deafferentation of neighbouring motor cortex areas does not further enhance saturated...
TRANSCRIPT

www.elsevier.com/locate/clinph
Clinical Neurophysiology 119 (2008) 886–891
Deafferentation of neighbouring motor cortex areas does not furtherenhance saturated practice-dependent plasticity in healthy adults q
Michael Walther a,*, Nicola Kuhnke a, Joachim Schessl b, Igor Delvendahl a,Nikolai Jung a, Dirk Kreml c, Ulf Ziemann d, Volker Mall a
a Department of Neuropediatrics and Muscle Disorders, University Children’s Hospital, Freiburg, Germanyb Division of Neurology, The Children’s Hospital of Philadelphia, University of Pennsylvania, Philadelphia, USA
c Department of Anaesthesia, University Hospital Freiburg, Germanyd Department of Neurology, Johann Wolfgang Goethe-University of Frankfurt, Germany
Accepted 8 December 2007Available online 20 February 2008
Abstract
Objective: To assess effects of deafferentation of the arm representation of primary motor cortex (M1) on practice-dependent plasticity inhealthy adults.Methods: Twelve healthy, right-handed adults (18–48 years, median 20.2 years) performed two consecutive experiments (exp. 1 and exp. 2).Exp. 1 consisted of a motor practice (MP) of repeated ballistic flexion movements of the left thumb. This was followed by exp. 2 consisting ofselective anaesthesia of the upper brachial plexus (SPA) to disinhibit the training M1 and a second period of the same MP. Peak accelerationof the trained thumb movement and the motor evoked potential (MEP) amplitude in the flexor pollicis brevis muscle elicited by single-pulsetranscranial magnetic stimulation of the training M1 were studied before and after exp. 1 and after exp. 2.Results: After exp. 1 all subjects demonstrated an increase of peak acceleration (baseline: 19.23 ± 3.81 m s�2; after exp. 1:43.28 ± 17.63 m s�2, p = 0.008) and MEP amplitude (from 0.46 ± 0.23 mV to 1.26 ± 0.77 mV, p = 0.03). There was no additionalincrease of these measures after exp. 2 (44.37 ± 19.56 m s�2, p = 0.78, 1.69 ± 1.21 mV (p = 0.07)).Conclusions: Training of ballistic thumb movements leads to behavioural improvement as well as to an increased excitability of the cor-responding M1 representation. These effects do not increase further during deafferentation of the training M1. In contrast to strokepatients [Muellbacher W, Richards C, Ziemann U, Wittenberg G, Weltz D, Boroojerdi B, et al. Improving hand function in chronicstroke. Arch Neurol 2002;59:1278–82], practice-dependent plasticity in healthy subjects cannot be enhanced by deafferentation of neigh-bouring motor cortex areas.Significance: Healthy subjects, in contrast to patients with central motor lesions, are capable of saturating practice-dependent plasticityto a level that cannot be further enhanced by experimental manipulation.� 2008 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.
Keywords: Neuronal plasticity; Motor learning; Motor practice; Temporary cortical deafferentation; Transcranial magnetic stimulation
1. Introduction
There is increasing evidence that bidirectional modifica-tion of intracortical pathways, namely long-term depres-sion (LTD)-like and long-term potentiation (LTP)-like
1388-2457/$34.00 � 2008 International Federation of Clinical Neurophysiolog
doi:10.1016/j.clinph.2007.12.006
q This study was supported by a grant of the Deutsche Forschungs-gemeinschaft (DFG MA 3306/1-1).
* Corresponding author. Tel./fax: +49 761 2704344.E-mail address: [email protected] (M. Walther).
mechanisms of layer II/III pyramidal cell synapses, is abasic mechanism of motor learning in animals and humans(Classen et al., 2004; Rioult-Pedotti et al., 1998; Rioult-Pedotti et al., 2000; Stefan et al., 2002; Ziemann, 2004; Zie-mann et al., 2006). Experiments in rats proved a direct linkbetween motor learning and LTP in M1 (Rioult-Pedottiet al., 1998; Rioult-Pedotti et al., 2000). Similarly, inhumans, transcranial magnetic stimulation (TMS) studiesand the paired associative stimulation (PAS) (Stefanet al., 2000; Wolters et al., 2003) protocol revealed LTP-
y. Published by Elsevier Ireland Ltd. All rights reserved.

