cytoplasmic cd3+ surface cd8+ lymphocytes develop as a thymus-independent lineage in chick-quail...
TRANSCRIPT

Eur. J. Immunol. 1989.19: 1449-1455
R. Pat Bucy', Monique Coltey", Chen-lo H. Chen', David Chap, Nicole M. Le Douarin" and Max D. Cooper"
Departments of Pathology', Microbiologym, Pediatrics' and Medicine', The University of Alabama at Birmingham and The Howard Hughes Medical Institute', Birmingham, and Institut d'Embryologie, Nogent-sur-Marne"
Thymus-independent development of CD3 'ITcR- cells 1449
Cytoplasmic CD3' surface CD8' lymphocytes develop as a thymus-independent lineage in chick-quail chimeras*
We have analyzed the embryonic development of a population of lymphoid cells that express a CD3 antigenic determinant in the cytoplasm but not on the cell surface. Since these cells lack T cell receptor (TcR) molecules, we have provisionally named them TCRO cells. Their development, expansion and distribution was investigated following transplantation of splenic and bursa1 fragments from chicken embryos into quail embryos. Since quail cells are not recognized by our panel of monoclonal antibodies against chicken TcRl, TcR2, TcR3, CD3, CD4 and CD8 molecules, these antibodies provided reliable markers for donor chick lymphocytes in the tissues of the quail recipients. Transplanted spleen and bursa both generated CD3+ cells, the number of which increased as a function of age. Notably, approximately half of these CD3' cells expressed surface CD8, but none acquired TcRl ( y /6 ) , TcR2 (a/@) or TcR3 expression. Since TCRO cells normally appear first in the spleen of 8-day chick embryos (E8), their generation in E6 splenic transplants indicated an extrathymic origin. The TCRO cells of chick splenic origin migrated to the spleen, bursa and thymus of the quail recipients. In six of seven chimeras acquiring CT3' cells in the recipient thymus, these cells were restricted to the medulla and displayed the typical TCRO phenotype: CD~+CD~+TCR~-TCR~-TCR~-. These intrathymic TCRO cells also lacked the CT1 thymocyte antigen. We conclude that the TCRO cells represent a thymus-independent lineage of lymphoid cells that can migrate into a receptive thy- mus but rarely, if ever, differentiate into conventional T cells.
1 Introduction
The development of mAb that recognize the chicken TcR complex and the CD4 and CD8 accessory molecules has allowed detailed analysis of avian T cell development [l-61. Cells expressing the CD3/TcR1 (y/6) receptor complex are generated first in the embryonic thymus, and CD3iTcR2 (a/@- bearing cells are acquired later. The TcR2 cells, like their mammalian counterparts, express both CD4 and CD8 as immature thymocytes, while mature T cells typically express one or the other. The histologic localization patterns of the TcRl+ (y/S) and TcR2' (a@) cells are distinctive in both thy- mus and peripheral lymphoid tissues [6]. TcRl cells are prefer- entially localized in the splenic sinusoids and the intestinal epithelium, while TcR2' cells are found in the splenic periar- teriolar sheaths and in the lamina propria of the intestine. A majority of the TcR1' cells acquire surface CD8 molecules in the chicken spleen and intestine but few, if any, express CD4 [3, 61. These distinctive tissue localization patterns and the expression of CD8 by TcRl cells in peripheral tissue are con- served in humans [7]. TcR3 is a distinct heterodimer found in association with CD3 on the surface of a subset of T cells which express neither TcRl nor TcR2 [8]. A mammalian homologue for TcR3 has not been identified. TcR3' cells are phenotypically similar to TcR2 cells, since mature cells express either CD4 or CD8 surface molecules and occupy similar his- tologic microenvironments.
[I 76361 *
A HHMI investigator.
Correspondence: R. Pat Bucy, Department of Pathology, Rm. 518 LHR, UAB Station, Birmingham, AL 35294, USA
Abbreviations: E9: Embryonic day 9 E3: Embryonic day 3, etc.
