current distribution not indicative of underlying …
TRANSCRIPT

CURRENTDISTRIBUTIONNOTINDICATIVEOFUNDERLYINGNICHEREQUIREMENTSOFANALPINEFORB:IMPLICATIONSFORPREDICTING
CLIMATE-INDUCEDRANGESHIFTS
EmmaSumner
AthesissubmittedinpartialfulfilmentoftherequirementsforthedegreeofBachelorofScience(WildlifeandConservationBiology)(Hons)
inthe
DepartmentofEcology,Environment&Evolution
LaTrobeUniversityBundoora,Victoria
24thApril2017wordcount:9690

II
Declaration
Icertifythattheattacheddocumentismyoriginalwork.Nootherperson’sworkhasbeen
usedwithoutdueacknowledgement.ExceptwhereIhaveclearlystatedthatIhaveused
someofthismaterialelsewhere,ithasnotbeenpresentedbymeforexaminationinany
othercourseorsubjectatthisoranyotherinstitution.Iunderstandthattheworksubmitted
maybereproducedand/orcommunicatedforthepurposeofdetectingplagiarism.
Noneoftheresearchundertakeninconnectionwiththisthesisrequiredapprovalbya
UniversityEthicsCommittee.
Name:EmmaElizabethSumner
Subject:EEE4HNA/B
Document:Thesis
StudentNumber:17724437
Date:24/4/2017

III
AcknowledgementsIwouldliketoacknowledgethefollowingpeople:Dr.JohnMorganforhissupervision,forprovidingmewithachallengingproject,andforhisguidancethroughoutthewholeprocess.ThePlantEcologyLab,forthethought-provokingweeklymeetingsandsupportthroughouttheyear.ThankstoSueByceson,Dr.PeteGreen,Dr.SusannaVennandDr.DeniseFernandofortheirastutesuggestionsregardingexperimentaldesign.ThanksalsotoZacWalker,GeorgeCollinsandHannaKapuscinski-Evansfortheirsupportandcomradery.MaxBartleyandNickMoorefortheirtechnicalassistanceandhardwork,andtheLaTrobeWildlifeSanctuaryforassistingwithseedpropagation.ThankstovolunteersStephJohnsonandJackTatefortheirassistanceinthefield.BevLawrenceandAPEASkiLodgefortheirhospitality,andMtHothamSkiResortforgraciouslyallowingmetoconductfieldresearchatMtHotham.Dr.SimonWatsonforintroducingmetostatisticalmodellingandguidingmethroughthedataanalysisprocess,andWarwickPapstforhisadviceonworkingonthehighplains.IwouldalsoliketothankandacknowledgetheResearchCentreforAlpineEcologyfortheirgenerousresearchgrant.Finally,thankyoutotheentireDepartmentofEcology,EnvironmentandEvolution.

IV
TableofContents
EXECUTIVESUMMARY.............................................................................................................1
INTRODUCTION.......................................................................................................................2FINGERPRINTSOFCHANGE...........................................................................................................2MODELLINGFUTUREPLANTDISTRIBUTIONS–CORRELATIVEAPPROACHES..............................................3THEROLEOFDISPERSALINDETERMININGRANGELIMITS....................................................................4AMORE‘IN-DEPTH’UNDERSTANDINGOFRANGELIMITS.....................................................................5THEROLEOFBIOTICINTERACTIONSINDETERMININGRANGELIMITS......................................................6THEROLEOFBIOTICINTERACTIONSINDETERMININGFUTUREDISTRIBUTIONS..........................................8
METHODS...............................................................................................................................10STUDYSPECIES........................................................................................................................10STUDYSITE.............................................................................................................................11SITECONDITIONS.....................................................................................................................12FIELDTRANSPLANTEXPERIMENT.................................................................................................14LABORATORYGERMINATIONEXPERIMENT.....................................................................................14LIGHTLIMITATIONEXPERIMENT..................................................................................................15FREEZINGRESISTANCEEXPERIMENT.............................................................................................16DATAANALYSIS.......................................................................................................................17
RESULTS..................................................................................................................................20SITECONDITIONS.....................................................................................................................20FIELDTRANSPLANTRESULTS.......................................................................................................22LABORATORYGERMINATIONRESULTS...........................................................................................24LIGHTLIMITATIONRESULTS........................................................................................................25FREEZINGRESISTANCERESULTS..................................................................................................27
DISCUSSION............................................................................................................................28GERMINATIONCHARACTERISTICS................................................................................................28FROSTTOLERANCEINSEEDLINGS.................................................................................................29EFFECTSOFFACILITATIONALONGANABIOTICSTRESSGRADIENT.........................................................30EFFECTSOFCOMPETITIONONSEEDLINGGROWTH..........................................................................32CONCLUSION..........................................................................................................................34
REFERENCES...........................................................................................................................36
APPENDICES...........................................................................................................................42APPENDIXI............................................................................................................................42APPENDIXII...........................................................................................................................45APPENDIXIII..........................................................................................................................46APPENDIXIV..........................................................................................................................47

V
ListofFiguresFIG1.OCCURRENCERECORDSOFALPINEPODOLEPIS(ATLASOFLIVINGAUSTRALIA)................................11FIG2.PODOLEPISROBUSTAINFLOWER(IMAGE:J.MORGAN).............................................................11FIG3.LOCATIONOFMTHOTHAMINVICTORIA,ANDSTUDYSITELOCATIONSATMTHOTHAM(1-10)REPRESENTINGTHEELEVATIONGRADIENTONSOUTH-EASTANDNORTH-WESTSITES..................................13FIG4.TOTALNUMBEROFFREEZINGDAYS(TEMPERATURES≤0ᵒC)RECORDEDATMTHOTHAM,INRELATIONTOALTITUDEDURINGSTUDYPERIOD(NOV2016–MAR207)ONNORTH-WEST(SOLIDLINE)ANDSOUTH-EAST(DASHEDLINE)EXPOSEDSLOPES......................................................................................................20FIG5.GROWINGDEGREEDAYS(GDD)RECORDEDFORSOUTH-EAST(A)ANDNORTH-WEST(B)EXPOSEDSITESDURINGSTUDYPERIOD(NOV2016–MAR2017)............................................................................21FIG6.EFFECTOFTREATMENTANDALTITUDEONSEEDLINGSURVIVAL.MEANPROBABILITYOFSURVIVAL(±95%
CI)OVERONEGROWINGSEASONFORSEEDLINGSPLANTEDALONGANALTITUDINALGRADIENTINCONTROL(SOLIDGREENLINE)ANDGAP(SOLIDORANGELINE)TREATMENTS...................................................................22FIG7.MEANRELATIVENEIGHBOUREFFECT(RNE)(±1SE)FORPAIREDTRANSPLANTEDSEEDLINGSINGAPANDCONTROLTREATMENTSON(A)NORTH-WESTAND(B)ANDSOUTH-EASTEXPOSEDASPECTSACROSSTHEELEVATIONGRADIENT...................................................................................................................23FIG8.EFFECTOFLIGHTANDTEMPERATUREONGERMINATION.FITTEDVALUESFORPROBABILITYOFGERMINATIONFOR(A)LIGHTCONDITIONSAND(B)DARKCONDITIONS(±95%CI)FORTHEGENERALISEDLINEARMODELAPPLIEDONCABINETGERMINATIONDATA..............................................................................24FIG9.EFFECTOFLIGHTLIMITATIONONNETPHOTOSYNTHESIS.FITTEDMEANRATEOFPHOTOSYNTHESIS(±95%
CI)OFSEEDLINGSGROWNUNDERFULLSUN,48%SHADEAND82%SHADE...........................................26FIG10.EFFECTOFLIGHTLIMITATIONONBIOMASS.FITTEDMEANDRY-WEIGHTOFBIOMASS(±95%CI)OFSEEDLINGSGROWNUNDERFULLSUN,48%SHADEAND82%SHADE.....................................................26FIG11.MEAN(±1SE)VARIABLEFLUORESCENCETOMAXIMUMFLUORESCENCE(FV/FM)RATIOSOFLEAFMATERIALUSEDINFREEZINGRESISTANCESTUDY,ATFOURTEMPERATURES:4,-5,-11,AND-20°C;FORALPINEPODOLEPISLEAVESOF(A)MATUREPLANTSAND(B)SEEDLINGS.............................................................27FIG12.SHADECASTBYRESIDENTVEGETATIONATNORTH-WESTSITESATMTHOTHAM,VICTORIA..............42FIG13.SHADECASTBYRESIDENTVEGETATIONATSOUTH-EASTSITESATMTHOTHAM,VICTORIA...............42FIG14.FREQUENCYOFVEGETATION(HERB,GRAMMINOID,SHRUB)HEIGHTSATMTHOTHAMONSOUTH-EASTEXPOSEDSITES(A-E).....................................................................................................................43FIG.15.FREQUENCYOFVEGETATION(HERB,GRAMMINOID,SHRUB)HEIGHTSATMTHOTHAMONNORTH-WESTEXPOSEDSITES(A-E).............................................................................................................44FIG16.EUCALYPTUSPAUCIFLORAWOODLANDWITHDENSEPODOLOBIUMALPESTREUNDERSTORYDOMINATE
NORTH-WESTEXPOSEDSLOPESATMTHOTHAM(1670-1700M).......................................................45

VI
ListofTablesTABLE1.SUMMARYINFORMATIONFORTHETENSITESALONGTHEMTHOTHAMTRANSECTINCLUDING
LOCATION,ALTITUDEANDASPECTFOREACHSITE...............................................................................13TABLE2.GERMINATIONATTRIBUTESOFALPINEPODOLEPISAT14°CWITHANDWITHOUTAPERIODOFCOLD
STRATIFICATION..........................................................................................................................25TABLE3.ANALYSISOFTHEEFFECTOFTREATMENT,ALTITUDEANDASPECTONSEEDLINGSURVIVAL.............46TABLE4.ANALYSISOFTHEEFFECTOFTREATMENTANDALTITUDEONSEEDLINGSURVIVAL.........................46TABLE5.TABLEOFCOEFFICIENTSFORTHEGLMUSEDTOFITSEEDLINGSURVIVALASAFUNCTIONOF
TREATMENTANDALTITUDE...........................................................................................................46TABLE6.STATISTICALANALYSISOFRELATIVENEIGHBOUREFFECTFORNORTH.......................................47

1
ExecutiveSummary
Astheclimatewarms,plantspeciesarepredictedtomoveupwardandpolewardinorderto
remainwithintheirclimaticenvelopes.Thesepredictionsaremadebyextrapolatingcurrent
distributionstofutureclimatescenariosundertheassumptionthatplantboundariesare
controlledbyclimatealone.However,species-specificresponsestoclimatechangearelikely
morecomplexthancanbeforecastwithsimpleclimatemodelling.Understandingcontrols
onspeciesgeographicalrangelimits,andhowthesemayrespondtorapidenvironmental
changehaspromptedcallsforgreater‘bruteforce’experimentation.Here,Iinvestigate
controlsonthedistributionofamodelalpinespeciesPodolepisrobusta(Asteraceae)by
conductingaseriesoffieldandlaboratoryexperiments.Germinationexperiments
demonstrateawidegerminationniche,withnolimitingdormancycharacteristicsorcold
stratificationrequirements.However,whenseedlingsweresubjectedtofreezing
temperatures,leavessufferedsignificantdamagetophotosyntheticapparatuscomparedto
adultplants.Hence,itislikelythatearlyseedlingestablishmentisstronglytiedto
occurrencesofearlyfrostduringthegrowingseason.Facilitativeinteractionsplayastrong
roleacrossrangeedges.Asdemonstratedbyatransplantexperimentalongatemperature
andmoisturegradient,closeinterspecificneighbourssignificantlyincreasedsurvivaland
growthofplantedseedlings,comparedtoseedlingsplantedincanopygaps.Thispatternwas
consistent,evenbelowthecurrentdistribution,contrarytoexpectationsofstronger
competitiveinteractions.Whilethefieldstudyfindsnoresponsetocompetitiononseedling
survival,experimentalshadingindicatesintoleranceatheavy(82%)lightinterception.This
hasconsequencesforestablishmentwithindensevegetation.Resultsconsistentlyindicate
thatP.robustaisabletooccupyafargreaternichethancanbeobservedcurrently.These
resultshighlighttheimportanceoffacilitativeinteractionsattheseedlingstageinthealpine
zone,demonstratingthatbioticfactorsacttoconstrainorwidenthetheoreticalniche.I
arguethatbioticinteractions,dispersallimitation,andrecruitmentprocessesmayenforce
strongerlimitstogeographicdistributionthanclimatictolerancesperse.Predictionsonhow
plantspeciesmayrespondtoclimatechangewillbenefitfromincorporatingthesefactors.

2
Introduction
Thenicheisafundamentaltenetinecology,andframesourunderstandingofspeciesrange
limits(Holt2009).Climateandotherabioticfactorsareoftendemonstratedashavingstrong
influencesonplantspeciesdistributions(KörnerandPaulsen2004).Hence,responsesto
climatechangeareoftencouchedintermsofclimatictolerances.Inalpineecosystems,
climatewarmingisexpectedtodrivespeciesdistributionstohigheraltitudes(Dullingeretal.
2012).Theserangeshiftshaveimportantconservationimplications,astheupwardmigration
ofmorecompetitive,lower-altitudeplants,andthereductionofsuitablehabitatforhigh-
altitudeplants,couldpotentiallyleadtorangecontractionsandspeciesextinctions(Thuiller
etal.2005;Dullingeretal.2012;Alexanderetal.2015).However,geographicdistributionsof
plantspeciesresultfromacombinationofabiotictolerancesandbioticfactorssuchas
speciesinteractions,dispersalanddemographicprocesses(Holt2009).Thecomplexityand
urgencyofpredictingthepotentialimpactofclimatechangeonspeciesdistributionshas
beenincreasinglymetwithcallsforgreater‘brute-force’experimentation(Holt2009;Munier
etal.2010;HilleRisLambersetal.2013;Alexanderetal.2015;Alexanderetal.2016).
Experimentalassessmentsofspeciesdistributionsprovideadirectapproachinattemptingto
quantifytheniche,andareapowerfultoolformakinginferencesaboutspatialheterogeneity
ofabioticfactorsandspeciesinteractions(Moore2009).
Fingerprintsofchange
Inmountains,duetoanaltitudinaltemperaturegradient,speciesareexpectedtomigrate
upslopeinresponsetoclimaticshiftsinvolvingwarmingtemperaturesandlongergrowing
seasons(Paulietal.1996).Revisitationofhistoricalsitesprovidesoverwhelmingevidencefor
globaltrendsinupwardspeciesmigration(Paulietal.2012),withincreasesinspecies
richnessreportedforsummitsintheEuropeanAlps(ParoloandRossi,2008;Paulietal.
2012),theHimalayas(Telwalaetal.2013),theAndes(Feeleyetal.2011)andtheAustralian
Alps(Vennetal.2012).Inparticular,speciesgainsaremorepronouncedonsummitsof
loweraltitudes,owingtolargernearbyspeciespoolsandsuggestingthatupwardshiftsare
drivenlargelybyleading-edgeexpansionsinresponsetoincreasingtemperature(Paulietal.