M. Walther et al. / Clinical Neurophysiology 119 (2008) 886–891 887
like mechanisms as an important mechanism of practice-dependent plasticity (Ziemann, 2004; Ziemann et al.,2006; Stefan et al., 2006). In these studies, the authors con-cluded, that changes of the MEP amplitude share similarmechanisms as LTP/LTD defined in slices of experimentalanimal preparations. Recent studies targeted the questionwhether practice-dependent plasticity and LTP-likechanges in MEP amplitude can be purposefully modulatedin order to enhance practice outcome.
Experimental selective deafferentation has been used inseveral settings to increase motor cortex excitability bymeans of disinhibition (Levy et al., 2002) in order toimprove motor learning. An ischemic nerve block of theforearm was used for experimental deafferentation reveal-ing a positive effect on motor learning as well as on LTP-like plasticity in healthy subjects (Ziemann et al., 2001).Muellbacher and colleagues used a motor practice (MP)consisting of repeated ballistic pinching movements anddemonstrated an increase of the MEP amplitude after atraining period of 60 min (Muellbacher et al., 2001). If thisMP was combined with selective upper arm deafferentation(SPA) in chronic stroke patients, who had performed thisMP extensively beforehand to reach a plateau level, anadditional significant gain of motor performance of thetrained movement and a further increase in MEP ampli-tude could be induced (Muellbacher et al., 2002).
This study was performed to assess to which extent prac-tice-dependent plasticity can be enhanced by SPA inhealthy subjects.
2. Methods
2.1. Subjects
Twelve healthy, right-handed adults (7 females, range:18–48 years; mean age: 20.2 years) were included. Thisstudy was approved by the Local Ethics Committee ofthe University hospital of Freiburg, Germany. All subjectsgave their written informed consent.
2.2. Motor practice (MP)
Subjects practiced fastest possible thumb flexion move-ments by performing brief contractions of their left flexorpollicis brevis (FPB) muscle (Muellbacher et al., 2001).Movements were externally paced by a metronome at a fre-quency of 0.25 Hz. Subjects sat in a chair, and to ensurethat training performance was not artificially upgradedby forearm pronation movements, the forearm was firmlystabilised in a neutral position between pronation and supi-nation in a table fixed cast. To guarantee MP specificallydriven by the FPB, the distal phalangeal joint was rigidlytaped. A piezo crystal was fixed on the end-phalanx ofthe practicing thumb to measure acceleration of the thumbflexion movements. The piezoelectric accelerometer and thesignal amplifier were custom-made by the technical labora-tory of the University of Freiburg. The sensitivity of the
accelerometer was 2 mV/g. The signal was digitized at arate of 5.000 Hz. Data were processed and stored in a per-sonal computer. Offline analysis was done with the Spike2�
software from CED (Cambridge Electronic Design Ltd.,Cambridge, UK). For visual feedback of MP performance,the acceleration of each thumb flexion appeared online ona screen in front of the subject. MP in experiment 1 andexperiment 2 was divided into blocks of 10 min duration(exp. 1: 6 blocks; exp. 2: 4 blocks), separated by 5 minbreaks to prevent fatigue.
