core.ac.ukeuteleosteans ognized by earlier authors. for example, in miuller's (1844)...
TRANSCRIPT

AMERICAN MUSEUMNovitatesPUBLISHED BY THE AMERICAN MUSEUMCENTRAL PARK WEST AT 79TH STREET,Number 2827, pp. 1-57, figs. 1-45
OF NATURAL HISTORYNEW YORK, N.Y. 10024
August 13, 1985
An Essay on Euteleostean Classification
DONN E. ROSEN'
ABSTRACT
The anatomy of the occipital region and rostralcartilage in euteleostean fishes is reviewed in somedetail. These data, in combination with other an-atomical features taken from the literature, haveled to a reassessment of interrelationships withinthe Euteleostei. This review supports the notionsthat the Salmoniformes, Aulopiformes, Mycto-phiformes, and Beryciformes are nonmonophy-
letic and raises questions about the monophyly ofthe fishes formerly grouped in the Osmeroidei.Evidence is presented on how the occipital regionmight be used in acanthomorph systematics, andincludes reasons for rejecting the concept of theParacanthopterygii, as this group was formerlyconstituted.
INTRODUCTION
The earliest general classification of fishesin which it is possible to pick out many ofthe main components of the Euteleostei isthat ofJohannes Muller (1844), in which theteleosts as a whole were presented as a ver-tebrate subclass, and their components as or-ders. These orders of Muller's bore namesthat may seem strange and unfamiliar to to-day's student, but the etymological charac-teristics of many of them were preserved forsome time, a few even to the present.Some of these names and their cross-
equivalents in different classifications werereviewed and explained in detail by Myers(1958) and now it remains only to emphasizethat these many different classifications[Muller (1844), Agassiz (1858), Gunther(1859-1870), Gill (1872), Boulenger (1904),Regan (1909, 1929), Goodrich (1909), Jor-
dan (1923), A. Smith-Woodward (1932),Norman (1934), Berg (1940) and its varioustranslations, reprinted editions and slightlymodified versions, and lastly Greenwood etal. (1966), McAllister (1968), and J. Nelson(1984)] went through an evolution from theearly nonsubordinated, ordinal classifica-tions of Muller (1844), Agassiz (1858), Gill(1872), Boulenger (1904), and Regan (1909,1929), to the complex subordinated, hierar-chical system of Goodrich (1909). It wasGoodrich (1909) who introduced the use ofuniform group endings, a practice that wasadopted by Berg (1940) and his successors,to ease recognition of the hierarchical posi-tion of a group in the general classification.During all ofthis history some ofthe fishes
now assigned to the Euteleostei were distrib-uted throughout most of the higher taxa rec-
1 Curator, Department of Ichthyology, American Museum of Natural History.
Copyright © American Museum of Natural History 1985 ISSN 0003-0082 / Price $5.25

AMERICAN MUSEUM NOVITATES
MULLER1844
REGAN19061929
FIG. 1. Branching diagrams representing the degree of resolution of teleostean classifications from1844 (Muller) to 1909 (Goodrich) and 1929 (Regan).
2 NO. 2827

ROSEN: EUTELEOSTEANS
ognized by earlier authors. For example, inMiUller's (1844) classification of fishlike ver-tebrates, the subclass Teleostei included sixorders, each of which included taxa now en-compassed by the Euteleostei. Thus, hisAcanthopteri included "perciform" fishes; theAnacanthini, the codfishes, cusk eels, andflatfishes; Pharyngognathi, the labrids, andtheir immediate allies; the Phystomi, the oto-physan Ostariophysi, percopsiforms, myc-tophids, salmonids, galaxiids, synbranchideels, pikes, and mudminnows; the Plectog-nathi, members of the modem Tetraodon-tiformes; and the Lophobranchii, the pipe-fishes, and seahorses. The Physostomi ofMuller also included some osteoglosso-morphs and the former is understood to bemore or less equivalent to the Malacopteryg-ii, Isospondyli, and Clupeiformes in classi-fications as recent as Berg's (1940). The threelatter "groups" were unnatural assemblagesof primitive teleosts, which included numer-ous taxa later treated by Greenwood et al.(1967) as euteleosteans in another unnaturalassemblage that Greenwood et al. (1966) hadpreviously termed the Protacanthopterygii.Progress has been slow, teleost classificationsgoing through a long period when all teleostswere assembled into one ofthree main kinds:lower, intermediate, or higher even as re-cently as Gosline (1971).The first comprehensive modem attempt
at detailed hierarchical synthesis since Good-rich (1909) was that of Greenwood et al.(1966), closely followed by that ofMcAllister(1968). This history is best appreciated byexamining a sprinkling of branching dia-grams extracted from the main componentsof each of the systems proposed since 1844(figs. 1 and 2).
It might be guessed that many major taxo-nomic problems have remained unsolvedonce an interest in cladistic methods of anal-ysis and classification was adopted by the ich-thyological community and most of theseproblems might be expected to be within theEuteleostei, as the largest of all recognizedteleostean assemblages. I do not regard theseproblems as close to solution since there ex-ists, still, significant disagreement amongstichthyologists on the interpretation of char-acter information and the delimitation ofnatural groups (Fink, 1984). This disagree-
ment is a sign of health and vigor in the fieldthat should serve as an example for othervertebrate systematists, some ofwhom haveabandoned the search for hierarchical orderin favor ofgeneral ecological research or lab-oratory studies of the behavior of selectedspecies.This paper is premised on certain new ob-
servations and interpretations of teleosteananatomy and the cladistic notions derivedfrom them. One of these notions, in agree-ment with Fink and Weitzman (1982) andFink (1984), is that the Salmoniformes ofRo-sen (1974) is not monophyletic. The secondis that the Aulopiformes of Rosen (1973) isalso not monophyletic. A third is that Poly-mixia and the acanthomorph neoteleosteansare defined, in part, by the absence of a re-sidual neural arch between the first vertebraand the occiput that is primitively present inmost halecostomes, many primitive eute-leosts, and in neoscopelids but not mycto-phids among the Myctophiformes, as rede-fined by Rosen (1973) to include themyctophids and neoscopelids.2 A fourth is
2 This and other characters conflict with four shared,derived features for myctophids and neoscopelids givenby Stiassny (Ms), but are consistent with the presence ofa subocular shelf, and single, medial, rostral cartilage inmost polymixiids and acanthomorphs (absent in neo-scopelids), and three or fewer predorsal bones in myc-tophids, polymixiids, and acanthomorphs (four in neo-scopelids). One of Stiassny's characters linking the twofamilies is the cone-shaped ventrally directed parapoph-yses for Baudelot's ligament that will, I believe, proveto be the primitive state of a similar structure in Poly-mixia and other acanthomorphs (fig. 18). Another ofStiassny's reasons for linking myctophids and neosco-pelids is described by Lauder (1983). In Lauder's paper,he stated that "all myctophiforms (including neoscope-lids; Rosen, 1973) possess a unique attachment of thebranchial skeleton to the urohyal." In myctophids, how-ever, the third hypobranchials have long anteroventraltips that clasp the urohyal laterally, whereas in neosco-pelids, the anteroventral tips ofthe third hypobranchialsextend forward above the urohyal to the dorsal edgewhere they are attached by ligament. The latter conditionis similar to that for primitive acanthomorphs (e.g., Poly-mixia) except that in the latter, the ligaments from thethird hypobranchials extend forward to contact the dor-sal edge of the urohyal more anteriorly. In any event, Isee no character here that clearly aligns myctophids withneoscopelids. This leaves two ligament characters ofStiassny's to align those taxa, as compared with the eight
1985 3