3
2012).However,contrastingtrendswerereportedforMediterraneanmountainregions
(Paulietal.2012),andmountainregionswithMediterranean-typeclimates(Crimminsetal.
2011),whereratherthantrackingtemperatures,plantspecieshaverespondedtoa
decreasedwaterdeficitbyshiftingtheiroptimalelevationsdownslope.Crimminsetal.(2011)
alsoreportthatspecies’entireranges–includinglowerboundaries–areshifting,ratherthan
justthroughupper-boundaryextensionsasobservedintemperateregions.Such
contradictoryresponsestoclimatechangeunderminetheassumptionthattemperatureis
theprinciplefactorindeterminingspeciesdistributions,andhighlightstheimportanceof
otherclimaticfactorssuchasenergyandwateravailability(Crimminsetal.2011;Cahilletal.
2012).Ultimately,theupwardtrendinspeciesmigrationsisexpectedtoresultinrange
contractionandextinctionofmountainplantspecies,manyofwhicharerareandendemic
(Dullingeretal.2012).However,long-termrevisitationstudiesarelimitedinspatialand
temporalscope,promptingmanyecologiststoemploycomputermodellinginorderto
predictpotentialresponsestoclimatechange(Brookeretal.2007).
Modellingfutureplantdistributions–correlativeapproaches
Typically,climateenvelopemodellingisemployedtopredictspeciesrangesinthecontextof
climatechange(Brookeretal.2007).Speciesrangelimitsareassumedfromsimple
correlationsofabioticconditionswithincurrentdistributions.These‘climaticenvelopes’are
thenextrapolatedtofutureclimatescenarios(Brookeretal.2007)inordertocreatea
‘statisticalnichemodel’(Holt2009).Frequently,speciesdistributionmodelspredict
substantialimpactsonalpineplantdiversity,owingtohighratesofhabitatloss(Dullingeret
al.2012).InaEuropean-scaleanalysis,aclimate-envelopebasedmodelpredictedhabitat
lossesofapproximately60percentofmountainspecies(Thuilleretal.2005).However,
estimatesofhabitatlossmaydependonthescaleofthemodel.Finescale(local)models
predictedthepersistenceofallhabitatsprojectedtonolongerexistatcoarseresolutions
(Europeanscale)(Randinetal.2009).Thisislikelybecausetopographyandmean
temperaturedatacontainmorevariabilityatfinescalethancanbeexpressedatlargerscales
(Thuilleretal.2005).Indeed,temperaturedatacollectedlocallyindicatesthathigh
topographicvariabilitywillconservetheclimaticnichesofmanyspeciesintheSwissAlps

4
(SherrerandKörner2010).Hence,inconsistenciesinspeciesdistributionmodelling
demonstratethatestimationsofhabitatlossdependupondataresolution.Thishas
implicationsfortheaccuracyofpredictionsofhabitatlossandspeciesextinctions.Further,
predictivemodelsignorecrucialelementsaffectingspeciesdistributionssuchasdispersal
traitsandbioticinteractions,andassuchprovideonlyaroughestimateofpossibleresponses
toclimatechange(Holt2009;Urbanetal.2012).
Manyecologicalassumptionsandtheoriescaninfluencepredictionsofspeciesdistributions
underclimatechange(Austin2002).Brookeretal.(2007)recognisetheshortcomingsof
climate-basedmodelling,andincorporatebioticinteractionssuchascompetitionand
facilitationbetweenplantspecies,andplantdispersalabilityintoaspatialmodelofa
theoreticalcommunity.Here,theinclusionofbioticprocessesprovidedcontrarypredictions
forfuturespeciesdistributionsthanthosebasedpurelyonclimaticcorrelation.However,
whilethisisusefulinhighlightingthequalitativeimportanceofnon-climaticvariablesinplant
distribution,itdoessowiththeuseofartificialdata.Assuch,isnotnecessarilyindicativeof
thepatternsandprocessesfoundinnaturalplantcommunities.Predictivemodelsrequire
greaterunderstandingoftheecologicalprocessesthatinfluencespeciesdistributionsinthe
contextofclimatechange.Whilespeciesdistributionmodelsassumethatrealisednicheswill
beretainedovertime,withspeciesmigratingtoremainwithincurrentclimaticenvelopes
(Thuilleretal.2005;Brookeretal.2007),limitationsofdispersalandestablishmentsuccess
maypreventaspeciesfromshiftingitsrangetokeeppacewithachangingclimate(Urbanet
al.2012).
Theroleofdispersalindeterminingrangelimits
Withongoingclimatechange,certainclimaticfiltersincludingtemperatureandsnow-
durationarerelaxedinalpineenvironments(Alexanderetal.2016).Thesechangeshavethe
potentialtoopenuphabitattoplantspeciesattheirleading-edge(Alexanderetal.2015).
Globally,alpinetree-linesarethoughttobestronglytemperature-controlled(Slatyerand
Noble1992;Harschetal.2009),andthereforepotentiallyresponsivetowarmer
temperatures(HoltmeierandBroll2005).However,ameta-analysisrevealedupward

5
advancementsatonlyhalftheexaminedtree-lines(Harschetal.2009).InAustralia,though
warmertemperatureshaveextendedthefundamentalnicheattheupperboundaryofsnow
gumsby100m,thetree-linehasremainedrelativelystable(Green2009).Evenafter
extensivefiresintheAustralianAlpsprovidedsuitableconditionsforseedlingestablishment,
mostrecruitmentwasseenwithinandbelowthetree-line,indicatingthatupslopemigration
maybelimitedbydispersalability.IntheItalianAlps,speciesthathavemigratedupslopethe
farthestsincethe1950sarethosethatpossessedlight,wind-disperseddiaspores(Paroloand
Rossi2008).
Aspreviouslydiscussed,plantdispersaltraitshavebeenincorporatedintosomespecies
distributionmodels,withwind-dispersaldenotinggreatermigrationsuccess(Austin2002;
Brookeretal.2007).However,thoughprimarilywind-dispersed,tree-linespeciesinthe
northernhemisphereshowlittlesignofupslopemigration,suggestingpost-dispersal
mechanismsmayinfluenceleading-edgeexpansions(HobbieandChapin1998).Similarly,
thoughdispersalhasbeenfoundtobesufficient,therehasbeenlittlesignofupslope
advancementintheScandinavianmountainsbymountainbirch,potentiallyduetothe
interactionofmicro-topography,climateandherbivory(Hofgaardetal.2009;Speedetal.
2010).Specieslimitedbypre-orpost-dispersalprocessesarethereforeatriskoffailingto
remainwithintheircurrentclimate-envelopeswithcontinuedclimatewarming.Certainly,
migrationisaprocessnotonlytiedtotraitssuchasseedsizeanddispersalability,butalso
contingentonpropaguleavailability,germinationniche,andseedlingestablishment.
Amore‘in-depth’understandingofrangelimits
Thereislittledoubtthatclimateunderpinsthelarge-scaledistributionsofmanyplant
species(Körner1999),andthisisindeedamajorassumptioninmostspeciesdistribution
models(Brookeretal.2007;Randinetal.2009;Dullingeretal.2012).Thisisparticularly
evidentformountainplantswheretheupperboundaryofalpineorsubalpinespeciescan
becorrelatedwithmeanseasonaltemperature(Körner1999).However,themechanism
behindtheseboundariesaremorecomplexthancanbeexpressedbyaveragegrowing
seasontemperaturestypicallyusedbyclimate-envelopemodelling(Körneretal.2016).In

6
mountainplants,irregularfreezingtemperaturesduringthegrowingseasonaremore
likelytoresultintissuedeathandplantmortalitythanaverageminimumtemperature
(Larcher1995).Körneretal.(2016)demonstratethatfrostdamageduringbudbreakand
leafemergenceduringthegrowingseasonareinstrumentalinestablishingthecold-edge
rangelimitsoffivetreespeciesinEurope.Freezingresistanceisalsospecies-specificand
relatedtolocation,withhigherfreezingresistancereportedforplantpopulationswith
increasingaltitude(Vennetal.2013).Inaddition,differentlifestagesofplantsexhibit
unequalfrostresistance:whileadultplantsmaybeabletore-sproutfromprotectedbuds,
seedlingsaremuchmoresensitivetofreezingtemperatures(Larcher1995).Indeed,entire
cohortsmaybedevastatedbyoneincidenceofearlyspringfrost(Larcher1995).Hence,
individuallife-historystagesmayresponddifferentlytoabioticconditions,andwill
contributeunequallytopopulationviabilitybeyondcurrentranges(WarrenandBradford
2011).
Theroleofbioticinteractionsindeterminingrangelimits
Competitionandfacilitationalsoplayimportantrolesinthestructureandassemblageof
alpineplantcommunities(Holt2009).Therelativeimportanceofsuchinteractionsmaybe
distinguishedbycomparingplantsurvivalorproductivityinthepresenceorabsenceof
benefactors,orwhenacomponent(suchasthecanopy)ofthebenefactorisaltered
(BertnessandCallaway1994).PositiveassociationswithinvasiveTaraxacumofficinaleand
thecushionplantAzorellamonanthaintheChileanAndeswasconfirmed,asT.officinale
seedlingsplantedwithinthecanopiesofA.monanthaexperiencedhighersurvival,net-
photosynthesisandstomatalconductancethanthoseplantedinbarepatches(Cavieresetal.
2005).Theacquisitionof,anduseofresourcesnecessaryforplantgrowthsuchasnitrogen
andwater,arelinkedtophotosyntheticperformance(Muraoka2002).Hence,amajor
limitationofalpineplantgrowthisphotosyntheticallyactiveradiation(PAR,400-700nm)
duringthegrowingseason(Körner1999).AreductioninincidentPARfromneighbouring
plants,whileconstituting‘resourcecompetition’,canbemitigated,orbalancedbyindirect
(positive)effectsofreducedsolarinsolationontopsoilmoisture(Körner1999).

7
Positivespatialassociationsbetweenmatureadultsofonespeciesandtheseedlingsof
anotherarewidelyreferredtoasthe‘nurseplant’syndrome,andtypicallyoccurinstressful
habitats(Callaway1995;Callawayetal.2002).Athighelevations,ameliorationofstressors
suchasstrongwinds,soilinstabilityanddroughtbystress-tolerantneighbours,facilitatesthe
persistenceoflessstress-tolerantspecies(BertnessandCallaway1994).Whileneighbours
positivelyinfluencedthegrowthoftargetspeciesintheTibetanPlateau,positiveresponses
wererelatedtospeciesdensity,suggestingthatbothcompetitionandfacilitationwere
operatinginparallel(Wangetal.2007).Thisisbecausethemagnitudeofpositiveand
negativeplantinteractionschangesacrossaspeciesrange(BertnessandCallaway1994).
Itisgenerallyacceptedthataspecies’lowerboundaryisconstrainedbycompetitive
interactionsandextendedatupperboundariesbyfacilitationbymorestress-tolerantspecies
(BertnessandCallaway1994).Theapparentinverserelationshipbetweencompetitiveability
andstress-tolerancewaspopularisedbyGrime(1977),andisneatlydemonstratedwith
manipulationsalonganenvironmentalgradient.Plantspeciesrespondedtoneighbour
removalsaccordingtotheirpositionalonganaltitudinalgradientintheFrenchAlps(Choler
etal.2001).Whenneighbourswereremovedfromtargetspeciesattheirtrailing-edge,
biomassincreased,whilebiomassdecreasedwithneighbourremovalattheleadingedge,
wherespeciesaretypicallymoreconstrained(Choleretal.2001).Thisstress-gradient
hypothesisissupportedbyevidencefromneighbourremovalexperimentsconducted
worldwidewheregenerally,interactionsshiftedfromcompetitiontofacilitationaselevation
andabioticstressincreased(Callawayetal.2002).Theselarge-scalemanipulative
experimentshighlighttheunderestimationofplant-plantinteractionsinecologicaltheory
(Choleretal.2001).And,whilethebalancebetweencompetitionandfacilitationshiftsacross
space,itmayalsoshiftacrosstime.
Potentiallypositiveornegativeresponsestoneighboursmaychangedependingonthelife
stagesoftheinteractingspecies(CallawayandWalker1997),ortemporalfluctuationsinthe
environment(Valiente-Banuet2008).Indeed,theaforementionednurse-plantsyndromeis
typicallyfavourableonlyforbeneficiariesduringearlylife-stagesandmayswitchtoa
competitiveinteractionastheseedlingmatures(McAuliffe1988;Callaway1995).
Interactionscanalsoshiftwithvariationinclimateoverthecourseofasinglegrowingseason

8
(Kikvidzeetal.2006),whereabioticstressorsarerelaxedorheightened(Bertnessand
Shumway1993).Furtherchangesinenvironmentalconditionsmayalterthebalanceof
facilitativeandcompetitiveinteractionsbetweenplantspecies.Inalpineenvironments,the
relaxationoralterationofcertainclimaticfilterswithongoingclimatechangeislikelyto
changethedirectionandstrengthofplantinteractions.
Theroleofbioticinteractionsindeterminingfuturedistributions
Aspredicted,specieshavebeguntoshifttheirdistributionsinresponsetoclimatechange.
Thisisespeciallyevidentinalpinesystemswheresteepelevationalgradientsprovideshort-
distanceescapes(Bertrandetal.2011).Transplantexperimentsarevaluableinteasingout
therelativeimportanceofbioticfactorsbeyondaspecies’currentrange.InArctictundra,the
growthandsurvivalofseveralspeciestransplantedabovecurrentelevationlimitswaslimited
bybelow-groundcompetitionfromtundraplants(HobbieandChapin1998).Conversely,
germinationandestablishmentofsugarmaplewaslowestbeyonditscurrentupper-
boundaryduetogranivorybythesouthernred-backedvole(BrownandVellend2014).In
contrast,rootpathogenslimitupwardexpansionofsugarmaple(BrownandVellend,2014),
whilefungalendophytesextendtherangeofBromuslaevipesduetoconferreddrought
resistance(Afkhamietal.2014).
Variationsinestablishmentsuccessbeyondcurrentrangeswilllikelyresultinthe
reorganisationofcommunitymembersintonovelplantassemblages(Urbanetal.2012).
Alexanderetal.(2015)testtheeffectsofasynchronousmigrationontheperformanceof
fourtargetalpineplants.Whenalpineplantsweretransplantedtoloweraltitudestosimulate
migrationfailures,performancewashinderedbynovelcompetitors.Conversely,
performanceofthesamespecieswasenhancedwhentransplantedintonovelhigh-altitude
communities.Thisasymmetryincompetitiveabilitybetweenplantsfromlowerandhigher
altitudessuggeststhatspeciesarelikelytobeaffectedatthetrailingedgeoftheirranges
(Alexanderetal.2015).

