cracking the fungal armor - studies on host defense ... · responses by antifungal therapy ......
TRANSCRIPT

Cracking the Fungal Armor -Studies on Host Defense Mechanisms
agains A. fumigatus
Tobias M. Hohl, MD, PhDMemorial Sloan-Kettering Cancer Center
hohlt@mskcc org [email protected]

A. fumigatus Germinationg
Intact pulmonary
Conidial clearance
Intact pulmonaryimmune defense
Defective pulmonaryDefective pulmonaryimmune defense
Tissue-invasive hyphae

Host Immune DefenseHost Immune Defense against A. fumigatus
• Recognition of inhaled spores by the innate immune systemy
• Modulation of inflammatory yresponses by antifungal therapy
• Monocytes and the initiation of CD4 T cell responsesp

Live Conidia induce Airway NeutrophilLive Conidia induce Airway Neutrophil Recruitment
36
42Total CellsMacrophagesNeutrophils
intratracheal
24
30
Neutrophils
12
18
6
Vehicle Heat-killed Live ConidiaResting conidia
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.

Live Conidia induce TNF/ CXCL2 SecretionLive Conidia induce TNF/ CXCL2 Secretion by Alveolar Macrophages
TNF CXCL2
18
24
45
60
6
12
15
30
ng/m
l
Live HK Medium LPSLive HK Medium LPS
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.

Killed Germinating Conidia are highly Inflammatoryg g y y
Conidial Swelling Germ Tube Formation
t = 0 3 h 5 h 7 h
9TNFCXCL2
3
6
ng/m
l
3 h 5 h 7 h
Heat-killed
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.

Killed swollen Conidia induce NeutrophilKilled swollen Conidia induce Neutrophil Influx into the BAL fluid
Total Cells30
MacrophagesNeutrophils
18
24
12
6
LiveHeat-killedConidia
Heat killedswollen conidia
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.
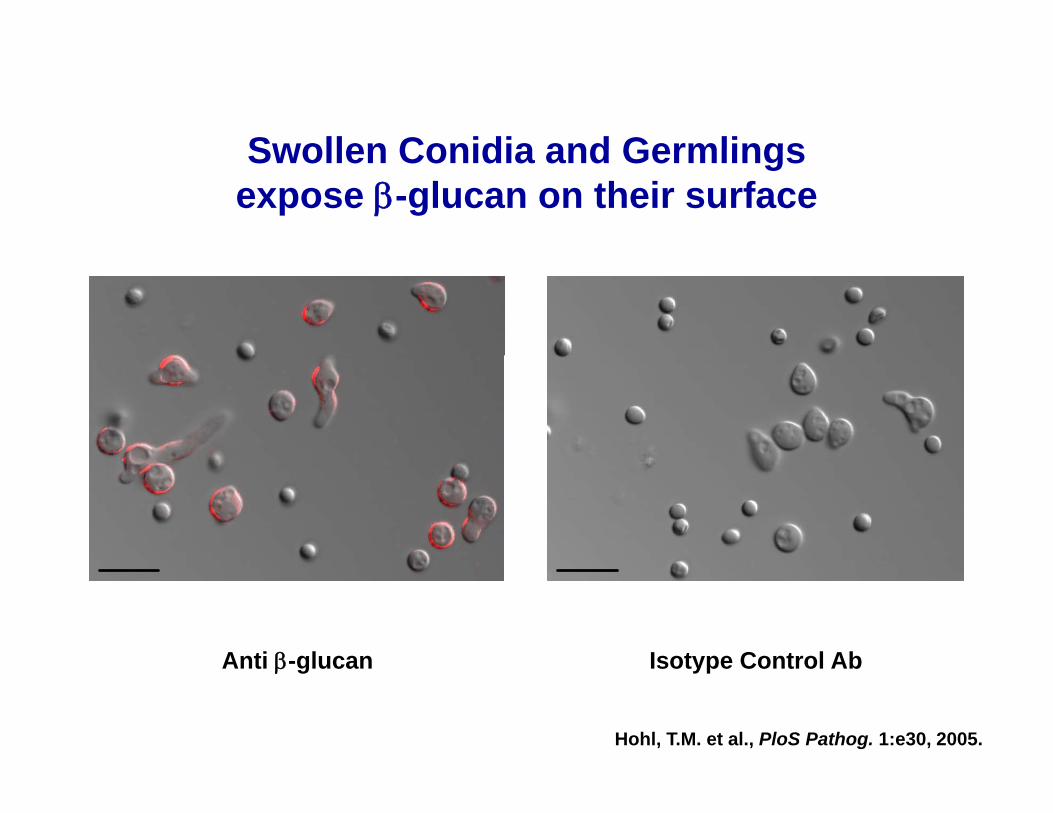
Swollen Conidia and GermlingsSwollen Conidia and Germlings expose β-glucan on their surface
Anti β-glucan Isotype Control AbAnti β-glucan Isotype Control Ab
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.

