diplomamunka klórozott szénhidrogén szennyez k...
TRANSCRIPT

Diplomamunka
Klórozott szénhidrogén szennyezők fitoplankton közösségekre gyakorolt hatásának vizsgálata
mikrokozmosz-kísérletekben.
Készítette:
Balogh Julianna Hidrobiológus (MSc) egyetemi hallgató
Témavezető: Dr. Bácsi István egyetemi adjunktus
Debreceni Egyetem
Természettudományi és Technológiai Kar Hidrobiológia Tanszék
Debrecen 2013

UNIVERSITY OF DEBRECEN o FACULTY OF SCIENCE AND TECHNOLOGY 1949
DEBRECENI EGYETEM
TERMÉSZETTUDOMÁNYIÉSTECHNOLÓGIAIKAR
NYILATKOZAT(A DE TTK hallgatóinak diplomadolgozatához/szakdolgozatához)
Alulírott Balogh Julianna (Neptun kód: F4QYGT) jelen nyilatkozat aláírásával kijelentem,hogya
Klórozott szénhidrogén szennyezők fitoplankton közösségekre gyakorolt hatásánakvizsgálata mikrokozmosz-kísérletekben.
című diplomadolgozat - a továbbiakban: dolgozat - önálló munkám, a dolgozat készítésesorán betartottam a szerzői jogról szóló 1999. évi LXXVI tv. szabályait, valamint az Egyetemáltal előírt, a dolgozat készítésére vonatkozó szabályokat, különösen a hivatkozások ésidézések tekintetében 1.
Kijelentem továbbá, hogy a dolgozat készítése során az önálló munka tekintetében atémavezetőt nem tévesztettem meg. Meg nem engedett segítséget nem vettem igénybe, adolgozatot más oktatási intézményben szakdolgozatként vagy diplomamunkaként korábbannem nyújtottam be.Jelen dolgozat aláírásával tudomásul veszem, hogy amennyiben bizonyítható, hogy a'dolgozatot nem magam készítettem, vagy a dolgozattal kapcsolatban szerzői jogsértés tényemerül fel, illetve a dolgozatot korábban más oktatási intézményben is benyújtottam, úgy aDebreceni Egyetem megtagadja a dolgozat befogadását és ellenem fegyelmi eljárást indíthat.A dolgozat befogadásának megtagadása és a fegyelmi eljárás indítása nem érinti a szerzőijogsértés miatti egyéb (polgári jog, szabálysértési jog, büntetőjogi) jogkövetkezményeket.
Debrecen, 2013. év május hó 22. nap
...~~\.~<~,,~ .Hallgató aláírása
Alulírott témavezető kijelentem, hogy az ezúton benyújtott szakdolgozat / diplomamunka azén szakmai irányításommal, a jelölttel folytatott rendszeres szakmai konzultáció mellettkészült, a hallgató önálló munkáját tükrözi. A dolgozatot áttekintettem, a záró- vizsgántörténő védésre bocsátásra megfelelőnek és a könyvtár ~?~kl.pj,~~afeltölthetőnek találtam.
ti..;:>Y; i é., , l'J;-,'9 0-'1-, ""'.
Debrecen, 2013. év május hó 22. nap ig1 -1 '..: ~etb,· 1.f 6 h'~;~ ~",. ',.0, ~ ,', .:·l~ ;~ ••••••• ':"".. JI. .L:9- .~\"0,'~\'J --;.0».,,( témavezető aláírása)ij~,~ <$~c:"Y/
°6i02<),",,:\ '\:~/
-"DE TTK, Hidrobiológiai Tanszék
I 1999. évi LXXVL tv. 34. § (1) A.mü részletet .....az átvevőmű által indokolt és azeredetihez híven a forrás, valamini az ott szerzö bárki idézheti.36. § (1) Nyilvánosan tartott előadások és más hasonló müvek . .' valamint .. . . beszédek
tájékoztatás céljára- a cél által indokolt terjedelemben szabadon felhasználhatók. Ilyen felhasználás eselén il
forrást a szerzö nevével együtt .....fel kell tüntetni, hacsak ez lehetetlennek nem bizonvul,
H4032 Debrecen,Egyetemtér L H4010 Debrecen,Pf: 18.Telefon: (+36)(52)316-012, Fax: (52) 533-677, E-mail: ttkdekanéüpuma.unideb.hu

Tartalomjegyzék 1. Bevezetés és célkitűzés 3
2. Irodalmi áttekintés 5
2.1. Kismolekulájú klórozott szénhidrogének 5
2.1.1. Tetraklóretán 5
2.1.1.1. A tetraklóretán sorsa a természetben 5
2.1.1.2. A tetraklóretán metabolizmusa és a toxicitás mechanizmusa 6
2.1.2. Tetraklóretilén 7
2.1.2.1. A tetraklóretilén sorsa a természetben 7
2.1.2.2. A tetraklóretilén metabolizmusa és a toxicitás mechanizmusa 8
2.1.3. Triklóretilén 9
2.1.3.1. A triklóretilén sorsa a természetben 10
2.1.3.2. A triklóretilén metabolizmusa és a toxicitás mechanizmusa 10
2.2. A klórozott szénhidrogének hatása a fitoplanktonra 11
2.3. Mikrokozmosz rendszerek 11
2.4. A biológiai sokféleség hatása az ökoszisztéma szennyezőkkel szembeni
érzékenységére 13
3. Anyag és módszer 15
3.1. A mikrokozmosz rendszerek összeállítása 15
3.2. A fitoplankton növekedésének nyomon követése 16
3.3. Az eredmények statisztikai értékelése 16
4. Eredmények és értékelésük 17
4.1. A háttérváltozókban megfigyelt változások az egyes vizsgálati időszakokban 17
4.1.1. Háttérváltozók - 2011 nyár 17
4.1.2. Háttérváltozók - 2012 tavasz 18
4.1.3. Háttérváltozók - 2012 nyár 19
4.2. Az egyedszámban megfigyelt változások 20
4.3. A klorofill-tartalomban megfigyelt változások 22
4.4. A taxonösszetételben megfigyelt változások 24
4.5. A fajszámban és diverzitásban bekövetkező változások 27
1

5. Összefoglalás 31
6. Köszönetnyilvánítás 33
7. Irodalomjegyzék 34
8. Függelék 43
2

1. Bevezetés és célkitűzés
Napjainkban több esetben mutattak ki nagyobb mértékű klórozott szénhidrogén
szennyezést a talajban, talajvízben az ipari hulladékká vált kismolekulájú klórozott
szénhidrogének nem megfelelő tárolása következtében. A legújabb vizsgálatok szerint a
talajvízmozgások eredményeként a kismolekulájú klórozott szénhidrogének felszíni
vizekben való megjelenésének kockázata megnőtt.
A tetraklóretán, a tetraklóretilén és a triklóretilén fizikai és kémiai tulajdonságaik
következtében (kis molekulatömeg, illékonyság, nagymértékű lipofilitás) gyorsan
abszorbeálódnak, miután kontaktusba kerülnek az élő szervezetekkel. A molekulák
toxicitásában kulcsszerepet játszanak a metabolizmusuk során keletkező reaktív termékek
(Costa és Ivanetich 1980; Bogen et al. 1988).
Annak ellenére, hogy a tetraklóretánt, a tetraklóretilént és a triklóretilént az ipar
széles körben alkalmazta, vagy alkalmazza napjainkban is, viszonylag kevés adat található
az irodalomban arra vonatkozóan, hogyan hatnak ezek az anyagok a vízi fotoszintetizáló
szervezetekre. E vegyületek fitoplankton fajokra gyakorolt hatása nem ismeretlen (Pearson
és McConnell, 1975; Bringmann és Kühn, 1980; Ward et al., 1986; Ando et al., 2003;
Lukavsky et al., 2011). Viszonylag alacsony azon tanulmányok száma, amelyek klórozott
szénhidrogén szennyezők természetes fitoplankton együttesekre gyakorolt hatását
vizsgálták sekély tavi ökoszisztémákban (Berglund et al., 2001; González et al. 2009).
Az egy fajt vizsgáló toxikológiai tesztek általánosan elterjedtek és nagyon jól
alkalmazhatók, ha fiziológiai hatások vizsgálata, toxikológiai határértékek megállapítása,
vagy különböző fajok érzékenységének összehasonlítása a cél (Moreire-Santos et al. 2005,
Liebig et al. 2008). A több fajt alkalmazó mesterséges rendszerek, vagy a természetes
közösséget tartalmazó mikrokozmosz ill. mezokozmosz vizsgálatok ökológiai relevanciája
azonban nagyobb, mert integrálják a természetes körülmények között zajló változásokat,
így a vizsgált ökoszisztéma változásainak pontosabb értékelését teszik lehetővé (Chappie
és Burton 2000, Culp et al. 2000, Moreira-Santos et al. 2004). Az algatesztek relevanciája
növelhető a természetben megjelenő fioplankton együttesek alkalmazásával (Moreira-
Santos et al. 2005). A fő előnye ezeknek a kísérleti rendszereknek a szennyezés ökológiai
hatásainak valósághű szimulációja vízi közösségekben. Így a legkülönbözőbb fajokra
gyakorolt hatások, illetve a különböző populációk közötti kölcsönhatások akár egyidejűleg
is tanulmányozhatók a közösségen belül (Lopez-Mancisidor et al. 2008).
3

Ismert, hogy az algaközösség kémiai stresszre adott válasza függ a szennyezés
pillanatában jelen lévő fajösszetételtől is (Guasch et al. 1997, Gurney és Robinson 1989,
Pesce et al. 2006, Tlili et al. 2008). McCann (2000) bevezette a "Biodiverzitás-stabilitás
hipotézist", mely szerint az ökoszisztéma ellenállása és rugalmassága növekszik a
közösségen belüli trofikus interakciók számával. Ez a hipotézis úgy is ismert, mint a
"biológiai biztosítás hipotézis" (Yatchi és Loreau, 1999), amely feltételezi, hogy diverzebb
ökoszisztémákban nagyobb valószínűséggel vannak jelen olyan fajok, amelyek képesek
ellenállni a zavarásnak és ez lehetővé teszi számukra, hogy ellensúlyozzák a más fajok
eltűnéséből származó funkcionális károsodást (Villeneuve et al. 2011).
Munkánk célja az volt, hogy in situ mikroalga teszttel (mikrokozmosz rendszerben)
értékeljük egyszeri „pontszerű” klórozott szénhidrogén szennyezések rövidtávú hatását
fitoplankton együttesek összetételére. A kísérleti összeállítás lehetővé tette annak
tanulmányozását, hogy a klórozott szénhidrogének oldódásának nem kedvező,
„természetközeli” körülmények között milyen változások zajlanak egy természetes
fitoplankton közösség szerkezetében. Munkánk során a következő kérdésekre kerestük a
választ:
- Hogyan változik különböző összetételű (diverzitású) algaközösségek produkciója
(klorofill-tartalom, egyedszám) a szennyezés hatására?
- Hogyan változik a közösségek diverzitása a szennyezés hatására a kiindulási
diverzitás függvényében?
A kísérletek eredményei egyrészt gazdagíthatják a klórozott szénhidrogének
természetkárosító hatásával kapcsolatos eddigi ismereteinket, másrészt rávilágíthatnak
arra, milyen mértékben használhatók a mikrokozmosz rendszerek szennyezések hatásainak
modellezésére, segítve az adott szennyezéssel szemben ellenálló fajok megtalálását,
melyeknek nagy szerepe lehet a későbbiekben a szennyezés biológiai eltávolítására
irányuló kutatásokban is.
4

2. Irodalmi áttekintés
2.1. Kismolekulájú klórozott szénhidrogének
2.1.1. Tetraklóretán
Az 1,1,2,2-tetraklóretán egy szintetikus, színtelen, sűrű folyadék, ami nem ég
könnyen. A kloroformhoz hasonló átható, édes szaga van. A molekula szerkezete az 1.
ábrán látható. Korábban nagy mennyiségben használták más vegyszerek előállítására,
festékekben és növényvédő-szerekben, valamint ipari oldószerként is, illetve zsírok és
olajok más anyagoktól való szétválasztására, fémek tisztítása és zsírtalanítása céljából,
továbbá szennyvíztisztítás során, vagy fotografikus filmekben található fémek
kivonására.(Lewis 2001). Ma már elérhető kevésbé mérgező vegyszer a helyettesítésére,
mivel manapság nagymértékben fejlődik a klórozott etilének előállításának technológiája.
Nagy léptékű kereskedelmi előállítása is megszűnt, bár néha még előfordul. Egyéb
elnevezései az acetilén-tetraklorid, diklór-2,2-diklóretán, s-tetraklóretán. Molekulatömege:
167,85g/M; olvadáspontja: -42,4°C; forráspontja: 145,2°C. Kémiai képlete: C2H2Cl4.
1. ábra. A tetraklóetán kémiai szerkezete
A tetraklóretánt olyan gyárakban állítják elő melléktermékként, ahol 1,1,1- és 1,1,2-
triklóretilént gyártanak (Rossberg et al. 2005). A más vegyületek előállítása során
megmaradó tetraklóretánt vagy a természetbe juttatják vagy szennyezőként jelenik meg a
végtermékekben.
2.1.1.1. A tetraklóretán sorsa a természetben
A felszíni vizekbe kerülő tetraklóretán, annak ellenére, hogy viszonylag nagy
mennyiség képes oldódni vízben (2,83g × L-1 25°C-on) általában napoktól hetekig tartó idő
alatt elpárolog, a víz mozgásától függően. Szennyezett felszíni vagy felszín alatti vizek
5

esetén ezért az egyik leggyakrabban alkalmazott tisztítási módszer a kilevegőztetés
(Kincannon et al. 1983). A víznél nagyobb sűrűsége miatt azonban nem mozgó víztérben a
vízfenékre süllyed, ahonnan folyamatosan a víztestbe kerülve károsíthatja a vízi
szervezeteket. A tetraklóretán vízben természetes pH-n bázis katalizált hidrolízisen megy
keresztül, melynek során triklóretilén keletkezik (Cervini-Silva 2003). A vízi élőlényekben
való felhalmozódása nem jellemző folyamat (a vízi állatok szervezetében metabolizálódik,
Franke et al. 1994).
Közepes gőznyomásának és a talajrészecskékhez való viszonylag alacsony
adszorpciójának köszönhetően a talaj felszínére kerülő tetraklóretán hamar elpárolog
(Swann et al. 1983). Folyamatos, hosszan tartó szennyezés esetén azonban a talajban és a
talajvízben is nagy mennyiségben jelen lehet, ahol hidrolízise és anaerob biodegradációja
is bekövetkezhet (Lorah és Olsen 1999).
2.1.1.2. A tetraklóretán metabolizmusa és a toxicitás mechanizmusa
A domináns útvonalak a diklórecetsav produkciót segítik elő, ami nem enzimatikus
degradáció, diklóracetil kloridot és diklóracetaldehidet eredményez köztes termékként.
Más útvonalakon a triklóretilén vagy tetraklóretilén keletkezése figyelhető meg, amelyek
trilkóretanolt, triklórecetsavat és oxálsavat eredményező reakciókat vonnak maguk után,
(Yllner 1971; Mitoma et al. 1985). Elképzelhető, hogy ezek az anyagcsereutak az
egysejtűekben is hasonló módon zajlanak.
A tetraklóretán reaktív termékekké történő bomlása valószínűleg kulcsfontosságú
szerepet játszik a toxikusságában. Mind a nukleáris, mind a mikroszomális citokróm P450
enzimek megjelennek az összetevők metabolizmusa során, amelyek valószínűleg számos
biológiailag aktív elemet szabadítanak fel, beleértve az aldehideket, alkéneket, savakat és
szabad gyököket (Mitoma et al. 1985; Hanley et al. 1988). A citokróm P450-ről kiderült,
hogy katalizálja mind a diklóracetilált fehérjék melléktermékeinek (Halpert 1982), mind a
dilkórecetsav keletkezését (Halpert és Neal 1981). Tomasi és munkatársai (1984) is
bemutatták, hogy a tetraklóretán metabolizmusa során szabad gyökök keletkeznek. Ezek a
felfedezések együttesen megmutatják, hogy a tetraklóretán anyagcseréje gyökök képzésébe
torkollik, ami lipid peroxidációs stimulációhoz vezet.
6

