correlation of giardia duodenalis assemblages with clinical and epidemiological data in cuban...
TRANSCRIPT

Infection, Genetics and Evolution 23 (2014) 7–12
Contents lists available at ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid
Correlation of Giardia duodenalis assemblages with clinicaland epidemiological data in Cuban children
http://dx.doi.org/10.1016/j.meegid.2014.01.0071567-1348/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author. Tel.: +53 255 3645.E-mail address: [email protected] (L.J. Puebla).
Luis Jerez Puebla a,⇑, Fidel A. Núñez a, Yenisey Alfonso Fernández a, Jorge Fraga a, Lázara Rojas Rivero a,Iraís Atencio Millán a, Lucía Ayllón Valdés b, Isabel Martínez Silva b
a Department of Parasitology, Tropical Medicine Institute ‘‘Pedro Kourí’’, Autopista Novia del Mediodía Km 6½ e/Autopista Nacional y Carretera Central, La Habana, Cubab Paediatric Hospital ‘‘William Soler’’, Avenida 100 y Perla, Altahabana, La Habana, Cuba
a r t i c l e i n f o
Article history:Received 3 September 2013Received in revised form 26 December 2013Accepted 6 January 2014Available online 22 January 2014
Keywords:Giardia duodenalisAssemblagesPCRChildrenCuba
a b s t r a c t
Giardia duodenalis is one of the most frequent intestinal parasitic infections in children worldwide. Todate, eight main assemblages of G. duodenalis have been described, but only A and B genetic groupsare known to infect humans. In Cuba, this parasite has most clinical impact on children. The aim of thisinvestigation was genetic characterization of G. duodenalis isolated from children with giardiasis diag-nosed at the Paediatric Hospital ‘‘William Soler’’ between 2010 and 2011, and to compare the geneticresults with clinical and epidemiological data. A total of 103 stool samples from 452 children were posi-tive for G. duodenalis and co-infections with other parasites were noted in 5 cases. Assemblage identifi-cation was carried out by the amplification of a fragment of the triosephosphate isomerase (tpi) gene.Sub-assemblages of assemblage A (AI and AII) were identified by a nested PCR using the intergenic spacer(IGS) region of ribosomal deoxyribonucleic acid gene as a target. DNA from 90 of 103 (87.4%) samples wassuccessfully amplified by PCR–tpi. The prevalence of assemblages A and B was 40% and 42%, respectively.Infections with both assemblages were reported in 16 cases. No associations between epidemiologicalinformation and assemblage was detected, but assemblage B was significantly (P < 0.01) more frequentlyfound in children with diarrhea, flatulence or abdominal pain than assemblage A. Sub-assemblage AIIaccounted for the majority of cases (86.5%).
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
Giardia duodenalis (syn. G. intestinalis, G. lamblia) is the only spe-cies of Giardia found in humans, although it is also found in othermammals, including pets and livestock (Thompson, 2004). Giardi-asis is one of the most common human intestinal protozoal infec-tions reported worldwide. It has been estimated that there are280 million symptomatic cases of giardiasis each year (Feng andXiao, 2011).
Children are one of the high risk groups for infection, particu-larly those in daycares settings, orphanages and primary schools(Feng and Xiao, 2011; Muhsen and Levine, 2012). In Cuba, the over-all prevalence of this protozoal infection in Cuban population isabout 7.2% (Escobedo et al., 2011); however, higher prevalencerates have been found among young children attending day-carecenters and primary schools (Cañete et al., 2012).
G. duodenalis consists of a highly heterogeneous group of organ-isms and is referred to as a species complex divided at the genetic
level into assemblages or genetic groups (A–H) with different hostspecificities. Of these, human infections are mainly caused byassemblage A and B (Monis et al., 2003; Al-Mohammed, 2011). G.duodenalis assemblage A was subdivided commonly into sub-assemblages AI which is mainly zoonotic and AII which is com-monly anthroponotic, although it was reported by a few studiesin animals (Cacciò and Ryan, 2008; Sprong et al., 2009).
A number of molecular genetic techniques have been employedsuccessfully to characterize Giardia from stool samples. The vastmajority of these studies have relied on the analysis of some genessuch as, the small subunit ribosomal RNA (ssu-rRNA) (Hopkinset al., 1997), the b-giardin (bg) (Lalle et al., 2005), the glutamatedehydrogenase (gdh) (Read et al., 2004), and the triosephosphateisomerase (tpi) (Bertrand et al., 2005). The intergenic spacer (IGS)regions of rDNA have also been successfully used for molecularcharacterization of G. duodenalis, including sub-genotyping (Healeyet al., 1990; Lee et al., 2006; Al-Mohammed, 2011).
The development of tools to dissect the molecular biology of dif-ferent Giardia isolates, and the knowledge of the spectrum of symp-toms associated with giardiasis, has led to the hunt for associationsbetween particular assemblages and defined symptom patterns.

8 L.J. Puebla et al. / Infection, Genetics and Evolution 23 (2014) 7–12
The current assimilation of results is inconclusive, with both assem-blages associated with diarrheal disease. Different symptom spectraare apparently associated with different assemblages in differentpopulations (Robertson et al., 2010). In a previous study made by Pe-layo et al. (2008) in a small group of children from La Habana, theprevalence rates in the positive cases of children in the study were55% for assemblage B and 45% for assemblage A, using the bg andgdh genes as molecular markers, although the successful rate ofgenotyping was low. In that study children harbouring assemblageB of Giardia were more likely to have symptomatic infections thanchildren with isolates from assemblage A.
