current immunity markers in insect ecolog
DESCRIPTION
paperTRANSCRIPT

Bulletin of Entomological Researchhttp://journals.cambridge.org/BER
Additional services for Bulletin of Entomological Research:
Email alerts: Click hereSubscriptions: Click hereCommercial reprints: Click hereTerms of use : Click here
Current immunity markers in insect ecological immunology: assumed tradeoffs and methodological issues
M. MorenoGarcía, A. CórdobaAguilar, R. Condé and H. LanzMendoza
Bulletin of Entomological Research / FirstView Article / August 2012, pp 1 13DOI: 10.1017/S000748531200048X, Published online:
Link to this article: http://journals.cambridge.org/abstract_S000748531200048X
How to cite this article:M. MorenoGarcía, A. CórdobaAguilar, R. Condé and H. LanzMendoza Current immunity markers in insect ecological immunology: assumed tradeoffs and methodological issues. Bulletin of Entomological Research, Available on CJO doi:10.1017/S000748531200048X
Request Permissions : Click here
Downloaded from http://journals.cambridge.org/BER, IP address: 132.248.9.8 on 30 Aug 2012

Current immunity markers in insectecological immunology: assumed
trade-offs and methodological issues
M. Moreno-García1,2, A. Córdoba-Aguilar1, R. Condé2and H. Lanz-Mendoza2*
1Departamento de Ecología Evolutiva, Instituto de Ecología, UniversidadNacional Autónoma de México, Apdo. P. 70-275, Circuito Exterior, Ciudad
Universitaria, 04510, Coyoacán, Distrito Federal, México: 2Centro deInvestigaciones Sobre Enfermedades Infecciosas, Instituto Nacional de Salud
Pública, Avenida Universidad 655, Santa María Ahuacatitlán,62100 Cuernavaca, Morelos, México
Abstract
The field of ecological immunology currently relies on using a number of immuneeffectors or markers. These markers are usually used to infer ecological trade-offs(via conflicts in resource allocation), though physiological nature of these markersremains elusive. Here, we review markers frequently used in insect evolutionaryecology research: cuticle darkening, haemocyte density, nodule/capsule formation,phagocytosis and encapsulation/melanization via use of nylon filaments and beads,phenoloxidase activity, nitric oxide production, lysozyme and antimicrobial peptideproduction. We also provide physiologically based information that may shed lighton the probable trade-offs inferred when these markers are used. In addition, weprovide a number of methodological suggestions to improve immune markerassessment.
Keywords: evolutionary ecology, insect immunity, immune markers
(Accepted 21 June 2012)
Introduction
Ecological immunology is a rapidly expanding, youngdiscipline (Rolff & Reynolds, 2009). Within this field, immuneresponse is seen as a set of traits that involve costs (Sheldon& Verhulst, 1996). Resources must be allocated between anorganism’s maintenance and proper functioning and theexpression of other traits linked with survival and reproduc-tion, resulting in ecological trade-offs among trait types. At themoment, there is growing empirical evidence of trade-offsassociated between life-history traits (size at birth, growth and
mortality rates, size and age at maturity, longevity and sur-vival, clutch size and reproductive effort: reviewed in Stearns,1992) and immune defense (e.g. Boots & Begon, 1993;Kraaijeveld & Godfray, 1997; Fellowes et al., 1998; Moret &Schmid-Hempel, 2000; Rigby & Jokela, 2000; Hoang, 2001).
The ecological immunity framework has been applied to anumber of disciplines, including sexual selection (i.e. differ-ential mating and fertilization success: Darwin, 1871) (e.g.Siva-Jothy, 2000; McKean &Nunney, 2001; Roberts et al., 2004)and, to a lesser extent, social evolution (e.g. Calleri et al., 2007;Bocher et al., 2008), predator-prey relationships (e.g. Packeret al., 2003; Roy & Holt, 2008) and parental investment (e.g.Hoi-Leitner et al., 2001; Brzek & Konarzewski, 2007). Theinfluence of ecological immunity has also extended to broaderfields, such as life-history theory (Schmid-Hempel, 2005a),parasite-host coevolution (e.g. Day et al., 2007; Duffy & Sivars-Becker, 2007), conservation biology (e.g. Stevenson, 2006;
*Author for correspondenceFax: +52 777 317 5485E-mail: [email protected]
Bulletin of Entomological Research, Page 1 of 13 doi:10.1017/S000748531200048X© Cambridge University Press 2012

Tarlow & Blumstein, 2007) and learning (e.g. Barnard et al.,2006), among others.
One animal group where ecological immunology hasbeen extensively investigated is insects (see, for example,Lawniczak et al., 2006). Insects have properties (e.g. small size,large sample sizes, short development times) that allow awiderange of experiments. Insect studies have measured a numberof immune effectors related to both cellular and humoraldefence reactions (hereafter referred to as immunemarkers), inwhich a cost of such immune response, measured throughthese markers, is assumed (see also Adamo, 2004). The logicof choosing such markers has been based on the assumptionthat they protect the insect, but their production negativelycorrelates with other life history or intermediate traits(e.g. colored traits, behavioral traits, etc.). Many feasible andhighly sensitive techniques in insect immunology (e.g. proteo-mic, transcriptomic, liquid chromatography/mass spectro-metry) have been employed at the individual-level, but costsconfine these approaches to exploratory or qualitative studiesat the population level. This turns into a restriction when theobjects of study are primarily numerous individuals in naturalpopulations, because differences among organisms in im-mune responses (and their trade-offs among other behaviouralor reproductive traits) could be an important cause of adaptivechange of the population.
Here, we review each of the most common immunemarkers used in ecological and evolutionary studies thatcan be utilized across large sample sizes. Our first aim is toidentify, when possible, the physiological processes behindeach marker. This may allow the inference of trade-offs withother functions through individual data. Second, we brieflyoutline the methodological inconveniences in the assessmentof such markers and suggest solutions. Our aim is to provide aset of more robust methodological practices. An outline ofthese two aims appears in table 1; we present each in greaterdetail in the text below. First, however, we present a basicshort review of how insect immunity works, which serves asthe framework for discussing the trade-offs and methodologi-cal issues.
Insect immune mechanisms
The insect immune system is formed by a set of cells,molecules and reactions. All of these features are continuouslyevolving to resist (attack and eliminate) pathogen invasionand to limit the negative consequences of the infection(Hoffman & Reichhart, 2002; Schmid-Hempel, 2005a; Ayres& Schneider, 2009). The first lines of defence include theexoskeleton cuticle, physical and chemical properties (e.g. pH)of the epidermis, gut epithelium, and male and female re-productive accessory glands (Gillespie et al., 1997; Casteels,1998). These tissues also secrete lysozymes and cytotoxicmolecules such as reactive oxygen species (ROS: e.g. super-oxide anions, peroxides, hydroxyl radicals) (Schmid-Hempel,2005a). Pathogens are mainly recognized by the membrane ofhaemocytes (which occur free in the haemolymph) or by themembranes of epithelial cells. These membranes bear proteinscalled pattern recognition receptors (PRRs) that recognizeconserved molecular features of pathogens called pathogen-associated molecular patterns (PAMPs: LPS, mannoses, β-1,3 glucans and peptidoglycans) (Gillespie et al., 1997). Oncepathogens are recognized as non-self, cellular and humoralimmune mechanisms are activated. Cellular responses in-clude phagocytosis, nodulation and encapsulation. During
encapsulation, small and large pathogens are surroundedand bound by haemocytes (Gillespie et al., 1997). Duringnodulation and encapsulation, a melanin layer is constructed(frequently referred to as melanotic nodulation/encapsula-tion) to cover foreign agents, which ultimately die by anoxia,toxic ROS or starvation (Nappi & Ottoviani, 2000; Narayanan,2004).
Melanin production is a consequence of the phenoloxidase(PO) cascade (Söderhäll & Cerenius, 1998). During thecascade, opsonic factors, ROS and cytotoxins, such asquinones and semiquinones, are produced. These importantintermediate molecules are highly reactive and toxic to patho-gens, and serve to amplify the immune response (Nappi &Ottovianni, 2000; Cerenius & Söderhäll, 2004). PO is involvedin various physiological processes, including cuticular scler-otization and immune defense (such as melanotic encapsula-tion and wound healing). Three kinds of PO exist (Sugumaran& Kanost, 1993): monophenol monooxigenase (also referredto as tyrosinase-type PO: Ashida & Brey, 1997), ο-diphenol-oxidase (also referred to as cathecoloxidase-type PO: Decker& Jaenicke, 2004) and ρ-diphenoloxidase (also referred toas laccase-type PO: Sugumaran & Kanost, 1993). These threekinds of PO and their activating systems are structurallyalmost indistinguishable; hence, the term phenoloxidase isoften used in the literature without distinguishing amongthem. PO associated with sclerotization and pigmentation ofcuticle (monophenol monooxigenase and ρ-diphenoloxidase)appears to be under hormonal control (Ashida & Brey, 1997),while ο-diphenoloxidase is triggered by recognition of non-self particles (Ashida & Brey, 1997). Haemolymph PO ismainly kept inside or near haemocytes; nevertheless, it can bea feature of the humoral response. The precursor of PO, pro-phenoloxidase (proPO) is primarily released from haemocytesinto the haemolymph after contact with non-self particles(Ling & Yu, 2006). It also contributes to humoral melanizationof pathogens. The general PO activation pathway is initiatedwhen phenylalanine is hydroxylated and converted intotyrosine. Then, proPO is converted by specific protease clea-vage into active form PO. PO catalyzes both the hydroxylationof tyrosine to dopa and the oxidation of dopa to dopaquinone.Finally, dopaquinone is converted to melanin, which is usedfor wrapping pathogens and wound clotting (Söderhäll &Cerenius, 1998; Christensen et al., 2005; Nappi & Christensen,2005).
