diabetes impairs stem cell and proangiogenic cell ...diabetes impairs stem cell and proangiogenic...
TRANSCRIPT

Diabetes Impairs Stem Cell andProangiogenic Cell Mobilizationin HumansGIAN PAOLO FADINI, MD
MATTIA ALBIERO, PHD
SAULA VIGILI DE KREUTZENBERG, MD, PHD
ELISA BOSCARO, BSC
ROBERTA CAPPELLARI, BSC
MARIACRISTINA MARESCOTTI, BSC
NICOL PONCINA, BSC
CARLO AGOSTINI, MD
ANGELO AVOGARO, MD, PHD
OBJECTIVEdDiabetes mellitus (DM) increases cardiovascular risk, at least in part, throughshortage of vascular regenerative cells derived from the bone marrow (BM). In experimentalmodels, DM causes morphological and functional BM alterations, but information on BM func-tion in human DM ismissing. Herein, we sought to assaymobilization of stem and proangiogeniccells in subjects with and without DM.
RESEARCH DESIGN AND METHODSdIn a prospective trial (NCT01102699), wetested BM responsiveness to 5mg/kg human recombinant granulocyte colony–stimulating factor(hrG-CSF) in 24 individuals with DM (10 type 1 and 14 type 2) and 14 individuals without DM.Before and 24 h after hrG-CSF, we quantified circulating stem/progenitor cells and total anddifferential white blood cell counts. We also evaluated in vivo the proangiogenic capacity ofperipheral blood mononuclear cells using the Matrigel plug assay.
RESULTSdIn response to hrG-CSF, levels of CD34+ cells and other progenitor cell pheno-types increased in subjects without DM. Patients with DM had significantly impaired mobiliza-tion of CD34+, CD133+, and CD34+CD133+ hematopoietic stem cells and CD133+KDR+
endothelial progenitors, independently of potential confounders. The in vivo angiogenic capac-ity of peripheral blood mononuclear cells significantly increased after hrG-CSF in control sub-jects without DM, but not in patients with DM. DM was also associated with the inability toupregulate CD26/DPP-4 on CD34+ cells, which is required for the mobilizing effect of granulo-cyte colony–stimulating factor.
CONCLUSIONSdStem and proangiogenic cell mobilization in response to hrG-CSF is im-paired in DM, possibly because of maladaptive CD26/DPP-4 regulation. These alterations mayhamper tissue repair and favor the development of cardiovascular complications.
Diabetes Care 36:943–949, 2013
D iabetes mellitus (DM) increases car-diovascular disease, and this is at-tributed, at least in part, to shortage
of vascular regenerative cells derived fromthe bone marrow (BM) (1). DM is associ-ated with reduced levels of several circu-lating progenitor cell phenotypes (2). Wehave previously shown that DM preventspostischemic progenitor cell mobilizationin rats, which translates into impairedvascular recovery after ischemia (3).
Recent data from experimental modelsof type 1 DM and type 2 DM highlight BMpathologies that includemicroangiopathy(4), neuropathy (5), altered gene expres-sion (6), and niche dysfunction (7). Thesechanges may account for an impairedmo-bilizing capacity in DM compared withcontrol animals (8). Data on BM functionin human DM are scant, whereas there isno information on BM structure. In a retro-spective case series of patients undergoing
BM autotransplantation, DM was statisti-cally associated with poor mobilization inresponse to chemotherapy plus human re-combinant granulocyte colony–stimulatingfactor (hrG-CSF) (7). Moreover, in supportof the existence of a BM defect in humanDM, we have shown a reduction in BMCD34+ cells, compared with nondiabeticsubjects (9).
The mechanism of action of the mobi-lizing factor granulocyte colony–stimulatingfactor (G-CSF) is complex and involvescleavage of stromal-derived factor (SDF)-1a through release of proteases, elastases,and matrix metalloprotease-9, suppressionof osteoblastic function, and modulation ofintegrins (10). The mechanism wherebyDM impairs stem cell mobilization may de-pend on altered local concentrations of thechemokine SDF-1a. It is noteworthy thatSDF-1a is a natural substrate of the proteaseCD26/DPP-4, the activity of which is dys-regulated in DM (11). The impaired stemcell mobilization in DM has important im-plications for the care of patients in the he-matology clinic. Furthermore, because theBM harbors a variety of regenerative non-hematopoietic progenitors, including en-dothelial progenitor cells (EPCs), BMdysfunction may contribute to the onsetof chronic DM complications (12). Unfor-tunately, exploration of BM structure andfunction in humans is limited by the intrin-sic low availability of BM samples fromnon-hematologic patients. Therefore, to confirmthe diabetic stem cell “mobilopathy” in hu-mans, we devised a pharmacologic test ofBM reserve in a prospective trial of BM stim-ulationwith a single subcutaneous injectionof hrG-CSF in individuals with DM andwithout DM.
RESEARCH DESIGN ANDMETHODS
Patients and treatmentThe study was approved by the localethics committee and is registered inClinicalTrials.gov (NCT01102699). Thiswas a prospective, parallel group study ofdirect BM stimulation with hrG-CSF insubjectswith andwithoutDM.Theprimaryend point was change in circulating
c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c c
From the Department of Medicine, University of Padova, Padova, Italy, and Venetian Institute of MolecularMedicine, Padova, Italy.