Supported by grants CA 16673 and CA 13148, awarded by the National Cancer Institute.
In the course of analyzing the ontogeny of T cell development in the chick, we* and others (Houssaint, E. and Boyd, R., personal communication) have observed a population of CD3' cells in peripheral lymphoid organs early in ontogeny. These cells express a CD3 determinant in the cytoplasm, but they express neither CD3 nor TcR on the cell surface. A sub- population of these cells express surface CD8, but none express CD4. For convenience, we have called the cytoplasmic CD3+/TcR- lymphocytes to TCRO cells. The TCRO cells con- stitute a sizeable population of cells in adult birds, although their anatomical distribution is restricted. Approximately 30%-40% of the CD3' lymphocytes in sections of the intesti- nal epithelium are TCRO cells which contain cytoplasmic CD3 but no surface CD3/TcR complexes*. Cells with this phenotype are relatively rare in the adult spleen, comprising approximately 1% of all CD3' splenocytes.
The origin and fate of the TCRO cells was examined in the present experiments by constructing interspecies chick-quail chimeras [9-111, taking advantage of the fact that the murine mAb to the chicken CD3, CD4, CD8, TcRl and TcR2 molecules do not cross-react with quail T cells [l-3, 51. The development and distribution of chick T cells can thus be traced in the chimeras. The key issue that we have examined is the lineal relationship between the TCRO cells, that are sur- face CD3-/cytoplasmic CD3', and conventional T cells that express TcWCD3 complexes on their surface. Three alterna- tives were considered. First, the TCRO cells could be the precursors of TcR-bearing cells. Second, TCRO cells could represent cells with aberrantly rearranged TcR genes which emigrate from the thymus as sterile T cells. Finally, they could represent an independent lymphoid cell lineage that is only distantly related to the thymus-derived T cell. This issue was addressed by examining quail embryo recipients of spleen and
* R. P. Bucy, C. H. Chen and M. D. Cooper, manuscript in prepara- tion.
0 VCH Verlagsgesellschaft mbH, D-6940 Weinheim, 1989 0014-2980/89/0808-1449$02.50/0

1450
bursal transplants obtained from chick embryos prior to their infiltration with T cells. Our observations suggest that TCRO cells are an independent cell lineage.
R. P. Bucy, M. Coltey, C. H. Chen et al. Eur. J. Immunol. 1989.19: 1449-1455
2 Materials and methods
2.1 Animals
Fertile eggs of FP white leghorn chickens were obtained from Hy-Line International, Dallas Center, IA, and were hatched and raised in our facility. Fertile Bob-White quail eggs were obtained from Moore’s quail farm, Cullman, AL.
2.2 mAb
Murine IgGl mAb CT3, CT4, CT8 and TcRl were produced in our laboratories and are specific for the chicken CD3, CD4, CD8 and TcR y/6 molecules, respectively [ 1-31. Murine mAb TcR2, the kind gift of Drs. J. Cihak and U. Losch, is specific for the chicken T c R d p [5]. Murine mAb TcR3 recognizes a third TcR in the chicken [8] for which a mammalian homolo- gue has not been identified. Murine mAb CT-1 and CT-la both recognize >90% of chicken thymocytes, but only the CT-1 cross-reacts with quail thymocytes [ 121.
2.3 Surgical procedure
The spleen and bursa were surgically removed from chick embryos (E6, E9 or E l l ) . The Ell organs were cut into frag- ments of 0.2-0.4 mm in diameter before transplantation, while the entire organ from the less mature embryos was trans- planted. The donor tissue was placed in the somatopleure of E3 quail embryos between the fore and hind limb buds. The eggs were closed with transparent adhesive tape and returned to the incubator.
