cyanobacteriaassociationsinshallowpolytrophiclakes ... associations in... · found in melangsee...
TRANSCRIPT

Original article
Cyanobacteria associations in shallow polytrophic lakes:influence of environmental factors
Ute Mischke
The Chair of Water Conservation, Brandenburg Technical University of Cottbus, Seestraße 45, 15526 Bad Saarow, GermanyIGB, Institute of Freshwater Ecology and Inland Fisheries, Department of Shallow Lakes and Lowland Rivers, Müggelseedamm 310,
12587 Berlin, Germany. Fax: +49-030-6418799.
Abstract
A 2 year field study in two shallow, polytrophic lakes (Langer See and Melangsee) in Brandenburg revealed the importance ofenvironmental factors in controlling the population dynamics of various cyanobacterial species which do not form layers or aggregations. Thespecies belonged to the toxin producing genera Cylindrospermopsis, Planktothrix, Aphanizomenon and Anabaena, and to the nontoxicmembers of Limnothrix and Pseudanabaena. In German waters, Cylindrospermopsis raciborskii is a neophyte, and exhibits a comparativelylow temperature requirement of about 17 °C for germination and growth. The nutrient concentrations were comparable in both the lakes, anddid not change measurably during the summer succession. The dissolved nitrogen fractions were well below 50 µg l–1 with total depletion ofnitrate. Soluble reactive phosphate concentrations never fell below 3 µg l–1 throughout the summer season. The light attenuation factors (Kd)of 3–5.8 m–1 indicate high self-shading. However, under calm weather conditions, the cyanobacteria were not concentrated in near surfacelayers and the species composition was relatively uniform in the vertical. In mid-summer, the diazotrophic Nostocales were replaced byOscillatoriales. Reduced light availability caused by mixing and self-shading is considered to be the main factor causing the summer declineof Nostocales. By means of two weather stations, more frequent, deeper and longer lasting mixing events could be observed in Langer See thanin the more shallow, but wind protected Melangsee. The success of Planktothrix agardhii may be due to the more turbulent mixing regime inLanger See. The algal association, S of Reynolds (Reynolds, C.S., 1997), should be modified, since the different species grouped together inS have different responses to turbulent mixing.
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.
Keywords: Phytoplankton succession; Cyanobacteria; Toxin algae; Cylindrospermopsis; Nostocales; Planktothrix; Net growth rates; Vertical distribution;Shallow lakes; Mixing regime
1. Introduction
Successions in shallow polytrophic lakes are highly ir-regular due to the overwhelming importance of meteorologi-cal conditions and episodic factors, such as wash out andsediment resuspension. Very shallow lakes (<5 m Zmax) are acommon lake type in northern Germany (Nixdorf andDeneke, 1997) and their phytoplankton is dominated bycyanobacteria. In such lakes, the summer successions of
phytoplankton are influenced by the duration of the intermit-tent stagnation phases, by the restricted mixing depth, nutri-ent availability being pulsed by resuspension, and a summerlight decrease by self-shading (Dokulil and Mayer, 1996;Nixdorf and Deneke, 1997). Thus, a high variability in theplanktonic summer succession can be expected. In contrast,Oscillatoriales either by Planktothrix or by Limnothrix spe-cies dominated several, very shallow polytrophic lakes inBrandenburg, Germany over several years (Rücker et al.,1997; Wiedner, 1999). Both Planktothrix agardhii and Lim-nothrix redekei are known to be turbulence tolerant species(Mur et al., 1993). In some study sites, however, the toxinproducing species P. agardhii is much more successful thanLimnothrix resulting in a higher toxicity (Wiedner and Nix-dorf, 1997). Small differences in terms of the extent ofsummer mixing, phosphate concentrations and light climate
Abbreviations: Cyl. rac, Cylindrospermopsis raciborskii; Pla. aga,Planktothrix agardhii; Lim./Pse, Limnothrix spp. and Pseudanabaena lim-netica; Aph. spp, Aphanizomenon spp; Other cyano, Other cyanobacterialspecies.
E-mail address: [email protected] (U. Mischke).
Acta Oecologica 24 (2003) S11–S23
www.elsevier.com/locate/actoec
© 2003 Éditions scientifiques et médicales Elsevier SAS. All rights reserved.DOI: 1 0 . 1 0 1 6 / S 1 1 4 6 - 6 0 9 X ( 0 3 ) 0 0 0 0 3 - 1

have been proposed as the key factors causing this domi-nance (Rücker et al., 1997; Wiedner, 1999).
The aim of this study is to quantify those differences in theenvironmental conditions which favor the occurrence ofPlanktothrix. For this purpose, in situ probes with data log-gers were used to characterize the mixing regime. Correla-tions between mixing, and cyanobacterial composition wereexplored. Even in frequently mixed shallow lakes, stagnationperiods occur. It is uncertain if the turbulence tolerant speciesare able to stratify in such situations. Thus, the depth profileof plankton composition during a stagnation period was alsoinvestigated in this study.
The recent colonization of shallow German lakes by thepotential toxin-producing cyanobacterium Cylindrosper-mopsis raciborskii (Krienitz and Hegewald, 1996; Wiedner,1999; Zippel, 1996) gave further impetus to this study. C.raciborskii is a successful invader (Padisák, 1997) worldwide. Low water temperatures were expected to be the mainbarrier of the northern expansion of this tropic and subtropi-cal species. We sought to establish if there is any graduallyadaptation of C. raciborskii to lower temperatures and thusan increasing propensity for it to replace other cyanobacteriain German lake waters.
2. Materials and methods
2.1. Study site and sampling
Melangsee and Langer See are two shallow lakes in theeastern part of Brandenburg in Germany. They are parts oftwo different lake systems with water retention times ofabout 30–60 d. The sampling stations were located near thelake centers, but the lake depth at the sampling site was notthe deepest point of Lake Langer See (Table 1). Samplingwas performed in the morning at 2–4 weekly intervals in1999, and usually every week in 2000 using a Ruttner-sampler. A depth profile was taken in 0.5 m steps and thesesamples were mixed together yielding a single-mixed (depth-integrated) sample for chemical and biological analyses. Onone occasion (25 July 2000), a vertical profile was sampled in0.2 m steps in both the lakes and the individual samples wereanalyzed separately.
