cutting edge: cxcr4-lo: molecular cloning and … edge: cxcr4-lo: molecular cloning and functional...
TRANSCRIPT

of June 26, 2018.This information is current as
Human CXCR4 Splice VariantNovelCloning and Functional Expression of a
Cutting Edge: CXCR4-Lo: Molecular
Shalley K. Gupta and Kodandaram Pillarisetti
http://www.jimmunol.org/content/163/5/23681999; 163:2368-2372; ;J Immunol
Referenceshttp://www.jimmunol.org/content/163/5/2368.full#ref-list-1
, 16 of which you can access for free at: cites 27 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Cutting Edge: CXCR4-Lo: MolecularCloning and Functional Expression of aNovel Human CXCR4 Splice Variant
Shalley K. Gupta1 and Kodandaram Pillarisetti
Human CXCR4 is a specific receptor for the CXC chemokinestromal cell-derived factor-1 (SDF-1) and a coreceptor for Tcell line tropic strains of HIV-1. Genetic knockouts of CXCR4and SDF-1 have delineated their critical role during embryoniccardiogenesis, leukopoiesis, and vasculogenesis. Herein, weused bioinformatics and differential strategies like isoform-specific RT-PCR and Northern blots to identify and clone anovel unspliced isoform of human CXCR4, termed CXCR4-Lo. CXCR4-Lo corresponds to a larger;4.0-kb mRNA tran-script and differs from the known human CXCR4 by the first9 aa in the functionally important NH 2-terminal extracellulardomain of the receptor. CXCR4-Lo-transfected rat basophilleukemia-2H3 cells responded to SDF-1 with a transient rise ofintracellular Ca 21 concentration and by undergoing chemo-taxis. Expression of CXCR4-Lo is noteworthy, as it may havedifferential affinity as a coreceptor for HIV strains in compar-ison with CXCR4. Furthermore, CXCR4-Lo may also providea functional backup to CXCR4 during embryogenesis. TheJournal of Immunology,1999, 163: 2368–2372.
Stromal cell-derived factor-1 (SDF-1)2 is a CXC chemokine(1) that uses CXCR4 as its specific receptor (2). SDF-1 (1,3), along with CXCR4 (4, 5), has widespread tissue ex-
pression and is a potent chemoattractant of PBLs (6) and endothe-lial cells (7). Recent studies with genetic knockouts of both SDF-1(8) and CXCR4 (9, 10) have underscored their vital role duringembryogenesis. Another noteworthy feature of CXCR4 (11) andCC chemokine receptors like CCR5 (12) is their role as corecep-tors for infection by HIV. The NH2-terminal extracellular (EC)domain is a major determinant of receptor subtype specificity andbinding to chemokine ligands (13–16). Among HIV coreceptorslike CXCR4 and CCR5, the EC domain mediates a crucial role
during their tripartite interaction with HIV-encoded gp120 and itscellular receptor CD4 (11, 17). In this context, it is noteworthy thatthe first 20 NH2-terminal residues of CCR5 confer coreceptorfunction when placed into the CCR2b background (17). Absagainst the NH2 terminus of human CXCR4 inhibit cell fusion andinfection with HIV-1 (11).
The intron splice junction of murine CXCR4 gene has an in-frame alternative GT splice donor site within exon 1, causing ex-pression of an isoform that is shorter by 2 aa in the NH2-terminaldomain (18, 19). Both murine CXCR4 isoforms are coexpressed inall tissues examined and use SDF-1 as their functional ligand. Thehuman CXCR4 gene also contains 2 exons of 103- and 1563-bpsize (20, 21), interrupted by a 2132-bp intron precisely betweencodons 5 (AGT/Ser) and 6 (ATA/Ile) of the NH2-terminal domain(20). Although an alternative splice donor site (GT) similar to themurine CXCR4 gene exists 9 bp downstream from the last codonof the first exon in human CXCR4 (20), there is no molecularevidence it causes expression of multiple transcripts (21, 22).