Dectin-1 binds and signals i t β (1 3) lin response to β-(1,3) glucan
Brown, G. D., Nat Rev Immunol 6:33-43, 2006.

Conidia Stimulate Dectin 1 and MyD88Conidia Stimulate Dectin-1- and MyD88-dependent Pathways
CXCL2TNF
2
3
/ml 2
3
1
ng/
1
WT MyD88-/-MyD88-/-WT
anti-Dectin - + - + - + - +
Hohl, T.M. et al., PloS Pathog. 1:e30, 2005.


Host Immune DefenseHost Immune Defense against A. fumigatus
• Recognition of inhaled spores by the innate immune systemy
• Modulation of inflammatory yresponses by antifungal therapy
• Monocytes and the initiation of CD4 T cell responsesp

Modulation of Host Inflammatory ResponsesModulation of Host Inflammatory Responses by Antifungal Therapy
• Echinocandins target fungal-β-D-glucan synthase• Echinocandins reduce A. fumigatus bulk β-glucan levels
(Kahn, J. et al., Antimicrob Agents Chemother 50:2214-2216, 2006)E hi di d t f ll i hibit A f i t th t i d• Echinocandins do not fully inhibit A. fumigatus growth, yet induce prominent morphologic changes at or above the MEC
1 x MEC Caspofungin (63 ng/ml) No Caspofungin

Caspofungin Decreases Macrophage Inflammatory Responses to ConidiaResponses to Conidia
1.2
1.8
ng/m
l)
No Caspofungin
0.6TNF
(n
**
* *No Caspofungin
Caspo (ng/ml) 0 4 8 16 31 63 125 250500
Caspofungin (500 ng/ml)
BMMφ TNF/CXCL2 release (500 ng/ml caspofungin vs. no drug exposure):
• TNF 0.49 ± 0.04* (range 0.46-0.54; n=4)• CXCL2 0.55 ± 0.10* (range 0.43-0.62; n=4)
Hohl, T.M. et al. J Infect Dis, 2008.

Caspofungin Enhances Macrophage InflammatoryCaspofungin Enhances Macrophage Inflammatory Responses to Hyphae
8
10
6
8
ml)
g/m
l)* * * * * * * *
2
4
6
2
4
TNF
(ng/
m
CXC
L2 (n
gC
0 4 8 16 31 63 125 250 500 0 4 8 16 31 63 125 250 500Caspo (ng/ml)
BMMφ TNF/CXCL2 release (500 ng/ml caspofungin vs. no drug exposure) • TNF 4.11 ± 2.39* (range 1.90-7.84; n=8)• CXCL2 2.90 ± 1.40* (range 1.53-5.41; n=8)
Hohl, T.M. et al. J Infect Dis, 2008.

Caspofungin Modulates Dectin-1-dependent Inflammatory Responses to Conidia Germlings and HyphaeResponses to Conidia, Germlings, and Hyphae
67
HyphaeConidia1.2 3.5
3
Germlings
23456
0 4
0.8
NF
(ng/
ml)
11.52
2.53
12
Caspofungin
0.4TN
+-
0.51
+- +-
Dectin-1-dependent TNF release
Dectin-1-independent TNF release
Hohl, T.M. et al. J Infect Dis, 2008.

Effects of Echinocandin Drugs on β-glucan ExposureEffects of Echinocandin Drugs on β-glucan Exposure
No Caspofungin
DIC anti-β-glucanDIC
Caspofungin
Aanti-β-glucan
8 h
10 h

Effects of Echinocandin Drugs on β-glucan Exposure
No CaspofunginCaspofungin No Caspofungin
DIC anti-β-glucanDIC
Caspofungin
Aanti-β-glucan
12 h
15 h
18 h
Hohl, T.M. et al. J Infect Dis, 2008.