2.3. Selective plexus anaesthesia (SPA)
Temporary deafferentation was done by selectiveanaesthesia of the upper brachial plexus accomplishedby a neurostimulator device (Stimuplex�, FA BraunMelsungen, Melsungen, Germany). To identify the correctplacement of the tip of an insulated needle, evoked con-tractions in the deltoid or biceps muscles indicated thatthe tip of the needle lies close to the upper roots of thebrachial plexus. SPA was done and controlled by an expe-rienced anaesthesiologist. The dosage of the anaestheticwas chosen to result in deafferentation of the upper armfor approximately 60 min (lidocaine (15 mg/ml), dosage:10 ml). Effectiveness and selectiveness of SPA were mea-sured 15 min after the injection of the anaesthetic andat the end of anaesthesia by a standardised protocol. Asuccessful SPA was defined as an increase of the sensorythreshold in the shoulder and lateral upper arm (derma-tome C5), but sparing the lower cervical dermatomes offorearm and hand. Sensory threshold was measured bySemmes–Weinstein monofilament diameter size (the diam-eters provide a logarithmic scale of force exerted, andthus, a linear and interval scale of perceived intensity)(Semmes–Weinstein Von Frey Monofilaments; StoeltingCo., Wood Dale, Ill). In addition, the following criteriawere required for a successful SPA: a decrease in strength(measured by the Medical Research Council [MRC] Scale)of shoulder and upper arm muscles, sparing the hand andforearm; no change in the pinch force between thumb andindex finger (measured by a pinch-gauge; measurement ofthree trials per time point); and no change in the MEPamplitude of the FPB.
2.4. Study design
Experiment 1 (exp. 1) consisted of 60 min of MP andexperiment 2 (exp. 2) of SPA followed by 40 min of MP.In Exp. 2 the MP duration was shorter because TMS eval-uation of M1 excitability was aimed at being completedwithin the duration of SPA effect, which vanished off after60 min. Exp. 2 followed exp. 1 immediately.
2.5. TMS
Focal single-pulse TMS was performed using a Mag-stim 200 magnetic stimulator (Magstim Company, Whit-

888 M. Walther et al. / Clinical Neurophysiology 119 (2008) 886–891
land, UK) with a monophasic current waveform. Sub-jects were seated up-right on a chair and asked to relax.Relaxation was controlled online via high-gain visualfeedback of the EMG signal, and trials contaminatedby voluntary EMG were discarded from analysis. A fig-ure-eight-shaped stimulation coil was centred tangentiallyon the scalp over the hand area of the right M1, andpositioned at the site where maximum MEP amplitudeswere consistently obtained in the target muscle (leftFPB). Coil position was kept constant throughout themeasurements. The coil handle pointed in a posteriordirection and laterally at a 45� angle away from the mid-line. Thus, the current induced by TMS flowed approxi-mately perpendicular to the line of the central sulcus inorder to activate the corticospinal neurones mainly trans-synaptically (Brasil-Neto et al., 1992; Kaneko et al.,1996). MEPs in the FPB were recorded by surfaceEMG using the belly-tendon electrode montage. The cor-ner frequencies of the bandpass-filter were set at 2 Hzand 10 kHz, respectively. Data were amplified by a Toen-nies universal amplifier (Jager Toennies, Hochberg, Ger-many) connected with a 1401 micro data acquisition unit(CED Ltd., Cambridge, UK) with 5 kHz sampling rateand stored on a standard personal computer for offlineanalyses. TMS intensity was adjusted to elicit a MEPof 200–800 lV in peak-to-peak amplitude before training(baseline) and this intensity was kept constant through-out all measurements. MEP amplitude was measuredpeak-to-peak in the single trials (Muellbacher et al.,2002; Rossini et al., 1994; Ziemann et al., 2001) and thenaveraged over 15 trials for each time point of investiga-tion. Each subject underwent measurement of corticalexcitability before the beginning of MP (baseline), after60 min of MP (exp. 1) and after another 40 min of MP(exp. 2).
MP performance was assessed by measurements of peakacceleration of the practiced thumb flexion movement (seeabove).
2.6. Analyses and statistics
As basic behavioural measure, we averaged therecorded peak acceleration within the first 1 min ofMP in exp. 1 (baseline) and compared it with thepeak acceleration averaged between minutes 50 and60 of MP in exp. 1, and minutes 30 and 40 ofMP+SPA in exp. 2. For motor cortical excitability,MEP amplitude in the FPB was obtained before MPin exp. 1 (baseline) and compared with MEP ampli-tudes directly after MP in exp. 1 and after MP + SPAin exp. 2.
Statistical evaluation was performed by the non-para-metric Friedman two-way analysis of variance, and bythe non-parametric Wilcoxon test for pair-wise compari-sons. The level of significance was set to p = 0.05. If notindicated otherwise, all values are given as means ± stan-dard deviation.