A tetraklóretán alacsony genotoxikus aktivitással bír. Bebizonyosodott, hogy DNS-
hez kötődhet (Colacci et al. 1987; Hanley et al. 1988), az abszorpciót követően szinte
teljesen lebomlik, feltételezhetően részben a citokróm P450 következtében.
2.1.2. Tetraklóretilén
A tetraklóretilén egy szintetikus, könnyen párolgó, szúrós, édes szagú,
szobahőmérsékleten folyékony halmazállapotú, színtelen, nem gyúlékony vegyület, a
molekula szerkezetét a 2. ábra mutatja. Gazdasági szempontból fontos klórozott
szénhidrogén oldószer és köztes termék. Mióta ismert, hogy a tetraklóretilén képes
feloldani számos szerves összetevőt, elválasztani szervetlen vegyületeket, azóta széles
körben alkalmazzák ruhaanyagok száraztisztítására és fémek zsírtalanítására (Verschueren
1983). Más vegyületek előállításakor is használják kezdő vegyületként, számos fogyasztói
termék esetén is. Egyéb elnevezései az etilén-tetraklorid, 1,1,2,2-tetraklóretilén,
tetraklóretén, perklóretilén (PCE), perklén és perklór. Molekula súlya 165,83g/M;
olvadáspontja: -19°C; forráspontja: 129°C. Kémiai képlete C2Cl4.
2. ábra. A tetraklóretilén kémiai szerkezete
Tetraklóretilént számos mérsékelt és szubtrópusi tengerben élő makroalga termel
0,0026-8,2 ng/g tiszta tömeg óránkénti mennyiségben. Azt is kimutatták, hogy ezek az
algák ennél nagyobb mennyiségben állítanak elő triklóretilént. Az algák által a légkörbe
juttatott triklóretilén és tetraklóretilén mennyisége olyan méreteket ölt, hogy azt nem lehet
figyelmen kívül hagyni a légkör teljes klór-tartalmának kiszámításakor (Abrahamsson et
al. 1995).
2.1.2.1. A tetraklóretilén sorsa a természetben
Az újabb szabályok tiltják az olyan klórozott oldószerek természetben való
elhelyezését, amelyek tetraklóretilént tartalmaznak. Minden olyan szilárd hulladék, amely
tetraklóretilént tartalmaz veszélyes hulladéknak minősül. A tetrtaklóretilén lebomlási és
7

eltávolítási hatékonyságának 99,95-osnak kell lennie, mivel elsődleges szerves
szennyezőnek számít. Mielőtt a szennyezőanyagokat kihelyeznénk a szeméttelepekre,
azokat be kell vizsgáltatni a természetvédelmi szervezetekkel (HSDB 1996).
A felszíni vizekbe kerülő tetraklóretilén nem alakul át azonnal. Vízoldhatósága
csekély (0,15g × L-1 25°C-on), de sűrűségének köszönhetően (1,6227g × cm3) a vízfenékre
süllyed, ahol változatlan formában hosszabb ideig jelen lehet, mivel a vizsgálatok azt
mutatják, hogy természetes körülmények között hidrolízise nem megy végbe (csak magas
hőmérsékleten és pH-n). Természetes vizekben a tetraklóretilén átalakulásának
legvalószínűbb módja a biodegradáció (Parsons et al. 1984, 1985). Laboratóriumi
kísérletek eredményei azt mutatják, hogy a tetraklóretilén reduktív dehalogénezésen esik
át, aminek eredményeként etilén keletkezik. Az átalakulás metanogén körülmények között
megy végbe (Freedman és Gossett 1989).
A talajba, talajvízbe kerülő tetraklóretilén biodegradációja csak speciális
körülmények között megy végbe, és lassú folyamat. Bizonyos kísérletekben a
tetraklóretilén koncentrációjának csökkenése mellett a triklóretilén koncentrációjának
növekedéséről számoltak be, ami azt sugallja, hogy ez a fő termék (Milde et al. 1988;
Freedman és Gossett 1989). A tetraklóretilén talajból való párolgása sokkal kisebb
mértékű, mint a vízből való párolgás. A talajból való párolgás mértékét befolyásolja a jelen
lévő tetraklóretilén koncentrációja és a talaj típusa is – magas szervesanyag tartalmú
talajokban a folyamat lényegesen lassabb (Yagi et al. 1992).
2.1.2.2. A tetraklóretilén metabolizmusa és a toxicitás mechanizmusa
A molekula metabolizmusa egysejtűekben nem ismert, leszámítva a metanogén
körülmények között zajló biodegradációt. A tetraklóretilén megváltoztatja a foszfolipidek
és aminosavak zsírsav szerkezetét (Kyrklund et al. 1990). A citoszkeletális elemek jóval
érzékenyebbek, mint a citoszol fehérjéi. A tetraklóretilén a membránok ATP-áz aktivitását
is csökkenti (Kukongviriyapan et al. 1990), ami arra enged következtetni, hogy a
tetraklóretilén hatásai a transzportra eredhetnek a csökkent ATP szintből és a sejtmembrán
ATP-áz gátlásából. A tetraklóretilén TCA-vá alakulása a P450 útvonalon telítődhet.
8

2.1.3. Triklóretilén
A triklóretilén egy nem gyúlékony, színtelen, szobahőmérsékleten folyékony, édes
szagú és égető ízű anyag. A molekula szerkezete a 3. ábrán látható. A triklóretilént főként
oldószerként használják fémek zsírtalanításakor, valamint más vegyszerek előállítása során
(Kuney 1986). Triklóretilént találhatunk néhány háztartási termékben is, mint például a
hibajavító folyadékban, festék eltávolítókban, ragasztókban és folttisztítókban
(Frankenberry et al. 1987). Egyéb elnevezései az acetilén triklorid, 1-klór-2,2-diklóretilén,
1,1-diklór-2-klóretilén, etilén triklorid, triklorid, 1,1,2-triklór-etilén, triklóretén, TCE.
Imserhetjük még triklén-ként vagy vitran-ként is. A triklóretilén molekulatömege
131,40g/M; olvadáspontja: -87,1°C; forráspontja: 86,7°C. Kémiai képlete: C2HCl3.
3. ábra. A triklóretilén kémiai szerkezete
A triklóretilén szennyező eliminálásának ajánlott módja az égetés (ezt azután
végzik, hogy valamilyen gyúlékony anyaggal összekeverték – Sittig, 1985). A foszgének
keletkezését meg kell gátolni az égetés során, így erre nagy gondot fordítanak (Sjoberg
1952). A tökéletlen égés során keletkező más mérgező vegyületek a policiklikus aromás
szénhidrogének és a perklóros aromás vegyületek (Blakenship et al. 1994). Az USA-ban
nagy hangsúlyt fektettek a triklóretilén újrahasznosítására, hogy csökkentsék ennek a
fotoreaktív vegyületnek a légkörbe jutását (CMR 1986). A fotooxidatív lebomlást
sikeresen alkalmazták a levegő ’kigőzölési’ technikával kapcsolatban, amellyel sikerült
elpárologtatni a triklóretilént a vízből és így nem mérgező vegyületeket sikerült előállítani
(Bhowmick és Semmens 1994). Amennyiben lehetséges a szemétlerakás helyett az
újrahasznosítást kell alkalmazni.
9

2.1.3.1. A triklóretilén sorsa a természetben
Számos formában a levegőbe és a vízbe juthat a vegyület, például a nem megfelelő
hulladékkezeléssel, festékekből való párolgással, ragasztókból és egyéb termékekből, vagy
gyárakból való felszabadulás során.
Vízi környezetben a triklóretilén oxidációja nem jellemző folyamat, hidrolízise
szintén lassan megy végbe, inkább csak magasabb hőmérsékleten és pH-n. A triklóretilén
degradációjának legjelentősebb módja a reduktív dehalogénezés mikroorganizmusok által
(Parsons et al. 1985; Wilson et al. 1986). A triklóretilén vízoldhatósága közepesnek
mondható a kismolekulájú klórozott szénhidrogének körében (1,366g × L-1 25°C-on). A
vízbe kerülő triklóretilén nagy része sűrűségének köszönhetően (1,465g × cm3) a
vízfenékre süllyed, ahonnan folyamatosan oldódva károsíthatja a fölötte lévő víztestben
előforduló élőlényeket.
Az irodalomban található vizsgálati eredmények azt mutatják, hogy a talajba kerülő
triklóretilén nem alakul át kémiailag és nem kötődik kovalensen a talajrészecskékhez. A
triklóretilén biodegradációja magas szervesanyag tartalmú talajokban gyorsabban megy
végbe (Barrio-Lage et al. 1987). A folyamat aromás szénhidrogének együttes
metabolizációjával zajlik, így koszubsztrát – fenol vagy toluol – jelenlétét igényli (Nelson
et al. 1987; Fan és Scow 1993). A vizsgálatok azt mutatják, hogy a triklóretilén
degradálására képes baktériumok metánt használnak energiaforrásként, és a triklóretilént
szimultán bontják a metán monooxigenáz enzim közreműködésével (Bowman et al. 1993).
A metánt hasznosító baktériumok aerob körülmények között végzik a triklóretilén bontását
(Wilson and Wilson 1985), a folyamat érzékeny egyensúly meglétét igényli a szintén
toxikus koszubsztrátok jelenléte miatt (Ensley 1991).
2.1.3.2. A triklóretilén metabolizmusa és a toxicitás mechanizmusa
A triklóretilén legfőbb metabolitjai a TCA, a triklóretanol és a konjugálódott
triklóretanol (Dekant et al. 1984). Arról is készültek feljegyzések, hogy a kloroform is a
triklóretilén egyik kisebb mennyiségben keletkező metabolitja (Pfaffenberger et al. 1980);
jóllehet ez az állítás megkérdőjelezhető és további bizonyítékokat igényel, mivel a
kloroform lehet az anyagcsere termékek meghatározására használt analatikai eljárások
mesterséges mellékterméke is.
10

Az anyagcsere fontos szerepet játszik a triklóretilén toxikusságában, mivel számos
metabolitja önmagában is mérgező. A különböző fajok eltérő módon reagálnak a
triklóretilénnek való kitettségre, ami betudható annak, hogy eltérő mértékben
metabolizálják az eredeti vegyületet (Dekant et al. 1986b).
2.2. A klórozott szénhidrogének hatása a fitoplanktonra
Bár a tetraklóretánt, a tetraklóretilént és a triklóretilént az ipar széles körben
alkalmazta, vagy alkalmazza napjainkban is, viszonylag kevés adat áll rendelkezésre az
irodalomban vízi ökoszisztémákra, különösen a termelő szervezetekre gyakorolt hatásaikra
vonatkozóan. Ismeretes, hogy ezek a vegyületek hatással lehetnek különböző algákra:
hatásaikat tanulmányozták tengeri környezetben (Pearson és McConnell 1975; Ward et al.
1986), valamint édesvízi cianobaktériumok és eukarióta algák esetében (Bringmann és
Kühn 1980; Ando et al. 2003; Lukavsky et al., 2011; Bácsi et al. 2012).
Viszonylag alacsony azon tanulmányok száma, amelyek klórozott szénhidrogén
szennyezők természetes fitoplankton együttesekre gyakorolt hatását vizsgálták sekély tavi
ökoszisztémákban. Berglund és munkatársai (2001) pozitív korelációt mutattak ki a tó
trofikus állapota és a poliklórozott bifenilek mennyisége között. Arra a következtetésre
jutottak, hogy ez a sekély, eutróf tavakra jellemző magasabb ülepedési rátának köszönhető.
A szénhidrogén szennyezők fitoplankton közösségre gyakorolt hatásának vizsgálata során
kimutatták, hogy a szennyezők vízoldható frakciója átmeneti, rövidtávú negatív hatással
volt a fitoplanktonra, amely később változásokat okozott a fitoplankton közösség
szerkezetében (González et al. 2009).
2.3. Mikrokozmosz rendszerek
Az egy fajt alkalmazó laboratóriumi toxicitási teszteket általánosan alkalmazzák
arra, hogy a szennyező anyagok vízi ökoszisztémára gyakorolt potenciális veszélyeit
értékeljék (Moreira-Santos et al. 2005). Ezek a tesztek előnyösek, ha az előírások
megkövetelik a szennyező anyag hatásaira vonatkozó adatok ismételhetőségét és
összehasonlíthatóságát, azonban nem tükrözik a természetes rendszerek elemei közötti
11