Other humoral responses include antimicrobial systemicmolecules, which are also synthesized mainly in the fat bodyand reproductive accessory glands, gut cells and haemocytes(Manetti et al., 1998; Schmid-Hempel, 2005a). Most of thesemolecules are secreted close to the cuticle or into the haemo-lymph, gut tract or malpighian tubes. Antimicrobial peptide(AMP) molecules induce a number of negative effects onpathogen membranes including membrane collapse, preven-tion of cell division and permeability disruption (Otvos Jr,2000; Bulet et al., 2003). Nitric oxide (NO) is another immune-relevant molecule; it is a highly reactive and unstable freeradical gas that crosses cell membranes to act on nearby targets(Müller, 1997). NO also inhibits protein catalytic activity andhas damaging effects on pathogen protein and DNA (re-viewed in Rivero, 2006). Molecules such as ROS can damagepathogen nucleic acids, proteins and cell membrane (Nappiet al., 2000; Herrera-Ortiz et al., 2004). Lysozymes arehydrolytic enzymes that cleave the glycosidic bond betweenN-acetylmuramic acid and N-acetylglucosamine in peptido-glycan, a major component of the Gram-wall polymer
M. Moreno-García et al.2

(Jollès, 1996). It must be mentioned that most of theinformation regarding the humoral response has beenobtained under laboratory conditions. Further investigation/characterization of these molecules must be done in the wildincluding the natural pathogens of each host.
Trade-offs among immune responses
Trade-offs within immune system effectors are alsopossible. In an individual, different traits are frequentlyfound to be genetically or phenotypically correlated. Geneticcorrelation arises because a single gene can influence multipletraits in positive or negative fashions (pleiotropy andantagonistic pleiotropy, respectively) or because of linkagedisequilibrium between genes affecting different characters(Falconer & Mackay, 1996). Phenotypic correlations includethe positive or negative influences of environmental factors ontraits (Roff, 1992). Reactions and products of immune responseare interconnected and the kind of response could be related tothe type of attacking pathogen. A number of studies haveexamined genetic and phenotypic correlations among encap-sulation, lytic activity, cuticular darkness, PO activity andhaemocyte load. In summary, these studies report positiveand negative correlations, resulting in a lack of a clear patternacross species or even populations (see Ryder & Siva-Jothy,2001; Rantala & Kortet, 2003; Cotter et al., 2004; Fedorka et al.,2004; Rantala & Roff, 2005; Roff et al., 2005; Moreno-Garcíaet al., 2010). This point will be reconsidered later.
Immune markers used in evolutionary ecology research
Cuticle darkness
The main components of insect cuticle – quinones andmelanin (see Phenoloxidase activity) – are also used for otherfunctions, thus strongly suggesting a trade-off (e.g. Barnes &Siva-Jothy, 2000; Armitage & Siva-Jothy, 2005). Quinone com-pounds determine cuticle pigmentation (via sclerotization) bymaking covalent links with proteins, resulting in colouredproducts, a process called quinone tanning (Riddiford &Hiruma, 1988). Quinones also form methide derivatives thatcrosslink cuticle proteins through the quinone side chain(β-sclerotization reaction) and are responsible for cuticlehardening (Saul & Sugumaran, 1988). Cuticle darkness, thus,is related to how much quinone is deposited in every shed orwound repair site.
Methodology
Cuticular colour is frequently assessed under a fluorescentwhite light and the emerging colour classes are analyzed usinga greyscale value (e.g. Barnes & Siva-Jothy, 2000; Thompsonet al., 2002; Rolff et al., 2005). However, since the cuticle struc-ture is composed of hundreds of proteins (Andersen et al.,1995), chitin and lipids (Chapman, 1998), and such proteincomposition greatly influences cuticle mechanical properties(e.g. flexibility: Haas et al., 2000; Gosline et al., 2002); therecould be a possible overestimation of quinine and melanintanning. Furthermore, since cuticles among insect taxa varyconsiderably in stiffness, hardness and pigmentation, it ispossible that the dark aspect of a particular tissue may notnecessarily reflect or be related to a reduced or heightenedimmune ability. In combinationwith cuticular colour analysis,we recommend the use of transverse section preparations(using a microtome) from different representative regions of
the insect body and cuticle width measurement. This wouldreflect how much the insect has invested in producing a thickcuticle and, presumably, quinines and melanin. Note, how-ever, that cuticular melanization is not always associated withgreater immunogenic response (see Robb et al., 2003).
Cellular responses
Total number of haemocytes (haemocyte density)
Haemocyte counting and/or activity is a straightforwardway of assessing cellular immune ability (e.g. Rantala et al.,2000; Kraaijeveld et al., 2001; Cotter et al., 2004). Haemocytesare generally responsible for immune activities, such asengulfing, surrounding and destroying infectious agents.After bacteria have penetrated the haemocoel, haemocytesare the first and most efficient bacteria-clearing mechanism,(Hillyer et al., 2003). However, there are different haemocytetypes, each playing a different function (recognition, phago-cytosis, nodulation, encapsulation or melanization) (see re-views by Lavine & Strand, 2002; Ribeiro & Brehélin, 2006).Differences in pathogen type (e.g. bacteria, virus, nematodes,etc.) among individuals, sexes or species must be considered,for example, for example, the mosquito Aedes aegypti mainlyphagocyte Escherichia coli. In contrast, other types of bacteria(e.g. Micrococcus luteus) are melanized (Hillyer et al., 2003).However, the factors eliciting phagocytic vs. melanizationresponses against bacteria are independent of Gram type(Hillyer et al., 2004).
Methodology
For total and viable counts, haemolymph is collected andincubated in humidity chamber, then counted using an hemo-cytometer using (phase contrast) microscopy (e.g. Rantalaet al., 2000; Kraaijeveld et al., 2001). For collection and incu-bation of haemolymph, we suggest the use of cell culturemedia, such as RPMI, Schneider or Grace rather than PBSbuffers. This media may allow a better viability of cells.Trypan blue can be used to differentiate between dead andviable cells. Haemolymph extraction may be accomplished byperfusion-bleed; a droplet can be collected from a puncture/cut on the thorax or by a removed leg. Although this mayappear an easy task, haemocytes can adhere to tissues such asthe fat body. One solution to this problem is to use anti-coagulant buffers of low pH (Pech et al., 1994). An accurateextraction of these cells can also be carried out by injecting aprotease inhibitor blend (e.g. PMSF, TLCK, leupeptine).However, optimal conditions for obtaining insect haemocytescannot be generalized; and, in each procedure of extraction,the use (or non-use) of anticoagulant must be adjusted.
Phagocytosis
Phagocytic activities require the internalization of thesurface membrane and action of several organelles, includingendosomes and lysosomes, which fuse with the plasma mem-brane and provide the membrane for phagosome formation(Desjardins et al., 2005). In mosquitoes, the synthesis of lipidsfor the membranes of the phagosome, plasma membrane andfor the production of cytotoxic proteins for the degradation ofphagocytised material decreases their reproductive output(Ferdig et al., 1993; Hogg&Hurd, 1995; Hillyer et al., 2003). Forthese reasons, trade-offs are expected among phagocytic
Insect immuno-ecology markers 3
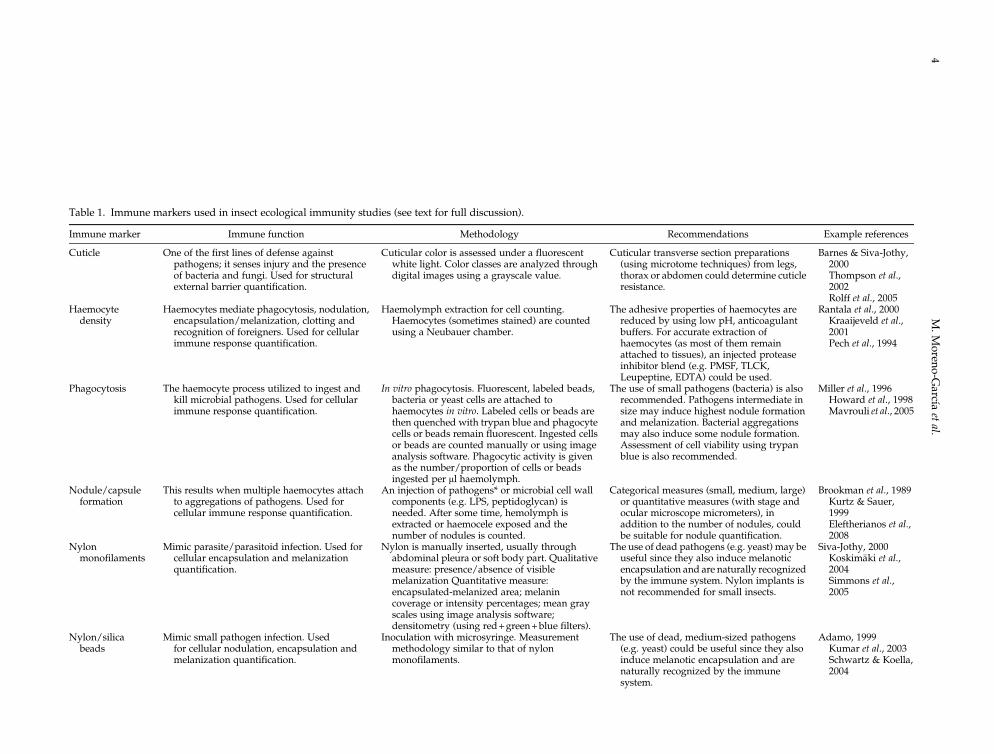
Table 1. Immune markers used in insect ecological immunity studies (see text for full discussion).
Immune marker Immune function Methodology Recommendations Example references
Cuticle One of the first lines of defense againstpathogens; it senses injury and the presenceof bacteria and fungi. Used for structuralexternal barrier quantification.
Cuticular color is assessed under a fluorescentwhite light. Color classes are analyzed throughdigital images using a grayscale value.
Cuticular transverse section preparations(using microtome techniques) from legs,thorax or abdomen could determine cuticleresistance.
Barnes & Siva-Jothy,2000Thompson et al.,2002Rolff et al., 2005
Haemocytedensity
Haemocytes mediate phagocytosis, nodulation,encapsulation/melanization, clotting andrecognition of foreigners. Used for cellularimmune response quantification.
Haemolymph extraction for cell counting.Haemocytes (sometimes stained) are countedusing a Neubauer chamber.
