degree antibody-independent activation classical ... · classical complement pathway. kl strains of...
TRANSCRIPT

Vol. 43, No. 2INFECTION AND IMMUNITY, Feb. 1984, p. 684-6920019-9567/84/020684-09$02.00/0Copyright © 1984, American Society for Microbiology
Degree of Antibody-Independent Activation of the ClassicalComplement Pathway by Ki Escherichia coli Differs with 0 Antigen
Type and Correlates with Virulence of Meningitis in NewbornsGERD PLUSCHKEt* AND MARK ACHTMAN
Max-Planck-Institut fir molekulare Genetik, D-1000 Berlin 33, Federal Republic of Germany
Received 27 September 1983/Accepted 10 November 1983
A total of 95 Kl Escherichia coli strains of the 0 (lipopolysaccharide) serotypes 01, 07, or 018 had beenanalyzed previously for the ability to cause bacteremia after colonizing the gut of newborn rats. In thisstudy, these strains were tested for their resistance to the bactericidal activity of rat serum. All strains thathad caused bacteremia in a high percentage of the inoculated rats were able to survive for several hours in90% adult rat serum. With only a few exceptions, 07:K1 and 018:K1 strains were serum resistant andvirulent, whereas 01:K1 strains were serum sensitive and avirulent. Serum sensitivity was due to theclassical complement pathway. Kl strains of all three 0 serotypes were resistant to the alternativecomplement pathway. 07:K1 and 018:K1 cells were killed efficiently after the classical pathway wastriggered by specific antilipopolysaccharide antibodies. However, killing of 01:K1 bacteria by the classicalpathway system did not require antibodies. Isolated 01-lipopolysaccharide fixed complement moreefficiently than did isolated 07- or 018-lipopolysaccharide, suggesting that the differences in the chemicalstructure of the 0 antigens are responsible for the observed differences in complement sensitivity. Incombination with epidemiological data, the results indicate that antibody-independent classical pathwayactivation provides an important defense mechanism for newborns against certain gram-negative infections.
Escherichia coli strains of the Ki capsular serotype repre-sent 80% of all E. coli strains isolated from cases ofmeningitis in newborns (23, 25). They are also frequentlyisolated from feces of healthy individuals as well as fromcases of urinary tract infections and septicemia (data sum-marized in references 22 and 23). Most of these Kl isolatesbelong to a limited spectrum of O-lipopolysaccharide (LPS)antigen types (18). In three previous studies (1, 12, 22), wehave described biochemical and virulence properties of alarge number of different Kl strains of the common LPStypes 01, 07, and 018. These strains had been obtainedfrom different sources and had been isolated from diseasedand healthy individuals. It appeared that most of thesestrains belong to a limited number of clones spread overEurope and the United States. One predominant clone wasdetected among 07:K1 strains, two subclones were detectedamong 018:K1 strains, and two unrelated clones weredetected among 01:K1 isolates (1, 12).
Distinct differences were observed between these groupswhen they were tested for the ability to cause bacteremiaafter colonizing the gut of newborn rats. Although most ofthe 018:K1 and 07:K1 strains were virulent, bacteremia wasonly rarely caused by 01:K1 bacteria, irrespective of theclonal descent-(22). A more detailed analysis of the differentsteps of the infection process showed that avirulent 01:K1and virulent 07:K1 and 018:K1 strains colonize the gut andtranslocate to the mesenteric lymph nodes with comparableefficiency. Further experiments suggested that the strainsthat caused bacteremia were able to multiply directly in thebloodstream of the infected animals, whereas the 01:K1bacteria were incapable of such multiplication (22).An analysis of published epidemiological data (Table 4 in
reference 22) revealed that these serotypes were all common
* Corresponding author.t Present address: Basel Institute for Immunology, CH-4005
Basel 5, Switzerland.
among fecal isolates. However, similar to their differingvirulence in the rat model, 07:K1 and 018:K1 bacteria arecommon and O1:K1 bacteria are relatively rare amongisolates from cases of meningitis in newborn children (22).Kl strains are less common among E. coli strains isolatedfrom cases of adult septicemia, and a higher proportion ofthese Kl isolates carry the 01 antigen. 018:K1 bacteria arerare and 01:K1 (and 07:K1) bacteria are common amongisolates from pyelonephritis (12). These groups of Kl bacte-ria thus seem to differ in the type of disease they cause ratherthan to represent virulent versus avirulent isolates.Due to the differing ability of 01:K1, 07:K1, and 018:K1
bacteria to cause meningitis in newborns and to multiply inthe bloodstream of newborn rats, experiments were initiatedto determine whether these differences in virulence correlatewith differences in serum sensitivity. It is commonly as-sumed that resistance to being killed by the serum comple-ment system is an important virulence determinant of inva-sive gram-negative bacteria (for a review see reference 27).For example, complement-dependent bactericidal activity isapparently critical for the defense of the host against inva-sive Neisseria species (16). There are, however, only limiteddata available about the relative importance of antibodies,complement, and phagocytes for host defense in many otherinvasive bacterial diseases.The bactericidal effects of serum are mediated by the
activated components of either the classical or the alterna-tive complement pathway. Both pathways lead to the gener-ation of the membrane attack complex (MAC), which isresponsible for membrane alterations that culminate in celldeath. The alternative pathway has been shown to beactivated by a number of bacterial organisms in the completeabsence of antibodies (24, 27) and can thus provide thenonimmune host with a natural defense against certainmicrobial infections. Whereas the classical pathway is typi-cally activated by immune complexes, it has been shownrecently that antibody-independent activation of the classi-
684
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERUM SENSITIVITY OF E. COLI 685
cal pathway by microorganisms can occur (5). However, ithas been suggested that this type of activation, is relevantonly for killing of rough rather than smooth strains (27), andthus the biological significance of non-immundlogical classi-cal pathway activation for host defense remained unclear.One mechanismn of serum resistance seems to be due to a
failure of the amphiphilic MAC to integrate into hydrophobicdomains on the bacterial cell surface (8, 9). Both the capsuleand the 0 antigen seem to unspecifically reduce the sensitiv-ity of the bacteria to the MAC (21). In addition, the Klcapsule and certain 0 antigens seem to interfere with anti-body-independent complement activation (21).