9
Currentandfuturedistributionsofplantspeciesarecontrolledbyacombinationofabiotic
andbioticfactors,actinginconcert.Theseforcesaredynamic,shiftingovertemporaland
spatialscalesbutareoftenignoredorminimisedinspeciesdistributionmodelling.
Therelative,andsimultaneousimportanceofclimate,bioticinteractions,andother
constraintsoncurrentandfuturedistributions,canonlybeseparatedbyexperimental
manipulation;however,empiricalstudiesofthistypearerare(Holt2009).Iconducteda
seriesoffieldandlaboratoryexperimentsontheecologyandphysicaltolerancesofamodel
speciesAlpinePodoelpis(Podolepisrobusta,Asteraceae).Iaimedtoinvestigatehowabiotic
andbioticfactorsaffectseedlingsurvival,inordertomakeinferencesonconstraintsto
currentandfuturedistributions.Thetypeandstrengthofplantinteractionswere
determinedwithatransplantexperimentaboveandbelowthecurrentdistributionofAlpine
PodolepisatMtHotham,alongatemperatureandmoisturegradient.Inaddition,laboratory
experimentsassessedgerminationcharacteristics,andfreezingandshadetoleranceof
seedlings.Specifically,Itestedthefollowinghypotheses:
(i)competitionwillreduceseedlingsurvivalandgrowthatloweraltitudes,andfacilitationwill
increasesurvivalandgrowthathigheraltitudes;
(ii)germinationnicheforAlpinePodolepisisnarrow;
(iii)AlpinePodolepissurvivalandgrowthisrestrictedbylightlimitation;and
(iv)earlyseedlingsurvivalisconstrainedbyfreezingtemperatures.

10
Methods
Studyspecies
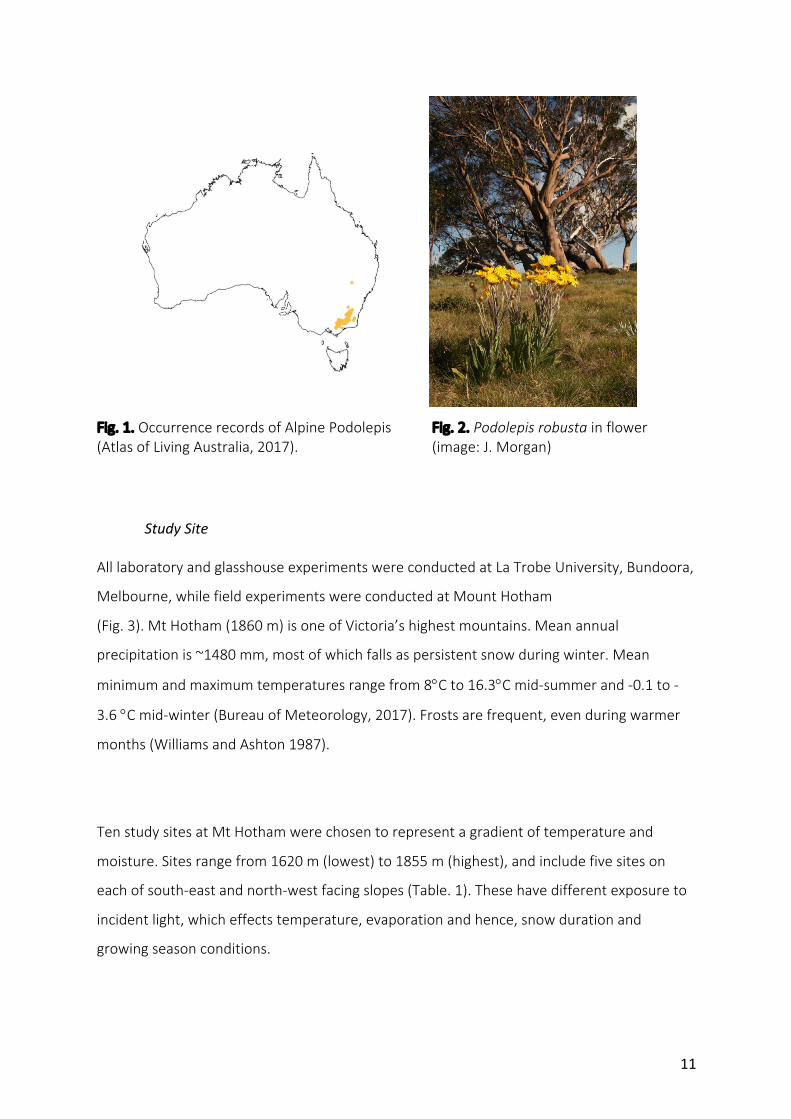
Podolepisrobusta(Maiden&Betche)J.H.Willis,commonlyknownasAlpinePodolepis,isa
small(20–60cm)erectperennialdaisy(Jeanes1999).AlpinePodolepisisrestrictedto,and
reasonablycommonthroughouttheAustralianAlpsBiogeographicRegion(Frood2015)(Fig.
1).Itgrowslargelyinassociationwithsnowgumwoodlandandalpinemeadowsattheupper
limit,orontheedgeofinvertedtree-linesassociatedwithcoldairdrainage,ataltitudes
between1200–1700m(Jeanes1999;Frood2015).AtMtHotham,plantsarelargely
associatedwiththewarmanddrynorth-westaspect(J.Morgan,unpublisheddata).
TheabundanceofAlpinePodolepiswasoncereducedatleastinpartthroughcattlegrazing
upuntilthe1950s(VanRees1982;Wahrenetal.1994),adisturbancewhichhassince
ceased(Wahrenetal.1994).Theeffectoffireisnotthoughttohavelong-lastinginfluences
ontheabundanceofAlpinePodolepisas,likemanyalpineforbsandgrasses,itrespondsby
resproutingrapidlypost-fire(Wahrenetal.2001).Inearlyspring,multipleshootsarisefrom
abasalrosetteofglabrous,lightgreenleaves(Frood2015).AlpinePodoelpisflowersin
summer(Dec-Mar)andarecharacterisedbybrightyellowcapitula(20-35mmindiameter),
clusteredindensecymes(Jeanes1999)(Fig2.).Comparedtootheralpinedaisies,seedsare
heavy(0.69mg±0.01mg)(Sommervilleetal.2013).Thoughpresumablywinddispersed,
thereiscurrentlynoexperimentalevidenceconcerningtheefficacyofwinddispersalby
AlpinePodolepis.
11
Fig.1.OccurrencerecordsofAlpinePodolepis Fig.2.Podolepisrobustainflower(AtlasofLivingAustralia,2017). (image:J.Morgan)
StudySite
AlllaboratoryandglasshouseexperimentswereconductedatLaTrobeUniversity,Bundoora,
Melbourne,whilefieldexperimentswereconductedatMountHotham
(Fig.3).MtHotham(1860m)isoneofVictoria’shighestmountains.Meanannual
precipitationis~1480mm,mostofwhichfallsaspersistentsnowduringwinter.Mean
minimumandmaximumtemperaturesrangefrom8°Cto16.3°Cmid-summerand-0.1to-
3.6°Cmid-winter(BureauofMeteorology,2017).Frostsarefrequent,evenduringwarmer
months(WilliamsandAshton1987).
TenstudysitesatMtHothamwerechosentorepresentagradientoftemperatureand
moisture.Sitesrangefrom1620m(lowest)to1855m(highest),andincludefivesiteson
eachofsouth-eastandnorth-westfacingslopes(Table.1).Thesehavedifferentexposureto
incidentlight,whicheffectstemperature,evaporationandhence,snowdurationand
growingseasonconditions.

12
Siteconditions
Airtemperatureat5cmabovegroundduringstudyperiod(Nov2016–Mar2017)was
measuredbi-hourlywithThermochroniButtons(MaximIntegrated,California,USA)attached
toawoodenstakeateachsite.Fromthesedata,growingdegreedays(GDDs)were
calculated(McMasterandWilhelm1997)asanindicatorofthermalaccumulation(in°C).
GDDsweresummedforeachsiteinordertorepresentacumulativeindexofavailableenergy
forgrowthanddevelopmentofseedlingsduringthestudyduration,accordingtothe
formula:
GDDs=[(TMAX+TMIN)/2]–TBASE,
whereTMAXandTMINarethedailymaximumandminimumtemperatures,respectively,and
TBASE=thetemperatureabovewhichmetabolicfunctionispossible.0°Cwaschosenasthe
basetemperature,asperBrownetal.(2006)andVennetal.(2013).
Ateachsite,lightinterceptionbydominantvegetationwasmeasuredwithaquantumlight
sensor(LI-COR,Nebraska,USA).Photosyntheticallyactiveradiation(PAR)wasmeasureda
totaloftentimesaboveandbelowthecanopyofvegetationalonga30mtransectat3m
intervals.Alongthesametransect,vegetationstructurewasmeasuredat1mintervals,
wherebycanopyinterceptionofherbs,graminoidsandshrubsalongastructurepolewas
recordedwithinheightclassesof10cmincrements.

13
Fig.3.LocationofMtHothaminVictoria,andstudysitelocationsatMtHotham(1-10)representingtheelevationgradientonsouth-eastandnorth-westsites(seeTable.1).Table1.SummaryinformationforthetensitesalongtheMtHothamtransectincludinglocation,altitudeandaspectforeachsite.(UTM=UniversalTransverseMercatorcoordinatesystem).
Site Location(UTM55H) Altitude(m) Aspect1 511370E,5907898N 1855 NW2 511433E,5907823N 1860 SE3 511218E,5907696N 1820 NW4 511297E,5907589N 1800 SE5 510990E,5907580N 1780 NW6 510924E,5907434N 1690 SE7 510900E,5907495N 1700 NW8 510816E,5907296N 1660 SE9 510792E,5907329N 1670 NW10 510783E,5907128N 1620 SE

14
Fieldtransplantexperiment
TodeterminethecontrolsonAlpinePodolepisseedlingsurvivalandgrowth,beyondthe
boundariesofitscurrentdistribution,atransplantexperimentwasundertakenatMt
Hotham.Theeffectsofaltitude,aspectandcompetitionwereexamined.
SeedwasgerminatedfromseedcollectedatMtHothaminMarch,2016.Seedwassurface
sownontoamixtureofsterilisedperlite,sandandpottingmixinOctober2016ina
glasshouse.Propagatedseedlingswerethenprickedoutintotrayswith4X4cmsoilplugs
andgrownfor6weeksunderglasshouseconditions.PriortoplantingatMtHotham,
seedlingswereplacedoutsidefor7daysatBundoora,andwerethentransportedtoMt
Hotham,acclimatedfor5days,andthenplantedovera3-dayperiod.Seedlingswerekeptin
theirtraysduringtransportationtoeachsite.Inmid-November2016,eachsitereceived26
seedlings,oneintoeachgap(cleared)andcontrolplot,arrangedalonga26mtransectwith
alternatingtreatmentsateachmetreinterval.Clearedplotswerecreatedfor‘gap’treatment
byremovingabove-groundvegetationfromwithin~15cmofthetransplantsitewith
secateursandweremaintainedmonthly.Thegaptreatmentswereprobablyconservative.
Seedlingsplantedintogapswerelikelystillgivensomeshelterfromwindsbyresident
vegetation.Atplanting,thenumberofleaves,andlengthoflongestleafwasrecorded.These
measurementswererecordedagainatthefinalsurveyinMarch2017.Seedlingsurvivalwas
recordedmonthly.
Laboratorygerminationexperiment
TodeterminewhetherseedgerminationofAlpinePodoelpishasanarrowverseswide
germinationniche,seedswereexposedtodifferenttemperatureandlightavailability
regimesinalaboratorygrowthcabinetstudy.
Treatmentsspannedarangeofdiurnaltemperaturesthatrepresentspring/autumn
conditions(14/10°C),earlytomid-summerconditions(20/12°C,24/16°C)andextreme
mid-summerconditions(30/20°C).Eachtreatmentwasconductedwith12hrlight/dark
cycles(ThermolineGrowthCabinets).Withineachtreatment,seedswereeitherexposedto

15
light/darkcyclesorwerekeptinconstantdarkconditionsbywrappingpetridishesintwo
layersofaluminiumfoil.Anadditionaltreatmentat4°Cwasimplementedfortwosetsof
replicatesinconstantdarkconditions.After30days,if0%germinationwasscored,replicates
weremovedintogrowthcabinetat14/10°Candplacedintorespectivelight/darkand
constantdarkconditions.
Foreachtreatment,therewerefivereplicates,eachconsistingof25seedsplacedinto90
mmpetridishesonWhatman#1filterpapermoistenedwithdistilledwaterandsealedwith
Parafilmtopreventlossofmoisture.Thesereplicateswereplaced,stacked,intogrowth
cabinets.Theorderofstackeddisheswasrandomisedafterscoring.
Replicateswerescoredforgerminationapproximatelyevery4daysforlight/darktreatments,
andapproximatelyevery8daysforreplicatesinconstantdarktreatmentsinorderto
minimizelightexposure.Seedwasconsideredgerminatedoncetheradiclehademerged
fromtheseedcoat,andwereremovedafterscoring.Filterpaperwasre-moistenedwhen
required.Once12dayspassedwithnonewgerminationfromwithinatreatment,remaining
seedsweretestedforviabilityusingtetrazoliumtesting(Moore1973).Attheendofthe
experiment,germinationpercentageswereadjustedforviability.
Lightlimitationexperiment
Theeffectoflightavailabilityonseedlinggrowthwasexaminedinaglasshouseexperiment.
Inthisexperiment,netphotosynthesisandbiomassaccumulationwereassessedinrelation
tothreelightregimes.Thetreatmentswere:(i)0%shade(treatedwithoutshadecloth);(ii)
48%shade,(iii)82%shade.PhotosyntheticallyActiveRadiation(PAR)intheglasshouseona
cleardayatmiddaywasapproximately1500µmolm-2s-1.
Treatmentswereconstructedusingcommercialknittedshadeclothattachedtothesidesand
topofarectangularenclosureconstructedof2.5mmfencewireandchickenwirenetting.
Seedwassurfacesownontoamixtureofsterilisedperlite,sandandpottingmix.Propagated
seedlingswerethenprickedoutinto40x100mmpotscontainingsterilisedpottingmixand