Corresponding author: Gian Paolo Fadini, [email protected] or [email protected] 7 June 2012 and accepted 6 September 2012.DOI: 10.2337/dc12-1084. Clinical trial reg. no. NCT01102699, clinicaltrials.gov.This article contains Supplementary Data online at http://care.diabetesjournals.org/lookup/suppl/doi:10
.2337/dc12-1084/-/DC1.© 2013 by the American Diabetes Association. Readers may use this article as long as the work is properly
cited, the use is educational and not for profit, and thework is not altered. See http://creativecommons.org/licenses/by-nc-nd/3.0/ for details.
care.diabetesjournals.org DIABETES CARE, VOLUME 36, APRIL 2013 943
P a t h o p h y s i o l o g y / C o m p l i c a t i o n sO R I G I N A L A R T I C L E
CD34+ cells from baseline. Secondary endpoints were changes in other progenitorcell phenotypes, proangiogenic capacityof peripheral blood mononuclear cells(PBMCs), white blood cells, and safety.The study was not designed and poweredto detect baseline differences in progenitorcell levels. DM patients were recruited atthe outpatient clinic of the University Hos-pital of Padova, and healthy control sub-jects were volunteers from the localcommunity. Both type 1 DM and type 2DM patients were eligible because preclin-ical studies have shown similar BM alter-ations and progenitor cell reductions inboth types of DM (4,5). Exclusion criteriawere as follows: age ,25 or .65 years;any acute disease or infection; recenttrauma, surgery, or cardiovascular event;chronic immune or infectious diseases;current or past hematological disordersor malignancy; leukocytosis, leukopenia,or thrombocytopenia; organ transplanta-tion or immune suppression; advanceddiabetic retinopathy; altered liver func-tion; severe renal failure (estimated glo-merular filtration rate ,30 mL/min/m2);anomalies in lymphocytes subpopula-tions; allergy to Filgrastim; bronchialasthma or other chronic lung disorders;and impossibility to provide informedconsent. For each patient, we collected
anthropometric measures, data on con-comitant risk factors, HbA1c, eventualDM complications, and therapy.
After providing informed consent,patients were subjected to baseline exam-ination and blood samples, including de-termination of the complete leukocytecounts and lymphocyte subpopulations,liver enzymes, renal function, plasmaprotein electrophoresis, erythrocyte sed-imentation rate, C-reactive protein, pro-thrombin time, uric acid, and standardurine examination. After verification of in-clusion and exclusion criteria, at 8:30 A.M.
after an overnight fast, eligible patients weresubjected to a baseline peripheral bloodsampling for circulating progenitor cellquantification and collection of PBMCs.Immediately after, they were injectedsubcutaneously with 5 mg/kg Filgrastim(Granulokine; Amgen). Twenty-fourhours later, another peripheral blood sam-ple was obtained to evaluate the effects ofFilgrastim. Study subjects were invited toregister and report any eventual side effectoccurred after Filgrastim injection. Dosageof the drug was chosen as the minimumeffective dose based on available pharma-codynamic data on Filgrastim, showingthat 5 mg/kg is sufficient to increase abso-lute count of circulating CD34+ cells inhealthy control subjects (13).