AMERICAN MUSEUM NOVITATES
that the Neoteleostei is characterized by atripartite occipital condyle (the basioccipitaland two exoccipital condyles), as describedand illustrated by Rosen and Patterson (1969),and which unites stomiiforms with them, aselaborated by Fink and Weitzman (1982) andFink (1984). A fifth is that, in disagreementwith Fink and Weitzman (1982), the presenceofa well-defined triple joint that incorporatestwo large exoccipital condyles is not evidencefor linking the Salmonidae with the neote-leosts since this type of joint has a limiteddistribution only in Recent salmonines andis, therefore, probably convergent. A sixth isthat neoteleosts primitively show a cervicalgap between the occiput and first vertebra. Aseventh is that the Acanthomorpha are de-fined by complete closure of the cervical gapvia two prezygapophyseal exoccipital facetsand a basioccipital facet from the body ofthevertebral centrum. An eighth is that Poly-mixia is the sister group to the Acantho-morpha, thus defined because it possesses ex-occipital facets but retains a part ofthe cervicalgap in the basioccipital position. Other "be-ryciforms" have a more derived "percoid-like" condition.
features that relate myctophids, but not neoscopelids, tothe acanthomorphs. (1) a large subocular shelf (like thatin Polymixia), (2) rostral cartilage a simple median struc-ture (with relic pairs of lateral cartilages, or no pairedcartilages), (3) an interarcual cartilage between the firstand second gill arches (small, when present, and absentin some species including Polymixia and some primitiveacanthomorphs), (4) only three predorsal bones, (5) par-tial closure ofthe cervical gap, (6) absence ofan accessoryneural arch in cervical region, (7) presence ofneural archprezygapophyses on the first vertebra, (8) direct connec-tion (via ligaments) of the autocentrum of first vertebrawith the exoccipital condyles. Character 3 is questionablebecause several groups ofprimitive acanthomorphs lackan interarcual cartilage and characters 6 through 8 mightbe manifestations of only a single developmental shift.Even allowing for the latter two ambiguities, there arefour trenchant features suggesting nonmonophyly of themyctophids plus neoscopelids (characters 1, 2, 4, and 5-8) as contrasted with the two remaining ligament featuresproposed by Stiassny. This state of affairs indicates tome that myctophids are the sister group of acantho-morphs (in a restricted ctenosquamata), but that neo-
scopelids, for reasons discussed in this paper, are bestregarded, at present, as part of major polychotomy im-mediately preceding the ctenosquamata in the cladogram(fig. 45).
Certain subsidiary notions also emerge asconsequences of this work, (1) that the "os-meroids" and the Salmonidae might not bemonophyletic groups. Pefiaz (1983, p. 370)recently attempted an ontogenetic diagnosisofthe Salmonidae, but pointed out that someof the diagnostic features may also be foundin other fish groups, (2) that Aulopus, andperhaps some closely allied forms excludingChlorophthalmus, might jointly form the sis-ter group to the Ctenosquamata based, in part,on the anatomy of the rostral cartilage, and(3) that within the groups that remain, theold Paracanthopterygii toadfishes and theirimmediate allies might be more closely linkedto cods and theirs than the cods are to cuskeels and brotulas based on both neurocranial,vertebral, and gill arch evidence.3
ACKNOWLEDGMENTS
I am grateful to the following for com-mentaries during the work and specimens forstudy, especially Daniel M. Cohen, S. and W.Fink, P. H. Greenwood, George Lauder, Co-lin Patterson, D. Siebert, K. Sulak, RichardVari, and S. Weitzman. Part ofthis work wasbegun with Colin Patterson and MelanieStiassny in 1983, both ofwhom supplied mewith many helpful comments and drawings.To the others, many thanks: R. Bailey, F.Berry, B. Collette, L. Grande, G. Nelson, andV. Springer. I thank Mary Rauchenberger,Christine Rossi, and Janice Lillien for muchtechnical assistance and specimen prepara-tion. This work was supported by NSF grantBSR 8100103.
ANATOMICAL ABBREVIATIONS
ACCNA, accessory neural archABAUDLIG, attachment for Baudelot's ligamentANA, ankylosed neural archARTPR, articular processASCPR, ascending processAUTLIG, autocentral ligamentAUTNA, autogenous neural archBAUDLIG, Baudelot's ligamentBO, basioccipitalBOC, basioccipital condyleBOF, basioccipital facetCG, cervical gap
3The gill arch evidence will be discussed in a subse-quent paper by Patterson and Rosen.
4 NO. 2827

ROSEN: EUTELEOSTEANS
A. SMITH WOODWARD1932
MCALLISTER1 968
FIG. 2. Branching diagrams representing the degree of resolution of teleostean classifications from1932 (A. Smith-Woodward) to the present time as summarized by Rosen in 1982.
1985 5