16
grownforsevendaysforanacclimationperiodandtocontrolformortalityduetotransplant
shock.Duringtheacclimationperiod,seedlingsweregrownunderfullambientlightinthe
glasshouseandirrigatedatregularintervals.Afterthisperiod,threeseedlingswereallocated
atrandomtoeachtreatmentofwhichtherewerefivereplicates.Thedry-weightofboth
aboveandbelowgroundbiomassof15seedlingswasmeasuredatthecommencementof
experimentaltreatments.Treatmentreplicateswererepositionedweeklytocontrolfor
glasshouseeffects,andseedlingswereirrigatedatregularintervals.
Imeasuredfoliargasexchangewithadifferentialinfraredgasanalyser(IRGA)(PP-Systems,
Massachusetts,USA),70to80daysafterseedlingswereinstalledintheirrespectivelight
regimes.Measurementsweretakenunderambienttemperatureconditions,whileCO2was
controlledforeachreplicateat400ppm.Measurementswererestrictedtothehours
between11:45amand1:45pmoncleardaystoreducetheinfluenceofpassingcloudand
sunposition.Fiverecordingsweretakenmanuallyonthreefullyexpandedleavesfromeach
seedling.EachleafwasinsertedinsidetheleafcuvetteoftheIRGA,whererecordsofnet-
photosynthesis,stomatalconductance,photosyntheticallyactiveradiation(PAR)intensity,
leafhumidityandtemperaturewereobtained.Atthetimeoffinalrecording,allseedlings
wereharvestedforaboveandbelowgroundbiomass,washedtoremoveexcesssubstrate
anddriedat80°Cfor48hinordertoobtainmeasurementsofdry-weightbiomass.
Freezingresistanceexperiment
Toassessthesensitivityofseedlingstoextremecold,theeffectsoffreezingtemperatures
wereassessedinthelaboratoryusingchlorophyllfluorescenceandthefluorescenceyieldof
PhotosystemII.
PlantmaterialforfreezingresistanceanalysiswascollectedfromthepopulationatMt
Hothaminmid-March2017.Duringcollection,15wholerosetteswereharvestedandsealed
inpolyethylenebags,andtransportedtothelaboratoryinMelbourneinachilledportable
cooler.Uponarrivalatthelaboratory,plantmaterialwaswrappedindamppapertowelin
ordertomaintainhydrationofplantmaterial,andstoredat4°Cinthedarkfor24htoallow

17
forcoldacclimation.Followingpre-treatment,threeleavesfromeachindividualwere
subjectedtooneoffourtemperaturetreatments(-5°C,-11°C,-20°C)for8husing
thermostaticallycontrolledfreezers.Similarly,controlsampleswereheldat4°C.Following
thefreezingtreatment,plantmaterialwaswrappedindamppapertowel,sealedin
polyethylenebagsandplacedinthedarkinaconstanttemperaturefacilityat18°C.
Theratioofvariabletomaximumfluorescenceoftheplantmaterial(FV/FM)wasthen
determinedafterthreedaysusingaPAMchlorophyllfluorometer(Walz,Effeltrich,
Germany).FV/FMratiosarereportedalongsidecontrol(4°C)samples,providinganestimate
ofphotosystemIIefficiencyduringtheassayprocedure.
Theaboveprocesswasalsoexecutedforseedlings.SeedcollectedfromMtHothamin2016
wassurfacesownonamixtureofsterilisedperlite,sandandpottingmix.Propagated
seedlingswereprickedoutandtransplantedinto40x100mmpotsfilledwithpottingmix
andgrownforthreeweeksunderglasshouseconditions.Afterthisperiod,seedlingswere
washedtoremoveexcesssubstratefromroots.Seedlingrootswereretainedduringfreezing
resistancetreatments.Ratherthanassessingfreezingdamagetothreeleavesperplant(as
withthematureplants),oneleafwasassessedfrom15replicateseedlingsforeach
treatmentduetothesmallnumberandsizeofleaves.
DataAnalysis
Fieldtransplantexperiment
Prediction:Ifthedirectionofplantinteractionchangesamongseedlingsalongatemperature
andmoisturegradient,Iwouldexpecttoseehighersurvivalingapsrelativetocontrolsatlow
altitudeswherecompetitiveprocessesaremorelikely,switchingtohighersurvivalincontrols
relativetogapsathigheraltitudes,wherefacilitativeprocessesbecomemorecommon
(Callaway1995).Additionally,ifthecurrentdistributionofAlpinePodolepisiscontrolledby
climate,Ipredictthattherewouldbelowersurvivalonthesouth-eastaspect.Therefore,in
themodelling,treatment,altitudeandaspectwereconsideredasfixedeffects.Totest
whetherseedlingsurvivaldifferedamongfixedeffects,generalisedlinearmodels(GLM)were

18
fitusingtheLaplaceapproximationinthepackagelme4(Batesetal.2015)inthe‘R’
statisticalpackage(RCoreTeam2013).ModelselectionwasbasedontheAkaikeinformation
criterion(AIC)(Akaike1974).
Similarly,plantperformance(growth)isexpectedtochangeaccordingtotreatment(gapor
control)alonganaltitudinalgradient.Thenetdirectionofpositiveandnegativeinteractions
wascalculatedbetweenpairsoftransplantedseedlingsingapandcontrolplotsusingthe
RelativeNeighbourEffect(RNE)Index(Brookeretal.2005;Vennetal.2009):
RNE=(Gap–Control/MAX(Gap,Control),
whereGapandControlcorrespondtotheperformanceofindividualsingaps(neighbours
removed)andcontrols(neighboursleftintact),respectively,andMAX(Gap,Control)refersto
thehighestperformancevaluebetweenthetwotreatments.Lengthoflongestleafwasthe
indiceusedtoquantifyseedlingperformanceandwascalculatedasfinallength/initial
lengthuponthefinalsurveyinMarch2017.RNEvaluesweremodifiedasperCallawayetal.
(2002)suchthatnegativevaluesindicatenegativeneighboureffectsonperformance
(competition),andpositivevaluesindicatepositiveneighboureffectsonperformance
(facilitation).PresentedaremeanRNEforseedlingsinpaired‘gap’and‘control’plots.Inthe
analysis,adeathinanypairofseedlingswasremovedfromfinalcalculations.Thismeantthat
inonecase,onthesouth-eastaspect,asitewasremovedfromanalysisduetolow
replicationofpairedseedlings.Inaddition,asiteonthenorth-westaspectwascouldnotbe
analysedduetomissingdata.RNEvalueswereanalysedwithone-wayANOVAaccordingto
sitewherebythelowestsiteoneachaspectwassetasthe‘dummy’thatallothersiteson
thatcorrespondingaspectwerecomparedto.North-westandsouth-eastaspectswere
analysedseparatelyinordertoteaseoutthedirectionofplantinteractionbyaltitudealone
withoutpotentialconfoundingfactorofaspect.
Lightlimitationexperiment
Prediction:IfcompetitionforlightisakeyfactorlimitingthedistributionofAlpinePodolepis,
thenIwouldexpecttoseereducedrateofphotosynthesisandreducedbiomasswith

19
decreasinglightavailability.Rateofphotosynthesisanddry-weightbiomasswerefittedin
separategeneralisedlinearmixedeffectsmodels(GLMM)usinglme4(Batesetal.2015)in
the‘R’statisticalpackage(RCoreTeam2013).Inthefirstinstance,shade-housereplicate,
plant,andleafwereusedasrandomfactors,andseedlingwasusedasarandomfactorinthe
latter.
Laboratorygerminationexperiment
Prediction:Ifspecificgerminationrequirementsarelimitingfactorsinthedistributionof
AlpinePodolepis,Iwouldexpecttoseeanarrowrangeofgermination.
TheinteractionbetweenlightandtemperatureonthegerminationofAlpinePodolepis
seedlingswasanalysedwithageneralisedlinearmodel(GLM)usinglme4(Batesetal.2015)
inthe‘R’statisticalpackage(RCoreTeam2013).Aquasi-binomialerrordistributionwasused
inordertoaccountfortheover-dispersionevidentingerminationdata(clusteringofbinary
outcomeswithintreatments).Finalgerminationofseedsindarkandlighttreatmentsat14°C
werecomparedtothosethatreceivedaperiodofcold-stratificationusingStudent’st-tests.
Freezingresistanceexperiment
Theextentoftissuedamageinresponsetofreezingtemperatures(-5°C,-11°C,-20°C)was
analysed.Ifrecruitmentdynamicsareinfluencedbyoccurrencesoffreezingtemperatures,
seedlingtissueisexpectedtoexperiencehigherdamagerelativetocontrol(4°C)andtoadult
tissue.Student’st-testswereusedforsimplecomparisonsbetweenFV/FMratiosofadultand
seedlingleaves,asagreatertemperaturerangeisrequiredinordertocalculatelevelof
‘freezingresistance’bylinearinterpolationasperBannister(2005)andVennetal.(2013).
Lethaltemperaturesareconsideredthoseatwhich50%damageoccurstothe
photosyntheticapparatusofleavesrelativetocontrol(4°C).
20
Results
Siteconditions
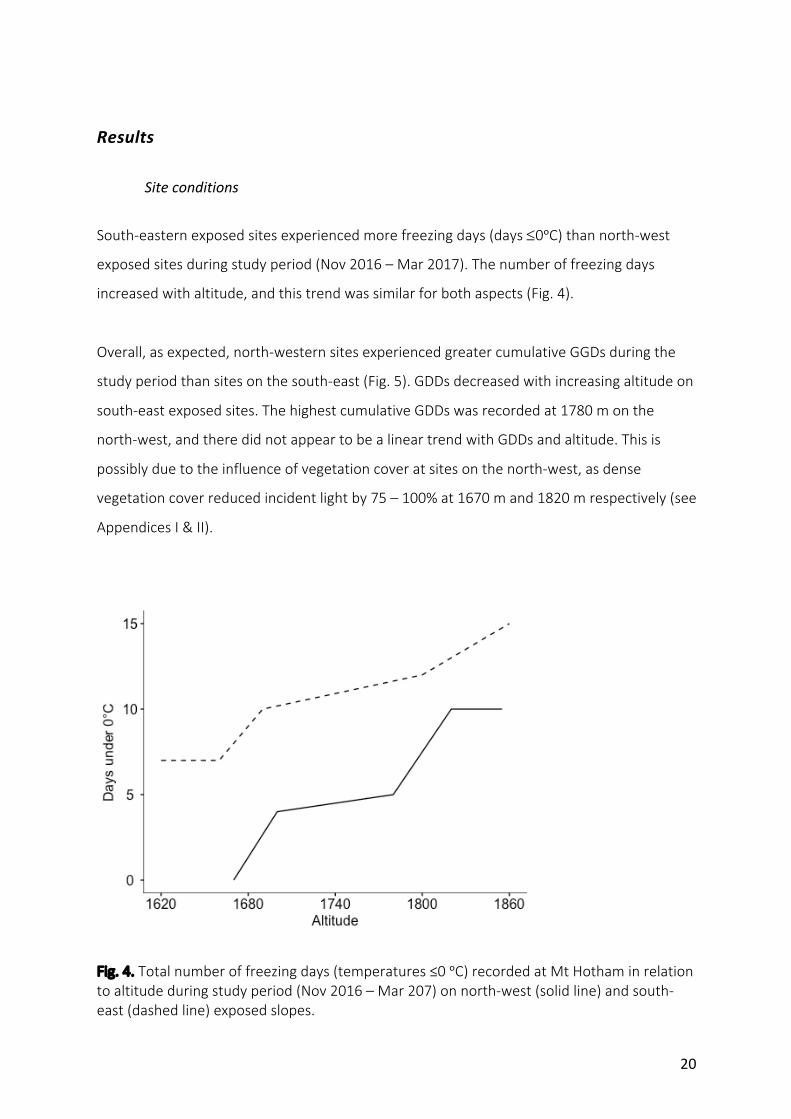
South-easternexposedsitesexperiencedmorefreezingdays(days£0ᵒC)thannorth-west
exposedsitesduringstudyperiod(Nov2016–Mar2017).Thenumberoffreezingdays
increasedwithaltitude,andthistrendwassimilarforbothaspects(Fig.4).
Overall,asexpected,north-westernsitesexperiencedgreatercumulativeGGDsduringthe
studyperiodthansitesonthesouth-east(Fig.5).GDDsdecreasedwithincreasingaltitudeon
south-eastexposedsites.ThehighestcumulativeGDDswasrecordedat1780monthe
north-west,andtheredidnotappeartobealineartrendwithGDDsandaltitude.Thisis
possiblyduetotheinfluenceofvegetationcoveratsitesonthenorth-west,asdense
vegetationcoverreducedincidentlightby75–100%at1670mand1820mrespectively(see
AppendicesI&II).
Fig.4.Totalnumberoffreezingdays(temperatures≤0ᵒC)recordedatMtHothaminrelationtoaltitudeduringstudyperiod(Nov2016–Mar207)onnorth-west(solidline)andsouth-east(dashedline)exposedslopes.

