defectivehumanleukocyteantigenclassi-associated ... · in rpmi 1640 medium supplemented with 10%...
TRANSCRIPT

Defective Human Leukocyte Antigen Class I-associatedAntigen Presentation Caused by a Novel �2-MicroglobulinLoss-of-function in Melanoma Cells*
Received for publication, October 24, 2005, and in revised form, April 27, 2006 Published, JBC Papers in Press, April 28, 2006, DOI 10.1074/jbc.M511525200
Chien-Chung Chang‡1, Takeshi Ogino‡, David W. Mullins§¶, Janine L. Oliver§¶, Galina V. Yamshchikov¶�,Nobuyuki Bandoh‡, Craig L. Slingluff, Jr.¶�2, and Soldano Ferrone‡3
From the ‡Department of Immunology, Roswell Park Cancer Institute, Buffalo, New York 14263 and the §Department ofMicrobiology, �Department of Surgery, and ¶Human Immune Therapy Center, University of Virginia, Charlottesville, Virginia 22908
The major histocompatibility complex class I molecules consistof three subunits, the 45-kDa heavy chain, the 12-kDa �2-micro-globulin (�2m), and an �8–9-residue antigenic peptide. Without�2m, themajor histocompatibility complex class Imolecules cannotassemble, thereby abolishing their transport to the cell membraneand the subsequent recognition by antigen-specific T cells. Here wereport a case of defective antigen presentation caused by the expres-sion of a �2m with a Cys-to-Trp substitution at position 25(�2mC25W). This substitution causes misfolding and degradation of�2mC25W but does not result in complete lack of human leukocyteantigen (HLA) class I molecule expression on the surface of mela-noma VMM5B cells. Despite HLA class I expression, VMM5B cellsare not recognized by HLA class I-restricted, melanoma antigen-specific cytotoxic T lymphocytes even following loading with exog-enous peptides or transduction with melanoma antigen-expressingviruses. Lysis of VMM5B cells is restored only following reconstitu-tion with exogenous or endogenous wild-type �2m protein.Together, our results indicate impairment of antigenic peptidepresentation because of a dysfunctional �2m and provide a mecha-nism for the lack of close association between HLA class I expres-sion and susceptibility of tumor cells to cytotoxic T lymphocytes-mediated lysis in malignant diseases.
The human leukocyte antigen (HLA)4 class I molecules, encoded bythe genes located in the major histocompatibility complex, are com-posed of three subunits including a 45-kDa HLA class I heavy chain(HC), a 12-kDa �2-microglobulin (�2m), and an �8–9-residue peptide(1). Expression of these molecules on the cell surface requires the step-wise assembly of HCs, �2m, and peptides in the endoplasmic reticulum(ER) followed by the transport of the trimeric molecule to the plasma
membrane. These processes are dependent on a functional antigenprocessingmachinery (APM), which includes the proteasome subunits,the peptide transporters TAP1 and TAP2, and a number of ER-residentchaperons such as calnexin, calreticulin, ERp57, and tapasin (2, 3). �2mplays an integral part in the assembly and transport of HLA class Imolecules because it stabilizes the HC-�2m heterodimer through non-covalent protein-protein interactions, thereby allowing binding ofendogenous antigenic peptides with the help of TAP and tapasin (4). Asa result, the assembled HC-�2m-peptide trimeric complexes can travelto the cell surface, where they are recognized by HLA class I-restricted,antigen-specific cytotoxic T lymphocytes (CTLs).The lack of HLA class I molecule expression on the cell surface
usually reflects defects in �2m protein synthesis caused by mutationsin the �2m gene, as has been found mostly in malignant cells thus far(5). This abnormality renders tumor cells resistant to tumor antigen-specific CTLs and may have a negative impact on the elimination oftumor cells by host CTLs. The defects underlying �2m loss have thusfar been found to be structural in nature, involving lack of translationbecause of small deletions or point mutations in most cases and lackof transcription because of large deletions in a few cases (5). Becauseof a lack of �2m expression, the resulting HLA class I loss cannot becorrected by interferon (IFN-�), a cytokine that up-regulates theexpression of most of the molecules participating in the assemblyand transport of HLA class I molecules. On the other hand, a lowlevel of HLA class I expression on cells usually reflects nonstructuraldefects in the APM components because this abnormality can becorrected by IFN-� (6).
In the present study, we have elucidated the mechanism underlyingHLA class I down-regulation identified in a melanoma cell line and inthe metastasis from which the cell line was derived (7). This HLA classI down-regulation phenotype cannot be corrected by IFN-� and wasunexpectedly found to be caused by an abnormal full-length �2m pro-tein that can neither form stable complexes with HCs nor assist inloading peptides onto the HLA class I peptide binding groove.
EXPERIMENTAL PROCEDURES
Cell Lines—The VMM5A and VMM5B melanoma cell lines wereestablished from two sequential metastatic melanoma deposits surgi-cally removed from patient VMM5 at two time points (7). Other mela-noma cell lines included DM6 cells (a gift from Dr. T. L. Darrow, DukeUniversity, Durham, NC), which express HLA-A2 and multiple mela-nocytic differentiation antigens including gp100, melanoma antigenrecognized by T cells (MART-1) and tyrosinase, and FO-1 cells, whichdo not express HLA class I molecules because of �2m loss (8). LG-2 is ahumanB lymphoblastoid cell line. All of these cell linesweremaintained
* This work was supported in part by Public Health Services Grants R01 CA67108 and P30CA160056 (to S. F.) and R01 CA57653 (to C. L. S.) awarded by NCI, National Institutes ofHealth, Department of Health and Human Services. The costs of publication of thisarticle were defrayed in part by the payment of page charges. This article must there-fore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
1 Recipient of the Susan G. Komen Breast Cancer Foundation dissertation researchaward.
2 An Elaine Shepard cancer investigator supported by the Cancer Research Institute.3 To whom correspondence should be addressed: Dept. of Immunology, Roswell Park
Cancer Inst., Elm and Carlton St., Buffalo, NY 14263. Tel.: 716-845-8534; Fax: 716-845-7613; E-mail: [email protected].