The purpose of this study was to determine the prevalence ofdifferent G. duodenalis assemblages and subtypes among childrenwith giardiasis, and investigate associations with clinical and epi-demiological data collected from children attending a PaediatricHospital from La Havana, Cuba.
2. Materials and methods
2.1. Clinical samples and surveillance data
A descriptive cross-sectional study in children attending at thePediatric Hospital ‘‘William Soler’’ was conducted from 2010 to2011. Stool samples from 452 symptomatic and asymptomatic chil-dren were examined for intestinal parasites by a wet smear stainedwith Lugol’s iodine and followed by formalin ethyl acetate concen-tration technique. All the stool samples were stained by modifiedacid-fast for Cryptosporidium spp., Cyclospora, and Cystoisospora(Garcia, 2001), and were tested for common bacterial pathogensusing standard culture methods (Murray and Baron, 2007).
Our samples came from a mixture of clinical cases as well asasymptomatic children enrolled in a program of school-based sur-veillance. These samples were divided in 163 asymptomatic chil-dren from day care centers corresponding to local surveillance tointestinal protozoa and 289 symptomatic children from primaryschools and day care assistants attending at the Pediatric Hospital‘‘William Soler’’ due to gastrointestinal symptoms.
All stool samples positive to G. duodenalis and in which bacterialpathogens were excluded were included in this research. Cases inwhich coinfection with other parasites, were not included in theinvestigation between infecting assemblages and clinical signs.
A questionnaire to obtain epidemiological and clinical data wascompleted by the parents or caregivers of the patients, followinginformed consent of their agreement to participate in this investi-gation. These surveillance data included information about someepidemiological variables (sex, age, residing area, ethnic group)and clinical symptoms (fever, flatulence, nausea, vomiting, head-ache, anorexia, fatigue, and loss of weight).
All Giardia positive samples were stored in potassium dichro-mate (2.5%) at 4 �C and transferred to ‘‘Pedro Kourí’’ Institute formolecular characterization.
2.2. Purification of G. duodenalis cyst and DNA extraction
Giardia cysts were purified and concentrated from stool samplesin a sucrose gradient with a specific gravity of 0.85 M and thenwashed with distilled water, following the protocol described byBabaei et al. (2011). The cyst wall was disrupted by 8–10 freeze–thaw cycles in liquid nitrogen alternated with a 95 �C water bath.After that purified cysts were mixed with 300 lL of buffer lysis(50 mM Tris–HCl, pH 7.5; 25 mM EDTA, 25 mM NaCl, and 1% of so-dium dodecyl sulfate (SDS)) and vortexed. After adding 100 lg/mLof proteinase K, the suspension was incubated at 56 �C for 2 h.
The DNA lysate was then treated with phenol/chloroform/iso-amyl alcohol (24:24:1), followed by chloroform/isoamyl alcohol
(24:1) according to Sambrook and Russell (2001). The DNA wasprecipitated by the addition of 1 mL chilled ethanol and stored at�20 �C until use. The dried DNA was re-suspended in 40 lL dis-tilled water and used as a template for PCR.
2.3. PCR amplification
Amplification of the tpi gene was performed as a single PCRwith a forward 50GGAGACCGACGAGCAAAGC30, and reverse 50
CTTGCCAAGCGCCTCAA30 primers for assemblage A (PCR–tpi A),and a forward 50AATAGCAGCACARAACGTGTATCTG30 and reverse50CCCATGTCCAGCAGCATC 30 for assemblage B (PCR–tpi B), as de-scribed by Bertrand et al. (2005). The PCR reaction mixture wasdone using a TopTaq Master Mix Kit (Qiagen, Germany) in a totalvolume of 25 lL, and comprised 10 lL of 10� PCR buffer (Qiagen,Germany), 0.2 mM of each deoxynucleoside triphosphate (dNTPs)(Applied Biosystems), 1U of Taq polymerase (Qiagen, Germany),0.4 lM of each primer, 5 lL of Q solution, and 5 lL of DNA tem-plate, with ultrapure water used as a negative control.
The DNA was amplified using a thermocycler (MJ Research,USA) following conditions described by Bertrand et al. (2005).The PCR products were visualized on 0.5 lg/mL of ethidium bro-mide-staining 2% agarose gels.
To amplify the IGS of rDNA, a nested PCR was done (Table 1) onall samples previously identified as being of assemblage A, using anested PCR as described by Lee et al. (2006), with 5 lL of templateused in the first PCR, and 2 lL of template (product from the firstPCR) in the second PCR. Primers are described in Table 1.
DNA from axenic cultures of G. duodenalis strains WB-C6(assemblage A), was used as positive controls, while ultrapurewater was included in negative controls.
2.4. Statistical analysis
All data were analyzed using EPINFO 6.04 and EPIDAT 3.1 statis-tical programmes. Chi square test and proportion tests were em-ployed to assess the significance of the associations. The Fisher’sexact test was used when required by data scarcity.
After normality tests described for Shapiro–Wilk and D’Agosti-no and Pearson, the medians of age in years, and duration of diar-rhea in days were compared using the Kruskal–Wallis nonparametric test. The P values less than 0.05 were considered as sta-tistically significant for all test. The odds ratio (OR) with 95% con-fidence interval (CI) were performed as measures of association.