21
(a)
(b)
Fig.5.GrowingDegreeDays(GDD)recordedforsouth-east(a)andnorth-west(b)exposedsitesduringstudyperiod(Nov2016–Mar2017).
1350
1450
1550
1650
1750
1670 1700 1780 1820 1855
GDD
Altitude
1350
1450
1550
1650
1750
1620 1660 1690 1800 1860
GDD
Altitude
22
Fieldtransplantexperiment
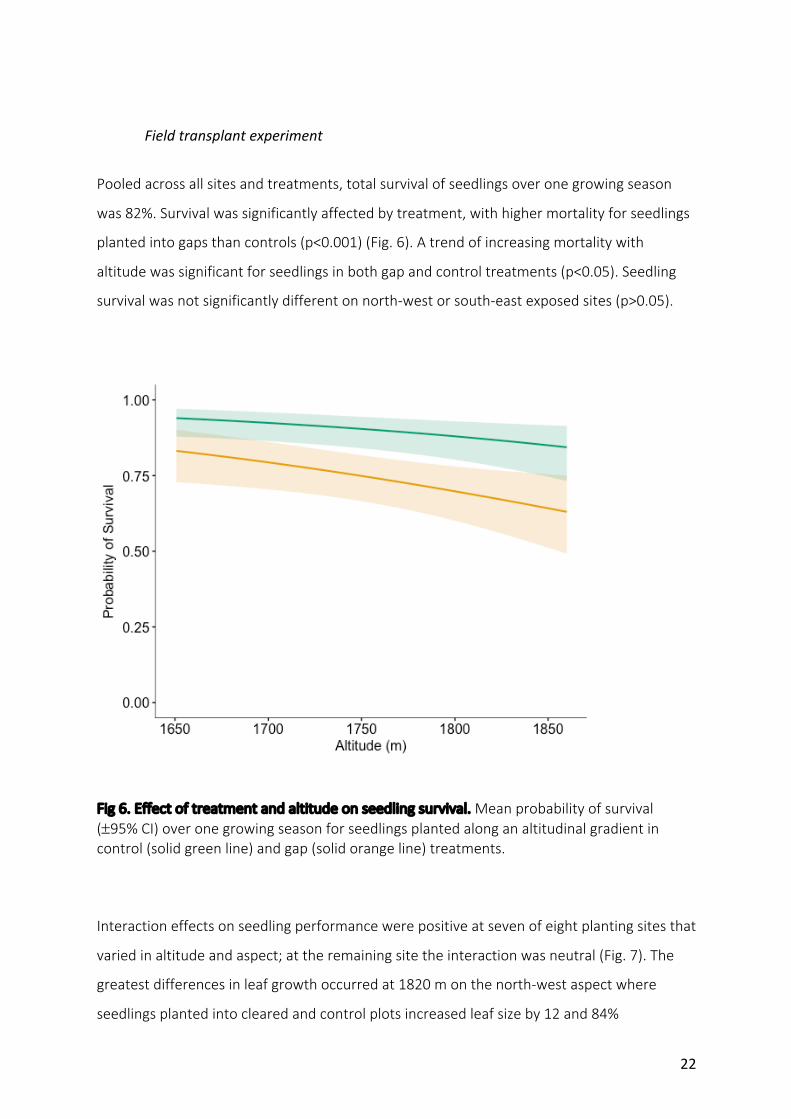
Pooledacrossallsitesandtreatments,totalsurvivalofseedlingsoveronegrowingseason
was82%.Survivalwassignificantlyaffectedbytreatment,withhighermortalityforseedlings
plantedintogapsthancontrols(p<0.001)(Fig.6).Atrendofincreasingmortalitywith
altitudewassignificantforseedlingsinbothgapandcontroltreatments(p<0.05).Seedling
survivalwasnotsignificantlydifferentonnorth-westorsouth-eastexposedsites(p>0.05).
Fig6.Effectoftreatmentandaltitudeonseedlingsurvival.Meanprobabilityofsurvival(±95%CI)overonegrowingseasonforseedlingsplantedalonganaltitudinalgradientincontrol(solidgreenline)andgap(solidorangeline)treatments.
Interactioneffectsonseedlingperformancewerepositiveatsevenofeightplantingsitesthat
variedinaltitudeandaspect;attheremainingsitetheinteractionwasneutral(Fig.7).The
greatestdifferencesinleafgrowthoccurredat1820monthenorth-westaspectwhere
seedlingsplantedintoclearedandcontrolplotsincreasedleafsizeby12and84%

23
respectively,hencethepositiveRNEvalue.Seedlingsat1670mshoweda153%increaseof
leafsizeinclearedplotsanda110%increaseofleafsizeincontrolplots,hencetheneutral
RNEvalue.NostatisticaldifferencesbetweenRNEvaluesweredetectedforplantedseedlings
onthesouth-eastsites;rather,positiveinteractionswithcloseneighbourswereconsistentat
eachsitealongtheelevationgradient.Onthenorth-westaspect,theneutralinteractionat
1670mdifferedsignificantlyto1820m(p<0.01)and1855m(p<0.05)wheregrowthwas
positivelyinfluencedbycloseneighbours(Fig.7a;AppendixIV,Table.6).
(a) (b)
Fig.7.MeanRelativeNeighbourEffect(RNE)(±1SE)forpairedtransplantedseedlingsingapandcontroltreatmentson(a)north-westand(b)andsouth-eastexposedaspectsacrosstheelevationgradient.Positivevaluesindicatefacilitativeinteractions,andnegativevaluesindicatecompetitiveinteractions.Neutralinteractionsareindicatedbyerrorbarscrossingtheyaxisat0.Differentsuperscriptsabovecolumnsdenotesignificantdifferencesbetweensites.NPlabelsindicatethatanalysiswasnotpossible.

24
Laboratorygerminationexperiment
Temperature-mediateddormancyisweak,withhighgermination(>50%)acrossalldiurnal
temperatureandlightconditions,andonlyconstrainedatthelowesttemperatureinthedark
(Fig.8).Therewasasignificantinteractionbetweenlightanddarktreatmentswith
temperature(p<0.05),asgerminationissuppressedindarkconditions(p<0.01),but
increaseswithtemperature(p<0.001),whilegerminationoccurredequallywellatall
temperatureregimesinlightconditions(14°C–30°C).Timeto50%germination(t50)
occurredwithoutstratificationandwithin30daysforbothlightanddarkconditions³20°C.
At14°Cinthedark,responsetocoldstratificationwaspositive(p<0.05),increasing
germinationby21.8%andreducingT50byatleast42days(Table2).Inthelighttreatment,
T50decreasedonlymarginallyby3daysaftercoldstratification.
(a) (b)
Fig.8.Effectoflightandtemperatureongermination.Fittedvaluesforprobabilityofgerminationfor(a)lightconditionsand(b)darkconditions(±95%CI)forthegeneralisedlinearmodelappliedoncabinetgerminationdata.Germinationpercentagesfromrawdataincludedaspoints.Notethedifferentscalesfortemperatureaxis.

25
Table2.GerminationattributesofAlpinePodolepisat14°Cwithandwithoutaperiodofcoldstratification.LAG(days)=thenumberofdaysfromthestartoftheexperimentuntilgerminationbegan.t50(days)=thenumberofdaysfor50%oftotalgerminationtooccur.Lighttreatment Coldstratification
periodLAG(days) t50(days)
Final%
germinationLight 30days 4-8 8-11 96.8Light 0days 7-10 14-20 99.2Dark 30days 11-14 13-23 83.8Dark 0days 10-13 62-65 62
Lightlimitationexperiment
Bothseedlingdrymass(aboveandbelowgroundbiomass)andnetphotosynthesis
respondedsimilarlytolightlimitationtreatments(Fig9.10.).Valueswerecomparablefor
seedlingsgrowninfullsun(0%shade)andat48%shadeforseedlingbiomass(p>0.05)and
netphotosynthesis(p>0.05).However,plantbiomass(Fig.10)wasreducedsignificantlyin
seedlingsgrownunder82%shadewhencomparedtoseedlingsgrowninfullsun(0%)
(p<0.001)and48%shade(p<0.001).Similarly,netphotosynthesis(Fig.9)wassignificantly
lowerforseedlingsgrownat82%shadewhencomparedtoseedlingsgrowninfullsun
(p<0.01)and48%shade(p<0.05).

26
Fig.9.Effectoflightlimitationonnetphotosynthesis.Fittedmeanrateofphotosynthesis(±95%CI)ofseedlingsgrownunderfullsun,48%shadeand82%shade.
Fig.10.Effectoflightlimitationonbiomass.Fittedmeandry-weightofbiomass(±95%CI)ofseedlingsgrownunderfullsun,48%shadeand82%shade.
27
Freezingresistanceexperiment
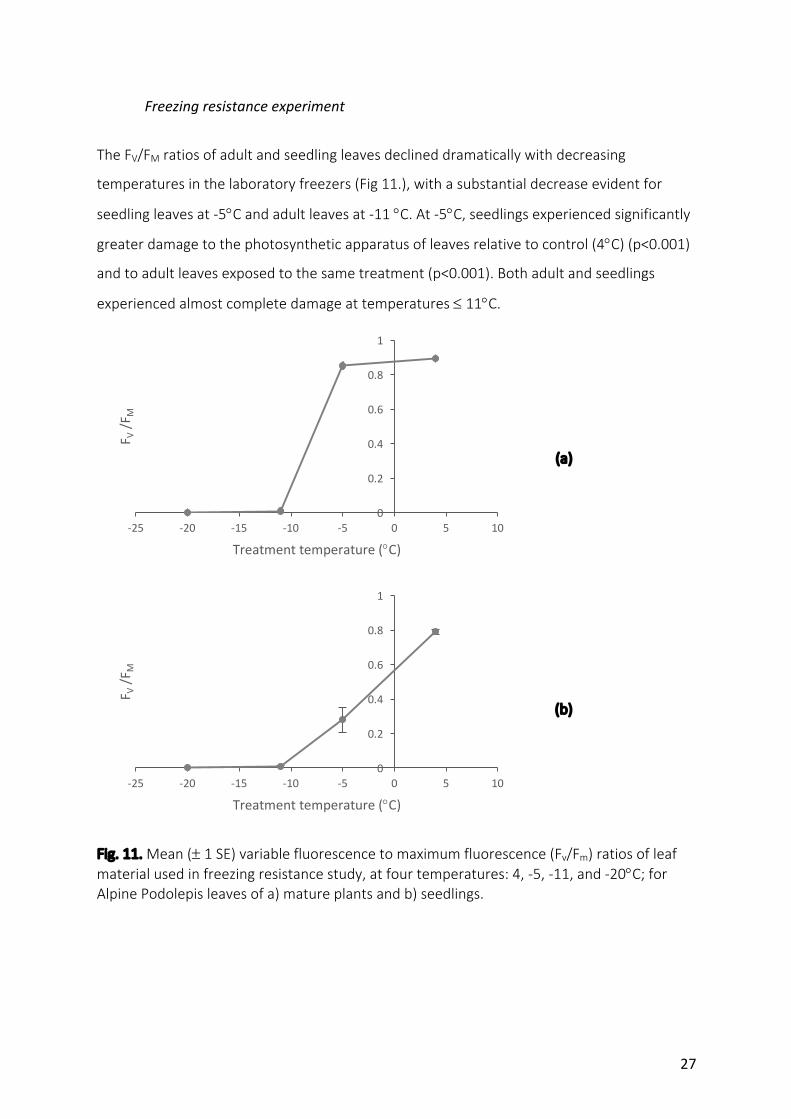
TheFV/FMratiosofadultandseedlingleavesdeclineddramaticallywithdecreasing
temperaturesinthelaboratoryfreezers(Fig11.),withasubstantialdecreaseevidentfor
seedlingleavesat-5°Candadultleavesat-11°C.At-5°C,seedlingsexperiencedsignificantly
greaterdamagetothephotosyntheticapparatusofleavesrelativetocontrol(4°C)(p<0.001)
andtoadultleavesexposedtothesametreatment(p<0.001).Bothadultandseedlings
experiencedalmostcompletedamageattemperatures£11°C.
Fig.11.Mean(±1SE)variablefluorescencetomaximumfluorescence(Fv/Fm)ratiosofleafmaterialusedinfreezingresistancestudy,atfourtemperatures:4,-5,-11,and-20°C;forAlpinePodolepisleavesofa)matureplantsandb)seedlings.
0
0.2
0.4
0.6
0.8
1
-25 -20 -15 -10 -5 0 5 10
F V/F
M
Treatmenttemperature(°C)
0
0.2
0.4
0.6
0.8
1
-25 -20 -15 -10 -5 0 5 10
F V/F
M
Treatmenttemperature(°C)
(a)
(b)

28
Discussion Facilitative(positive)interactionsinfluencedthesurvivalandgrowthofseedlingsoverone
growingseasonatMtHothamintheVictorianAlps.Seedlingsurvivalwasnotinfluencedby
aspect,butatrendofincreasingmortalitywithaltitudewasdetected.Therateof
photosynthesisandbiomassaccumulationmayalsoberestrictedinthefielddueto
competitionforlightinheavilyshadedsites,assuggestedbyglasshouseexperiments.
Germinationwasobservedinthelaboratorytobebroad,attemperatures³14°Cinlightand
darkconditions.Whilecoldstratificationwasnotrequired,theresponsetostratificationwas
positiveindarkconditions,wherehigherpercentgerminationandfastergerminationrates
likelyreflectssynchronousgerminationinthefieldaftersnowmelt.Youngseedlingssuffered
greaterdamagetofreezingtemperaturesthanadultplants.Asfreezingtemperaturesare
commonandunpredictableinthealpinezone,vulnerabilityattheemergingseedlingsstage
maybealimitingfactorinrecruitmentanddistribution.Resultssuggestthatwhatis
observedtodayastherealisednicheofAlpinePodolepisisnotreflectiveofitspotential
range,andhighlightstheimportanceoffacilitativeinteractionsinalpineenvironments.
Germinationcharacteristics
Germinationisoneofthemostfundamentaldevelopmentalprocessesinthelifecycleof
plants,andvarieswidelyamongspecies(Korner1999).AlpinePodolepisexhibitedhigh
germinationatabroadrangeoftemperatures;however,itwassuppressedinthedarkat
4°C.Responsetocoldstratificationwaspositive,increasingrateofgerminationandfinal
germinationinthedark.Thoughlong-termdormancyisconsideredrareinalpinelandscapes,
somealpinespeciesrequiretheexperienceoflowtemperaturesoverwinterinorderto
germinate(BaskinandBaskin2003).ThisisthecasewithAustralianalpinespeciesAciphylla
glacialis(VennandMorgan2009),Euphrasiacollinassp.(Sommervilleetal.2013),and
Eucalyptuspauciflora(BeardsellandMullett1984),whereunlessseedsaresubjectedtoa
periodofcold-wetstratification,germinationispoorornotpossible.Alackofdormancy
(excludingafterripening),andawidegerminationrangeindicatesthatAlpinePodolepisare

