determination and analysis of the primary structure of the nerve
TRANSCRIPT

The EMBO Journal vol.5 no. 12 pp.3167-3173, 1986
Determination and analysis of the primary structure of the nerveterminal specific phosphoprotein, synapsin I
Cheryl A.McCaffery and Louis J.DeGennaroMax Planck Institute for Psychiatry, Department of Neurochemistry, D-8033Martinsried, FRG
Communicated by H.Thoenen
A rat brain cDNA clone containing an open reading frameencoding the neuron-specific protein synapsin I has been se-quenced. The sequence predicts a protein of 691 amino acidswith a mol. wt of 73 kd. This is in excelent agreement withthe size of rat brain synapsin lb measured by SDS-polyacryl-amide gel electrophoresis. Inspection of the predicted primarystructure has revealed the probable sites for synapsin I phos-phorylation by the cAMP-dependent and Ca2 +/calhodulin-dependent protein kinases. All of the biochemically observedintermediates of synapsin I digestion by collagenase can beverified by inspection of the sequence, and the collagenase-resistant fragment has been defined as the amino-terminal439 amino acids of the molecule. Predictions of sequencesecondary structure and hydrophobicity suggest that a centraldomain of - 270 amino acids may exist as a folded, globularcore. The carboxyl-terminal domain of the protein (the regionsensitive to collagenase digestion) contains sites for Ca2+/cal-modulin-dependent protein kinase phosphorylation. Thesesites are flanked by three regions of repeating amino acidsequence that are proposed to be the synaptic vesicle-bindingdomain of synapsin I. This region also shares homology withthe actin-binding proteins profilin and villin. The character-istics of the synapsin I sequence do not support extensive hom-ology with the erythrocyte cytoskeletal protein 4.1.Key words: cDNA/cytoskeleton/neurotransmitter vesicle/proteinkinase/protein structure
IntroductionThe search for the molecular mechanisms underlying the nervecell-specific function of synaptic transmission has led to two majorareas of investigation. One is the characterization of neuron-specific proteins. The second is the study of the role of proteinphosphorylation in the regulation of events involved in synaptictransmission. Synapsin I is a neuron-specific phosphoproteinwhose characteristics overlap both these areas of investigation,and suggest that it may play an important role in synaptic trans-mission.
Synapsin I is composed of two very similar polypeptides re-ferred to as synapsin Ia and synapsin Tb, respectively (Ueda andGreengard, 1977). Both proteins have similar amino acid com-positions and basic isoelectric points (> 10). They both have arelatively high content of proline and glycine and are substratesfor degradation by highly purified collagenase. It has been sug-gested that synapsin I is composed of a globular collagenase-resistant domain and a highly elongated collagenase-sensitivedomain (Ueda and Greengard, 1977; Schiebler et al., 1986).
Immunohistochemical studies have localized synapsin I to the pre-synaptic terminals of virtually all neurons, where it is associatedwith the cytoplasmic surface of small synaptic vesicles (DeCamilliet al., 1983a,b; Navone et al., 1984). The protein is phosphoryl-ated by both cAMP-dependent and Ca2+/calmodulin-dependentprotein kinases, and its state of phosphorylation is altered by con-ditions that affect neuronal activity (for a review, see Nestler andGreengard, 1984). Nerve cell depolarization has been shown toinduce the phosphorylation of synapsin I, by Ca2+/calmodulin-dependent protein kinase II, at specific sites in the collagenase-sensitive domain of the molecule (Huttner and Greengard, 1979;Huttner et al., 1981), thereby reducing its association with syn-aptic vesicle membranes (Huttner et al., 1983; Schiebler et al.,1986). A more detailed molecular description of these sites istherefore of particular interest with regard to the elucidation ofsynapsin I function.
Additional insight into the functional role of synapsin I in thenerve terminal has come from the studies of Baines and Bennett(1985). They demonstrated that synapsin I is immunologicallyrelated to the erythrocyte cytoskeletal protein 4.1 and is also aspectrin-binding protein. These results have led to the proposalthat synapsin I may interact with cytoskeletal elements in the pre-synaptic terminal.The characteristics of synapsin I suggest that it may play a role
in synaptic transmission through interactions with both cytoskel-etal elements and synaptic vesicles in the presynaptic terminal.In order to make a more detailed analysis of the molecular struc-ture of synapsin I we cloned cDNA molecules which are comp-lementary to rat brain synapsin I mRNA (Kilimann andDeGennaro, 1985). The clones were shown to be synapsin Ispecific by differential colony hybridization and hybridization-selected translation. Here we report the nucleotide sequence ofone of these synapsin I cDNAs, and the predicted primary struc-ture of the synapsin I protein. The synapsin I amino acid sequenceis analyzed with regard to the structure and function of themolecule and possible homology with other known proteins.
0 1000
_ta & 0 &0
5'lIII I I I 1It
-2 -
,_i
2000
X0 - 0c _
I I I
3000
>O
c uII I l
2t
~~~-b 4-~~~~~~~~~~~~~
poly(dA)
0. 0.I 3
Fig. 1. DNA restriction map of synapsin I cDNA clone pSyn 5 (heavyline). Restriction endonuclease cleavage sites are shown above. Horizontalarrows show the strategy (direction and extent of sequencing) used togenerate the sequence presented in Figure 2. Vertical arrows indicate thepredicted initiation (1) and termination (2) sites of translation. Scale at thetop is base pairs.