4 The abbreviations used are: HLA, human leukocyte antigen; APM, antigen processingmachinery; �2m, �2-microglobulin; CTL, cytotoxic T lymphocyte; ER, endoplasmicreticulum; HC, HLA class I heavy chain; IFN-�, interferon-�; LMP, low molecular masspolypeptide; LOH, loss of heterozygosity; mAb, monoclonal antibody; MFI, mean flu-orescence intensity; TA, tumor antigen; TAP, transporter associated with antigenprocessing; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TyrVac, tyrosin-ase-expressing vaccinia strain.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 281, NO. 27, pp. 18763–18773, July 7, 2006© 2006 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18763
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

in RPMI 1640 medium supplemented with 10% heat-inactivated fetalcalf serum in a 5% CO2 atmosphere at 37 °C. A tyrosinase369–377D-spe-cific5 CTL line was derived from patient VMM119 who had been vac-cinated with a mixture of four peptides including tyrosinase369–377D
(YMDGTMSQV) (9).Monoclonal and Polyclonal Antibodies—The following mAbs were
developed and characterized as described: the mAbW6/32, which rec-ognizes a monomorphic determinant expressed on �2m-associatedHLA-A, -B, and -C heavy chains (10); the mAb LG III-147.4.1, whichrecognizes a determinant expressed on �2m-associated HLA-A heavychains except A23, A24, A25, and A32 (11); the mAb B1.23.2, whichrecognizes a determinant restricted to HLA-B and -C antigens (12); themAbHC-10, which recognizes a determinant expressed on all�2m-freeHLA-B and -C heavy chains and on �2m-free HLA-A10, -A28, -A29,-A30, -A31, -A32, and -A33 heavy chains (13–15); the �2m-specificmAbs L368 (16) andKJ-2, which recognizeHLA class I heavy chain-free�2m6; the delta (Y)-specific mAb SY-5; theMB1 (X)-specific mAb SJJ-3;the Z-specific mAb NB-1; the LMP2-specific mAb SY-1; the LMP7-specific mAb HB-2; the LMP10-specific mAb TO-7 (17); the TAP1-
specific mAb NOB-1 (18); the TAP2-specific mAb NOB-2 (18); thecalnexin-specific mAb TO-5 (19); the calreticulin-specific mAb TO-11(19); the ERp57-specific mAb TO-2 (19); and the tapasin-specific mAbTO-3 (19). The human actin-specific mAb (sc-8432) was purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA). The anti-idiotypicmAb MK2–23 (20) and mouse IgG2a (BD Biosciences) were used asisotype controls. R-phycoerythrin-conjugated goat anti-mouse Fc�
F(ab�)2 fragments and goat anti-mouse IgG antibodies were purchasedfrom Dako (Carpinteria, CA) and GE Healthcare, respectively.
Peptides, IFN-�, Wild-type Human �2m, and PharmacologicalInhibitors—The HLA class I-associated peptides HER2/neu369–377,KIFGSLAFL, and tyrosinase369–377D YMDGTMSQV were synthesizedwith a free amide N terminus and free acid C terminus by standardFmoc (N-(9-fluorenyl)methoxycarbonyl) chemistry using a modelAMS422 peptide synthesizer (Gilson Co. Inc.,Middleton,WI). Peptideswere purified to �98% purity by reverse-phase high pressure liquidchromatography on a C-8 column (Vydac, Hesperia, CA) at the Univer-sity of Virginia biomolecular core facility. Purity and identity were con-firmed using a triple quadrupolemass spectrometer (Finnigan, San Jose,CA). Recombinant human IFN-� was purchased from Roche AppliedScience. The wild-type human �2m was purchased from Sigma. Theproteasome inhibitor MG132 and the trans-Golgi-to-secretory vesicles
5 D means a post-translational modification from N to D at position 371 when the proteintyrosinase is processed inside the cells.
6 S. Ferrone, unpublished results.
FIGURE 1. HLA class I down-regulation on VMM5B melanoma cells. A, control- (thin line) and IFN-�-treated (thick line) VMM5A, VMM5B, and FO-1 cells were stained with mAb W6/32and analyzed by flow cytometry. B, VMM5B cells were treated with citric acid (pH 3.0) before being added with either medium alone (pH 3.0) or the combination of HER2/neu369 –375
peptide (40 �g/ml) with human �2m (2.5 �g/ml) (�2m � peptide) and cultured for 3 h at room temperature. Subsequently, cells were harvested, stained with mAb W6/32, andanalyzed by flow cytometry. The untreated VMM5B cells (pH 7.4) were used as a negative control. The numbers above each open profile are -fold MFI. Representative results of oneof the three experiments are shown. The variability in results between experiments is less than 5%.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
18764 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from
traffic inhibitormonensin were purchased from Sigma andAlexis Corp.(San Diego, CA), respectively.
Low pH Treatment of Cells and Restabilization of Cell Surface HLAClass I Molecules—Low pH treatment of cells was performed asdescribed previously (21). Briefly, cell pellets containing �1–10 � 106
cells were resuspendedwith 0.5ml of stripping buffer (0.13 M citric acid,66 mM Na2HPO4, 1% bovine serum albumin, pH 3.0) for 2 min on iceand then neutralized immediately with 50 ml of RPMI 1640 medium.After three washes, cells (1 � 106) were pulsed with 40 �g/ml peptideand 2.5 �g/ml human �2m (Sigma) for 3 h at room temperature andthen washed three times with 1% bovine serum albumin/phosphate-buffered saline. Subsequently, cells were subjected to staining and flowcytometric analysis as described.
FlowCytometry—Cell surface staining and intracellular stainingwereperformed as described (22). Results are presented as -fold increase inmean fluorescence intensity over the control (-fold MFI).
Reverse Transcription-PCR—Total RNA isolation from cells wasperformed with TRIzol (Invitrogen) according to the manufacturer’sinstructions. First-strand cDNA synthesis was conducted using theSuperScriptTM system (Invitrogen) according to the manufacturer’sinstructions. The PCR was performed using �2m-specific primers,forward 261 (5�-CCTGAAGCTGACAGCATTCG-3�) and reverse262 (5�-ACCTCCATGATGCTGCTTAC-3�); GAPDH-specific prim-ers, GAPDH-F (5�-TGAAGGTCGGAGTCAACGGATTTGGT-3�)and GAPDH-R (5�-CATGTGGGCCATGAGGTCCACCAC-3�). PCRproducts were run on a 1.2% agarose gel (Roche Applied Science) andvisualized by ethidium bromide staining. The predicted sizes of PCRproducts were 401 bp for �2m and 983 bp for GAPDH. The PCR prod-ucts were gel-purified, and their sequences were analyzed by a DNAsequencer (ABI PRISM model 377, Applied Biosystems).