3. Results
3.1. Selective plexus anaesthesia
In 9 of 12 subjects, selective plexus anaesthesia was suc-cessful according to the criteria defined in Section 2. In theother three subjects, the increase in sensory threshold in theC5 was too small, or the distal dermatomes were notspared, and therefore the criteria of a successful SPA werenot fulfilled. These three subjects were excluded from fur-ther data analysis. SPA was well tolerated by all subjectsand that there were no serious adverse events. The pair-wise comparison of the tip-pinch before and directly afterSPA onset revealed no significant change in the pinch-force(baseline: 7.06 ± 1.11 kg, after SPA onset: 6.49 ± 1.51 kg,p = 0.32).
3.2. Motor cortical excitability and behavioural
measurements
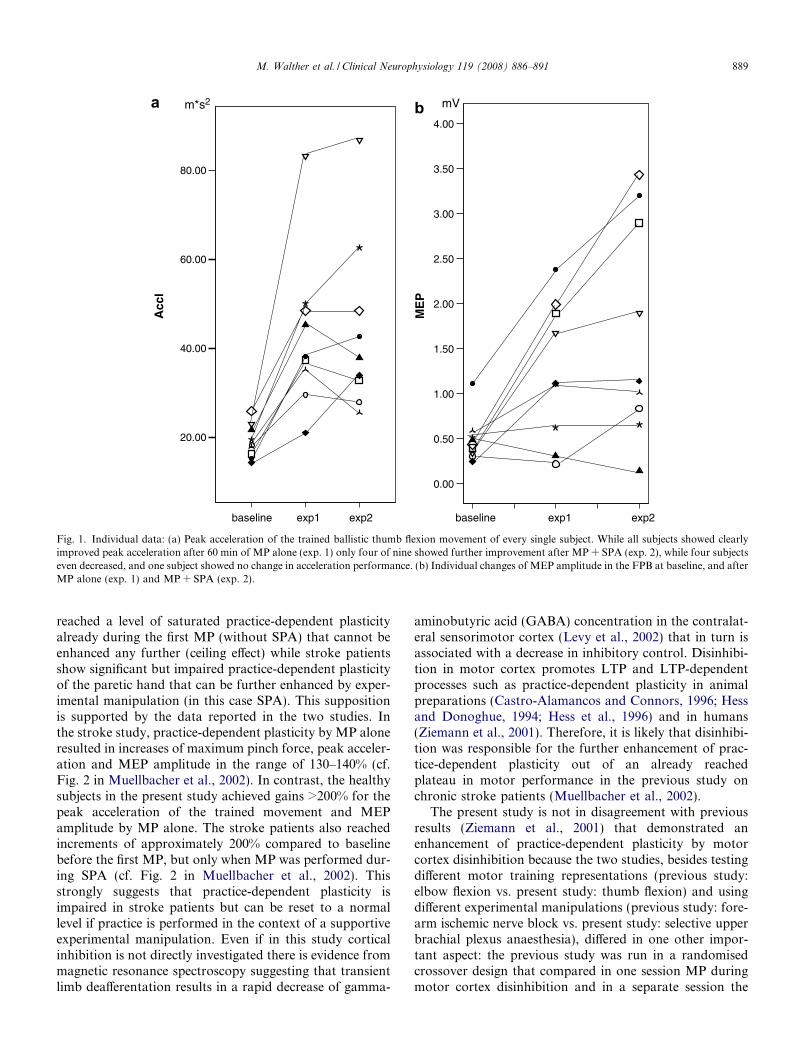
There was a significant effect of TIME of MEP amplitudein the FPB (p = 0.02), and on peak acceleration of thepracticed thumb flexion movement (p = 0.001). Post hocpair-wise comparisons showed after 60 min of MP in exp.1 a significantly increased MEP amplitude (baseline:0.46 ± 0.23 mV, after exp. 1: 1.26 ± 0.77 mV, p = 0.03),and a significantly increased peak acceleration (baseline:19.23 ± 3.81 m s�2; after exp. 1: 43.28 ± 17.63 m s�2,p = 0.008) (Figs. 1 and 2). Pair-wise comparisons of thedata after exp. 2 with those after exp. 1 did not show signif-icant further change in MEP amplitude (after exp. 2:1.68 ± 1.21 mV, p = 0.07) or peak acceleration (after exp.2: 44.37 ± 19.56 m s�2, p = 0.78) (Figs. 1 and 2).