3. Results
Of the 452 stool samples examined, 103 were positive to G. duo-denalis and co-infection with other parasites was found in 5 symp-tomatic children (Entamoeba histolytica/E. dispar, Ascarislumbricoides, Enterobius vermicularis, Cyclospora cayetanensis, andTrichuris trichiura). No bacterial pathogens were found in thosecases. These samples were not included in the investigation ofassociations between assemblages and clinical characteristics.Among 163 asymptomatic children investigated from day care cen-ters only 8 individuals were infected by Giardia, the other 95 wererepresented by symptomatic children from day care centers or pri-mary schools.
Of the samples from 103 children with giardiasis, PCR productsof the expected size were generated from 90 of them by PCR–tpi(87.4%). The prevalence of assemblages showed that 36 (40%) iso-lates belonged to assemblage A and 38 (42.2%) to assemblage B.The remaining 16 (17.8%) isolates demonstrated mixed infectionsof both assemblages. Among the 52 positive samples classified asassemblage A (including those in mixed infections), 45 (86.5%)
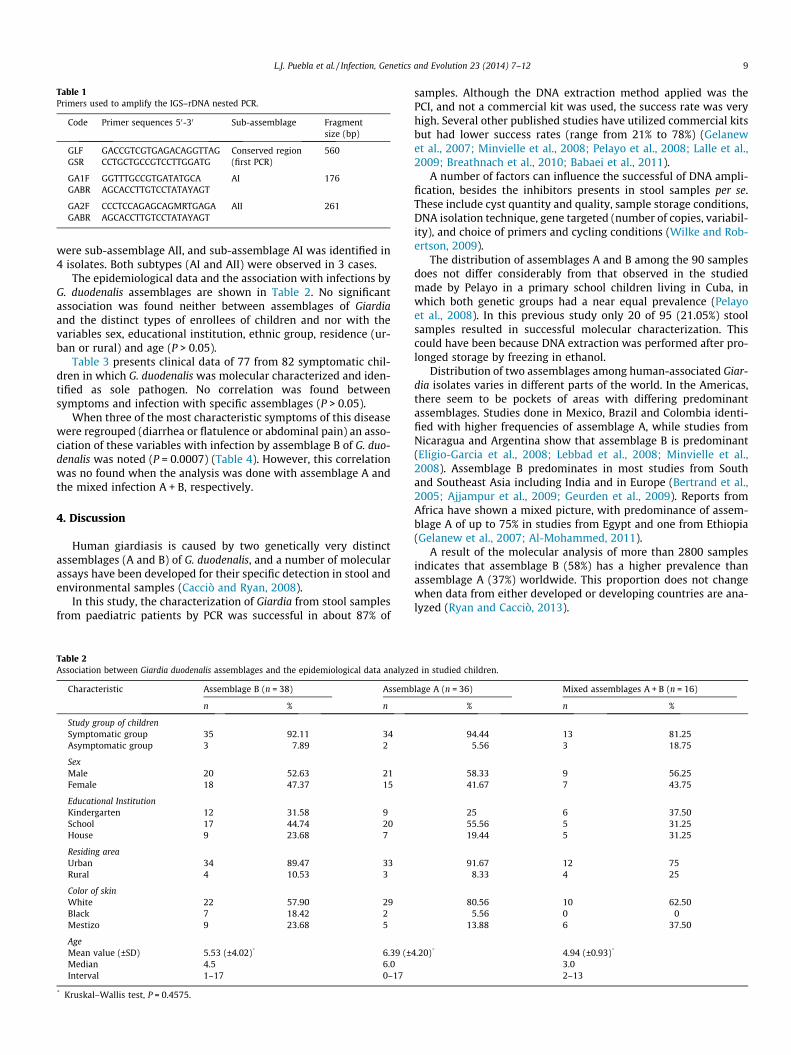
Table 1Primers used to amplify the IGS–rDNA nested PCR.
Code Primer sequences 50-30 Sub-assemblage Fragmentsize (bp)
GLF GACCGTCGTGAGACAGGTTAG Conserved region(first PCR)
560GSR CCTGCTGCCGTCCTTGGATG
GA1F GGTTTGCCGTGATATGCA AI 176GABR AGCACCTTGTCCTATAYAGT
GA2F CCCTCCAGAGCAGMRTGAGA AII 261GABR AGCACCTTGTCCTATAYAGT
L.J. Puebla et al. / Infection, Genetics and Evolution 23 (2014) 7–12 9
were sub-assemblage AII, and sub-assemblage AI was identified in4 isolates. Both subtypes (AI and AII) were observed in 3 cases.
The epidemiological data and the association with infections byG. duodenalis assemblages are shown in Table 2. No significantassociation was found neither between assemblages of Giardiaand the distinct types of enrollees of children and nor with thevariables sex, educational institution, ethnic group, residence (ur-ban or rural) and age (P > 0.05).
Table 3 presents clinical data of 77 from 82 symptomatic chil-dren in which G. duodenalis was molecular characterized and iden-tified as sole pathogen. No correlation was found betweensymptoms and infection with specific assemblages (P > 0.05).