kapcsolatokat, mint pl. a ragadozó-zsákmány kapcsolat, vagy a tápanyagok körforgalma
(Liebig et al. 2008).
A laboratóriumi, sok fajt használó, meghatározott összetételű mikrokozmosz
rendszerek átmenetet képeznek az egy fajt alkalmazó tesztek és a természetes élőlény-
együtteseket tartalmazó laboratóriumi, vagy terepi kísérletek között. A laboratóriumi több
fajt alkalmazó tesztek hasznosak lehetnek a magasabb szintű kockázatbecslésnél, segítenek
a terepi mezokozmosz vizsgálatok megtervezésében, illetve segítenek a mezokozmoszban
megfigyelt eredmények ok-okozati összefüggéseinek tisztázásában (Campbell et al., 1999).
Az ilyen modell ökoszisztémák, melyek utánozzák az édesvízi környezeteket (pl.
mikrokozmosz és mezokozmosz) egyre gyakrabban használt eszközök a
szennyezőanyagok ökotoxikológiai veszélyeinek felmérésére (Touart 1988; Graney et al.
1994; Hill et al. 1994). Ezeknek a kísérleti rendszereknek nagy előnye, hogy reális
ökológiai hatásokat szimulálnak a szennyező anyagok a vízi közösségekbe jutása után. A
mikrokozmosz, illetve mezokozmosz rendszerek lehetővé teszik a szennyezőanyagok
hatásainak tanulmányozását a fajok széles skáláján, miközben populációs kölcsönhatások
is érvényesülnek a közösségen belül (Lopez-Mancisidor 2008). A több fajt vizsgáló
rendszerek lehetővé teszik a lehetséges közvetett hatások vizsgálatát, beleértve az egymást
követő hatásokat is (Lawler, 1993; Kooi, 2003).
Az in situ terepi vizsgálatok ökológiai relevanciája a legnagyobb, mert a
természetesen változó feltételeket integrálják minimális beavatkozással (Chappie és Burton
2000; Moriera-Santos et al. 2004). Külön előnye ezeknek a rendszereknek, hogy a
vizsgálatok a helyi faj-együtteseken alapulnak, amelyek a helyi körülményekhez
alkalmazkodtak és nem kell őket alávetni a laboratóriumba való szállítás és akklimatizáció
során fellépő stressznek. A módszer ökológiai jelentőségét növeli az is, hogy nincsenek
nem őshonos fajok használatából adódó komplikációk (Chappie és Burton 2000).
Az a felismerés, hogy minden egyes toxikológiai eszköznek saját értéke van,
vezetett el az integrált toxikológiai vizsgálatok kialakulásához, melyek egyesítik a
laboratóriumban születő információkat és az in situ eszközöket, és lehetőséget adnak az
ökoszisztéma minőségének még pontosabb felmérésére (Clup et al. 2000). A laboratóriumi
algateszteket már régóta rutinszerűen alkalmazzák a szennyező anyagok ökoszisztémára
gyakorolt hatásainak felmérésére (OECD 1984; USEPA 1994), míg az algák in situ
vizsgálata még kevésbé elterjedt (Moreisa-Santos et al. 2004).
12

2.4. A biológiai sokféleség hatása az ökoszisztéma szennyezőkkel szembeni
érzékenységére
A közelmúltban a biológiai sokféleség ökológiai rendszerek működésében betöltött
szerepe egyre nagyobb figyelmet kap az egyre fokozódó aggodalomnak köszönhetően,
miszerint a biológiai sokféleség csökkenése ronthatja az ökoszisztéma működését (Ehrlich
és Wilson, 1991; Schulze és Mooney, 1993; Vitousek et al., 1997; Chapin et al., 1997;
Costanza et al., 1997).
McCann (2000) bevezette a "Biodiverzitás-stabilitás hipotézist", mely szerint az
ökoszisztéma ellenállása és rugalmassága növekszik a közösségen belüli trofikus
interakciók számával. Ez a hipotézis úgy is ismert, mint a "biológiai biztosítás hipotézis"
(Yatchi és Loreau, 1999). Bár a biológiai sokféleség hatása az ökoszisztéma
működésére az ökológia egyik legkutatottabb területévé vált, a diverzitás
jelentősége a változó környezetben még kevéssé érthető. A „biztosítás hipotézis” szerint, a
biodiverzitás biztosítja, hogy az ökoszisztéma működése ne sérüljön, hiszen sok faj
nagyobb garanciát biztosít arra, hogy fenntartja a rendszer működését akkor is, ha bizonyos
fajok kiesnek (Yatchi és Loreau, 1999). A tapasztalat azt mutatja, hogy hosszú távon
minden ökológiai rendszer ki van téve környezeti változásoknak (Chesson és Chase, 1986).
Ezért a kritikus kérdés az, hogyan hat a biodiverzitás, vagy a biodiverzitás csökkenése az
ökológiai rendszer működésére változó környezetben (Yatchi és Loreau, 1999). A
„biztosítás hipotézis” feltételezése, hogy a diverzebb ökoszisztémákban nagyobb
valószínűséggel vannak jelen olyan fajok, amelyek képesek ellenállni a zavarásnak és ez
lehetővé teszi számukra, hogy ellensúlyozzák a más fajok eltűnéséből származó
funkcionális károsodást. Ez a feltételezés azért állhatja meg a helyét, mert a különböző
fajok eltérően reagálnak a környezeti változásokra, míg egyes fajok „hozzájárulása” a
rendszer működéséhez csökkenhet, addig másoké nőhet a változó környezeti feltételek
között. Nagyobb fajgazdagság a rendszerben zajló folyamatok kisebb változatosságához
vezethet, a fajok közötti kompenzációs folyamatok miatt. (Yatchi és Loreau, 1999).
Yatchi és Loreau (1999) elméleti modelljének kísérletes ellenőrzésére
algaközösségekre vonatkozóan a következőkben összefoglalt tanulmányok említhetők.
O’Connor és Crowe (2005) terepi zárt rendszerben végzett megfigyeléseik alkalmával nem
találtak összefüggést az ökológiai rendszer működése, és a diverzitás között, azonban az
egyes fajok sajátos hatásait lehetett kimutatni. Goodsell és Underwood (2008) hasonló
megfigyelésre jutottak tengerparti makroalga-közösség megfigyelése során.
13

Li és munkatársai (2010) laboratóriumi mikrokozmosz kísérletekben vizsgálták a
kadmium hatását. Eredményeik azt mutatták, hogy a nem szennyezett közösségben nem
volt pozitív korreláció a fajgazdagság és a produktivitás között, míg kadmium szennyezés
hatására a nagyobb diverzitással jellemezhető közösségben egyértelműen nagyobb volt a
produktivitás is. Külön érdekesség, hogy nemcsak kadmium toleráns fajok, hanem
érzékeny fajok is nagymértékben hozzá tudtak járulni a közösség primer produkciójához.
A szerzők azt a következtetést vonták le a kísérleti eredményekből, hogy a biodiverzitás
megőrzése segíthet csökkenteni az ökológiai rendszerek működését (pl. elsődleges
termelést) érő stresszt, illetve diverz algaközösségek kialakítása szennyezett vizek
kezelésében is előnyös lehet (Li et al. 2010).
Villeneuve és munkatársai (2011) a kémiai és fizikai faktorok perifiton
szerkezetére, sokféleségére és működésére gyakorolt hatásait vizsgálták egy szabadtéri
mezokozmosz kísérletben. A bentikus mikrobiális közösségek (biofilm vagy perifiton)
fontos szerepet játszanak a felszíni vizek ökológiai működésében (Hansson, 1992; Munoz
et al., 2001; Pusch et al.,1998; Romani és Sabater, 1999; Woodruff et al., 1999). A
perifiton kialakulása, szerkezete, taxonómiai sokfélesége és működése nagymértékben
függ a fizikai és kémiai tényezőktől (Biggs, 2000; Hillebrand és Sommer, 2000; Sabater et
al., 1998; Villeneuve et al., 2010). Ezen különböző tényezők bármilyen módosítása, akár
természetes eredetű vagy emberi tevékenységből adódó, ronthatja a biofilm képességét
arra, hogy betöltse funkcióját az ökológiai rendszerben. Ismert, hogy az algaközösség
kémiai stresszre adott válasza függ a szennyezés pillanatában jelen lévő fajösszetételtől is
(Guasch et al. 1997, Gurney és Robinson 1989, Pesce et al. 2006, Tlili et al. 2008). A
kutatócsoport eredményei azt mutatták, hogy a beállított kísérleti körülmények között a
nagyobb biológiai sokféleséggel bíró közösségben nem nőtt a biofilm szennyezőkkel
(peszticidek) szembeni ellenálló képessége, éppen ellenkezőleg, ezen közösségek
érzékenysége a peszticid szennyezésre valójában fokozódott Villeneuve et al. 2011).
Úgy tűnik tehát, hogy a vizsgált közösség tulajdonságaitól, illetve a zavarás
típusától függ, hogy a kezdeti diverzitás hogyan befolyásolja a közösség „válaszát” a
zavarásra. A kérdés minél alaposabb megértése érdekében az ilyen irányú vizsgálatok
végzése mindenképpen indokolt, figyelembe véve a napjainkban az ökológiai rendszereket
érő mind „változatosságában”, mind mértékében egyre fokozódó terhelést.
14

3. Anyag és módszer 3.1. A mikrokozmosz rendszerek összeállítása
A „terepi” vizsgálatokat a Debreceni Egyetem Botanikus Kertjében, a Botanikus
Kerti Tóban végeztük. A kerti tó egy sekély mesterséges víztér (átlagos mélység: 0,7 m;
terület: kb. 100 m2), amely jól reprezentálja Magyarország felszíni vizeinek egyik
leggyakoribb típusát. A tóból 12 l vízmintát vettünk, amelyet szétosztottunk 4 műanyag
(polimetilpentén - PMP) főzőpohárba (3 l vízminta főzőpoharanként). 1 főzőpohár szolgált
kontrollként, 1-1 főzőpohárban végeztük az egyes klórozott szénhidrogénekkel a
kezeléseket. Az elméletileg telített oldat eléréséhez szükséges mennyiségű klórozott
szénhidrogént, azaz 5,4 ml tetraklóretánt, 276 µl tetraklóretilént, illetve 2,805 ml
triklóretilént adtunk a főzőpoharakba vett vízmintákhoz. A főzőpoharakat műanyag
kosárba helyeztük, amelyet a tó 20 cm mélységű parti zónájába állítottunk. A kosarat
átlátszó műanyag (poliészter) tetővel láttuk el, mely lehetővé tette a fotoszintézist,
ugyanakkor megóvta a kosár tartalmát a károsodástól (4. ábra). Az expozíciós idő alatt a
reggeli órákban WTW Multi 340i készülékkel mértük a tóban és az egyes
összeállításokban a hőmérsékletet, pH-t, vezetőképességet, O2 koncentrációt és O2
telítettséget. A kísérlet 0., 24., 48. és 72. órájában 20 ml mintát vettünk minden
főzőpohárból, illetve a tóból a kosár környékéről.
4. ábra. A mikrokozmosz rendszerek összeállítása.
15

3.2. A fitoplankton növekedésének nyomon követése
A klorofill-a tartalom meghatározásához a mintákból 5 ml-t centrifugáltunk (Micro-
centrifuge Type-320a, 5 perc, 10000 rpm), a felülúszó eltávolítása után a pelletet
liofilizáltuk (Christ Alpha 1-2 LD plus), a klorofill-a tartalmat metanolos extrakcióval,
spektrofotometriásan határoztuk meg (Felföldy 1987).
A maradék mintát (15 ml) Lugol-oldatban tartósítottuk a fitoplankton mennyiségi
és minőségi meghatározásához, amit az Utermöhl módszer (1958), valamint az EN 15204
(2006) Európai Standard alapján végeztünk. A mikroszkópos határozás és számlálás
Olympos CKX31 fordított mikroszkóppal, 200 illetve 400× nagyításon történt. Az
azonosított fajok rendszertani besorolását az AlgaeBase internetes adatbázis alapján
végeztük el (Guiry, 2013).
3.3. Az eredmények statisztikai értékelése
A méréseket és meghatározásokat háromszoros ismétlésben végeztük el. Az
eredmények statisztikai értékelésére kétutas ANOVA-t és Tukey-tesztet alkalmaztunk
(SigmaStat-3.10 szoftver).
A diverzitásmutatók közül a ritka fajokra érzékeny Shannon-Wiener indexet (H) és
a legtömegesebb fajra érzékeny Berger-Parker indexet (d) használtuk (Tóthmérész 1997),
számításukat a PAST program segítségével végeztük.
16

4. Eredmények és értékelésük
4.1. A háttérváltozókban megfigyelt változások az egyes vizsgálati időszakokban
4.1.1. Háttérváltozók - 2011 nyár
A 2011 nyarán végzett mikrokozmosz kísérletek során a Botanikus-kerti tóban,
valamint a kontroll és klórozott szénhidrogénekkel kezelt edényekben a hőmérséklet
szignifikánsan alacsonyabb volt a kísérlet végén (72. óra), mint a kísérlet indításakor (4a.
ábra). A közel 6 °C-os eltérés oka egy nyári vihar volt. A különböző összeállítások és a tó
hőmérséklete nem tért el egymástól (4a. ábra). Míg a pH és a vezetőképesség nem tért el
szignifikánsan egymástól az egyes edényekben, valamint a tóban (4b-c. ábra), addig a
tóban mért O2 koncentráció és O2 telítettség már az első naptól kezdve szignifikánsan
alacsonyabb volt a kontroll összeállításhoz képest (4d-e. ábra). A különböző klórozott
szénhidrogénekkel kezelt közösségek O2 termelése a kontroll és tó közösségének O2
termelése közé esett, viszont ezek már nem tértek el szignifikánsan sem egymástól sem a
tótól és a kontrolltól (4d-e. ábra).
4. ábra. A háttérváltozók alakulása a Botanikus kerti tóban és az egyes összeállításokban a
2011 nyarán végzett kísérlet során. a) vízhőmérséklet, b) pH, c)vezetőképesség, d oxigén-
17

koncentráció, e) oxigén telítettség. TeCa: tetraklóretán, PCE: tetraklóretilén
(perchloroethylen), TCE: triklóretilén (trichloroethylen)
4.1.2. Háttérváltozók - 2012 tavasz
A 2012 tavaszi kísérletek során a kontroll és a kezelt edényekben a hőmérséklet
szignifikánsan nem tért el egymástól, a tó hőmérséklete a 2. naptól kezdve szignifikánsan
magasabb volt, mint az edényekben (5a. ábra). Ennek oka feltehetően abban keresendő,
hogy a mintavételekre a korai órákban (reggel 8-9 óra) került sor, az edényekben a
viszonylag kis térfogat miatt nagyobb lehetett a hőingás, mint a tóban: így éjszaka
gyorsabban és jobban le tudott hűlni a víz az edényekben. Hasonlóan a 2011 nyarán
tapasztaltakhoz, 2012 tavaszán sem volt szignifikáns különbség a tóban és az edényekben
mérhető pH és vezetőképesség értékek között (5b-c. ábra). Ezzel szemben a tó O2 tartalma
és O2 telítettsége már az első naptól kezdve szignifikánsan alacsonyabb volt, mint a
kontroll és kezelt közösségeké (5d-e. ábra).
5. ábra. A háttérváltozók alakulása a Botanikus kerti tóban és az egyes összeállításokban a
2012 tavaszán végzett kísérlet során. a) vízhőmérséklet, b) pH, c)vezetőképesség, d
18

oxigén-koncentráció, e) oxigén telítettség. TeCa: tetraklóretán, PCE: tetraklóretilén
(perchloroethylen), TCE: triklóretilén (trichloroethylen)
4.1.3. Háttérváltozók - 2012 nyár
A 2012 nyarán végzett mikrokozmosz kísérletek során a tó, a kontroll és a klórozott
szénhidrogénekkel kezelt edények hőmérséklete, pH-ja és vezetőképessége nem tért el
szignifikánsan egymástól (6a-c. ábra). A tó O2 tartalma és O2 telítettsége a kísérlet ideje
alatt szignifikánsan alacsonyabb volt az edényekénél (6d-e. ábra), ahogyan az a korábbi
kísérletek során is megfigyelhető volt.
6. ábra. A háttérváltozók alakulása a Botanikus kerti tóban és az egyes összeállításokban a
2012 nyarán végzett kísérlet során. a) vízhőmérséklet, b) pH, c)vezetőképesség, d oxigén-
koncentráció, e) oxigén telítettség. TeCa: tetraklóretán, PCE: tetraklóretilén
(perchloroethylen), TCE: triklóretilén (trichloroethylen)
19