Western Blot Analysis—Western blot analysis of cell lysates with�2m-, HLA class I heavy chain-, andAPMcomponent-specific antibod-ies was performed as described (22).
Genomic DNA Extraction and PCR—Genomic DNA extraction wasperformed with blood and cell culture DNA midi kit (Qiagen Inc.,Valencia, CA) according to themanufacturer’s instructions. PCR ampli-fication of �2m gene exon 2 was conducted with the primers, 491 intron1-specific (5�-CCTGGCAATATTAATGTG-3�) and 462 intron 2-spe-cific (5�-CATACACAACTTTCAGCAGCT-3�). The predicted PCRproduct size is 362 bp. The PCR products were gel-purified and theirsequences analyzed by a DNA sequencer (ABI PRISM model 377,Applied Biosystems).
Loss of Heterozygosity (LOH) Analysis of the �2m Gene—LOH analy-sis of the �2m gene was performed as described previously (23) withminor modifications. Briefly, purified genomic DNA (100 ng) was sub-jected to a PCR amplification using two pairs of primers specific to thetwo short tandem repeat markers (D15S126 and D15S209) located nearthe �2m gene. The sequences of the primers are as follows: D15S126F,5�-GTGAGCCAAGATGGCACTAC-3�; D15S126R, 5�-GCCAGC-AATAATGGGAAGTT-3�; D15S209F, 5�-AAACATAGTGCTCTG-GAGGC-3�; and D15S209R, 5�-GGGCTAACAACAGTGTCTGC-3�.The amplification conditions are as follows: 95 °C for 12 min, 94 °C for30 s, 55 °C for 30 s, 72 °C for 30 s for 10 cycles; 89 °C for 30 s, 55 °C for30 s, 72 °C for 30 s for 20 cycles; and a final extension at 72 °C for 10min.PCR products were then fractionated on a 4% agarose gel and visualizedby ethidium bromide staining. The intensity of the staining was deter-mined by the AlphaImagerTM 2200 system (Alpha Innotech Corp., SanLeandro, CA). LOH is determined by the following formula: (intensityof tumor allele one/intensity of tumor allele two)/(intensity of normal
allele one/intensity of normal allele two) (referred to as the percent LOHindex). An LOH index less than 50% is considered significant.
Radiolabeling of Cells, Indirect Immunoprecipitation, and SDS-PAGE—Radiolabeling of cells, indirect immunoprecipitation, andSDS-PAGE were performed as described (24).
Transfection—Cells were transfected with a plasmid encoding thewild-type �2m or the mutant �2m (�2mC25W) utilizing Lipofectamine-mediated gene transfer (Invitrogen) according to the manufacturer’sinstructions. Briefly, pcDNA3.1-b2m, pcDNA3.1-b2mC25W, or theempty plasmid pcDNA3.1-neo (Invitrogen) was mixed with Lipo-fectamineTM 2000 before being added to melanoma cells grown inmonolayers with a 90% confluence. Cells were selected 2 days aftertransfection in medium containing G418 sulfate (1 mg/ml) (Calbio-chem). After �2–3 weeks of selection, G418-resistant clones werepicked, expanded, and screened by flow cytometry for HLA class Iexpression. Positive clones were then further expanded in completemedia supplemented with a maintenance dose (0.3 mg/ml) of G418.
Recombinant Vaccinia Virus—Virus encoding full-length humantyrosinase (TyrVac) was constructed as described previously (25). Puri-fied recombinant vaccinia virus stock was titered and tested for properexpression of tyrosinase using specific HLA-A2-restricted CTL (datanot shown).
Cytotoxicity Assay in Vitro—Standard 51Cr release assays wereperformed to determine CTL recognition of the HLA-A2-restricted
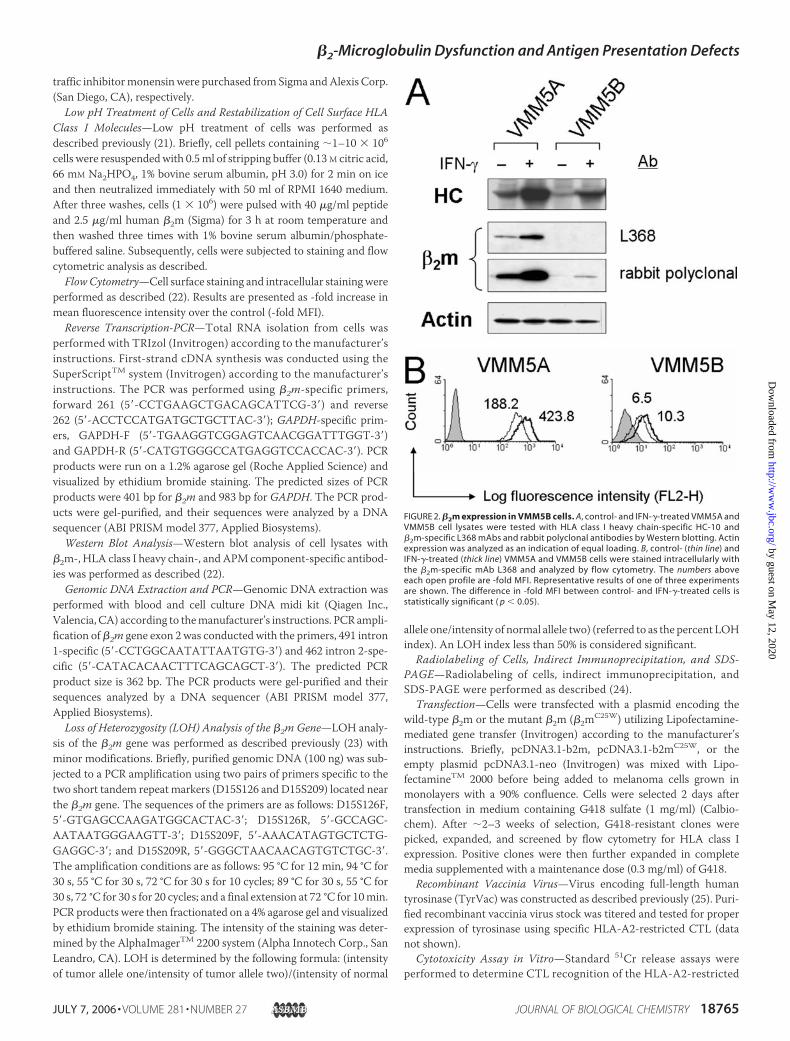
FIGURE 2. �2m expression in VMM5B cells. A, control- and IFN-�-treated VMM5A andVMM5B cell lysates were tested with HLA class I heavy chain-specific HC-10 and�2m-specific L368 mAbs and rabbit polyclonal antibodies by Western blotting. Actinexpression was analyzed as an indication of equal loading. B, control- (thin line) andIFN-�-treated (thick line) VMM5A and VMM5B cells were stained intracellularly withthe �2m-specific mAb L368 and analyzed by flow cytometry. The numbers aboveeach open profile are -fold MFI. Representative results of one of three experimentsare shown. The difference in -fold MFI between control- and IFN-�-treated cells isstatistically significant ( p � 0.05).