2.2. Chemical and physical parameters
Water temperatures, oxygen saturation and pH were mea-sured in situ with a multi-parameter probe (Hydrolab Corpo-ration). Isotherm graphs were generated using the softwareISO (© Aquasoft, Clasen, Siegburg, Germany). Underwaterphotosynthetically active radiation (PAR) was measured us-
ing two scalar LI 193 SB Spherical Quantum Sensors (Li-Cor). Climatic data (air temperature, global radiation, windspeed and precipitation) were kindly provided by the meteo-rological station in Lindenberg located 15–30 km from thetwo lakes. In the summer of 2000, a weather station (fittedwith a data logger) was installed at each lake near the sam-pling station to measure the 15 min values of wind speed(m s–1), wind direction (sorted to the four main wind direc-tions) and the water temperature (°C) at three different waterdepths (Melangsee 0.5, 1.0 and 1.7 m; Langer See 0.5, 1.5and 2.7 m).
Mixed samples were analyzed for chlorophyll a, dissolvednutrients, soluble reactive phosphorus (SRP), nitrate, ammo-nia, and total phosphorus (TP) and total nitrogen (TN) ac-cording to standard methods. The sum of the dissolved nitro-gen fractions is given as DIN.
2.3. Phytoplankton analysis
Phytoplankton species were identified by light micro-scopic observation (Zeiss Axioskop) of living and Lugol-fixed samples. Commonly used taxonomic monographs re-lated to cyanobacteria (Anagnostidis and Komárek, 1985,1988; Komárek and Anagnostidis, 1989), were supple-mented with recently published original literature for speciesdiscrimination (Hindák, 1988; Horecká and Komárek, 1979;Kohl et al., 1985; Meffert, 1988; Meffert et al., 1981). Phy-toplankton biomass was determined in Lugol-fixed sub-samples by determination of cell concentration according tothe method of Utermöhl (1958) and estimation of cell vol-umes according to the method of Rott (1981) using an in-verted microscope (Zeiss Axiovert). For cell counts, allLugol-fixed samples from May to October were diluted 1:19by filtered drinking water and 10 ml volume of that dilutionwere used for settling in counting chambers (Utermöhl,1958) to achieve an equal distribution of algal filaments.
3. Results
3.1. Hydrophysical and hydrochemical conditions
Daily mean air temperature at the weather station Linden-berg first exceeded 20 °C at the end of May 1999, and morefrequently in July 1999 followed by a sharp decrease inmid-August. In 2000, high temperatures (T > 20 °C) wereobserved for short periods in April, May and June followedby an unusually cold July. The summer periods of decreasingair temperatures were accompanied by periods of decreasedglobal radiation (Fig. 1).
In winter 1998/1999, ice cover was observed for 66 d, andfor 48 d at Melangsee in winter 1999/2000. In Langer See,
Table 1Morphometric and hydrological parameters of the investigated lakes
Zmax (m) Zmean (m) Zsampling point (m) Volume (106 m3) Area (km2) Mixing typeMelangsee 3.3 1.6 1.6 0.17 0.11 PolymicticLanger See 3.8 2.1 2.7 3.27 1.55 Polymictic
S12 U. Mischke / Acta Oecologica 24 (2003) S11–S23

the ice cover persisted for 66 and 25 d, respectively. In boththe lakes, the water temperatures exceeded 20 °C only forshort periods during the investigations (Fig. 2). The firstwarming was observed at the end of June in 1999, and earlierin May in the subsequent year. In the summer of 2000, a moredetailed record of the lake temperatures and weather condi-tions, at the study sites is given by the two continuously-measuring data loggers (Fig. 3). Both the lakes werepolymictic, with two periods of stable stagnation each lastingmore than 3 d at the beginning of, and in the middle of,August. In Langer See, summer mixing events were weaklycorrelated with wind speeds observed on the previous day(correlation factor = corr. = 0.511 in July-September). At this
study site, much higher wind speeds (maximum: 4.6 m s–1;and average: 2.0 m s–1) were observed than those measured atthe more sheltered Melangsee (2.7 m s–1; 1.4 m s–1, respec-tively; corr. = 0.240). Therefore, in Melangsee, sharper andmore stable water temperature differences could be estab-lished than in Langer See (Fig. 3). Winds from the north andeast were rare. The number of daily mixing events affectedby night cooling was different in both the lakes: from 25 Julyto 23 October, the in situ temperature differences (15 minvalues of near bottom and near surface probe) were morethan 0.2 °C for 38 complete days in Melangsee and only for28 d in Langer See (data not shown). A comparable numberof days without complete mixing delivered a threshold of0.75 °C for the daily averaged temperature differencesmarked in Fig. 3. A change in the main wind directions didnot significantly influence the mixing in the water columns(Fig. 3). In Melangsee, periods of stratification were termi-nated by decreasing air temperatures rather than by highwind speeds as in Langer See (see above). This interpretationis supported by the positive correlation between the gradientof water temperature and the daily averaged air temperature(corr. = 0.69 in Melangsee).
The underwater light climate, as reflected in the lightattenuation factor (Kd), was strongly influenced by the algalbiomass in both the lakes (corr. = 0.90 and 0.86; Fig. 4) andwas between 2 and 4 m–1 in Melangsee and even above3–5 m–1 in Langer See in summer. The euphotic depth (Zeu)reached the lake bottom (apart from July) until mid-September, when it decreases to a minimum of 1.23 m inLanger See, and to a minimum of 1.60 m in Melangsee. Highwater transparencies were restricted to March and April,since even in winter, secchi depths were reduced by highphytoplankton biomass to about 1 m (Fig. 4). Since waterexchange is low in summer, turbid inflowing waters did notreduce the turbidity of both the lakes then. The large increasein turbidity is due to the enormous algal mass development(Fig. 4).