In contrast, our initial observation of two distinct mRNA tran-scripts in HUVECs (7) strongly revealed the possible existenceand expression of a unique human CXCR4 splice variant. In thisreport, we demonstrate that the human CXCR4 gene indeed has alarger unspliced form of the receptor, CXCR4-Lo, that differs withCXCR4 in the NH2-terminal region by 9 aa. We also show thatCXCR4-Lo is functional and differentially expressed in humantissues.
Materials and MethodsIsolation of human CXCR4-Lo cDNA, RT-PCR, and Northernblot analysis
The CXCR4-Lo cDNA clone was identified from the SmithKline BeechamcDNA database by searching for expressed sequence tags (ESTs) withidentity to human CXCR4 and analyzing their open reading frames (ORFs)for NH2-terminal sequence variants using the FASTA and BLAST algo-rithms of the GCG software package (Genetics Computer Group, Madison,WI). Total RNA was isolated from HL-60 cells by the Tri-reagent proce-dure (Molecular Research Center, Cincinnati, OH), and CXCR4-Lo spe-cific cDNA was amplified with two sets of specific primers, Lo-346 andLo-1071 (Lo-346, forward: 59AGAGAGAGAACTAGTCTCGC, reverse:59AGAGGCAAAGGAATGGACAT; and Lo-1071, forward: 59ATGTCCATTCCTTTGCCTCT, reverse: 59AGCTGGAGTGAAAACTTGAAG)using the RNA-PCR kit from Perkin-Elmer (Foster City, CA), according tothe manufacturer’s protocol. The 346-bp PCR product amplified with Lo-346 primers was used as a CXCR4-Lo-specific cDNA probe, while the515-bp cDNA fragment of CXCR4 (7) was used as a common probe forboth isoforms. Human tissue Northern blots were purchased from Clontech(Palo Alto, CA). The GAPDH probe (Clontech) was used to normalize theamount of total RNA loaded in all lanes. [a-32P]dCTP-labeled DNA probeswere used for high-stringency hybridizations as described earlier (7).Northern blots were analyzed using phosphorimager densitometric scans(Molecular Devices, Menlo Park, CA).
Department of Cardiovascular Pharmacology, SmithKline Beecham Pharmaceuticals,King of Prussia, PA 19406
Received for publication March 17, 1999. Accepted for publication July 6, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Shalley K. Gupta, MailCode UW2511, Department of Cardiovascular Pharmacology, SmithKline BeechamPharmaceuticals, King of Prussia, PA 19406. E-mail address: [email protected] Abbreviations used in this paper: SDF-1, stromal cell-derived factor-1; EC, extra-cellular; EST, expressed sequence tag; FLIPR, fluorometric imaging plate reader;ORF, open reading frame; RBL, rat basophil leukemia cells; [Ca21]i, intracellularCa21 concentration.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
●
●
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Transfection of rat basophil leukemia (RBL)-2H3 cells
Full-length CXCR4 and CXCR4-Lo cDNAs were subcloned into pCRR3.1(Invitrogen, San Diego, CA) eukaryotic expression vectors, and their ori-entation was confirmed by sequencing. The resultant expression constructswere transfected into RBL cells (American Type Culture Collection, Ma-nassas, VA) by electroporation, and transfected cells were selected by us-ing RPMI 1640 media containing 800mg/ml G-418 (Life Technologies,Rockville, MD). CXCR4 and CXCR4-Lo specific mRNA expression wasconfirmed by Northern blot analysis. FACS analysis to compare surfaceexpression of CXCR4 and CXCR4-Lo in transfected RBL cell lines wasdone with the CXCR4-specific 12G5 mAb as described before for endo-thelial cells (7).
Functional characterization of CXCR4-Lo in stably transfectedRBL cells
Mobilization of [Ca21]i in transfected RBL cells was measured byfluorometric imaging plate reader (FLIPR; Molecular Devices)
analysis (23). Briefly, 63 104 cells were added per well of a96-well plate and grown for 24 h. Cells were loaded with Fluo-3AM dissolved in dye-loading buffer (Eagle’s minimum essentialmedium with 0.1% BSA and 15 mM sulpinpyrozone) for 60 minat 37°C. Following this, the dye-loading buffer was removed, and100 ml hydrolysis buffer (15 mM sulfinpyrozone in Eagle’s min-imum essential medium) was added to each well and incubated for10 min at 37°C. Relative changes in fluorescence counts as a re-sponse to [Ca21]i flux after addition of SDF-1a (R&D Systems,Minneapolis, MN) are expressed as FLIPR units.