AMERICAN MUSEUM NOVITATES
CTNOT, connective tissue sheath of notochord?CART, questionably cartilaginousDNA, depression in autocentrum for neural arch
baseENR, epineural ribEP, epuralEXO, exoccipitalEXOC, exoccipital condyleEXOF, exoccipital facetEXO FRAGMENTS, exoccipital fragments from
site of attachment of autocentral ligamentHYPI-6, hypural 1 to 6LEXO, left exoccipitalMEDCART, medial upper jaw cartilageMX, maxillaNA, neural archNAPZYG, prezygapophyseal neural archNOT, notochordNPU2, 3, neural spines on second and third pleuralcentrum
PAL, palatinePAR, parapophysis?PAR, questionably a parapophysisPARHYP, parhypuralPD 1, first predorsal bonePMX, premaxillaPOPMYO, posterior opening of posterior myo-dome
PMXCART, premaxillary cartilagePR, pleural ribPTMXPR, postmaxillary process of premaxillaPU1, 2, first or second preural centrumPZYG, prezygapophysisRCART, rostral cartilageRETDORS TENDON, tendinous origin of re-
tractor dorsalis muscleREXO, right exoccipitalRV1NA, right halfofneural arch on first vertebraRVINSP, right half of neural arch and spine on
first vertebraSACBUL, saccular bullaSEXOF, site for development of exoccipital facetSTEG, steguralU 1, 2, first or second ural centrumUN 1, 2, first or second uroneuralV 1, 2, 3, first, second or third vertebraVIANA, ankylosed neural arch of first vertebraV1NSP, neural arch and spine of first vertebraX, foramen for vagus nerve
INSTITUTIONAL ABBREVIATIONSAMNH, American Museum of Natural HistoryMCZ, Museum ofComparative Zoology, Harvard
University
ANATOMICAL EVIDENCE
THE NEUROCRANIAL JOINT WITH THE FIRSTVERTEBRA: It has been proposed (Patterson,
1964; Rosen and Patterson, 1969; and Finkand Weitzman, 1982) that advanced eute-leosts can be characterized by the presencein the posterior neurocranium ofan invertedY-shaped junction between the basioccipitaland the exoccipitals. This configuration canbe seen in primitive myctophids, stomi-iforms, tCtenothrissa radians, and Polymix-ia (Patterson, 1964). This condition appearsto differ from that of primitive teleosts inwhich the basioccipital occupies the entirearea of these three bones for contact with thecentrum of the first vertebra. When that basi-occipital contact is reduced, the exoccipitalsenter the posterior neurocranial surface toform a tripartite condylar surface for contactwith the first vertebra.Fink and Weitzman (1982), citing the ear-
lier paper by Rosen and Patterson (1969),called attention to the tripartite occipital con-dyle as a synapomorphy ofneoteleosts. Find-ing a similar occipital joint in the gonosto-matid stomiiform, Diplophos, they proposedthat this is one of two features that unitesstomiiforms with neoteleosts. The other fea-ture, a rostral premaxillary cartilage, is dis-cussed below. Fink and Weitzman also usedthe tripartite condyle and rostral cartilage topropose a sister-group relationship ofthe Sal-monidae with the neoteleosts plus stomi-iforms.The nature of the occipital joint with the
first vertebra has attracted the attention ofseveral investigators. Ridewood (1904, 1905)held the view that a tripartite joint is prim-itive for teleosts, but is masked by the fusionof the first vertebral centrum (V1) to the oc-ciput so as to exclude the exoccipitals fromthe joint surface. In that view, what is iden-tified in most teleosts as the basioccipital isactually a vertebra fused to the braincase.Removal of this vertebra should, therefore,reveal the primitive tripartite arrangement.Patterson (1975, p. 318) proposed that thebasioccipital condyle, rather than being a ver-tebra, is made up of a plug of osteoid tissuerepresenting the ossification of the small an-terior part of the notochord that penetratesthe basioccipital bone, and that it is the growthof this osteoid plug that excludes the exoc-cipitals from the posterior face ofthe occiput.Cavender and Miller (1972) also reviewed
the origin of the salmonid occipital joint and
NO. 28276

ROSEN: EUTELEOSTEANS
FIG. 3. Salmonine occipital regions show character and disposition of the condyles that articulatewith the first vertebra. A. Salmo salar Linnaeus, AMNH 39098, posterior view ofskull ofadult specimen.B and C. Oncorhynchus tshawytscha (Walbaum), AMNH 21719. B. Posterior view of exoccipital andbasioccipital bones. C. As in B, but in lateral view with first vertebra in place (see fig. 4).
concluded, correctly in my view, that the tri-partite condition of salmonids is present ornot as a consequence of whether a vertebrafuses or does not fuse with the braincase.
Cavender and Miller took an interest in thismatter after finding and describing a largemiddle Pliocene salmonid from westernNorth America (tSmilodonichthys rastro-
71985

AMERICAN MUSEUM NOVITATES
EXOF
EXOF
FIG. 4. The first vertebra of Oncorhynchus tshawytscha (Walbaum), AMNH 21719, in posterior (A)and three-quarter posterodorsal (B) views. This vertebra contains a pair of wells on its dorsal surfaceunderlying a poorly developed (accessory) neural arch that does not bear a neural spine.
sus); they proposed that it has a close rela-tionship to the species of Oncorhynchus. Af-ter noting that Oncorhynchus has a tripartitejoint (figs. 3B, C, 4, 5) and tSmilodonichthysbut a simple basioccipital condyle, they re-viewed the distribution ofthese different kindsof articulations in a variety of teleosts. Ca-vender and Miller concluded that there is evi-dence of a vertebra fusing with the braincasein the Pliocene fossil and that such fusion isby no means unusual or restricted to just afew taxa. In fact, they report that "in Core-gonus two centra may be fused with the basi-occipital" and that the "condition in Proso-pium williamsoni and [Thymallus, Norden(1961)] is somewhat intermediate" betweenthe tripartite condyle and the single one inCoregonus.4 They also write that "close in-
4 Fink (1984) argued that because Prosopium has theexoccipitals participating in the occipital condyle alongwith Thymallus and the Salmoninae that the tripartitecondition is primitive for the Salmonidae in general, and
can therefore be used as evidence to link the salmonidswith the neoteleosts. But in my material, exoccipital par-ticipation is not true of Prosopium williamsoni, P. cy-lindraceum, or any other coregonine examined (figs. 7,8, 13) and the condition in Thymallus is hardly differentfrom that of Albula (fig. 9) and Pterothrissus, amongother teleosts, in which only a small extension of theexoccipital is visible posteriorly without noticeably af-fecting the shape of the basioccipital. The tripartite oc-cipital condyle of salmonines is more derived than thesimple, inverted Y-shaped morphology in primitivectenosquamates (except during early ontogeny, fig. 12A),resembling in the adult state that of an advanced per-comorph (e.g., Lutjanus, cf. figs. 3B and 24, 25). Thusthe resemblance is probably secondary. This conclusionpredicts a different ontogeny for the salmonine and cten-osquamate conditions which, if found, would indicatetheir nonhomology (see below, p. 54). Perhaps the prob-lem is, as stated by Fink (1984), that the monophyly ofthe Salmonidae "is based primarily on a single character,apparent polyploidy of the karyotype . . ." and that, asenunciated by him, many of the salmoniform taxa areunnatural (e.g., "salmonids," "osmeroids," and "eso-coids").
8 NO. 2827