�2-Microglobulin Dysfunction and Antigen Presentation Defects
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18765
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

epitope from the melanocyte differentiation protein tyrosinase(tyrosinase369–377D). Target cells were prepared by either loadingwith tyrosinase369–377D peptide (40 �g/ml) for 1 h at room temper-ature or by infecting with TyrVac (10 plaque-forming units/cell, 106
target cells) in 1 ml of Hanks’ balanced salt solution supplementedwith 0.1% bovine serum albumin, 1.6 mM MgSO4, and 1.8 mM CaCl2for 1 h and then 4 ml of RPMI 1640 medium supplemented with 10%fetal bovine serum for 8 h to allow for expression followed by labelingwith 100 �Ci of Na51CrO4 for 1 h for a standard 4 h 51Cr releaseassay. Percent cytotoxicity was calculated by the formula: % specificlysis � ((experimental release � spontaneous release)/(totalrelease � experimental release)) � 100.
RESULTS
MarkedHLAClass IDown-regulation onVMM5BMelanomaCells—Fluorescence-activated cell sorting analysis of cells stained with mAbW6/32 showed that HLA class I molecules were barely detectable onmelanoma cells VMM5B as compared with autologous VMM5Amela-noma cells (Fig. 1A). The low level of staining ofVMM5BcellswithmAbW6/32 does not represent nonspecific background staining because nostaining was detected when the �2m-deficient FO-1 cells (8) werestained with mAbW6/32. Incubation of VMM5B cells with IFN-� (300units/ml) for 48 h at 37 °C had no detectable effect on the staining ofVMM5B cells, as well as FO-1 cells, by mAbW6/32.
To corroborate the specificity of the staining ofVMM5Bcells bymAbW6/32, cells were treated with low pH (pH 3.0) and then stained withmAb W6/32, a method used to disintegrate the trimeric HLA class Icomplex on the cell surface (21). As shown in Fig. 1B, following treat-ment with pH 3.0, VMM5B cells were not stained by mAb W6/32.These results imply the dissociation of peptides and �2m from the HCsbecause the antigenic determinant recognized by mAbW6/32 requiresthe association of HCs with �2m for its expression (10). This possibilityis supported by the restoration of the staining with mAbW6/32 follow-ing loading of low pH-treated VMM5B cells with wild-type �2m alongwith a HLA-A2-binding peptide (HER-2/neu369–397, KIFGSLAFL, t1⁄2 �
481.2 min). This result reflects the reassembly of the trimeric HLA classI complex on the cell surface. It is noteworthy that the level of therestored HLA class I expression is similar to that on untreated cells,remaining at a low level, unlike the full restoration ofHLAclass I expres-sion on low pH-treated autologous VMM5A cells following loadingwith exogenous peptide and �2m (data not shown). These results sug-gest that the amount of HCs transported to the plasmamembrane is thelimiting factor in VMM5B cells. This finding is different from the effi-cient transport to the cell surface of open-form (peptide-free or lowaffinity peptide-bound) HCs, which can subsequently be refolded totheir native, bioactive conformation following loading with a high affin-ity peptide, as has been observed in the TAP-deficient T2 cells (3).
FIGURE 3. Identification of a C-to-G point mutation in exon 2 of the �2m gene in VMM5B cells. A, �2m mRNA expression in VMM5B cells was analyzed by reverse transcription-PCR.GAPDH mRNA expression served as an internal control. B, �2m cDNA and gene prepared from VMM5B cells were subjected to nucleotide sequence analysis. The downward trianglesindicate the location (exon 2, nt 135) of the C-to-G transversion mutation. C, the C-to-G mutation site in the �2m gene is schematically illustrated.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
18766 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

To determine whether the low level of HLA class I molecules onthe membrane of VMM5B cells is caused by a reduced level of allHCs and/or by a defect in �2m, HC and �2m expression in VMM5Aand VMM5B cell lysates were analyzed by Western blotting. Asshown in Fig. 2A, HCs were detected in both cell lysates and weremarkedly up-regulated by IFN-�. In contrast, �2m was barely detect-able with �2m-specific rabbit polyclonal antibodies in a VMM5B celllysate following IFN-� treatment. �2m was not detected in aVMM5B cell lysate with a panel of �2m-specific mouse mAb (Fig. 2Aand data not shown). The latter results may reflect the insufficientsensitivity of the Western blotting technique because �2m wasdetected in VMM5B cells by intracellular staining with the �2m-specific mAb L368 (Fig. 2B). These results in conjunction with thelack of up-regulation of HLA class I molecules by IFN-� suggest thatthe abnormal HLA class I phenotype of VMM5B cells is caused by adefect in �2m.
Cys-to-Trp substitution in �2m Caused by a Point Mutation in the�2m Gene in VMM5B Cells—To investigate whether structuralmutation(s) underlie the low �2m level and its functional abnormal-ities, we amplified the full-length �2m cDNA fragments (Fig. 3A) andperformed nucleotide sequencing of the open reading frame of the�2m gene in VMM5B cells. As shown in Fig. 3, B and C, a cytosine-to-guanine transversion mutation at position 135 in exon 2 wasdetected. This mutation changes codon 25 from Cys to Trp(�2mC25W) (Fig. 4A), abolishing the disulfide bond between residue25 (Cys-25) and 80 (Cys-80) of the full-length �2m protein (B). It isnoteworthy that this mutation was not acquired during in vitro cul-ture of the VMM5B cell line because the identical mutated nucleo-tide was detected in the genomic DNA extracted from the melanomametastasis from which the cell line had been derived (Fig. 5A). More-over, the wild-type sequence of the �2m gene in VMM5B cells wasnot detected suggesting that one �2m gene copy was lost. This pos-
FIGURE 4. Nucleotide and deduced amino acid sequences of �2m cDNA synthesized from VMM5A and VMM5B cells. A, sequence conservation is indicated by dashes.Downward triangles indicate the exon-exon junctions. Bold underlined nucleotide letters indicate the mutated codon. Bold nucleotide letters indicate the stop codon. Italic amino acidletters indicate the signal peptide sequence. Bold amino acid letters indicate the residues where the substitution occurs. B, the lack of the disulfide bond in the �2m protein in VMM5Bcells is schematically shown.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18767
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

sibility is supported by the LOH identified at two chromosome 15short tandem repeat sites (D15S126 and D15S209) (23) flanking the�2m gene in genomic DNA extracted from VMM5B cells and thecorresponding melanoma metastasis (Fig. 5, B and C).