The high concentrations of dissolved nutrients in winterreduced drastically during the growing season in both thelakes (Fig. 5). In summer DIN never exceeded 50 µg l–1, andnitrate was always less than 5 µg l–1, except for one occasionat the beginning of September in Langer See. The averagesummer concentrations of ammonia declined to 16.1 µg l–1 inLanger See, and to 8.6 µg l–1 in Melangsee. SRP concentra-tions were low throughout summer (Fig. 5). SRP concentra-tions (averaged from June to October 2000) were about10.5 µg l–1 in Melangsee and 7.1 µg l–1 in Langer See. Theconcentrations of TP and TN increased gradually duringsummer to maxima >170 µg l–1 TP and >2000 µg l–1 TN(Fig. 5). From June to October, the concentration ranges, andthe average concentrations of TP and TN were very similar inboth the lakes, and were strongly correlated with the totalalgal biovolume (Table 2). TN/TP ratio ranged between 10.0and 18.1 throughout summer.
Fig. 1. Daily averages of air temperature and monthly averages of the dailysum of global radiation at the weather station Lindenberg, located 12 and30 km from the respective study sites. The horizontal line indicates the 20 °Cair temperature. Summer decreases in air temperature are indicated by blackarrows.
S13U. Mischke / Acta Oecologica 24 (2003) S11–S23
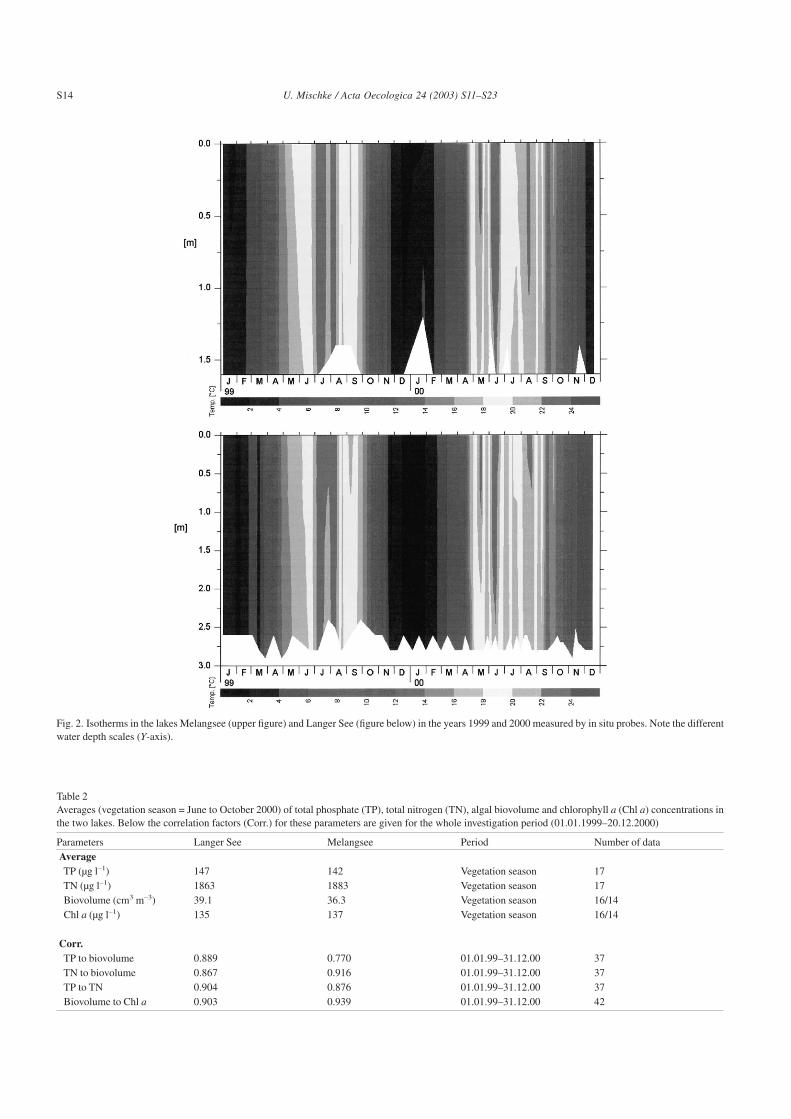
Fig. 2. Isotherms in the lakes Melangsee (upper figure) and Langer See (figure below) in the years 1999 and 2000 measured by in situ probes. Note the differentwater depth scales (Y-axis).