MATERIALS AND METHODSBacterial strains. The E. coli strains used here have been
described elsewhere (1, 12, 22) in detail. Standard andexceptional strains are listed in Table 1. The followingstrains (referred to by the sequential numbers used in Table 1of reference 1) had formerly been tested in the rat bacteremiamodel (22) and were tested for serum sensitivity: O1:K1strains 1, 2, 4, 5, 8-13, 20-22, 26, 28-30, 32, 35-44, 46, 52-55, and 58; 07:K1 strains 152-155, 157-172, 174, 178, 180,181, 189-191; and 018:K1 strains 64-66, 68-78, 80, 81, 83,87, 91, 92, 94, 96-99, 101-103, 105-108, and 117.
Sera. Sera were prepared by allowing freshly drawn bloodto clot for 1 h at room temperature, followed by 3 h at 4°Cand low-speed centrifugation. Small samples were stored at-70°C until use. Inactivated serum was obtained by heatingfor 30 mim at 56°C. Classical pathway activity was selectivelyinhibited by treatment of serum with 10 mM ethylene glycol-bis(,-aminoethyl ether)-N,N'-tetraacetic acid (EGTA) and 5mM MgC12.
TABLE 1. Partial list of bacterial strainsMem- % Serum
Straina erotypebrane Bacte- sens-uearmStran' eroypeprotein remic ses-Rmrs
pteb rtc tivity,
Standard strains41 01:K1 9 0 S165 07:K1 3 42 R64 018:K1 9 51 R
Clonal exceptions22 01:K1 3 0 S Unique LPS58 01:K1 6 0 S180 07:K1 23 28 R190 07:K1 16 20 R191 07:K1 24 25 I Unique LPS117 018:K1 10 17 R Unique LPS
Exceptions in serumsensitivity orresistance
19 01:K1 5 11 I108 018:K1 6 7 S Low level of
capsulea Numbers refer to the sequential numbers given in Table 1 of
reference 1.b Major outer membrane protein pattern on SDS-PAGE as de-
fined in reference 1.c Three days after feeding 4 x 10' bacteria to 5- to 7-day-old rats.
Data are from reference 22.d R, Resistant; S, sensitive; I, intermediate.These refer to exceptions in LPS migration patterns and differ-
ences in the amount of capsule produced. Data are taken fromreference 12.
Pooled normal rat serum (N-RS) was obtained from 100specific pathogen-free outbred Wistar adult rats obtainedfrom the Bundesgesundheitsamt, Berlin. Serum from germ-free BDIX rats (GF-RS) was a kind gift from N. C. Juhr(Zentrale Tierlaboratorien der Freien Universitat, Berlin).Immune rat sera (I-RS) were obtained from rats immunizedwith boiled bacteria of strains 41, 165, or 64 and are referredto as O1-I-RS, 07-I-RS, and 018-I-RS, respectively. Forimmunization E. coli cells were grown in L broth to a celldensity of 2 x 108/ml, harvested by centrifugation, washedwith and suspended in phosphate-buffered saline (PBS), andboiled for 2 h. The boiled cells were centrifuged and resus-pended in PBS to a cell density of 2 x 109/ml. For eachspecific antiserum, three 6-week-old rats were given six 0.2-ml injections of boiled bacteria every 3 days via a tail vein.Blood was collected 5 days after the last injection. The serafrom each group of three animals were pooled. Control seraobtained from untreated rats did not differ in their bactericid-al activities from N-RS.Media and buffers. L broth contained 10 g of tryptone, 5 g
of yeast extract, 10 g of NaCl, and 0.2 g ofNaOH per liter ofwater. PBS contained 8.5 g of NaCl, 0.8 g of Na2HPO4, and0.145 g of KH2PO4 per liter of water (pH 7.4). DGVB2+contained o.i5 mM CaCl2, 0.5 mM MgCI2, 75 mM NaCl, 2.5mM Veronal buffer, 0.1% gelatin, and 2.5% glucose (pH 7.5).Enzyme-linked immunosorbent assay (ELISA) buffer wasPBS containing 20 mM MgCl2. ELISA substrate contained0.01% o-phenylenediamine dihydrochloride and 0.003%H202 per liter of water. Transfer buffer contained 20 mMTris base, 150 mM glycine, and 20% methanol. Blottingbuffer was PBS containing 0.05% Tween 20. Staining buffercontained 29.5 g of sodium barbital and 19.5 g of sodiumacetate trihydrate per liter of water (pH 10). Staining solu-tion (Milan Blake, Rockefeller University, personal commu-nication) was staining buffer containing 50 mM MgCl2,0.01% Nitro Blue Tetrazolium (Sigma Chemical Co., St.Louis, Mo.), and 0.005% 5-bromo-4-chloro-3-indolyl phos-phate (Sigma).LPS preparation. LPS was extracted with hot phenol
water by the method of Westphal and Jann (28) from the K-strains A691 (018-LPS), A487 (07-LPS), and A26 (01/9-LPS) and from strain no. 9 (O1/5-LPS). Sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (12)of these preparations showed that almost all of the lipid A-core molecules were substituted with smooth long-chainedoligosaccharides. O1/5-LPS yields a doublet band pattern onSDS-PAGE, whereas O1/9-LPS yields a singlet band pattern(12).ELISA. Microtiter plates (code no. 655101; Greiner, Nur-
tingen, Federal Republic of Germany) were sequentiallysubjected to: (i) overnight incubation at 37°C with ELISAbuffer containing 5 jig of LPS per ml, (ii) three washes withELISA buffer, (iii) incubation with ELISA buffer containing0.5% bovine serum albumin for 1 h at room temperature, (iv)three washes with ELISA buffer, (v) incubation with serialdilutions of rat serum in ELISA buffer containing 0.5%bovine serum albumin for 4 h at room temperature, (vi) threewashes with ELISA buffer, (vii) incubation with goat anti-ratimmunoglobulin (Ig) G or anti-rat IgM serum (dilution factor1:500; Nordic Immunological Laboratories, Tilburg, Nether-lands) at room temperature overnight, (viii) three washeswith ELISA buffer, (ix) incubation with conjugated rabbitanti-goat IgG antibodies (IgG fraction of antiserum, peroxi-dase conjugated, dilution factor 1:1,000; Sigma) for 4 h atroom temperature, (x) five washes with ELISA buffer, and(xi) incubation with ELISA substrate for 30 min at room
VOL. 43, 1984
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

686 PLUSCHKE AND ACHTMAN
temperature. A 25-,u amount of 9 N sulfuric acid was addedto stop the reaction. The optical density was measured byusing a Microelisa Autoreader MR 580 (Dynatech, Plochin-gen, Federal Republic of Germany) at a wavelength of 490nm and by using as control a well treated identically exceptthat no rat serum had been added.