SDF-1-induced migration of transfected RBL cells was done asdescribed for HUVECs (7). Briefly, 53 105 cells in serum-freeRPMI 1640 medium (with 0.25% BSA) were added in the topchamber of a 6.5-mm diameter, 5-mm pore poly carbonate trans-well culture insert (Costar, Cambridge, MA). Incubation was con-ducted at 37°C in a 5% CO2 incubator for 18 h. Migrated cells in
FIGURE 1. Sequence of a novelCXCR4-Lo isoform and the proposedsplicing mechanism of the human CXCR4gene.A, Partial nucleotide sequence of the59-untranslated region and deduced aminoacid sequence of the neo-NH2-terminal endof the full-length CXCR4-Lo cDNA clone.The unique 9-aa extension of the EC do-main is shown in circles. The AG spliceacceptor site used for splicing out the in-tron for CXCR4 (20) is boxed. The cDNAsequence distal to ATA codon (Ile) at po-sition 370 is identical with CXCR4 cDNA.Nucleotide sequences of the Lo-346 prim-ers used to amplify the 346-bp CXCR4-Lo-specific fragment are highlighted withdirectional arrows.B, Schematic represen-tation of the origin of human CXCR4-LocDNA from CXCR4 gene. The splice do-nor (GT) and splice acceptor (AG) sitesthat flank the 2.13-kb intron of CXCR4 areencircled. In the proposed origin ofCXCR4-Lo, the normal splicing does notoccur and the intron is retained and ex-pressed as part of the unspliced;4.0 kbmRNA. The in-frame alternative ATG startcodon within the “coding intron” used bythe CXCR4-Lo variant is boxed and ini-tiates the unique 9-aa extension to theNH2-terminal EC domain.C, RT-PCR ofCXCR4-Lo ORF with isoform-specificprimers (Lo-1071) from HL-60 cells andits hybridization with the specific 346-bpDNA probe. The reverse transcriptase re-action was initiated with either the oli-go(dT) or the 39-Lo-1071 primers.D, Mul-tiple-tissue Northern blot hybridizationwith the CXCR4-Lo-specific 346-bpcDNA probe.
2369The Journal of Immunology
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
the lower chamber were counted with a ZM Coulter counter(Coulter Diagnostics, Hialeah, FL). Percent migration was calcu-lated based on the total initial input of cells per well.
Results and DiscussionMolecular cloning and analysis of the novel un-spliced CXCR4-Lo variant cDNA
We previously demonstrated the expression of two distinctmRNAs, ;1.7-kb and ;4.0-kb transcripts for CXCR4 inHUVECs, and had speculated on the putative existence of an al-ternatively spliced isoform (7). Subsequently, we used bioinfor-matics to identify homologous ESTs with sequence variation in theNH2-terminal region of the EC domain of CXCR4. Two variantcDNA clones, designated as CXCR4-Lo, were isolated from a hu-man neutrophil cDNA library and sequenced. Unlike CXCR4, the1071-bp ORF of CXCR4-Lo cDNA clone is encoded by a singleexon. The coding region is initiated from an alternate in-frameATG start codon found within the intron sequence characterized inthe recently published genomic structure for CXCR4 (20, 21) and25-bp upstream of the known AG acceptor site used for intronsplicing (Fig. 1,A andB). Significantly, the predicted full-length357-aa CXCR4-Lo receptor (not shown, GenBank accession no.AF147204) contains a longer neo-NH2-terminal EC domain that isdifferent from the corresponding CXCR4 sequence by the first 9 aa
residues M-S-I-P-L-P-L-L-Q (Fig. 1,A andB), while the remain-ing sequence is identical. The 39 untranslated region of CXCR4-Lo(not shown) has an AATAAA polyadenylation consensus signaland is identical with the published genomic sequence of humanCXCR4 (20).