2.4 Immunoperoxidase analysis
Individual quail embryos with chick tissue transplants were removed from the egg, and the chick transplant, quail spleen, bursa, thymus and intestine were frozen in a block of OCT compound (Fisher Scientific, Pittsburgh, PA) in liquid nitro- gen and stored at -80°C. Four-micron frozen sections were cut, placed on glass slides, immediately fixed in 100% acetone for 5 min and dried at -30°C. Slides were then placed in a humidified staining chamber, at room temperature and
covered for 20 min with PBS (pH 7.4) containing 10% heat- inactivated normal horse serum to block nonspecific protein binding sites. The sections were incubated successively with the primary mAb, biotinylated horse anti-mouse IgG (Vector Labs, Burlingame, CA), the avidin-biotin peroxidase complex (ABC elite, Vector Labs), and 3,3‘-diaminobenzidine (Sigma, St. Louis, MO) with 0.01% Hz02 [13]. Between each incuba- tion the slides were washed 3 times with PBS. In some experi- ments, endogenous peroxidase activity was inhibited by incu- bation with 0.3% H202 in methanol for 30 min after the secondary antibody and before the ABC step. The sections were counterstained with 1% methyl green (Sigma), dehy- drated in graded alcohols, cleared in xylene and mounted for microscopical observation. Serial sections of each set of tissues were individually stained with different mAb and examined microscopically using a calibrated ocular grid. Cells with a “rim pattern” of peroxidase reaction product in each tissue were counted in multiple fields and the numbers of nuclei were counted to allow calculation of the percentage of positive cells. Cells which contained endogenous peroxidase enzyme were excluded from analysis on the basis of the morphological stain- ing pattern and inhibition with 0.3% H202 in methanol. Cells with endogenous peroxidase were stained with a dense, granu- lar staining pattern and were present on control slides with no primary mAb.
2.5 Simultaneous staining of surface and cytoplasmic antigen
Suspensions of viable cells were indirectly stained using unlabeled primary mAb followed by FITC-conjugated anti- mouse IgG [Southern Biotechnology Associates (SBA), Bir- mingham, AL]. These cells were cytocentrifuged onto glass slides, air-dried and fixed in 100% acetone. The cell prepara- tions were then stained with a second unlabeled mAb and, after washing, TRITC-conjugated, anti-mouse IgG (SBA) was applied. The cells were examined with a microscope equipped with epifluorescence optics. Cells which contained only cyto- plasmic antigen were identified as cells that stained positive with TRITC but not with the FITC surface stain.
3 Results
3.1 Development and distribution of TCRO cells in quail embryos receiving chick spleen or bursal transplants
Quail embryos transplanted with splenic fragments from day 9 chick embryos (E9) were examined every 2 days, beginning 6 days after transplantation, until the expected time of hatch-
A A Spleen C T 3 + (B) C T 8 + .--a W-W Bursa C T 3 + A
i \ T C R ~ + n --A 0.20 ci T C R Z + v -c 0 3.0
Figure 1 . Development of CT3’ cells in transplants of embryonic chick spleen (E9) and in the quail embryo recipient. Percentage of CT3+, CTV, TcRlt and TcR2’ cells in the transplanted chick
6 8 10 12 1 4 16 18 11 T4 -1’, spleen (A) and quail tissues (B) in a series of E3 quail embryos transplanted with E9 chick splenic tissue.
, . 0. 0.05 ..