When three of the most characteristic symptoms of this diseasewere regrouped (diarrhea or flatulence or abdominal pain) an asso-ciation of these variables with infection by assemblage B of G. duo-denalis was noted (P = 0.0007) (Table 4). However, this correlationwas no found when the analysis was done with assemblage A andthe mixed infection A + B, respectively.
4. Discussion
Human giardiasis is caused by two genetically very distinctassemblages (A and B) of G. duodenalis, and a number of molecularassays have been developed for their specific detection in stool andenvironmental samples (Cacciò and Ryan, 2008).
In this study, the characterization of Giardia from stool samplesfrom paediatric patients by PCR was successful in about 87% of
Table 2Association between Giardia duodenalis assemblages and the epidemiological data analyze
Characteristic Assemblage B (n = 38) Assemb
n % n
Study group of childrenSymptomatic group 35 92.11 34Asymptomatic group 3 7.89 2
SexMale 20 52.63 21Female 18 47.37 15
Educational InstitutionKindergarten 12 31.58 9School 17 44.74 20House 9 23.68 7
Residing areaUrban 34 89.47 33Rural 4 10.53 3
Color of skinWhite 22 57.90 29Black 7 18.42 2Mestizo 9 23.68 5
AgeMean value (±SD) 5.53 (±4.02)* 6.39 (±Median 4.5 6.0Interval 1–17 0–17
* Kruskal–Wallis test, P = 0.4575.
samples. Although the DNA extraction method applied was thePCI, and not a commercial kit was used, the success rate was veryhigh. Several other published studies have utilized commercial kitsbut had lower success rates (range from 21% to 78%) (Gelanewet al., 2007; Minvielle et al., 2008; Pelayo et al., 2008; Lalle et al.,2009; Breathnach et al., 2010; Babaei et al., 2011).
A number of factors can influence the successful of DNA ampli-fication, besides the inhibitors presents in stool samples per se.These include cyst quantity and quality, sample storage conditions,DNA isolation technique, gene targeted (number of copies, variabil-ity), and choice of primers and cycling conditions (Wilke and Rob-ertson, 2009).
The distribution of assemblages A and B among the 90 samplesdoes not differ considerably from that observed in the studiedmade by Pelayo in a primary school children living in Cuba, inwhich both genetic groups had a near equal prevalence (Pelayoet al., 2008). In this previous study only 20 of 95 (21.05%) stoolsamples resulted in successful molecular characterization. Thiscould have been because DNA extraction was performed after pro-longed storage by freezing in ethanol.
Distribution of two assemblages among human-associated Giar-dia isolates varies in different parts of the world. In the Americas,there seem to be pockets of areas with differing predominantassemblages. Studies done in Mexico, Brazil and Colombia identi-fied with higher frequencies of assemblage A, while studies fromNicaragua and Argentina show that assemblage B is predominant(Eligio-Garcia et al., 2008; Lebbad et al., 2008; Minvielle et al.,2008). Assemblage B predominates in most studies from Southand Southeast Asia including India and in Europe (Bertrand et al.,2005; Ajjampur et al., 2009; Geurden et al., 2009). Reports fromAfrica have shown a mixed picture, with predominance of assem-blage A of up to 75% in studies from Egypt and one from Ethiopia(Gelanew et al., 2007; Al-Mohammed, 2011).
A result of the molecular analysis of more than 2800 samplesindicates that assemblage B (58%) has a higher prevalence thanassemblage A (37%) worldwide. This proportion does not changewhen data from either developed or developing countries are ana-lyzed (Ryan and Cacciò, 2013).
d in studied children.
lage A (n = 36) Mixed assemblages A + B (n = 16)
% n %
94.44 13 81.255.56 3 18.75
58.33 9 56.2541.67 7 43.75
25 6 37.5055.56 5 31.2519.44 5 31.25
91.67 12 758.33 4 25
80.56 10 62.505.56 0 0
13.88 6 37.50
4.20)* 4.94 (±0.93)*
3.02–13
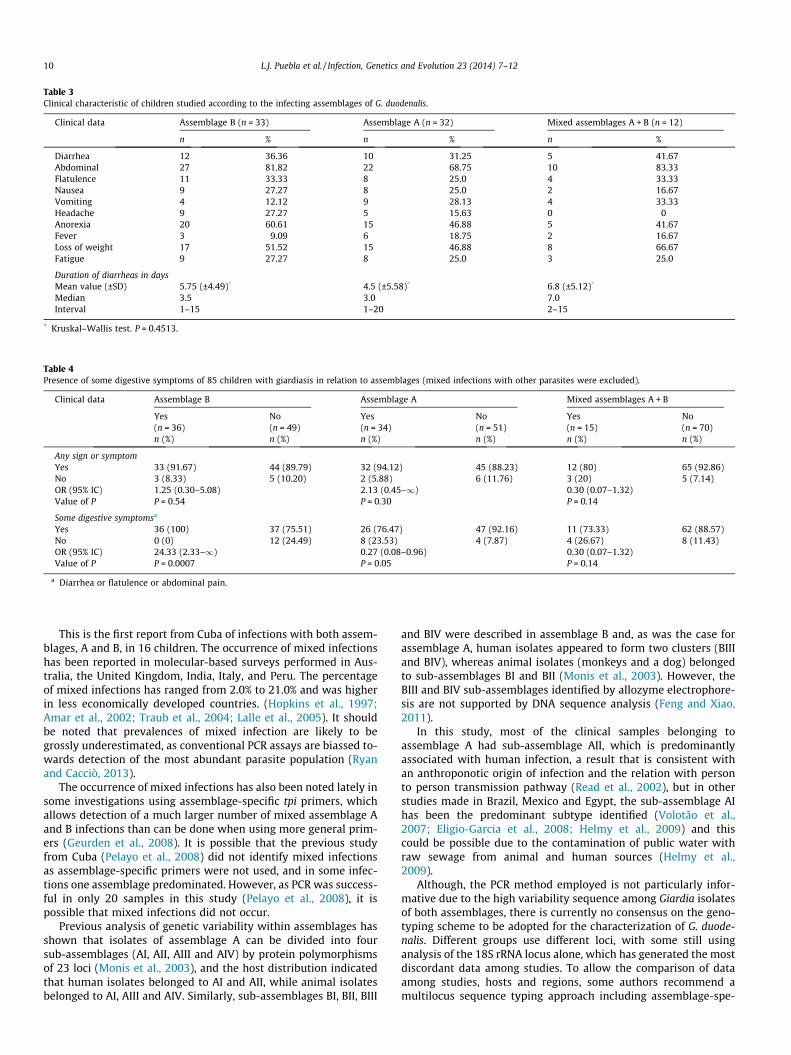
Table 3Clinical characteristic of children studied according to the infecting assemblages of G. duodenalis.