Although alternative splicing was recently demonstrated for mu-rine CXCR4 (18, 19, 22), the human CXCR4-Lo we have discov-ered is structurally distinct and generated by a unique mechanism.In the case of murine CXCR4b (18, 19, 22), a different GT splicedonor site is used to yield an isoform that is shorter by 2 aa, whilethe overall functional activity, genomic structure, and intron splic-ing mechanism are conserved (18, 22). In contrast, in humanCXCR4-Lo, the known intron remains unspliced and the entiregene is expressed as a single exon to yield the longer 4.0-kb tran-script (Fig. 1B). Other cases of retention and read through of “codingintrons” have also been observed, most recently with the generation ofa new CTL Ag from a tyrosinase-related protein mRNA (24).
To further test this hypothesis, and also address the questionwhether the two human CXCR4 isoforms are products of a singlegene, RT-PCR primers (Lo-346 and Lo-1071) were designed toselectively amplify from the unique 59-end of CXCR4-Lo tran-scripts from HL-60 cells. Hybridization of amplified cDNA withthe CXCR4-Lo cDNA-specific 346-bp DNA probe revealed theexpected specificity of the 1071-bp PCR product (Fig. 1C). In
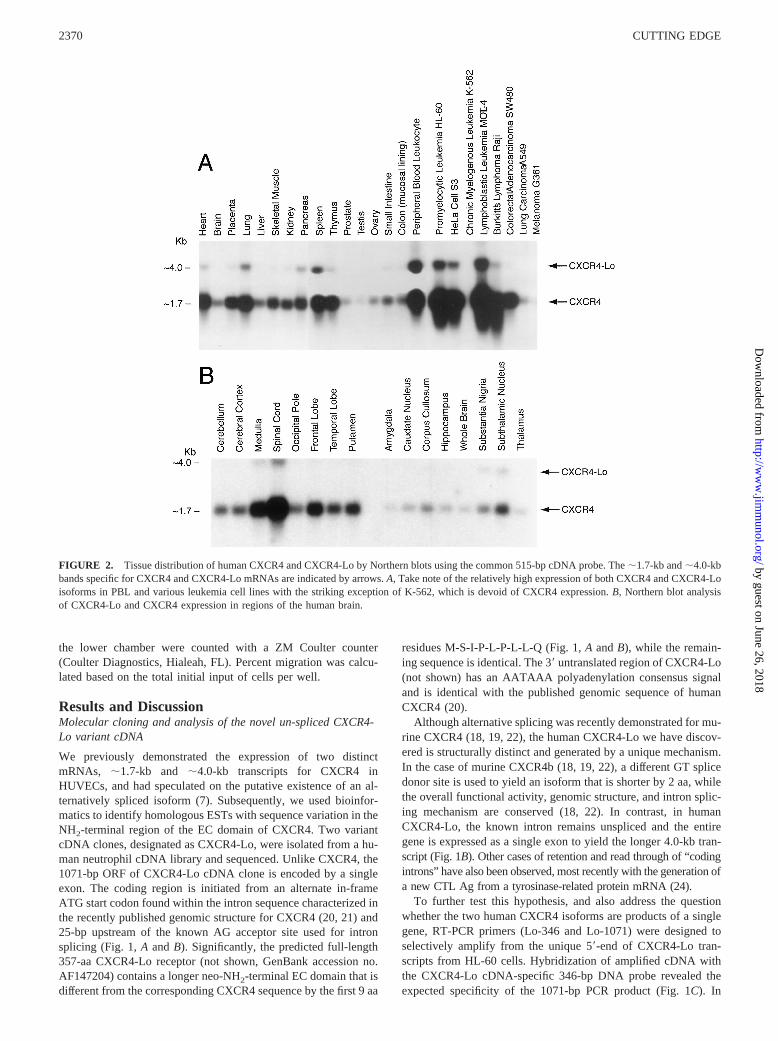
FIGURE 2. Tissue distribution of human CXCR4 and CXCR4-Lo by Northern blots using the common 515-bp cDNA probe. The;1.7-kb and;4.0-kbbands specific for CXCR4 and CXCR4-Lo mRNAs are indicated by arrows.A, Take note of the relatively high expression of both CXCR4 and CXCR4-Loisoforms in PBL and various leukemia cell lines with the striking exception of K-562, which is devoid of CXCR4 expression.B, Northern blot analysisof CXCR4-Lo and CXCR4 expression in regions of the human brain.