I C
I -
Days af ter Transplantat ion
Eur. J . Immunol. 1989.19: 1449-1455
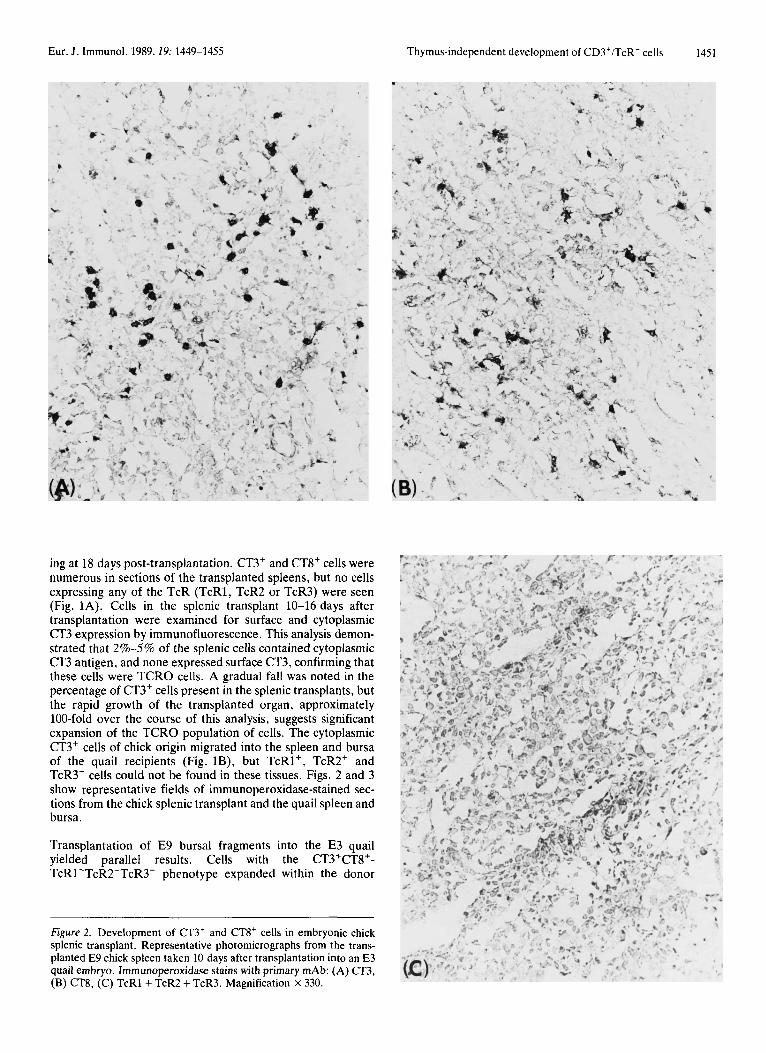
ing at 18 days post-transplantation. CT3+ and CT8' cells were numerous in sections of the transplanted spleens, but no cells expressing any of the TcR (TcR1, TcR2 or TcR3) were seen (Fig. 1A). Cells in the splenic transplant 10-16 days after transplantation were examined for surface and cytoplasmic CT3 expression by immunofluorescence. This analysis demon- strated that 2%-5% of the splenic cells contained cytoplasmic CT3 antigen, and none expressed surface CT3, confirming that these cells were TCRO cells. A gradual fall was noted in the percentage of CT3' cells present in the splenic transplants, but the rapid growth of the transplanted organ, approximately 100-fold over the course of this analysis, suggests significant expansion of the TCRO population of cells. The cytoplasmic CT3' cells of chick origin migrated into the spleen and bursa of the quail recipients (Fig. lB), but TcRl', TcR2' and TcR3' cells could not be found in these tissues. Figs. 2 and 3 show representative fields of immunoperoxidase-stained sec- tions from the chick splenic transplant and the quail spleen and bursa.
Transplantation of E9 bursa1 fragments into the E3 quail yielded parallel results. Cells with the CT3+CT8+- TcRl-TcR2-TcR3- phenotype expanded within the donor
Thymus-independent development of CD3'ITcR- cells 1451
Figure 2. Development of CT3' and CT8' cells in embryonic chick splenic transplant. Representative photomicrographs from the trans- planted E 9 chick spleen taken 10 days after transplantation into an E3 quail embryo. Immunoperoxidase stains with primary mAb: (A) CT3, (B) CT8, (C) TcRl + TcR2 + TcR3. Magnification x 330.