Q tit ti A l i f β l I ti it i t dQuantitative Analysis of β-glucan Immunoreactivity associated with Caspofungin-treated and Untreated Hyphae
Integrated Fluorescence Intensity/Fungal Mass(Arbitrary Units)
Caspofungin-treated Hyphae Untreated Hyphae
Expt. 1 21.4 ± 8.3* 1.83 ± 0.73
Expt. 2 43.7 ± 7.0* 2.96 ± 4.67
Each value represents the average ratio (± SD) of β-glucan immunofluorescence intensity normalized to hyphal mass as calculated from 4-5 fields of view per condition.
* p <0.02 compared to control condition (untreated hyphae).
Hohl, T.M. et al. J Infect Dis, 2008.

Echinocandin Drugs have an Immunopharmacologic Mechanism of
Action

Host Immune DefenseHost Immune Defense against A. fumigatus
• Recognition of inhaled spores by the innate immune systemy
• Modulation of inflammatory yresponses by antifungal therapy
• Monocytes and the initiation of CD4 T cell responsesp

Monocyte-derived Populations in HostMonocyte-derived Populations in Host Defense against A. fumigatus

Development of an ExperimentalDevelopment of an Experimental System to Track and Ablate Monocytes

Recruitment of GFP+ monocytes and myeloid DCs into the lungs of A. fumigatus-infected mice
14.6
A. fumigatusg g
31.4
MDCs (2.7 M)
60.6 Mo (5.2 M)
29.17.2
Uninfected
48D11
c
GFP
MDCs(0.4 M)
Mo (0 7 M)48CD
CD11b
G
Ly6G
Mo (0.7 M)
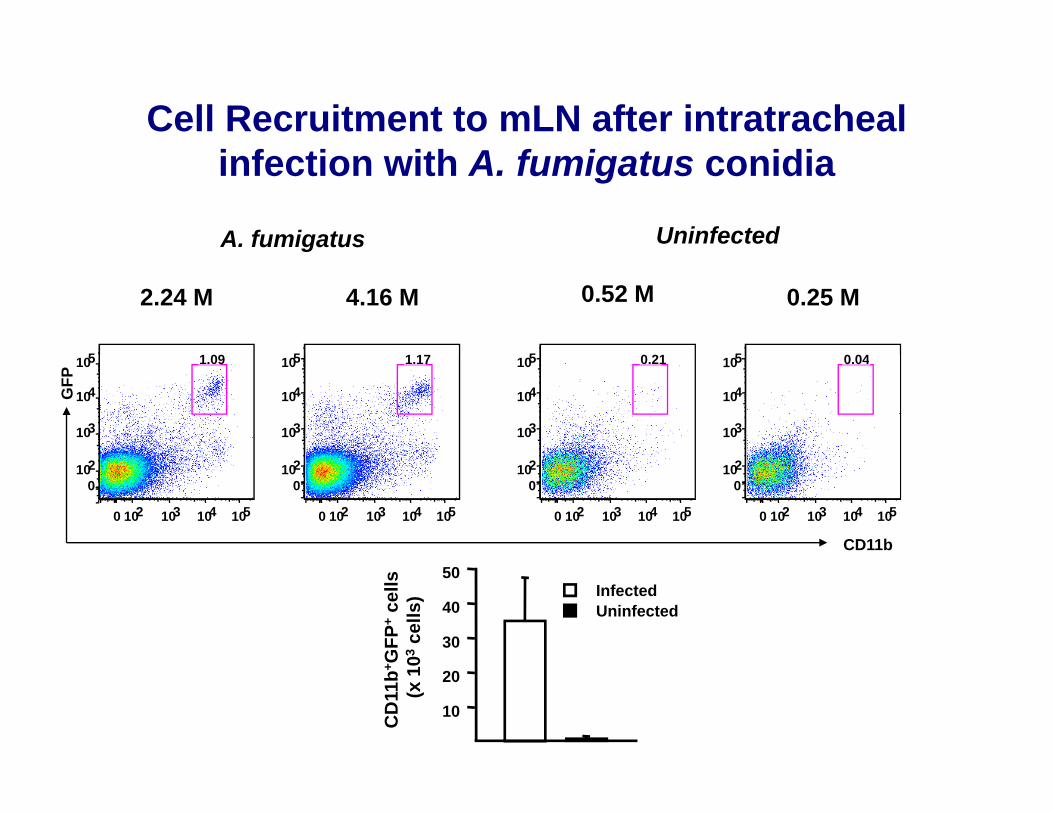
Cell Recruitment to mLN after intratracheali f ti ith A f i t idiinfection with A. fumigatus conidia
A. fumigatus Uninfected
2.24 M 4.16 M 0.52 M 0.25 M
5 1 09 5 1 17 5 0 21 5 0 04
103
104
105
GFP
1.09
103
104
105 1.17
103
104
105 0.21
103
104
105 0.04
0 102 103 104 105
CD11b
0102
0 102 103 104 105
0102
0 102 103 104 105
0102
0 102 103 104 105
0102
30
40
50InfectedUninfected
GFP
+ce
lls03
cells
)
10
20
CD
11b+
(x 1
0