Blotting procedure. Crude membrane fractions were ob-tained by sonication (80 s at 50% cycle, Branson celldisrupter B-30; Branson Sonic Power Co., Danbury, Conn.)from bacteria grown to mid-exponential phase in L broth.Unbroken cells were removed by low-speed centrifugation,and the membranes were pelleted by high-speed centrifuga-tion as described previously (1). SDS-PAGE was carried outas described previously (12) by using a discontinuous gelsystem in slab gels containing 4 M urea and an exponentialgradient of acrylamide (7.5 to 25% [wt/vol]) and bis-acryl-amide (0.2 to 0.67% [wt/vol]). The gels were subjected toelectrophoretic transfer with a TE42 Transphor Electropho-resis Unit (Hoefer Scientific Instruments, San Francisco,Calif.). For electrophoretic transfer, a sandwich was con-structed consisting of a perforated polyvinyl chloride sheet,a Dacron sponge, three layers of filter paper, a nitrocellulosefilter membrane (NCM) (BA 85; 0.45 Rm; Schleicher andSchiill, Federal Republic of Germany), and a second layer offilter paper, sponge, and polyvinyl chloride sheet. Transferwas accomplished in a chamber filled with transfer buffer atca. 6 V/cm (1 A) for 2 h.
After transfer, NCM strips at room temperature weresequentially subjected to: (i) incubation with blotting buffercontaining 1% casein for 1 h, (ii) incubation with diluted ratserum for 2 h, (iii) three washes with blotting buffer, (iv)incubation with conjugated goat anti-rat IgG antibodies (IgGfraction of antiserum, alkaline phosphatase conjugated, dilu-tion factor 1:1,000; Sigma) for 2 h, (v) five washes withblotting buffer, and (vi) one final wash with staining buffer.The NCM strips were stained by incubation at 37°C instaining solution for 2 h.
Detection of precipitating antibodies by bacterial agglutina-tion. Bacteria grown overnight in L broth were incubated ina boiling water bath for 2 h. A 0.2-ml portion of thesuspension was mixed in a glass test tube with 0.2 ml ofserial dilutions of antiserum in 0.15 M NaCl. The sealedtubes were incubated for 20 h at 50°C and examined foragglutination. Agglutinated material should remain as asuspension of particles when the tube is agitated.
Bactericidal assay. Bacteria were growth in L broth at 37°Cto a density of 108 cells per ml, washed with and diluted inPBS at room temperature, and incubated in 90% serum at37°C. The viable cell counts were determined by platingserial dilutions of the reaction mixtures on L broth agarplates. In some experiments monoclonal rat IgM specific forthe 018 antigen (21; G. Pluschke and M. Achtman, manu-script in preparation) was added to the assay mixture (finalconcentration 20 Rg per ml). The killing experiments wereperformed at least three times (see Fig. 1, 2, 3, and 5). Onlyminor differences in the time course of killing were observedin the different experiments.IComplement fixation assays. A 90-p.l portion of serum wasmixed with 10 RI of isolated LPS suspended in distilledwater. After incubation of the mixtures for 1 h at 37°C,residual complement activity was determined as describedpreviously (13) by hemolysis of sheep erythrocytes sensi-tized with rabbit anti-sheep erythrocyte serum. Briefly, theserum dilution necessary to yield 50% hemolysis in DGVB2"with 1.25 x 107 erythrocytes in a total volume of 0.25 mlwithin 1 h of incubation at 37°C was determined. Hem(vlytic
units are defined as the volume of undiluted serum calculat-ed to be sufficient to lyse 0.625 x 107 erythrocytes underthese conditions. The data on complement fixation arepresented as the percentage of activity lost relative to acontrol serum incubated without bacteria.
RESULTS
Sensitivity of Kl E. coli strains to N-RS. The sensitivity ofKl E. coli strains to the bactericidal activity of 90% N-RSwas analyzed by determining the changes in the numbers ofviable cells. The 07:K1 bacteria (Fig. 1, curve C) and the018:K1 bacteria (curve B) were able to multiply, but the01:K1 cells were killed after a delay of about 1 h (curve E).Resistance of 01:K1 bacteria to heat-inactivated serum(curve A) indicated that heat-labile complement componentswere required for killing. Chelation of Ca2+ ions by additionof MgEGTA also abolished the bactericidal effect (curve D),suggesting that the classical complement pathway compo-nent Cl was involved in complement killing by N-RS.Similar to these results, O1:K1 cells were also sensitive tonormal human and guinea pig sera but resistant to C2-deficient human and C4-deficient guinea pig sera, which lackclassical pathway activity (21).
io7 ~~~~~~A
BC
106~~~~~~~/ Y / ~~~~D
z X1E~~~~~~~~
Ei 105
°104
l1 2 3 4 5
FIG. 1. Sensitivity of three selected representative Ki E. colistrains to the bactericidal activity of 90% N-RS. Bacteria wereincubated in serum at 37°C, and the numbers of viable cells weredetermined at different times by plating serial dilutions on L brothagar plates. (A) Strain 41 (O1:K1) in heat-inactivated N-RS, (B)strain 64 (018:K1) in N-RS, (C) strain 165 (07:K1) in N-RS, (D)strain 41 (O1:K1) in MgEGTA-chelated N-RS, (E) strain 41 (01:K1)in N-RS. Time is given in hours.