© IRL Press Limited, Oxford, England
4 --*
4-4
--'O 4-
3167
C.A.McCaffery and LjJ.DeGennaro
1 AACTTC ATG CCC AAT CTG GGT AAT GGG TAT ATG ACA GAG CTG GAG CC-C CC CGAA CCA CCT CCC CCC' CCG CCC TCA CCC GCC AGC CCA COOMet Ala Asn Leu Pro Ass Gly Tyr Met Thr Asp Leu Gin Arg Pro Gin Pro Pro Pro Pro Pro Pro 5cr Ala Ala 5cr Pro GLy 28
91 GCC ACT CCC GGA TCC OCT OCTr GCC TCT OCT GAG AGO GCCC(SACA OCT GCT CCA GTG GCC TCT CCA GCA GCC CCT_ ACT CCC COG FCC TCGALa Thr Pro Cly Ser Ala Ala ALa 5cr ALa Gtu Arg Ala \Sr/Thr Ala Ala Pro Vat ALa 5cr Pro Ala Ala Pro 5cr Pro Gly Ser 5cr 56
181 GCOO GGC COT GCO TTC TTC TCC1 TCG CTC TCT AGC GCG GTC AAA CAG ACC ACA GGA GCT GCA GCC GCC ACC TTC AGT GAO GAG GTG GCC GOTGly Gly Oly Gly Phe Phe 5cr Ser Leu 5cr Ass Ala Vat Lys GIn Thr Thr Ala Ala Ala Ala Ala Thr Phe 5cr CIu Gin Vat GLy Cly 88
271 GCO TCT COO GGC GCA GCC GCG GGG GCCG CCG CCC CCA 000 TOGC TGC TOO TCA TCG ACGO AOGC COGC AGA CCG ACT COO CAA AAT ACT CCA ACGCly Ser Gty CLy ALa GLy Ala GLy Ala Pro Pro Pro Gly Cys Cys Trp Scr 5cr Thr 5cr Arg Thr Pro Thr Oly GIn Ass Thr Pro Lys 118
361 GGA AGA ACA TGC ATO GAO AAA TTG AGA TTA AAG0 AAG AGC AAG0 CTO AAT TCT CCG ATC TCA ATC TTC TOO CTC ATG CCA ATG GTO GAT TCTCly Arg Arg Ser Mel Gtu Lys Leo Thr Leu Lys Lys Ser Lys Leu Ass Ser Pro lIe 5cr tIle Leu Trp Leu Mel Pro Mel Vat Asp Ser 148
451 CCG TOO AGA TCC AAO TTC TTC GOGA ATG COO TGC AAA OTT GIG AGO TCT CTG AAG0 GCA GAG TTT GTO CTG ATG CGCCAGA CAT GCC TIC AGC- P ro Trp Thr Trp Lys Phe Phe Cly Met Gly 5er Lys Val Vat Ary 5cr Leu Lys Pro Asp Phe Vat Leo Ite Arg GILn His Ala Phe 5cer 1178
h- 541 ATG GCA COT AAT GCA GAG TAG COGC POT TTG GCT ATT COO GIG CGA TAT GCGT GGG PIG CCC POT OTT AAG TCT TTC CAT TCT GTC TAG AAICMel ALa Arg Asn Gly Asp Tyr Prg See Leo Vat lIe Oly Leo GILn Tyr Ala GLy tIce Pro 5cr Vat Ass Ser Leo His 5cer Vat Tyr Ass 208
WI 631 TTT TOT GAG AAIA CCC TOG CTC TTT GCC CGA ATC OTT CGA CTA GAG /CAAG/AGCGTT GGA ACA GAO GAA TTC GCT CTG ATT GAT CGA ACT TTC0-,
Phe Cys Asp Lys Pro Trp Vat Phe Ala Gin Mel Vat Arg Leo His Lys Lys Leu Oly Thr Gtu CIa Phe Pro Leo IlIe Asp GIn Thr Phe 238
C/I.~< 721 TAT CCC AAT GAT AAA GAO ATG CTC AGC POGC ACA ACA TAO CGT GTA OTT GTG AAG ATOCGGGCAG GGA GAG TCT CCC ATrG GCO AAG GTG AAG0__ Tyr Pro Ass His Lys Gbu Met Leo 8cr 5cr Thr__Thr TXye Pro Vat Vat Vat Lys. Met Gly His Ala His Ser Gly Mel Gty Lys Vatl Lys 268
-1Cc 811 GTA GAG A/IC CA/I CAT GAG TTC GAG GAT ATT GCA ACT CTT GTG GCA CTC ACT A/IC ACA TAT GCC ACT OCT GAO CCC TTC ATT GAT OCT A/AA-jVat Asp Ass Gin His Asp Phe GCn Asp tIle Ala 5cr Vat Vat Ala Leu Thr Lys Thr Tyr A L a Thr Ala Gtu Pro Phe IlIe Asp Ala Lys 298
901 TACGCAT GTG COT CTC CGA AAC ATT COO CGA AAG TAG AAG GCC TAG ATG AOO AGA TCA CTG TCA GGG AAIC TGC A/I ACAGGA/T AGA GCO TCGTTye Asp Vat Prg Vat Cin Lys IlIe Gly Oln Ass Tyr Lys Ala Tyr Met Prg Thr 5cr Vat 5cr Gly Ass Trp Lys The Ass Thr Gly 5cr 328
891 OCT ATG CTT GAO CGA ATT OCT ATG TCT GAG AGG TAAAGPCITOG TOG GTA GAG ACGC TOGC TGA GAG ATT TTT GGG GGA CTT GAG ATC TOGC GCC/Ala Met Leo Gtu GIn ItIc Ala Met 5cr Asp Arg Tyr Lys Leo Trp Vat Asp Thr Cys 5cr GLu IlIe Phe Cly Gly Leu Asp tIe Cys Ala 358