ROSEN: EUTELEOSTEANS
DNA
EXOF
krvB
C
FIG. 5. First vertebra of Salmo gairdneri Richardson, AMNH 40268, parr stage (ca. 10 cm totallength). A. Dorsal view to show wells for accessory neural arch. B. Three-quarter posterior view. C.Anterior view to show extent of development of facets that articulate with exoccipital condyle. Notelarge notochordal canal in centrum.
spection of the condyle [in tSmilodonich-thys] shows that it is ... a fused centrum thatsupported a neural arch [in] a pair of inden-tations ... on its dorsal surface" (cf. figs. 3-8). And further, that this "basicranial-verte-bral joint is similar to that found in Tarpon[sic] atlanticus, Megalops cyprinoides(Greenwood, 1970), [and] Albula vulpes...."Their claim is problematical because onto-genetic data illustrating the course of verte-bral fusion are lacking for most cited exam-ples. Such data are available for Megalopsatlanticus, however (fig. 9A, B). Strong cir-cumstantial support for the idea can be foundin other elopomorphs, as illustrated by Forey(1973), where the part claimed to be a fusedcentrum not only bears a neural arch but par-apophyses as well (Forey, 1973, figs. 3, 5, 21-23, 31).
In Elops, which appears to have a vertebrathat is ontogenetically a part of the basioc-cipital (fig. 10 and illustrations in Forey,1973), Baudelot's ligament is attached to theventrolateral aspect of the first free vertebra.Whitehead and Teugels (in press) describe asituation much like that of Elops in a fresh-water herring, Sierrathrissa. They state that"the posterior halfofa first vertebral centrum
[appears to have] become trapped by flangesfrom the exoccipital and basioccipital . . . ,"thus agreeing with Ridewood's (1904) inter-pretation of the clupeid occipital region (andsee Greenwood, 1968, on Denticeps). Addi-tional circumstantial support for the primi-tive ontogenetic incorporation of vertebraewith the braincase in modem halecostomescomes from the correlation ofthe occurrenceof accessory, free-floating neural arches withfusion of vertebrae to the occiput [e.g., inAmia (Jollie, 1984a, p. 431)].The clupeocephalan first neural arch is re-
duced and incomplete (figs. 11 A, 12B) ascompared with the neural arch associated withthe basioccipital in some elopomorphs (fig.10), and might, therefore, be another syn-apomorphy of the Clupeocephali. This lineof argument depends on an assumption thatthe position of Baudelot's ligament is a reli-able landmark for the identification ofa givenvertebra. In osteoglossomorphs there is noevidence, direct or circumstantial, that a cen-trum is primitively fused with the braincase(e.g., in Hiodon or Scleropages), but in a morederived condition several vertebrae either areclosed adherent to the basioccipital (Osteo-glossum) or are included in a complex an-
1985 9

AMERICAN MUSEUM NOVITATES
AR
PAR
DNA
B
SEXOF
A
C D
FIG. 6. First vertebra ofjuvenile lake trout, Cristivomer namaycush (Walbaum), AMNH 39269 (ca.6 cm total length). A. Lateral view (anterior to left). B. Dorsal view (anterior up) to show position ofwells for accessory neural arch. C. Posterior view. D. Anterior view to show areas where exoccipitalfacets develop in larger specimens. Note large notochordal canal in centrum.
kylosed structure (Arapaima, Notopterus). InHiodon, Scleropages, and Osteoglossum,Baudelot's ligament arises on VI as opposedto its attachment to the basicranium in taxawith fused centra. That correlation is not trueof ostariophysans, however, where there isno evidence of a centrum fused to the brain-case; yet, as pointed out by Fink and Fink(1981), the ligament attaches to V1 in cypri-noids, and to the basicranium in the otherotophysans.S All one can say about this sit-
5 All one can say for the Otophysi is that the onto-genetic mechanism for transferring the ligament fromVI to the basioccipital is unknown but the characterappears to be consistent and therefore usable taxonom-ically. The alternative is that there is no such impliedcharacter transformation because the two kinds of lig-aments in the Otophysi are not homologous. In fact, ifone envisions a whole series of ligaments arising prim-itively on the basioccipital and V 1, and inserting on theshoulder girdle-and the disappearance of one or moreof these in the ontogeny of different taxa-the ligamentthat is left becomes a retained primitive character andthe absence of ligaments from certain areas, the derivedcondition. Under such circumstances, comprehensivesurvey of shoulder girdle support ligaments would haveto be made before one could use the character in a cla-distic sense. Nevertheless, I am inclined to treat the lig-aments as homologous when the insertion on the shoul-der girdle is as precisely similar as illustrated by Finkand Fink (1981).
uation in the osteoglossomorphs and elopo-cephalans is that the ontogenetic mechanismfor moving the ligament between V1 and thebasicranium is unknown. But if the positionof Baudelot's ligament in elopocephalans iscorrectly judged to be primitively V1 and isstable, then two possible explanations for itsvariable attachment are: (1) the nonhomol-ogy of the vertebral and basicranial liga-ments,4 and (2) the presence or absence of acentrum fused to the braincase, as in tSmi-lodonichthys and some coregonines (accord-ing to Cavender and Miller, 1972).In a recent account of the development of
the syncranium ofsalmonines, Jollie (1984b)states that there is no vertebra fused with thebraincase, but he did not comment on Ca-vender and Miller's (1972) paper, nor did heillustrate the presence of an accessory neuralarch between the braincase and the first cer-vical vertebra (bearing Baudelot's ligament),which appears to be a general feature of mymaterial. I cannot resolve these inconsis-tencies.An accessory neural arch is present in
primitive neoteleosts (figs. 14B, 15), and mostprimitive neoteleosts exhibit a gap betweenthe braincase and the first vertebra. This cer-vical gap is progressively smaller in more de-rived neoteleosts (figs. 14-17), being takenup by ligament or bony facets from the au-
lo NO. 2827