4.2. Az egyedszámban megfigyelt változások
A 2011 nyarán végzett mikrokozmosz kísérletek során a 24. órára mind a tó, mind a
kontroll, mind pedig a klórozott szénhidrogénekkel kezelt közösségek egyedszáma
szignifikánsan alacsonyabb volt, mint a kísérlet indulásakor (7a. ábra). Ez feltehetően
annak a nyári viharnak volt köszönhető, ami a 6 °C-os hőmérséklet csökkenést okozta (4a.
ábra). Azonban míg a tó és kontroll közösségek egyedszáma nem tért el egymástól
szignifikánsan, addig a kezelt edényekben szignifikánsan alacsonyabb volt az algaszám,
mint a tóban és kontrollban (7a. ábra). A második napra minden közösségben nőtt az
egyedszám (7a. ábra), azonban a tetraklór-származékokkal kezelt alga-együttes
egyedszáma szignifikánsan alacsonyabb volt a többinél. A kísérlet legvégén a kezelt
edényekben az algaszám minden esetben alacsonyabb volt, mint a kontrollban, vagy a
tóban, szignifikáns különbség csak a tetraklóretánnal (TeCa) kezelt közösség esetén volt
kimutatható (7a. ábra).
A 2012 tavaszi kísérletek alkalmával nőtt az egyedszám a tóban, valamint a
kontroll és a kezelt együttesekben is az első 24 órában (7b. ábra), a közösségek
egyedszámai között ekkor még szignifikáns különbségek nem voltak kimutathatók (7b.
ábra). A második napra a TeCa kivételével minden együttesben tovább emelkedett az
algaszám. A TeCa-val kezelt közösségben azonban 70%-kal csökkent az egyedszám (7b.
ábra). A TeCa-val kezelt együttes algaszáma a 2. naptól kezdve a kísérlet további részében
szignifikánsan alacsonyabb volt, mint a tó, a kontroll, vagy a többi kezelt együttes
algaszáma (7b. ábra). A második naptól a tetraklóretilénnel (PCE) kezelt közösség
egyedszáma pedig szignifikánsan magasabb volt a többi közösségénél (7b. ábra).
A 2012 nyarán végzett mikrokozmosz kísérletekben az algaszám az első 24. órában
minden esetben valamelyest csökkent (max. csökkenés a triklóretilénnel (TCE) kezelt
együttesben - 34%; 7c. ábra). Szignifikáns különbségek azonban nem voltak kimutathatók
az egyes közösségek között. A 2. naptól kezdve a kontroll közösség egyedszáma
szignifikánsan magasabb volt, mint a többi közösségé (7c. ábra). A tó és a klórozott
szénhidrogénekkel kezelt közössége egyedszáma között azonban nem volt kimutatható
különbség (7c. ábra).
20

7. á
bra.
Egy
edsz
ámvá
ltoz
ások
a B
otan
ikus
ker
ti tó
ban
és a
z eg
yes
kísé
rlet
i ös
szeá
llít
ások
ban
a) 2
011
nyar
án;
b) 2
012
tava
szán
és
c) 2
012
nyar
án. T
eCa:
tetr
akló
retá
n, P
CE
: tet
rakl
óret
ilén
(pe
rchl
oroe
thyl
en),
TC
E: t
rikl
óret
ilén
(tr
ichl
oroe
thyl
en)
21

4.3. A klorofill-tartalomban megfigyelt változások
A 2011 nyarán végzett kísérletek során általánosan megfigyelhető volt, hogy a
klorofill-tartalom mind a tóban, mind a kontroll edényekben, mind pedig a kezelt
edényekben csökkent az első 24 órában (8a. ábra). Azonban míg a tóban és a kontroll
közösségben a csökkenés mértéke 50-60% volt, addig a kezelt közösségekben ennél
magasabb volt (70-90%; 8a. ábra). A csökkenés oka, ahogy arra már az egyedszám-
változások esetében utaltunk (7a. ábra), feltehetően az első éjszakai nyári vihar volt. Ezt
követően a 2. napra a kontroll közösségben és a tóban emelkedett, a klórozott
szénhidrogénekkel kezelt közösségekben azonban tovább csökkent a klorofill-tartalom (8a.
ábra). A kezelt alga-együttesek klorofill-tartalma a 2. naptól kezdve szignifikánsan
alacsonyabb volt, mint a tóé, és a kontroll közösségé (8a. ábra). A TCE-vel kezelt közösség
klorofill-tartalma azonban szignifikánsan magasabb volt, mint a TeCa-val, és PCE-vel
kezelt együttesé (8a. ábra).
A 2012 tavaszi kísérletek során azt tapasztaltuk, hogy a tó és a kontroll közösség
klorofill-tartalma már az első naptól kezdve szignifikánsan magasabb volt, mint a kezelt
közösségeké (8b. ábra). A PCE-vel kezelt együttes klorofill-tartalma a 2. naptól kezdve
szignifikánsan magasabb volt, mint a TeCa-val, vagy TCE-vel kezelteké. A TeCa kezelés
hatására a közösség klorofill-tartalma a 48. órától kezdve szignifikánsan alacsonyabb volt a
többinél (8b. ábra).
2012 nyarán a közösségek klorofill-tartalma, hasonlóan az egyedszámban
bekövetkezett változásokhoz (7c. ábra), jelentős mértékben csökkent az első napra (max.
86% - kontroll; 8c. ábra). A 2. naptól a kontroll közösség klorofill-tartalma, hasonlóan az
egyedszámban megfigyelt változásokhoz (7c. ábra) szignifikánsan magasabb volt mind a
tónál, mind a kezelt együttesekénél (8c. ábra). A PCE-vel kezelt együttes klorofill-tartalma
a kontroll kivételével minden közösségénél szignifikánsan magasabb volt (8c. ábra).
A 2012 évben mind a tavaszi, mind pedig a nyári kísérletek során összefüggést
lehetett kimutatni a közösségek klorofill-tartalom változása, valamint a tesztelt vegyületek
vízoldhatósága között: a leginkább vízoldható TeCa estében volt a legalacsonyabb a
közösség klorofill-tartalma, míg a legkevésbé vízoldható PCE-vel kezelt együttesé a
legmagasabb a kezelt közösségek közül.
22
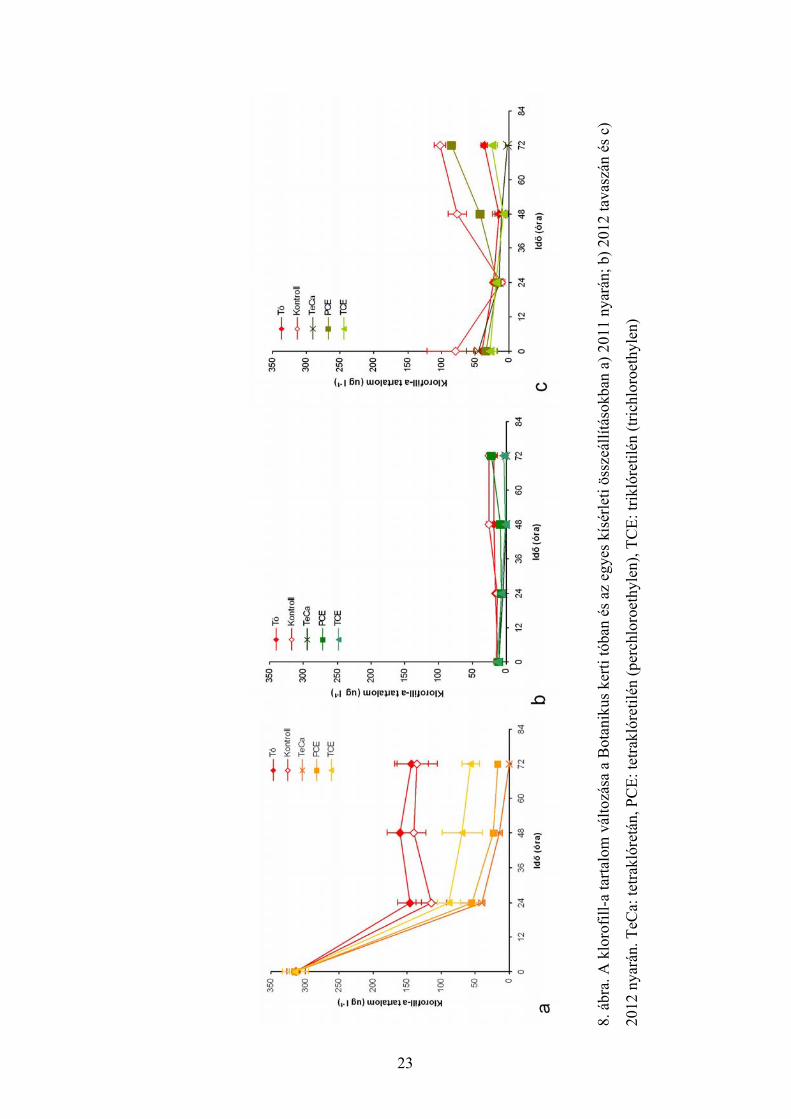
8. á
bra.
A k
loro
fill
-a ta
rtal
om v
álto
zása
2012
nya
rán.
TeC
a: te
trak
lóre
tán,
PC
E: t
etra
klór
etil
én (
per
a B
otan
ikus
ker
ti tó
ban
és a
z eg
yes
kísé
rlet
i öss
zeál
lítá
sokb
an a
) 20
11 n
yará
n; b
) 20
12 ta
vasz
án é
s c)
chlo
roet
hyle
n), T
CE
: tri
klór
etil
én (
tric
hlor
oeth
ylen
)
23

4.4. A taxonösszetételben megfigyelt változások
A Botanikus-kerti tóban 2011 nyarán egyértelmű Trachelomonas volvocinopsis
dominancia volt kimutatható (Euglenophyceae/Euglenales). A tóban a 0. napon 70%-ban, a
kísérleti edényekben pedig 70-80%-ban volt jelen (9a. ábra, függelék 1. táblázat). Ezen
kívül viszonylag nagy egyedszámmal voltak még jelen a 0. napon a tóban és az
edényekben is a Cyclotella fajok (C. meneghiniana, Cyclotella sp. -
Bacillariophyceae/Thalassiosirales), Cryptomonas fajok (C. ovata, Cryptomonas sp. -
Cryptophyceae/Cryptomonadales), különböző zöldalga fajok (Monoraphidium spp.,
Scenedesmus spp., Kirchneriella spp., Pedisatrum spp. - Chlorophyceae/Sphaeropleales),
valamint Chrysophyta-k (Kephyrion sp., Chrysococcus rufescens -
Chrysophyceae/Chromulinales; függelék 1. táblázat). Eredményeink azt mutatták, hogy a
klórozott szénhidrogénekkel szemben igen nagy az euglenoid T. volvocinopsis
rezisztenciája. Az első napra, feltehetően az első éjszakai viharnak köszönhetően, minden
közösségben megemelkedett a gyakoriságuk. Azonban míg a tóban és a kontroll
közösségben ezt követően gyakoriságuk folyamatosan csökkent, addig a kezelt
tenyészetekben gyakoriságuk ~90% volt. Megfigyelhető, hogy a kísérlet ideje alatt a
legnagyobb átlagos dominanciát (több, mint 92%) a leginkább vízoldható TeCa-val kezelt
közösségben, míg a „legalacsonyabbat” a legkevésbé vízoldható PCE-vel kezelt
együttesben (~88%) érték el. A Trachelomonas fajon kívül még egyes zöldalga fajok (pl.
Chlorella sp., Coelastrum microporum, Scenedesmus spp.), valamint a Cyclotella fajok
bizonyultak kevésbé érzékenynek a klórozott szénhidrogénekkel szemben (9a. ábra). Ezzel
szemben a Cryptomonas fajok, valamint a Chrysophyceae fajok érzékenyen reagáltak a
kezelésre: gyakoriságuk a szennyező vegyületek jelenlétében töredékére csökkent,
előfordult, hogy nem is tudtuk kimutatni jelenlétüket (9a. ábra). Itt kell megjegyezni, hogy
a legkevésbé vízoldható PCE-vel kezelt közösségekben csökkent legkisebb mértékben
ezen érzékeny taxonok gyakorisága (9a. ábra).
A 2012 tavaszán a 0. napon a Botanikus-kerti tóban, valamint a kísérleti
edényekben a Cryptomonas fajok (min. rel. gyak.: 23% - TeCa; max. rel. gyak.: 57% - tó),
valamint „Centrales” (nehezen határozható Stephanodiscus - Cyclostephanus fajok és
Cyclotella fajok, többek között C. meneghiniana) fajok (min. rel. gyak.: 8% - tó; max. rel.
gyak.: 44% - TeCa) voltak a legnagyobb gyakorisággal jelen (9b. ábra, függelék 2.
táblázat). Ezen taxonok mellett nagy számban fordultak elő különböző zöldalga fajok (pl.
Chlorella sp. - Trebouxiophyceae/Chlorellales; Scenedesmus spp., Monoraphidium spp. -
24