Table 2Averages (vegetation season = June to October 2000) of total phosphate (TP), total nitrogen (TN), algal biovolume and chlorophyll a (Chl a) concentrations inthe two lakes. Below the correlation factors (Corr.) for these parameters are given for the whole investigation period (01.01.1999–20.12.2000)
Parameters Langer See Melangsee Period Number of dataAverageTP (µg l–1) 147 142 Vegetation season 17TN (µg l–1) 1863 1883 Vegetation season 17Biovolume (cm3 m–3) 39.1 36.3 Vegetation season 16/14Chl a (µg l–1) 135 137 Vegetation season 16/14
Corr.TP to biovolume 0.889 0.770 01.01.99–31.12.00 37TN to biovolume 0.867 0.916 01.01.99–31.12.00 37TP to TN 0.904 0.876 01.01.99–31.12.00 37Biovolume to Chl a 0.903 0.939 01.01.99–31.12.00 42
S14 U. Mischke / Acta Oecologica 24 (2003) S11–S23

3.2. Phytoplankton composition
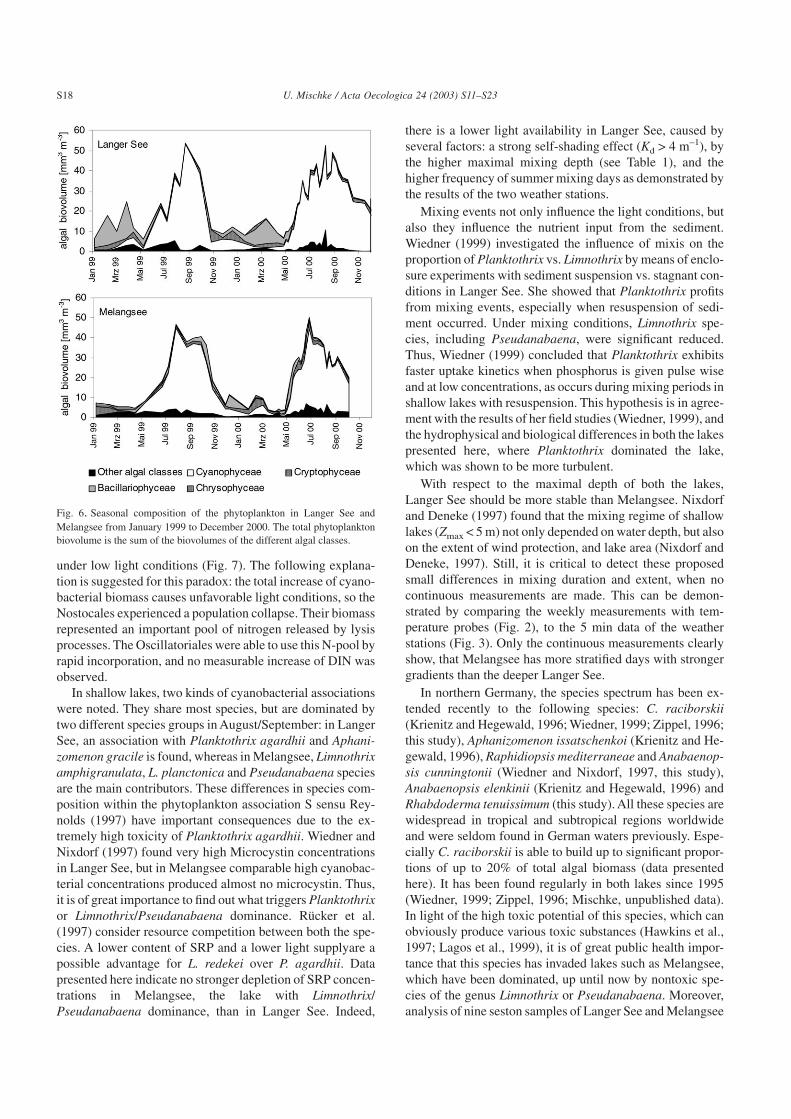
In total, 167 algal taxa were distinguished in the phy-toplankton of the Melangsee and Langer See. The lakesshared 89% of the total taxa. Only 43 species had a biovol-ume of more than 1 cm3 m–3 at any one sampling date(Table 3). In summer and autumn, cyanobacteria were themost common species (Fig. 6), while in winter and spring,diatoms, cryptophytes and chrysophytes species were impor-tant (Table 3).
The plankton of the two lakes were dominated by differentcyanobacterial species. In Langer See, Planktothrix agardhii
(GOM.) ANAGN. et KOM. constituted the greatest specificpopulation biovolume (Table 3). Species with very smallfilaments (1.2–2.1 µm width) of Limnothrix, Pseudanabaenaand Skujanema sp. were most important in Melangsee. Theircumulated biovolume was comparable to that of P. agardhiiin Langer See. L. redekei was important only in winter/springand L. amphigranulata was much more significant.
The composition of the cyanobacterial phytoplankton wassimilar in both the years of investigation (Fig. 7). In LangerSee, species of the Nostocales such as Aphanizomenon grac-ile (LEMMERM.) LEMMERM., A. issatschenkoi (USAC.)
Fig. 3. Differences of water temperatures in the near surface and near lake bottom layer (daily averages; shaded areas) as inverse indicators of completel summermixing events in lake Langer See (upper graph; differences between 0.5 and 2.7 m) and in Melangsee (graph below; differences between 0.5 and 1.7 m) inrelation to daily averages of wind speed (lines) and main wind direction (rhomb symbol) in the year 2000. The dotted horizontal lines indicate temperaturedifferences of 0.75 °C.
S15U. Mischke / Acta Oecologica 24 (2003) S11–S23

PROSK.-LAVR. and Anabaena solitaria KLEB. increasedin June and July, followed bya sharply and continuouslyincreasing biovolume of the sole Oscillatoriales speciesP. agardhii. In 2000, the total cyanobacterial biovolumestarted at a much higher level than in the previous year andspecies of the Chroococales, such as Microcystis wesen-bergii, M. aeruginosa and Synechocystis were less signifi-cant. In Melangsee, the very small Limnothrix species L.amphigranulata (VAN GOOR) MEFFERT was the mostcommon species accompanied by other Oscillatoriales, suchas Pseudanabaena limnetica LEMM., Limnothrix redekei(VAN GOOR) MEFFERT and L. planktonica (WOLOSZ.)MEFFERT. Typical morphotypes of L. redekei were mainlyobserved in autumn/winter and near the lake bottom. Nosto-cales were represented mainly by A. gracile KLEBAHN andby the new invading species C. raciborskii (WOLSZYN-SKA) SEENAYA & SUBBA RAJU. Additional cyanobacte-rial species, such as Rhaphidiopsis mediterranea SKUJA,Rhabdoderma tenuissimum KOMÁREK et KLING, Plank-tolyngbya limnetica ANAGNOSTIDIS & KOMÁREK 1988,Anabaenopsis cunningtonii TAYLOR 1932 and Anabaenacompacta (NYGAARD) HICKEL, which are seldom foundin Germany (Krienitz and Hegewald, 1996), were deter-mined at the study sites. They were found mainly in Langer
See. The ratio of diazotrophic to other filamentous cyanobac-teria changed from about 1:1 in June to 1:10 and maximal1:20 in September in all four observed summer successions(Fig. 7). The Nostocales showed positive net biomasschanges in periods of water temperatures above 15 °C(Fig. 8). The populations of Planktothrix and Limnothrixgrew in spring/autumn at low water temperatures also(Fig. 8). The calculated net growth rates of all the specieswere comparable to each other (Table 4). In Langer See andin Melangsee, P. agardhii grew more slowly than in pub-lished data, but more continuously over a longer period.From 14 March to 6 September 2000 P. agardhii showedpositive net growth rates for 113 d in Langer See, and only for95 d, and interrupted with decreasing periods, in Melangsee.