Fluorescence-activated cell sorteranalysisCirculating progenitor cells were quanti-fied using flow cytometry as previouslydescribed in detail (14). In brief, aftererythrocyte lysis, 150 mL peripheralblood were stained with 10 mL fluores-cein isothiocyanate-conjugated anti-human CD34 mAb (Becton Dickinson),10 mL phycoethrin-conjugated anti-humanKDR mAb (R&D Systems), and 10 mLallophycocyanin-conjugated anti-CD133mAb (Miltenyi Biotech). The frequencyof peripheral blood cells positive for thesereagents was determined by a two-dimensional side-scatter fluorescence dotplot analysis after appropriate gating.We gated CD34+ or CD133+ peripheralblood cells in the mononuclear cell frac-tion and then examined the resulting pop-ulation for the dual expression of KDR. Atthe intersection of the CD34 and CD133gates, we identified CD34+CD133+ cells,which were examined for KDR expression.In all patients, we also quantified theexpression of CD26/DPP-4 on CD34+
cells using a phycoethrin-labeled anti-CD26mAb (BectonDickinson). In separateanalyses, CD45 costaining was performedand showed that .90% of CD34+ cellsare CD45dim. For fluorescence-activatedcell sorter analysis, 5 3 105 cells were
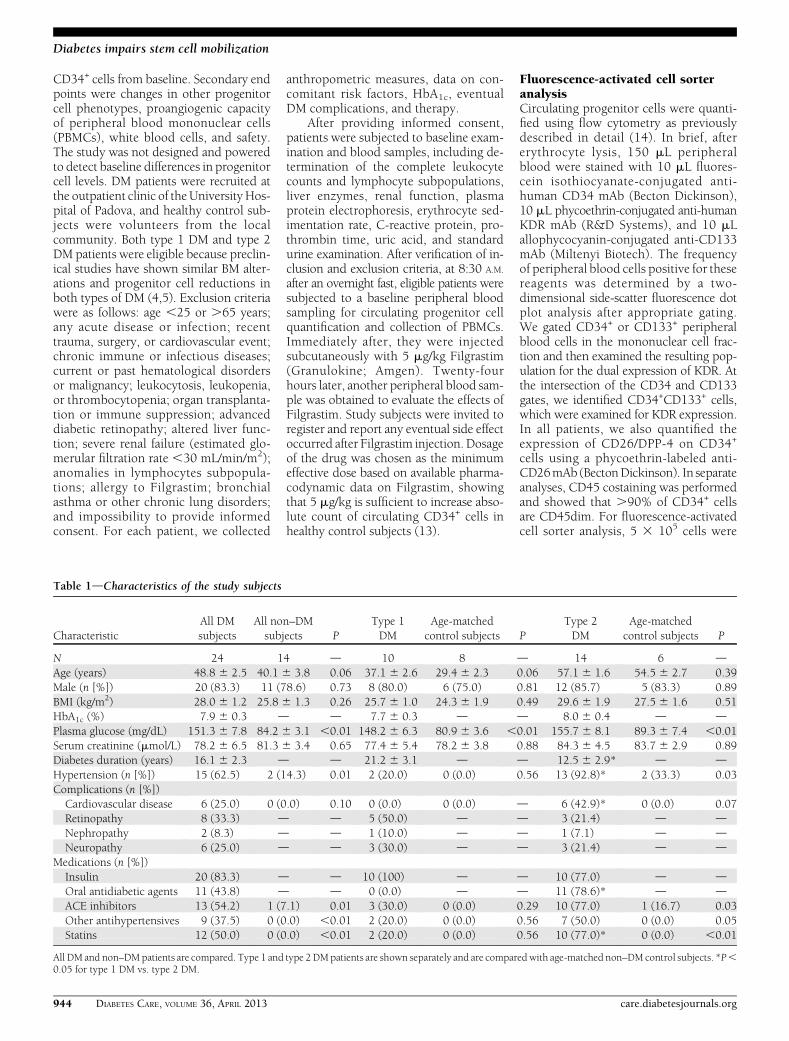
Table 1dCharacteristics of the study subjects
CharacteristicAll DMsubjects
All non–DMsubjects P
Type 1DM
Age-matchedcontrol subjects P
Type 2DM
Age-matchedcontrol subjects P
N 24 14 d 10 8 d 14 6 dAge (years) 48.8 6 2.5 40.1 6 3.8 0.06 37.1 6 2.6 29.4 6 2.3 0.06 57.1 6 1.6 54.5 6 2.7 0.39Male (n [%]) 20 (83.3) 11 (78.6) 0.73 8 (80.0) 6 (75.0) 0.81 12 (85.7) 5 (83.3) 0.89BMI (kg/m2) 28.0 6 1.2 25.8 6 1.3 0.26 25.7 6 1.0 24.3 6 1.9 0.49 29.6 6 1.9 27.5 6 1.6 0.51HbA1c (%) 7.9 6 0.3 d d 7.7 6 0.3 d d 8.0 6 0.4 d dPlasma glucose (mg/dL) 151.3 6 7.8 84.2 6 3.1 ,0.01 148.2 6 6.3 80.9 6 3.6 ,0.01 155.7 6 8.1 89.3 6 7.4 ,0.01Serum creatinine (mmol/L) 78.2 6 6.5 81.3 6 3.4 0.65 77.4 6 5.4 78.2 6 3.8 0.88 84.3 6 4.5 83.7 6 2.9 0.89Diabetes duration (years) 16.1 6 2.3 d d 21.2 6 3.1 d d 12.5 6 2.9* d dHypertension (n [%]) 15 (62.5) 2 (14.3) 0.01 2 (20.0) 0 (0.0) 0.56 13 (92.8)* 2 (33.3) 0.03Complications (n [%])Cardiovascular disease 6 (25.0) 0 (0.0) 0.10 0 (0.0) 0 (0.0) d 6 (42.9)* 0 (0.0) 0.07Retinopathy 8 (33.3) d d 5 (50.0) d d 3 (21.4) d dNephropathy 2 (8.3) d d 1 (10.0) d d 1 (7.1) d dNeuropathy 6 (25.0) d d 3 (30.0) d d 3 (21.4) d d
Medications (n [%])Insulin 20 (83.3) d d 10 (100) d d 10 (77.0) d dOral antidiabetic agents 11 (43.8) d d 0 (0.0) d d 11 (78.6)* d dACE inhibitors 13 (54.2) 1 (7.1) 0.01 3 (30.0) 0 (0.0) 0.29 10 (77.0) 1 (16.7) 0.03Other antihypertensives 9 (37.5) 0 (0.0) ,0.01 2 (20.0) 0 (0.0) 0.56 7 (50.0) 0 (0.0) 0.05Statins 12 (50.0) 0 (0.0) ,0.01 2 (20.0) 0 (0.0) 0.56 10 (77.0)* 0 (0.0) ,0.01
All DM and non–DMpatients are compared. Type 1 and type 2DMpatients are shown separately and are compared with age-matched non–DM control subjects. *P,0.05 for type 1 DM vs. type 2 DM.