1081 GTG GAAI GCA CGT CAT GCO AAG GACGOGA AOG GAT GAG ATT ATT GAO GTG GTG GCO TCC FCC ATG CCA CTC ATT GGC GAT GAG CGA GCG GATVat Glo Ala Leu His CLy Lys Asp Cly Arg Asp His tIle tIc Gbu Vat Vat Gly Ser See Mel Pro Leo lIe GIly Asp His Cmn Gly Asp 388
1171 GAA GAG AAGCAGA CTG ATC CTC GA/I CTT GTG CTC AAIC AAG ATG ACT CGA GCT CTC CCT COO GAG COO GAT OCT TCC CCT GGC ACG OCT TCCGlo Asp Lys ClIor Leu tIc Vat GIu Leu Vat Vat Ass Lys Mel Thr Gin Ala Leo Pro Arg GLsn Arg Asp Ala 5cr Pro Gly Arg Gty 5cer 418
1261 GAG AGC GAG ACT CCA TGCCCGGACC GCCC CTG CCC TTG GCO CGC CGA ACC TGC A .AiTO~AC1 OTT%A A A00COPA
- His 5cer Gin Thr Pro 5cer Pro Gly Ala Leu Pro Leu Oly Arg Gin Thr 5cr r.AatyPr wAl Iv41---A Gef r 4411
1351 CCTOGGAI~E GGA CA/I CCA WGC%CA GGA OVQ.IA. P. OWOACZCPCA P.A A
01in0HHiss1441 CTT TGT GCO CTTGACC CCA OCT GGC POGC CCT CTG CCC CGA CC-C CTA CCA POT CCC ACA GCA GCA CCT GAG GAG TCC GCC TCT CGA GCC
Leo 5cr Gly Leo t Pro Pro Ala Gty 5cr Pro Leu Pro Gtn Arg Leo Pro 5cr Pro The Ala Ala Pro Gin Gin 5cr ALa 5cr Gis Ala 508
iI.1531 ACA GCA ATG ACC CGA OCT CAA GCO COGC CGA TCA COO CCA GTG GCA O4GA FEGGC IcOcGA GCA CCT CCA GGA GCC CCC CCC CCG 0CC TCC CCA- Thr Pro Met Thr Gin Gly G in Cly Arg Cmn 5cer Ary Pro Vat Ala GlOly Pro Oly Ala Pro Pro Ala Ala Ar9 Pro Pro Ala 5cer Pro 538
t/) 1621 TGT CGA CGA COT CGA GCA CCCACGAGGP0 CTA CCC GTC PG0 CAT CTA ..CTG GTC CGA CTC GAG CGA AGO TCT CGA GAO CCT GAG CCCwf 5cr Pro Gis Arg Gis Ala GyPro His Ary Leo Pro Vat Arg His Lcu Leo Val Oln Leo His Arg Arg 5cer Cin Gu Pro His Pro 5618wL 1711 GAO 0CC AOC AGCGCCA ACG CCC TCC CCA GAAI ACC CCC_ AGC CCC TCC TOG TCC CAT TCC TCA GGC CGA CCA CCC AGG TCC COG AGO TCG GAGC/) CIa Oly Ser See Ala Arg Pro 5cer Pro Cmu Ther Pro Arg Pro Cys Trp 5cr His 5cr 5cr Gly Gts Pro Oty Arg 5cr Airg Th 5cSr H is 598
1851 TOO GCC ACC GAG GAG AGA GCA GCC CCC CCC AOC CCC AjGGT CCTI OCT GGA CGOT CCC ACC A/IAvCOAC- GGGTGG A C lOC5 T r p Ala The His His The Ala Ala Pro Pro 5cer IGy ProiCly Prol Ala Cly Arg Pro Thr Lys ?oGtCaAs tei eeS t 628
-l 1891 GAT OTO COP 00/I CCC AIlS PIT GOTO~~~~~CCT CC (C OGA CCG GAG CCC CGA CTC A.AC PAAA TCG CAO(TT'CTG ACC PAT GCC TTC A/IC CTT
1961 GCA GAO CCA GCCC CCC CCC AGO CCC AGO CT7 ACG AGA GAT CIAC OTG AAA GCOT GAG AGO ATC CCC AGC CTG AGO AAG FCT TTC GCC AOL CTCCPin Gini Pro Ala Pro Pro Ar q P r 5cSr Leii Seri GIrin Asp ClI i Vat Lwyi Ala G Ik Thi Ilie Ary Sc r L.iei Argl Lys 5Cci Phc AlIa SirLtkr 6611
207 Tlc rCC GAG TGA CACC-TAC-TG-TOAC("AACCCC2T'AAAIX:CC- T(AG,ACA/ICOC CTCTGt;rO~3,TCCTG,'AGTCCAI'TITC-TCACCTTTTIO.A/I TC'-TCC,-%A/IA'CGC FT-GAGAACCCrC1T CGTC-CT- Phc Sir Asp Soili Full
2159- OC-,T TOTAlA TC(.,`Ac TTrCTOA T rT':," r:AJT,CCT. .AC
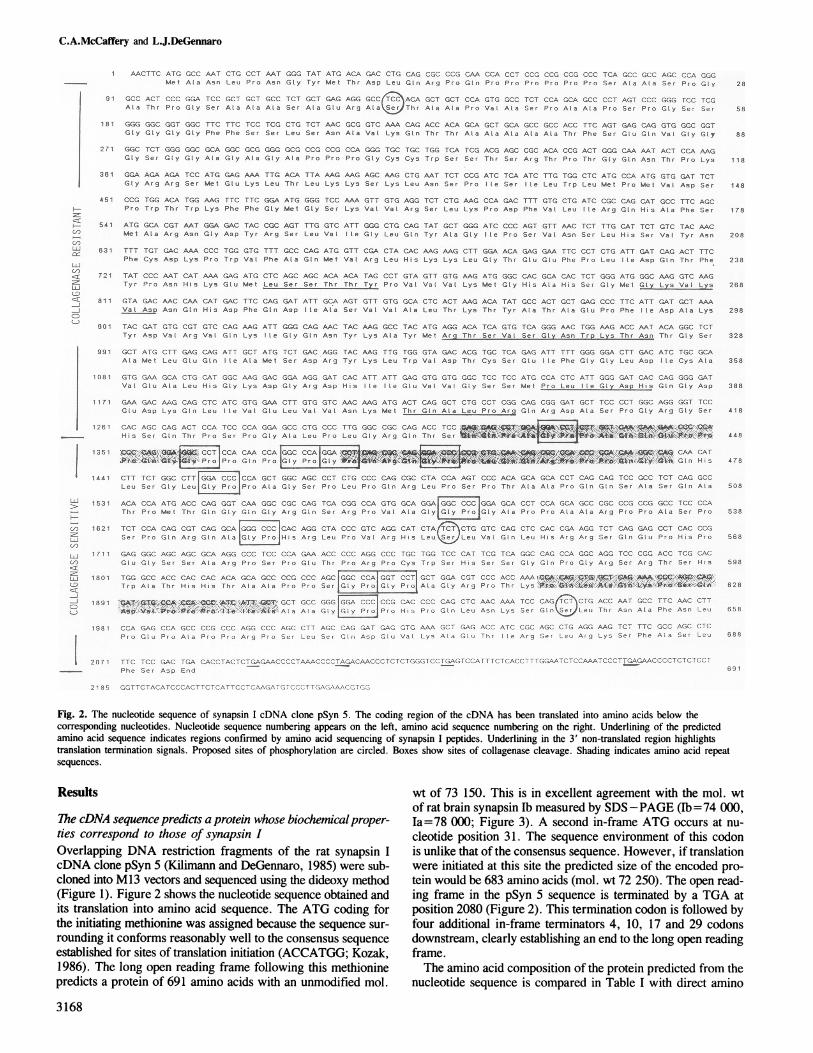
Fig. 2. The nucleotide sequence of synapsin I cDNA clone pSyn 5. The coding region of the cDNA has been translated into amino acids below thecorresponding nucleotides. Nucleotide sequence numbering appears on the left, amino acid sequence numbering on the right. Underlining of the predictedamino acid sequence indicates regions confirmed by amino acid sequencing of synapsin I peptides. Underlining in the 3' non-translated region highlightstranslation termination signals. Proposed sites of phosphorylation are circled. Boxes show sites of collagenase cleavage. Shading indicates amino acid repeatsequences.