1452 R. P. Bucy, M. Coltey, C. H. Chen et al. Eur. J. Immunol. 1989.19: 1449-1455
Figure 3. Development of CT3' chick cells in embryonic quail reci- pients of chick spleen transplant. CT3 mAb immunoperoxidase stains of quail spleen (A) and quail bursa (B) with from chimeras trans-
bursal fragments, and they migrated to the quail spleen and bursa (Figs. 4 and 5). Here too, cells expressing TcR were not seen in either donor or recipient tissues. Since E l l is the ear- liest embryonic age at which CT3+ cells can be detected in the normal chick bursa*, these results indicate the presence of the relevant precursor cells in the E9 bursa of Fabricius.
Precursor stem cells normally enter the chick thymus begin- ning at E6.5 [ll], whereas CT3' cells appear first in the E8 spleen*. Thus, to examine further the possible extrathymic
* R. P. Bucy, C. H. Chen and M. D. Cooper, manuscript in prepara- tion.
planted with E9 chick spleen. Magnification X 330. Examination of serial sections revealed a few (38 ' cells, but no TcRl', TcR2+ or TcR3' cells (not shown).
origin for TCRO cells, fragments of E6 chick spleen were transplanted to E3 quails. In these experiments, TCRO cells were generated in the transplanted E6 chick spleen (Figs. 6 and 7) and migration of these cells to the quail spleen and bursa was observed.
3.2 Migration of chick TCRO cells to the quail thymus
In quail embryos receiving E9 or E l l chick spleen or bursal transplants, donor CT3' cells were often observed in the re- cipient thymus. In six of seven such instances, the CT3+ cells were confined to the medullary region of the lobe and lacked
Figure 4. Percentages of CT3+, CI'8+, TcRl+ and TcR2' cells in the trans- planted recipient chick bursa (A) and quail tissues (B). E3 quail embryos were transplanted with E9 chick bursal tissue fragments. The frequency of CT3+ cells in the chick bursa is relatively high but gradually decreases with growth of the tissue. Note the relatively low frequency of CT8 expression in the chick bursa compared to the splenic transplants. The same lower CT8 expression is also seen in the normal chick bursa (not shown).

Eur. J. Immunol. 1989.19: 1449-1455
TcR expression (Fig. 8, upper panel). Approximately 50% of these CT3+ cells were CT8+, but none expressed the chicken thymocyte antigen (CT-la) that is normally expressed by both cortical and medullary thymocytes [12]. These cells with the TCRO phenotype were observed in multiple thymic lobes in each of the six chick-quail chimeras.
In the seventh quail embryo that received an E9 splenic trans- plant, a small cluster of chick T cells was observed in the cortex of a single thymus lobe (Fig. 8, lower panel). These cells were CT3+CT4'CT8+TcW+CTla+ and TcR1-, the expected phenotype and histological location of a chick prothymocyte which has generated a TcR2 lineage colony.
4 Discussion
A population of lymphoid cells that can be identified by cyto- plasmic staining with the CT3 antibody and the absence of CD3/TcR complexes on their surface can be identified early in chick embryonic development. The cytoplasmic CD3+ lym- phocytes (TCRO), which frequently express surface CD8 molecules, first appear in the chick spleen on the 8th embry-
Thymus-independent development of CD3+EcR- cells 1453
Figure 5. Photomicrographs from the transplanted E9 chick bursa taken 14 days after transplantation into an E3 quail embryo. Magnifi- cation X 330. Immunoperoxidase stains with primary mAb: (A) CT3, (B) CT8, (C) TcRl + TcR2 + TcR3.