944 DIABETES CARE, VOLUME 36, APRIL 2013 care.diabetesjournals.org
Diabetes impairs stem cell mobilization

acquired and scored using a FACSCalibur(BD). Data were processed using the Mac-intosh CELLQuest software program(BD). The same trained operators, blindto the clinical status of the patients, per-formed the tests throughout the study.Absolute progenitor cell counts per unitof blood were derived by multiplyingfractional data per white blood cellcount. We previously have shown thatreproducibility of CD34+ cell quantifica-tion with this method is high (intraclasscorrelation coefficient 0.94; 95% CI0.88–0.96; coefficient of variation [CV]6.3%) (14).
In vivo proangiogenic cell functionTo gather information on the presence offunctional circulating proangiogenic cellsand how they are modulated by hrG-CSFin subjects with and without DM, we usedthe in vivo Matrigel plug angiogenesisassay with patients’ PBMCs. Data suggestthat diverse monocyte subsets, includingmonocytic EPCs and Tie2-expressingmonocytes, have proangiogenic capacity
(15,16). In brief, PBMCs were isolatedwith Histopaque (Sigma-Aldrich). Cellcount and viability were assayed with anautomated BioRad TC20 cell counter, andthen, 3 3 106 PBMCs were resuspendedin 500 mL phenol-free Matrigel (catalognumber 356237; BD) and implanted sub-cutaneously into the dorsum of immuno-deficient RAG-2/g(c) double knockoutmice (in-house colony). The experimentwas performed with pre-hrG-CSF andpost-hrG-CSF PBMCs of five non–DMcontrol subjects, five type 1 DM patients,and five type 2 DM patients. To minimizevariability, the same mouse received pre-GCSF and post-GCSF PBMCs of the samesubject. Plugs were explanted 10 dayslater for macroscopic inspection, histol-ogy (hematoxylin and eosin staining),and determination of the hemoglobin-to-protein content ratio (Drabkin solu-tion and Bradford reagent, respectively;Sigma-Aldrich), which is a surrogate ofperfusion. Hemoglobin-to-protein ratio wasadjusted for the change in monocyte countafter hrG-CSF to gather information on
the proangiogenic capacity of circulatingPBMCs at each time point.
Statistical analysisData are expressed as mean6 SE. Normaldistribution of the variables of interestwas verifiedwith theKolmogorov-Smirnovtest. Comparisons between the diabeticgroup and nondiabetic group were per-formed using Student t test for normalvariables, Mann-Whitney U test for non-normal variables, and the x2 test for cate-gorical variables. Because CD34+ cellcount is a normally distributed variable,the change in CD34+ cell count frombaseline to 24 h after Filgrastim was as-sessed using paired Student t test. Wethen calculated the mean 6 SE changeof CD34+ cells in the diabetic and non-diabetic groups, which were comparedusing unpaired Student t test. Linear as-sociations were assayed using the Pearsonr correlation coefficient. To control forvariables that were different between thetwo groups at P# 0.10 and that may biasresults, a multiple linear regression
Figure 1dCD34+ cell mobilization after hrG-CSF. A: Absolute pre-G-CSF and post-G-CSF CD34+ cell count in all DM and all non–DM control(Ctrl) subjects (study primary end point). *P , 0.05 vs. baseline. Absolute pre-G-CSF and post-G-CSF CD34+ cell count in type 1 DM (T1D)compared with young control subjects (B) and in type 2 DM (T2D) compared with age-matched control subjects (C). *P , 0.05 vs. baseline.D: Changes in the absolute levels of circulating CD34+ cell counts. *P , 0.05 vs. DM.
care.diabetesjournals.org DIABETES CARE, VOLUME 36, APRIL 2013 945
Fadini and Associates

analysis was performed with change in pro-genitor cell levels as the dependent variable.Nonnormal dependent variables in second-ary endpoint analyseswere log transformed.Statistical significance was accepted at P ,0.05; SPSS version 17.0 was used.
RESULTS
Characteristics of the studypopulationA total of 24 DM patients (10 type 1 DMand 14 type 2 DM patients) and 14control subjects without DM have beenenrolled and treated. DM patients had ahigher prevalence of hypertension andtended to be older than control subjects.Type 1 DM patients had significantlylonger disease duration and lower prev-alence of cardiovascular disease com-pared with type 2 DM patients (Table 1).
Stem and progenitor cell mobilizationIn subjects without DM, absolute CD34+
cell level significantly increased 2.2-foldafter hrG-CSF, whereas CD34+ cells com-pletely failed to mobilize in DM patients(mean 6 SE change vs. baseline; non–DM: 3,475 6 800 cells/mL; DM: 52 6332 cells/mL; P = 5 3 1025) (Fig. 1A).Results were similar when type 1 or
type 2 DM patients, considered sepa-rately, were compared with age-matchednon–DM control subjects (Fig. 1B and C).Clinical characteristics of the subgroupsare shown in Table 1.
After adjusting for potential con-founders that were different between thetwo groups at P # 0.10 (age, hyperten-sion, and cardiovascular disease), DM re-mained significantly associated withreduced CD34+ cell increase after hrG-CSF (P = 0.002).
In response to hrG-CSF, DM was asso-ciated with impaired mobilization of all theother progenitor cell phenotypes, such asCD133+, CD34+CD133+, CD34+KDR+,CD133+KDR+, and CD34+CD133+KDR+
cells (Fig. 2). On correction for potentialconfounders, DM remained significantlyassociated with defective mobilization ofCD133+ (P = 0.015), CD34+CD133+ (P =0.011), and CD133+KDR+ (P = 0.013) cells,whereas the association between defec-tive CD34+KDR+ mobilization and DMwas marginally significant (P = 0.056) andblunted by age (P = 0.024).