Results wt of 73 150. This is in excellent agreement with the mol. wt
7he DNAseqenc prdict a rotin hos biohemcalproer-of rat brain synapsin Ib measured by SDS -PAGE (Ib z=74 000,
ThecDN seuene pedits prtei whse iocemial ropr- la= 78 000; Figure 3). A second in-frame ATG occurs at nu-ties correspond to those of synapsin I cleotide position 31. The sequence environment of this codonOverlapping DNA restriction fragments of the rat synapsin I is unlike that of the consensus sequence. However, if translationcDNA clone pSyn 5 (Kilimann and DeGennaro, 1985) were sub- were initiated at this site the predicted size of the encoded pro-cloned into M13 vectors and sequenced using the dideoxy method tein would be 683 amino acids (mol. wt 72 250). The open read-(Figure 1). Figure 2 shows the nucleotide sequence obtained and ing frame in the pSyn 5 sequence is terminated by a TGA atits translation into amino acid sequence. The ATG coding for position 2080 (Figure 2). This termination codon is followed bythe initiating methionine was assigned because the sequence sur- four additional in-frame terminators 4, 10, 17 and 29 codonsrounding it conforms reasonably well to the consensus sequence downstream, clearly establishing an end to the long open readingestablished for sites of translation initiation (ACCATGG; Kozak, frame.1986). The long open reading frame following this methionine The amino acid composition of the protein predicted from thepredicts a protein of 691 amino acids with an unmodified mol. nucleotide sequence is compared in Table I with direct amino
3168

Synapsin I primary structure
TIME (mins)0 10 20 30 40
105
la1bU_68 o62
52
48 -
ji92-5
66-2
%I 45
Fig. 3. SDS-PAGE of synapsin I collagenase digestion intermediates.Collagenase digestion and gel electrophoresis were carried out as describedin Materials and methods. Mol. wt standards: collagenase: 105 kd,phosphorylase B: 92.5 kd, bovine serum albumin: 66.2 kd and ovalbumin:45 kd.
acid composition data available for bovine synapsin lb (Ueda andGreengard, 1977). The values agree well with the exception ofserine for which the cDNA sequence predicts a considerablyhigher value. We believe this discrepancy may be explained bythe fact that serine content is often underestimated in the deter-mination of amino acid composition due to its lability during pep-tide bond hydrolysis. The difference in serine content, and othersmall variations in amino acid composition between the predictedprotein and literature composition values, may also be the resultof species differences. The predicted composition confirms therelatively high proline content of synapsin lb (13.6%), and ex-plains its basic isoelectric point due to a relatively high contentof basic amino acids (Arg:5.3%; Lys:4.3%; His:3.0%).To confirm the authenticity of the predicted amino acid se-
quence it was compared with amino acid sequence derived frompeptides of synapsin I. Purified rat brain synapsin I was subjectedto cleavage by cyanogen bromide and the resulting peptides wereseparated by reverse-phase h.p.l.c. N-Terminal amino acid se-quence analyses were performed on selected peptide peaks. Allsequences obtained (Figure 2) were found to the carboxyl-ter-minal side of methionine residues in the predicted synapsin I se-quence.
Table I. Comparison of amino acid compositions
Amino acid Amino acid compositionTotal Mol %A B C
Ala 67 9.7 10.4Phe 17 2.5 2.5Lys 30 4.3 4.4Pro 94 13.6 12.4Thr 36 5.2 4.5Cys 6 0.9 1.9Gly 65 9.4 11.1Leu 44 6.4 6.0Glu 20 2.9 11.6aGln 51 7.4 NDVal 34 4.9 5.8Asp 23 3.3 6.7bAsn 17 2.4 NDHis 21 3.0 3.0Met 17 2.5 1.8Arg 37 5.3 4.8Trp 9 1.3 1.8Ile 19 2.7 3.4Ser 73 10.5 6.2Tyr 11 1.6 2.0
Amino acid composition of synapsin Ib. Column A: total number of eachresidue predicted by nucleotide sequence. Column B: mole percent of eachresidue calculated from data in A. Column C: mole percent of each residuefrom amino acid composition determination (Ueda and Greengard, 1977).ND, not detected.aGlu + Gln.bAsp + Asn.
Comparison of codon usage frequency in the synapsin I se-quence with previously compiled codon usage data for rat (Maru-yama et al., 1986) shows no unusual bias, unlike that seen inthe sequences encoding some other proline-rich proteins (Annand Carlson, 1985). Based on the mol. wt and amino acid com-position of the predicted protein we conclude that the sequencerepresents that of synapsin lb.Structural andfunctional predictions ofthe synapsin I sequenceCollagenase sensitivity ofsynapsin I. Synapsin I has been shownto be a substrate for degradation by collagenase (Ueda and Green-gard, 1977). Figure 3 shows that the protein is digested in a step-wise fashion through major mol. wt intermediates of 68, 62 and52 kd to a final collagenase-resistant polypeptide of 48 kd. Elevenpotential sites for collagenase digestion (cleavage at the Y-Glybond in the sequence Y-Gly-Pro) can be found in the predictedsynapsin I sequence (boxes in Figure 2). Proteins with mol. wtssimilar to those of the major collagenase intermediates could resultby preferential collagenase cleavage after residue 639 (67.0 kd),residue 544 (57.4 kd), residue 482 (51.4 kd) and residue 439(47.0 kd). Additional intermediates which appear in smalleramounts during the digestion may be explained by less frequentcleavage after residue 611 (64.7 kd) and residue 524 (55.5 kd).No sites for collagenase digestion occur between amino acids1 and 439. This corresponds well with the size of the collagenase-resistant fragment of synapsin 1 (48 kd measured versus 47.0 kdpredicted). Our data now defines the collagenase-sensitive portionof synapsin I as the carboxyl-terminal third of the molecule (resi-dues 440-691), and the collagenase-resistant portion as the re-maining amino-terminal sequence.Synapsin Iphosphorylation. Synapsin I is known to be a substratefor the cAMP-dependent protein kinase and Ca2+/calmodulin-
3169

C.A.McCaffery and L.J.DeGennaro
tOo 40D
p
GOD
III - - 1111 -4111---1P P
Fig. 4. Hydropathy plot and secondary structure predictions for the synapsin I sequence. Plot was made by the algorithm of Kyte and Doolittle (1982) withhydrophobicity values averaged over nine amino acids. Secondary structure predictions, shown below the hydropathy plot, were made according to Chou andFasman (1978). Predicted (B-structure is indicated by vertical lines, a-helical structure by horizontal lines, spanning the sequence regions involved. Location ofproposed sites of phosphorylation are indicated by a P.