Chlorophyceae/Sphaeropleales; Chlamydomonas sp. -
Chlorophyceae/Chlamydomonadales), Trachelomonas fajok (Euglenophyceae/Euglenales)
és Chrysophyta fajok (pl. Chrysococcus rufescens, Kephyrion sp. -
Chrysophyceae/Chromulinales) (9b. ábra, függelék 2. táblázat). A kezelések hatására -
különösen a jobb vízoldhatóságú TeCa és TCE esetében - a Cryptomonas fajok, ahogy azt
már 2011 nyarán is megfigyelhettük, szinte teljesen eltűntek a közösségekből. Továbbá
összességében lecsökkent a „Centrales” kovaalgák száma; annak ellenére is, hogy a C.
meneghiniana (Bacillariophyceae/Thalassiosirales) egyedszáma jelentősen megnőtt
(Bacillariophyceae/Thalassiosirales) (9a. ábra). Vagyis míg a kisebb „Centrales” fajok
érzékenyebben reagáltak a szerves szennyezők jelenlétére, addig a C. meneghiniana ezen
kísérlet során sem bizonyult érzékenynek. A 2011 nyarán végzett kísérletekhez hasonlóan
itt is megnőtt a zöldalgák gyakorisága a kezelt közösségekben (9a-b. ábra). Viszont a
tavaszi kísérlet során nemcsak a Sphaeropleales rendbe tartozó taxonok (pl.
Monoraphidium spp., Scenedesmus spp.), hanem a Chlorellales és Chlamydomonadales
rendbe tartozó taxonok egyedszáma is jelentősen emelkedett. A 2011 nyarán
tapasztaltakkal ellentétben, ahol a Chrysophyceae fajok gyakorisága töredékére csökkent a
klórozott szénhidrogénekkel kezelt közösségekben (9a. ábra), 2012 tavaszán jelentősen
nőtt a gyakoriságuk a kezelt együttesekben. Feltehetően nem arról van szó, hogy a
Chrysophyceae fajok érzékenyek a klórozott szénhidrogének jelenlétére (ahogyan azt csak
a 2011-es adatok ismeretében feltételezhetnénk), hanem arról, hogy 2011 nyarán az alga
taxonok közti interakciós kapcsolatok miatt (pl. fény/tápanyag kompetíció, allelopátia)
nem tudtak nagy számban elszaporodni.
A 2012 nyarán végzett mikrokozmosz kísérletek során a 0. napon a következő
rendekbe tartozó fajok relatív gyakorisága volt a legnagyobb: Cryptomonadales,
Euglenales, Sphaeropleales, Achnanthales, Thalassiosirales, és Chlorellales. Emellett még
nagy számban voltak jelen korábban Pennales rendbe sorolt kovaalga fajok is (pl.
Fragilaria spp., Nitzschia spp., Gomphonema spp.; 9c. ábra, függelék 3. táblázat).
Hasonlóan az eddigi eredményekhez, 2012 nyarán is a kezelések hatására jelentősen
csökkent a Cryptomonas fajok gyakorisága (9c. ábra). Felhívnánk még a figyelmet arra,
hogy a Chrysophyceae fajok gyakorisága, hasonlóan a 2011 nyarán tapasztaltakhoz,
viszont eltérően a 2012 tavaszi eredményektől, itt is jelentősen lecsökkent a kezelések
során (9c. ábra). Ezzel szemben megnőtt a kovaalgák és egyes zöldalga taxonok aránya a
kezelt együttesekben (9c. ábra).
25

9.
ábra
. A
ta
xonö
ssze
téte
l
vált
ozás
a a
Bot
anik
us
kert
i
tóba
n és
az
eg
yes
kísé
rlet
i
össz
eáll
ítás
okba
n a)
20
11
nyar
án;
b) 2
012
tava
szán
és
c) 2
012
nyar
án.
Tó
0.,
1.,
2.
és
3.
nap;
K
0,
1,
2,
3:
kont
roll
0.,
1.,
2. é
s 3.
nap
;
TeC
a 0,
1,
2, 3
: te
trak
lóre
tán
0.,
1.,
2. é
s 3.
nap
; P
CE
0,
1,
2,
3:
tetr
akló
reti
lén
(per
chlo
ro-e
thyl
en)
0.,
1.,
2.
és 3
. na
p; T
CE
0,
1, 2
, 3:
trik
lóre
tilé
n (t
rich
loro
ethy
l-
en)
0., 1
., 2.
és
3. n
ap.
26

4.5. A fajszámban és diverzitásban bekövetkező változások
A 2011 nyarán végzett kísérletek során mind a tóban, mind a kontroll, mind pedig a
kezelt edényekben csökkent a fajszám az első két napban (10a. ábra). Azonban a fajszám-
csökkenés a kezelt, különösképpen pedig a tetraklór-származékokkal kezelt együttesekben
sokkal intenzívebb és nagyobb mértékű volt, mint a tóban, vagy a kontrollban (10a. ábra).
A különböző diverzitási mutatók alapján a kezelt együttesek diverzitása a tó együtteséhezz
képest már az első naptól kezdve, a kontroll közösségéhez képest pedig a második naptól
kezdve szignifikánsan csökkent (10a. ábra).
A 2012 tavaszi mikrokozmosz kísérletek során nem tapasztaltunk olyan mértékű
fajszám-csökkenést, mint 2011 nyarán (10a-b. ábra). A diverzitási mutatók alapján is azt
lehet mondani, hogy a tavaszi együttesek sokkal diverzebbek voltak mind a tóban, mind a
kezelt együttesekben, mint a megelőző év nyarán. Bár szignifikáns különbségek nem
voltak kimutathatók az együttesek diverzitási mutatói között, azt meg kell említeni, hogy a
TeCa-val kezelt közösség diverzebbé vált a közösség (10b. ábra). Amennyiben megnézzük
a közösség taxonösszetételét (9b. ábra), jól látható, hogy az érzékeny Cryptomonas fajok
eltűnésével más taxonok összegyedszáma nőtt meg (pl. Chromulinales rendbe tartozó
fajok, különböző kovaalga fajok, Trachelomonas spp. és zöldalga fajok) (9b. ábra).
A 2012 nyári kísérletek esetében volt a legdiverzebb a közösség összetétele mind a
tóban, mind a kontroll, mind pedig a kezelt edényekben (10c. ábra). A fajszám nem
csökkent olyan mértékben, mint azt az egy évvel korábban végzett vizsgálatok esetében
megfigyelhettük (10c ábra). A tavaszi vizsgálatokkal ellentétben ezen vizsgálatok során azt
tapasztaltuk, hogy a kontroll, vagyis kezeletlen edényben a közösség diverzitása már az
első napon szignifikánsan alacsonyabb volt, mint a tóban, vagy a kezelt együttesekben
(10c. ábra). A kezelt együttesek diverzitása pedig a 2. napon szignifikánsan magasabb volt
mind a kontrollénál, mind pedig a tó közösségénél. A 3. napon a kezelt együttesek
diverzitása szignifikánsan magasabb volt a kontrollénál, viszont a tóénál már csak a TCE-
vel kezelt együttesé volt magasabb (10c. ábra). Az, hogy a kezelt tenyészetek diverzebbek,
mint akár a tó, akár a kontroll közösségek, feltehetően annak köszönhető, hogy míg a tóban
a Cryptomonas fajok, a kontrollban pedig emellett még a Cyclotella fajok fordultak elő
nagy egyedszámmal, addig a kezelt tenyészetekben a Cryptomonas fajok száma
minimálisra csökkent, a Cyclotella fajok mellett pedig más kovaalga taxonok és zöldalga
taxonok is nagy számban jelentek meg (9c. ábra).
27

10. ábra. A fajszám és a diverzitás (H; d) változása a Botanikus kerti tóban és az egyes
kísérleti összeállításokban a) 2011 nyarán; b) 2012 tavaszán és c) 2012 nyarán. Tó 0., 1., 2.
és 3. nap; K 0, 1, 2, 3: kontroll 0., 1., 2. és 3. nap; TeCa 0, 1, 2, 3: tetraklóretán 0., 1., 2. és
3. nap; PCE 0, 1, 2, 3: tetraklóretilén (perchloro-ethylen) 0., 1., 2. és 3. nap; TCE 0, 1, 2, 3:
triklóretilén (trichloroethylen) 0., 1., 2. és 3. nap.
28

A fizikia-kémiai háttérváltozókkal kapcsolatos eredményeink azt mutatják, hogy az
algaközösségekben, azok faj-, és egyedszám összetételében végbement változások nem a
háttérváltozókban (pl.: T, pH, vezetőképesség) bekövetkezett változásoknak, hanem a
kezeléseknek köszönhetők.
A csaknem 70%-ban Trachelomonas volvocinopsis uralta 2011-es nyári közösségben
a fajösszetétel változatossága csökkent, mivel a domináns faj nem mutatkozott
érzékenynek a kezelésekre, és az érzékeny fajok eltűnésével a keletkezett „űrt” is kitöltötte
(feltehetően a kezeléssel szembeni ellenállóképessége mellett jó kompetíciós készséggel is
bírt a jelen lévő többi fajjal szemben). A Chrysophyták eltűnése tehát nem a kezelésre való
érzékenységnek köszönhető (lásd 2012 tavaszi vizsgálatok eredményei), hanem a
domináns fajjal szembeni gyengébb kompetíciós készségnek. A vizsgálatsorok
eredményeit összevetve elmondható, hogy a klórozott szénhidrogének hatása természetes
rendszerekben nagymértékben függ a fitoplankton együttesek összetételétől. A változatos
fajösszetételű 2012 tavaszi és nyári együttesekben a domináns, és egyben érzékeny
Cryptophyta fajok számának drasztikus csökkenése után a kevésbé érzékeny fajok aránya
megnőtt, továbbra is változatos fajeggyüttest létrehozva a kezelt összeállításokban is. A
három vizsgálatsor eredményeit összevetve elmondható, hogy a Cryptophyták rendkívül
érzékenyek a klórozott szénhidrogének jelenlétére, míg a zöldalgák közül a Sphaeropleales
rendbe tartozó fajok (Coelastrum, Monoraphidium, Pediastrum, Scenedesmus, Tetrastrum
genuszok) mérsékelt érzékenységet, a Chlorellales és Chlamydomonadales rendek fajai
pedig alacsony érzékenységet mutattak. Összefüggés látszik az egyes fajok érzékenysége
és valamilyen speciális sejtburok jelenléte között: az ún. lorika a Trachelomonas fajoknál,
a nyálkaanyagok a Chlorella fajok esetében, kolóniaképzés vagy vastag sejtfal jelenléte
jellemző az alacsonyabb érzékenységet mutató fajokra.
Mindhárom évben nagy számban jelentek meg bentikus fajok is a fitoplankton
mintákban. Ennek oka az, hogy a tó vízinövényzettel gazdagon benőtt, nyáron
gyakorlatilag alig látható szabad vízfelület. A 2010-es évben a tó mederátalakításon esett
át, 2011-ben így lényegesen alacsonyabb botrítottság jellemezte, mint a következő évben,
amely során intenzív vízinövény-telepítés is történt. Minden bizonnyal ennek köszönhető a
2012-es tavaszi és nyári minták lényegesen alacsonyabb klorofill-tartalma és alga-
egyedszáma is.
Jelen eredmények alátámasztani látszanak Yatchi és Loreau (1999) "biológiai
biztosítás hipotézis"-ét, hiszen a diverzebb (esetünkben 2012 tavaszi és nyári) rendszerek
változatossága a kezelés után is megmaradt. Ezekben feltehetően nagyobb arányban voltak
29

jelen olyan fajok, amelyek ellenálltak a zavarásnak és ez lehetővé tette, hogy
ellensúlyozzák a más fajok eltűnéséből származó funkcionális károsodást. Li és
munkatársai (2010) eredményeihez hasonlóan, mi is tapasztaltunk olyan esetet, amikor a
kezelt együttesekben magasabb volt a diverzitás, mint a kontrollban, vagy a tóban (pl.:
2012 nyarán, 10c ábra; ebben az esetben a TCE-vel kezelt együttesben szignifikánsan
magasabb volt a diverzitás a 3. napon, mint kontrollban, vagy a tóban). Azt azonban meg
kell jegyezni, hogy a Li és munkatársai által kimutatott nagyobb diverzitással
egyértelműen együtt járó nagyobb produktivitást nem sikerült kimutatnunk sem klorofill-
tartalom, sem össz-egyedszám alapján.
Eredményeink azt mutatják, hogy a fitoplankton együtteseket alkalmazó in situ
algatesztek, mikrokozmosz kísérletek jól alkalmazhatók a természetes közösségek toxikus
hatásra adott válaszainak vizsgálatára. Az adott kezelésre egyértelműen érzékeny, illetve
egyértelműen rezisztens fajok nagy pontossággal kiszűrhetők, az egyes fajok
érzékenységének megállapításán túl a mikrokozmosz kísérletek alkalmazása során
lehetőség nyílik a kompetíciós viszonyok tanulmányozására is. A „bizonytalan”
érzékenységű fajok esetében azonban a szennyezés pontos hatásának felmérésére a tiszta
tenyészetekkel végzett laboratóriumi toxikológiai tesztek sem nélkülözhetők.
30

5. Összefoglalás
A korábbi ipari tevékenység eredményeként a talajban, talajvízben felhalmozódott
kismolekulájú klórozott szénhidrogének problémája napjainkban újra a figyelem
középpontjába került, a legújabb vizsgálatok ugyanis kimutatták, hogy a talajvízmozgások
következtében a szennyezőanyagok felszíni vizekben való megjelenésének kockázata
megnőtt. A klórozott szénhidrogének fotoszintetizáló szervezetekre gyakorolt hatásai sem
ismeretlenek, hatásaikat tanulmányozták tengeri környezetben (Pearson és McConnell
1975; Ward et al. 1986), valamint édesvízi cianobaktériumok és eukarióta algák esetében
(Bringmann és Kühn 1980; Ando et al. 2003; Lukavsky et al., 2011; Bácsi et al. 2012).
Viszonylag alacsony azon tanulmányok száma, amelyek klórozott szénhidrogén
szennyezők természetes fitoplankton együttesekre gyakorolt hatását vizsgálták sekély tavi
ökoszisztémákban (Berglund et al. 2001; Gonzalez et al., 2009). Bár a biológiai sokféleség
hatása az ökoszisztéma működésére az ökológia egyik legkutatottabb területévé vált, a
diverzitás jelentősége a változó környezetben még kevéssé érthető.
Munkánk során in situ mikroalga teszttel (mikrokozmosz rendszerben) vizsgáltuk
egyszeri „pontszerű” klórozott szénhidrogén szennyezések rövidtávú hatását fitoplankton
együttesek összetételére. Vizsgálatainkat eltérő fajgazdagsággal jellemezhető, és
különböző szerkezetű fajegyütteseket tartalmazó kísérleti összeállításokban végeztük 2011
nyarán, illetve 2012 tavaszán és nyarán. A célkitűzések során feltett kérdésekre
eredményeink alapján a következő válaszok adhatók:
- A különböző fitoplankton közösségek diverzitása és produkciója között nem
tudtunk egyértelmű összefüggést kimutatni: a magasabb diverzitású 2012-es tavaszi
és nyári közösségek alacsonyabb egyedszámmal és alacsonyabb össz-klorofill-
tartalommal voltak jellemezhetők, ez azonban sokkal inkább a 2011-ben és 2012-
ben zajló folyamatos vízinövény-telepítésnek köszönhető, minthogy a
fajgazdagsággal lenne közvetlen összefüggésben.
- A három vizsgálatsor eredményeit összevetve elmondható, a diverzebb (esetünkben
2012 tavaszi és nyári) rendszerek változatossága a kezelés után is megmaradt, míg
a közel 70%-ban Trachelomonas volvocinopsis uralta 2011-es nyári közösségben a
fajösszetétel változatossága csökkent. Ez annak köszönhető, hogy a 2012-es
vizsgálatok alkalmával 40-60%-os gyakorisággal jelen lévő Cryptomonas fajok
rendkívül érzékenyen reagáltak a kezelésekre, csaknem teljesen eltűntek a kezelt
31

közösségekből. Ezzel szemben a 2011-ben domináns Trachelomonas
volvocinopsis, vélhetően jó kompetíciós készségének köszönhetően is, betöltötte az
érzékeny fajok visszaszorulásával keletkezett „űrt”.
Eredményeink azt mutatják, hogy az adott kezelésre egyértelműen érzékeny, illetve
egyértelműen rezisztens fajok nagy pontossággal kiszűrhetők mikrokozmosz kísérletek
alkalmazásával, a természetes fitoplankton együtteseket alkalmazó in situ algatesztek jól
alkalmazhatók a közösségek toxikus hatásra adott válaszainak vizsgálatára.
32