INFECT. IMMUN.
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
SERUM SENSITIVITY OF E. COLI 687
VI btz
z 0.1L
C1:0~~~~~~~~~
0.01
1 2 3 4TIME (hours)
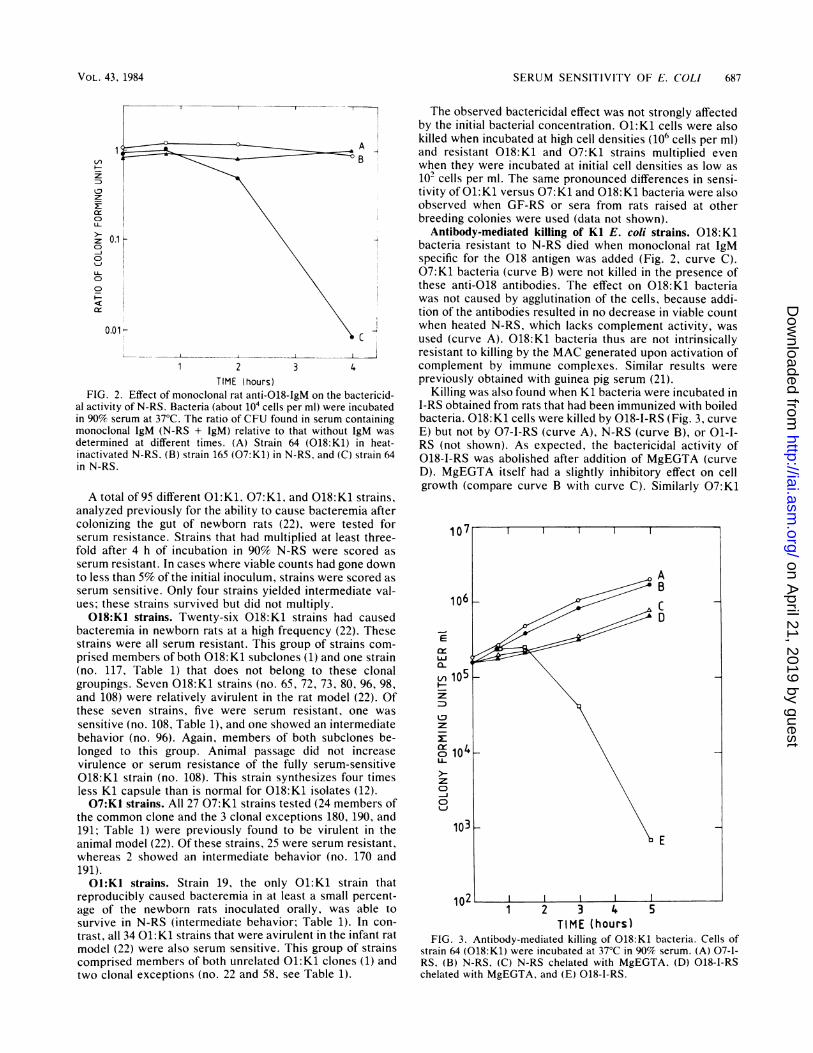
FIG. 2. Effect of monoclonal rat anti-018-IgM on the bactericid-al activity of N-RS. Bacteria (about 104 cells per ml) were incubatedin 90% serum at 37°C. The ratio of CFU found in serum containingmonoclonal IgM (N-RS + IgM) relative to that without 1gM wasdetermined at different times. (A) Strain 64 (018:K1) in heat-inactivated N-RS. (B) strain 165 (07:K1) in N-RS. and (C) strain 64in N-RS.
A total of 95 different O1:K1, 07:K1, and 018:K1 strains,analyzed previously for the ability to cause bacteremia aftercolonizing the gut of newborn rats (22), were tested forserum resistance. Strains that had multiplied at least three-fold after 4 h of incubation in 90% N-RS were scored asserum resistant. In cases where viable counts had gone downto less than 5% of the initial inoculum, strains were scored asserum sensitive. Only four strains yielded intermediate val-ues; these strains survived but did not multiply.018:K1 strains. Twenty-six 018:K1 strains had caused
bacteremia in newborn rats at a high frequency (22). Thesestrains were all serum resistant. This group of strains com-prised members of both 018:K1 subclones (1) and one strain(no. 117, Table 1) that does not belong to these clonalgroupings. Seven 018:K1 strains (no. 65, 72, 73, 80, 96, 98,and 108) were relatively avirulent in the rat model (22). Ofthese seven strains, five were serum resistant, one was
sensitive (no. 108, Table 1), and one showed an intermediatebehavior (no. 96). Again, members of both subclones be-longed to this group. Animal passage did not increasevirulence or serum resistance of the fully serum-sensitive018:K1 strain (no. 108). This strain synthesizes four timesless Kl capsule than is normal for 018:K1 isolates (12).07:K1 strains. All 27 07:K1 strains tested (24 members of
the common clone and the 3 clonal exceptions 180, 190, and191; Table 1) were previously found to be virulent in theanimal model (22). Of these strains, 25 were serum resistant,whereas 2 showed an intermediate behavior (no. 170 and191).O1:K1 strains. Strain 19, the only O1:K1 strain that
reproducibly caused bacteremia in at least a small percent-age of the newborn rats inoculated orally, was able tosurvive in N-RS (intermediate behavior; Table 1). In con-
trast, all 34 01:K1 strains that were avirulent in the infant ratmodel (22) were also serum sensitive. This group of strainscomprised members of both unrelated O1:K1 clones (1) andtwo clonal exceptions (no. 22 and 58, see Table 1).
The observed bactericidal effect was not strongly affectedby the initial bacterial concentration. 01:K1 cells were alsokilled when incubated at high cell densities (106 cells per ml)and resistant 018:K1 and 07:K1 strains multiplied evenwhen they were incubated at initial cell densities as low as102 cells per ml. The same pronounced differences in sensi-tivity of 01:K1 versus 07:K1 and 018:K1 bacteria were alsoobserved when GF-RS or sera from rats raised at otherbreeding colonies were used (data not shown).