29
notdependentuponaspecifictemperatureregimeforgermination,acommontrait
throughoutAsteraceae(BaskinandBaskin2003;Sommervilleetal.2013).
Positiveresponsestocoldstratificationhavebeenreportedforawidearrayofalpinespecies,
andinsomecaseswidensthegerminationrangeandimprovesfinalgermination
(Sommervilleetal.2013).Hence,thoughgerminationwasreducedindarkconditions,thisis
likelyovercomeinsitubyacoldperiodexperiencedoverwinter.Coldstratificationalso
reducesvariabilityingerminationtime,synchronisingseedlingemergenceaftersnowmelt
(ShimonoandKudo2005).Assuch,itisunlikelythatAlpinePodolepisarecapableofstoring
dormantseedinthesoil.ThiscontrastswithCarexspecieswheregerminationeventsare
spreadovertwoyears,reducingtheriskofcompleterecruitmentfailure(Schütz2002).
Hence,whileitisunlikelythatthedistributionofAlpinePodolepisiscontrolledbynarrow
germinationrequirementsseeninotheralpinespecies,anopportunisticandlikely
synchronousgerminationinearlyspringindicatesthatrecruitmenthingesupontheamount
ofseeddispersedinthepreviousseason,andtheenvironmentalconditionsupon
germination.
Frosttoleranceinseedlings
GivenrequirementsforgerminationarerelativelybroadforAlpinePodolepis,limitstoits
distributionmustbecontrolledbyfactorsotherthananarrowgerminationniche.Indeed,
whilegerminationiscommonintheVictorianalpinezone,halftheamountofnaturally
emergingseedlingsdiesbeforeestablishment(VennandMorgan2009).AlpinePodolepis
seedlingswerefoundtobemoresensitivetofreezingtemperaturesthanadults,suffering
significantdamagetophotosyntheticapparatusat-5°C.Cold-edgeboundariesaresetby
occurrencesoffrostduringthegrowingseason,coupledwithspecies-specificfreezing
resistance(abilitytosurvivefreezingtemperatureswithoutdamage)(Körneretal.2016).
Freezingtemperaturesareanimportantfilterongermination,recruitmentandpersistencein
alpineplantcommunities(KörnerandPaulsen2004)and,assuch,areprincipledeterminants
ofgeographicdistributions(SakaiandWeiser1973).FrostsintheAustralianAlpsare
common,evenduringwarmerseasons(WilliamsandAshton1987).Duringthisstudy,

30
freezingtemperatureswererecordedasbeingmorecommononsouth-easternaspects,and
increasingwithaltitude.Resistancetofreezingtemperaturesofalpineplantsoverthe
growingseasonisreportedlyquitehighinAustralianalpinespecies,withapatternof
increasingtolerancewithaltitude(Vennetal.2013).Whileadultsmaybelesssusceptibleto
springorsummerfrosts,seedlingshavelowertoleranceandlimitedrecuperationability,as
recentlyemergingseedlingsareyettodevelopmeristematicregionsunderground(Korner
1999).However,whileitisunknownwhenthethresholdforfrostresistanceisreachedfor
AlpinePodolepis,occurrencesoffrostduringseedlingemergence,andduringtheearly
stagesofestablishment,arelikelytoreducesurvival.Hence,seedlingestablishmentmay
thereforebelimitedtorelativelyrare,favourableseasons.However,successful
establishmentmaybepossible,givensuitablemicrosites(Vennetal.2009).
Effectsoffacilitationalonganabioticstressgradient
Duringthisstudy,seedlingmortalitywaslow,andwasnotaffectedbyaspect,contraryto
expectations,giventhecurrent,stronglynorth-westerndistributionofAlpinePodolepisatMt
Hotham.Whileseedlingmortalityincreasedslightlywithaltitude,mortalitywaslowacross
theentiregradientofbeyond-rangetransplants.Althoughouraltitudinalgradientwassmall
(235m),thechangesinenvironmentalconditionswerenoticeablewithmorefreezing
temperaturesrecordedonsouth-easternaspect,whichincreasedonbothaspectswith
altitude.Asindicatedbyourfreezingresistanceexperiment,earlyseedlingsurvivalisstrongly
affectedbyfreezingtemperatures.Ourplantedseedlingsweresixweeksoldattimeof
planting,likelyenoughtimeforseedlingstodeveloprootreservesandaleveloffrost
resistance,unlikeinnewlyemergentseedlings(Körner1999).Hence,ifconditionsduringthe
earlygrowingseasonallowforseedlingestablishment,seedlingsaretoleranttoarangeof
conditionsatMtHotham,contrarytowhatthecurrent,stronglynorth-westerndistribution
mightsuggest.Conclusionscanonlybebasedoffofonegrowingseason,andperhapsthe
strengthofaspectandaltitudemaybecomemoreapparentwithtime.
Incontrast,similarstudieshavereportedhighseedlingmortalityduringthefirstgrowing
season(UrbanskaandShutz1986;Vennetal.2009).Indeed,theseedlingstageiscriticalin

31
thelifecycleofplants(ForbisandDoak2004),andpresumably,highsurvivaloverasingle
growingseasonisindicativeofahightolerancetovariedclimaticconditionsthatwouldbe
experiencedalongthealtitudinalgradient.Inalpineenvironments,plantsareexposedto
stressfulabioticconditionsincludingfreezingtemperatures,needleice,andscouringwinds
(Körner1999).However,substantialvariationinmicrohabitatinfluencesthecompositionof
localvegetation(SherrerandKörner2011).Suchvariation,whiledrivenbytopography,
altitude,andslopeorientation(SherrerandKörner2011),isalsodrivenbyvegetation(Körner
1999;Vennetal.2009).
Whileseedlingsurvivalwasgenerallyhighatallsites,survivalwasconsistentlyhigherin
control(uncleared)plotsthaningaps.Numerousstudieshighlightfacilitationasanimportant
factorinseedlingsurvivalinnaturalcommunities(Callawayetal.1997;Brookeretal.2007;
Vennetal.2009:WarrenandBradford2010),asneighbouringplantsamelioratestressful
abioticconditions,creatingamicroclimatesuitableforseedlingestablishment(Callawayetal.
1997).Indeed,whilesomespeciesrequirebarepatchesforgerminationandestablishment,
seedlingsinalpineenvironmentsarecommonlyreportedclumpedtogetherintheleeof
largerneighbours(Callawayetal.2002).Insuchmicrosites,‘nurseplants’bufferestablishing
seedlingsfromscouringwinds,increaselocaltemperatureandsoilmoisture(Callaway1995)
andaidinthereductionofdisturbancessuchassoilfrostheave(Vennetal.2012).However,
unlikemanysimilartransplantstudieswhichtestthestressgradienthypothesis(Choleretal.
2001:Callawayetal.2002;Cavieresetal.2005),aswitchinthetypeofinteraction
(facilitationorcompetition)wasnotevident.Instead,consistentlypositiveinteractionswere
demonstratedatallsites,highlightingtheimportanceofpositiveinteractionsatoneof
Australia’shighestmountains.Perhapsthisisunsurprising,giventhegradientsinvestigated
byotherecologistsareoftengreaterandtransplantsaremonitoredovermorethanone
growingseason(seeCholeretal.2001:Alexanderetal.2015).However,Vennetal.(2009)
andAlexanderetal.(2015)reporteffectsofcompetitionduringthefirstgrowingseason.
Whilecompetitioncertainlyisanimportantfactorinthedistributionofspecies(Callawayet
al.1997;DunnettandGrime1999),itdoesnotappeartobeakeydriverofAlpinePodolepis
establishmentduringthefirstgrowingseason.

32
FacilitativeinteractionspositivelyinfluencedthegrowthofAlpinePodolepisseedlingsat
mostsitesoverthecourseofonegrowingseason,consistentwithseedlingsurvivalresponse.
Thisiscountertoexpectationsthatthedirectionofplantinteractionsshouldshiftalongan
abioticstressgradient(Callaway1995;Choleretal.2001;Callawayetal.2002).One
occurrenceofneutralinteraction(whereseedlinggrowthwasnotaffectedbythepresence
orabsenceofneighbours)wasreportedonthenorth-westaspectatthelowestsite.Here,
abioticconditionsdidnotnegativelyaffectseedlingsinclearedsitesasitdidelsewhere,a
signalofrelativelybenignconditions,whichissupportedbylocaltemperaturedata.Here,no
freezingtemperatureswererecordedduringthestudyperiod.Hadsitesextendedtoeven
loweraltitudes,competitiveinteractionsmayhavebecometheprevailinginteractionas
reportedelsewhere(Callawayetal.2002;Alexanderetal.2015).Thelackofcompetitive
interactionsatMtHothammaybeindicativeofabioticstressorsotherthantemperature.
Facilitativeinteractionsareoftenreportedduetolowavailablemoistureondryaspects
(Hillier1990)andheatstress(Korner1999).
PositiveinteractionsmaydominateforAlpinePodolepisduringearlylifestage.Indeed,the
patternsandphenomenaof‘nurseplant’syndromeareoftenstrongestduringearlylife
stages(Callaway1995).Competitiveinteractionsmaydominateoncetheplantreaches
maturity(Callaway1995)orduringmildseasonswhereabioticstressisreduced(Kikvidzeet
al.2006).Alongasimilaraltitudinalgradient,positiveinteractionsinfluencedthegrowthof
Brachyschomeridigulaseedlings,butthestrengthandbalanceofinteractionsshifted
temporally(Vennetal.2009).Withenvironmentalvariationoverthecourseofone,ormany
growingseasons,thedirectionandstrengthoffacilitationandcompetitionislikelytochange
inapredictablemanner(BertnessandCallaway1994;BrookerandCallaghan1998).Nurse-
plantsmaybecomeout-competedbybenefactorsorthereverse(Callaway1995;Kikvidzeet
al.2006),whilefavourableseasonalconditionsmaytipotherwisefacilitativeinteractionsinto
competitiveones(TielborgerandKadmon2000;Valiente-Banuet2008).Hence,while
positiveinteractionspredominatethegrowthandsurvivalofAlpinePodolepis,overthefirst
growingseason,thistrajectorymaychangethroughsubsequentseasons.
Effectsofcompetitiononseedlinggrowth

33
Themechanismsbywhichseedlingsinteractwithnaturalcommunitiesarenumerous,and
canonlybeseparatedbyexperimentalmanipulation(Brunoetal.2003).Theresultssuggest
thata“suitable”environmentforAlpinePodolepisdependsonlightavailability,andindicates
thatmoderateshadeisnotnecessarilynegativeforseedlinggrowth.Thelow-light
environmentthatoccursbeneaththestandofdominantPodolobiumalpestreatlow-altitude,
north-westernsitesforexample,wouldleadtodecreasednetphotosynthesisandbiomass.
Incontrast,high-lightorpartyshadedenvironmentsareequallyfavourableforgrowth.Such
lightconditionscanbefoundwithintherelativelywell-illuminatedunderstoryoftheupper
limitsofsnow-gumwoodlands,andthroughoutmuchofthetreelessalpinemeadows.
However,insituconditionsareusuallynotoptimal,andnetphotosynthesisisoftenbelowfull
capacityduetocombinationsofwaterstress,temperatureextremesorlowhumiditycausing
partialstomatalclosure(Muraokaetal.2002).Moreover,damagecausedbyextreme
temperatures,drought,orherbivoryalsomustincreasemaintenancecosts,anadded
constraintonnetgrowth(Körner1999).
Whilecertainabioticstressorscanbemediatedbymicrositescreatedintheleeorcanopyof
other,morestress-tolerantplants,astheplantreachesmaturity,competitionforresources
becomesmoreimportantbelow-ground(AguiarandSala1994).Wilson(1988)foundthat
rootcompetitionwasthemoreintenseformofplantinteraction,occurringdirectlythrough
allelochemicalexudates,andindirectlythroughresourcedepletion.IntheTibetanPlateau,
interactionsofrootcompetitionvarybetweenspecies(Songetal.2006).Hence,theeffects
ofcompetitionandmayplayoutinthelongertermforAlpinePodolepisseedlings.Indeed,
competitionmaybeactingalongthegradientinconcertwithfacilitation,suchthatweare
notyetabletoobservetheeffectsofaboveorbelow-groundcompetition.Howthisbalance
ofcompetitionandfacilitationwillplayoutthroughtimewillbeimportantforunderstanding
howinteractionsinfluencetherangeboundariesofalpineplants.
Asdemonstratedwithlaboratorygerminationandfieldtransplants,AlpinePodolepisappear
tohaveatheoreticalnichefargreaterthancanbeobservedatMtHothamcurrently,
indicatingpossiblelimitationofdispersaland/orpropaguleavailability.Itiscommonfor
plantstolose90-100%ofproducedseedtoseedpredation(Louda1989).Pre-and-postseed

34
predatorscandeterminenetfecundityoftheirplanthosts(Mckoneetal.1998),having
consequencesforpopulationdensity(Louda1982)anddistribution(BrownandVellend
2014).Pre-dispersalseedpredationiscommonamongalpineAsteraceaeandhasbeen
reportedforAlpinePodolepisinMtKosciuszko(Pickering,2009).However,theeffectsof
herbivoryandsoilpathogensaredifficulttodetermine.
Thereisapaucityofevidenceregardingtheimportanceofinsectsandpathogensinthe
populationdynamicsandrangelimitsofhostplants.Differentialratesofinsectorpathogen
attackmayalterorreversecompetitiverelationshipsbetweenplants(LeeandBazzaz1980;
RaiandTripathi1985).Indoingso,theyhavethepotentialtoexcludeplantspeciesfrom
communities(Crawley1989),andthereforemayplayamajorroleinthestructuringofplant
boundaries.Inaddition,plantresponsetoherbivoryislikelytovarywithlifestageandmay
interactwithresourceavailability.Seedlingsarelikelytohavelowerdefenseand/ortolerance
toattackthanestablishedplants(BoegeandMarquis2005)andmayalsoplayarolein
definingspeciesdistributions.Futureresearchmightconsiderhowdispersal,seedpredation
and/orherbivoryandpathogensmayinfluencepopulationdynamicsanddistributionof
plants,andhowtheseinteractionsmightchangewithcontinuedclimatewarming.
Conclusion
Typically,facilitativeinteractionsarereportedasoccurringmostlyattheextremelimitsofa
speciesenvironmentaltolerance,leadingtorangeexpansionandalteringtherealisedniche
(Brunoetal.2003).Here,residentplantspositivelyinfluencedsurvivalandgrowthofAlpine
Podolepisseedlingsbeyondthecurrent,observedrange.However,facilitationoccurrednot
onlyattheupperlimitstoitsdistribution,butalsobelowthetrailingedge.Habitat
ameliorationisanimportantstructuringforceinstressfulenvironments(Brunoetal.2003).
Forseedlingsurvivalinthealpinelandscape,thisistrueforAlpinePodolepis,regardlessof
theabioticstressgradient,duringthefirstgrowingseason.Althoughtheconsistenttrendof
positiveinteractionsmaychangeovertime,earlyseedlingestablishmentisacriticalstage
(Körner1999).Highoverallsurvivalindicatesawiderangeofphysiologicaltolerancealongan
abioticstressgradient.Highoverallsurvivalalsohighlightspossiblelimitationsindispersal

35
and/orpropaguleavailability.Inaddition,earlyseedlingsurvivalislikelyrestrictedby
occurrencesoffrostduringtheearlygrowingseason.Whilethereissomeevidenceofshade
intolerance,whichmayacttorestrictdistributionsatloweraltitudes,furtherexperimental
researchwillberequiredinordertobetterunderstandcompetitiveinteractionsinalpine
plants.Simultaneousinvestigationintopropaguleavailability,dispersalabilityandinsitu
recruitmentwillalsoaidinferencesontheroleofpopulationdynamicsondistributionand
migrationrates.Whetherconsistent,positiveinteractionsreportedherewillchangeover
timewillprovidefurtherinsightintothemechanismsofspeciesinteractions,andtheir
influenceonspeciesdistributions.