ROSEN: EUTELEOSTEANS
EXO
BO B
FIG. 7. Occipital views ofneurocranium. A. Coregonus artedi Lesueur, AMNH 20096 (ca. 7 cm totallength). B. Thymallus arcticus (Pallas), after Norden (1961). Note postenor opening of posterior myo-dome in A and compare with fig. 3A and apparent absence of same in B. Note also that the exoccipitalsare masked posteriorly by basioccipital condyle in A and are almost excluded from the condylar surfacein B. Compare with figures 3 and 8.
tocentrum in ctenosquamates (myctophoidsplus acanthomorphs). The complete closureof this gap is correlated with (1) the absenceof the accessory neural arch (figs. 16, 17), (2)the formation of vertebral facets for the ex-occipitals, and (3) the presence of prezyg-apophyses on the neural arches. These threefeatures are synapomorphies ofthe Acantho-morpha (cf. figs. 14-20).
Patterson (1975, p. 318) argued that theappearance of a small centrum (or part of acentrum) fused to the braincase is merely afalse impression produced by the centrumlikeossification of the notochord in the basioc-
cipital and the growth dorsally of this cen-trumlike disc to exclude the exoccipitals. Hisargument carries with it the implication thatthe tripartite arrangement of bones on theposterior face of the braincase is a primitivefeature that has been restored in neoteleostsand some primitive euteleosts by a means sofar unknown.The question ofwhich is the first and which
the second centrum in elopocephalans is, asWhitehead and Teugels (in press) remarked,"left open." Detailed histological investiga-tions of early ontogeny might "close" thequestion.
1985 I1I

AMERICAN MUSEUM NOVITATES
EXO
Bua 0
FIG. 8. Occipital region. A. Prosopium cylindraceum (Pallas), AMNH 31044 (ca. 10 cm total length),shown in three-quarter view with the arrow representing the anteroposterior axis. B. Prosopium wil-liamsoni (Girard), AMNH 37967 (ca. 27 cm total length), in posterior view, showing position ofaccessoryneural arch above basioccipital condyle. Note that, in both, the exoccipitals are masked posteriorly bybasioccipital condyle. Compare with figures 3, 7, and 13B.
12 NO. 2827

ROSEN: EUTELEOSTEANS
BOBAUDLIG
DNA
B
BO
ACCNAEXO
FIG. 9. Occipital regions in the elopomorphs, Megalops atlanticus Valenciennes (A, B) and Albulavulpes (Linnaeus) (C). A. A 10 cm total length juvenile (AMNH uncataloged). B. An 80 cm subadult,AMNH 55321. C. Subadult specimen, AMNH 21516. Note in B the ankylosis of the first vertebra withthe basioccipital and the presence of wells on its dorsal margin for the small neural arch shown in A.In C, note especially the presence of an accessory neural arch articulating between the exposed tips ofthe exoccipitals which resemble those of Thymallus (fig. 7B). Compare also with figure 10.
Neoscopelids, chlorophthalmids, and au-lopids have a large notochordal gap and an
accessory neural arch (Rosen and Patterson,1969, figs. 61-63). (Rosen and Patterson mis-
1985 13

AMERICAN MUSEUM NOVITATES
ACCNA
FIG. 10. Occipital region and first vertebra of a 6 cm Elops saurus Linnaeus, AMNH 51485. Theleft exoccipital is separated from the basioccipital to illustrate the latter's resemblance to a foreshortenedcervical vertebra (demarcated by the sculpturing around the ventral half). The accessory neural archcorresponds with a pair of dorsal indentations or wells as in Megalops, figure 9B.
takenly labeled the first of four predorsalbones in their fig. 61A as a neural arch inNeoscopelus, however.) Myctophids have aremnant of the gap which is being closed byligaments and bone from the autocentrum(figs. 16-17) in the position ofacanthomorphprezygapophyses (figs. 18-20) and they andacanthomorphs lack an accessory neural arch.These shared derived states of myctophidsand acanthomorphs, and the absence ofsamein Neoscopelus suggest that the Ctenosqua-mata should be restricted (as noted above) tothe Myctophidae and the Acanthomorpha.2The primitive position of Baudelot's liga-ment on the first cervical (fig. 17) is retainedin some primitive acanthomorphs such asamblyopsids (Woods and Inger, 1957), butBaudelot's ligament has made its way ontothe basicranium ofmany acanthomorphs, in-cluding atherinomorphs (Woods and Inger,1957). A survey needs to be conducted todiscover the anatomical position and possi-ble ontogenetic correlatives of this ligament
within the immense acanthomorph assem-blage.Thus, the tripartite joint of neoteleosts is
a very old feature and the primitive and wide-spread presence of an accessory neural archis inferred to be the remains of an ontogenythat had incorporated vertebral fusion withthe occiput. In the fossil salmonine describedby Cavender and Miller (1972), V1 is fusedto the occiput to produce a Coregonus-likesingle condylar articulation. A situation likethat in some species of Salmo and Onco-rhynchus was previously described by Gos-line (1969, fig. 7) in a species of the argen-tinoid genus Alepocephalus, but I know ofnone of the above described conditions inesocoids or ostariophysans, except to notethat in Esox the back end ofthe basioccipitalforms a disclike ossification resembling thearticular surface of a centrum (rings of acel-lular bone that constrict the notochord), whichgrows rapidly, occluding the exoccipitals fromthe joint. This disc resembles that ofOsmerus
14 NO. 2827

ROSEN: EUTELEOSTEANS
x
ol0
o
ACCNA
00~~~~~~~
a
0 / " X1
'0 00
0 00a
SACBUBAUDLIG
EXO BO
B
SACBULFIG. 11. Osmeroid occipital regions. A. Spirinchus thaleichthys (Ayres), ca. 8 cm, AMNH 51363,
showing the extent ofthe cervical gap just below an accessory neural arch. B. Osmerus mordax (Mitchill),AMNH 21727; occipital region in specimens ranging in size from 1 to 3 cm in length. In B, the dimensionsof the saccular bullae and their relation to the basioccipital condyle in a 2 cm individual in ventral vieware shown at left; in the upper row, the middle figure shows the conjunction of the exoccipital andbasioccipital; the same area appears at right in dorsal view in which articulation points for an accessoryneural arch are also shown; the same area is again shown at lower right to illustrate where the basioccipitalcondyle is deformed by the neural arch bearing exoccipitals. Compare with figure 1 2B ofa 3 cm individual.
1985 15