Percentage expression of CD26/DPP-4on CD34+ cells significantly increased af-ter hrG-CSF in non–DM control subjects(d = +14.16 3.9%), consistent with pre-vious findings in vitro (17). In DM,
CD26/DPP-4 expression was elevatedat baseline and tended to decline afterhrG-CSF treatment (d = 28.8 6 5.5%;P = 0.013 vs. non–DM) (Fig. 3).
In the DM groups, progenitor cellmobilization was not significantly corre-lated to HbA1c, disease duration, patternof complications, or treatment regimen.
After treatment with hrG-CSF, whiteblood cell counts, neutrophil counts, andmonocyte counts significantly increasedin both groups, and there were no differ-ences between patients with DM andcontrol subjects without DMs (Table 2and Supplementary Fig. 1). This suggeststhat DM affects immature, but not ma-ture, cell mobilization.
Proangiogenic cell function in vivoMononuclear cells collected from patientsbefore and after hr-GSCF administrationwere embedded into Matrigel plugs andimplanted in immunodeficient mice toassess the presence of proangiogenic cells.Baseline PBMC from non–DM subjectsshowed higher neovascularization capac-ity compared with DM patients, whichwas statistically significant versus type 1DM. Plugs with non–DM PBMCs beforehrG-CSF (baseline) showed vascular inva-sion at gross inspection and the presence of
Figure 2dMobilization of other progenitor cell phenotypes after hrG-CSF. A–E: Absolute pre-G-CSF and post-G-CSF cell counts of circula-ting hematopoietic stem cells (CD133+, CD34+CD133+) and endothelial progenitor cells (CD34+KDR+, CD133+KDR+, and CD34+CD133+KDR+) inDM and non–DM patients. *P, 0.05 vs. baseline. F: Changes in the absolute levels of stem/progenitor cell phenotypes in DM and non–DM patients.*P , 0.05 vs. non–DM control subjects.
946 DIABETES CARE, VOLUME 36, APRIL 2013 care.diabetesjournals.org
Diabetes impairs stem cell mobilization

vascular structures containing erythrocytesat histology. Plugs implanted with type 1DM PBMCs at baseline showed almost novascularization, lower hemoglobin con-tent, and no evidence of perfused vascularstructures. Plugs containing baseline type 2DMPBMCs showed nonsignificantly lowervascularization capacity compared withnon–DM at baseline. After hrG-CSF, theproangiogenic capacity of patients’ cells sig-nificantly increased in non–DM, but not inDM groups, and post-hrG-CSF neovascu-larization capacity was lower in type 1 DM(P = 0.058) and type 2 DM (P = 0.045)versus non–DM (Fig. 4). Matrigel plug neo-vascularization capacity was significantlycorrelated with circulating CD34+ (r =0.47; P = 0.003), CD34+KDR+ (r = 0.38;P = 0.024), CD133+KDR+ (r = 0.46; P =
0.005), and CD34+CD133+KDR+ (r =0.41; P = 0.014) cell levels (SupplementaryFig. 2). These data suggest that hrG-CSFmobilizes functional proangiogenic cells innon–DM subjects but not in DM patients.
SafetyTreatment with hrG-CSF was safe anduneventful. Five subjects in the non–DMgroup (35.7%) and five patients in theDM group (20.8%; P = 0.53 DM vs. non–DM) reported mild back pain 12 to 18 hafter Filgrastim injection, which resolved at24 h and required analgesic therapy withacetaminophen in three non–DM cases.
CONCLUSIONSdWe show that DMis associated with impaired stem andprogenitor cell mobilization after directBM stimulation, independently of poten-tial confounders. Remarkably, this wastrue for both hematopoietic stem cells andEPCs despite baseline cell levels not beingreduced in this DM cohort. Therefore, thepresent prospective trial substantiates theexistence of a BM defect in human DM andsuggests that the BM mobilization failureprecedes reduction of circulating progeni-tors. Both type 1 DM and type 2 DMpatients showed almost complete unre-sponsiveness to stem/progenitor cell mobi-lization, suggesting that this complication isindependent of DM etiology; however, therole of autoimmunity in determining BMresponse may be worthy of investigation.
In mice, long-term DM causes BMmicroangiopathy and altered oxygen gra-dients (4) that, in addition to reduced ex-pression of prosurvival genes (6), lead to apauperization of the stem cell pool. Mi-crovascular BM alterations in experimen-tal DM include capillary rarefaction,increased permeability, endothelial cellapoptosis, and dysfunction (4), featuresthat resemble diabetic microangiopathyof other organs, such as the kidney andretina. These histopathological aspectssuggest that the BM is a hitherto unrecog-nized site of DM complication and islikely responsible for the mobilizationfailure. Data on the amount of BM stemcells in DM are discordant, with somestudies showing normal (3,18) or even in-creased (7) primitive Sca-1+c-kit+Lin2
hematopoietic progenitors. Therefore,the low CD34+ cell count in BM aspiratesfrom type 2 DM patients that we haveshown previously (9) might reflect truestem cell deficiency or reduced accessibil-ity of the niches to aspiration, attributableto the sticky property of the diabeticniche, which is more prone to stem cellretention thanmobilization (7). It is note-worthy that DM did not impair mobiliza-tion of mature leukocytes, which aremore loosely retained by the BM stromathan stem cells.