The net growth rates of most cyanobacterial species inves-tigated were distributed evenly along the whole spectrum oflight intensities found in these two shallow lakes, expressedas the ratio of maximal depth (Zm) to the secchi depth (Zs) inFig. 8. Anabaena solitaria did not grow under very low lightconditions (>7 Zm/Zs). The onset of growth of C. raciborskiiwas comparatively late in summer, when light resource wasalready reduced by self-shading (>3 Zm/Zs). Restricted to ashort period, it was still able to grow with net growth ratescomparable to maximum growth rates published byIstvánovics et al. (2000) (Table 4).
3.3. Vertical distribution of cyanobacteria
Even in polymictic lakes, periods with stratification can beobserved (see Fig. 3), which enable the cyanobacteria tostratify. The vertical distribution of the phytoplankton wasstudied (Fig. 9) during the strongest stagnation period at theend of July in 2000, when calm conditions with increasing airtemperature occurred on the preceding day as well. Maxi-mum temperature differences (top/bottom) were 5 °C inMelangsee, and 3.8 °C in Langer See. Oxygen saturation andpH values were highest near the lake surface (Fig. 9C). Thephytoplankton biovolume and chlorophyll a concentrationswere distributed unevenly in both the lakes (Fig. 9A). At07:00 a.m. in Langer See most of the phytoplankton wasobserved in the upper part of the water column with a maxi-mum at around 1 m water depth. A different distribution wasfound in Melangsee with a maximum concentration at 1.4 mwater depth and low near-surface concentrations. The com-position of the phytoplankton did not change significantlyalong the vertical axis in both the lakes (Fig. 9B), except forthe increased proportion of P. agardhii and Chroococalesnear the lake bottom in Melangsee. SRP concentrations wereabove 5 µg l–1 at all water depths in both the lakes. Ammoniawas depleted to below the detection level (<19.4 µg l–1) in thesubsurface layers. Only near the lake sediment, concentra-tions above 200 µg l–1 were detected in Langer See. Nitrateconcentrations ranged between 1.4 and 8.8 µg l–1.
Fig. 4. Light climate given as the light attenuation factor (Kd), the euphoticdepth (Zeu (m)) and secchi depth (m) in comparison to the phytoplanktonbiovolume in the lakes Langer See (upper graph) and Melangsee (below).
S16 U. Mischke / Acta Oecologica 24 (2003) S11–S23

4. Discussion
4.1. The cyanobacterial associations in Melangseeand Langer See
In both the lakes investigated, the summer association ofthe phytoplankton is composed of cyanobacterial species.This is the result of the very high trophic state and thedepletion of dissolved nutrients. In the sense of Mur et al.(1993), turbulence tolerant species are favored by mixingconditions occurring intermittently with stratification peri-ods in shallow lakes during summer. Those species belong-ing to the Oscillatoriales are known to tolerate very low lightconditions (Foy and Gibson, 1982). Planktothrix and Limno-thrix species are grouped together to the plankton associationS by Reynolds (1997), to characterize the cyanobacterialassemblage of shallow, wind exposed lakes with reducedlight availability.
The second governing environmental factor is the deple-tion of dissolved inorganic nitrogen in both the lakes, whichshould favor N-fixing cyanobacteria (Mur et al., 1993). In-deed, on the onset of summer succession, N-fixing Nosto-cales are an important component (about 50%) of the totalcyanobacterial biomass. Colony forming and stratifyingtypes such as Aphanizomenon flos-aquae and Microcystisspecies were rare due to the mixing regime and the low lightclimate.
The replacement of N-fixing cyanobacteria by turbulencetolerant species in all four summer plankton successions isthe most striking fact during summer succession (Fig. 7). Areduced light availability by self-shading is likely to be re-sponsible for this succession. Most Nostocales have a highlight and temperature demand (Dokulil and Teubner, 2000),especially under N depletion (Schlangstedt et al., 1987). Incontrast, the Oscillatoriales satisfied their nitrogen demandanyway, and they were able to increase their population size
Fig. 5. Concentrations of dissolved inorganic nitrogen (nitrate, ammonia both upper graphs), of SRP (center graphs) and of total phosphate (TP) and totalnitrogen (TN, lowest graphs) in the lakes Langer See (left side) and Melangsee (right side) from January to December 2000.
S17U. Mischke / Acta Oecologica 24 (2003) S11–S23
under low light conditions (Fig. 7). The following explana-tion is suggested for this paradox: the total increase of cyano-bacterial biomass causes unfavorable light conditions, so theNostocales experienced a population collapse. Their biomassrepresented an important pool of nitrogen released by lysisprocesses. The Oscillatoriales were able to use this N-pool byrapid incorporation, and no measurable increase of DIN wasobserved.
In shallow lakes, two kinds of cyanobacterial associationswere noted. They share most species, but are dominated bytwo different species groups in August/September: in LangerSee, an association with Planktothrix agardhii and Aphani-zomenon gracile is found, whereas in Melangsee, Limnothrixamphigranulata, L. planctonica and Pseudanabaena speciesare the main contributors. These differences in species com-position within the phytoplankton association S sensu Rey-nolds (1997) have important consequences due to the ex-tremely high toxicity of Planktothrix agardhii. Wiedner andNixdorf (1997) found very high Microcystin concentrationsin Langer See, but in Melangsee comparable high cyanobac-terial concentrations produced almost no microcystin. Thus,it is of great importance to find out what triggers Planktothrixor Limnothrix/Pseudanabaena dominance. Rücker et al.(1997) consider resource competition between both the spe-cies. A lower content of SRP and a lower light supplyare apossible advantage for L. redekei over P. agardhii. Datapresented here indicate no stronger depletion of SRP concen-trations in Melangsee, the lake with Limnothrix/Pseudanabaena dominance, than in Langer See. Indeed,
there is a lower light availability in Langer See, caused byseveral factors: a strong self-shading effect (Kd > 4 m–1), bythe higher maximal mixing depth (see Table 1), and thehigher frequency of summer mixing days as demonstrated bythe results of the two weather stations.