Antibody-mediated killing of KI E. coli strains. 018:K1bacteria resistant to N-RS died when monoclonal rat IgMspecific for the 018 antigen was added (Fig. 2, curve C).07:K1 bacteria (curve B) were not killed in the presence ofthese anti-018 antibodies. The effect on 018:K1 bacteriawas not caused by agglutination of the cells, because addi-tion of the antibodies resulted in no decrease in viable countwhen heated N-RS, which lacks complement activity, wasused (curve A). 018:K1 bacteria thus are not intrinsicallyresistant to killing by the MAC generated upon activation ofcomplement by immune complexes. Similar results werepreviously obtained with guinea pig serum (21).
Killing was also found when Kl bacteria were incubated inI-RS obtained from rats that had been immunized with boiledbacteria. 018:K1 cells were killed by 018-I-RS (Fig. 3, curveE) but not by 07-I-RS (curve A), N-RS (curve B), or 01-I-RS (not shown). As expected, the bactericidal activity of018-I-RS was abolished after addition of MgEGTA (curveD). MgEGTA itself had a slightly inhibitory effect on cellgrowth (compare curve B with curve C). Similarly 07:K1
10'
AB
0-~~~~~~~~~
LU
104
I-
1 2 3 4 5TlIME ( hours )
FIG. 3. Antibody-mediated killing of 018:K1 bacteria. Cells ofstrain 64 (018:Kl) were incubated at 37°C in 90% serum. (A) 07-1-RS, (B) N-RS, (C) N-RS chelated with MgEGTA, (D) 018-1-RSchelated with MgEGTA, and (E) 018-1-RS.
VOL. 43, 1984
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

688 PLUSCHKE AND ACHTMAN
H i 1
co C (01-IgM) e D (018-IgM)
d
b
1:-'128 1:164 1:32 1:16 1:128 1:64 1:32 1:161:256 1:256
1:2048 1:2048DILUTION IN BUFFER
FIG. 4. Titers of LPS-specific antibodies in I-RS and N-RS as determined by ELISA. First antibody: (a) N-RS, (b) N-RS containing 1% I-RS, (c) N-RS containing 5% I-RS, (d) N-RS containing 20% I-RS, and (e) I-RS. In A and C, plates were coated with 01-LPS, and 01-I-RS wasused as I-RS. In B and D, plates were coated with 018-LPS, and 018-I-RS was used as I-RS. Second antibody: goat anti-rat-IgG serum in Aand B; goat anti-rat-IgM serum in C and D. Third antibody: peroxidase-labeled anti-goat-IgG antibodies.
cells were killed by 07-I-RS but not by 018-I-RS or by 01-1-RS (data not shown). Thus, strains resistant to N-RS werekilled only by immune sera raised against boiled bacteria ofthe homologous 0 antigen type. Ol:K1 cells were sensitiveto all three I-RS, and killing of these bacteria was notenhanced in 01-I-RS as compared with N-RS.A plausible explanation of these results was that the
observed classical complement pathway killing of 01:K1bacteria by sera from nonimmunized rats was triggered bynatural antibodies specific for the 01 antigen. Thereforeattempts were made to detect and quantitate LPS-specificantibodies.
Antibody content of rat sera. The titer of precipitating anti-0 antigen antibodies was determined by 0 agglutinationtests with boiled bacteria. Whereas N-RS and GF-RS causedno agglutination of boiled 01, 07, or 018 bacteria, all threeI-RS reacted specifically up to a dilution of 1:512 with cellsof the corresponding 0 antigen type.IgG and IgM antibodies that can bind to LPS were
quantitated separately by ELISA. LPS extracted with hotphenol water was adsorbed onto microtiter plates. Serialdilutions of rat sera (first antibody), diluted goat anti-rat-IgGor goat anti-rat-IgM serum (second antibody), and peroxi-dase-labeled rabbit anti-goat-IgG antibodies (third antibody)were sequentially added to the wells, and the amount of thirdantibody bound was measured by enzymatic assay. Onlyweak responses (probably caused by unspecific binding of
rat immunoglobulins to the wells) were observed when N-RSwas assayed against the antigens 01-LPS (Fig. 4A, curve a;Fig. 4C, curve a), 018-LPS (Fig. 4B, curve a; Fig. 4D, curvea), or 07-LPS (not shown). The responses for all three LPStypes were comparable. Addition of 1% I-RS to N-RSresulted in a distinct response to the corresponding LPS (bcurves). Low values similar to those obtained with N-RSwere detected with GF-RS against all three LPS antigens orwhen the immune sera were tested against the heterologousantigens.
Figure 5 shows that 01:K1 cells were killed more efficient-ly by N-RS alone (curve D) than 018:K1 cells were killed byN-RS containing 10% 018-I-RS (curve C). Addition of 1%018-I-RS to N-RS caused a distinct response in the ELISA(Fig. 4) but did not affect the growth of 018:K1 cells (Fig. 5,curve B). Thus, the bactericidal assay is much less sensitivethan the ELISA for anti-LPS antibodies. Because no anti-01-LPS antibodies were detected by the ELISA in N-RS orGF-RS, it would be necessary to invoke an exquisite sensi-tivity of 01:K1 bacteria to anti-LPS antibodies to accountfor the bactericidal activity of N-RS or GF-RS. However, noaccelerated killing was observed when anti-Ol antibodieswere added to N-RS or GF-RS (see above). Thus it seemsthat the presence of trace amounts of anti-01-LPS antibod-ies cannot account for the bactericidal activity seen withthese sera.A highly sensitive blotting technique was used to investi-
INFECT. IMMUN.