4. Discussion
Our results demonstrate that in healthy adults practice-dependent plasticity induced in a repetitive thumb flexionmovement protocol cannot be enhanced by deafferentationof the training motor cortex.
These results are in contrast to previous findings instroke patients. Muellbacher and colleagues showed thatSPA enhanced practice-dependent plasticity as measuredby MEP amplitude and peak acceleration of the practicedmovement in stroke patients, after they had already per-formed extensive MP to reach a plateau in performanceof the trained pinching movement (Muellbacher et al.,2002). Even though there are some minor differences inthe experimental setup (movement rate in the stroke study0.3–0.5 Hz vs. 0.25 Hz in the present study; practice dura-tion 2 � 30 min in the stroke study vs. 6 � 10 min in thepresent study) it is unlikely that these differences played amajor role in explaining why SPA enhanced practice-dependent plasticity in chronic stroke patients but not inthe healthy subjects of the present study. The most likelyexplanation for the failure of SPA to enhance practice-dependent plasticity in healthy subjects further is that they
80.00
60.00
40.00
20.00
4.00
3.50
3.00
2.50
2.00
1.50
1.00
0.50
0.00
ME
P
Acc
l
m*s2 mV
baseline baselineexp1 exp1exp2 exp2
a b
Fig. 1. Individual data: (a) Peak acceleration of the trained ballistic thumb flexion movement of every single subject. While all subjects showed clearlyimproved peak acceleration after 60 min of MP alone (exp. 1) only four of nine showed further improvement after MP + SPA (exp. 2), while four subjectseven decreased, and one subject showed no change in acceleration performance. (b) Individual changes of MEP amplitude in the FPB at baseline, and afterMP alone (exp. 1) and MP + SPA (exp. 2).
M. Walther et al. / Clinical Neurophysiology 119 (2008) 886–891 889
reached a level of saturated practice-dependent plasticityalready during the first MP (without SPA) that cannot beenhanced any further (ceiling effect) while stroke patientsshow significant but impaired practice-dependent plasticityof the paretic hand that can be further enhanced by exper-imental manipulation (in this case SPA). This suppositionis supported by the data reported in the two studies. Inthe stroke study, practice-dependent plasticity by MP aloneresulted in increases of maximum pinch force, peak acceler-ation and MEP amplitude in the range of 130–140% (cf.Fig. 2 in Muellbacher et al., 2002). In contrast, the healthysubjects in the present study achieved gains >200% for thepeak acceleration of the trained movement and MEPamplitude by MP alone. The stroke patients also reachedincrements of approximately 200% compared to baselinebefore the first MP, but only when MP was performed dur-ing SPA (cf. Fig. 2 in Muellbacher et al., 2002). Thisstrongly suggests that practice-dependent plasticity isimpaired in stroke patients but can be reset to a normallevel if practice is performed in the context of a supportiveexperimental manipulation. Even if in this study corticalinhibition is not directly investigated there is evidence frommagnetic resonance spectroscopy suggesting that transientlimb deafferentation results in a rapid decrease of gamma-
aminobutyric acid (GABA) concentration in the contralat-eral sensorimotor cortex (Levy et al., 2002) that in turn isassociated with a decrease in inhibitory control. Disinhibi-tion in motor cortex promotes LTP and LTP-dependentprocesses such as practice-dependent plasticity in animalpreparations (Castro-Alamancos and Connors, 1996; Hessand Donoghue, 1994; Hess et al., 1996) and in humans(Ziemann et al., 2001). Therefore, it is likely that disinhibi-tion was responsible for the further enhancement of prac-tice-dependent plasticity out of an already reachedplateau in motor performance in the previous study onchronic stroke patients (Muellbacher et al., 2002).