Mixing events not only influence the light conditions, butalso they influence the nutrient input from the sediment.Wiedner (1999) investigated the influence of mixis on theproportion of Planktothrix vs. Limnothrix by means of enclo-sure experiments with sediment suspension vs. stagnant con-ditions in Langer See. She showed that Planktothrix profitsfrom mixing events, especially when resuspension of sedi-ment occurred. Under mixing conditions, Limnothrix spe-cies, including Pseudanabaena, were significant reduced.Thus, Wiedner (1999) concluded that Planktothrix exhibitsfaster uptake kinetics when phosphorus is given pulse wiseand at low concentrations, as occurs during mixing periods inshallow lakes with resuspension. This hypothesis is in agree-ment with the results of her field studies (Wiedner, 1999), andthe hydrophysical and biological differences in both the lakespresented here, where Planktothrix dominated the lake,which was shown to be more turbulent.
With respect to the maximal depth of both the lakes,Langer See should be more stable than Melangsee. Nixdorfand Deneke (1997) found that the mixing regime of shallowlakes (Zmax < 5 m) not only depended on water depth, but alsoon the extent of wind protection, and lake area (Nixdorf andDeneke, 1997). Still, it is critical to detect these proposedsmall differences in mixing duration and extent, when nocontinuous measurements are made. This can be demon-strated by comparing the weekly measurements with tem-perature probes (Fig. 2), to the 5 min data of the weatherstations (Fig. 3). Only the continuous measurements clearlyshow, that Melangsee has more stratified days with strongergradients than the deeper Langer See.
In northern Germany, the species spectrum has been ex-tended recently to the following species: C. raciborskii(Krienitz and Hegewald, 1996; Wiedner, 1999; Zippel, 1996;this study), Aphanizomenon issatschenkoi (Krienitz and He-gewald, 1996), Raphidiopsis mediterraneae and Anabaenop-sis cunningtonii (Wiedner and Nixdorf, 1997, this study),Anabaenopsis elenkinii (Krienitz and Hegewald, 1996) andRhabdoderma tenuissimum (this study). All these species arewidespread in tropical and subtropical regions worldwideand were seldom found in German waters previously. Espe-cially C. raciborskii is able to build up to significant propor-tions of up to 20% of total algal biomass (data presentedhere). It has been found regularly in both lakes since 1995(Wiedner, 1999; Zippel, 1996; Mischke, unpublished data).In light of the high toxic potential of this species, which canobviously produce various toxic substances (Hawkins et al.,1997; Lagos et al., 1999), it is of great public health impor-tance that this species has invaded lakes such as Melangsee,which have been dominated, up until now by nontoxic spe-cies of the genus Limnothrix or Pseudanabaena. Moreover,analysis of nine seston samples of Langer See and Melangsee
Fig. 6. Seasonal composition of the phytoplankton in Langer See andMelangsee from January 1999 to December 2000. The total phytoplanktonbiovolume is the sum of the biovolumes of the different algal classes.
S18 U. Mischke / Acta Oecologica 24 (2003) S11–S23

in summer 2000 provided the first proof of the toxin Cylin-drospermopsin in German and even in European waters(Eaglesham, unpublished data).
In German waters, C. raciborskii has already started sea-sonal growth at water temperatures above 17 °C (Fig. 8),clearly well below the critical value of 22 °C, which wasthought to be the lower limit. It showed a significant reducedtemperature requirement for growth in comparison to thesubtropical and tropical strains (Padisák, 1997). Up untilnow, only Dokulil and Mayer (1996) found a strain of C.raciborskii in central Europe, with such a low temperaturerequirement. Since the temperature can influence the toxincontent of C. raciborskii (Saker and Griffiths, 2000), a hightoxin production can be expected.
Surprisingly, the growth of the C. raciborskii populationwas not strongly correlated with water temperatures in theinvestigated lakes (corr. 0.335 in Langer See; corr. 0.324 inMelangsee). Dokulil and Teubner (2000) found positive net
changes of C. raciborskii at much lower light climate(Zm/Zs > 10) and water temperatures (<15 °C) than thosefound in the present study. Thus, Padisák and Reynolds(1998) created the new algal association, Sn, demonstratingthe affiliation of C. raciborskii with the non aggregating, lowlight tolerant species. Its ability to fix nitrogen is of second-ary importance. Strong nutrient depletion (Dokulil andMayer, 1996) and/or low light caused by deep mixing couldprevent the growth of C. raciborskii, as was observed byHawkins and Griffiths (1983). Since the low water depthsprevent deep mixing in both the study sites, very low nitrogenavailability is proposed to be responsible for the populationdecline of C. raciborskii in summer.