1454 R. P. Bucy, M. Coltey, C. H. Chen et al.
(i \ I I
0 0 5
\
Eur. J. Immunol. 1989.19: 1449-1455
I
I sues (B) in a senes of E3 quail embryos transplanted with E6 chick splenic tissue
~~
\
' / fragments The relative abundance of CT3+ cells is less in the transplanted E6
J A A '
4'
onic day*. Cytoplasmic CT3' thymocytes first appear on the ninth day of embryonic life (E9), around 3 days after precur- sor cells begin to enter the thymus [l l] . Cells expressing the TcRUCD3 complex then appear on day El2 [3], TcR2/CD3+ cells on El5 [5] and TcR31CD3' cells on E18. TCRO cells are thus acquired in peripheral lymphoid tissues prior to the devel- opment of recognizable pre-T cells and T cells in the thymus. Our present identification of TCRO cells in quail embryos receiving E6 chick spleen transplants demonstrates that these cells are generated from CD3- precursors outside of the thymus.
Since the TCRO population of cells arise extrathymically very early in development, it seemed possible that they represent prothymocytes. The concept of a cytoplasmic CD3' prothy- mocyte has some appeal, since immature cortical thymocytes may exhibit this phenotype [14] and the phenotype of acute T cell leukemias (T-ALL) considered to be the most immature in humans is the cytoplasmic CD3' leukemic cell [15, 161. By direct transfer assays in rodents, thymocyte precursors have been found in very low frequency within the BM (0.01% to 0.08% of BM cells; [ 17-20]). The possible expression of T cell antigens in the cytoplasm of the donor cells has not been examined in the direct transfer assays, but these cells lack both the CD4 and CD8 molecules [21].
Several characteristics of the TCRO cells argue strongly that they are not prothymocytes. First, there is no evidence that prothymocytes can express the CD8 surface molecule, whereas 50%-70% of TCRO cells have this phenotype. Sec- ond, TCRO cells comprise around 10% of E l4 splenocytes*, while thymocyte precursors are < 0.1% of bone marrow cells and are even more infrequent in spleen [ 17-20]. Third, TCRO cells generated in the chick spleen or bursa often migrate to the thymus of the quail embryo recipients where they fail to differentiate into conventional TcR expressing T cells. In six of the seven chimeric hosts acquiring CD3' in their thymus, the cells of chick spleen or bursa1 origin had the CT3+CT8+- TcRl-TcR2-TcR3- phenotype, failed to express the common thymocyte antigen CTla and were restricted to the medulla of the quail thymus. Cells with this phenotype were present in multiple thymic lobes in each of the six embryonic recipients. In the exceptional case, a solitary focus of cortical C D ~ + C D ~ + C D ~ + T C R ~ ' cells was identified. Since whole splenic tissue was transplanted, the phenotype of the precursor
* R. P. Bucy, C. H. Chen and M. D. Cooper, manuscript in prepara-
Figure 7. Photomicrographs from the transplanted E6 chick bursa taken 14 days after transplantation into an E3 quail embryo. Magnifi- cation X 400. Immunoperoxidase stains with primary mAb: (A) CT3, (B) TcRl + TcR2 + TcR3. tion.

Eur. J. Immunol. 1989.19: 1449-1455 Thymus-independent development of CD3+/TcR- cells 1455
Figure 8. The two patterns of chick T cells found in recipient quail thymus. The top panel represents the pattern in six of seven quail embryos which contain CT3' cells. These organs contain CT3' cells restricted to the medulla, but no TcR1' or TcR2' cells. These photomicrographs depict the thymus population pattern in a chimera transplant 16 days after transplantation of an E9 bursa. The lower panels demonstrate the single exception to the above pattern, and demonstrate a quail thymic lobe with a cortical focus of CT3+TcMC cells, but no TcRl' cells. Serial sections demonstrated the presence of CT4', CT8+ and CTla' cells in this focus (not shown).
of this TcR2' thymocyte colony cannot be established in the present experiment beyond the fact that it originated in the transplanted chick E9 spleen. The observation confirms, how- ever, that the embryonic quail thymus is receptive to chick prothymocytes and can support their differentiation along T cell lines [lo, 111. Thus, the observed failure of TCRO cells to undergo thymocyte differentiation, despite their frequent mi- gration to a receptive thymus, strongly suggests that T C R O cells are not prothymocytes.