6. Köszönetnyilvánítás
Ezúton köszönöm meg és tiszteletemet fejezem ki mindazoknak, akik
diplomamunkám elkészítésében a segítségemre voltak.
Köszönetem fejezem ki témavezetőimnek, Dr. Bácsi István egyetemi adjunktusnak
a munkámban nyújtott támogatásáért és segítségéért.
Köszönöm Kovácsné Gábor Anikó vegyésztechnikus segítségét a laboratóriumi
mérésekben.
Köszönettel tartozom Édesanyámnak, aki munkám során mellettem állt és segített
munkám során.
A kutatás az Európai Unió és Magyarország támogatásával a TÁMOP-
4.2.4.A/2-11-1-2012-0001 azonosító számú „Nemzeti Kiválóság Program – Hazai
hallgatói, illetve kutatói személyi támogatást biztosító rendszer kidolgozása és
működtetése konvergencia program” című kiemelt projekt keretei között valósult
meg.
33

7. Irodalomjegyzék
ANDO T., OTSUKA S., NISHIYAMA M., SENOO K., WATANABE M.M. AND
MATSUMOTO S. 2003. Toxic effects of dichlomethane and trichloroethylene on the
growth of planktonic green algae, Chlorella vulgaris NIES227, Selenastrum
capricornutum NIES35, and Volvulina steinii NIES545. Microbes and Environments
8: 43–46.
ABRAHAMSSON K., EKDAHL A., COHEN J., ET AL. 1995. Marine algae-a source of
trichloroethylene and perchloroethylene. Limnology and Oceanography 40(7): 1321-
1326.
BÁCSI I., TÖRÖK T., B-BÉRES V., TÖRÖK P., TÓTHMÉRÉSZ B., NAGY S.A. AND
VASAS G. 2013. Laboratory and microcosm experiments testing the toxicity of
chlorinated hydrocarbons on a cyanobacterium strain (Synechococcus PCC 6301)
and on natural phytoplankton assemblages. Hydrobiologia, 710 (1): 189-203.
BARRIO-LAGE G., PARSONS F.Z., NASSAR R.S. 1987. Kinetics of the depletion of
trichloroethene. Environmental Science and Technology 21:366-370.
BERGLUND O., LARSSON P., EWALD G. AND OKLA L. 2001. Influence of trophic
status on PCB distribution in lake sediments and biota. Environmental Pollution 113:
199-210.
BHOWMICK M. AND SEMMENS M.J. 1994. Ultraviolet photooxidation for the
destruction of VOCs in air. Water Res 28:2407-2415.
BIGGS B.J.F., 2000. New Zealand Periphyton Guideline: Detecting, Monitoring and
Managing Enrichment of Streams. Ministry for the Environment, NZ, pp. 1–121.
BRINGMANN G. AND KÜHN R. 1980. Comparison of the toxicity thresholds of water
pollutants to bacteria, algae and protozoa in the cell multiplication inhibition test.
Water Research 14: 231–241.
BLANKENSHIP A., CHANG D.P.Y., JONES A.D., ET AL. 1994. Toxic combustion by-
products from the incineration of chlorinated hydrocarbons and plastics.
Chemosphere 28: 183-196.
BOGEN K.T., HALL L.C., PERRY L., FISHER, MCKONE T.E., DOWD P., PATTON
S.E., MALLON B. 1988. Health risk assessment of trichloroethylene (TCE) in
California drinking water. Livermore, CA: University of California, Lawrence
34

Livermore National Laboratory, Environmental Sciences Division. NTIS No. DE88-
005364.
BOWMAN J.P., JIMENEZ L., ROSARIO I., ET AL. 1993. Characterization of the
methanotrophic bacterial community present in a trichloroethylene-contaminated
subsurface groundwater site. Appl Environ Microbial 59:2380-2387.
CAMPBELL P.J., ARNOLD D.J.S., BROCK T.C.M., GRANDY N.J., HEGER W.,
HEIMBACH F., MAUND S.J., STRELOKE M. 1999. Guidance document on
higher-tier aquatic risk assessment for pesticides (HARAP). Report from the
SETAC-Europe/OECD/EC Workshop, 19–22 April 1998, Lacanau Oce´ an, France.
SETAC-Europe, Brussels.
CERVINI-SILVA J. 2003. Linear free-energy relationship analysis of the fate of
chlorinated 1- and 2-carbon compounds by redox-manipulated smectite clay
minerals. Environ Toxicol Chem 22:2298-2305.
CHAPIN F.S., WALKER B.H., HOBBS R.J., HOOPER D.U., LAWTON J.H., SALA
O.E. AND TILMAN, D. 1997. Science 277, 500–504.
CHAPPIE D.J. AND BURTON G.A. JR. 2000. Applications of aquatic and sediment
toxicity testing in situ. Soil Sed Cont 9: 219-245.
CHESSON P.L. AND CASE T.L. 1986. in Community Ecology, eds. Diamond, J.&Case,
T. J. (Harper&Row, New York), pp. 229–239.
CMR. 1986. Chemical Profile-Trichloroethylene. Chemical Marketing Reporter, January
27,1986.
COSTANZA R., D’ARGE R., DE GROOT R., FARBER S., GRASSO M., HANNON B.,
LIMBURG K., NAEEM S., O’NEILL R.V., PAUELO J., ET AL. 1997. Nature
(London) 387, 253–260.
CULP J.M., LOWELL R.B. AND CASH K.J. 2000. Integrating mesocosm experiments
with field and laboratory studies to generate weight-of-evidence risk assessments for
large rivers. Environmental Toxicology and Chemistry 19: 1167-1173.
COLACCI A., GRILLI S., LATTANZI G., ET AL. 1987. The covalent binding of 1,1,2,2-
tetrachloroethane to macromolecules of rat and mouse organs. Teratog Carcinog
Mutagen 7:465-474.
COSTA A.K., IVANETICH K.M. 1980. Tetrachloroethylene metabolism by the hepatic
microsomal cytochrome P-450 system. Biochem Pharmacol 29:2863-2869.
35

CULP J.M., LOWELL R.B. AND CASH K.J. 2000. Integrating mesocosm experiments
with field and laboratory studies to generate weight-of-evidence risk assessments for
large rivers. Environmental Toxicology and Chemistry 19: 1167-1173.
DEKANT W., METZLER M., HENSCHLER D. 1984. Novel metabolites of
trichloroethylene through dechlorination reactions in rats, mice and humans.
Biochem Pharmacol33:2021-2027.
DEKANT W., SCHULZ A., METZLER M., ET AL. 1986b. Absorption, elimination and
metabolism of trichloroethylene: A quantitative comparison between rats and mice.
Xenobiotica 16:143-152.
ENSLEY B.D. 1991. Biochemical diversity of trichloroethylene metabolism. Annu Rev
Microbial 45:283-299.
EHRLICH P.R. AND WILSON E.O. (1991) Science 253, 758–762.
FAN S., SCOW K.M. 1993. Biodegradation of trichloroethylene and toluene by indigenous
microbial populations in soil. Appl Environ Microbial 59:1911-1918.
FELFÖLDY L., 1987. A biológiai vízminősítés. Vízügyi Hidrobiológia 16. VGI,
Budapest, 258.
FRANKE C., STUDINGER G., BERGER G., ET AL. 1994. The assessment of
bioaccumulation. Chemosphere 29(7):1501-1514.
FRANKENBERRY M., KENT R., STROUP C., ET AL. 1987. Household products
containing methylene chloride and other chlorinated solvents: A shelf survey.
Rockville, MD: Westat, Inc. 4-l to 4-29.
FREEDMAN D.L., GOSETT J.M. 1989. Biological reductive dechlorination of
tetrachloroethylene and trichloroethylene to ethylene under methanogenic conditions.
Appl Environ Microbial 55:2144-2151.
GOODSELL P.J., UNDERWOOD A.J. 2008. Complexity and idiosyncrasy in the
responses of algae to disturbance in mono- and multi-species assemblages.
GONZÁLEZ J.F., FIGUEIRAS G., ARANGUREN-GASSIS M., CRESPO B.G.,
FERNÁDEZ E., MORÁN X.A.G. AND NIETO-CID M. 2009. Effect of a simulated
oil spill on natural assemblages of marine phytoplankton enclosed in microcosms.
Estuarine, Coastal and Shelf Science 83: 265–276.
GUASCH H., MUNOZ I., ROSÉS N. AND SABATER S. 1997. Changes in atrazine
toxicity throughout succession of stream periphyton communities. Journal of Applied
36

Phycology 9: 132–146.
GUIRY M.D. 2013. In: Guiry, M.D. & Guiry, G.M. AlgaeBase. World-wide electronic
publication, National University of Ireland, Galway. http://www.algaebase.org.
GURNEY S.E. AND ROBINSON G.C. 1989. The influence of two triazine herbicides on
the productivity, biomass and community composition of freshwater marsh
periphyton. Aquatic. Botany 36: 1–22.
GRANEY R.L., KENNEDY J.H., RODGERS J.H. 1994. Aquatic mesocosm studies in
ecological risk assessment. Lewis, Boca Raton, FL, USA.
HALPERT J. 1982. Cytochrome P-450 dependent covalent binding of 1,1,2,2-
tetrachloroethane in vitro. Drug Metab Dispos 10:465-468.
HALPERT J., NEAL R.A. 1981. Cytochrome P-450 metabolism of 1,1,2,2-
tetrachloroethane to dichloroacetic acid in vitro. Biochem Pharmacol 30:1366-1368.
HANLEY T.R., QUAST J.F., SCHUMANN A.M. 1988. The metabolism and hepatic
macromolecular interactions of 1,1,2,2-tetrachloroethane (TCE) in mice and rats.
Dow Chemical Company. Submitted to the U.S. Environmental Protection Agency
under TSCA Section 8D. OTS0514187.
HANSSON L.A. 1992. Factors regulating periphytic algal biomass. Limnol. Oceanogr. 37,
322–328.
HILLEBRAND H., SOMMER U. 2000. In: Biggs, B.J.F. (Ed.), New Zealand Periphyton
Guideline: Detecting, Monitoring and Managing Enrichment of Streams. Ministry for
the Environment, NZ, pp. 1–121.
HILL I.A., HEIMBACH F., LEEUWANGH P., MATTHIESEN P. (EDS) 1994.
Freshwater field tests for hazard assessment of chemicals. Lewis, Boca Raton, FL,
USA.
HSDB. 1996. Hazardous Substance Data Bank. National Library of Medicine, National
Toxicology Information Program, Bethesda, MD.
KINCANNON D.F., WEINERT A., PADORR R., ET AL. 1983. Predicting treatability of
multiple organic priority pollutant wastewater from single-pollutant treatability
studies. In: Bell MR, ed. Proceedings 37th Industrial Waste Conference. Ann Arbor,
MI: Ann Arbor Science, 641-650.
KOOI B.W., 2003. Numerical bifurcation analysis of ecosystems in a spatially
homogeneous environment. Acta Biotheor. 51, 189–222.
37

KUKONGVIRIYAPAN V., KUKONGVIRIYAPAN U., STACEY N.H. 1990. Interference
with hepatocellular substrate uptake by l,l,1-trichloroethane and tetrachloroethylene.
Toxicol Appl Pharmacol 102(1):80-90.
KUNEY J.H. 1986. Chemcyclopedia, Volume 5. Washington, DC: American Chemical
Society, 116.
KYRKLUND T., KJELLSTRAND P., HAGLID K.G. 1990. Long-term exposure of rats to
perchloroethylene, with and without a post-exposure solvent-free recovery period:
Effects on brain lipids. Toxicology Letters 52:279-285.
LAWLER S.P. 1993. Direct and indirect effects in microcosm communities of protists.
Oecologia 93, 184–190.
LEWIS R.J. 2001. Hawley’s condensed chemical dictionary. 14th ed. New York, NY: John
Wiley & Sons, Inc., 1079.
LI J.T. , DUAN H.N., LI S.P., KUANG J.L., ZENG Y., SHU W.S. 2010. Cadmium
pollution triggers a positive biodiversity-productivity relationship: evidence from a
laboratory microcosm experiment. Journal of Applied Ecology 47 (4): 890-898.
LIEBIG M., SCHMIDT G., BONTJE D., KOOI B.W., STRECK G., TRAUNSPURGER
W., KNACKER T. 2008. Direct and indirect effects of pollutants on algae and
algivorous ciliates in an aquatic indoor microcosm. Aquatic Toxicology 88: 102–
110.
LOPEZ-MANCISIDOR P., CARBONELL G., FERNÁNDEZ C., TARAZONA J.V. 2008.
Ecological impact of repeated applications of chlorpyrifos on zooplankton
community in mesocosms under Mediterranean conditions. Ecotoxicology 17: 811–
825.
LORAH M.M., OLSEN L.D. 1999. Degradation of 1,1,2,2-tetrachloroethane in a
freshwater tidal wetland: Fields and laboratory evidence. Environ Sci Technol
33:227-234.
LUKAVSKY J., FURNADZHIEVA S., DITTRT F. 2011. Toxicity of Trichloroethylene
(TCE) on Some Algae and Cyanobacteria. Bull Environ Contam Toxicol 86: 226–
231.
McCANN K. 2000. The diversity–stability debate. Nature 405: 228–233.
MILDE G., NENGER M., MENGLER R. 1988. Biological degradation of volatile
chlorinated hydrocarbons in groundwater. Meeting on Groundwater Microbiology:
38