The adhesive properties of haemocytes arereduced by using low pH, anticoagulantbuffers. For accurate extraction ofhaemocytes (as most of them remainattached to tissues), an injected proteaseinhibitor blend (e.g. PMSF, TLCK,Leupeptine, EDTA) could be used.
Rantala et al., 2000Kraaijeveld et al.,2001Pech et al., 1994
Phagocytosis The haemocyte process utilized to ingest andkill microbial pathogens. Used for cellularimmune response quantification.
In vitro phagocytosis. Fluorescent, labeled beads,bacteria or yeast cells are attached tohaemocytes in vitro. Labeled cells or beads arethen quenched with trypan blue and phagocytecells or beads remain fluorescent. Ingested cellsor beads are counted manually or using imageanalysis software. Phagocytic activity is givenas the number/proportion of cells or beadsingested per μl haemolymph.
The use of small pathogens (bacteria) is alsorecommended. Pathogens intermediate insize may induce highest nodule formationand melanization. Bacterial aggregationsmay also induce some nodule formation.Assessment of cell viability using trypanblue is also recommended.
Miller et al., 1996Howard et al., 1998Mavrouli et al., 2005
Nodule/capsuleformation
This results when multiple haemocytes attachto aggregations of pathogens. Used forcellular immune response quantification.
An injection of pathogens* or microbial cell wallcomponents (e.g. LPS, peptidoglycan) isneeded. After some time, hemolymph isextracted or haemocele exposed and thenumber of nodules is counted.
Categorical measures (small, medium, large)or quantitative measures (with stage andocular microscope micrometers), inaddition to the number of nodules, couldbe suitable for nodule quantification.
Brookman et al., 1989Kurtz & Sauer,1999Eleftherianos et al.,2008
Nylonmonofilaments
Mimic parasite/parasitoid infection. Used forcellular encapsulation and melanizationquantification.
Nylon is manually inserted, usually throughabdominal pleura or soft body part. Qualitativemeasure: presence/absence of visiblemelanization Quantitative measure:encapsulated-melanized area; melanincoverage or intensity percentages; mean grayscales using image analysis software;densitometry (using red+green+blue filters).
The use of dead pathogens (e.g. yeast) may beuseful since they also induce melanoticencapsulation and are naturally recognizedby the immune system. Nylon implants isnot recommended for small insects.
Siva-Jothy, 2000Koskimäki et al.,2004Simmons et al.,2005
Nylon/silicabeads
Mimic small pathogen infection. Usedfor cellular nodulation, encapsulation andmelanization quantification.
Inoculation with microsyringe. Measurementmethodology similar to that of nylonmonofilaments.
The use of dead, medium-sized pathogens(e.g. yeast) could be useful since they alsoinduce melanotic encapsulation and arenaturally recognized by the immunesystem.
Adamo, 1999Kumar et al., 2003Schwartz & Koella,2004
M.M
oreno-García
etal.4
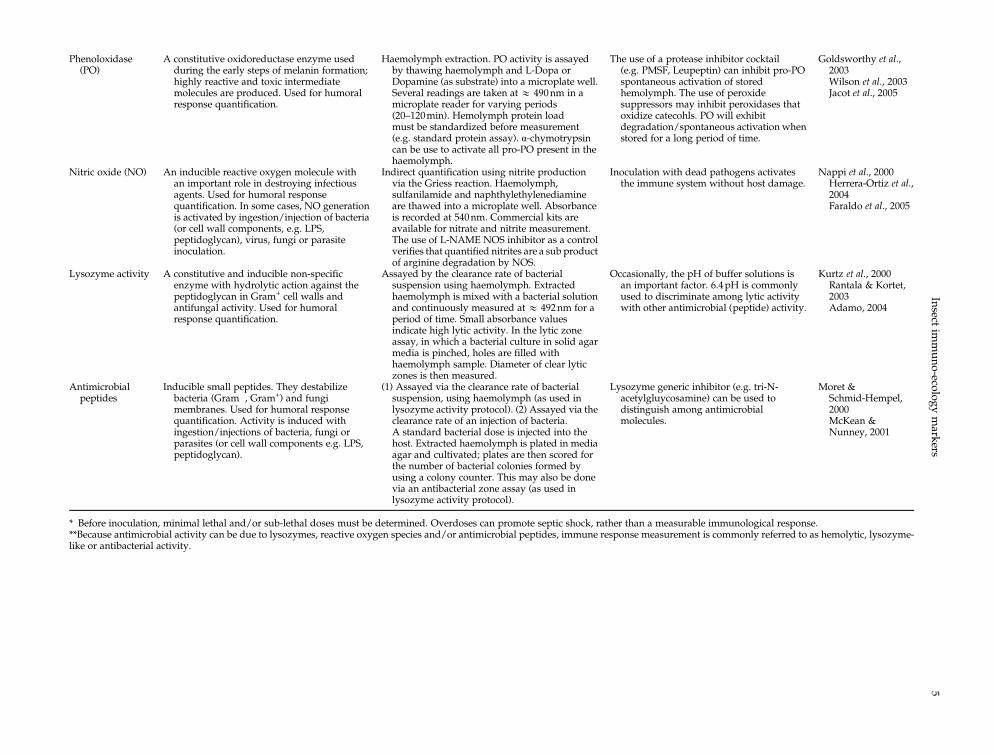
Phenoloxidase(PO)
A constitutive oxidoreductase enzyme usedduring the early steps of melanin formation;highly reactive and toxic intermediatemolecules are produced. Used for humoralresponse quantification.
Haemolymph extraction. PO activity is assayedby thawing haemolymph and L-Dopa orDopamine (as substrate) into a microplate well.Several readings are taken at & 490nm in amicroplate reader for varying periods(20–120min). Hemolymph protein loadmust be standardized before measurement(e.g. standard protein assay). α-chymotrypsincan be use to activate all pro-PO present in thehaemolymph.
The use of a protease inhibitor cocktail(e.g. PMSF, Leupeptin) can inhibit pro-POspontaneous activation of storedhemolymph. The use of peroxidesuppressors may inhibit peroxidases thatoxidize catecohls. PO will exhibitdegradation/spontaneous activation whenstored for a long period of time.
Goldsworthy et al.,2003Wilson et al., 2003Jacot et al., 2005
Nitric oxide (NO) An inducible reactive oxygen molecule withan important role in destroying infectiousagents. Used for humoral responsequantification. In some cases, NO generationis activated by ingestion/injection of bacteria(or cell wall components, e.g. LPS,peptidoglycan), virus, fungi or parasiteinoculation.
Indirect quantification using nitrite productionvia the Griess reaction. Haemolymph,sulfanilamide and naphthylethylenediamineare thawed into a microplate well. Absorbanceis recorded at 540nm. Commercial kits areavailable for nitrate and nitrite measurement.The use of L-NAME NOS inhibitor as a controlverifies that quantified nitrites are a sub productof arginine degradation by NOS.
Inoculation with dead pathogens activatesthe immune system without host damage.
Nappi et al., 2000Herrera-Ortiz et al.,2004Faraldo et al., 2005
Lysozyme activity A constitutive and inducible non-specificenzyme with hydrolytic action against thepeptidoglycan in Gram+ cell walls andantifungal activity. Used for humoralresponse quantification.
Assayed by the clearance rate of bacterialsuspension using haemolymph. Extractedhaemolymph is mixed with a bacterial solutionand continuously measured at & 492nm for aperiod of time. Small absorbance valuesindicate high lytic activity. In the lytic zoneassay, in which a bacterial culture in solid agarmedia is pinched, holes are filled withhaemolymph sample. Diameter of clear lyticzones is then measured.
Occasionally, the pH of buffer solutions isan important factor. 6.4pH is commonlyused to discriminate among lytic activitywith other antimicrobial (peptide) activity.
Kurtz et al., 2000Rantala & Kortet,2003Adamo, 2004
Antimicrobialpeptides
Inducible small peptides. They destabilizebacteria (Gram�, Gram+) and fungimembranes. Used for humoral responsequantification. Activity is induced withingestion/injections of bacteria, fungi orparasites (or cell wall components e.g. LPS,peptidoglycan).
(1) Assayed via the clearance rate of bacterialsuspension, using haemolymph (as used inlysozyme activity protocol). (2) Assayed via theclearance rate of an injection of bacteria.A standard bacterial dose is injected into thehost. Extracted haemolymph is plated in mediaagar and cultivated; plates are then scored forthe number of bacterial colonies formed byusing a colony counter. This may also be donevia an antibacterial zone assay (as used inlysozyme activity protocol).
Lysozyme generic inhibitor (e.g. tri-N-acetylgluycosamine) can be used todistinguish among antimicrobialmolecules.
Moret &Schmid-Hempel,2000McKean &Nunney, 2001
* Before inoculation, minimal lethal and/or sub-lethal doses must be determined. Overdoses can promote septic shock, rather than a measurable immunological response.**Because antimicrobial activity can be due to lysozymes, reactive oxygen species and/or antimicrobial peptides, immune response measurement is commonly referred to as hemolytic, lysozyme-like or antibacterial activity.
Insectim
muno-ecology
markers
5

activities and fitness traits. Furthermore, it is also probable thathaemocytes undergo apoptosis after phagocytosis, whichreduces the number of circulating haemocytes (as occurs inAe. aegypti: Hillyer et al., 2005). This would lead to anothertrade-off, as it would generate a decrease in the homeostasisand remodelling of tissues, as well as the capacity for fightingpathogens. Phagocytosis is an important mechanism to re-move dead or damaged cells, as well as to remodel organsduring metamorphosis (Abrams et al., 1993; Franc et al., 1996).Thus, a delay in growth seems necessary for pathogen clear-ing, via releasing haemocytes from their immune activity andfacilitating the wound repairing process before cuticle shed-ding (Lavine & Strand, 2002).