We conclude that the extrathymic development and expansion of chick TCRO cells and their extensive migration into the various lymphoid organs of the quail imply that these cells represent a thymus-independent and bursa-independent lineage of widely disseminated lymphoid cells. The physiologic role of this lineage of cells is currently being examined.
We wish to thank Drs. J . Cihak and U. Losch for the TcR2 mAb, Jimin Li and Jane Schrimpf for excellent technical assistance, and Jeannie Cornelim and Ann Brookshire for help in preparing the manuscript.
Received May 11, 1989.
5 References
1 Chen, C. H., Ager, L. L., Gartland, G. L. and Cooper, M. D., J .
2 Chan, M. M., Chen, C. H., Ager, L. L. and Cooper, M. D.,
3 Sowder, J . T., Chen, C. H., Ager, L. L., Chan, M. M. and
Exp. Med. 1986. 164: 375.
J . Immunol. 1988. 140: 2133.
Cooper, M. D., J . Exp. Med. 1988. 167: 315.
4 Cihak, J., Ziegler-Heitbrook, H. W., Schranner, I., Mer- kenschlager, M. and Losch, U., Eur. J . Immunol. 1988. 18: 533.
5 Chen, C. H., Cihak, J., Lbsch, U. and Cooper, M. D., Eur. J . lm- munol. 1988. 18: 539.
6 Bucy, R. P., Chen, C. H., Cihak, J., Losch, U. and Cooper, M. D., J . Immunol. 1988. 141: 2200.
7 Bucy, R. P., Chen, C. H. and Cooper, M. D., J . lmmunol. 1989. 142: 3045.
8 Chen, C. H., Sowder, J. T., Lahti, J . M., Cihak, J., Losch, U. and Cooper, M. D., Proc. Natl. Acad. Sci. USA 1989. 86: 2352.
9 Le Douarin, N. and Jotereau, F., J . Exp. Med. 1975. 142: 17. 10 Jotereau, F. V., Houssaint, E. and Le Douarin, N., Eur. J . Immu-
11 Joterau, F. V. and Le Douarin, N., J . Immunol. 1982. 129: 1869. 12 Chen, C. H., Chanh, T. C. and Cooper, M. D., Eur. J . Immunol.
1984. 14: 385. 13 Hsu, S.-M., Raine, L. and Fanger, H., J . Histochem. Cytochem.
1981. 29: 477. 14 Swerdlow, S . H., Angermeier, P. A. and Hartman, A. L., Lab.
Invest. 1988. 58: 421. 15 Campana, D., Thompson, J. S. , Amlot, P., Brown, S. and
Janossy, G., J . Immunol. 1987. 138: 648. 16 Van Dongen, J. J. M., Krissansen, G. W., Wolvers-Tettero, I. L.
M., Comans-Bitter, W. M., Adriaansen, H. J . , Hooijkaas, H., Van Wering, E. R. and Terhorst, C., Blood 1988. 71: 603.
17 Katsura, Y., Kina, T., Amagai, T., Tsubata, T., Hirayoshi, K . , Takaoki, Y. , Sado, T. and Nishikawa, S . I., J . Immunol. 1986. 137: 2434.
nol. 1980. 10: 620.
18 Kadish, J. L. and Basch, R. S . , J . Exp. Med. 1976. 143: 1082. 19 Boersma, W., Betel, I . , Daculsi, R. and Van der Westen, G., Cell
20 Lepault, F. and Weissman, I. L., Nature 1981. 293: 151. 21 Fowlkes, B. J . , Edison, L., Mathieson, B. J. and Chused, T. M., J .
Tissue Kinet. 1981. 14: 179.
Exp. Med. 1985. 162: 802.