Clinical data Assemblage B (n = 33) Assemblage A (n = 32) Mixed assemblages A + B (n = 12)
n % n % n %
Diarrhea 12 36.36 10 31.25 5 41.67Abdominal 27 81.82 22 68.75 10 83.33Flatulence 11 33.33 8 25.0 4 33.33Nausea 9 27.27 8 25.0 2 16.67Vomiting 4 12.12 9 28.13 4 33.33Headache 9 27.27 5 15.63 0 0Anorexia 20 60.61 15 46.88 5 41.67Fever 3 9.09 6 18.75 2 16.67Loss of weight 17 51.52 15 46.88 8 66.67Fatigue 9 27.27 8 25.0 3 25.0
Duration of diarrheas in daysMean value (±SD) 5.75 (±4.49)* 4.5 (±5.58)* 6.8 (±5.12)*
Median 3.5 3.0 7.0Interval 1–15 1–20 2–15
* Kruskal–Wallis test. P = 0.4513.
Table 4Presence of some digestive symptoms of 85 children with giardiasis in relation to assemblages (mixed infections with other parasites were excluded).
Clinical data Assemblage B Assemblage A Mixed assemblages A + B
Yes No Yes No Yes No(n = 36) (n = 49) (n = 34) (n = 51) (n = 15) (n = 70)n (%) n (%) n (%) n (%) n (%) n (%)
Any sign or symptomYes 33 (91.67) 44 (89.79) 32 (94.12) 45 (88.23) 12 (80) 65 (92.86)No 3 (8.33) 5 (10.20) 2 (5.88) 6 (11.76) 3 (20) 5 (7.14)OR (95% IC) 1.25 (0.30–5.08) 2.13 (0.45–1) 0.30 (0.07–1.32)Value of P P = 0.54 P = 0.30 P = 0.14
Some digestive symptomsa
Yes 36 (100) 37 (75.51) 26 (76.47) 47 (92.16) 11 (73.33) 62 (88.57)No 0 (0) 12 (24.49) 8 (23.53) 4 (7.87) 4 (26.67) 8 (11.43)OR (95% IC) 24.33 (2.33–1) 0.27 (0.08–0.96) 0.30 (0.07–1.32)Value of P P = 0.0007 P = 0.05 P = 0.14
a Diarrhea or flatulence or abdominal pain.
10 L.J. Puebla et al. / Infection, Genetics and Evolution 23 (2014) 7–12
This is the first report from Cuba of infections with both assem-blages, A and B, in 16 children. The occurrence of mixed infectionshas been reported in molecular-based surveys performed in Aus-tralia, the United Kingdom, India, Italy, and Peru. The percentageof mixed infections has ranged from 2.0% to 21.0% and was higherin less economically developed countries. (Hopkins et al., 1997;Amar et al., 2002; Traub et al., 2004; Lalle et al., 2005). It shouldbe noted that prevalences of mixed infection are likely to begrossly underestimated, as conventional PCR assays are biassed to-wards detection of the most abundant parasite population (Ryanand Cacciò, 2013).
The occurrence of mixed infections has also been noted lately insome investigations using assemblage-specific tpi primers, whichallows detection of a much larger number of mixed assemblage Aand B infections than can be done when using more general prim-ers (Geurden et al., 2008). It is possible that the previous studyfrom Cuba (Pelayo et al., 2008) did not identify mixed infectionsas assemblage-specific primers were not used, and in some infec-tions one assemblage predominated. However, as PCR was success-ful in only 20 samples in this study (Pelayo et al., 2008), it ispossible that mixed infections did not occur.
Previous analysis of genetic variability within assemblages hasshown that isolates of assemblage A can be divided into foursub-assemblages (AI, AII, AIII and AIV) by protein polymorphismsof 23 loci (Monis et al., 2003), and the host distribution indicatedthat human isolates belonged to AI and AII, while animal isolatesbelonged to AI, AIII and AIV. Similarly, sub-assemblages BI, BII, BIII
and BIV were described in assemblage B and, as was the case forassemblage A, human isolates appeared to form two clusters (BIIIand BIV), whereas animal isolates (monkeys and a dog) belongedto sub-assemblages BI and BII (Monis et al., 2003). However, theBIII and BIV sub-assemblages identified by allozyme electrophore-sis are not supported by DNA sequence analysis (Feng and Xiao,2011).