The present study is not in disagreement with previousresults (Ziemann et al., 2001) that demonstrated anenhancement of practice-dependent plasticity by motorcortex disinhibition because the two studies, besides testingdifferent motor training representations (previous study:elbow flexion vs. present study: thumb flexion) and usingdifferent experimental manipulations (previous study: fore-arm ischemic nerve block vs. present study: selective upperbrachial plexus anaesthesia), differed in one other impor-tant aspect: the previous study was run in a randomisedcrossover design that compared in one session MP duringmotor cortex disinhibition and in a separate session the

0
1
2
3
mVa bm*s-2
0
20
40
60
baseline exp.1 exp.2 exp.1
Acc
l
ME
Pbaseline exp.2
Fig. 2. Motor cortical excitability and motor performance: Time course of MEP amplitudes in the primary practicing muscle, the flexor pollicis brevis (a)and peak acceleration of the trained ballistic thumb flexion movement (b) before (baseline) and after experimental manipulation (exp. 1 and exp. 2). Dataare depicted in box-plots with medians and quartiles. Abbreviations: m s�2, peak acceleration measured in the SI-unit m s�2; mV, millivolts; exp. 1, afterexp. 1 (60 min of MP); exp. 2, after exp. 2 (40 min MP + SPA).
890 M. Walther et al. / Clinical Neurophysiology 119 (2008) 886–891
same MP without motor cortex disinhibition. MP was per-formed at a low rate of 0.1 Hz that in the session withoutmotor cortex disinhibition resulted in only a very moderateincrease in MEP amplitude and no change in peak acceler-ation of the trained movement (Ziemann et al., 2001). If thesame MP was performed during motor cortex disinhibi-tion, it resulted in a significant gain in practice-dependentplasticity (Ziemann et al., 2001). In the present study, wechose the MP protocol of Muellbacher et al. (2001) whichby itself led already to a strong increase in MEP amplitudeand peak acceleration. We were interested in learning towhich extent fully expressed practice-dependent plasticitycan be further enhanced in the same session, if subse-quently combined with deafferentation of neighbouringmotor cortex areas. The findings indicate that this is notpossible in healthy subjects. Practice-dependent plasticity,once it has occurred and reached a plateau, cannot be dri-ven beyond the already achieved limits.
The observed saturation of practice-dependent plasticityin healthy adults is in accordance to the Bienenstock–Coo-per–Munro (BCM) theory (Bienenstock et al., 1982). TheBCM theory predicted bidirectionally modifiable synapses,being able to express both, LTP and LTD. The thresholdfor induction of LTP and LTD varies with the history ofpostsynaptic neuronal activity. If LTP has occurredrecently then this will result in an increase of the LTPinduction threshold while induction of LTD becomes morelikely (Bienenstock et al., 1982). This form of homeostaticmetaplasticity has been demonstrated recently at the sys-tems level of human motor cortex (Siebner et al., 2004; Ste-
fan et al., 2006; Ziemann et al., 2004). Therefore, it may beargued that homeostatic metaplasticity contributed to theinability of SPA to further enhance practice-dependentplasticity in the present experiments, because substantialMP-induced LTP-like plasticity had already occurred inthe first MP block that directly preceded the SPAintervention.
5. Conclusion
In contrast to chronic stroke patients, healthy adults arecapable of saturating practice-dependent plasticity in repet-itive movement tasks, and this effect cannot be enhancedfurther by motor cortical deafferentation. Therefore resultsfrom studies on plasticity in healthy subjects cannot be sim-ply transferred to neurological patients, and vice versa.
References
Bienenstock EL, Cooper LN, Munro PW. Theory for the development ofneuron selectivity: orientation specificity and binocular interaction invisual cortex. J Neurosci 1982;2:32–48.
Brasil-Neto JP, Cohen LG, Panizza M, Nilsson CG, Roth BJ, Hallett M.Optimal focal transcranial magnetic activation of the human motorcortex: effects of coil orientation, shape of the induced current pulse,and stimulus intensity. J Clin Neurophysiol 1992;9:132–6.
Castro-Alamancos MA, Connors BW. Spatiotemporal properties ofshort-term plasticity sensorimotor thalamocortical pathways of therat. J Neurosci 1996;16:2767–79.