Methodology
Phagocytic activity has been measured by exposing thehost to microbial pathogens, such as bacteria and yeast cells orpolystyrene beads, and assessing in vitro phagocytosis (Kurtz& Sauer, 1999; Kurtz et al., 2000; Kurtz, 2002). One way toassess this process is by inoculating fluorescent, labelled beadsor bacteria in vivo (Kurtz, 2002). In either process, the phago-cytised intruders are counted manually or by using imageanalysis software. Care must be taken to ensure the beads donot become obstructed by melanin. Recovery of remainingbeads (if possible) from the haemolymph is a useful method toestimate phagocytosis activity. Trypan blue also can furtherassess the viability of labelled cells or molecules. The effect ofthis colorant is that only the ingested particles retain theirfluorescence, while non-ingested particles become coatedwithtrypan blue dye (see Kurtz et al., 2000). Another method isto use pHrodo dye conjugated bacteria. Bacteria becomefluorescent only in the acidic environment of the haemocytephagosome (see Luce-Fedrow et al., 2009), while extracellularnon-phagocytosed bacteria do not fluoresce. When using livebacteria to evaluate the efficiency of phagocytosis, or any ofthe immune parameters referred to in this paper, we rec-ommend first to assess the bacterial clearance of the infectedhost. To do this, supernatant of haemolymph (or homogen-ized/macerated individuals) can be plated (e.g. LB-agar,blood-agar) and colony forming units counted (see Phamet al., 2007).
Capacity to attach to non-self surfaces(nodule/capsule formation)
Quite frequently, researchers have introduced pathogensor microbial cell wall components to insect hosts to assesscellular immune response (e.g. Howard et al., 1998). Thisinduces the production of nodules/capsules, which are thencounted when haemolymph is extracted after the challenge ordirectly in the haemocoel (e.g. Miller et al., 1996; Goldsworthyet al., 2003). Nodules can persist in insects for long periods oftime (Carton & Nappi, 1997), which means that the haemo-cytes used for nodule formation remain attached, therebydecreasing haemocyte density during insect lifetime. Thisreduction in haemocyte number could in turn limit the avail-ability of haemocytes that become relevant during moultingand/or metamorphosis. Not all adult insects produce haemo-cytes since haematopoietic organs are not always present atthis stage (Chapman, 1998). In lepidopterans and dipterans,hematopoietic organs have been identified exclusively in theembryonic, larval and pupal stages (Gardiner & Strand, 2000;Holz et al., 2003; Nardi et al., 2003; Williams, 2007). This means
that any haemocytes present in the adult stage were producedduring embryogenesis, larval stages and metamorphosis.However, the presence of hematopoietic organs in adultgrasshoppers has been reported (Hoffman, 1973; Hoffmanet al., 1974). Natural history differences among insect groups(e.g. the presence of metamorphosis) likely leads to variationin haemocyte production (consequently cell number).
Methodology
Pathogens such as bacteria or yeast cells are inoculateddirectly to haemocoel (Brookman et al., 1989). Also, injectedparasites can be used to assess nodule/encapsulation for-mation (see Eleftherianos et al., 2008). Haemolymph iscollected and incubated in a humidity chamber; nodules areobserved using amicroscope and counted. Caremust be takensince pathogens can be destroyed before nodulation takesplace, for example, if pathogens enter the midgut (Luckhartet al., 1998). Furthermore, nodules must be counted evenwhentheir size varies, which can occur even within the sameindividual (see Howard et al., 1998). Enumeration of differentsizes can provide further information on differential immuneability among hosts.We recommenddirectly assessing the sizeof nodules either qualitatively (by categorizing the differentsizes and counting how many of these fall into the distinctcategories) or quantitatively with the use of image analysissoftware.
Melanotic nodulation/encapsulation (nylon monofilamentand beads insertion)
For encapsulation/melanotic encapsulation assessment,inoculationwith sephadex or polystyrene beads or insertion ofa nylon monofilament into the haemocoel is frequently used.In this method, nylon and beads are thought to mimic naturalinfection (like that of a parasite/parasitoid) while avoidingthe damaging effects of pathogens (e.g. Siva-Jothy, 2000;Koskimäki et al., 2004; Simmons et al., 2005; Rantala & Roff,2007). Cellular and humoral responses are activated, whichcould lead to trade-offs similar to when nodule formation andphagocytosis are elicited. In the melanin layer, which isfrequently produced during nodulation/encapsulation, syn-thesis of melanin is required. Melanin is costly to produce, notonly because of the resources used (see Phenoloxidase Activitybelow), but because melanin is required for cuticle formation,humoral toxic molecule production, and colouring andhardening of other structural traits (e.g. wings, eggs, sper-matheca: Li & Christensen, 1993; Ilango, 2005).
Methodology
After insertion, filament/beads are retrieved and thevolume of melanized and non-melanized cell masses thatencapsulate the filament are measured (Siva-Jothy, 2000).In vitro encapsulation assays can be also be performed. Forthis, haemolymph is collected from each individual andmixedwith beads. Encapsulation and melanization are observedafter incubation by microscopy. Three methods have beendeveloped for measuring such masses. One method observesthe mean gray scale of the covered area using image analysissoftware (Simmons et al., 2005), while a second uses imagessoftware filters to measure the density of the covered area(Koskimäki et al., 2004). Another approach measures the en-capsulated/melanized area (Contreras-Garduño et al., 2006).
M. Moreno-García et al.6

All measures, however, are unable to differentiate the func-tional aspects of haemocytes, as cellular nodulation/encapsu-lation and humoral melanization are two separate but relatedprocesses (Ling & Yu, 2006). Haemocytes release or containsurface proPO that, when activated to PO, causes melaniza-tion. This cellular encapsulation also enhances interactionsbetween haemolymph and PO. Ling & Yu (2006) noticed thatmelanization did not occur when isolated haemocytes wereused. Rather, melanization occurred only in the presence ofplasma. This suggests that factors in haemolymph are requiredfor PO cascade activation and, hence, melanization. Therefore,cellular encapsulation could be implicated in both cellular andhumoral immune responses. A possible adverse effect ofmelanization is the production of toxic molecules that couldaffect self tissues or cells. Also, melanotic capsules consists ofan inner area of necrotic hemocytes with melanin formation atthe interface with the foreign object (Hernández-Martínezet al., 2002); this loss of haemocytes could be detrimental forthe host.
Trade-offs among immunity and other traits (e.g. devel-opmental time) using implants have been found (see Rantala& Rolff, 2005). The use of non-natural infections seems to be asuitable index of host immune activation-recognition and itsecological costs (e.g. Rantala & Roff, 2007). For the methodsdiscussed above, unfortunately, the distribution of mela-nized/encapsulated areas is often so irregular that it impedesthorough assessment (i.e. it depends on the position of theartificial object during area measurement). We recommendseveral measures of the same implant. Additionally, insertinga nylon piece is a difficult task for small insects. In this case, werecommend the use of inoculated beads. We also recommendusing live pathogens that induce melanotic encapsulation orother immune responses. This can be useful, for example,when testing for immune tolerance and behaviour (see Ayres& Schneider, 2009). Tolerance theory predicts a possibleincrease in the pathogen burden, but the immune responsewill limit the health or fitness consequences of this pathogenload (sensu Read et al., 2008; Schneider & Ayres, 2008). Onlylive pathogens can replicate and continuously damage a host.It is also possible that live pathogens activate more immunepathways (compared with inert implants) and, therefore, givea more complete picture of an immune response. However,when using live pathogens, it is important to first estimatethe dosage of pathogen needed for accurate measurement ofa particular immune response. It has been observed that insome insects, low to medium doses elicit phagocytosis; mod-erate to high doses elicit nodulation; and overdoses canpromote septic shock (S. Hernández-Martínez, personalcommunication). Before a challenge, we recommend discover-ing the immune response that is elicited in relation to the typeand dose of infectious agent (natural or artificial) model.
Humoral responses
Phenoloxidase activity
PO is the most widely used immune marker in ecologicalimmunology studies (e.g. Goldsworthy et al., 2003; Wilsonet al., 2003; Jacot et al., 2005). The PO enzyme is used for differ-ent physiological processes (wounding, clotting, cuticle com-position, melanotic encapsulation, production of cytotoxicmolecules) and probably spermatheca formation (Ilango,2005). Due to these different functions, PO is constantlysynthesized; therefore, it could be costly to produce for the
organism (see González-Santoyo & Córdoba-Aguilar, 2012).Recall that proPO molecules are produced mainly by haemo-cytes (other tissues, such as fat body and midgut epithelium,may also produce proPO in smaller quantities), and that someof thesemolecules remain attached to haemocytes, while someproPO might end up in several other tissues (i.e. cuticle,midgut, salivary glands, eggs: Ashida & Brey, 1997). If patho-gens are detected during proPO transportation from haemo-cytes to other tissues, it is possible that this proPO getsactivated in the haemocele for clearing pathogens, conse-quently leaving target tissues with a proPO deficiency. Thismeans that a trade-off may arise because of a proPO deficiency(but not of other components, such as PO substrates; seebelow), as has been assumed in general studies where PO andmelanin have been involved (Siva-Jothy, 2000). This differentperspective of a possible trade-off when measuring POactivity has not been put forward nor examined previouslyand needs further empirical study.
The PO substrate tyrosine (TYR) and tyrosine subproductsare used for the expression and manufacture of other traits.TYR is obtained through food and is naturally synthesized byorganisms or by the hydroxylation of phenylalanine (PHE)(Christensen et al., 2005). Since PHE is an essential aminoacid for protein synthesis, its importance lies in the cost ofacquiring this resource. TYR is also involved in a variety offunctions. One of these, for example, is the colouring of the eggchorion in some insects (Li & Christensen, 1993). In mosqui-toes, melanization responses against worms generate a delayin TYR accumulation in the ovaries and, therefore, a decreasein the number of eggs produced after an immune challenge(Li & Christensen, 1993) and a delay in oviposition (Ferdiget al., 1993). Examples like this are suggestive of substrates ofPO and melanin – PHE and TYR – being restrictive resourcesthat may lead to trade-offs between the immune response andother key functions, including moulting, basal metabolic ratesand protein synthesis. Although TYR can be stored (astyrosine-O-phosphate, beta-alanyl-L-tyrosine, tyrosine gluco-side: Mitchell & Lunan, 1964; Levenbook et al., 1969; Chenet al., 1978), trade-offs could be detected if not enough TYR isgathered in order to provide this amino acid to all the meta-bolic requirements through the insect lifetime. However, notall insects necessarily use TYR or PHE in the same way, POmeasures need to be carefully interpreted in every insectmodel. Melanin, the final product of the PO cascade, is alsoinvolved in other morphological traits, such as cuticle for-mation and pigmentation, which may affect functions relatedto aposematism and crypsis, thermoregulation, resistance toUV radiation and coloured sexual traits. Many of thesefunctions and their supposed trade-offs have not beenadequately investigated.