In this study, most of the clinical samples belonging toassemblage A had sub-assemblage AII, which is predominantlyassociated with human infection, a result that is consistent withan anthroponotic origin of infection and the relation with personto person transmission pathway (Read et al., 2002), but in otherstudies made in Brazil, Mexico and Egypt, the sub-assemblage AIhas been the predominant subtype identified (Volotão et al.,2007; Eligio-Garcia et al., 2008; Helmy et al., 2009) and thiscould be possible due to the contamination of public water withraw sewage from animal and human sources (Helmy et al.,2009).
Although, the PCR method employed is not particularly infor-mative due to the high variability sequence among Giardia isolatesof both assemblages, there is currently no consensus on the geno-typing scheme to be adopted for the characterization of G. duode-nalis. Different groups use different loci, with some still usinganalysis of the 18S rRNA locus alone, which has generated the mostdiscordant data among studies. To allow the comparison of dataamong studies, hosts and regions, some authors recommend amultilocus sequence typing approach including assemblage-spe-

L.J. Puebla et al. / Infection, Genetics and Evolution 23 (2014) 7–12 11
cific detection. As a minimum, the gdh, tpi and bg loci should beused (Cacciò and Sprong, 2009; Ryan and Cacciò, 2013).
Given the genetic and phenotypic differences between assem-blages, it might be expected that there would be correlating clini-cal/epidemiological differences. However, our study did not findsignificant differences in the epidemiological aspects evaluated.
G. duodenalis is known for causing gastrointestinal symptoms,the most common are, acute or chronic diarrhea, flatulence, andstomach cramps, where as asymptomatic infections may occur,particularly in highly endemic areas (Ignatius et al., 2012). Severalstudies have reported correlations between assemblages andsymptoms, but there has been a lack of concordance in the data ob-tained. We associate these more predominant variables with theinfecting assemblage and found a correlation between infectionby assemblage B and the presence of these symptoms (diarrheaor flatulence or abdominal pain).
In a previous study by Pelayo et al. (2008) assemblage B wasmore prevalent among symptomatic children than assemblage A.In agreement with the present study results, Gelanew et al.(2007) stated that symptomatic giardiasis was more significantlyassociated with assemblage B in Ethiopia. Recently, MohammedMahdy et al. (2009) proved that assemblage B was more signifi-cantly present in symptomatic giardiasis patients in Malaysia. Leb-bad et al. (2011) straight for find a correlation between symptomsand assemblages and was found only for flatulence, which was sig-nificantly more common in children less than six years of age in-fected with assemblage B. In our continent, Minvielle et al.(2008) in Argentina identified assemblage B with polysymptomaticpeople, many of whom presented diarrhea, as well as Molina et al.(2011) in the same country found a relationship between assem-blage B with some clinical manifestations in children such as, diar-rhea, vomiting and weakness.
Other studies, have found a correlation between assemblage Aand symptomatic infection (Read et al., 2002; Ajjampur et al.,2009), while others have found relationship between assemblageB and asymptomatic infection in children less than 5 years of agein Australia and in children less than 10 years in Bangladesh,respectively (Read et al., 2002; Haque et al., 2005).
Differences in symptomatology of giardiasis with differentassemblages were initially described in Dutch patients, amongwhom assemblage A was associated with mild intermittent diseaseand assemblage B with severe persistent disease (Homan andMank, 2001). Since then, several studies have found an associationof diarrhea with assemblage A infection; in contrast, some studieshave found an association of symptoms with assemblage B. Inter-estingly, most studies where assemblage A was associated withdisease or more severe symptoms were from regions where assem-blage B predominated (Laishram et al., 2012).
To date, the studies in Giardia focus on the question of whetherthe G. duodenalis infectivity and pathogenicity vary in hosts of dif-ferent ages, or whether there is an association of clinical symp-toms, pathogenicity, or zoonotic potential with assemblages andsub-assemblages.
To the best of our knowledge, this is the second study to provideinformation on the assemblages associated with human infection,but is the first research aimed to know the sub-assemblages inCuba, representing the first study in the country that identifiedsub-assemblages by a PCR method.
5. Conclusions
This study confirms the similarity in distribution of assem-blages A and B in Giardia cases in Cuba. Both assemblages had sim-ilar epidemiological features, although assemblage B infectionswere statistically associated with clinical manifestations.
Acknowledgement
We would like to thank Dr. Lucy Robertson for the excellent re-view of this manuscript.
References
Ajjampur, S.S., Sankaran, P., Kannan, A., Sathyakumar, K., Sakar, R., Gladstone, B.,Kang, G., 2009. Giardia duodenalis assemblages associated with diarrhea inchildren in South India identified by PCR–RFLP. Am. J. Trop. Med. Hyg. 80, 16–19.
Al-Mohammed, H.I., 2011. Assemblages of Giardia intestinalis clinical isolates ofgastrointestinal symptomatic and asymptomatic Saudi children. Parasitol. Res.108, 1375–1381.