Instability of �2mC25W in VMM5B Cells—To test whether loss of adisulfide bond in the mutant �2m identified in VMM5B cells causedconformational instability of the full-length protein, we examinedthe effect of Cys-to-Trp substitution at residue 25 on the stability of�2m structure using molecular modeling. The wild-type �2m is a�-sandwich structure composed of two anti-parallel �-pleatedsheets connected by a disulfide bond between Cys-25 and Cys-80(Fig. 6A). Each of the �-pleated sheets contains three � strands. Inthe wild-type Cys-25–Cys-80 hydrophobic core, the linked sulfuratoms exhibit favorable van der Waals contacts with respect to the
surrounding atoms 4.43, 4.81, and 3.88 Å for Cys25SG-Val27CG1,Cys25SG-Gln8C�, and Cys80SG-Val82CG2, respectively (Fig. 6B).However, when Cys-25 is replaced with Trp-25, the bulky indole ringof Trp side chain displays drastic steric clashes with the side chains ofthe neighboring residues. These clashes occur with all 14 possibleTrp-25 side chain rotameric conformations analyzed. Fig. 6C shows onerepresentative conformation in which the ring carbon and nitrogenmem-bers arewithin the unfavorable van derWaals distanceswith the neighbor-ingatoms2.63, 1.62, 2.74, and1.95ÅforTrp25CZ3-Tyr66CD1,Trp25CZ2-Val27CG1, Trp25NE1-Val82CG2, and Trp25CD1-Cys80SG, respectively.Because all of these distances are below 3.0Å, the minimum distancebetween two nonbonded carbon atoms, the free energy is drasticallyincreased, leading toa thermodynamicallyunstable stateof themutant�2m(�2mC25W).
FIGURE 5. Detection of C-to-G point mutation and LOH of the �2m locus in genomic DNA extracted from VMM5B metastasis. A, the nucleotide sequence analysis shows a mixof C (blue) and G (black) at position 135 in genomic DNA prepared from VMM5B metastasis. B, the heterozygosity status of the �2m locus in VMM5A cells, VMM5B cells, cryopreservedVMM5B tumor (VMM5B-T ), and cryopreserved peripheral blood lymphocytes (PBL) of the patient was subjected to LOH analysis. C, the degree of LOH of the �2m locus in VMM5B cellsand VMM5B metastasis is quantitatively presented.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
18768 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Degradation of�2mC25W and Lack of StableHC-�2mC25WAssociationin VMM5B Cells—Next we tested whether �2mC25W was degraded bythe proteasome in VMM5B cells, especially because it was present at avery low level intracellularly, approximately at a level 29-fold and41-fold lower than that in VMM5A cells under basal conditions andfollowing incubation with IFN-� (300 units/ml), respectively (Fig. 2B).To this end, VMM5B cells were sequentially incubated with IFN-� (300units/ml) for 24 h at 37 °C and with the proteasome inhibitorMG132 (5�M) for 12 h at 37 °C. The intracellular levels of the steady-state free�2mC25W, free HCs, and HC-�2mC25W complexes in IFN-�/MG132-treated and in IFN-�-treated VMM5B cells were compared utilizingfluorescence-activated cell sorting analysis of cells intracellularlystained with mAbs. As shown in Fig. 7, A and B, both the �2mC25W andHCs were increased �2-fold following incubation of cells with IFN-�,but the level of HC-�2mC25W complexes remained unchanged. In thepresence of IFN-� and MG132, the level of �2mC25W increased mark-edly (5-fold above that in untreated cells) alongwith an�3-fold increasein HCs, but the level of HC-�2mC25W complexes still remainedunchanged. On the other hand, increased secretion of �2mC25W did notappear to play a role in its low intracellular accumulation because thelevel of �2mC25W, HCs, and HC-�2mC25W complex expression was notincreased by the addition, 4 h before harvest, of monensin, an inhibitorof trans-Golgi-to-secretory vesicles traffic, to MG132-treated cells cul-tured in the presence of IFN-�. Therefore, �2mC25W was removedthrough proteasome-mediated degradation. Even when it accumulates,the mutant �2m cannot form stable complexes with HCs. This conclu-sion is corroborated by the lack of immunoprecipitation ofHC-�2mC25W complexes with mAb W6/32 from MG132 and IFN-�-treated VMM5B cell lysates (Fig. 7C). Attempts to obtain biochemicalevidence for the degradation of �2mC25W in pulse-chase experimentsfailed because the available methods were not sufficiently sensitive todetect the mutant �2m in immunoprecipitation (Fig. 7C and data notshown).