Methodology
PO activity is assayed by thawing haemolymph ando-diphenols (dopa or dopamine) as substrate into a microplatewell. This assay will not distinguish the three different POactivities, since it is inferred that the ο-diphenoloxidaseactivity of all three enzymes will be detected (Sugumaran &Kanost, 1993). Care must be taken when using o-diphenols,since there could also be a peroxidase-mediated melaninformation (Christensen et al., 2005), which can result inan overestimation of PO activity. We recommend the useof a peroxidase suppressor (e.g. hydrogen peroxide inmethanol or commercially available). Another current
Insect immuno-ecology markers 7

problem with assessing PO is that protein load standardiz-ation is needed before measurement, using a standard proteinassay (Contreras-Garduño et al., 2007), a practice that is rarelycarried out and without which there may result disparate andunreliable PO readings. Since, occasionally, real protein loadcan be masked if there are differences among individuals (e.g.bigger size of females than males, inaccurate extract of totalamount of haemolymph). We recommend using the proteinload practice mentioned above. Furthermore, there might be arapid degradation of PO if stored for a relatively long time.This issue can be resolved by using a protease inhibitorcocktail (e.g. PMSF, Leupeptin). Another problem is thatsmall-sized insects may not provide PO readings, since POquantities are so small. This problem can be solved by usingmore than one individual for a single sample to obtain POreadings. Also, the total PO activity can be estimated using theenzyme α-chymotrypsin to activate all pro-PO present in thehaemolymph (see Bailey & Zuk, 2010).
Nitric oxide
NO quantification is a relatively new immune marker inecological immunity studies (see, for example, Moreno-Garcíaet al., 2010). NO damages pathogens and is also used as asignalling molecule. In Drosophila, NO is indispensable foractivation of the immune deficiency (Imd) pathway (Foley &O’Farrel, 2003). This is one of the three pathways (the othertwo being Toll and JAK/STAT; see Ferrandon et al., 2007) thatproduce transcriptional factors in the fat body, leading to thesynthesis of antimicrobial peptides and other factors thatprevent self tissue damage and control haemocyte prolifer-ation and differentiation (Foley & O’Farrell, 2003; Agaisse &Perrimon, 2004). Another interesting point is the fact that theNO synthase (NOS) (an enzyme that converts L-Arginine toL-Citrulline and generates NO) is constitutive and inducible(Müller, 1997). NO is also a signalling molecule, so a constantproduction of NOS is necessary for homeostatic purposes,while inducible NOS is only synthesized after an immunechallenge (Nappi et al., 2000). It is, therefore, expected thatNO production increases after an immune challenge.Nevertheless, Krishnana et al. (2006) found that NO pro-duction was similar in both non-stimulated and stimulatedhaemocytes in lepidopteran larvae. In this particular case, theauthors indicate that NO can be continuously synthesized bythe induced NOS for long periods of time (hours to days). Thismeans that the effectiveness for killing pathogens with NOmay not only reside on NO concentration but in the durationof NO synthesis (see Laurent et al., 1996).
NO is produced during the oxidation of L-Arginine(Müller, 1997). Arginine is an amino acid, which must beobtained from the diet (Rivero, 2006). Arginine is also im-portant for sperm maturation (Osanai & Chen, 1993), eggproduction (Uchida, 1993), long term memory, chemosensory(antennal lobes, olfaction) and visual information processing(Müller, 1997). Trade-offs are thus expected among these traitsand the immune response.
Methodology
Basal levels of NO are normally used, which provides anincomplete picture of host immune ability. A better alternativeis to assess NO production after host injection or ingestion ofpathogens. The standard method to measure NO is to use theGriess reaction that measures nitrite production, an indirect
measure of NO (Bredt & Snyder, 1994). The use of controlssuch as an L-NAME (a NOS-inhibitory arginine analogue; seeRivero, 2006) and its inactive enantiomer D-NAME isrecommended. Controls are needed because nitrites can alsobe sub-products of other reactions not related with immunity.For this method, however, it is unknown whether haemo-lymph protein load standardization is needed before measure-ment. This is a technical problem that must be resolved in thenear future. Similar to PO, more than a single individual maybe needed if an individual host sample is too small.
Lysozyme activity and antimicrobial peptides
Lysozyme is an enzyme with hydrolytic action, mainlyagainst the peptidoglycan of Gram+ cell walls. It can be in-duced or constitutively expressed in the gut tract (Hetru et al.,1997), haemocytes and fat body (Gillispie et al., 1997). AMPsshow great structural diversity (more than 170 isoforms havebeen found in insects: Bulet et al., 1999) and are produced soonafter foreign recognition (1–4h) with an efficient pathogen-specific killing action (Schmid-Hempel, 2005b). Some AMPsmay remain in haemolymph for up to three weeks (Schmid-Hempel, 2005b), which is convenient during subsequentpathogen encounters. AMPs are generally short peptides,containing fewer than 150–200 amino acids (Bulet et al., 2004),so they are considered energy efficient, quick and economicalto produce (Otvos Jr, 2000). However, the overall AMPsconcentration in Drosophila haemolymph can reach 200μM(Otvos Jr, 2000). At this concentration, antimicrobial peptidesdo seem costly to produce. It is possible that resources(proteins) gathered through ontogeny are indispensable forantimicrobial peptide generation as well as for the synthesis ofother molecules. This fact could potentially lead to trade-offsbetween immunity and synthesis of other products (e.g.spermatophalyx peptides) and tissue conformation.
Methodology
Lysozyme activity is commonly assayed via the clearancerate of bacterial suspension using haemolymph. The methodis relatively simple; after the haemolymph is extracted andmixed with a bacterial solution, a turbidity assay is then per-formed (e.g. Adamo, 2004). Small absorbance values indicatehigh lytic activity. Another way is to add small drops ofhaemolymph to bacterial culture, measuring the area of lyticactivity after some time (12h). The lytic activity appears asclear circular zones in the culture, which indicates that bacteriahave been cleared. The diameter of the hole is measured andtaken as the indicator of lytic effectiveness.
AMPs have been measured by injection or ingestionof complete bacteria, fungi, parasites or their fractions (e.g.LPS, peptidoglycane). AMP activity is assessed in three ways:(i) measurement of the clearance rate of a bacterial suspension(via a turbidity assay, similar to the lysozyme protocol de-scribed above); (ii) measurement of the rate of bacterial growthon a plate, after the same bacteria have been injected into thehost and the haemolymph has been extracted after some time;and (iii) the antibacterial zone assay, via measurement of thehole area, similar to the lysozyme protocol described above.A problem with these different assays is that antimicrobialactivity can be due to other molecules and not necessarilyAMPs. Since lysozyme needs a pH 8 to 6.5 for optimal activity,this can be used to distinguish between other AMP activities
M. Moreno-García et al.8

(Rantala & Kortet, 2004; Ahtiainen et al., 2005; but see da Silvaet al., 2000; Adamo & Parsons, 2006).
Some concluding remarks
One intention of this review is to highlight the ecologicaland physiological context of insect immune markers and theirvalue for estimating trade-offs between immunity and othertraits. Thus, current (and new) knowledge of immune phy-siological mechanisms must be incorporated into ecologicaldata to allow for a deeper understanding of the reasonsfor evolutionary correlations and constraints. The use ofeach method mentioned here must be determined by eachresearcher in terms of its overall cost (in terms of time-consumption or price), differences in the kind of pathogens orelicitors used to induce immune response, and the immunemarker that can address questions via an appropriate experi-mental design. Another important consideration for eachresearcher is the potential to make repeated immunemeasurements on the same sample. Unfortunately, some ofthe measurement methods referred to here are not suitableto be repeated more than once. For example, PO and NOcontained in haemolymph extraction gradually degrade evenwhen stored in an ultrafreezer (–70°C). Also, unfreezing andre-freezing samples accelerates degradation. Occasionally,haemocyte membranes can become disrupted even in a buffersolution or culture medium if stored for long periods of time(e.g. weeks to months). We recommend extracting enoughsamples to do all measurements required for the experimentaldesign. If a large number of individuals are required, they(or extractions) can be stored and measurements should onlybe done for the exact number of tests that can be processed andquantified in a single day.
Finally, there are two related points that we would like tohighlight: (i) some immune markers mentioned here areusually seen as responses to attack and kill pathogens; and(ii) the existing correlations among immune responses. Asmentioned before, negative and positive correlations havebeen found among immune traits, but correlations seem to behost-specific. Commonly, resource limitation has been used toexplain the negative correlations; however, the non-expressionof an immune effector does not necessarily mean that there isno response. Reactions and products of immune responses areinterconnected; and the type of response is related to thepathogens virulence. As mentioned before, not all pathogenswill induce the same reaction; or, at least, the magnitude of aresponse can differ depending on the host-pathogen inter-action type. Occasionally, tolerance could be the best strategyto cope with infections. The resistance or tolerance strategiesadopted by an individual will depend on the force and courseof the infection. There could be a critical host damage thre-shold; thus, when the intensity of infection is below thisthreshold (i.e. no significant damage to the host), the insectcould tolerate it. Above the threshold, it then pays to allocatemore resources into resisting the pathogen. For example, pha-gocytic activity could be turned on, while PO activity couldremain down regulated; in this case, probably the host isavoiding autoreactivity molecules (generated through the POcascade). In this example, the immune strategy is enoughin order to limit pathogen and self damage; nevertheless,the pathogen is not eliminated. Since the immune system ishighly complex, several aspects of immune response must bemeasured to comprehend the insect immunity strategies. Also,occasionally, some immune responses are not suitable to test
some hypothesis; for example, pro-PO cascade and PO activityoccasionally are not affected by the nutritional condition, butother responses, such as lysozyme, are affected (see Jacot et al.,2005; Moreno-García et al., 2010). Therefore, it is rec-ommended to use more than one immune marker whenpossible. Finally, the starting point in insect immunoecology isthat it must incorporate the great range of environmental fac-tors involved using a trade-off framework. Resource allocationand investment strategies should be studied by taking intoaccount evidence of the real cost in natural conditions andtheir effects in the expression of immune response andgenerational changes.