36
References
Afkhami,M.E.,McIntyre,P.J.,&Strauss,S.Y.(2014).Mutualist-mediatedeffectsonspecies'rangelimitsacrosslargegeographicscales.EcologyLetters,17(10),1265-1273.doi:10.1111/ele.12332
Aguiar,M.R.,&Sala,O.E.(1994).Competition,facilitation,seeddistributionandtheoriginofpatchesinaPatagoniansteppe.Oikos,26-34.
Akaike,H.(1974).Anewlookatthestatisticalmodelidentification.IEEETransactionsOnAutomaticControl,19(6),716-723.
Alexander,J.M.,Diez,J.M.,&Levine,J.M.(2015).Novelcompetitorsshapespecies'responsestoclimatechange.Nature,525(7570),515-518.doi:10.1038/nature14952
Alexander,J.M.,Lembrechts,J.J.,Cavieres,L.A.,Daehler,C.,Haider,S.,Kueffer,C.,.Liu,G.,McDougal,K.,Milbau,A.,&Pauchard,A.(2016).Plantinvasionsintomountainsandalpineecosystems:currentstatusandfuturechallenges.AlpineBotany,126(2),89-103.
AtlasofLivingAustralia(2017).Podolepisrobustaoccurrencerecordsonline.Availablewww.http://spatial.ala.org.au/?q=lsid:http://id.biodiversity.org.au/node/apni/2908666[Verified15March2017].
Austin,M.P.(2002).Spatialpredictionofspeciesdistribution:aninterfacebetweenecologicaltheoryandstatisticalmodelling.EcologicalModelling,157(2–3),101-118.doi:http://dx.doi.org/10.1016/S0304-3800(02)00205-3
AustralianBureauofMeteorology(2017).HothamHeightsclimatedataonline.Availablewww.bom.gov.au[Verified20April2017].
Bates,D.,Maechler,M.,Bolker,B.,&Walker,S.(2015).Fittinglinearmixed-effectsmodelsusinglme4.JournalofStatisticalSoftware,67(1),1-48.doi:doi:10.18637/jss.v067.i01.
Bannister,P.(2005).FrostresistanceoftheNewZealandnarrow-leavedsnowtussockgrass,Chionochloarigida.NewZealandJournalofBotany,43(2),425-430.
Baskin,C.C.,&Baskin,J.M.(2003).Whenbreakingseeddormancyisaproblemtryamove-alongexperiment.NativePlantsJournal,4(1),17-21.
Bertness,M.D.,&Callaway,R.(1994).Positiveinteractionsincommunities.TrendsinEcology&Evolution,9(5),191-193.
Bertness,M.D.,&Shumway,S.W.(1993).Competitionandfacilitationinmarshplants.TheAmericanNaturalist,142(4),718-724.
Bertrand,R.,Lenoir,J.,Piedallu,C.,Riofrio-Dillon,G.,deRuffray,P.,Vidal,C.,Pierrat,J.C.,&Gegout,J.C.(2011).Changesinplantcommunitycompositionlagbehindclimatewarminginlowlandforests.Nature,479(7374),517-520.doi:10.1038/nature10548
Boege,K.,&Marquis,R.J.(2005).Facingherbivoryasyougrowup:theontogenyofresistanceinplants.TrendsinEcology&Evolution,20(8),441-448.
Brooker,R.W.(2006).Plant–plantinteractionsandenvironmentalchange.NewPhytologist,171(2),271-284.doi:10.1111/j.1469-8137.2006.01752.x

37
Brooker,R.W.,&Callaghan,T.V.(1998).Thebalancebetweenpositiveandnegativeplantinteractionsanditsrelationshiptoenvironmentalgradients:amodel.Oikos,196-207.
Brooker,R.W.,Travis,J.M.,Clark,E.J.,&Dytham,C.(2007).Modellingspecies'rangeshiftsinachangingclimate:theimpactsofbioticinteractions,dispersaldistanceandtherateofclimatechange.JournalofTheoreticalBiology,245(1),59-65.doi:10.1016/j.jtbi.2006.09.033
Brown,C.D.,&Vellend,M.(2014).Non-climaticconstraintsonupperelevationalplantrangeexpansionunderclimatechange.ProceedingsoftheRoyalSocietyB:BiologicalSciences,281(1794).doi:10.1098/rspb.2014.1779
Brown,C.S.,Mark,A.F.,Kershaw,G.P.,&Dickinson,K.J.(2006).Secondarysuccession24yearsafterdisturbanceofaNewZealandhigh-alpinecushionfield.Arctic,Antarctic,andAlpineResearch,38(3),325-334.
Bruno,J.F.,Stachowicz,J.J.,&Bertness,M.D.(2003).Inclusionoffacilitationintoecologicaltheory.TrendsinEcology&Evolution,18(3),119-125.
Cahill,A.E.,Aiello-Lammens,M.E.,Fisher-Reid,M.C.,Hua,X.,Karanewsky,C.J.,YeongRyu,H.,Sbeglia,G.C.,SpagnoloF.,WaldronJ.B.,WarsiO.,&Wiens,J.J.(2012).Howdoesclimatechangecauseextinction?ProceedingsoftheRoyalSocietyB:BiologicalSciences.doi:10.1098/rspb.2012.1890
Callaway,R.M.(1995).Positiveinteractionsamongplants.TheBotanicalReview,61(4),306-349.
Callaway,R.M.,Brooker,R.,Choler,P.,Kikvidze,Z.,Lortie,C.J.,Michalet,R.,PaoliniL.,Pugnaire,F.I.,Newingham,B.,&Aschehoug,E.T.(2002).Positiveinteractionsamongalpineplantsincreasewithstress.Nature,417(6891),844-848.
Callaway,R.M.,&Walker,L.R.(1997).Competitionandfacilitation:asyntheticapproachtointeractionsinplantcommunities.Ecology,78(7),1958-1965.
Cavieres,L.A.,Quiroz,C.L.,Molina-Montenegro,M.A.,Muñoz,A.A.,&Pauchard,A.(2005).NurseeffectofthenativecushionplantAzorellamonanthaontheinvasivenon-nativeTaraxacumofficinaleinthehigh-AndesofcentralChile.PerspectivesinPlantEcology,EvolutionandSystematics,7(3),217-226.
Choler,P.,Michalet,R.,&Callaway,R.M.(2001).Facilitationandcompetitionongradientsinalpineplantcommunities.Ecology,82(12),3295-3308.
Crawley,M.J.(1989).Insectherbivoresandplantpopulationdynamics.AnnualReviewofEntomology,34(1),531-562.
Crawley,M.J.(1992).Seedpredatorsandplantpopulationdynamics.Pages157–191inM.Fenner,(Ed.).Seeds:theecologyofregenerationinplantcommunities.Wallingford,UK:CABInternational
Crimmins,S.M.,Dobrowski,S.Z.,Greenberg,J.A.,Abatzoglou,J.T.,&Mynsberge,A.R.(2011).Changesinclimaticwaterbalancedrivedownhillshiftsinplantspecies’optimumelevations.Science,331(6015),324-327.

38
Dullinger,S.,Gattringer,A.,Thuiller,W.,Moser,D.,Zimmermann,N.E.,Guisan,A.,Wilner,W.,PlutzarC.,LeitnerM.,&Mang,T.(2012).Extinctiondebtofhigh-mountainplantsundertwenty-first-centuryclimatechange.NatureClimateChange,2(8),619-622.
Feeley,K.J.,Silman,M.R.,Bush,M.B.,Farfan,W.,Cabrera,K.G.,Malhi,Y.,MeirP.,RevillaN.S.,QuisiyupanquiM.N.R.,&Saatchi,S.(2011).UpslopemigrationofAndeantrees.JournalofBiogeography,38(4),783-791.
Fenner,M.,Cresswell,J.,Hurley,R.,&Baldwin,T.(2002).Relationshipbetweencapitulumsizeandpre-dispersalseedpredationbyinsectlarvaeincommonAsteraceae.Oecologia,130(1),72-77.
Forbis,T.A.,&Doak,D.F.(2004).Seedlingestablishmentandlifehistorytrade-offsinalpineplants.AmericanJournalofBotany,91(7),1147-1153.
Frood,D.(2015).DescriptionofanewspeciesalliedtoPodolepisrobusta(Asteraceae:Gnaphalieae)fromthesouth-easternAustralianAlps.Muelleria,33,61-65.
Green,K.(2009).CausesofstabilityinthealpinetreelineintheSnowyMountainsofAustralia–anaturalexperiment.AustralianJournalofBotany,57(3),171-179.
Grime,J.P.(1977).Evidencefortheexistenceofthreeprimarystrategiesinplantsanditsrelevancetoecologicalandevolutionarytheory.AmericanNaturalist,1169-1194.
Harsch,M.A.,Hulme,P.E.,McGlone,M.S.,&Duncan,R.P.(2009).Aretreelinesadvancing?Aglobalmeta-analysisoftreelineresponsetoclimatewarming.EcologyLetters,12(10),1040-1049.
HilleRisLambers,J.,Harsch,M.A.,Ettinger,A.K.,Ford,K.R.,&Theobald,E.J.(2013).Howwillbioticinteractionsinfluenceclimatechange–inducedrangeshifts?AnnalsoftheNewYorkAcademyofSciences,1297(1),112-125.
Hillier,S.(1990).Gaps,seedbanksandplantspeciesdiversityincalcareousgrasslands.InS.HHillier,D.W.H.Walton,&D.A.Wells,(Eds.),Calcareousgrasslands-ecologyandmanagement(pp.57-66).Huntington,UK:BluntishamBooks.
Hobbie,S.E.,&Chapin,F.S.(1998).AnexperimentaltestoflimitstotreeestablishmentinArctictundra.JournalofEcology,86(3),449-461.
Hofgaard,A.,Dalen,L.,&Hytteborn,H.(2009).Treerecruitmentabovethetreelineandpotentialforclimate-driventreelinechange.JournalofVegetationScience,20(6),1133-1144.
Holt,R.D.(2009).BringingtheHutchinsoniannicheintothe21stcentury:ecologicalandevolutionaryperspectives.ProceedingsoftheNationalAcademyofSciences,106(2),19659-19665.
Holtmeier,F.-K.,&Broll,G.(2005).Sensitivityandresponseofnorthernhemispherealtitudinalandpolartreelinestoenvironmentalchangeatlandscapeandlocalscales.GlobalEcologyandBiogeography,14(5),395-410.doi:10.1111/j.1466-822X.2005.00168.x
Jeanes,J.A.(1999).Asteraceae.In:Walsh,N.G.,Entwisle,T.J.(Eds.),FloraofVictoriaVol.4,CornaceaetoAsteraceae.Melbourne,Australia:InkataPress.

39
Kikvidze,Z.,Khetsuriani,L.,Kikodze,D.,&Callaway,R.M.(2006).SeasonalshiftsincompetitionandfacilitationinsubalpineplantcommunitiesofthecentralCaucasus.JournalofVegetationScience,17(1),77-82.
Körner,C.(1999).Alpineplantlife:Functionalplantecologyofhighmountainecosystems.Heidelberg,Germany:Springer.
Körner,C.,&Paulsen,J.(2004).Aworld-widestudyofhighaltitudetreelinetemperatures.JournalofBiogeography,31(5),713-732.doi:10.1111/j.1365-2699.2003.01043.x
Körner,C.,Basler,D.,Hoch,G.,Kollas,C.,Lenz,A.,Randin,C.F.,VitasseY.,&Zimmermann,N.E.(2016).Where,whyandhow?Explainingthelow-temperaturerangelimitsoftemperatetreespecies.JournalofEcology,104(4),1076-1088.
Larcher,W.(1995).Photosynthesisasatoolforindicatingtemperaturestressevents.InE.DSchulze&M.MCaldwell(Eds.),EcophysiologyofPhotosynthesis(pp.261-277).Heidelberg,Germany:Springer.
Lee,T.D.,&Bazzaz,F.A.(1980).EffectsofdefoliationandcompetitionongrowthandreproductionintheannualplantAbutilontheophrasti.TheJournalofEcology,813-821.
Louda,S.M.1989.Predationinthedynamicsofseedregeneration.InM.A.Leck,V.T.Parker,&R.L.Simpson(Eds.),Ecologyofsoilseedbanks(pp.25-51).NewYork,USA:AcademicPress.
McAuliffe,J.R.(1988).Markoviandynamicsofsimpleandcomplexdesertplantcommunities.AmericanNaturalist,459-490.
McMaster,G.S.,&Wilhelm,W.(1997).Growingdegree-days:oneequation,twointerpretations.AgriculturalandForestMeteorology,87(4),291-300.
Moore,R.1973.Tetrazoliumstainingforassessingseedquality.SeedEcology,11-23.Moore,K.A.(2009).Fluctuatingpatchboundariesinanativeannualforb:therolesofniche
anddispersallimitation.Ecology,90(2),378-387.Munier,A.,Hermanutz,L.,Jacobs,J.D.,&Lewis,K.(2010).Theinteractingeffectsof
temperature,grounddisturbance,andherbivoryonseedlingestablishment:implicationsfortreelineadvancewithclimatewarming.PlantEcology,210(1),19-30.
Muraoka,H.,Tang,Y.,Koizumi,H.,&Washitani,I.(2002).EffectsoflightandsoilwateravailabilityonleafphotosynthesisandgrowthofArisaemaheterophyllum,ariparianforestunderstoreyplant.JournalofPlantResearch,115(6),419-427.
Parolo,G.,&Rossi,G.(2008).UpwardmigrationofvascularplantsfollowingaclimatewarmingtrendintheAlps.BasicandAppliedEcology,9(2),100-107.
Pauli,H.,Gottfried,M.,Dullinger,S.,Abdaladze,O.,Akhalkatsi,M.,Alonso,J.L.B.,ColdeaG.,DickJ.,ErschbamerB.,&Calzado,R.F.(2012).RecentplantdiversitychangesonEurope’smountainsummits.Science,336(6079),353-355.
Pickering,C.M.(2009).Pre-dispersalseedpredationbyTephritidaeiscommonamongspeciesofAustralianalpineAsteraceae.Arctic,Antarctic,andAlpineResearch,41(3),339-346.
RCoreTeam(2013).R:Alanguageandenvironmentforstatisticalcomputing.R