Normally, G-CSF stimulates expres-sion and activity of CD26/DPP-4 andother proteases, with subsequent degra-dation of the chemokine and retentionsignal SDF-1a (17,19). Thus, stem/pro-genitor cells migrate to the peripheral cir-culation following SDF-1a gradients.Herein, we suggest a possible mechanismof stem cell unresponsiveness to G-CSF inDM by showing a maladaptive CD26/DPP-4 response. Systemic CD26/DPP-4activity is increased in DM (11), and ournew data indicate that G-CSF fails to up-regulate CD26/DPP-4 on BM-derivedcells in DM, likely preventing modifica-tion of the SDF-1a gradient. Studiesshow that DM mice mobilize stem cellsafter treatment with the SDF-1a receptorCXCR4 antagonists AMD3100 (7,18,20)andNIBR1816 (21). Therefore, the vascularniche containing stem cells that can be read-ily mobilized on disruption of the SDF-1aretention signal seems to be preserved, butresponsiveness to CXCR4 antagonistsshould be confirmed in DM patients.
The existence of a BMmobilopathy inDM might be ascribed to both structuralalterations affecting the stem cell niche(microangiopathy) and functional defectspreventing the cells from being mobilized
Figure 3dEffects of hrG-CSF on CD26/DPP-4expression. Percentage CD26/DPP-4 expres-sion on CD34+ cells was significantly (*P ,0.05) increased in non–DM control subjects,whereas it was reduced in DM patients.
Table 2dHematological parameters and absolute progenitor cell counts in DM andnon–DM patients before and after administration of hrG-CSF
Variable
Diabetic patients(n = 24)
Nondiabetic subjects(n = 14)
Baseline Post-hrG-CSF Baseline Post-hrG-CSF
White blood cells(3103/mL) 6.53 6 0.32 21.88 6 0.82* 6.05 6 0.34 23.54 6 1.14*
Neutrophils (3103/mL) 3.57 6 0.26 17.91 6 0.74* 3.29 6 0.22 19.94 6 0.97*Lymphocytes (3103/mL) 2.20 6 0.10 2.60 6 0.14 2.02 6 0.12 2.34 6 0.20Monocytes (3103/mL) 0.55 6 0.03 1.06 6 0.04* 0.52 6 0.05 1.00 6 0.04*Eosinophils (3103/mL) 0.18 6 0.02 0.28 6 0.03 0.19 6 0.04 0.27 6 0.06Basophils (3103/mL) 0.03 6 0.003 0.04 6 0.004 0.03 6 0.005 0.04 6 0.003Erythrocytes (3106/mL) 4.88 6 0.07 4.82 6 0.08 5.04 6 0.11 4.91 6 0.12Platelets (3103/mL) 235.4 6 7.5 231.5 6 8.7 243.0 6 12.4 237.6 6 13.1CD34+ cells/mL 3,236 6 269 3,289 6 269 2,837 6 351 6,312 6 743*CD133+ cells/mL 1,884 6 185 2,128 6 212 937 6 184 2,476 6 409*CD34+CD133+ cells/mL 1,204 6 179 1,423 6 193 719 6 156 2,152 6 374*CD34+KDR+ cells/mL 253 6 34 223 6 38 201 6 46 515 6 145*CD133+KDR+ cells/mL 287 6 55 274 6 40 249 6 84 754 6 266*CD34+CD133+KDR+
cells/mL 31 6 8 27 6 7 38 6 11 86 6 27
*P , 0.05 vs. baseline.
care.diabetesjournals.org DIABETES CARE, VOLUME 36, APRIL 2013 947
Fadini and Associates

(e.g., the CD26/SDF-1a/CXCR4 axis). Inaddition, DM alters the activity of endo-thelial nitric oxide synthase (22), which ispivotal for EPC mobilization (23), and al-though G-CSF activity is mainly endothe-lial nitric oxide synthase–independent(24), this may be another mechanism ac-counting for depressed mobilization inDM. Among strategies to reverse BM dys-function, our data showing no correlationbetween HbA1c and mobilization suggestthat glucose control might not be effec-tive, whereas experimental data indicatethat boosting the antioxidative defenseis a suitable strategy to prevent BM alter-ations (4). Although CD26/DPP-4 inhibi-tion increases EPCs (25), whether thisrepresents a therapeutic target to restoreBM responsiveness in DM needs to be as-certained.
We also found that DM impairs neo-vascularization by mononuclear cells
mobilized by hrG-CSF, as shown by the invivo Matrigel plug assay. It is noteworthythat type 1 DM patients already had de-fective neovascularization capacity at base-line. This is possibly related to the longerdisease duration in type 1 DM comparedwith type 2 DM patients (Table 1). Bothtype 1 DM and type 2 DM patients wereunable to increase significantly theirPBMCs proangiogenic capacity after hr-GCSF, again suggesting that mobilizationfailure precedes reduction of proangiogeniccells. Althoughwe did not identify the sub-population of PBMCs involved (15,16),change in neovascularization capacity wascorrelated with stem/progenitor mobiliza-tion. These important results indicate thatthe stem cell mobilization failure can bepathophysiologically linked to impairedtissue repair anddevelopment of cardiovas-cular DM complications, which are charac-terized by defective angiogenesis (26,27).