Amar, C.F.L., Dear, P.H., Pedraza-Diaz, S., Looker, N., Linnane, E., McLauchlin, J., 2002.Sensitive PCR restriction fragment length polymorphism assay for detectionand genotyping of Giardia duodenalis in human feces. J. Clin. Microbiol. 40, 446–452.
Babaei, Z., Oormazdi, H., Rezaie, S., Rezaeian, M., Razmjou, E., 2011. Giardiaintestinalis: DNA extraction approaches to improve PCR results. Exp. Parasitol.128, 159–162.
Bertrand, I., Albertini, L., Schwartzbrod, J., 2005. Comparison of two target genes fordetection and genotyping of Giardia lamblia in human feces by PCR and PCR–restriction fragment length polymorphism. J. Clin. Microbiol. 43, 5940–5944.
Breathnach, A.S., McHugh, T.D., Butcher, P.D., 2010. Prevalence and clinicalcorrelations of genetic subtypes of Giardia lamblia in an urban setting.Epidemiol. Infect. 138, 1459–1467.
Cacciò, S.M., Ryan, U., 2008. Molecular epidemiology of giardiasis. Mol. Biochem.Parasitol. 160, 75–80.
Cacciò, S.M., Sprong, H., 2009. Giardia duodenalis: genetic recombination and itsimplications for taxonomy and molecular epidemiology. Exp. Parasitol. 124,107–112.
Cañete, R., Díaz, M.M., Avalos García, R., Laúd Martinez, P.M., Manuel Ponce, F.,2012. Intestinal parasites in children from a day care centre in Matanzas city,Cuba. PLoS One 7 (12), e51394.
Eligio-Garcia, L., Cortes-Campos, A., Cota-Guajardo, S., Gaxiola, S., Jimenez-Cardoso,E., 2008. Frequency of Giardia intestinalis assemblages isolated from dogs andhumans in a community from Culiacan, Sinaloa, Mexico using beta-giardinrestriction gene. Vet. Parasitol. 156, 205–209.
Escobedo, A.A., Almirall, P., Alfonso, M., Avila, I., Cimerman, S., Salazar, Y., Dawkins,I.V., García, R.M., 2011. Caregiver perspectives for the prevention, diagnosis andtreatment of childhood giardiasis in Havana city, Cuba. A qualitative study. ActaTrop. 119 (2–3), 99–106.
Feng, Y., Xiao, L., 2011. Zoonotic potential and molecular epidemiology of Giardiaspecies and giardiasis. Clin. Microbiol. Rev. 24 (1), 110–140.
Garcia, L.S., 2001. Intestinal protozoa coccidia and microspordia. In: Garcia, L. (Ed.),Diagnostic Medical Parasitology, 4th ed. American Society of Microbiology,Washington.
Gelanew, T., Lalle, M., Hailu, A., Pozio, E., Caccio, S.M., 2007. Molecularcharacterization of human isolates of Giardia duodenalis from Ethiopia. ActaTrop. 102, 92–99.
Geurden, T., Geldhof, P., Levecke, B., Martens, C., Berkvens, D., Casaert, S.,Vercruysse, J., Claerebout, E., 2008. Mixed Giardia duodenalis assemblage Aand E infections in calves. Int. J. Parasitol. 38, 259–264.
Geurden, T., Levecke, B., Cacció, S.M., Visser, A., De Groote, G., Casaert, S., Vercruysse,J., Claerebout, E., 2009. Multilocus genotyping of Cryptosporidium and Giardia innon-outbreak related cases of diarrhea in human patients in Belgium.Parasitology 136, 1161–1168.
Haque, R., Roy, S., Kabir, M., Stroup, S.E., Mondal, D., Houpt, E.R., 2005. Giardiaassemblage A infection and diarrhea in Bangladesh. J. Infect. Dis. 192, 2171–2173.
Healey, A., Mitchell, R., Upcroft, J.A., Boreham, P.F., Upcroft, P., 1990. Completenucleotide sequence of the ribosomal RNA tandem repeat unit from Giardiaintestinalis. Nucleic. Acids Res. 18, 4006.
Helmy, M.M., Abdel-Fattah, H.S., Rashed, L., 2009. Real-time PCR/RFLP assay todetect Giardia intestinalis assemblages in human isolates with diarrhea in Egypt.J. Parasitol. 95, 1000–1004.
Homan, W.L., Mank, T.G., 2001. Human giardiasis: assemblage linked differences inclinical symptomatology. Int. J. Parasitol. 31, 822–826.
Hopkins, R.M., Meloni, B.P., Groth, D.M., Wetherall, J.D., Reynoldson, J.A., Thompson,R.C., 1997. Ribosomal RNA sequencing reveals differences between theassemblages of Giardia isolates recovered from humans and dogs living in thesame locality. J. Parasitol. 83, 44–51.
Ignatius, R., Gahutu, J.B., Klotz, C., Steininger, C., Shyirambere, C., Lyng, M.,Musemakweri, A., Aebischer, T., Martus, P., Harms, G., Mockenhaupt, F.P.,2012. High prevalence of Giardia duodenalis assemblage B infection andassociation with underweight in Rwandan children. PLoS Negl. Trop. Dis. 6,e1677.
Laishram, S., Kang, G., Ajjampur, S.S., 2012. Giardiasis: a review on assemblagedistribution and epidemiology in India. Indian J. Gastroenterol. 31 (1), 3–12.