Restoration of HLA Class I Expression and Peptide Presentation onVMM5B Cells following Exogenous and Endogenous Reconstitution ofWild-type �2m Expression—Despite HLA class I expression, VMM5Bcells were resistant to lysis by HLA-A2-restricted, tyrosinase-derivedpeptide tyrosinase369–377D (YMDGTMSQV, t1⁄2 � 212.6 min)-specificCTLs, even following pulsing target cells with the tyrosinase369–377D
peptide (Fig. 8, A andD). To determine whether reconstitution of wild-type �2m expression is sufficient to restore HLA class I expression andits ability to present peptides to CTLs, we transfected VMM5B cellswith a wild-type �2m cDNA. As shown in Fig. 8B, expression of HLAclass I molecules was restored on stable clones of �2m-transfectedVMM5B cells (VMM5B.�2.7G) compared with mock-transfectedVMM5B cells (VMM5B.neo). Similarly, the gene products of HLA-Aand HLA-B, HLA-C loci were restored, although the latter moleculeswere expressed at a level �10-fold lower than the former ones (Fig. 8B).To test whether the lack of peptide presentation is caused by themutated �2m (�2mC25W) associated with HC, VMM5B.neo cells weretreated with low pH to dissociate the HC-�2mC25W complexes on thecell surface before being loaded with the wild-type �2m together withthe tyrosinase369–377D peptide and tested for susceptibility to lysis bytyrosinase369–377D-specific CTLs. VMM5B.�2.7G cells were treated in asimilar fashion and analyzed in parallel as a control. Although acid-stripped VMM5B.�2.7G cells became sensitive to CTL lysis followingloading with either the tyrosinase369–377D peptide by itself or both, thewild-type �2m and the tyrosinase369–377D peptide (Fig. 8C, left panel),VMM5B.neo cells were sensitive to CTL lysis only following loadingwith both thewild-type�2m and the tyrosinase369–377D peptide (Fig. 8C,right panel). These results indicate that peptide presentation is restoredon VMM5B cells only when both the wild-type �2m and thetyrosinase369–377D peptide are present. Therefore, the presence of�2mC25W does not allow exogenous peptide loading onto HLA class Imolecules.To test whether �2mC25W affects endogenous peptide loading,
VMM5B.neo cells were transduced with TyrVac, a tyrosinase-express-ing vaccinia strain, and analyzed for susceptibility to lysis by HLA-A2-restricted, tyrosinase369–377D peptide-specific CTLs. As shown in Fig.8D, TyrVac-infected VMM5B.neo cells were resistant to CTL lysis,whereas TyrVac-infected VMM5B.�2.7G cells were sensitive. In theseexperiments, the tyrosinase-expressing DM6 cells were used as a posi-tive control, whereas the untransduced VMM5B, VMM5B.neo, andVMM5B.�2.7G cells were used as specificity controls. Therefore, inVMM5B cells, the endogenously processed tyrosinase369–377D peptidecannot be presented by�2mC25W associated withHLA-A2 heavy chainsunless the wild-type HLA-A2-�2m dimer is available in sufficientamount following reconstitution with wild-type �2m expression. These
FIGURE 6. Molecular modeling of �2m with a Cys-to-Trp substitution at position 25 (�2mC25W). A,the three-dimensional structure of the HLA-A2-�2m-human immunodeficiency virus polypeptide com-plex (Protein Data Bank code 1AKJ), in which the�2m is enlarged on the right, is shown in a schematicdiagram. The disulfide bond between Cys-25 andCys-80 of �2m is highlighted. Close-up views of Cys-25–Cys-80 (B) and Trp-25–Cys-80 (C ) show the vander Waals distances in relation to the neighboringatoms. Pictures were generated with the PyMOLmolecular graphics program (DeLano Scientific LLC).
�2-Microglobulin Dysfunction and Antigen Presentation Defects
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18769
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE 7. Lack of stable HC-�2mC25W association despite accumulation of �2mC25W following inhibition of proteasome function in VMM5B cells. A, VMM5B cells were treatedwith the indicated combination of IFN-�, MG132, and monensin or mock-treated before being intracellularly stained with �2m-specific mAb KJ-2, HC-specific mAbs HC-A2 and HC-10,and HLA class I-specific mAb W6/32 and analyzed by flow cytometry. The numbers above each histogram profile indicate the -fold MFI increase above the background. B, the -folddifferences in the levels of free �2m, free HC-A, -B, and -C and HC-�2m complexes among the indicated treated groups are shown. C, lysates of VMM5B cells treated with IFN-� and/orMG132 were immunoprecipitated with mAb W6/32 and analyzed by SDS-PAGE. A VMM5A cell lysate was used as a positive control. The bottom panel shows equal loading of proteinsin each lane as monitored by Western blot analysis of an equal portion of each lysate preparation with calnexin-specific mAb TO-5. CNX, calnexin.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
18770 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

FIGURE 8. Restoration of HLA class I expression and function following reconstitution of wild-type �2m expression on VMM5B cells. A, VMM5B cells (VMM5B.neo) wereanalyzed for susceptibility to lysis by HLA-A2-tyrosinase369 –377D-specific CTL in cytotoxicity assays following loading with the tyrosinase369 –377D peptide (YMDGTMSQV). VMM5A,VMM5B.�2.7G, and DM6 cells were used as positive controls. B, �2m-transfected VMM5B cells (VMM5B.�2.7G) and their mock-transfected counterpart VMM5B.neo cells were stainedwith HLA class I-specific mAb W6/32 (thick line), HLA-A2-specific mAb KS-1 (thin line), and HLA-B and -C-specific mAb B1.23.1 (dotted line) and analyzed by flow cytometry. C,acid-treated VMM5B.�2.7G (left panel) or VMM5B.neo cells (right panel) loaded with the tyrosinase369 –377D peptide or with the tyrosinase369 –377D peptide and wild-type �2m wereanalyzed for susceptibility to lysis by HLA-A2-tyrosinase369 –377D-specific CTLs in cytotoxicity assays. Acid-treated VMM5B.�2.7G and VMM5B.neo cells incubated in medium wereused as controls. D, VMM5B.neo and VMM5B.�2.7G cells infected with a TyrVac were analyzed for susceptibility to lysis by HLA-A2-tyrosinase369 –377D-specific CTLs in cytoxicity assays.Control-infected cells and DM6 cells serve as a specificity control and a positive control, respectively. E:T, effector-to-target ratio.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18771
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

data indicate that although expressed on the cell membrane, HLA classI heavy chains, associated with �2mC25W, cannot present peptides.
To exclude the possibility that defects in antigen presentation byVMM5B cells were caused by APM abnormalities, the level of APMcomponent expression in VMM5A and VMM5B cells was compared.No marked difference was detected, except for lower LMP7, LMP10,calnexin, and tapasin expression inVMM5B cells than inVMM5Acells,as measured by Western blotting (Fig. 9) and by flow cytometry (forresults, see Ref. 7). However, LMP7, LMP10, calnexin, and tapasin down-regulation did not appear to have an impact on antigen presentationbecause peptide presentation was restored in VMM5B cells transfectedwith awild-type�2m cDNA (VMM5B.�2.7G) (Fig. 8D). This observationin conjunction with the failure of exogenous peptide loading ontoVMM5B cells, as compared with the wild-type �2m-transfectedVMM5B.�2.7G cells, indicates that defective HLA class I-associatedantigen presentation in VMM5B cells is caused by a dysfunctional�2m.