Characterization of GFP+ cells in the mLN48 h t i f ti ith A f i t48 h post-infection with A. fumigatus
1.17
1.17 Mo
P
R249.7
38Mo
MDC
11b
MDC
GFP
CD11b
49.7
CD
CD11c MDCCD11b CD11c
Class II CD86 Ly6C

CD11b+CD11c+GFP+ cells transportCD11b CD11c GFP cells transport labelled conidia to mLN
10 4
10 5
P
0.4
15
20
25
2
2.5
100
150
51.8
0
10 2
10 3GFP
0
5
10
15
# C
ells
2.28
0
0.5
1
1.5
# C
ells
95.2
0
50
100
# C
ells
48.2
200
0 10 2 10 3 10 4 10 5
CD11b
10 51.4
0 10 2 10 3 10 4 10 5
AF633-Conidia
0
20
25
0 10 3 10 4 10 5
CD11c
00 10 3 10 4 10 5
CD11c
0
50
100
150
# C
ells
52.2
47.8
10 2
10 3
10 4
GFP
5
10
15
20#
Cel
ls
0.018
0 10 3 10 4 10 5
CD11c
00 10 2 10 3 10 4 10 5
CD11b
0
0 10 2 10 3 10 4 10 5
Conidia
0
5

Depletion of CCR2-expressing cellsDepletion of CCR2-expressing cells reduces conidial trafficking to the mLN

Ablation of Lung DC subsets in CCR2 Depleter

CCR2 Depleter mice cannot prime A.
f i t ifi CD4 T llfumigatus-specific CD4 T cell responsesCCR2 depleter orC57BL/6 (Thy 1.2) Infection via i.t. route
AF-specific CD4 T cells (Thy 1.1/1.2)
mLN CD4 Gate
+6-1 0 +1
0.06
0.09
0.12
Cells
103
104
105
hy 1
.1
0.15
CCR2 Depleter(DT t t d)
mLN CD4 GateDT DT
0 103 104 105
CFSE
0
0.03
#
0 102 103 104 105
Thy 1.2
0
102
10
Th(DT-treated)
6
9
12
Cells
103
104
105
y 1.
1
3.66
Non-Tg Control(DT t t d)
0 103 104 105
CFSE
0
3
6
#
0 102 103 104 105
Thy 1.2
0
102
10
Th(DT-treated)

Summary
• CCR2 reporter and depleter mice represent valuable tools to dissect the role of monocytes and monocyte derived cells in microbial defenseand monocyte-derived cells in microbial defense
• Monocyte-derived lung DCs (CD11b+) transport conidia to draining lymph nodesg y p
• Ablation results in loss of A. fumigatus-specific CD4 T cell primingM t i t ib t t i i d A• Monocytopenia may contribute to impaired A. fumigatus CD4 T cell responses in patients undergoing HSCTg g

AcknowledgementsAcknowledgements
MSKCC Albert Einstein College of Medicine• Eric Pamer• Heather Van Epps• Amariliz Rivera• Monica Mircescu
• Marta Feldmesser• Patrick Chen
P bli H lth R h I tit t• Alena Gallegos• Katharina Brandl• Ting Jia• Natalya Serbina
Public Health Research Institute, Newark, NJ
• David Perlin
• Natalya Serbina
• Alexander Lesokhin• Alan Houghton
University of Cape Town• Gordon Brown
• Mabel Ryder• James Fagin
Research Support: Charles F. Revson Foundation, NIH K08 Award