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERUM SENSITIVITY OF E. COLI 689
E
L&J
C-E;103C1:1 2
z
102
TIME (hours)FIG. 5. Bactericidal effect of 90% N-RS with and without addi-
tion of I-RS. (A) Strain 64 (018:Kl) in N-RS, (B) strain 64 in N-RScontaining 1% 018-I-RS, (C) strain 64 containing 10%o 018-1-RS, and(D) strain 41 (01:Kl) in N-RS.
gate whether antibodies specific for bacterial antigens otherthan LPS are responsible for complement activation by01:K1 cells in sera from nonimmune rats (Fig. 6). A crudemembrane preparation of the 01:K1 strain 41 was subjectedto separation by SDS-PAGE and electrophoretic transfer toNCM. Under the conditions used, LPS was not bound to theNCM. NCM strips were sequentially incubated with ratserum and alkaline phosphatase-conjugated goat anti-rat-IgGantibodies. Binding of conjugated antibodies was monitoredby staining for phosphatase activity. All three I-RS showed astrong cross-reactivity at a 1:1,000 dilution to a multitude ofantigens (lanes A through C), N-RS reacted somewhat at amuch higher concentration (1:50 dilution) (lane F), whereasGF-RS gave no response (lane E). Similar results wereobtained with membranes of 07:K1 and 018:K1 cells andalso when a double sandwich technique for the detection ofrat IgM was used (data not shown). Because immune seraraised against bacteria of heterologous 0 serotype did notkill 07:K1 and 018:K1 cells, although they contained cross-reactive protein antibodies, it appears that these antibodiesare irrelevant to the initiation of complement killing of thesesmooth encapsulated bacteria. Possibly the protein antigensare not accessible when present in an intact cell membrane(shielding by capsule and LPS), or the antibodies detectedhave a low affinity. Although anti-Kl capsule antibodieswere not assayed, such antibodies should have been equallybactericidal against all Kl strains and are probably lacking inN-RS or GF-RS. Thus GF-RS apparently contains no anti-bodies specific for E. coli antigens, and we conclude that the
bactericidal activity of GF-RS against Ol:K1 cells (which iscomparable to that of N-RS) is antibody independent. Wenote that colostrum-deprived calf serum, which containsonly trace amounts of antibodies, is also bactericidal toO1:K1 but not to 07:K1 or 018:K1 E. coli strains (reference21 and unpublished data).Complement fixation by isolated LPS. Recent evidence
indicates that the classical complement pathway componentCl is readily activated even in the complete absence ofantibodies by a diversity of substances, including the lipid Apart of LPS (5, 6). As complement resistance of Kl E. colistrains seems to be related to certain 0 antigens and 01 cellsseem to activate the classical complement pathway moreefficiently than do 018 bacteria (21), we tested whether 01-,07-, and 018-LPS differ in their capacity to activate comple-ment. Isolated LPS was incubated for 1 h in 90% GF-RS, andthe residual complement activity was measured by hemoly-sis of sensitized sheep erythrocytes. It has been shown (12)that the LPS of the two 01 clonal groupings (01/5 and 01/9)
4
mm
-
p
--e
A BC DE FFIG. 6. Binding of rat antibodies to NCM-bound membrane
antigens of strain 41 (01:Kl). (A) 07-1-RS diluted 1:1,000, (B) 018-I-RS diluted 1:1,000, (C) 01-1-RS diluted 1:1,000, (D) buffer, (E) GF-RS diluted 1:50, and (F) N-RS diluted 1:50.
VOL. 43, 1984
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

690 PLUSCHKE AND ACHTMAN
is structurally somewhat different. Each isolated 01-LPShad a far higher anticomplementary activity than did isolated07- or 018-LPS (Fig. 7). The 01 antigen thus seems to bedirectly responsible for the increased complement activationobserved with 01:K1 cells.
DISCUSSIONMany gram-negative bacterial strains are generally sensi-
tive to the bactericidal effects of vertebrate sera, whereasothers are resistant, at least in the absence of specificantibodies (27). However, no generally accepted techniquehas yet emerged for distinguishing serum-resistant fromserum-sensitive strains. Thus, serum resistance has beenclaimed for laboratory strains of E. coli K-12 after theacquisition of certain plasmids, based upon a small reductionin killing by highly diluted serum. In contrast, serum sensi-tivity has been claimed for smooth, natural isolates that firstshowed a reduction in viable counts after 1 h of exposure to90% serum (27). We have chosen the stringent conditionsrecommended by Taylor (27) and have called strains re-sistant when they showed no reduction in viable counts afterseveral hours of incubation in 90% serum and sensitive whenthe viable counts dropped to less than 5% of the initialinoculum. Most of the strains tested here fell into one ofthese categories, whereas a few yielded intermediate values.A total of 95 01:K1, 07:K1, or 018:K1 E. coli strains had
been analyzed previously for the ability to cause bacteremiaafter colonizing the gut of newborn rats (22). These strainswere tested for their resistance to serum obtained fromnonimmunized adult rats. All strains that had caused bacte-remia in at least 10% of the rats tested were scored asresistant or intermediate in the serum sensitivity test. Noserum-sensitive strain was found which had been able tocause bacteremia. Thus, under the stringent assay condi-tions used, serum resistance or sensitivity correlated withvirulence or lack of virulence in the animal model. A few018:K1 strains which rarely caused bacteremia were serum
100 _I01/5oi/9
11030~~~~~~~~~100
Ln
uJ2:C-
X:
50~~~~~~~~~~~~~1
CONCENTRATION OF LPS (pg/nI)
FIG. 7. Fixation of complement by isolated LPS. The figureshows the percentage of hemolytic activity of GF-RS consumedupon incubation with isolated LPS for 1 h relative to control samplesincubated without LPS. The trend of the results was the same, butdifferences were less pronounced when N-RS was used instead ofGF-RS. The concentration of LPS is given as final concentration inthe assay.