2370 CUTTING EDGE
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
addition, Northern analysis with the 346-bp probe also confirmedexpression of the;4.0-kb CXCR4-Lo mRNA in PBLs and othertissues (Fig. 1D).
The qualitative comparison of CXCR4 and CXCR4-Lo mRNAexpression in tissues was done using the common 515-bp cDNAprobe (7). As noted previously (4, 5), CXCR4 is highly expressedin most human tissues upon Northern blot analysis (Fig. 2). Whileoverall CXCR4-Lo expression is either proportionally lower orabsent in all tissues (Fig. 2A), there is relatively more expressionof its mRNA in spleen, lung, PBLs, and cancer cell lines likeHL-60 and MOLT-4. Such selective differences in tissue-specificexpression of the unspliced CXCR4-Lo isoform implies that itsexpression is regulated, rather than being caused by a random orpassive absence of normal splicing. However, additional studiesincluding the role of the “coding intron” in modulating transcrip-
tion are desirable to assess the functional significance ofCXCR4-Lo expression. Furthermore, although CXCR4 expressionis low in whole brain (Fig. 2,A and B), especially high mRNAexpression was observed in spinal cord, medulla, and frontal lobe,with moderate to low expression in putamen, temporal lobe, cer-ebellum, and cerebellar cortex. Significant, though lower, concom-itant expression of the;4.0-kb CXCR4-Lo mRNA was also ob-served in the spinal cord, medulla, substantia nigra, andsubthalamic nucleus (Fig. 2B). The functional consequence of suchan expression pattern of both CXCR4 and CXCR4-Lo in brain isnot known, although it was recently shown that fetal cerebellardevelopment is impaired in the CXCR4-knockout mice (10).
Heterologous functional expression of CXCR4-Lo andcomparison with CXCR4
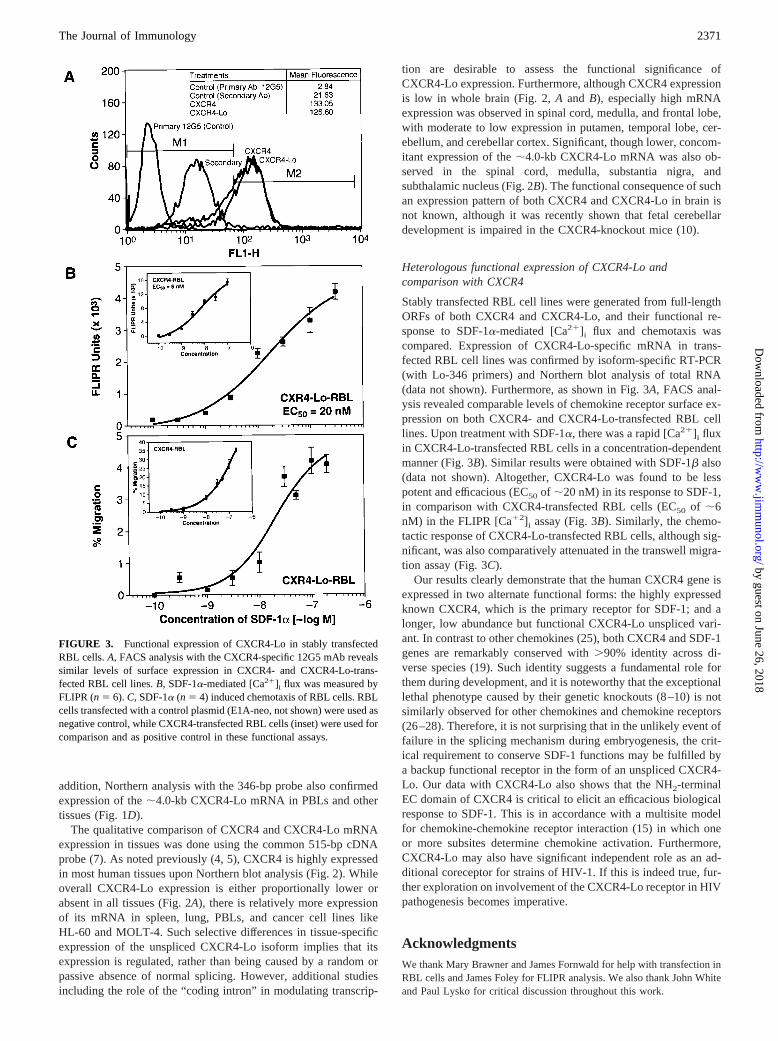
Stably transfected RBL cell lines were generated from full-lengthORFs of both CXCR4 and CXCR4-Lo, and their functional re-sponse to SDF-1a-mediated [Ca21]i flux and chemotaxis wascompared. Expression of CXCR4-Lo-specific mRNA in trans-fected RBL cell lines was confirmed by isoform-specific RT-PCR(with Lo-346 primers) and Northern blot analysis of total RNA(data not shown). Furthermore, as shown in Fig. 3A, FACS anal-ysis revealed comparable levels of chemokine receptor surface ex-pression on both CXCR4- and CXCR4-Lo-transfected RBL celllines. Upon treatment with SDF-1a, there was a rapid [Ca21]i fluxin CXCR4-Lo-transfected RBL cells in a concentration-dependentmanner (Fig. 3B). Similar results were obtained with SDF-1b also(data not shown). Altogether, CXCR4-Lo was found to be lesspotent and efficacious (EC50 of ;20 nM) in its response to SDF-1,in comparison with CXCR4-transfected RBL cells (EC50 of ;6nM) in the FLIPR [Ca12]i assay (Fig. 3B). Similarly, the chemo-tactic response of CXCR4-Lo-transfected RBL cells, although sig-nificant, was also comparatively attenuated in the transwell migra-tion assay (Fig. 3C).