Classen J, Wolters A, Stefan K, Wycislo M, Sandbrink F, Schmidt A,et al. Paired associative stimulation. Suppl Clin Neurophysiol2004;57:563–9.

M. Walther et al. / Clinical Neurophysiology 119 (2008) 886–891 891
Hess G, Aizenman CD, Donoghue JP. Conditions for the induction oflong-term potentiation in layer II/III horizontal connections of the ratmotor cortex. J Neurophysiol 1996;75:1765–78.
Hess G, Donoghue JP. Long-term potentiation of horizontal connectionsprovides a mechanism to reorganize cortical motor maps. J Neuro-physiol 1994;71:2543–7.
Kaneko K, Kawai S, Fuchigami Y, Morita H, Ofuji A. The effect ofcurrent direction induced by transcranial magnetic stimulation on thecorticospinal excitability in human brain. Electroencephalogr ClinNeurophysiol 1996;101:478–82.
Levy LM, Ziemann U, Chen R, Cohen LG. Rapid modulation of GABAin sensorimotor cortex induced by acute deafferentation. Ann Neurol2002;52:755–61.
Muellbacher W, Richards C, Ziemann U, Wittenberg G, Weltz D,Boroojerdi B, et al. Improving hand function in chronic stroke. ArchNeurol 2002;59:1278–82.
Muellbacher W, Ziemann U, Boroojerdi B, Cohen L, Hallett M. Role ofthe human motor cortex in rapid motor learning. Exp Brain Res2001;136:431–8.
Rioult-Pedotti MS, Friedman D, Donoghue JP. Learning-induced LTP inneocortex. Science 2000;290:533–6.
Rioult-Pedotti MS, Friedman D, Hess G, Donoghue JP. Strengthening ofhorizontal cortical connections following skill learning. Nat Neurosci1998;1:230–4.
Rossini PM, Barker AT, Berardelli A, Caramia MD, Caruso G, CraccoRQ, et al. Non-invasive electrical and magnetic stimulation of thebrain, spinal cord and roots: basic principles and procedures forroutine clinical application. Report of an IFCN committee. Electro-encephalogr Clin Neurophysiol 1994;91:79–92.
Siebner HR, Lang N, Rizzo V, Nitsche MA, Paulus W, Lemon RN, et al.Preconditioning of low-frequency repetitive transcranial magneticstimulation with transcranial direct current stimulation: evidence forhomeostatic plasticity in the human motor cortex. J Neurosci2004;24:3379–85.
Stefan K, Kunesch E, Benecke R, Cohen LG, Classen J. Mechanisms ofenhancement of human motor cortex excitability induced by interven-tional paired associative stimulation. J Physiol 2002;543:699–708.
Stefan K, Kunesch E, Cohen LG, Benecke R, Classen J. Induction ofplasticity in the human motor cortex by paired associative stimulation.Brain 2000;123:572–84.
Stefan K, Wycislo M, Gentner R, Schramm A, Naumann M, Reiners K,et al. Temporary occlusion of associative motor cortical plasticity byprior dynamic motor training. Cereb Cortex 2006;16:376–85.
Wolters A, Sandbrink F, Schlottmann A, Kunesch E, Stefan K, CohenLG, et al. A temporally asymmetric Hebbian rule governing plasticityin the human motor cortex. J Neurophysiol 2003;89:2339–45.
Ziemann U. TMS and drugs. Clin Neurophysiol 2004;115:1717–29.Ziemann U, Ilic TV, Jung P. Long-term potentiation (LTP)-like
plasticity and learning in human motor cortex-investigations withtranscranial magnetic stimulation (TMS). Suppl Clin Neurophysiol2006;59:19–25.
Ziemann U, Ilic TV, Pauli C, Meintzschel F, Ruge D. Learning modifiessubsequent induction of long-term potentiation-like and long-termdepression-like plasticity in human motor cortex. J Neurosci2004;24:1666–72.
Ziemann U, Muellbacher W, Hallett M, Cohen LG. Modulation ofpractice-dependent plasticity in human motor cortex. Brain2001;124:1171–81.