resistant. These may be blocked in early stages of infectionsuch as during colonization or translocation (see reference22) but have not yet been analyzed further.Except for 1 strain with intermediate resistance to serum
and intermediate ability to cause bacteremia, all 34 other01:K1 strains were serum sensitive and avirulent. In con-trast, except for 1 018:K1 strain which synthesized lowlevels of Ki capsule, was serum sensitive, and rarely causedbacteremia, all 59 other 07:K1 and 018:K1 strains testedwere intermediate or resistant to 90% serum. 01:K1 cellsproduce as much or more Kl capsular polysaccharide as do07:K1 or 018:K1 bacteria (12), and differences in capsularcontent thus do not seem to be responsible for the observeddifferences in complement sensitivity. Thus serum sensitiv-ity or resistance correlated with the 0 serotype of thesebacteria, and this correlation is a plausible explanation forthe virulence differences observed in the disease model.018:K1 and 07:K1 cells were not intrinsically resistant tothe bactericidal action of the terminal MAC but were effi-ciently killed once the complement system was activated byaddition of specific anti-LPS antibodies. In the absence ofsuch antibodies, however, the cells were complement re-sistant. In a previous report (21), we showed that 018:K1 E.coli strains require both the Kl capsule and the 0-antigen forinterference with non-immunological complement activa-tion. Sialic acid seems to specifically impede alternativepathway activation by changing the affinity between certaincomplement components (B or H to C3b) (11, 17, 20).Therefore strains that possess the KI (sialic acid) capsuleseem to be generally resistant to killing (21) and opsonization(2, 26) by sera deficient in classical pathway activity(MgEGTA-chelated or genetically C4 or C2 deficient sera).The data presented here and in a previous report (21)
demonstrate that antibody-independent classical pathwayactivation accounts for complement sensitivity of 01:K1cells. Although it is now known that the ability to activatethe classical pathway in the complete absence of antibodiesis shared by many substances, it is unclear which structuralcharacteristics lead to initiation and amplification of thistype of activation (4, 5, 10, 14). Whereas deeper structureson the cell surface seem to be responsible for non-immuno-logical classical pathway activation by rough or unencapsu-lated mutants (4, 21), it appears that the 01 antigen itselfactivates complement efficiently and thus causes serumsensitivity of wild-type 01:K1 E. coli bacteria. It should benoted that 01:K1 cells have as much LPS as do 07:K1 or018:K1 cells and that the length distribution of LPS in01:K1 cells is the same as that of 07:K1 cells and onlyslightly different from that of 018:K1 cells (12). Thus thechemical composition of the LPS rather than the chainlength or amount seems to be responsible for the observeddifferences in complement activation. The LPS of the two01 clonal groupings is, however, structurally and antigeni-cally somewhat different (12; M. Achtman, A. Moll, B.Kusecek, G. Pluschke, B. Slawig, B. Jann, and K. Jann,manuscript in preparation). Furthermore strain 22 (01:K1),whose LPS is dissimilar to that of the clonal groupings (12),was also avirulent and complement sensitive. The differ-ences in the structural composition of these three 01-LPSantigens are currently unknown but do not seem to berelevant for complement activation.The fact that one moderately complement-resistant 01:K1
strain was found that synthesized normal amounts of capsuleand LPS (12) indicates that still other cell structures may beable to modulate the nature of the interactions of comple-ment components with bacterial cell surfaces. Although
INFECT. IMMUN.
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

SERUM SENSITIVITY OF E. COLI 691
many of the bacteria tested here possess F type or CoIVplasmids (1), this does not seem to be very relevant to thesituation described here: some O1:K1 and 018:K1 strains ofmembrane protein pattern 9, which are indistinguishable inmany independent properties (1), also contain an identical Ftype plasmid. Plasmid-free derivatives did not differ in theircomplement resistance properties from those of their parents(manuscript in preparation).The difference in sensitivity of O1:K1 versus 07:K1 or
018:K1 E. coli strains to classical pathway killing was foundin rat, guinea pig, and calf sera (21), demonstrating that thisdifference is related to mechanisms that are not only ratspecific. Furthermore, a comparable difference was foundupon analysis of the relative frequency of isolation of 01:K1,07:K1, and 018:K1, bacteria from cases of meningitis innewborn children (22), suggesting that antibody-independentclassical pathway activation protects newborns against men-
ingitis due to 01:K1 bacteria. Whereas this activation initi-ates killing of O1:K1 cells by the MAC in adult rat serum,
opsonization leading to phagocytotic ingestion may be moreimportant in newborns (3; Pluschke and Achtman, manu-
script in preparation), where the complement system is notyet fully developed (15, 19).The results presented here and in previous reports (21)
indicate that the chemical compositions of both the capsularpolysaccharide and the 0 antigen determine the degree ofcomplement activation and thus the virulence of E. colistrains in meningitis in newborns. The ability to impedealternative pathway activation may well be the major reason
for the clear dominance of Kl strains in meningitis innewborns (7, 23; 25). Differing capacities for antibody-independent classical pathway activation seem to accountfor 0 antigen-related differences in virulence of Kl strains.Additional immune mechanisms (transfer of maternal anti-bodies) may protect most newborns colonized by potentiallyinvasive E. coli bacteria within the first days of life againstsepsis and meningitis. In fact, LPS-specific antibodies notonly initiate killing of otherwise complement-resistant Klstrains in adult rat serum but also protect newborn ratsagainst bacterial invasion (Pluschke and Achtman, manu-
script in preparation).
ACKNOWLEDGMENTSG.P. was supported by grant AC 36/4 from the Deutsche Fors-
chungsgemeinschaft.We gratefully acknowledge receiving the details of the staining
method for blotting from Milan Blake before publication, the gift ofgermfree rat serum from N. F. Juhr, and helpful discussions with A.Mercer, C. Galanos, and R. P. Levine.
LITERATURE CITED1. Achtman, M., A. Mercer, B. Kusecek, A. Pohl, M. Heuzen-
roeder, W. Aaronson, A. Sutton, and R. P. Silver. 1983. Sixwidespread bacterial clones among Escherichia coli Kl isolates.Infect. Immun. 39:315-335.
2. Bortolussi, R., R. Ferrieri, B. Bjorksten, and P. G. Quie. 1979.Capsular Kl polysaccharide of Escherichia coli: relationship tovirulence in newborn rats and resistance to phagocytosis.Infect. Immun. 25:293-298.
3. Bortolussi, R., P. Ferrieri, and L. W. Wannamaker. 1978.Dynamics of Escherichia coli infection and meningitis in infantrats. Infect. Immun. 22:480-485.