AMERICAN MUSEUM NOVITATES
FIG. 12. Occipital region in a salmonine and two osmeroids. A. Salmo gairdneri Richardson, AMNH40308 (ca. 6 cm). B. Osmerus mordax (Mitchill), AMNH 108093 (ca. 3 cm). C. Spirinchus thaleichthys(Ayres), AMNH 51363 (ca. 7 cm). An accessory neural arch is shown in B. In A, B, and C, there is acervical gap between the occiput and first vertebra occupied by unconstricted notochord and its connectivetissue sheath, as indicated. The basioccipital part of the vertebral joint is incompletely developed ineach. In B and C, the posterior cartilage cores of the exoccipitals are visible through the still poorlyossified, rounded basioccipital condyle; bits of the exoccipitals are visible above it and remain so toproduce an effect like that in some coregonines.
16 NO. 2827

1985 ROSEN: EUTELEOSTEANS
VINSP
V2 -
PAR
17
V3
A
\- PRx
C-000: ACCNA000 o 00
00 0.oo00a
&00 o, oo
RVINSRNAa0 o0
~000.0
.0 lb eo * 000
FIG. 13. A. Neural arches and spines of first three vertebrae of Salmo gairdneri Richardson, AMNH40508, to illustrate the absence of neural arch prezygapophyses and the extent of the ventral cartilagebase seated in dorsal vertebral wells; vertebral body of first vertebra and left half of its arch and spineomitted. B. Occipital region and cervical vertebral elements in Prosopium williamsoni (Girard), AMNH37967 (ca. 7 cm) to show a vertebralike basioccipital condyle and its association with a differentiatedneural arch without a spine. This neural arch resembles the accessory arch in other primitive euteleosts.Compare with figure 8 and figure 10 of Elops.

AMERICAN MUSEUM NOVITATES
x
FIG. 14. Occipital regions. A. Chlorophthalmus agassizi Bonaparte, AMNH 27402. B. Aulopus ja-ponicus Gunther, AMNH 28635. The cervical gap between the occiput and the first vertebra is occupiedby an unconstricted notochord and its connective tissue sheath, as indicated. In both species, theexoccipitals are exposed posteriorly above the basioccipital.
mordax, described below. Its removal ineither Osmerus or Esox would expose, not atripartite joint, but the empty interior of thebasioccipital and the formerly occluded ends
of the exoccipitals (i.e., a dual, rather thantripartite, articular surface).
Study of the occipital region of Osmerusmordax at different sizes (figs. 1 1B, 12B) (1-
18 NO. 2827

ROSEN: EUTELEOSTEANS
ACCNA
CG
/ X ~~~~~~~~CTNOT
CG PAR~~
'I ~~~~~~~~~~~~B
FIG. 15. The occipital region of Aulopus japonicus Gunther, AMNH 28635, to show the extent ofthe cervical gap between the occiput and the first vertebra and its relation to the accessory neural arch.A. Posterior quartering view. B. Lateral view. The connective tissue in the region of the gap shows aslight degree of staining with alizarine dye, indicating the presence of some calcification or ossification.This slightly stained, transparent connective tissue gap is hypothesized here to be what remains of asingle basioccipital facet that would normally occlude the exoccipitals from the posterior face of theocciput as in Elops and Prosopium (figs. 8, 10, and 13).
5 cm in standard length) suggests how the At 2 cm, neural arches are well developedneoteleostean condition might have arisen. along the vertebral axis and a small less well-
1985 19

AMERICAN MUSEUM NOVITATES
BAUDLIG
ENR
EXO FRAGMENTSA
BAUDLIG
FIG. 16. Anterior vertebrae in myctophids. A. Rhinoscopelus tenuiculus (Garman), AMNH 1915. B.Myctophum nitidulum Garman, AMNH 25022. In B, separation of Vl from the occiput caused thedense autocentral ligaments to break away with fragments from the exoccipitals. In A, a bit of neuralarch base of V1 has grown forward into such ligaments (not shown here, but see fig. 17).
formed arch is present between the occiputand VI. At 5 cm, the accessory neural archis ossified except at its dorsal and ventral tips
and it articulates ventrally with a notochor-dal-connective tissue plug in a distinct gapbetween the occiput and the first cervical ver-
20 NO. 2827
ATITT T TN-'

ROSEN: EUTELEOSTEANS
ENR
EXO
A
? PAR
EXO
B
ABAUD LIG
? PAR
FIG. 17. Myctophid occipital regions to show the extent of the cervical gap and the autocentralligaments to the exoccipitals just below prezygapophyses on the neural arches. These ligaments arehypothesized to be the primitive state of exoccipital facets (as prezygapophyses) in more derived cteno-squamate conditions. A. Myctophum obtusirostreTining, AMNH 25022. B. M. aurolaternatum Garman,AMNH 15975.
tebra (fig. 1 1, 12B). At 2 cm, the small archarticulates ventrally with a flat disc that isadherent to the basioccipital and is somewhatindented dorsolaterally by the exoccipitals.
The anteroposterior dimension of this disc isvery small (fig. 11 B), indicating that if it is acentrum, that centrum has formed onlyaround a narrow anterior part of the noto-
211985

AMERICAN MUSEUM NOVITATES
NAPZYG
, ENR
A
BAUDLIG
EXO
CG ? PAR(CTNOT) (ABAUD HG)
FIG. 18. The cervical region of Polymixia lowei Gunther, AMNH 37335. A. First two vertebraeshowing the exoccipital facets and a persisting notochordal plug on V1. B. The vertebrae in A in theirarticulated position. Note that the cervical gap is closed dorsally by autocentral prezygapophyses to theexoccipitals, but still open ventrally.
chordal sheath which is easily deformed bythe exoccipitals.As hypothesized above, the presence of an
accessory neural arch, in the absence of anunderlying centrum, is inferred to representa retention of the neural arch component ofa vertebral segment that either is incompleteor had been incorporated indistinguishablyinto the braincase.
The origin ofthe ctenosquamate triple jointappears to follow a direct course involvingthe following steps: (1) formation of a gapbetween the occiput and VI and exposure ofthe basi- and exoccipitals as attachment orarticular surfaces (figs. 14, 15); (2) loss of theaccessory neural arch in all myctophids andacanthomorphs (figs. 16-20); (3) attachmentof the dorsolateral part of V 1 with the ex-
22 NO. 2827