Problems and Biological Treatment held at the International Association on Water
Pollution Research and Control Symposium, Kuopio, Finland, August 4-6, 1987.
Water Sci Technol 20:67-73.
MITOMA C., STEEGER T., JACKSON S.E., WHEELER K.P., ROGERS J.H., MILMAN
H.A. 1985. Metabolic disposition study of chlorinated hydrocarbons in rat and mice.
Drug Chem Toxicol 3:183-194.
MOREIRA-SANTOS M., DA SILVA E.M., SOARES A.M.V.M. AND RIBEIRO R.
2005. In Situ and Laboratory Microalgal Assays in the Tropics: A Microcosm
Simulation of Edge-of-Field Pesticide Runoff. Bulletin of Environmental
Contamination and Toxicology 74: 48–55.
MOREIRA-SANTOS M., SOARES A.M.V.M. AND RIBEIRO R. 2004. A phytoplankton
growth assay for routine in situ environmental assessments. Environmental
Toxicology and Chemistry 23: 1549-1560.
MUNOZ, I., REAL, M., GUASCH, H., NAVARRO, E., SABATER, S. 2001. Effects of
atrazine on periphyton under grazing pressure. Aquat. Toxicol. 55, 239–249.
NELSON M.J.K., MONTGOMERY S.O., MAHAFFEY W.R., ET AL. 1987.
Biodegradation of trichloroethylene and involvement of an aromatic biodegradative
pathway. Appl Environ Microbial 53:949-954.
O’CONNOR N.E., CROWE T.P. 2005. Biodiversity loss and ecosystem functioning:
distinguishing between number and identity of species. Ecology 86:1783–1796.
OECD 1984. Algal growth inhibition test. OECD guidelines for testing of chemicals 201.
Organization for Economic Cooperation and Developemnet, Paris.
PARSONS F., BARRIO-LAGE G., RICE R. 1985. Biotransformation of chlorinated
organic solvents in static microcosms. Environ Toxicol Chem 4:739-742.
PARSONS F., WOOD P.R., DEMARCO J. 1984. Transformation of tetrachloroethene and
trichloroethene in microcosms and groundwater. J Am Water Works Assoc 76:56-59.
PEARSON C.R., MCCONNELL G. 1975. Chlorinated Cl and C2 hydrocarbons in the
marine environment. Proc R Sot Lond [Biol] 189:305-332.
PESCE S., FAJON C., BARDOT C., BONNEMOY F., PORTELLI C. AND BOHATIER
J. 2006. Effects of the phenylurea herbicide diuron on natural riverine microbial
communities in an experimental study. Aquatic Toxicology 78: 303–314.
PFAFFENBERGER C.D., PEOPLES A.J., ENOS H.F. 1980. Distribution of volatile
39

halogenated organic compounds between rat blood serum and adipose tissue. Int J
Environ Anal Chem 8:55-65.
PUSCH, M., FIEBIG, D., BRETTAR, I., EISENMANN, H., ELLIS, B.K., KAPLAN,
L.A., LOCK, M.A., NAEGELI, M.W., TRAUNSPURGER, W. 1998. The role of
micro-organisms in the ecological connectivity of running waters. Freshw. Biol. 40,
453–495.
ROMANI A.M. AND SABATER S. 1999. Effect of primary producers on the
heterotrophic metabolism of a stream biofilm. Freshw. Biol. 41, 729–736.
ROSSBERG M., AKTIENGESELLSCHAFT H., LENDLE W., ET AL. 2005. Chlorinated
hydrocarbons. In: Ullman's encyclopedia of industrial chemistry. Weinheim,
Germany: Wiley-VCH Verlag, 1-184.
SABATER S., GREGORY S.V., SEDELL J.R. 1998. Community dynamics and
metabolism of benthic algae colonizing wood and rock substrata in forest stream. J.
Phycol. 34, 561–567.
SCHULZE E.-D. AND MOONEY H. A. (EDS.) 1993. Biodiversity and Ecosystem
Function (Springer, Berlin).
SITTIG M. 1985. Handbook of Toxic and Hazardous Chemicals and Carcinogens. Park
Ridge, New Jersey: Noyes Publications, 843-845.
SJOBERG B. 1952. Thermal decomposition of chlorinated hydrocarbons. Sven Kern
Tidskr 64:63-79.
SWANN R.L., LANKOWSKI D.A., MCCALL P.J., ET AL. 1983. A rapid method for the
estimation of the environmental parameters octanol, water partition coefficient, soil
sorption constant, water to air ratio, and water solubility. Res Rev 85:17-28.
TLILI A., DORIGO U., MONTUELLE B., MARGOUM C., CARLUER N., GOUY V.,
BOUCHEZ A. AND BÉRARD A. 2008. Responses of chronically contaminated
biofilms to short pulses of diuron. An experimental study simulating flooding events
in a small river. Aquatic Toxicology 87: 252–263.
TOMASI A., ALBANO E., BINI A., ET AL. 1984. Free radical intermediates under
hypoxic conditions in the metabolism of halogenated carcinogens. Toxicol Pathol
12:240-246.
TÓTHMÉRÉSZ B. 1997. Diverzitási rendezések. – Scientia Kiadó, Budapest.
40

TOUART L. 1988. Guidance document for aquatic mesocosm tests to support pesticide
registrations. EPA 540/09//88/035. Environmental Protection Agency, Washington,
DC.
USEPA 1994. Short-term methods for estimating the chronic toxicity of effluents and
receiving waters to freshwater organisms. EPA 600/7-91-002. Unites States
Environmental Protection Agency, Washington, DC.
UTERMÖHL H. 1958. Zur Vervollkommung der quantitativen Phytoplankton-Methodik.
Mitt. Int. Vr. Limnol. 9: 113-118
VERSCHUEREN K. 1983. Handbook of environmental data on organic chemicals. New
York: Van Nostrand Reinhold Co. 1131-l 135.
VILLENEUVE A., MONTUELLE B., BOUCHEZ A. 2010. Influence of slight differences
in environmental conditions (light, hydrodynamics) on the structure and function of
periphyton. Aquat. Sci. 72, 33–44.
VILLENEUVE A., MONTUELLE B. AND BOUCHEZ A. 2011. Effects of flow regime
and pesticides on periphytic communities: Evolution and role of biodiversity.
Aquatic Toxicology 102: 123–133.
VITOUSEK P.M., H. A. MOONEY, J. LUBCHENCO, AND J. M. MELILLO. 1997.
Human domination of Earth's ecosystems. Science 277:494-499.
WARD G.S., TOLMSOFF A.J. AND PETROCELL S.R. 1986. Acute toxicity of
trichloroethylene to saltwater organisms. Bulletin of Environmental Contamination
and Toxicology 37: 830–836.
WILSON B.H., SMITH G.B., REES J.F. 1986. Biotransformations of selected
alkylbenzenes and halogenated aliphatic hydrocarbons in methanogenic aquifer
material: A microcosm study. Environmental Science and Technology 20:997-1002.
WILSON J.T., WILSON B.H. 1985. Biotransformation of trichloroethylene in soil. Appl
Environ Microbial 49~242-243.
WOODRUFF S.L., HOUSE W.A., CALLOW M.E., LEADBEATER B.S.C. 1999. The
effects of biofilms on chemical processes in surficial sediments. Freshw. Biol. 41,
73– 89.
YAGI O., UCHIYAMA H., IWASAKI K. 1992. Biodegradation rate of chloroethylene in
soil environment. Water Science and Technology 25:419-424.
YATCHI S. AND LOREAU M. 1999. Biodiversity and ecosystem productivity in a
41

fluctuating environment: the insurance hypothesis. Proceedings of the National
Academy of Sciences U.S.A. 96: 1463–1468.
YLLNER S. 1971. Metabolism of 1,1,2,2-tetrachloroethane-14C in the mouse. Acta
Pharmacol Toxicol 29:499-512.
42

8. Függelék
Függelék 1. táblázat. A 2011 nyarán zajlott kísérletek során kimutatott fajok és
rendszertani besorolásuk (a színek a 9a. ábra jelmagyarázatában látható színkódolással
egyezőek).
Kimutatott taxon Rendszertani besorolás Chrysococcus rufescens Ochrophyta - Chrysophyceae/Chromulinales Chrysococcus sp. Ochrophyta - Chrysophyceae/Chromulinales Dinobyron sp. Ochrophyta - Chrysophyceae/Chromulinales Kephyrion sp. Ochrophyta - Chrysophyceae/Chromulinales CCyyccllootteellllaa mmeenneegghhiinniiaannaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioossiirraalleess CCyyccllootteellllaa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioossiirraalleess EEppiitthheemmiiaa aaddnnaattaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//RRhhooppaallooddiiaalleess EEppiitthheemmiiaa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//RRhhooppaallooddiiaalleess Navicula radiosa Ochrophyta - Bacillariophyceae/Naviculales Navicula sp. Ochrophyta - Bacillariophyceae/Naviculales Melosira varians Ochrophyta - Bacillariophyceae/Melosirales Asterionella formosa Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria acus Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria capucina Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria sp. Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria ulna acus Ochrophyta - Bacillariophyceae/Fragilariales Cymbella affinis Ochrophyta - Bacillariophyceae/Cymbellales Cymbella sp. Ochrophyta - Bacillariophyceae/Cymbellales Didymosphenia geminata Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema acuminatum Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema parvulum Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema sp. Ochrophyta - Bacillariophyceae/Cymbellales Encyonema silesiacum Ochrophyta - Bacillariophyceae/Cymbellales Rhoicosphenia abbreviata Ochrophyta - Bacillariophyceae/Cymbellales Nitzschia acicularis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia linearis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia sp. Ochrophyta - Bacillariophyceae/Bacillariales Aulacoseira distans Ochrophyta - Bacillariophyceae/Aulacoseirales Coccoenis placentura Ochrophyta - Bacillariophyceae/Achnanthales Achantales sp. Ochrophyta - Bacillariophyceae/Achnanthales Lepocinclis sp. Euglenophyta - Euglenophyceae/Euglenales Trachelomonas globularis Euglenophyta - Euglenophyceae/Euglenales Trachelomonas rugulosa Euglenophyta - Euglenophyceae/Euglenales Trachelomonas volvocina Euglenophyta - Euglenophyceae/Euglenales Trachelomonas volvocinopsis Euglenophyta - Euglenophyceae/Euglenales Limnothrix sp. Cyanobacteria - Cyanophyceae/Oscillatoriales Cryptomonas ovata Cryptophyta - Cryptophyceae/Cryptomonadales Cryptomonas sp. Cryptophyta - Cryptophyceae/Cryptomonadales Chlorella sp. Chlorophyta - Trebouxiophyceae/Chlorellales Oocystis sp. Chlorophyta - Trebouxiophyceae/Chlorellales Acutodesmus acuminatus Chlorophyta - Chlorophyceae/Sphaeropleales Coelastrum microporum Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus armatus Chlorophyta - Chlorophyceae/Sphaeropleales
43

Desmodesmus communis Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus intermedius Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus spinosus Chlorophyta - Chlorophyceae/Sphaeropleales Kirchneriella obesa Chlorophyta - Chlorophyceae/Sphaeropleales Kirchneriella sp. Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium circinale Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium contortum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium griffithii Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium pusillum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium tortile Chlorophyta - Chlorophyceae/Sphaeropleales Pediastrum duplex Chlorophyta - Chlorophyceae/Sphaeropleales Pseudopediastrum boryanum Chlorophyta - Chlorophyceae/Sphaeropleales Tetraedron caudatum Chlorophyta - Chlorophyceae/Sphaeropleales Tetrastrum staurogeniaeforme Chlorophyta - Chlorophyceae/Sphaeropleales Sphaerellopsis sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Chlamydomonas sp. Chlorophyta - Chlorophyceae/Chlamydomonadales
44

Függelék 2. táblázat. A 2012 tavaszán zajlott kísérletek során kimutatott fajok és
rendszertani besorolásuk (a színek a 9b. ábra jelmagyarázatában látható színkódolással
egyezőek).
Kimutatott taxon Rendszertani besorolás Synura sp. Ochrophyta - Synurophyceae/Synurales Mallomonas sp. Ochrophyta - Chrysophyceae/Synurales Kephyrion sp. Ochrophyta - Chrysophyceae/Chromulinales Chrysococcus rufescens Ochrophyta - Chrysophyceae/Chromulinales Dinobyron sp. Ochrophyta - Chrysophyceae/Chromulinales CCyyccllootteellllaa mmeenneegghhiinniiaannaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioossiirraalleess AAmmpphhoorraa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioopphhyyssaalleess Cymatopleura solea Ochrophyta - Bacillariophyceae/Surirellales Surirella sp. Ochrophyta - Bacillariophyceae/Surirellales Entomoneis sp. Ochrophyta - Bacillariophyceae/Surirellales Epithemia adnata Ochrophyta - Bacillariophyceae/Rhopalodiales Amphipleura pellucida Ochrophyta - Bacillariophyceae/Naviculales Gyrosigma acuminatum Ochrophyta - Bacillariophyceae/Naviculales Hippodonta capitata Ochrophyta - Bacillariophyceae/Naviculales Navicula sp. Ochrophyta - Bacillariophyceae/Naviculales Pinnularia microstauron Ochrophyta - Bacillariophyceae/Naviculales Fragilaria capucina Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria ulna acus Ochrophyta - Bacillariophyceae/Fragilariales Ulnaria capitata Ochrophyta - Bacillariophyceae/Fragilariales Ulnaria ulna Ochrophyta - Bacillariophyceae/Fragilariales Cymbella sp. Ochrophyta - Bacillariophyceae/Cymbellales Didymosphenia geminata Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema parvulum Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema sp. Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema truncatum Ochrophyta - Bacillariophyceae/Cymbellales Rhoicosphenia abbreviata Ochrophyta - Bacillariophyceae/Cymbellales Nitzschia acicularis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia linearis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia reversa Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia sp. Ochrophyta - Bacillariophyceae/Bacillariales Coccoenis placentura Ochrophyta - Bacillariophyceae/Achnanthales Achantales sp. Ochrophyta - Bacillariophyceae/Achnanthales Achanthidium exiguum Ochrophyta - Bacillariophyceae/Achnanthales Achanthidium minutissimum Ochrophyta - Bacillariophyceae/Achnanthales "Centrales" Ochrophyta - Bacillariophyceae/"Centrales" Gymnodium sp. Myzozoa - Dinophyceae/Gymnodiniales Peridinium cinctum Myzozoa - Dinophyceae/Peridiniales Euglena sp. Euglenozoa - Euglenophyceae/Euglenales Lepocinclis sp. Euglenozoa - Euglenophyceae/Euglenales Trachelomonas bernardiensis bernardinis Euglenozoa - Euglenophyceae/Euglenales Trachelomonas oblonga Euglenozoa - Euglenophyceae/Euglenales Trachelomonas playfairi playfairi Euglenozoa - Euglenophyceae/Euglenales Trachelomonas rugulosa Euglenozoa - Euglenophyceae/Euglenales Trachelomonas sp. Euglenozoa - Euglenophyceae/Euglenales Trachelomonas verrucosa Euglenozoa - Euglenophyceae/Euglenales
45