Our results clearly demonstrate that the human CXCR4 gene isexpressed in two alternate functional forms: the highly expressedknown CXCR4, which is the primary receptor for SDF-1; and alonger, low abundance but functional CXCR4-Lo unspliced vari-ant. In contrast to other chemokines (25), both CXCR4 and SDF-1genes are remarkably conserved with.90% identity across di-verse species (19). Such identity suggests a fundamental role forthem during development, and it is noteworthy that the exceptionallethal phenotype caused by their genetic knockouts (8–10) is notsimilarly observed for other chemokines and chemokine receptors(26–28). Therefore, it is not surprising that in the unlikely event offailure in the splicing mechanism during embryogenesis, the crit-ical requirement to conserve SDF-1 functions may be fulfilled bya backup functional receptor in the form of an unspliced CXCR4-Lo. Our data with CXCR4-Lo also shows that the NH2-terminalEC domain of CXCR4 is critical to elicit an efficacious biologicalresponse to SDF-1. This is in accordance with a multisite modelfor chemokine-chemokine receptor interaction (15) in which oneor more subsites determine chemokine activation. Furthermore,CXCR4-Lo may also have significant independent role as an ad-ditional coreceptor for strains of HIV-1. If this is indeed true, fur-ther exploration on involvement of the CXCR4-Lo receptor in HIVpathogenesis becomes imperative.
AcknowledgmentsWe thank Mary Brawner and James Fornwald for help with transfection inRBL cells and James Foley for FLIPR analysis. We also thank John Whiteand Paul Lysko for critical discussion throughout this work.
FIGURE 3. Functional expression of CXCR4-Lo in stably transfectedRBL cells.A, FACS analysis with the CXCR4-specific 12G5 mAb revealssimilar levels of surface expression in CXCR4- and CXCR4-Lo-trans-fected RBL cell lines.B, SDF-1a-mediated [Ca21]i flux was measured byFLIPR (n5 6). C, SDF-1a (n 5 4) induced chemotaxis of RBL cells. RBLcells transfected with a control plasmid (E1A-neo, not shown) were used asnegative control, while CXCR4-transfected RBL cells (inset) were used forcomparison and as positive control in these functional assays.
2371The Journal of Immunology
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

References1. Tashiro, K., H. Tada, R. Heilker, M. Shirozu, T. Nakano, and T. Honjo. 1993.
Signal sequence trap: a cloning strategy for secreted proteins and type I mem-brane proteins.Science 261:600.