40
FoundationforStatisticalComputing,Vienna,Austria.ISBN3-900051-07-0,URLhttp://www.R-project.org/.
Rai,J.,&Tripathi,R.(1985).Effectofherbivorybytheslug,Mariaelladussumieri,andcertaininsectsongrowthandcompetitivesuccessoftwosympatricannualweeds.Agriculture,Ecosystems&Environment,13(2),125-137.
Randin,C.F.,Engler,R.,Normand,S.,Zappa,M.,Zimmermann,N.E.,Pearman,P.B.,VittozP.,ThuillerW.,&Guisan,A.(2009).Climatechangeandplantdistribution:localmodelspredicthigh-elevationpersistence.GlobalChangeBiology,15(6),1557-1569.doi:10.1111/j.1365-2486.2008.01766.x
Sakai,A.,&Weiser,C.(1973).FreezingresistanceoftreesinNorthAmericawithreferencetotreeregions.Ecology,54(1),118-126.
Scherrer,D.,&Koerner,C.(2010).Infra-redthermometryofalpinelandscapeschallengesclimaticwarmingprojections.GlobalChangeBiology,16(9),2602-2613.
Schütz,W.(2002).DormancycharacteristicsandgerminationtimingintwoalpineCarexspecies.BasicandAppliedEcology,3(2),125-134.
Shimono,Y.,&Kudo,G.(2005).Comparisonsofgerminationtraitsofalpineplantsbetweenfellfieldandsnowbedhabitats.EcologicalResearch,20(2),189-197.
Slatyer,R.,&Noble,I.(1992).Dynamicsofmontanetreelines.InA.JHansen&F.DiCastri(Eds.),Landscapeboundaries:Consequencesforbioticdiversityandecologicalflows(pp.346-359):NewYork,USA:Springer.
Sommerville,K.D.,Martyn,A.J.,&Offord,C.A.(2013).CanseedcharacteristicsorspeciesdistributionbeusedtopredictthestratificationrequirementsofherbsintheAustralianAlps?BotanicalJournaloftheLinneanSociety,172(2),187-204.
Song,M.,Tian,Y.,Xu,X.,Hu,Q.,&Ouyang,H.(2006).InteractionsbetweenrootandshootcompetitionamongfourplantspeciesinanalpinemeadowontheTibetanPlateau.ActaOecologica,29(2),214-220.
Speed,J.D.,Austrheim,G.,Hester,A.J.,&Mysterud,A.(2010).Experimentalevidenceforherbivorelimitationofthetreeline.Ecology,91(11),3414-3420.
Telwala,Y.,Brook,B.W.,Manish,K.,&Pandit,M.K.(2013).Climate-inducedelevationalrangeshiftsandincreaseinplantspeciesrichnessinaHimalayanbiodiversityepicentre.PLoSOne,8(2),e57103.
Thuiller,W.,Lavorel,S.,Araújo,M.B.,Sykes,M.T.,&Prentice,I.C.(2005).ClimatechangethreatstoplantdiversityinEurope.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,102(23),8245-8250.
Tielbörger,K.,&Kadmon,R.(2000).Temporalenvironmentalvariationtipsthebalancebetweenfacilitationandinterferenceindesertplants.Ecology,81(6),1544-1553.
Urban,M.C.,Tewksbury,J.J.,&Sheldon,K.S.(2012).Onacollisioncourse:competitionanddispersaldifferencescreateno-analoguecommunitiesandcauseextinctionsduringclimatechange.ProceedingsoftheRoyalSocietyofLondonB:BiologicalSciences,279(1735),2072-2080.

41
Urbanska,K.,&Schutz,M.(1986).Reproductionbyseedinalpineplantsandrevegetationresearchabovetimberline.BotanicaHelvetica,96,43-60.
Valiente-Banuet,A.,&Verdú,M.(2008).Temporalshiftsfromfacilitationtocompetitionoccurbetweencloselyrelatedtaxa.JournalofEcology,96(3),489-494.
VanRees,H.(1982).Thedietoffree-rangingcattleontheBogongHighPlains,Victoria.TheRangelandJournal,4(1),29-33.
Venn,S.,Pickering,C.,&Green,K.(2012).Short-termvariationinspeciesrichnessacrossanaltitudinalgradientofalpinesummits.BiodiversityandConservation,21(12),3157-3186.doi:10.1007/s10531-012-0359-2
Venn,S.E.,Green,K.,Pickering,C.M.,&Morgan,J.W.(2011).UsingplantfunctionaltraitstoexplaincommunitycompositionacrossastrongenvironmentalfilterinAustralianalpinesnowpatches.PlantEcology,212(9),1491-1499.doi:10.1007/s11258-011-9923-1.
Venn,S.E.,&Morgan,J.W.(2009).GerminationCharacteristicsofMountainCelery'AciphyllaGlacialis'(F.Muell.)Benth.(Apiaceae).TheVictorianNaturalist,126(1),4.
Venn,S.E.,Morgan,J.W.,&Green,P.T.(2009).DofacilitativeinteractionswithneighboringplantsassistthegrowthofseedlingsathighaltitudesinalpineAustralia?Arctic,Antarctic,andAlpineResearch,41(3),381-387.
Venn,S.E.,Morgan,J.W.,&Lord,J.M.(2013).FoliarfreezingresistanceofAustralianalpineplantsoverthegrowingseason.AustralEcology,38(2),152-161.
Wahren,C.,Papst,W.,&Williams,R.(1994).Long-termvegetationchangeinrelationtocattlegrazinginsub-alpinegrasslandandheathlandontheBogongHigh-Plains:ananalysisofvegetationrecordsfrom1945to1994.AustralianJournalofBotany,42(6),607-639.
Wahren,C.H.,Papst,W.,&Williams,R.(2001).Earlypost-fireregenerationinsubalpineheathlandandgrasslandintheVictorianAlpineNationalPark,south-easternAustralia.AustralEcology,26(6),670-679.
Wang,Y.,Chu,C.,Maestre,F.T.,&Wang,G.(2008).Ontherelevanceoffacilitationinalpinemeadowcommunities:anexperimentalassessmentwithmultiplespeciesdifferingintheirecologicaloptimum.ActaOecologica,33(1),108-113.
Warren,R.J.,&Bradford,M.A.(2011).Theshapeofthingstocome:woodlandherbnichecontractionbeginsduringrecruitmentinmesicforestmicrohabitat.ProceedingsoftheRoyalSocietyofLondonB:BiologicalSciences,278(1710),1390-1398.
Williams,R.,&Ashton,D.(1987).EffectsofdisturbanceandgrazingbycattleonthedynamicsofheathlandandgrasslandcommunitiesontheBogongHighPlains,Victoria.AustralianJournalofBotany,35(4),413-431.
Wilson,J.B.(1988).Shootcompetitionandrootcompetition.JournalofAppliedEcology,279-296.

42
AppendixI
Fig12.Shadecastbyresidentvegetationatnorth-westsitesatMtHotham,Victoria
Fig13.Shadecastbyresidentvegetationatsouth-eastsitesatMtHotham,Victoria

43
(a)
(b) (c) (d) (e)
00.20.40.60.81
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1660m
00.20.40.60.81
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1620m
00.20.40.60.81
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1690m
00.20.40.60.81
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1800m
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1860m
Fig.14.Frequencyofcanopyinterceptionofherb,gramminoidandshrublife-formsatMtHothamonsouth-eastexposedsites(a-e).
Freq
uencyofto
uche
s
Heightincrement(cm)

44
(a)(b)(c)(d)(e)
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1855m
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1820m
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1780m
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1700m
0
0.2
0.4
0.6
0.8
1
0-10 11-20 21-30 31-40 41-50 51-60 61-70
1670m
Fig.15.Frequencyofcanopyinterceptionofherb,gramminoidandshrublife-formsatMtHothamonnorth-westexposedsites(a-e).
Freq
uencyofto
uche
s
Heightincrement(cm)

45
AppendixII
Fig.16.EucalyptuspauciflorawoodlandwithdensePodolobiumalpestreunderstorydominatenorth-westexposedslopesatMtHotham(1670-1700m).
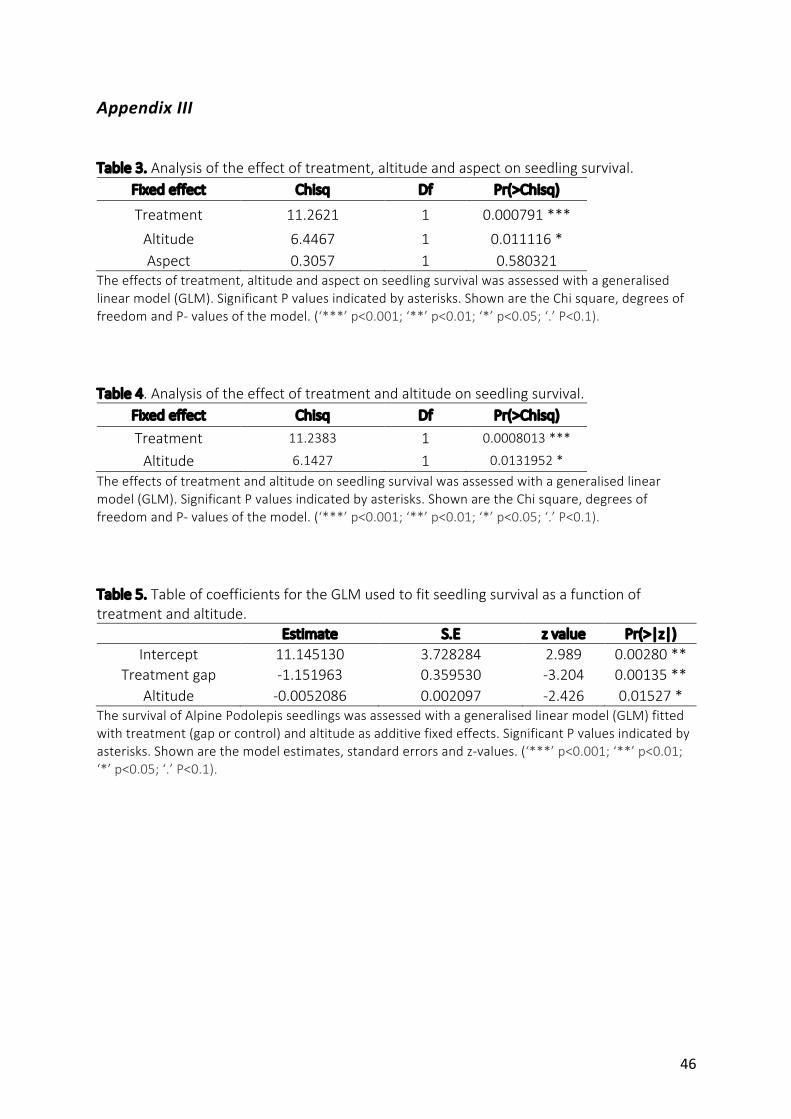
46
AppendixIIITable3.Analysisoftheeffectoftreatment,altitudeandaspectonseedlingsurvival.
Fixedeffect Chisq Df Pr(>Chisq)
Treatment 11.2621 1 0.000791***Altitude 6.4467 1 0.011116*Aspect 0.3057 1 0.580321
Theeffectsoftreatment,altitudeandaspectonseedlingsurvivalwasassessedwithageneralisedlinearmodel(GLM).SignificantPvaluesindicatedbyasterisks.ShownaretheChisquare,degreesoffreedomandP-valuesofthemodel.(‘***’p<0.001;‘**’p<0.01;‘*’p<0.05;‘.’P<0.1).Table4.Analysisoftheeffectoftreatmentandaltitudeonseedlingsurvival.
Fixedeffect Chisq Df Pr(>Chisq)Treatment 11.2383 1 0.0008013***Altitude 6.1427 1 0.0131952*
Theeffectsoftreatmentandaltitudeonseedlingsurvivalwasassessedwithageneralisedlinearmodel(GLM).SignificantPvaluesindicatedbyasterisks.ShownaretheChisquare,degreesoffreedomandP-valuesofthemodel.(‘***’p<0.001;‘**’p<0.01;‘*’p<0.05;‘.’P<0.1).Table5.TableofcoefficientsfortheGLMusedtofitseedlingsurvivalasafunctionoftreatmentandaltitude.
Estimate S.E zvalue Pr(>|z|)Intercept 11.145130 3.728284 2.989 0.00280**
Treatmentgap -1.151963 0.359530 -3.204 0.00135**Altitude -0.0052086 0.002097 -2.426 0.01527*
ThesurvivalofAlpinePodolepisseedlingswasassessedwithageneralisedlinearmodel(GLM)fittedwithtreatment(gaporcontrol)andaltitudeasadditivefixedeffects.SignificantPvaluesindicatedbyasterisks.Shownarethemodelestimates,standarderrorsandz-values.(‘***’p<0.001;‘**’p<0.01;‘*’p<0.05;‘.’P<0.1).

47
AppendixIVTable6.StatisticalanalysisofRelativeNeighbourEffectfornorth-westandsouth-eastsites.
RNEnorth-west coefficients estimate s.e. zvalue Pr(>|t|) 1670m-1700m -0.27689 0.16143 -1.715 0.0954 1670m-1820m -0.45308 0.14534 -3.117 0.0037** 1670m-1855m -0.34817 0.14968 -2.326 0.0261*
RNEsouth-east 1620m-1660m -0.07716 0.17118 -0.451 0.6547 1620m-1690m 0.07056 0.16204 0.435 0.6656 1620m-1800m 0.10457 0.16204 0.645 0.5225
Shownareone-wayANOVAcomparisonsbetweensitesaccordingtoaspect.AsterisksindicatesignificantP-values.(‘***’p<0.001;‘**’p<0.01;‘*’p<0.05;‘.’P<0.1).