5. Conclusions
The comparison of cyanobacterial communities in twoshallow lakes revealed that small differences in mixing and
Table 3Algal taxa, with more than 1 cm3 m–3 specific biovolume at one sampling date with their maximal biovolume (Max in cm3 m–3) during the investigation periodfrom 01.01.1999 to 01.11.2000 in both the lakes
Langer See b. Dolgenbrodt MelangseeTaxon name Max Taxon name MaxCyanobacteria CyanobacteriaPlanktothrix agardhii 43.6 Limnothrix amphigranulata 25.5Aphanizomenon gracile 9.8 Limnothrix planctonica 16.2Pseudanabaena spp. 8.8 Aphanizomenon gracile 10.0Cylindrospermopsis raciborskii 5.3 Cylindrospermopsis raciborskii 9.2Limnothrix redekei 5.2 Pseudanabaena spp. 8.8Limnothrix planctonica 4.9 Limnothrix sp. 5.8Skujanema spp. 1.6 Anabaena solitaria 4.5Aphanizomenon issatschenkoi 1.5 Chroococcus sp. 4.3Limnothrix sp. 1.8 Limnothrix redekei 4.2Synechocystis 2.3 Planktothrix agardhii 3.9Anabaena solitaria 2.2 Aphanizomenon sp. 1.2Anabaena flos-aquae 1.4 Planktolyngbya limnetica 1.0Aphanizomenon sp. 1.4 Synechocystis sp. 2.6
Other algal classes Other algal classesCryptomonas erosa 5.5 Pinnularia divergens 5.6Cryptomonas marssonii 1.8 Nitzschia cf. acicularis 5.0Rhodomonas lacustris 1.7 Fragilaria ulna var acus 2.8Rhodomonas minuta 1.5 Fragilaria ulna var ulna 2.2Ceratium hirundinella 3.0 Fragilaria crotonensis 1.2Mougeotia sp. 2.6 Ephithemia sp. 1.1Stephanodiscus parvus/minutulus 11.5 Carteria sp. 4.4Centric diatoms 12–18 µm 5.9 Cryptomonas erosa 3.0Centric diatoms >18 µm 9.2 Cryptomonas marssonii 2.5Asterionella formosa 1.9 Peridinium aciculiferum 2.1
Peridinium cf. cinctum 1.5Chrysochromulina spec. 2.2Uroglena sp. 1.5Synura sp. 1.4Dinobryon sertularia 1.2Mougeotia sp. 1.7Chlamydomonas sp. 1.2
S19U. Mischke / Acta Oecologica 24 (2003) S11–S23

light regime can lead to switching between toxin producingcyanobacteria or others. It is insufficient to derive the mixingbehavior of a lake from its maximal or mean water depth.Lakes with low water depths, but with substantial windprotection such as Melangsee, can be more stably stratified
than deeper lakes. C. raciborskii recently invaded both thelakes, but it regularly declined in summer similar to otherNostocales, when Planktothrix or Limnothrix/Pseudana-baena became dominant. The latter species are known to betolerant to turbulent conditions. P. agardhii is much more
Fig. 7. Contribution of cyanobacterial species (areas) to total biovolume of cyanobacteria (line with open circles) in the lakes Langer See (left) and Melangsee(right) during summer succession from June to October in 1999 and 2000.
Table 4Mean and maximal positive net biomass changes (>0.02) calculated from field data (net growth rates, d–1) for the investigated species. Published maximumgrowth rates in the laboratory or from field data (Dokulil and Mayer, 1996; Istvánovics et al., 2000; Schlangstedt et al., 1987)
Species/Net growth rate Melangsee Langer See Maximum publishedMean Maximum Mean Maximum
Anabaena solitaria 0.107 0.298 0.126 0.303 1.18Aphanizomenon gracile 0.079 0.490 0.084 0.259Aphanizomenon issatschenkoi – – 0.119 0.188Cylindrospermopsis raciborskii 0.087 0.149 0.105 0.226 1.00Pseudanabaena limnetica 0.088 0.514 0.113 0.554Limnothrix spp. 0.063 0.176 0.091 0.307Planktothrix agardhii 0.090 0.174 0.058 0.144 0.24
S20 U. Mischke / Acta Oecologica 24 (2003) S11–S23

successful under a strong mixing regime. The increasedknowledge about the taxonomic differentiation of cyanobac-teria allows a deeper analysis of the effect of resource condi-tions on their population dynamics. This is underlined by theobviously different resource demands of L. redekei found inwinter, and of L. amphigranulata found during the summerperiod. Laboratory studies on L. redekei revealed its ability togrow under extremely low light and low nutrient conditions,but in fact, L. redekei is not relevant in the complex mixtureof species with similar morphotypes in summer. Further fieldand laboratory studies should take cognizance of theenormous diversity of cyanobacteria occurring in shallowlakes.
Acknowledgements
This work was carried out at the research station BadSaarow of the Brandenburg Technical University of Cottbus.The author thanks all the members of the laboratory staff,especially Gudrun Lippert, Tina Hankem, Ute Abel and Kor-nelia Tahedel for the chemical analysis including chloro-phyll. Ingo Henschke and Wolfgang Terlinden are greatlythanked for the reliable field work and the technical care ofthe weather stations. Claudia Wiedner and Jutta Fastnergreatly assisted this work by their discussions and taxonomichints. The comments of two unknown reviewers and of Phil-lip Ford, who gave linguistic help additionally, greatly im-
Fig. 8. Two-dimensional representations of the positive net biomass changes to water temperature and Zm/Zs for the species indicated. Maximal biovolume ofspecies are indicated by larger circles for Melangsee (filled black) and for Langer See (filled white). Cyl. rac. = Cylindrospermopsis raciborskii; Ana. sol. =Anabaena solitaria; Pla. aga. = Planktothrix agardhii; Aph. gra. = Aphanizomenon gracile; Lim. spp. = Limnothrix spp.; Pse. lim. = Pseudanabaena limnetica.
S21U. Mischke / Acta Oecologica 24 (2003) S11–S23

Fig. 9. Depth profile of (A) total phytoplankton biovolume (white bars), of Cylindrospermopsis raciborskii (black bars) and of chlorophyll a concentrations, (B)the contribution of selected species to total algal biovolume and (C) the depth profiles of water temperature, oxygen saturation and pH value in the lakes LangerSee (upper graphs; 07:30 a.m.) and Melangsee (graphs below; 11.00 a.m.) on 25 July 2000.
S22 U. Mischke / Acta Oecologica 24 (2003) S11–S23

proved the paper. This work was financially supported by theGerman Research Foundation (DFG) contract AZNI347/5-1.
References
Anagnostidis, K., Komárek, J., 1985. Modern approach to the classificationsystem of cyanophytes - 1 - Introduction. Arch. Hydrobiol. 1/2 (Supple-ment 71), 291–302.
Anagnostidis, K., Komárek, J., 1988. Modern approach to the classificationsystem of cyanophytes - 3 - Oscillatoriales. Arch. Hydrobiol. 1/4(Supplement 80), 327–472.
Dokulil, M.T., Mayer, J., 1996. Population dynamics and photosyntheticrates of a Cylindrospermopsis - Limnothrix association in a highlyeutrophic urban lake, Alte Donau, Vienna, Austria. Algol Stud 83,179–195.
Dokulil, M.T., Teubner, K., 2000. Cyanobacterial dominance in lakes.Hydrobiologia (The Hague) 438, 1–12.