Restoration of Low Level of HLA Class I Expression on �2m-deficientCells Transfected with a cDNA Construct Encoding the Mutant�2mC25W—Toexclude the possibility that the observed low level ofHLAclass I expression on VMM5B cells is a cell line-specific phenomenon,we transfected a �2mC25W-expressing cDNA construct into FO-1 cells,a well characterized �2m-deficient cell line harboring a large deletion inthe �2m gene (8). Fourteen stable clones were established and analyzedfor HLA class I expression on the cell surface by flow cytometry withmAbW6/32. Ten of them were stained weakly by mAbW6/32 as com-pared with the mock-transfected (FO-1.neo) and wild-type �2m-trans-fected (FO-1.�2) controls (Fig. 10 and data not shown). These resultsindicate that the mutant �2mC25W alone is able to restore low level ofHLA class I expression on the cell surface regardless of the differences inthe genetic background.
DISCUSSION
In this report, a naturally occurring, full-length mutant �2m(�2mC25W) in the melanoma cell line VMM5B and the metastasis fromwhich the cell line originated was found to cause HLA class I down-regu-lation rather than loss because of its impaired association with HCs. Nota-bly, the HC-�2mC25W dimer does not appear to constitute a conventionalpeptide-receptive conformation because neither exogenous nor endoge-nouspeptides canbepresentedon it, as indicatedby the lackof inductionoflysis by the cognate CTLs. These findings represent the first example of a�2m structural abnormality that does not cause total HLA class I loss but
causes defects in antigen presentation associated with their down-regula-tion. Importantly, the abilityof themutant�2mC25W to restore a low level ofHLA class I expression is not restricted to VMM5B cells because the samephenotype was observed in the allogeneic �2m-deficient cell line FO-1transfected with a �2mC25W-expressing construct.
Several examples of single amino acid substitutions in proteinsinvolved in HLA class I-associated antigen presentation, such as TAP1,
FIGURE 9. APM component expression in con-trol- and IFN-�-treated VMM5A and VMM5Bcells. Lysates of control- and IFN-�-treatedVMM5A and VMM5B cells were tested with APMcomponent-specific mAb by Western blotting.Actin expression was used to monitor loading ofeach lane with an equal amount of protein. AnLG-2 lymphoid cell lysate was used as a positivecontrol.
FIGURE 10. Restoration of low level of HLA class I expression following reconstitu-tion of mutant �2mC25W expression in FO-1 cells. Mock-transfected (FO-1.neo),�2mC25W-transfected (FO-1.�2.C25W.13, clone 13), and wild-type �2m-transfected (FO-1.�2) FO-1 cells were stained with an isotype-matched control mAb and the HLA classI-specific mAb W6/32 and analyzed by flow cytometry. The numbers above each openprofile are -fold MFI. Representative results of one of two experiments are shown. Thevariability in results between experiments is less than 5%.
�2-Microglobulin Dysfunction and Antigen Presentation Defects
18772 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 281 • NUMBER 27 • JULY 7, 2006
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

have shown that such mutations do not affect the level of proteinexpression (26). However, this is not the case for �2mC25W presentedhere. The steady-state level of �2mC25W in VMM5B cells is �29-foldlower than that of its wild-type counterpart inVMM5Acells. Such a lowlevel of accumulation could be explained by the increased degradationof �2mC25W because of the structural instability caused by the Cys-to-Trp substitution at position 25. It is likely that a drastic alteration in themode of folding of the nascent �2m polypeptide chain occurs, resultingin either a folded protein with an altered three-dimensional conforma-tion or an unfolded protein with a thermodynamically unstable struc-ture. The latter possibility is supported by the results of our structuralanalysis because: (i) the strong steric hindrance by the Trp side chain islikely to interfere with the hydrophobic interaction between the two�-pleated sheets, and (ii) the disappearance of the disulfide link betweenCys-25 and Cys-80 may not keep �2m in an energetically favored con-formation. The accumulation of �2mC25W in VMM5B cells incubatedwith a proteasome inhibitor further suggests that �2mC25W is unstableand ready for proteasome-mediated degradation. On the other hand,the predicted unfolding effect of the C25W substitution may not occurwhen Cys is replaced with a small, hydrophobic amino acid, such asalanine. In this situation, no apparent steric hindrance would take place,and the two �-pleated sheets may remain closely contacted, resulting inthe proper folding of the �2m protein. The importance of disulfidebonds in the structure and function of a protein has been indicated inseveral examples; one is represented by the replacement of Cys at posi-tion 634 of the RET (rearranged during transfection) proto-oncogeneproduct. This type of mutation is present in 85% of individuals sufferingfrom multiple endocrine neoplasia 2A (27).mAb W6/32, which recognizes a conformational epitope expressed
by the �2m-associated HCs, detected a low level of HLA class I mole-cules expressed on the surface of VMM5B cells and inside them. Thisphenomenon is likely to result from a stable interaction between a smallpopulation of aberrantly folded �2mC25W and HCs because the amountof HC-�2mC25W complexes was not increased significantly in IFN-�-andMG132-treated VMM5B cells despite themarked elevation of indi-vidual HC and �2mC25W. The inability of the HCs associated with aber-rantly folded �2mC25W to receive their peptide ligands, even when thecognate peptide has a high binding affinity (e.g. tyrosinase369–377D,YMDGTMSQV, t1⁄2 � 212.6 min), may result from the lack of properlyconformed peptide binding grooves on the HC-�2mC25W heterodimer.This possibility is supported by the lack of refolding on the cell surface ofthemousemajor histocompatibility complexmoleculeH-2DdHC asso-ciated with an artificially mutated �2m (�2mW60A) following loadingwith exogenous peptides (28). In the latter situation, �2mW60A acts in adominant-negative fashion. On the other hand, we do not know at pres-ent whether the formation of HC-�2mC25W complexes occurs at thestage of co-translational translocation of individual HC/calnexin and�2mC25W into the ER or at a later stagewhen each of themature proteinsare present.It is noteworthy that not all the tumor cells in the metastasis, from
which the VMM5B cell line had been derived, carry the described anti-gen presentation defect because the freshly isolated tumor cells showeda certain degree of susceptibility to tyrosinase-specific CTLs, asreported in our previous study (7). In this regard, the VMM5B cell line islikely to represent a subpopulation of tumor cells in the metastasis thathas escaped control by CTLs in the course of the disease. Also worthy ofmentioning is that the patient did not receive any type of immunother-
apy in the course of the disease. Therefore, the elicited CTL responsewas spontaneous in nature as opposed to the response introduced by Tcell-based immunotherapy. This is also reflected by the location of theidentified �2m gene mutation (TGC to TGG in exon 2), outside themutational hot spot in exon 1, which in melanoma has been foundmostly in patients exposed to T cell-based immunotherapy-relatedimmune selective pressure (29). Whether this type of mutation willrepresent a common one among malignant cells exposed to spontane-ous CTL immune selective pressure remains to be determined. From apractical viewpoint, the resistance of VMM5B cells to lysis by CTLsdespite HLA class I expression on the cell surface represents anotherexample of lack of correlation betweenHLAclass I expression andCTL-mediated lysis in malignant cells (30). Understanding the mechanismsunderlying this phenomenon as reported here may contribute to defin-ing the molecular basis of tumor escape and optimization of the criteriato select patients to be treated with immunotherapy.