Our findings have clinical implica-tions. Reduced progenitor cell levels arepowerful predictors of future cardiovas-cular events (28), and replenishment ofprogenitor cells may lower cardiovascularrisk. Although the pathogenesis of dia-betic vascular complications is complexand possibly diversified in type 1 DMand type 2 DM, it is remarkable that alter-ations in BM-derived cells have been con-sistently reported for both type 1 DM (29)and type 2 DMpatients (30). Understand-ing the causes of progenitor cell reductionand the role of BM can identify novelstrategies to reverse this defect and pre-vent vascular disease. Moreover, the im-munomodulatory activity of mobilizedBM-derived progenitors may be impor-tant in type 1 DM (21). In addition, clini-cians should be aware of the likelihood ofmobilization failure in DM patients un-dergoing stem cell collection for BM au-totransplantation or for angiogenic celltherapy (7,8).
The study has limitations. First, rep-lication in other, possibly larger, co-horts is needed and comparison betweenrecent-onset and long-term type 1 DMpatients would allow a description ofthe natural history of BM dysfunction.Second, whereas the imbalances in clinicalcharacteristics between groups were ad-justed by multivariate analyses, residualconfounding may be present. Finally,whereas a single-dose hrG-CSF wasused in this study, DM patients mightrespond to a full 5-day course of hrG-CSF,but it would not be ethical to performmaximal BM stimulation for research pur-poses only. Several DM patients have beentreated with high-dose hrG-CSF in celltherapy protocols (31), but a formal com-parison of the mobilization effect betweenDM and non–DM patients never hasbeen performed. Nonetheless, our datarepresent a proof-of-concept of diabeticBM dysfunction in humans. As the BMemerges as a novel target organ in DM,intensive investigation to reverse thiscomplication becomes compelling.
AcknowledgmentsdThis study was sup-ported by grants from the University of Padova(60A07-8452) and the European Foundationfor the Study of Diabetes (EFSD)/Lilly Fel-lowship and EFSD/Sanofi Clinical InvestigatorFellowship to G.P.F.No potential conflicts of interest relevant to
this article were reported.G.P.F. designed the study and wrote the
manuscript. G.P.F., M.A., S.V.d.K., E.B., R.C.,M.M., and N.P. researched data. C.A. designed
Figure 4dIn vivo proangiogenic cell function. Patients’ PBMCs before collected pre-G-CSF andpost-G-CSF were embedded into Matrigel plugs and implanted in immunodeficient mice. Rep-resentative gross appearance of the plugs (scale bar = 1 cm) and histological sections (scale bar =100mm) are shown. In the bottom, quantification of hemoglobin content, a quantitative surrogateof perfusion, adjusted for mobilized monocytes is shown. *P, 0.05 vs. pre-G-CSF. †P, 0.05 vs.non–DM.
948 DIABETES CARE, VOLUME 36, APRIL 2013 care.diabetesjournals.org
Diabetes impairs stem cell mobilization

the study and contributed to discussion. A.A.reviewed and edited the manuscript, contrib-uted to discussion, and supervised the project.G.P.F. is the guarantor of this work and, assuch, had full access to all the data in the studyand takes responsibility for the integrity of thedata and the accuracy of the data analysis.
References1. Fadini GP, Avogaro A. It is all in the blood:
the multifaceted contribution of circulatingprogenitor cells in diabetic complications.Exp Diabetes Res 2012;2012:742976
2. Fadini GP. An underlying principle forthe study of circulating progenitor cellsin diabetes and its complications. Dia-betologia 2008;51:1091–1094
3. Fadini GP, Sartore S, Schiavon M, et al.Diabetes impairs progenitor cell mobilisa-tion after hindlimb ischaemia-reperfusioninjury in rats. Diabetologia 2006;49:3075–3084
4. Oikawa A, Siragusa M, Quaini F, et al.Diabetes mellitus induces bone marrowmicroangiopathy. Arterioscler ThrombVascBiol 2010;30:498–508
5. Busik JV, Tikhonenko M, Bhatwadekar A,et al. Diabetic retinopathy is associatedwith bone marrow neuropathy and a de-pressed peripheral clock. J ExpMed 2009;206:2897–2906
6. Orlandi A, Chavakis E, Seeger F, Tjwa M,Zeiher AM, Dimmeler S. Long-term di-abetes impairs repopulation of hemato-poietic progenitor cells and dysregulatesthe cytokine expression in the bone mar-row microenvironment in mice. Basic ResCardiol 2010;105:703–712
7. Ferraro F, Lymperi S, Mendez-Ferrer S,et al. Diabetes impairs hematopoietic stemcell mobilization by altering niche func-tion. Sci Transl Med 2011;3:104ra101
8. DiPersio JF.Diabetic stem-cell “mobilopathy”.N Engl J Med 2011;365:2536–2538