Lalle, M., Bruschi, F., Castagna, B., Campa, M., Pozio, E., Caccio, S.M., 2009. Highgenetic polymorphism among Giardia duodenalis isolates from Sahrawichildren. Trans. R. Soc. Trop. Med. Hyg. 103, 834–838.

12 L.J. Puebla et al. / Infection, Genetics and Evolution 23 (2014) 7–12
Lalle, M., Pozio, E., Capelli, G., Bruschi, F., Crotti, D., Caccio, S.M., 2005. Geneticheterogeneity at the b-giardin locus among human and animal isolates ofGiardia duodenalis and identification of potentially zoonotic sub-assemblages.Int. J. Parasitol. 35, 207–213.
Lebbad, M., Ankarklev, J., Tellez, A., Leiva, B., Andersson, J.O., Svard, S., 2008.Dominance of Giardia assemblage B in Leon, Nicaragua. Acta Trop. 106, 44–53.
Lebbad, M., Petersson, I., Karlsson, L., Botero-Kleiven, S., Andersson, J.O.,Svenungsson, B., Svärd, S.G., 2011. Multilocus genotyping of human Giardiaisolates suggests limited zoonotic transmission and association betweenassemblage B and flatulence in children. PLoS Negl. Trop. Dis. 5 (8), e1262.
Lee, J.-H., Lee, J., Park, S.-J., Yong, T.-S., Hwang, U.-W., 2006. Detection andgenotyping of Giardia intestinalis isolates using intergenic spacer (IGS)-basedPCR. Korean J. Parasitol. 44, 343–353.
Minvielle, M.C., Molina, N.B., Polverino, D., Basualdo, J.A., 2008. First genotyping ofGiardia lamblia from human and animal feces in Argentina, South America.Mem. Inst. Oswaldo Cruz. 03, 98–103.
Molina, N., Pezzani, B., Ciarmela, M., Orden, A., Rosa, D., Apezteguía, M., Basualdo, J.,Minvielle, M., 2011. Intestinal parasites and assemblages of Giardia intestinalisin school children from Berisso, Argentina. J. Infect. Dev. Ctries. 5 (7), 527–534.
Mohammed Mahdy, A.K., Surin, J., Wan, K.L., Mohd-Adnan, A., AlMekhlafi, M.S., Lim,Y.A., 2009. Giardia intestinalis assemblages: risk factors and correlation withclinical symptoms. Acta Trop. 112, 67–70.
Monis, P., Anderws, R., Mayrhofer, G., Ey, P., 2003. Genetic diversity within themorphological species Giardia intestinalis and its relationship to host origin.Infect. Genet. Evol. 3, 29–38.
Muhsen, K., Levine, M.M., 2012. A systematic review and meta-analysis of theassociation between Giardia lamblia and endemic pediatric diarrhea indeveloping countries. Clin. Infect. Dis. 55 (4), S271–S293.
Murray, P., Baron, E., 2007. Manual of Clinical Microbiology, 9th ed. ASM,Washington.
Pelayo, L., Nuñez, F.A., Rojas, L., Furuseth Hansen, E., Gjerde, B., Wilke, H., Mulder, B.,Robertson, L., 2008. Giardia infections in Cuban children: the assemblagescirculating in a rural population. Ann. Trop. Med. Parasitol. 102 (7), 585–595.
Read, C., Walters, J., Robertson, I.D., Thompson, R.C., 2002. Correlation betweenassemblage of Giardia duodenalis and diarrhea. Int. J. Parasitol. 32, 229–231.
Read, C.M., Monis, P.T., Thompson, R.C.A., 2004. Discrimination of all assemblages ofGiardia duodenalis at the glutamate dehydrogenase locus using PCR–RFLP.Infect. Genet. Evol. 4, 125–130.
Ryan, U., Cacciò, S.M., 2013. Zoonotic potential of Giardia. Int. J. Parasitol. 43 (12–13), 943–956.
Robertson, L.J., Hanevik, K., Escobedo, A.A., Mørch, K., Langeland, N., 2010. Giardiasis– why do the symptoms sometimes never stop? Trends Parasitol. 26 (2), 75–82.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory Manual. ColdSpring Harbor Laboratory, New York.
Sprong, H., Caccio, S., van der Giessen, J., 2009. Identification of zoonoticassemblages of Giardia duodenalis. PloS Negl. Trop. Dis. 3 (12), e558.
Thompson, R.C.A., 2004. The zoonotic significance and molecular epidemiology ofGiardia and giardiasis. Vet. Parasitol. 126, 15–35.
Traub, R.J., Monis, P., Robertson, I., Irwin, P., Mencke, N., Thompson, R.C.A., 2004.Epidemiological and molecular evidence support the zoonotic transmission ofGiardia among humans and dogs living in the same community. Parasitology128, 253–262.
Volotão, A.C., Costa-Macedo, L.M., Haddad, F.S., Brandão, A., Peralta, J.M., Fernandes,O., 2007. Genotyping of Giardia duodenalis from human and animal samplesfrom Brazil using beta-giardin gene: a phylogenetic analysis. Acta. Trop. 102,10–19.
Wilke, H., Robertson, L.J., 2009. Preservation of Giardia cysts in stool samples forsubsequent PCR analysis. J. Microbiol. Meth. 78 (3), 292–296.