dependent protein kinases I and II (Huttner et al., 1981; Kennedyand Greengard, 1981). These enzymes phosphorylate synapsinI on serine residues. The cAMP-dependent kinase phosphorylatessynapsin I at one site located in the collagenase-resistant portionof the molecule (Huttner et al., 1981). Inspection of the predictedprimary structure reveals three potential recognition sequences
for the cAMP-dependent kinase within this domain (Figure 2).The first is at amino acid residues 40-42. The sequence, Arg-Ala-Ser, has the cAMP-dependent kinase consensus sequence
(Kemp et al., 1975) and serine as the acceptor amino acid. Thissite lies near the amino-terminal end of the collagenase-resistantportion of synapsin I in a region of predicted a-helical secondarystructure (see below).A second site (residues 315-317) lies within a portion of the
synapsin I molecule that is predominantly hydrophobic and pre-dicted to adopt :-like secondary structure. We hypothesize thatthis region of the molecule may participate in the formation ofa folded hydrophobic core (see below), possibly making the siteunavailable for phosphorylation.The third potential cAMP-dependent kinase phosphorylation
site is at position 416-418. It lies in a hydrophilic region locatedimmediately to the carboxyl-terminal side of the proposed hydro-phobic core of synapsin I. The site is bordered by two potentialCa2+/calmodulin-dependent kinase phosphorylation sites whichappear not to be active as substrates for that enzyme (see below).Given the primary and secondary structure predictions discussedabove, and previous peptide mapping data (Huttner et al., 1981;DeGennaro, unpublished observations), we propose that residues40-42 (circled in Figure 2) most likely provide the physiologi-cally relevant site for phosphorylation by the cAMP-dependentprotein kinase. We cannot, however, completely exclude phos-phorylation at the other two sites.The collagenase-resistant domain of synapsin I also contains
a site of phosphorylation by Ca2+/calmodulin-dependent proteinkinase I (Kennedy and Greengard, 1981). Little is known aboutthe substrate specificity of this enzyme. However, the serine itphosphorylates in synapsin I is known to occur in the same tryp-tic/chymotryptic peptide as that phosphorylated by the cAMP-dependent kinase (Huttner et al., 1981). The predicted sequencereveals a number of serine residues, including those proposedas cAMP-dependent kinase phosphorylation sites, which couldbe substrates for Ca2+ kinase I.
Biochemical evidence indicates that Ca2+/calmodulin-depen-dent kinase II phosphorylates synapsin I on at least two siteswithin the collagenase-sensitive portion of the molecule (Kennedyand Greengard, 1981; Huttner et al., 1981). Three potentialCa2+ kinase H recognition sequences have been identified by in-spection of the synapsin I amino acid sequence in that region.They occur at positions 493 -496 (site 1), 552 -555 (site 2) and648-651 (site 3). All three sites have serine as the acceptor amino
3170
acid. Sites 1 and 2 are most homologous to the Ca2+/calmodulin-dependent kinase recognition sequence (Arg-X-X-Ser) determinedby model substrate studies to have the highest activity (Pearsonet al., 1985). Site 3 has the correct form (a basic amino acidthree residues to the amino-terminal side of the acceptor serine)and could also provide a site for phosphorylation by Ca2+ kinaseII.
Results of previous experiipents (Huttner et al., 1981) indicatethat one of the physiologically relevant Ca2+ kinase II phosphor-ylation sites lies within the region of the synapsin I moleculewhich is cleaved by collagenase digestion resulting in the 68-kdintermediate. As discussed above, this cleavage probably occurs
after amino acid 639, and would liberate phosphorylation site3. Likewise, the second Ca2+ kinase II phosphorylation site isknown to be cleaved from the synapsin I molecule by collagenasedigestion resulting in the 62-kd intermediate. Our sequence sug-gests that this intermediate is produced by collagenase cleavageafter amino acid 544, and would liberate phosphorylation site2. The collagenase digestion data therefore indicate that Ca2 +kinase II recognition sites 2 and 3 (circled in Figure 2) are thosephosphorylated in vivo, and that despite the occurrence of a properkinase recognition sequence at site 1, phosphorylation does notnormally occur on serine 496. In this regard it should be notedthat three potential sites for Ca2+ kinase II phosphorylation alsooccur within the collagenase-resistant domain of the synapsin Imolecule (residues 132-135, residues 410-413 and residues432 -435). These potential sites are not phosphorylated in vivoor in vitro by purified Ca2+ kinase II. The latter two sites liein the same region of the synapsin I molecule as one of the poten-tial cAMP-dependent kinase phosphorylation sites.
Hydrophobicity plot and predicted secondary structure ofsynapsin IThe hydrophobicity plot (Figure 4) of the synapsin I sequence,generated by the method of Kyte and Doolittle (1982), revealsthree general domains in the protein molecule. The amino-ter-minal domain of the protein (residues 1-137) is relatively non-
descript with regard to hydrophobic or hydrophilic character.Four separate regions, comprising -40% of this domain, are
predicted by the method of Chou and Fasman (1978) to havethe potential to form oa-helix. The proposed site for cAMP-dependent kinase phosphorylation (serine 42) lies within the sec-ond of these a-helical regions (Figure 4). Not shown in the hydro-phobicity plot, due to the method of averaging employed, is thefact that the first nine amino-terminal residues of the moleculeare hydrophobic (average hydropathy = 0.53). This may indicatethat the amino-terminal end of synapsin I is sequestered fromthe aqueous environment.The central domain of the protein, amino acids 137-410, con-
tains the majority of the hydrophobic residues of the synapsin
Hydropathy
SecondaryStructure
Pebic
Phir

Synapsin I primary structure
A Q K P S D V P P P I A1*be181* } I *1' 1
P P G 6 G
-I P P L QG R
Fig. 5. Comparison of the internal amino acid repeat sequences of synapsinI. Numbers indicating the beginning and end of each sequence are shown on
the left and right, respectively. Shading indicates the region of greatestsequence homology. Asterisks mark amino acids shared by all threesequences. Vertical lines indicate same-residue conservation, dots indicateconservative amino acid substitutions (Dayhoff et al., 1978).