2. Oberlin, E., A. Amara, F. Bachelerie, C. Bessia, J.-L. Virelizier,F. Arenzana-Seisdedos, O. Schwartz, J.-L. Heard, I. Clark-Lewis, D. F. Legler,M. Loetscher, M. Baggiolini, and B. Moser. 1996. The CXC chemokine SDF-1is the ligand for LESTR/fusin and prevents infection by T-cell-line-adaptedHIV-1. Nature 382:833.
3. Shirozu, M., T. Nakano, J. Inazawa, K. Tashiro, H. Tada, T. Shinohara, andT. Honjo. 1995. Structure and chromosomal localization of the human stromalcell-derived factor 1 (SDF1) gene.Genomics 28:495.
4. Rimland, J., W. Xin, P. Sweetnam, K. Saijoh, E. J. Nestler, and R. S. Duman1991. Sequence and expression of a neuropeptide Y receptor cDNA.Mol. Phar-macol. 40:869.
5. Federsppiel, B., I. G. Melhado, A. M. V. Duncan, A. Delaney, K. Schappert,I. Clark-Lewis, and F. R. Jirik. 1993. Molecular cloning of the cDNA and chro-mosomal localization of the gene for a putative seven-transmembrane segment(7-TMS) receptor isolated from human spleen.Genomics 16:707.
6. Bleul, C. C., R. C. Fuhlbrigge, J. M. Casanovas, A. Aiuti, and T. A. Springer.1996. A highly efficacious lymphocyte chemoattractant, stromal cell-derived fac-tor 1 (SDF-1).J. Exp. Med. 184:1101.
7. Gupta, S. K., P. G. Lysko, K. Pillarisetti, E. Ohlstein, and J. M. Stadel. 1998.Chemokine receptors in human endothelial cells: functional expression ofCXCR4 and its transcriptional regulation by inflammatory cytokines.J. Biol.Chem. 273:4282.
8. Nagasawa, T., S. Hirota, K. Tachibana, N. Takakura, S. Nishikawa, Y. Kitamura,N. Yoshida, H. Kikutani, and T. Kishimoto. 1996. Defects of B-cell lymphopoi-esis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1.Nature 382:635.
9. Tachibana, K., S. Hirota, H. Iizasa, H. Yoshida, K. Kawabata, Y. Kataoka,Y. Kitamura, K. Matsushima, N. Yoshida, S. Nishikawa, T. Kishimoto, andT. Nagasawa. 1998. The chemokine receptor CXCR4 is essential for vascular-ization of the gastrointestinal tract.Nature 393:591.
10. Zou, Y.-R., A. H. Kottman, M. Kuroda, I. Taniuchi, and D. R. Littman. 1998.Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellardevelopment.Nature 393:595.
11. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entrycofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupledreceptor.Science 272:872.
12. Alkhatib, G., C. Comadiere, C. C. Broder, Y. Feng, P. E. Kennedy,P. M. Murphy, and E.A. Berger. 1996. CC CKR5: a RANTES, MIP-1a, MIP-1breceptor as a fusion cofactor for macrophage-tropic HIV-1.Science 272:1955.
13. LaRosa, G. J., K. M. Thomas, M. E. Kaufmann, R. Mark, M. White, L. Taylor,G. Gray, D. Witt, and J. Navarro. 1992. Amino terminus of the interleukin-8receptor is a major determinant of receptor subtype specificity.J. Biol. Chem.267:25402.
14. Hebert, C., A. Chuntharapai, M. Smith, T. Colby, J. Kim, and R. Horuk. 1993.Partial functional mapping of the human interleukin-8 type A receptor: identifi-cation of a major ligand binding domain.J. Biol. Chem. 268:18549.
15. Monteclaro, F. S., and I. F. Charo. 1996. The amino-terminal extracellular do-main of the MCP-1 receptor, but not the RANTES/MIP-1 alpha receptor, confers
chemokine selectivity: evidence for a two-step mechanism for MCP-1 receptoractivation.J. Biol. Chem. 271:19084.
16. Pease, J. E., J. Wang, P. D. Ponath, and P. M. Murphy. 1998. The N-terminalextracellular segments of the chemokine receptors CCR1 and CCR3 are deter-minants for MIP-1a and eotaxin binding, respectively, but a second domain isessential for efficient receptor activation.J. Biol. Chem. 273:19972.