ROSEN: EUTELEOSTEANS
AUTNA
..- EXOC
B11BOF
FIG. 19. Holocentrid cervical anatomy. A. Occiput and first vertebra ofHolocentrus rufus (Walbaum),AMNH 35497. B. Anterior quartering view of the first vertebra of H. ascensionis (Osbeck), AMNH22006, showing the single, continuous exoccipital facet and autogenous neural arch and spine.
occipital, initially by ligament (figs. 16, 17);(4) the growth posteriorly of the exoccipitalinto this ligamentous network as in Chlo-rophthalmus and at least one myctophid (fig.16) accompanied by the growth of autocen-tral prezygapophyses toward the exoccipitals(fig. 18) and the closure of the gap by thebasioccipital and the body of the centrum
(figs. 19, 20); and (5) finally the full devel-opment of bone-to-bone condylar articula-tions between the occipital region and facetson VI.
Primitively, V1 in ctenosquamates has aneural arch with a ventral cartilage tip seatedin dorsal depressions in the centrum (figs. 4-6), or is at least sutured, rather than anky-
231985

AMERICAN MUSEUM NOVITATES
ENR
PR
BOC \ %RET DORSBOF TENDON
FIG. 20. Holocentrid cervical anatomy. A. Holocentrus rufus (Walbaum), AMNH 35456, first twovertebrae, showing, in lateral view, the exoccipital facet and autogenous neural arch and spine on V1.B. H. rufus, AMNH 35477, showing the first two vertebrae in normal articulation with the occiput. Thetendon for the retractor dorsalis muscle is shown on V2 in A and B.
losed with the autocentrum. There is somequestion about the generality of that laststatement, since my specimen of adult Myc-tophum aurolaternatum (fig. 17B) shows thesuture line clearly, whereas Jollie's (1954) fig-ure 22 of the first vertebra of Lampanyctus
leucopsarus shows the arch to be completelyankylosed with the centrum. Yet anothermyctophid, Rhinoscopelus, is consistent withJollie's figure ofLampanyctus (fig. 16A). Theneural arch of V1 is unfused in holocentrids(figs. 19, 20) and primitive "percoids" such
24 NO. 2827
ROSEN: EUTELEOSTEANS
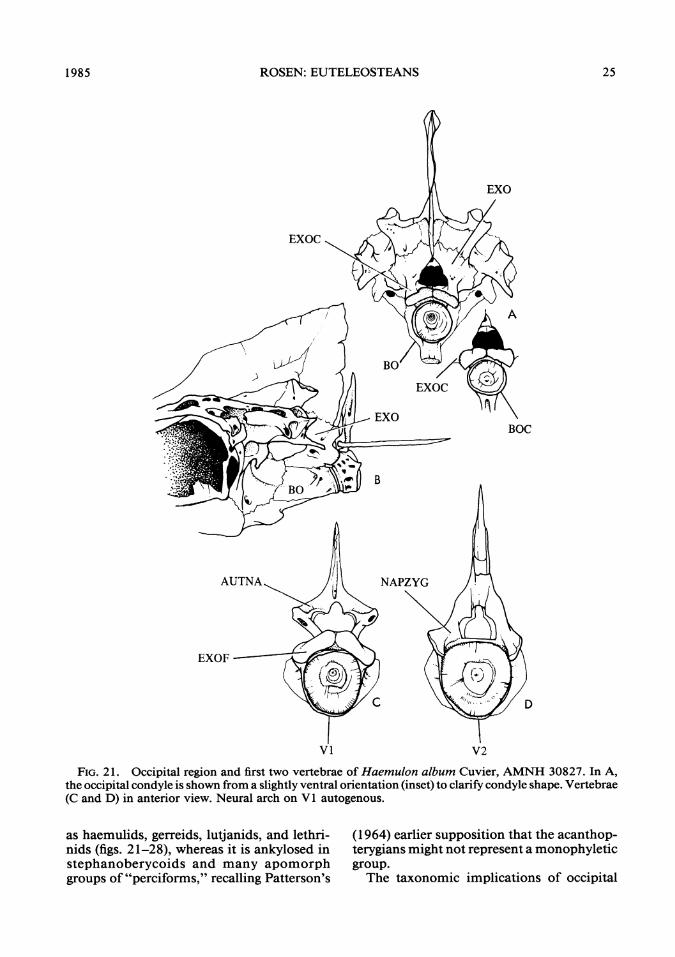
FIG. 21. Occipital region and first two vertebrae of Haemulon album Cuvier, AMNH 30827. In A,the occipital condyle is shown from a slightly ventral orientation (inset) to clarify condyle shape. Vertebrae(C and D) in anterior view. Neural arch on VI autogenous.
as haemulids, gerreids, lutjanids, and lethri-nids (figs. 21-28), whereas it is ankylosed instephanoberycoids and many apomorphgroups of "perciforms," recalling Patterson's
(1964) earlier supposition that the acanthop-terygians might not represent a monophyleticgroup.The taxonomic implications of occipital
1985 25

AMERICAN MUSEUM NOVITATES
EXO
D
VI V2
FIG. 22. Occipital region and first two vertebrae of Gerres cinereus (Walbaum), AMNH 21732. InA, the occipital condyle (inset) is shown from a slightly ventral orientation to clarify condyle shape. B.Lateral view as in A. Vertebrae (C and D) in anterior view. Neural arch on V1 autogenous.
anatomy do not stop there, however. Prim-itively, the exoccipital condyles meet in themidline just above the notochordal canal. Inholocentrids (fig. 29A), they are united by a
delicately interdigitating suture (fig. 29A). Asimilar arrangement has also been found intrachichthyids and berycids. Reference toPatterson's (1964) account of the acanthop-
26 NO. 2827

ROSEN: EUTELEOSTEANS
AUITNA
EXOFC
BOF
FIG. 23. Occipital region (A,arch on Vl autogenous.
B) and first vertebra (C) of Pomatomus saltatrix (Linnaeus). Neural
terygian fishes of the English Chalk showsthat the pattern formed by these two condylesis very old and occurs in such primitive crea-tures as tAulolepus typus. The arrangementproduced by these condyles is best described
as an elongate rectangle that forms a concavearc posteriorly. A single convex facet on thefirst cervical vertebra of similar shape artic-ulates with these medially united condyles.The exoccipital condyles retain this basic form
1985 27