9. Fadini GP, Boscaro E, de Kreutzenberg S,et al. Time course and mechanisms ofcirculating progenitor cell reduction inthe natural history of type 2 diabetes. Di-abetes Care 2010;33:1097–1102
10. Roberts AWG-CSF. a key regulator ofneutrophil production, but that’s not all!Growth Factors 2005;23:33–41
11. Fadini GP, Albiero M, Menegazzo L, deKreutzenberg SV, Avogaro A. The in-creased dipeptidyl peptidase-4 activity is
not counteracted by optimized glucosecontrol in type 2 diabetes, but is lower inmetformin-treated patients. Diabetes ObesMetab 2012;14:518–522
12. Fadini GP. Is bone marrow another targetof diabetic complications? Eur J Clin In-vest 2011;41:457–463
13. van Der Auwera P, Platzer E, Xu ZX, et al.Pharmacodynamics and pharmacoki-netics of single doses of subcutaneouspegylated human G-CSF mutant (Ro 25-8315) in healthy volunteers: comparisonwith single and multiple daily doses offilgrastim. Am JHematol 2001;66:245–251
14. Fadini GP, de Kreutzenberg SV, CoracinaA, et al. Circulating CD34+ cells, meta-bolic syndrome, and cardiovascular risk.Eur Heart J 2006;27:2247–2255
15. Rehman J, Li J, Orschell CM, March KL.Peripheral blood “endothelial progenitorcells” are derived from monocyte/macro-phages and secrete angiogenic growthfactors. Circulation 2003;107:1164–1169
16. Pucci F, Venneri MA, Biziato D, et al. Adistinguishing gene signature shared bytumor-infiltrating Tie2-expressing mono-cytes, blood “resident” monocytes, andembryonic macrophages suggests com-mon functions and developmental rela-tionships. Blood 2009;114:901–914
17. Christopherson KW 2nd, Uralil SE,Porecha NK, Zabriskie RC, Kidd SM,Ramin SM. G-CSF- and GM-CSF-inducedupregulation of CD26 peptidase down-regulates the functional chemotactic re-sponse of CD34+CD38- human cordblood hematopoietic cells. Exp Hematol2006;34:1060–1068
18. Tepper OM, Carr J, Allen RJ Jr, et al.Decreased circulating progenitor cellnumber and failed mechanisms of stromalcell-derived factor-1alpha mediated bonemarrow mobilization impair diabetic tissuerepair. Diabetes 2010;59:1974–1983
19. Christopherson KW 2nd, Cooper S,Broxmeyer HE. Cell surface peptidaseCD26/DPPIV mediates G-CSF mobiliza-tion of mouse progenitor cells. Blood2003;101:4680–4686
20. Nishimura Y, Ii M, Qin G, et al. CXCR4antagonist AMD3100 accelerates im-paired wound healing in diabetic mice.J Invest Dermatol 2012;132:711–720
21. Fiorina P, Jurewicz M, Vergani A, et al.Targeting the CXCR4-CXCL12 axis mo-bilizes autologous hematopoietic stem
cells and prolongs islet allograft survivalvia programmed death ligand 1. J Im-munol 2011;186:121–131
22. Du XL, Edelstein D, Dimmeler S, Ju Q, SuiC, Brownlee M. Hyperglycemia inhibitsendothelial nitric oxide synthase activity byposttranslational modification at the Aktsite. J Clin Invest 2001;108:1341–1348
23. Aicher A, Heeschen C, Mildner-Rihm C,et al. Essential role of endothelial nitricoxide synthase for mobilization of stemand progenitor cells. Nat Med 2003;9:1370–1376
24. de Resende MM, Huw LY, Qian HS,Kauser K. Role of endothelial nitric oxidein bone marrow-derived progenitor cellmobilization. Handb Exp Pharmacol2007;(108):37–44
25. Fadini GP, Boscaro E, Albiero M, et al.The oral dipeptidyl peptidase-4 inhibitorsitagliptin increases circulating endothe-lial progenitor cells in patients with type 2diabetes: possible role of stromal-derivedfactor-1alpha. Diabetes Care 2010;33:1607–1609
26. Boodhwani M, Sodha NR, Mieno S, et al.Functional, cellular, and molecular char-acterization of the angiogenic response tochronic myocardial ischemia in diabetes.Circulation 2007;116(Suppl):I31–I37
27. Caporali A, Meloni M, Völlenkle C, et al.Deregulation of microRNA-503 contrib-utes to diabetes mellitus-induced impair-ment of endothelial function and reparativeangiogenesis after limb ischemia. Circula-tion 2011;123:282–291
28. Fadini GP, Maruyama S, Ozaki T, et al.Circulating progenitor cell count for car-diovascular risk stratification: a pooledanalysis. PLoS ONE 2010;5:e11488
29. Loomans CJ, de Koning EJ, Staal FJ, et al.Endothelial progenitor cell dysfunction:a novel concept in the pathogenesis ofvascular complications of type 1 diabetes.Diabetes 2004;53:195–199
30. Tepper OM, Galiano RD, Capla JM, et al.Human endothelial progenitor cells fromtype II diabetics exhibit impaired pro-liferation, adhesion, and incorporationinto vascular structures. Circulation 2002;106:2781–2786
31. Fadini GP, Agostini C, Avogaro A. Au-tologous stem cell therapy for peripheralarterial disease meta-analysis and sys-tematic review of the literature. Athero-sclerosis 2010;209:10–17
care.diabetesjournals.org DIABETES CARE, VOLUME 36, APRIL 2013 949
Fadini and Associates