Ideally, an ecological study of insect immunity shouldstart with thoughtful design regarding appropriate immunemarkers. Regarding methodological issues, in many cases,methodological information is available, but it is possible thatthe status quo of how other evolutionary ecology researchershave used such markers leads others not to think twice aboutthe best way to proceed. Lastly, we encourage ecological andevolutionary biologists alike to incorporate useful moleculartechniques (e.g. microassays, proteomics, quantitative ReverseTranscriptase-PCR, etc.) to identify the possible mechanismsand the genetic basis underlying the trade-offs betweenimmunity and life history traits. It is hoped that this infor-mation proves constructive in paving theway to amore robustfield of ecological immunology.
Acknowledgements
We thank Salvador Hernández-Martínez for providingassistance with protocols and for helpful suggestions. Wethank Gloria Tavera and Chris Anderson for revising thispaper in English, and providing thoughtful comments. MM-Gwas supported by the Posgrado en Ciencias Biomédicas(CONACYT: grant no. 172947), Instituto de Ecología, (Univer-sidad Nacional Autónoma de México), and the Centro deInvestigación Sobre Enfermedades Infecciosas, InstitutoNacional de Salud Pública, México. AC-A was supportedby a grant provided by PAPIIT-UNAM (Project No. 211506).An anonymous reviewer provided corrections to the manu-script.
References
Abrams, J.M., White, K., Fessler, L.I. & Steller, H. (1993)Programmed cell death during Drosophila embryogenesis.Development 117, 29–43.
Adamo, S.A. (1999) Evidence for adaptive changes in egg laying incrickets exposed to bacteria and parasites. Animal Behaviour57, 117–124.
Adamo, S.A. (2004) Estimating disease resistance in insects:phenoloxidase and lysozyme-like activity and disease re-sistance in the cricket Gryllus texensis. Journal of InsectPhysiology 50, 209–216.
Adamo, S.A. & Parsons, N.M. (2006) The emergency life-historystage and immunity in the cricket, Gryllus texensis. AnimalBehaviour 72, 235–244.
Agaisse, H. & Perrimon, N. (2004) The roles of JAK/STATsignaling in Drosophila immune responses. ImmunologicalReviews 198, 77–82.
Ahtiainen, J.J., Alatalo, R.V., Kortet, R. & Rantala, M.J. (2005) Atrade-off between sexual signalling and immune function in anatural population of the drumming wolf spiderHygrolycosarubrofasciata. Journal of Evolutionary Biology 18, 985–991.
Insect immuno-ecology markers 9

Andersen, S.O., Hojrup, P. & Roepstorff, P. (1995) Insectcuticular proteins. Insect Biochemistry and Molecular Biology25, 153–176.
Armitage, S.A.O. & Siva-Jothy, M.T. (2005) Immune functionresponds to selection for cuticular colour in Tenebrio molitor.Heredity 94, 650–656.
Ashida, M. & Brey, P.T. (1997) Recent advances in research on theinsect prophenoloxidase cascade. pp. 135–172 in Brey, P.T. &Hultmark, D. (Eds)MolecularMechanisms of Immune Responsesin Insects. New York, USA, Chapman & Hall.
Ayres, J.S. & Schneider, D.S. (2009) The role of anorexia in re-sistance and tolerance to infections inDrosophila. PLoS Biology7, e1000150.
Bailey, N.W. & Zuk, M. (2010) Changes in immune effort of malefield crickets infestedwithmobile parasitoid larvae. Journal ofInsect Physiology 54, 96–104.
Barnard, C.J., Collins, S.A., Daisley, J.N. & Behnke, J.M. (2006)Odour learning and immunity costs in mice. BehaviouralProcesses 72, 74–83.
Barnes, A.I. & Siva-Jothy, M.T. (2000) Density-dependent pro-phylaxis in the mealworm beetle Tenebrio molitorL. (Coleoptera: Tenebrionidae): cuticular melanization is anindicator of investment in immunity. Proceedings of the RoyalSociety of London, Series B 267, 177–182.
Bocher, A., Doums, C., Millot, L. & Tirard, C. (2008)Reproductive conflicts affect labor and immune defense inthe queenless ant diacam sp ‘Nilgiri’. Evolution 62, 123–134.
Boots, M. & Begon, M. (1993) Trade-offs with resistance to agranulosis virus in the indian meal moth, examined by a lab-oratory evolution experiment. Functional Ecology 7, 528–534.
Bredt, D.S. & Snyder, S.H. (1994) Nitric oxide: a physiologicalmessenger molecule. Annual Review of Biochemistry 63, 175–195.
Brookman, J.L., Rowley, A.F. & Ratcliffe, N.A. (1989) Studies onnodule formation in locusts following injection of microbialproducts. Journal of Invertebrate Pathology 53, 315–323.
Brzek, P. & Konarzewski, M. (2007) Relationship between aviangrowth rate and immune response depends on food avail-ability. Journal of Experimental Biology 210, 2361–2367.
Bulet, P., Charlet, M. & Hetru, C. (2003) Antimicrobial peptidesin insect immunity. pp. 89–108 in Ezekowitz, R.A.B. &Hoffmann, J.A. (Eds) Innate Immunity. New Jersey, USA,Humana Press.
Bulet, P., Stöcklin, R. &Menin, L. (2004) Anti-microbial peptides:from invertebrates to vertebrates. Immunological Review 198,169–184.
Bulet, P., Hetru, C., Dimarq, J.L. & Hoffmann, D. (1999)Antimicrobial peptides in insects: structure and function.Developmental and Comparative Immunology, 23, 329–344.
Calleri, D.V., Rosengars, R.B. & Traniello, J.F.A. (2007)Immunity and reproduction during colony foundation in thedampwood termite, Zootermopsis angusticollis. PhysiologicalEntomology 32, 136–142.
Carton, Y. & Nappi, A.J. (1997) Drosophila cellular immunityagainst parasitoids. Parasitology Today 13, 218–227.
Casteels, P. (1998) Immune response in Hymenoptera. pp. 92–110in Brey, P.T. & Hultmark, D. (Eds) Molecular Mechanisms ofImmune Responses in Insects. New York, USA, Chapman &Hall.
Cerenius, L. & Söderhäll, K. (2004) The prophenoloxidase.activating system in invertebrates. Immunological Review 198,116–126.
Chapman, R.F. (1998) The Insects: Structure and Function.Cambridge, UK, Cambridge University Press.
Chen, P.S., Mitchell, H.K. & Neuweg, M. (1978) Tyrosineglucoside in Drosophila busckii. Insect Biochemistry 8, 279–286.
Christensen, B.M., Li, J., Chen Ch.-Ch. & Nappi, A.J. (2005)Melanization immune responses in mosquito vectors. Trendsin Parasitology 21, 192–199.
Contreras-Garduño, J., Canales-Lazcano, J. & Córdoba-Aguilar, A. (2006) Wing pigmentation, immune ability, fatreserves and territorial status in males of the rubyspotdamselfly,Hetaerina americana. Journal of Ethology 24, 165–173.
Contreras-Garduño, J., Lanz-Mendoza, H. & Córdoba-Aguilar, A. (2007) The expression of a sexually selected traitcorrelates with different immune defense components andsurvival in males of the American rubyspot. Journal of InsectPhysiology 53, 612–621.
Cotter, S.C., Hails, R.S., Cory, J.S. & Wilson, K. (2004) Density-dependent prophylaxis and condition-dependent immunefunction in Lepidopteran larvae: a multivariate approach.Journal of Animal Ecology 73, 283–293.
da Silva, C.C.A., Dunphy, G.B. & Rau, M.E. (2000) Interaction ofXenorhabdus nematophilus (Enterobacteriaceae) with theAntimicrobial Defenses of the House Cricket, Acheta domes-ticus. Journal of Invertebrate Pathology 76, 285–292.
Darwin, C. (1871) TheDescent ofMan and Selection in Relation to Sex.London, UK, John Murray.
Day, T., Graham, A.L. & Read, A.F. (2007) Evolution of parasitevirulence when host responses cause disease. Proceedings ofthe Royal Society of London, Series B 274, 2685–2692.
Decker, H. & Jaenicke, E. (2004) Recent findings on phenolox-idase activity and aintimicrobial activity of hemocyanins.Developmental and Comparative Immunology 28, 673–687.
Desjardins,M., Houde,M. &Gagnon, E. (2005) Phagocytosis: theconvoluted way from nutrition to adaptive immunity.Immunological Reviews 207, 158–165.
Duffy, M.A. & Sivars-Becker, L. (2007) Rapid evolutionand ecological host-parasite dynamics. Ecology Letters 10,44–53.
Eleftherianos, I., Baldwin, H., ffrench-Constant, R.H. &Reynolds, S.E. (2008) Developmental modulation of im-munity: Changes within the feeding period of the fifth larvalstage in the defence reactions ofManduca sexta to infection byPhotorhabdus. Journal of Insect Physiology 54, 309–318.
Falconer, D.S. & Mackay, T.F. (1996) Introduction to QuantitativeGenetics. New York, USA, Dover.
Faraldo, A.C., Sá-Nunes, A., Del Bel, E.A., Faccioli, L.H. &Lello, E. (2005) Nitric oxide production in blowfly hemo-lymph after yeast inoculation. Nitric Oxide 13, 240–246.
Fedorka, K.M., Zuk, M. & Mousseau, T.A. (2004) Immune su-pression and the cost of reproduction in the ground cricket,Allonemobius socius. Evolution 58, 2478–2485.
Fellowes, M.D.E., Kraaijeveld, A.R. & Godfray, H.C.J. (1998)Trade-off associated with selection for increased ability toresist parasitoid attack in Drosophila melanogaster. Proceedingsof the Royal Society of London, Series B 265, 1553–1558.
