, D.J., Gems, D., Harshman, L.G., Oldham, S., Stocker, H., Hafen,E., Leevers, S.J., Partridge, L., 2001. Extension of life-span by loss ofCHICO, a Drosophila insulin receptor substrate protein. Science 292,104–106.
Gargano, J.W., Martin, I., Bhandari, P., Grotewiel, M.S., 2005. Rapiditerative negative geotaxis (RING): a new method for assessingage-related locomotor decline in Drosophila. Exp. Gerontol. 40,386–395.
Grotewiel, M.S., Martin, I., Bhandari, P., Cook-Wiens, E., 2005.Functional senescence in Drosophila melanogaster. Ageing Res. Rev.4, 372–397.
Martin, I., Gargano, J.W., Grotewiel, M.S., 2005. A proposed set ofdescriptors for functional senescence data. Aging Cell 4, 161–164.
Martin, I., Grotewiel, M.S., 2006. Oxidative damage and age-relatedfunctional declines. Mech. Ageing Dev. 127, 411–423.
Novoseltsev, V.N., Novoseltseva, J.A., Boyko, S.I., Yashin, A.I., 2003.What fecundity patterns indicate about aging and longevity: insightsfrom Drosophila studies. J. Gerontol. A. Biol. Sci. Med. Sci. 58, 484–494.
Rogina, B., Reenan, R.A., Nilsen, S.P., Helfand, S.L., 2000. Extended life-span conferred by cotransporter gene mutations in Drosophila. Science290, 2137–2140.
Stadtman, E.R., 2002. Importance of individuality in oxidative stress andaging. Free Radic. Biol. Med. 33, 597–604.
Tu, M.P., Epstein, D., Tatar, M., 2002. The demography of slow aging inmale and female Drosophila mutant for the insulin-receptor substratehomologue chico. Aging Cell 1, 75–80.