I sequence. Secondary structure prediction indicates that - 40%of this region has the potential to form repeating stretches offl-structure (Figure 4). These regions are separated by shortstretches of a-helix, random coil and,B-turn. By analogy withother proteins which exhibit regions of,-structure, such as myelinbasic protein (Martenson, 1986), we propose that this centraldomain may organize into relatively stable fl-sheet and form a
compact, hydrophobic, globular core. This would result in theremoval of a maximum of non-polar surface area from the aque-
ous milieu.The carboxyl-terminal domain of synapsin I (residues 410-
691) contains the collagenase-sensitive region of the moleculeand is notably hydrophilic for most of its length. Previous studieshave suggested that this domain adopts an elongated conformation(Ueda and Greengard, 1977; Schiebler et al., 1986). Secondarystructure predictions from our sequence indicate that the majorityof this region would have a tendency to adopt fl-turn structure.
This may be explained in part by the large number of prolineand glycine residues (32.5%). Two exceptions to this rule are
the regions containing the predicted sites for Ca2+ kinase II
phosphorylation which are both associated with small segments
of f-structure (Figure 4).
Amino acid sequence repeats occur in the synapsin I sequence
Figure 5 shows the primary structure of three similar amino acidsequences repeated in the synapsin I molecule. The shaded se-
quences in the right half of the figure (also shaded in Figure 2)display the greatest homology. In this region 35% of the aminoacids occur in the same position in all three sequences. Compari-son of repeat IH with repeat I, or repeat HI with repeat II, reveals41 % same-residue conservation in this region. Repeats I and Hdisplay 65 % exact conservation. When conservative amino acidsubstitutions are considered (Dayhoff et al., 1978) the homologyin the shaded region becomes 59% (III/I), 47% (HI/H) and 65%(Ill), respectively.The homologous nature of the repeats extends beyond this
region of highest similarity. Thus, over the entire illustrated se-
quence, same-residue conservation between the repeats is 39%(II/I), 33% (mI/H) and 39% (I/H). Taking conservative substi-tutions into account the homologies are 67% (III/I), 50% (III/H1)and 56% (Ill). The amino acid sequence homology is not carriedover to homology at the DNA base sequence level.
Homology between synapsin I and other proteins
Computer-assisted comparisons (George et al., 1986) of thededuced synapsin I amino acid sequence with those of other pro-
teins revealed slight homology to a class of proline-rich phospho-proteins synthesized in the salivary gland (Ann and Carlson,1985), and to regions of the collagen alpha 1 and alpha 2 chains.However, these homologies appear to be a consequence of theproline-rich composition of the respective proteins. A comparisonof greater interest is that between synapsin I and the cytoskeletalprotein 4.1. Experiments reported by Baines and Bennett (1985)have indicated that synapsin I is related at the level of immuno-
PROFILIN 20 A I V G Y K D S P S V WAAVPGKTFVN T P AEVG L V G KDRS 56
I III* 11. . -SYNAPSIN 399 V N K M TQ A L P R Q R D A S P G R G S H S Q T P S P G A L P L G R Q T S 435
I .. I I . I .. . -VILLIN 22 V N T A A E D L P R GV D PSRKENHL I D E DFKA V F G M T R S A F 58
Fig. 6. Region of potential homology between rat synapsin I, bovine profilinand chicken villin. Numbers indicate residue positions within the respectiveamino acid sequence. Vertical lines indicate same-residue conservation, dotsindicate conservative amino acid substitutions.
logical cross-reactivity and spectrin-binding activity to protein4.1. Our sequence data do not support homology between theproteins beyond these indirect observations. The amino acid com-position of rat brain synapsin I and human erythrocyte protein4.1 (Tyler et al., 1980) are very different. Synapsin I does notdisplay clustered cysteine residues, nor do our data support thedivision of synapsin I into acidic and basic domains, both of whichare defining characteristics of protein 4.1 (Leto and Marchesi,1984). Finally, a comparison between the deduced synapsin Isequence and the sequence of a 67 amino acid peptide of human.erythrocyte protein 4.1 previously shown to be involved in pro-moting spectrin-actin associations (Correas et al., 1986a,b),reveals no significant homology. We cannot exclude that as yetunsequenced regions of protein 4.1 might display meaningfulhomology with synapsin I.
Prompted by the possible functional similarity between synapsinI and protein 4.1, we compared the synapsin I amino acid se-quence with the sequences of known actin-binding proteins. Fig-ure 6 shows a region of synapsin I that displays some homologyto sequences of bovine profilin (Nystrom et al., 1979) and thecarboxyl-terminal domain of chicken villin (Glenny et al., 1981).Exact amino acid position conservation is small: 21 % with pro-filin and 24% with villin. However when conservative aminoacid substitutions are considered the homology becomes 43 % and49%, respectively. The fact that both actin-binding proteins sharehomology with the same region of synapsin I may be indicativeof domains involved in similar functions. The illustrated se-quences of both profilin and villin are part of larger sequencescontaining the actin-binding domain of each protein. In addition,some of the amino acids common between synapsin I and villin(Asp-41 1, Ser-413 and Ser420) are thought to be involved incalcium binding by villin (Hesterberg et al., 1983). This segmentof the synapsin I molecule (residues 399 -435) lies within thehydrophilic sequence immediately bordering the carboxyl-ter-minal side of the ,3-structure-forming region discussed above,and overlaps partially with the first of three amino acid repeatunits found in the synapsin I molecule (residues 421-456, Figure5).