REFERENCES1. Saper, M. A., Bjorkman, P. J., and Wiley, D. C. (1991) J. Mol. Biol. 219, 277–3192. Cresswell, P., Bangia, N., Dick, T., and Diedrich, G. (1999) Immunol. Rev. 172, 21–283. Yewdell, J. (guest ed.) (2002)Mol. Immunol. 39, 125–2594. Hansen, T. H., and Lee, D. R. (1997) Adv. Immunol. 64, 105–1375. Ferrone, S. (guest ed.) (2002) Semin. Cancer Biol. 12, 1–866. Seliger, B., Maeurer, M. J., and Ferrone, S. (2000) Immunol. Today 21, 455–4647. Yamshchikov, G. V., Mullins, D. W., Chang, C. C., Ogino, T., Thompson, L., Presley,
J., Galavotti, H., Aquila, W., Deacon, D., Ross, W., Patterson, J. W., Engelhard, V. H.,Ferrone, S., and Slingluff, C. L., Jr. (2005) J. Immunol. 174, 6863–6871
8. D’Urso, C. M., Wang, Z. G., Cao, Y., Tatake, R., Zeff, R. A., and Ferrone, S. (1991)J. Clin. Investig. 87, 284–292
9. Yamshchikov, G. V., Barnd, D. L., Eastham, S., Galavotti, H., Patterson, J.W., Deacon,D. H., Teates, D., Neese, P., Grosh,W.W., Petroni, G., Engelhard, V. H., and Slingluff,C. L., Jr. (2001) Int. J. Cancer 92, 703–712
10. Barnstable, C. J., Bodmer, W. F., Brown, G., Galfre, G., Milstein, C., Williams, A. F.,and Ziegler, A. (1978) Cell 14, 9–19
11. Wang, X., Liang, B., Rebmann, V., Lu, J., Celis, E., Kageshita, T., Grosse-Wilde,H., andFerrone, S. (2003) Tissue Antigens 62, 139–147
12. Rebai, N., and Malissen, B. (1983) Tissue Antigens 22, 107–11413. Stam, N. J., Vroom, T. M., Peters, P. J., Pastoors, E. B., and Ploegh, H. L. (1990) Int.
Immunol. 2, 113–12214. Sernee, M. F., Ploegh, H. L., and Schust, D. J. (1998)Mol. Immunol. 35, 177–18415. Perosa, F., Luccarelli, G. M., Favoino, E., Ferrone, S., and Dammacco, F. (2003) J. Im-
munol. 171, 1918–192616. Lampson, L. A., Fisher, C. A., and Whelan, J. P. (1983) J. Immunol. 30, 2471–247817. Bandoh, N., Ogino, T., Cho, H. S., Hur, S. Y., Shen, J., Wang, X., Kato, S., Miyokawa,
N., Harabuchi, Y., and Ferrone, S. (2005) Tissue Antigens 66, 185–19418. Wang, X., Campoli, M., Cho, H. S., Ogino, T., Bandoh, N., Shen, J., Hur, S. Y., Kages-
hita, T., and Ferrone, S. (2005) J. Immunol. Methods 299, 139–15119. Ogino, T., Wang, X., Kato, S., Miyokawa, N., Harabuchi, Y., and Ferrone, S. (2003)
Tissue Antigens 62, 385–39320. Kusama, M., Kageshita, T., Chen, Z. J., and Ferrone, S. (1989) J. Immunol. 143,
3844–385221. Sugawara, S., Abo, T., and Kumagai, K. (1987) J. Immunol. Methods 100, 83–9022. Ogino, T., Wang, X., and Ferrone, S. (2003) J. Immunol. Methods 278, 33–4423. Ramal, L. M., Feenstra, M., van der Zwan, A. W., Collado, A., Lopez-Nevot, M. A.,
Tilanus, M., and Garrido, F. (2000) Tissue Antigens 55, 443–44824. Bangia, N., Lehner, P. J., Hughes, E. A., Surman, M., and Cresswell, P. (1999) Eur.
J. Immunol. 29, 1858–187025. Mosse, C. A., Meadows, L., Luckey, C. J., Kittlesen, D. J., Huczko, E. L., Slingluff, C. L.,
Shabanowitz, J., Hunt, D. F., and Engelhard, V. H. (1998) J. Exp. Med. 187, 37–4826. Chen, H. L., Gabrilovich, D., Tampe, R., Girgis, K. R., Nadaf, S., and Carbone, D. P.
(1996) Nat. Genet. 13, 210–21327. Eng, C. (1999) J. Clin. Oncol. 17, 380–39328. Hill, D. M., Kasliwal, T., Schwarz, E., Hebert, A. M., Chen, T., Gubina, E., Zhang, L.,
and Kozlowski, S. (2003) J. Biol. Chem. 278, 5630–563829. Chang, C. C., Campoli,M., Restifo, N. P.,Wang, X., and Ferrone, S. (2005) J. Immunol.
174, 1462–147130. Chang, C. C., Campoli,M., and Ferrone, S. (2004)Curr. Opin. Immunol. 16, 644–650
�2-Microglobulin Dysfunction and Antigen Presentation Defects
JULY 7, 2006 • VOLUME 281 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 18773
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from

Yamshchikov, Nobuyuki Bandoh, Craig L. Slingluff, Jr. and Soldano FerroneChien-Chung Chang, Takeshi Ogino, David W. Mullins, Janine L. Oliver, Galina V.
-Microglobulin Loss-of-function in Melanoma Cells2βCaused by a Novel Defective Human Leukocyte Antigen Class I-associated Antigen Presentation
doi: 10.1074/jbc.M511525200 originally published online April 28, 20062006, 281:18763-18773.J. Biol. Chem.
10.1074/jbc.M511525200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/281/27/18763.full.html#ref-list-1
This article cites 30 references, 7 of which can be accessed free at
by guest on May 12, 2020
http://ww
w.jbc.org/
Dow
nloaded from