Ferdig,M., Beerntsen, B., Spray, F., Li, J. & Christensen, B. (1993)Reproductive costs associated with resistance in a mosquito-filarial worm system. American Journal of Tropical Medicine &Hygiene 49, 756–762.
Ferrandon, D., Imler, J.L., Hetru, C. & Hoffman, J.A. (2007) TheDrosophila systemic immune response: a sensing and signal-ling during bacterial and fungal infections. Nature Reviews inImmunology 7, 863–874.
Foley, E. & O’Farrell, P.H. (2003) Nitric oxide contributes to in-duction of innate immune responses to gram negative bac-teria in Drosophila. Genes & Development 17, 115–125.
M. Moreno-García et al.10

Franc, N.C., Dimarcq, J.-L., Lagueux, M., Hoffmann, J. &Ezekowitz, R.A.B. (1996) Croquemort, a novel Drosophilahemocyte/macrophage receptor that recognizes apoptoticcells. Immunity 4, 431–443.
Gardiner, E.M. & Strand, M.R. (2000) Hematopoiesis in larvalPseudoplusia includens and Spodoptera frugiperda. Archives ofInsect Biochemistry and Physiology 43, 147–164.
Gillespie, J.P., Kanost, M.R. & Trenzeck, T. (1997) Biologicalmediators of insect immunity. Annual Review of Entomology42, 611–643.
Goldsworthy, G., Mullen, L., Opoku-Ware, K. &Chandrakant, S. (2003) Interactions between the endocrineand immune systems in locusts. Physiological Entomology 28,54–61.
González-Santoyo, I. & Córdoba-Aguilar, A. (2012)Phenoloxidase: a key component of the insect immune sys-tem. Entomologia Experimentalis et Applicata 142, 1–16.
Gosline, J., Lillie, M., Carrington, E., Guerette, P., Ortlepp, C. &Savage, K. (2002) Elastic proteins: biological roles andmechanical properties. Philosophical Transactions of the RoyalSociety of London, Series B 357, 121–132.
Haas, F., Gorb, S. & Blickhan, R. (2000) The function of resilin inbeetle wings. Proceedings of the Royal Society of London, Series B267, 1375–1381.
Hernández-Martínez, S., Lanz, H., Rodrguez, M.H., González-Ceron, L. & Tsutsumi, V. (2002) Cellular-mediated reactionsto foreign organisms inoculated into the hemocoel ofAnopheles albimanus (Diptera: Culicidae). Journal of MedicalEntomology 39, 61–69.
Herrera-Ortiz, A., Lanz-Mendoza, H., Martínez-Barnetche, J.,Hernández-Martínez, S., Villarreal-Treviño, C., Aguilar-Marcelino, L. & Rodríguez, M.H. (2004) Plasmodium bergheiookinetes induces nitric oxide production in Anopheles pseu-dopunctipennis midguts cultured in vitro. Insect Biochemistryand Molecular Biology 34, 893–901.
Hetru, C., Hoffman, D. & Bulet, P. (1997) Antimicrobial peptidesfrom insects. pp. 40–66 in Brey, P.T. & Hultmark, D. (Eds)Molecular Mechanisms of Immune Responses in Insects.New York, USA, Chapman & Hall.
Hillyer, J.F., Schmidt, S.L. & Christensen, B.M. (2003) Rapidphagocytosis and melanization of bacteria and Plasmodiumsporozoites by hemocytes of the mosquito Aedes aegypti.Journal of Parasitology 89, 62–69.
Hillyer, J.F., Schmidt, S.L. & Christensen, B.M. (2004) The anti-bacterial innate immune response by the mosquito Aedesaegypti is mediated by hemocytes and independent ofGram type and pathogenicity. Microbes and Infection 6,448–459.
Hillyer, J.F., Schmidt, S.L., Fuchs, J.F., Boyle, J.P. &Christensen, B.M. (2005) Age-associated mortality in im-mune challenged mosquitoes (Aedes aegypti) correlateswith a decrease in haemocyte numbers. Cellular Microbiology7, 39–51.
Hoang, A. (2001) Immune response to parasitism reduces resist-ance of Drosophila melanogaster to desiccation and starvation.Evolution 55, 2352–2358.
Hoffmann, D., Brehelin, M. & Hoffmann, J.A. (1974)Modifications of the hemogram and of the hemocytopoietictissue of male adults of Locusta migratoria (Orthoptera) afterinjection of Bacillus thuringiensis. Journal of InvertebratePathology 24, 238–247.
Hoffmann, J.A. (1973) Blood-forming tissues in orthopteran in-sects: An analogue to vertebrate hemopoietic organs. Cellularand Molecular Life Sciences 29, 50–51.
Hoffmann, J.A. & Reichhart, J.M. (2002) Drosophila innate im-munity: an evolutionary perspective. Nature Immunology 3,121–126.
Hogg, J.C. & Hurd, H. (1995) Malaria-induced reduction offecundity during the first gonotrophic cycle of Anophelesstephensi mosquitoes. Medical and Veterinary Entomology 9,176–180.
Hoi-Leitner, M., Romero-Pujante, M., Hoy, H. & Pavlova, A.(2001) Food availability and immune capacity in serin(Serinus serinus) nestlings. Behavioral Ecology and Sociobiology49, 333–339.
Holz, A., Bossinger, B., Strasser, T., Janning, W. & Klapper, R.(2003) The two origins of hemocytes in Drosophila.Development 130, 4955–4962.
Howard, R.W., Miller, J.S. & Stanley, D.W. (1998) The influenceof bacterial species and intensity of infections on noduleformation in insects. Journal of Insect Physiology 44, 157–164.
Ilango, K. (2005) Structure and function of the spermathecalcomplex in the phlebotomine sandfly Phlebotomus papatasiScopoli (Diptera: Psychodidae): I. ultrastructure and his-tology. Journal of Bioscience 30, 101–131.
Jacot, A., Scheuber, H., Kurtz, J. & Brinkhof, M.W.G.(2005) Juvenile immune system activation induces a costlyupregulation of adult immunity in field crickets Grylluscampestris. Proceedings of the Royal Society of London, Series B272, 63–69.
Jollès, P. (1996) Lysozymes: Model Enzymes in Biochemistry andBiology. Switzerland, Birkhäuser Verlag.
Koskimaki, J., Rantala, M.J., Taskinen, J., Tynkkynen, K. &Suhonen, J. (2004) Immunocompetence and resource hold-ing potential in the damselfly, Calopteryx virgo L. BehavioralEcology 15, 169–173.
Kraaijeveld, A.R. & Godfray, H.C.J. (1997) Trade-off betweenparasitoid resistance and larval competitive ability inDrosophila melanogaster. Nature 389, 278–280.
Kraaijeveld, A.R., Limentani, E.C. &Godfray, H.C.J. (2001) Basisof the trade-off between parasitoid resistance and larvalcompetitive ability in Drosophila melanogaster. Proceedings ofthe Royal Society of London, Series B 268, 259–261.
Krishnana, N., Hyršl, P. & Šimek, V. (2006) Nitric oxide pro-duction by hemocytes of larva and pharate prepupa ofGalleria mellonella in response to bacterial lipopolysaccharide:Cytoprotective or cytotoxic? Comparative Biochemistry andPhysiology, Part C 142, 103–110.
Kumar, S., Christophides, G.K., Cantera, R., Charles, B., SooHan, Y., Meister, S., Dimopoulos, G., Kafatos, F.C. &Barillas-Mury, C. (2003) The role of reactive oxygen specieson Plasmodiummelanotic encapsulation in Anopheles gambiae.Proceedings of the National Academy of Science USA 100,14139–14144.
Kurtz, J. (2002) Phagocytosis by invertebrate hemocytes: Causes ofindividual variation in Panorpa vulgaris scorpionflies.Microscopy Research and Technique 57, 456–468.
Kurtz, J. & Sauer, K.P. (1999) The immunocompetence handicaphypothesis: testing the genetic predictions. Proceedings of theRoyal Society of London, Series B 266, 2515–2522.
Kurtz, J., Wiesner, A., Götz, P. & Sauer, K.P. (2000) Genderdifferences and individual variation in the immune systemof the scorpionfly Panorpa vulgaris (Insecta: Mecoptera).Developmental and Comparative Immunology 24, 1–12.
Lackie, A.M. (1988) Hemocyte behaviour. Advances in InsectPhysiology 21, 85–178.
Laurent,M., Lepoivre,M.&Tenu, J.P. (1996) Kineticmodelling ofthe nitric oxide gradient generated in vitro by adherent cells
Insect immuno-ecology markers 11

expressing inducible nitric oxide synthase. BiochemistryJournal 314, 109–113.
Lavine, M.D. & Strand, M.R. (2002) Insect hemocytes and theirrole in immunity. Insect Biochemistry and Molecular Biology 32,1295–1309.
Lawniczak, M.K.N., Barnes, A.I., Linklater, J.R., Boone, J.M.,Wigby, S. & Chapman, T. (2006) Mating and immunity ininvertebrates. Trends in Ecology and Evolution 22, 48–55.
Levenbook, L., Bodnaryk, R.P. & Spande, T.F. (1969) P-alanyl-L-tyrosine: Chemical synthesis, properties and occurrence inlarvae of the fleshfly Sarcophaga bullata Parker. BiochemicalJournal 113, 837–841.
Li, J. & Christensen, B.M. (1993) Involvement of L-tyrosine andphenoloxidase in the tanning of Aedes aegypti eggs. InsectBiochemistry and Molecular Biology 23, 739–748.
Ling, E. & Yu, X.Q. (2006) Cellular encapsulation and melaniza-tion are enhanced by immulectins, pattern recognitionreceptors from the tobacco hornworm Manduca sexta.Developmental and Comparative Immunology 30, 289–299.
Luce-Fedrow, A., Von Ohlen, T. & Chapes, S.K. (2009) Ehrlichiachaffeensis infections in Drosophila melanogaster. Infection andImmunity 77, 4815–4826.