4. Clas, F., and M. Loos. 1982. Requirement for an additionalserum factor essential for the antibody-independent activationof the classical complement sequence by gram-negative bacte-ria. Infect.Immun. 37:935-939.
5. Cooper, N. R. 1983. Activation and regulation of the first
complement component. Fed. Proc. 42:134-138.6. Cooper, N. R., and D. C. Morrison. 1978. Binding and activa-
tion of the first component of human complement by the lipid Aregion of lipopolysaccharides. J. Immunol. 120:1862-1868.
7. Glode, M. P., A. Sutton, E. R. Moxon, and J. B. Robbins. 1977.Pathogenesis of neonatal Escherichia coli meningitis: inductionof bacteremia and meningitis in infant rats fed E. coli Kl. Infect.Immun. 16:75-80.
8. Joiner, K. A., C. H. Hammer, E. J. Brown, R. J. Cole, andM. M. Frank. 1982. Studies on the mechanism of bacterialresistance to complement-mediated killing. I. Terminal comple-ment components are deposited and released from Salmonellaminnesota S218 without causing bacterial death. J. Exp. Med.155:797-808.
9. Joiner, K. A., C. H. Hammer, E. J. Brown, and M. M. Frank.1982. Studies on the mechanism of bacterial resistance tocomplement-mediated killing. II. C8 or C9 release C5b67 fromthe surface of Salmonella minnesota S218 because the terminalcomplex does not insert into the bacterial outer membrane. J.Exp. Med. 155:809-819.
10. Kawakami, M., I. Ihara, A. Suzuki, and Y. Harada. 1982.Properties of a new complement-dependent bactericidal factorspecific for Ra chemotype salmonella in sera from conventionaland germ-free mice. J. Immunol. 129:2198-2201.
11. Kazatchkine, M. D., D. T. Fearon, and K. F. Austen. 1979.Human alternative complement pathway: membrane-associatedsialic acid regulates the competition between B and 13lH for cellbound C3b. J. Immunol. 122:75-81.
12. Kusecek, B., H. Wloch, A. Mercer, V. Vaisanen, G. Pluschke, T.Korhonen, and M. Achtman. 1984. Lipopolysaccharide, cap-sule, and fimbriae as virulence factors among 01, 07, 016, 018,or 075 and Kl, K5, or K100 Escherichia coli. Infect. Immun.43:368-379.
13. Lachmann, P. J., M. J. Hobart, and W. P. Aston. 1973. Com-plement technology, p. 5A.1-5A.23. In D. M. Weir (ed.), Hand-book of experimental immunology, vol. I. Blackwell ScientificPublications, Ltd., Oxford.
14. Loos, M., B. Wellek, R. Thesen, and W. Opferkuch. 1978.Antibody-independent interaction of the first component ofcomplement with gram-negative bacteria. Infect. Immun. 22:5-9.
15. Michalek, S. M., J. R. McGhee, and V. K. Ghanta. 1974. Com-plement levels of malnourished animals: quantification of serumcomplement in rat dams and their offsprings. RES J. Reticuloen-dothel. Soc. 16:204-212.
16. Nicholson, A., and I. H. Lepow. 1979. Host defense againstNeisseria meningitidis requires a complement-dependent bacte-ricidal activity. Nature (London) 205:298-299.
17. Okada, N., T. Yasuda, and H. Okada. 1982. Restriction ofalternative complement pathway activation by sialosylglycolip-ids. Nature (London) 299:261-263.
18. 0rskov, I., F.0rskov, B. Jann, and K. Jann. 1977. Serology,chemistry, and genetics of 0 and K antigens of Escherichia coli.Bacteriol. Rev. 41:667-710.
19. Pabst, H. F., and H. W. Kreth. 1980. Ontogeny of the immuneresponse as a basis of childhood disease. J. Pediatr. 97:519-534.
20. Pangburn, M. K., and H. J. Muller-Eberhard. 1978. Comple-ment C3 convertase: cell surface restriction of,1H control andgeneration of restriction on neuraminidase-treated cells. Proc.Natl. Acad. Sci. U.S.A. 75:2416-2420.
21. Pluschke, G., J. Mayden, M. Achtman, and R. P. Levine. 1983.Role of the capsule and the 0 antigen in resistance of018:K1Eschgrichia coli to complement-mediated killing. Infect. Im-mun. 42:907-913.
22. Pluschke, G., A. Mercer, B. Kusecek, A. Pohl, and M. Achtman.1983. Induction of bacteremia in newborn rats by Escherichiacoli Kl is correlated with only certain 0 (lipopolysaccharide)antigen types. Infect. Immun. 39:599-608.
23. Robbins, J. B., G. H. McCracken, E. C. Gotschlich, F.0rskov,I.0rskov, and L. A. Hanson. 1974. Escherichia coli Kl capsularpolysaccharide associated with neonatal meningitis. N. Engl. J.Med. 290:1216-1220.
24. Schreiber, R. D., D. C. Morrison, E. R. Podack, and H. J.
VOL. 43, 1984
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

692 PLUSCHKE AND ACHTMAN
Muller-Eberhard. 1979. Bactericidal activity of the alternativecomplement pathway generated from 11 isolated plasma pro-teins. J. Exp. Med. 149:870-882.
25. Siegel, J. D., and G. H. McCracken. 1981. Sepsis neonatorum.N. Engl. J. Med. 304:642-647.
26. Stevens, P., S. N.-Y. Huang, W. D. Welch, and L. S. Young.1978. Restricted complement activation by Escherichia coli with
INFECT. IMMUN.
the K-1 capsular serotype: a possible role in pathogenicity. J.Immunol. 121:2174-2180.
27. Taylor, P. W. 1983. Bactericidal and bacteriolytic activity ofserum against gram-negative bacteria. Microbiol. Rev. 47:46-83.
28. Westphal, O., and K. Jann. 1965. Bacterial lipopolysaccharides.Methods Carbohydr. Chem. 5:83-91.
on April 21, 2019 by guest
http://iai.asm.org/
Dow
nloaded from