17. Ruckers, J., M. Samson, B. J. Doranz, F. Libert, J. F. Berson, C. C. Broder,G. Vassart, R. W. Doms, and M. Parmentier. 1996. Regions inb-chemokinereceptors CCR5 and CCR2b that determine HIV-1 cofactor specificity.Cell 87:437.
18. Heesen, M., M. A. Berman, U. E. Hopken, N. P, Gerard, and M. E. Dorf. 1997.Alternate splicing of mouse fusin/CXC chemokine receptor-4: stromal cell-de-rived factor-1a is a ligand for both CXC chemokine receptor-4 isoforms.J. Im-munol. 158:3561.
19. Moepps, B., R. Frodl, H. Rodewald, M. Baggiolini, and P. Gierschik. 1997. Twomurine homologues of the human chemokine receptor CXCR4 mediating stromalcell-derived factor 1a activation of Gi2 are differentially expressed in vivo.Eur.J. Immunol. 27:2102.
20. Wegner, S.A., P. K. Ehrenberg, G. Chang, D. E. Dayhoff, A. L. Sleeker, andN. L. Michael. 1998. Genomic organization and functional characterization of thechemokine receptor CXCR4, a major entry co-receptor for human immunodefi-ciency virus type 1.J. Biol. Chem. 273:4754.
21. Caruz, A., M. Samsom, J. M. Alonso, J. Alcami, F. Baleux, J. L. Virelizier,M. Parmentier, and F. Arenzana-Seisdedos. 1998. Genomic organization and pro-moter characterization of human CXCR4 gene.FEBS Lett. 426:271.
22. Frodl. R., P.Gierschik, and B. Moepps. 1998. Genomic organization and expres-sion of the CXCR4 gene in mouse and man: absence of a splice variant corre-sponding to mouse CXCR4-b in human tissues.J. Recept. Signal Transduct. Res.18:321.
23. Dooley, D. J., J. J. Geer, S. J. Haleen, A.W. Probert, and K. M. Welch. 1998.Assessment of endothelin receptor subtype-mediated increases of [Ca21]i in dis-tinct rat cell types using fluorimetric imaging.J. Cardiovasc. Pharmacol.31(Suppl. 1):S192.
24. Lupetti, R., P. Pisarra, A. Verrecchia, C. Farina, G. Nicolini, A. Anichini,C. Bordignon, M. Sensi, G. Parmiani, and C. Travesari. 1998. Translation of aretained intron in tyrosinase-related protein (TRP) 2 mRNA generates a newcytotoxic T lymphocyte (CTL)-defined and shared human melanoma antigen notexpressed in normal cells of the melanocytic lineage.J. Exp. Med. 188:1005.
25. Crump, P. M., J.-H. Gong, P. Loetscher, K. Rajarathanam, A. Amara,F. Arenzana-Seisdedos, J.-L. Virelizier, M. Baggiolini, B. D. Sykes, andI. Clark- Lewis. 1997. Solution structure and basis for functional activity ofstromal cell-derived factor-1: dissociation of CXCR4 activation from binding andinhibition of HIV-1. EMBO J. 16:6996.
26. Cacalano, G., J. Lee, K. Kikly, A. M. Ryan, S. Pitts-Meek, B. Hultgren,W. I. Wood, and M. W. Moore. 1994. Neutrophil and B cell expansion in micethat lack the murine IL-8 receptor homolog.Science 265:682.
27. Rothenberg, M. E., J. A. MacLean, E. Pearlman, A. D. Luster, and L. Philip.1997. Targeted disruption of the chemokine eotaxin partially reduces antigen-induced tissue eosinophilia.J. Exp. Med. 185:785.
28. Boring, L., J. Gosling, M. Cleary, and I. F. Charo. 1998. Decreased lesion for-mation in CCR22/2 mice reveals a role for chemokines in the initiation of ath-erosclerosis.Nature 394:894.
2372 CUTTING EDGE
by guest on June 26, 2018http://w
ww
.jimm
unol.org/D
ownloaded from