DiscussionIn this report we have presented the nucleotide and predictedamino acid sequence of a cDNA clone representing rat brainsynapsin I (Figure 2). The open reading frame is initiated at asequence similar to the established translation initiation consensussequence, and is terminated by a TGA termination codon. Thepredicted protein is proposed to be synapsin Ib on the basis ofits molecular size, amino acid composition and agreement withamino acid sequence analysis of synapsin I peptides. The aminoterminus of synapsin I is known not to be amenable to Edmandegradation (DeGennaro, unpublished observations). In thisregard the amino-terminal residue exposed after removal of theinitiator methionine is predicted by the cDNA sequence to bealanine. Alanine often undergoes acetylation which leads to sucha block (Tsunasawa and Sakiyama, 1984). Nevertheless, the ac-
3171

C.A.McCaffery and L.J.DeGennaro
tual start site of translation may occur 5' to the sequence deter-mined. Given the excellent agreement between the predicted andmeasured synapsin I mol. wts, however, any additional aminoacid sequence must be relatively short.The biochemical difference between synapsin Ib and synapsin
Ia remains an open question. Both proteins exhibit nearly ident-ical peptide maps and undergo phosphorylation at identical sites.Rat synapsin I is encoded by a single gene in the haploid gen-ome (D.P.Carroll, and L.J.DeGennaro, in preparation). ClonedcDNA molecules hybridize with two bands in Northern blotanalysis of RNA from 14 day old rat brain (Kilimann and De-Gennaro, 1985), but only a single band in RNA from adult ratbrain (Haas and DeGennaro, unpublished observations). There-fore the difference between synapsin Ia and lb at the polypeptidelevel is probably generated during RNA processing. Previousdata indicate that this difference occurs in the collagenase-sensi-tive region of the molecule (Huttner et al., 1981). Based on oursequence data, and these previous observations, we propose thatsynapsin Ia may differ from synapsin Ib only at its carboxyl-ter-minal end; the difference being --40 additional amino acids. Thefunctional significance of this difference remains unexplained.
All of the known biochemical and structural characteristics ofsynapsin I can be explained by inspection of the predicted aminoacid sequence. The clustering of hydrophobic amino acids and,1-structure-forming residues in a central domain of the proteinsuggests the possible formation of a globular core composed of,B-sheet. The collagenase-resistant domain of the molecule is nowdefined as the amino-terminal 439 amino acids. Each of the col-lagenase digestion intermediates of synapsin I can be explainedby the location of amino acid digestion sites in the remainingcarboxyl portion of the molecule (residues 440-691). The mostlikely sites for phosphorylation by the cAMP-dependent andCa2+/calmodulin-dependent protein kinases have been identified.A number of apparently cryptic sites for both enzymes are alsopresent in the sequence.Although experiments of others (Baines and Bennett, 1985;
Goodman and Zagon, 1986) have suggested some similarity be-tween synapsin I and the erythrocyte cytoskeletal protein 4.1,our results indicate that any primary structure homology betweenthese proteins must be limited. The question of functional hom-ology remains open. The fact that synapsin I seems to containsequences related to some actin-binding proteins (Figure 6) mayhelp to explain the limited cross-reactivity of protein 4.1 anti-bodies with synapsin I. It may also continue to point to an inter-action between synapsin I and the cytoskeleton of the presynap-tic nerve terminal.Among the most interesting features of the predicted synapsin
I primary structure is the occurrence of three amino acid sequencerepeats, 36 amino acids in length (Figure 5). These repeats liein the collagenase-sensitive domain of the molecule. In this regionthey flank the sites we propose to be phosphorylated in vivo byCa2+/calmodulin-dependent kinase II. Previous experiments haveshown that the collagenase-sensitive domain of synapsin I is theregion of the molecule through which interactions with synapticvesicles are mediated (Ueda, 1981; Huttner et al., 1983; Schiebleret al., 1986). These interactions are thought to be of an electro-static nature, and phosphorylation of sites in this domain by Ca2+kinase II has been shown to reduce the tendency of synapsin Ito remain associated with synaptic vesicles in vitro (Huttner etal., 1983; Schiebler et al., 1986). The location of the synapsinIamino acid repeats, and their proximity to the Ca22+ kinase IIphosphorylation sites, make them prime candidates for involve-ment in synapsin I interactions with the binding domain of synap-
3172
tic vesicles. It would be of interest to investigate the role of theserepeats in synapsin I function. To this end, our amino acid se-quence predicts that cleavage of synapsin I by cyanogen bromideshould produce two peptides containing these repeat sequences.These peptides could be tested for vesicle-binding activity, andantibodies raised against them might be used as tools to probesynapsin I function. Ultimately, through site-directed mutagenicalterations of the cDNA sequences, specific amino acid changescan be introduced into the repeat sequences. The vesicle-bindingability of the altered synapsin I protein could then be tested. Thesame procedure might be used to assess the involvement of Ca2 +kinase II phosphorylation sites, present in the collagenase-sensi-tive domain, in the regulation of synapsin I -vesicle interactions.
Materials and methodscDNA clones and DNA sequencingSynapsin I cDNA clone pSyn 5 was isolated from a rat brain cDNA libraryprepared with poly(A)+ RNA enriched with synapsin I mRNA (Kilimann andDeGennaro, 1985). DNA restriction fragments of this cDNA were subclonedin M 13 vectors (Messing et al., 1981), and were sequenced by the dideoxy chaintermination technique (Sanger et al., 1977) essentially as described by Messing(1983). DNA sequencing was primed using the M13 universal primer.Amino acid sequencing of synapsin IRat brain synapsin I was purified by a modification of the original procedure(Ueda and Greengard, 1977). Briefly, the supernatant of the pH 6 step was ad-justed to pH 8, adsorbed to a carboxymethyl-cellulose column, and synapsinI was eluted with a gradient of 0-0.2 M NH4CI. Peak fractions were pooled,adsorbed to an hydroxylapatite column and eluted with 0.5 M NH4Cl. The finalstep of purification was molecular sieve chromatography through a column ofSephadex G-100. Purified synapsin I (2 mg) was cleaved by cyanogen bromidein 70% formic acid. The resulting peptides were separated by h.p.l.c. on a C18column using 0.05% trifluoroacetic acid (TFA) and 0.05% TFA in acetonitrileas the elution solvents. N-Terminal amino acid analyses were performed on anApplied Biosystems gas phase sequencer 470 A. The phenylthiohydantoin aminoacid derivatives were analyzed by an h.p.l.c. system which separates all com-ponents isocratically (Lottspeich, 1985).Collagenase digestion of rat brain synapsin IRat brain synapsin I was prepared as described above. Synapsin I (100 isg) wascombined with collagenase (100 units, Biofactures Corp., Form III) in 20 mMTris-HCl, pH 7.4, 10 mM CaCl2. Incubation was at 30°C. Aliquots were with-drawn from the reaction at 0, 10, 20, 30 and 40 min, mixed with SDS sampleapplication buffer (Huttner etal., 1981), and boiled immediately for 5 min. SDS-PAGE was according to Laemmli (1970). After electrophoresis the gel was fixedin 50% methanol, 10% trichloroacetic acid (20 min), stained in 25% methanol,10% trichloroacetic acid, 0.2% Coomassie blue (2 h) and destained in 10% aceticacid.