Foy, R.H., Gibson, C.E., 1982. Photosynthetic characteristics of planktonicblue-green algae: the response of 20 strains grown under high and lowlight. Br. Phycol. J 17, 169–182.
Hawkins, P.R., Chandrasena, N.R., Jones, G.J., Humpage, A.R., Fal-coner, I.R., 1997. Isolation and toxicity of Cylindrospermopsis racibor-skii from an ornamental lake. Toxicon 35, 341–346.
Hawkins, P.R., Griffiths, D.J., 1983. Artificial destratification of a smalltropical reservoir: effects upon the phytoplankton. Hydrobiologia (TheHague) 254, 169–181.
Hindák, F., 1988. Planktonic species of two related genera Cylindrosper-mopsis and Anabaenopsis from Western Slovakia. Algological Studies50-53, 283–302.
Horecká, M., Komárek, J., 1979. Taxonomic position of three planktonicblue-green algae from the genera Aphanizomenon and Cylindrosper-mopsis. Preslia 51, 289–312.
Istvánovics, V., Shafik, H.M., Présing, M., Juhos, S., 2000. Growth andphosphate uptake kinetics of the cyanobacterium, Cylindrospermopsisraciborskii (Cyanophyceae) in throughflow cultures. Freshwat. Biol 43,257–275.
Kohl, J.G., Dudel, G., Schlangstedt, M., Kühl, H., 1985. Zur morphologis-chen Abgrenzung von Aphanizomenon flos-aquae RALF ex. BORN. etFLAH. and A. gracile (LEMM.) LEMM. Arch. Protistenk. 130,119–131.
Komárek, J., Anagnostidis, K., 1989. Modern approach to the classificationsystem of cyanophytes - 4 - Nostocales. Arch. Hydrobiol (Supplement82), 247–345.
Krienitz, L., Hegewald, E., 1996. Über das Vorkommen von wärmelie-benden Blaualgenarten in einem norddeutschen See. Lauterbornia 26,55–63.
Lagos, N., Onodera, H., Zagatto, P.A., Andrinolo, D., Azevedo, S.M.,Oshima, Y., 1999. The first evidence of paralytic shellfish toxins in thefreshwater cyanobacteria Cylindrospermopsis raciborskii, isolated fromBrazil. Toxicon 37, 1359–1373.
Meffert, M.E., 1988. Limnothrix MEFFERT nov. gen. Arch. Hydrobiol.(Supplement 80), 269–276.
Meffert, M.E., Oberhäuser, R., Overbeck, J., 1981. Morphology and Tax-onomy of Oscillatoria redekei (Cyanophyta). Brit. Phycol. J 16,107–114.
Mur, L.R., Schreurs, H., Visser, P., 1993. How to control undesirable cyano-bacterial dominance. In: Giussani, G., Callieri, C. (Eds.), Strategies forLake Ecosystems Beyond 2000, Proceedings of the Fifth InternationalConference on Conservation and Management of Lakes, Stresa.pp. 565–569.
Nixdorf, B., Deneke, R., 1997. Why ’very shallow’ lakes are more success-ful opposing reduced nutrient loads. Hydrobiologia (The Hague)342/343, 269–284.
Padisák, J., 1997. Cylindrospermopsis raciborskii (WOLOZYNSKA)SEENAYYA et SUBBA RAJU, an expanding, highly adaptivecyanobacterium: worldwide distribution and review of its ecology. Arch.Hydrobiol. (Supplement 107), 563–693.
Padisák, J., Reynolds, C.S., 1998. Selection of phytoplankton associations inLake, Balaton, Hungary, in response to eutrophication and restorationmeasures, with special reference to the cyanoprokaryotes. Hydrobiolo-gia (The Hague) 384, 41–53.
Reynolds, C.S., 1997. Vegetation processes in the pelagic: a model forecosystem theory. Ecology Institute, D-21385 Oldendorf, Germany,pp. 7–18.
Rott, E., 1981. Some results from phytoplankton counting intercalibrations.Schweiz. Z. Hydrol 8, 34–62.
Rücker, J., Wiedner, C., Zippel, P., 1997. Factors controlling the dominanceof Planktothrix agardhii and Limnothrix redekei in eutrophic shallowlakes. Hydrobiologia (The Hague) 342/343, 107–115.
Saker, M.L., Griffiths, D.J., 2000. The effect of temperature on growth andcylindrospermopsin content of seven isolates of Cylindrospermopsisraciborskii (Nostocales, Cyanophyceae) from water bodies in northernAustralia. Phycologia 39, 349–354.
Schlangstedt, M., Bisen, P.S., Dudel, G., Kohl, J.G., 1987. Interaction ofcombined nitrogen availability and light in the regulation of growth,heterocyst differentiation and dinitrogen fixation of the planktic blue-green alga Anabaena solitaria KLEB. Arch. Protistenk 134, 389–396.
Utermöhl, H., 1958. Zur Vervollkommnung der quantitativenPhytoplankton-Methodik. Mitt. Internat. Verein. Limnol 9, 1–38.
Wiedner, C., 1999. Toxische und nicht-toxische Cyanobakterien inGewässern der Scharmützelseeregion: Ihr Vorkommen in Gewässernunterschiedlicher Trophie und Morphometrie und Steuermechanismenihrer Dynamik in polymiktischen Flachseen. Dissertation, BrandenburgTechnical University of Cottbus 131 p.
Wiedner, C., Nixdorf, B., 1997. Verbreitung und Steuerung der Entwicklungvon toxischen und nicht-toxischen Cyanobakterien in ostbrandenburgis-chen Gewässern unterschiedlicher Trophie und Hydrologie im Zuge derreduzierten Belastung. Informationshefte des Instituts für Wasser-,Boden- Lufthygiene in Berlin 4, 17–26.
Zippel, P., 1996. Phytoplanktonsukzession und -dynamik in den Gewässerndes Scharmützelseegebietes. In: Nixdorf, B., Kleeberg, A. (Eds.),Gewässerreport (Teil II). Aktuelle Reihe 2/96 Cottbus University Press,pp. 44–60.
S23U. Mischke / Acta Oecologica 24 (2003) S11–S23