Trachelomonas volvocina Euglenozoa - Euglenophyceae/Euglenales Trachelomonas volvocinopsis Euglenozoa - Euglenophyceae/Euglenales Phacus sp. Euglenozoa - Euglenophyceae/Euglanales Cyanogranis ferruginea Cyanobacteria - Cyanophyceae/Synechococcales Cyanogranis libera Cyanobacteria - Cyanophyceae/Synechococcales Phormidium sp. Cyanobacteria - Cyanophyceae/Oscillatoriales Nostocales Cyanobacteria - Cyanophyceae/Nostocales Gloeocapsa sp. Cyanobacteria - Cyanophyceae/Chroococcales Cryptomonas curvata Cryptophyta - Cryptophyceae/Cryptomonadales Cryptomonas marssonii Cryptophyta - Cryptophyceae/Cryptomonadales Cryptomonas sp. Cryptophyta - Cryptophyceae/Cryptomonadales FFoonnaallaass zzööllddaallggaa CChhlloorroopphhyyttaa Koliella longistea Chlorophyta - Trebouxiophyceae/Prasiolales Crucigenia tetrapedia Chlorophyta - Trebouxiophyceae/Trebouxiophyceae
ordo incertae sedis Actinastrum hantzschii Chlorophyta - Trebouxiophyceae/Chlorellales Chlorella sp. Chlorophyta - Trebouxiophyceae/Chlorellales Crucigeniella crucifera Chlorophyta - Trebouxiophyceae/Chlorellales Dictyosphaerium pulchellum Chlorophyta - Trebouxiophyceae/Chlorellales Didymocystis inermis Chlorophyta - Trebouxiophyceae/Chlorellales Oocystis lacustris Chlorophyta - Trebouxiophyceae/Chlorellales Oocystis sp. Chlorophyta - Trebouxiophyceae/Chlorellales Acutodesmus obliquus Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium tortile Chlorophyta - Chlorophyceae/Sphaeropleales Acutodesmus acuminatus Chlorophyta - Chlorophyceae/Sphaeropleales Coelastrum microporum Chlorophyta - Chlorophyceae/Sphaeropleales Coelastrum pseudomicroporum Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus armatus Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus intermedius Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus opoliensis var. carinatus Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus communis Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus spinosus Chlorophyta - Chlorophyceae/Sphaeropleales Kirchneriella irregularis Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium arcuatum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium circinale Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium contortum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium dybowskii Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium griffithii Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium komarkovae Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium minutum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium pusillum Chlorophyta - Chlorophyceae/Sphaeropleales Pediastrum duplex Chlorophyta - Chlorophyceae/Sphaeropleales Pseudodidymocystis inconspicua Chlorophyta - Chlorophyceae/Sphaeropleales Rraphidocelis rotunda Chlorophyta - Chlorophyceae/Sphaeropleales Scenedezmus dactylococcopsis Chlorophyta - Chlorophyceae/Sphaeropleales Scenedesmus sp. Chlorophyta - Chlorophyceae/Sphaeropleales Schroederia setigera Chlorophyta - Chlorophyceae/Sphaeropleales Tetrastrum staurogeniaeforme Chlorophyta - Chlorophyceae/Sphaeropleales Tetrastrum triangulare Chlorophyta - Chlorophyceae/Sphaeropleales Phacotus sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Sphaerellopsis sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Chlamydomonas sp. Chlorophyta - Chlorophyceae/Chlamydomonadales
46

Chlorogonium sp. Chlorophyta - Chlorophyceae/Chlamydomonadales CCoossmmaarriiuumm sspp.. CChhaarroopphhyyttaa -- ZZyyggnneemmoopphhyycceeaaee//DDeessmmiiddiiaalleess
47
48
Függelék 3. táblázat. A 2012 nyarán zajlott kísérletek során kimutatott fajok és
rendszertani besorolásuk (a színek a 9c. ábra jelmagyarázatában látható színkódolással
egyezőek).
Kimutatott taxon Rendszertani besorolás GGoonniioocchhlloorriiss mmuuttiiccaa OOcchhrroopphhyyttaa -- XXaannttoopphhyycceeaaee//MMiisscchhooccooccccaalleess Synura sp. Ochrophyta - Synurophyceae/Synurales Mallomonas sp. Ochrophyta - Chrysophyceae/Synurales Kephyrion sp. Ochrophyta - Chrysophyceae/Chromulinales Chrysococcus rufescens Ochrophyta - Chrysophyceae/Chromulinales Dinobyron sp. Ochrophyta - Chrysophyceae/Chromulinales CCyyccllootteellllaa mmeenneegghhiinniiaannaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioossiirraalleess CCyyccllootteellllaa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioossiirraalleess AAmmpphhoorraa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//TThhaallaassssiioopphhyyssaalleess CCyymmaattoopplleeuurraa ssoolleeaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//SSuurriirreellllaalleess SSuurriirreellllaa sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//SSuurriirreellllaalleess EEnnttoommoonneeiiss sspp.. OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//SSuurriirreellllaalleess EEppiitthheemmiiaa aaddnnaattaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//RRhhooppaallooddiiaalleess RRhhooppaallooddiiaa ggiibbbbaa OOcchhrroopphhyyttaa -- BBaacciillllaarriioopphhyycceeaaee//RRhhooppaallooddiiaalleess Amphipleura pellucida Ochrophyta - Bacillariophyceae/Naviculales Carticula cuspidata Ochrophyta - Bacillariophyceae/Naviculales Gyrosigma acuminatum Ochrophyta - Bacillariophyceae/Naviculales Gyrosigma attenuatum Ochrophyta - Bacillariophyceae/Naviculales Hippodonta capitata Ochrophyta - Bacillariophyceae/Naviculales Navicula sp. Ochrophyta - Bacillariophyceae/Naviculales Pinnularia microstauron Ochrophyta - Bacillariophyceae/Naviculales Melosira varians Ochrophyta - Bacillariophyceae/Melosirales Asterionella formosa Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria capucina Ochrophyta - Bacillariophyceae/Fragilariales Fragilaria ulna acus Ochrophyta - Bacillariophyceae/Fragilariales Ulnaria capitata Ochrophyta - Bacillariophyceae/Fragilariales Ulnaria ulna Ochrophyta - Bacillariophyceae/Fragilariales Cymbella sp. Ochrophyta - Bacillariophyceae/Cymbellales Didymosphenia geminata Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema acuminatum Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema augur Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema brebissonii Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema parvulum Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema sp. Ochrophyta - Bacillariophyceae/Cymbellales Gomphonema truncatum Ochrophyta - Bacillariophyceae/Cymbellales Rhoicosphenia abbreviata Ochrophyta - Bacillariophyceae/Cymbellales Nitzschia acicularis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia linearis Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia reversa Ochrophyta - Bacillariophyceae/Bacillariales Nitzschia sp. Ochrophyta - Bacillariophyceae/Bacillariales Aulacoseira granulata Ochrophyta - Bacillariophyceae/Aulacoseirales Lemnicola hungarica Ochrophyta - Bacillariophyceae/Aulacoseirales Coccoenis placentura Ochrophyta - Bacillariophyceae/Achnanthales Achantales sp. Ochrophyta - Bacillariophyceae/Achnanthales Achanthidium exiguum Ochrophyta - Bacillariophyceae/Achnanthales

Achanthidium minutissimum Ochrophyta - Bacillariophyceae/Achnanthales "Centrales" Ochrophyta - Bacillariophyceae/"Centrales" Gymnodium sp. Myzozoa - Dinophyceae/Gymnodiniales Peridinium cinctum Myzozoa - Dinophyceae/Peridiniales Euglena sp. Euglenozoa - Euglenophyceae/Euglenales Lepocinclis sp. Euglenozoa - Euglenophyceae/Euglenales Trachelomonas bernardiensis bernardinis Euglenozoa - Euglenophyceae/Euglenales Trachelomonas oblonga Euglenozoa - Euglenophyceae/Euglenales Trachelomonas playfairi playfairi Euglenozoa - Euglenophyceae/Euglenales Trachelomonas rugulosa Euglenozoa - Euglenophyceae/Euglenales Trachelomonas scabra Euglenozoa - Euglenophyceae/Euglenales Trachelomonas sp. Euglenozoa - Euglenophyceae/Euglenales Trachelomonas verrucosa Euglenozoa - Euglenophyceae/Euglenales Trachelomonas volvocina Euglenozoa - Euglenophyceae/Euglenales Trachelomonas volvocinopsis Euglenozoa - Euglenophyceae/Euglenales Phacus sp. Euglenozoa - Euglenophyceae/Euglanales Cyanogranis ferruginea Cyanobacteria - Cyanophyceae/Synechococcales Cyanogranis libera Cyanobacteria - Cyanophyceae/Synechococcales Phormidium sp. Cyanobacteria - Cyanophyceae/Oscillatoriales Oscillatoria sp. Cyanobacteria - Cyanophyceae/Oscillatoriales Nostocales Cyanobacteria - Cyanophyceae/Nostocales GGllooeeooccaappssaa sspp.. CCyyaannoobbaacctteerriiaa -- CCyyaannoopphhyycceeaaee//CChhrrooooccooccccaalleess Plagioselmis lacustris Cryptophyta - Cryptophyceae/Pyrenomonadales Cryptomonas curvata Cryptophyta - Cryptophyceae/Cryptomonadales Cryptomonas marssonii Cryptophyta - Cryptophyceae/Cryptomonadales Cryptomonas sp. Cryptophyta - Cryptophyceae/Cryptomonadales FFoonnaallaass zzööllddaallggaa CChhlloorroopphhyyttaa Koliella longistea Chlorophyta - Trebouxiophyceae/Prasiolales Crucigenia tetrapedia Chlorophyta - Trebouxiophyceae/Trebouxiophyceae
ordo incertae sedis Actinastrum hantzschii Chlorophyta - Trebouxiophyceae/Chlorellales Chlorella sp. Chlorophyta - Trebouxiophyceae/Chlorellales Closteriopsis acicularis Chlorophyta - Trebouxiophyceae/Chlorellales Crucigeniella crucifera Chlorophyta - Trebouxiophyceae/Chlorellales Dictyosphaerium pulchellum Chlorophyta - Trebouxiophyceae/Chlorellales Didymocystis inermis Chlorophyta - Trebouxiophyceae/Chlorellales Lagerheimina chodatii Chlorophyta - Trebouxiophyceae/Chlorellales Oocystis lacustris Chlorophyta - Trebouxiophyceae/Chlorellales Oocystis sp. Chlorophyta - Trebouxiophyceae/Chlorellales VVoollvvooxx sspp.. CChhlloorroopphhyyttaa -- CChhlloorroopphhyycceeaaee//VVoollvvooccaalleess Acutodesmus obliquus Chlorophyta - Chlorophyceae/Sphaeropleales Acutodesmus acuminatus Chlorophyta - Chlorophyceae/Sphaeropleales Characium sp. Chlorophyta - Chlorophyceae/Sphaeropleales Coelastrum microporum Chlorophyta - Chlorophyceae/Sphaeropleales Coelastrum pseudomicroporum Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus armatus Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus intermedius Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus opoliensis var. carinatus Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus communis Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus spinosus Chlorophyta - Chlorophyceae/Sphaeropleales Desmodesmus subspicatus Chlorophyta - Chlorophyceae/Sphaeropleales Enallax acutiformis Chlorophyta - Chlorophyceae/Sphaeropleales
49

50
Kirchneriella arcuata Chlorophyta - Chlorophyceae/Sphaeropleales Kirchneriella irregularis Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium arcuatum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium circinale Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium contortum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium dybowskii Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium griffithii Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium komarkovae Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium minutum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium pusillum Chlorophyta - Chlorophyceae/Sphaeropleales Monoraphidium tortile Chlorophyta - Chlorophyceae/Sphaeropleales Pediastrum biradiatum Chlorophyta - Chlorophyceae/Sphaeropleales Pediastrum duplex Chlorophyta - Chlorophyceae/Sphaeropleales Pseudodidymocystis inconspicua Chlorophyta - Chlorophyceae/Sphaeropleales Pseudopediastrum borianum Chlorophyta - Chlorophyceae/Sphaeropleales Rraphidocelis rotunda Chlorophyta - Chlorophyceae/Sphaeropleales Scenedesmus dactylococcopsis Chlorophyta - Chlorophyceae/Sphaeropleales Scenedesmus sp. Chlorophyta - Chlorophyceae/Sphaeropleales Schroederia setigera Chlorophyta - Chlorophyceae/Sphaeropleales Stauridium tetras Chlorophyta - Chlorophyceae/Sphaeropleales Tetraedron minimum Chlorophyta - Chlorophyceae/Sphaeropleales Tetraedron regulare Chlorophyta - Chlorophyceae/Sphaeropleales Tetraedron triangulare Chlorophyta - Chlorophyceae/Sphaeropleales Tetrastrum staurogeniaeforme Chlorophyta - Chlorophyceae/Sphaeropleales Tetrastrum triangulare Chlorophyta - Chlorophyceae/Sphaeropleales Phacotus sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Sphaerellopsis sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Eudorina sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Chlamydomonas sp. Chlorophyta - Chlorophyceae/Chlamydomonadales Chlorogonium sp. Chlorophyta - Chlorophyceae/Chlamydomonadales CCoossmmaarriiuumm sspp.. CChhaarroopphhyyttaa -- CCoonnjjuuggaattoopphhyycceeaaee//DDeessmmiiddiiaalleess CClloosstteerriiuumm sspp.. CChhaarroopphhyyttaa -- CCoonnjjuuggaattoopphhyycceeaaee//DDeessmmiiddiiaalleess CClloosstteerriiuumm eehhrreennbbeerrggiiii CChhaarroopphhyyttaa -- CCoonnjjuuggaattoopphhyycceeaaee//DDeessmmiiddiiaalleess CClloosstteerriiuumm aacciiccuullaarriiss CChhaarroopphhyyttaa -- CCoonnjjuuggaattoopphhyycceeaaee//DDeessmmiiddiiaalleess SSppiirrooggyyrraa sspp.. CChhaarroopphhyyttaa -- CCoonnjjuuggaattoopphhyycceeaaee//ZZyyggnneemmaattaalleess