Luckhart, S., Vodovotz, Y., Cui, L. & Rosenberg, R. (1998) Themosquito Anopheles stephensi limits malaria parasite devel-opmentwith inducible synthesis of nitric oxide. Proceedings ofthe National Academy of Science USA 95, 5700–5705.
Manetti, A.G.O., Rosetto, M. &Marchini, D. (1998) Antibacterialpeptides of the insect reproductive tract. pp. 67–91 in Brey,P.T. & Hultmark, D. (Eds) Molecular Mechanisms of ImmuneResponses in Insects. New York, USA, Chapman & Hall.
Mavrouli,M.D., Tsakas, S., Theodorou, G.L., Lampropoulou,M.&Marmaras, V.J. (2005) MAP kinases mediate phagocytosisand melanization via prophenoloxidase activation in medflyhemocytes. Biochimica et Biophysica Acta, Molecular CellResearch 1744, 145–156.
McKean, K.A. & Nunney, L. (2001) Increased sexual activityreduces male immune function in Drosophila melanogaster.Proceedings of the National Academy of Science USA 98, 7904–7909.
Miller, J.S., Howard, R.W., Nguyen, A., Rosario, R.M.T. &Stanley-Samuelson, D.W. (1996) Eicosanoids mediatenodulation responses to bacterial infections in larvae of thetenebrionid beetle, Zophobas atratus. Journal of InsectPhysiology 42, 3–12.
Mitchell, H.K. & Lunan, K.D. (1964) Tyrosine-O-phosphatein Drosophila. Archives of Biochemistry and Biophysics 106,219–222.
Moreno-García, M., Lanz-Mendoza, H. & Córdoba-Aguilar, A.(2010) Genetic variance and genotype-by-environmentinteraction of immune response in Aedes aegypti (Diptera:Culicidae). Journal of Medical Entomology 47, 111–120.
Moret, Y. & Schmid-Hempel, P. (2000) Survival for immunity: theprice of immune system activation for bumblebee workers.Science 290, 1166–1168.
Müller, U. (1997) The nitric oxide system in insects. Progress inNeurobiology 51, 363–381.
Nappi, A.J. & Christensen, B.M. (2005) Melanogenesis andassociated cytotoxic reactions: applications to insect innateimmunity. Insect Biochemistry and Molecular Biology 35,443–459.
Nappi, A.J. & Ottaviani, E. (2000) Cytotoxicity and cytotoxicmolecules in invertebrates. BioEssays 22, 469–480.
Nappi, A.J., Vass, E., Frey, F. & Carton, Y. (2000) Nitric oxideinvolvement in Drosophila immunity. Nitric Oxide 4, 423–430.
Narayanan, K. (2004) Insect defence: its impact on microbialcontrol of insect pests. Current Science 86, 800–814.
Nardi, J.B., Pilas, B., Ujhelyi, E., Garsha, K. & Kanost, M.R.(2003) Hematopoietic organs of Manduca sexta and hemo-cyte lineages. Development, Genes and Evolution 213,477–491.
Osanai,M.&Chen, P. (1993) A comparative study on the argininedegradation cascade for sperm maturation of Bombyx moriand Drosophila melanogaster. Amino Acids 5, 341–350.
Otvos, L. Jr (2000) Antibacterial peptides isolated from insects.Journal of Peptide Science 6, 497–511.
Packer, C., Holt, R.D., Hudson, P.J., Lafferty, K.D. & Dobson, A.P. (2003) Keeping the herds healthy and alert: implicationsof predator control for infectious disease. Ecology Letters 6,797–802.
Pech, L.L., Trudeau, D. & Strand, M.R. (1994) Separation andbehavior in vitro of hemocytes from the moth, Pseudoplusiaincludens. Cell Tissue Research 277, 159–167.
Pham, L.N., Dionne, M.S., Shirasu-Hiza, M. & Schneider, D.S.(2007) A specific primed immune response in Drosophila isdependent on phagocytes. PLoS Pathogens 3, e26.
Rantala, M.J. & Kortet, R. (2003) Courtship song and immunefunction in the field cricket Gryllus bimaculatus. BiologicalJournal of the Linnean Society 79, 503–510.
Rantala, M.J. & Kortet, R. (2004) Male dominance and im-munocompetence in a field cricket. Behavioral Ecology 15,187–191.
Rantala, M.J. & Roff, D.A. (2005) An analysis of trade-offs inimmune function, body size, and developmental time in theMediterranean Field Cricket, Gryllus bimaculatus. FunctionalEcology 19, 323–330.
Rantala, M.J. & Roff, D.A. (2007) Inbreeding and extreme out-breeding cause sex differences in immune defence and lifehistory traits in Epirrita autumnata. Heredity 98, 329–336.
Rantala, M.J., Koskimäki, J., Taskinen, J., Tynkkynen, K. &Suhonen, J. (2000) Immunocompetence, developmentalstability and wingspot size in the damselfly Calopteryxsplendens L. Proceedings of the Royal Society of London, Series B267, 2453–2457.
Read, A.F., Graham, A.L. & Råberg, L. (2008) Animal defensesagainst infectious agents: is damage control more importantthan pathogen control. PLoS Biology 6, e1000004.
Ribeiro, C. & Brehélin,M. (2006) Insect haemocytes:What type ofcell is that? Journal of Insect Physiology 52, 417–429.
Riddiford, L.M. & Hiruma, K. (1988) Regulation of melanizationin insect cuticle. Program of Clinical Biological Research 256,423–436.
Rigby, M. & Jokela, J. (2000) Predator avoidance and immunedefence: costs and trade-offs in snails. Proceedings of the RoyalSociety of London, Series B 267, 171–176.
Rivero, A. (2006) Nitric oxide: an antiparasitic molecule ofinvertebrates. Trends in Parasitology 22, 219–225.
Robb, T., Forbes, M.R. & Jamieson, I.G. (2003) Greater cuticularmelanism is not associated with greater immunogenic re-sponse in adults of the polymorphic mountain stone weta,Hemideina maori. Ecological Entomology 28, 738–746.
Roberts, M.L., Buchanan, K.L. & Evans, M.R. (2004) Testing theimmunocompetence hypothesis: a review of the evidence.Animal Behaviour 68, 227–239.
Roff, D.A. (1992) The Evolution of Life Histories. New York, USA,Chapman & Hall.
Rolff, J., Armitage, S.A.O. & Coltman, D.W. (2005) Geneticconstraints and sexual dimorphism in immune defence.Evolution 59, 1844–1850.
M. Moreno-García et al.12

Rolff, J. & Reynolds, S. (Eds) (2009) Insect Infection and Immunity:Evolution, Ecology and Mechanisms. Oxford, UK, OxfordUniversity Press.
Roy,M. &Holt, R.D. (2008) Effects of predation on host-pathogendynamics in SIR models. Theoretical Population Biology 73,319–331.
Ryder, J.J. & Siva-Jothy, M.T. (2001) Quantitative geneticsof immune function and body size in the house cricket,Acheta domesticus. Journal of Evolutionary Biology 14,646–653.
Saul, S. & Sugumaran, M. (1988) A novel quinone: quinone me-thide isomerase generates quinone methides in insect cuticle.FEBS Letter 237, 155–158.
Schmid-Hempel, P. (2005a) Evolutionary ecology of insectimmune defences. Annual Review of Entomology 50, 529–551.
Schmid-Hempel, P. (2005b) Natural insect host-parasite systemsshow immune priming and specificity: puzzles to be solved.BioEssays 27, 1026–1034.
Schneider, D.S. & Ayres, J.S. (2008) Two ways to surviveinfection: what resistance and tolerance can teach us abouttreating infectious diseases. Nature Reviews Immunology 8,889–895.
Schwartz, A. & Koella, J.C. (2004) The cost of immunity in theyellow fever mosquito, Aedes aegypti depends on immuneactivation. Journal of Evolutionary Biology 17, 834–840.
Sheldon, B.C. & Verhulst, S. (1996) Ecological immunology:costly parasite defences and trade offs in evolutionary ecol-ogy. Trends in Ecology and Evolution 11, 317–321.
Simmons, L.W., Zuk, M. & Rotenberry, J.T. (2005) Immunefunction reflected in calling song characteristics in a naturalpopulation of the cricket Teleogryllus commodus. AnimalBehaviour 69, 1235–1241.
Siva-Jothy, M.T. (2000) A mechanistic link between parasite re-sistance and expression of a sexually selected trait in adamselfly. Proceedings of the Royal Society of London, Series B267, 2523–2527.
Söderhäll, K. & Cerenius, L. (1998) Role of the prophenoloxidase-activating system in invertebrate immunity. Current Opinionin Immunology 10, 23–28.
Stearns, S.C. (1992) The Evolution of Life Histories. Oxford, UK,Oxford University Press.
Stevenson, R.D. (2006) Ecophysiology and conservation: thecontribution to energetics – introduction to the symposium.Integrative and Comparative Biology 46, 1088–1092.
Sugumaran, M. & Kanost, M.R. (1993) Regulation of insecthaemolymph phenoloxidases. pp. 317–342 in Beckage, N.E.,Thompson, S.N. & Federico, B.A. (Eds) Parasites and Pathogensof Insects. Volume I. Parasites. San Diego, CA, USA, AcademicPress.
Tarlow, E.M. & Blumsteid, D.T. (2007) Evaluating methods toquantify anthropogenic stressors on wild animals. AppliedAnimal Behaviour Science 102, 429–451.
Thompson, J.J.W., Armitage, S.A.O. & Siva-Jothy, M.T.(2002) Cuticular colour change after imaginal eclosion istime constrained: blacker beetles darken faster. PhysiologicalEntomology 27, 136–141.
Uchida, K. (1993) Balanced amino acid composition essential forinfusion-induced egg development in the mosquito (Culexpipiens pallens). Journal of Insect Physiology 39, 615–621.
Williams, M.J. (2007) Drosophila hemopoiesis and cellular im-munity. Journal of Immunology 178, 4711–4715.
Wilson, K., Knell, R., Boots,M. &Koch-Osborne, J. (2003) Groupliving and investment in immune defence: an interspecificanalysis. Journal of Animal Ecology 72, 133–143.
Insect immuno-ecology markers 13