AcknowledgementsThe authors wish to thank Drs Correas, Speicher and Marchesi for sharing theirsequence data before publication. We thank Dr M.Schleicher-Noegel for helpwith the purification of synapsin I peptides, Dr F.Lottspeich for carrying out theN-terminal amino acid sequence analyses, and Eva Erhard-Ott for expert technicalassistance. Thanks are also due to Drs Carroll, Barde and Lottspeich, and MsKirchgessner for critical reading of the manuscript, and Dr Lottspeich for helpwith computer search. C.A.M. was supported by the Deutscher AkademischerAustauschdienst.
ReferencesAnn,D.K. and Carlson,D.M. (1985) J. Biol. Chem., 260, 15863-15872.Baines,A.J. and Bennett,V. (1985) Nature, 315, 410-413.Chou,P.Y. and Fasman,G.D. (1978) Adv. Enzymol., 47, 45-148.Correas,I., Leto,T.L., Speicher,D.W. and Marchesi,V. (1986a) J. Biol. Chem.,
261, 3310-3315.Correas,I., Speicher,D.W. and Marchesi,V. (1986b) J. Biol. Chem., in press.Dayhoff,M.O., Schwartz,R.M. and Orcott,B.L. (1978) In Dayhoff,M.O. (ed.),
Atlas ofProtein Sequence and Structure. National Biomedical Research Foun-dation, Washington, DC, Vol. 5, Suppl. 3, pp. 359-362.
DeCamilli,P., Cameron,R. and Greengard,P. (1983a) J. Cell Biol., 96, 1337-1354.
DeCamilli,P., Harris,S.M., Huttner,W.B. and Greengard,P. (1983b) J. Cell Biol.,96, 1209-1211.

Synapsin I primary structure
George,D.G., Barker,W.C. and Hunt,LT. (1986) Nucleic Acids Res., 14, 11 - 15.Glenny,J.R., Jr., Geisler,N., Kaulfus,P. and Weber,K. (1981) J. Biol. Chem.,
256, 8156-8161.Goodman,S.R. and Zagon,I.S. (1986) Am. J. Physiol., 250, c347-c360.Hesterberg,L.K. and Weber,K. (1983) J. Biol. Chem., 258, 365-369.Huttner,W.B. and Greengard,P. (1979) Proc. Natl. Acad. Sci. USA, 76, 5402-
5406.Huttner,W.B., DeGennaro,L.J. and Greengard,P.(1981) J. Biol. Chem., 256,
1482-1488.Huttner,W.B., Schiebler,W., Greengard,P. and DeCamilli,P. (1983) J. Cell Biol.,%, 1374-1388.
Kennedy,M.B. and Greengard,P. (1981) Proc. Natl. Acad. Sci. USA, 78, 1293-1297.
Kemp,B.E., Bylund,D.B., Huang,T.S. and Krebs,E.G. (1975) Proc. Natl. Acad.Sci. USA, 72, 3448-3452.
Kilimann,M.W. and DeGennaro,L.J. (1985) EMBO J., 4, 1997-2002.Kozak,M. (1986) Cell, 44, 283-292.Kyte,J. and Doolittle,R.L. (1982) J. Mol. Biol., 157, 105-132.Laemmli,U.K. (1970) Nature, 227, 680-685.Leto,T.L. and Marchesi,V. (1984) J. Biol. Chem., 259, 4603-4608.Lottspeich,F. (1985) J. Chromatogr., 326, 321-327.Messing,J. (1983) Methods Enzymol., 101, 20-78.Messing,J., Crea,R. and Seeburg,P.H. (1981) Nucleic Acids Res., 9, 309-322.Martenson,R.E. (1986) J. Neurochem., 46, 1612-1622.Maruyama,T., Gojobori,T., Aota,S. and Ikemura,T. (1986) Nucleic Acids Res.,
14, r151-r197.Navone,F., Greengard,P. and DeCamilli,P. (1984) Science, 226, 1209-1211.Nestler,E.J. and Greengard,P. (1984) Protein Phosphorylation in the Nervous
System. Wiley, NY.Nystrom,L.E., Lindberg,U., Kendrick-Jones,J. and Jakes,R. (1979) FEBS Len.,
101, 161-165.Pearson,R.B., Woodgett,J.R., Cohen,P. and Kemp,B.E. (1985) J. Biol. Chem.,
260, 14471-14476.Sanger,F., Nicklen,S. and Coulson,A.R. (1977) Proc. Natl. Acad. Sci. USA,
74, 5463-5467.Schiebler,W., Jahn,R., Doucet,J.-P., Rothlein,J. and Greengard,P. (1986) J. Biol.
Chem., 261, 8383-8390.Tyler,J.M., Reinhardt,B.N. and Branton,D. (1980) J. Biol. Chem., 255, 7034-
7039.Tsunasawa,S. and Sakiyama,F. (1984) Methods Enzymol., 106, 165-171.Ueda,T. (1981) J. Neurochem., 36, 297-300.Ueda,T. and Greengard,P. (1977) J. Biol. Chem., 252, 5155-5163.
Received on 14 July 1986; revised on I September 1986
Note added in proofWe have sequenced a second synapsin I cDNA clone, pSyn 16 (Kilimann andDeGennaro, 1985) that overlaps with and is homologous to pSyn 5 from nucleotide1733 to 2234, except for a 44 base in-frame deletion between nucleotides 1943and 1986. The sequence suggests the existence of synapsin I mRNA and proteinvariants. Further investigation of synapsin I gene expression and function willbe required for a full understanding of such variants.
3173