AMERICAN MUSEUM NOVITATES
EXO
EXOC
A
BO
PZYGF(EXOF)
FIG. 24. Occipital region and first two vertebrae ofLutjanus campechanus (Poey). A. AMNH 21632.B. AMNH 21688. In B, the autogenous neural arch ofVI had fallen offand was lost during preparation.
and remain medially united in a variety offishes that have been regarded as "basal per-coids" (see figs. 21-28, 29B, C). Thus, the"percoid" condition includes two primitive
features that are shared with at least some"berycoids" (e.g., holocentrids): the mediallyunited condyles forming a long concave rect-angle and an autogenous neural arch on the
NO. 282728

ROSEN: EUTELEOSTEANS
DNA
DNA
BOF PZYG
FIG. 25. Anterior (A) and lateral (B) views of the first vertebra of Lutjanus campechanus (Poey),AMNH 21632. The autogenous neural arch was removed to show the articular wells on the dorsalsurface.
first cervical. Derived transformations ofthese conditions involve lateral displacementofthe condyles accompanied by an alterationof their rectangular shape and the shape andposition of the articular facets on the firstcervical vertebra (figs. 29D, 30-32), and an-kylosis of the neural arch with the centrumof this vertebra (figs. 33, 34). A comprehen-sive survey needs to be made before the taxo-nomic usefulness of this area can be judgedfully, but, in at least two cases, modificationsofthe occiput and first vertebra suggest affin-ities between major fish groups that were ma-jor components of the Paracanthopterygii(figs. 35-39). In each of the batrachoid-lo-phiiform and ophidiiform-gadiform groups,the exoccipital condyles primitively recededfrom the posterior occipital margin and con-sist ofwidely separated, cartilage-filled tubesto which prezygapophyses from the first cer-vical articulate. In all of these fishes the neu-ral arch and spine ofthe first cervical vertebrais ankylosed to the centrum and both arefirmly joined to the back end of the skull.THE ROSTRAL CARTILAGE: The rostral car-
tilage is another feature that has been givenprominent attention as a neoteleostean syn-apomorphy. Here again, Fink and Weitzman(1982) have found paired elements in somesalmonines and one coregonine that are smalldiscs of cartilage affixed to the inner face of
the premaxillae (fig. 40A), and hypothesizedthat the elements are a synapomorphy of sal-monids and neoteleosts. But not all salmo-nids, or even salmonines, have such carti-lages (I have not found one in any species ofSalvelinus) and at least one osmeroid, Os-merus mordax, does have them (fig. 41A).Among neoteleosts, however, Chlorophthal-mus has them well developed in connectionwith a series of premaxillary processes (figs.40C, 41B) almost certainly synapomorphicfor a group defined previously as the Euryp-terygii (Rosen, 1973). One stomioid also hasa Chlorophthalmus-like upper jaw with asimilar, though slight, indication of cartilagedevelopment. By this, I mean that it stainsprominently for mucopolysaccharides withalcian blue, as does cartilage, and the stainedarea appears to contain a few cells as well asconnective tissue fibers (fig. 40B). This sto-mioid is Maurolicus muelleri, but Fink andWeitzman (1982) treat this as homoplasiouson the grounds that Maurolicus is a derivedmember ofan apomorph group of stomioids(the sternoptychids)-ruling out the possi-bility of a retained primitive feature.They claimed, however, that in Diplophos
there is a rostral cartilage which, in their view,corroborates the placement ofstomioids withneoteleosts. What they illustrated in this in-stance, though, is a flat, median domino-
1985 29

AMERICAN MUSEUM NOVITATES
EXO
EXOC
BOC
NAPZYG
D
FIG. 26. Occipital region and anterior two vertebrae ofLethrinus sp., AMNH 30872. InA the condylesare shown from a slightly ventral orientation (inset) to clarify condyle shape. Neural arch of VI autog-enous.
shaped element not firmly bound to thepremaxillaries or oriented like the medianelement in neoteleosts. But S. Weitzman in-forms me (in litt.) that the cartilage is actually
a long cylinder surrounded by connective tis-sue, so that as redescribed it still does not fitthe anatomical requirements of a neoteleos-tean rostral cartilage.
NO. 282730

ROSEN: EUTELEOSTEANS
- EXOC
BOC
EXO
BO
EXOF
BOF
FIG. 27. Occipital region and anterior two vertebrae of Seriola sp., AMNH 30856. In A, the condylesare shown from a slightly ventral orientation (inset) to clarify condyle shape. Neural arch of VI autog-enous.
Nevertheless, Fink and Weitzman (1982)deserve much credit for making these initialobservations ofthe cartilaginous skeleton andthereby opening up new avenues for profit-able research.The manner in which paired premaxillary
cartilages transform into the median elementofneoteleosts is suggested by a series of struc-tures found amongst neoscopelids and somectenosquamates (figs. 24-25). (K. Sulak, per-sonal commun., informs me that paired car-tilages occur also in Ateleopus, a fish that has
311985

AMERICAN MUSEUM NOVITATES
B
AUTNA NAPZYG
ENR
FIG. 28. Occipital region and anterior two vertebrae of Archosargus probatocephalus (Walbaum),AMNH 21663. In A, the condyles are shown from a slightly ventral orientation (inset) to clarify condyleshape. Neural arch of VI autogenous.
defied, placement so far-see comments inRosen, 1973.) But within neoscopelids (figs.42, 43A), the paired elements retain their firmassociation with the premaxillary ascendingprocesses and a median element with whichthey become fused in Neoscopelus (fig. 42B)
appears to form as an extension of chondri-fication into the interpremaxillary ligament,or perhaps is best described as a sesamoidcartilage that develops in response to thestresses on that ligament. In all events, inNeoscopelus, what appear to be three distinct
32 NO. 2827

ROSEN: EUTELEOSTEANS
BO
FIG. 29. Comparison of primitive and derived acanthopterygian occipital regions. A. Holocentrusascensionis (Osbeck), AMNH 22086, showing the planar exoccipital condyles suturally united medially.B. Centropomus undecimalis (Bloch), AMNH 28058, showing the laterally expanded exoccipital condylesretaining the medial suture. C. Morone chrysops (Rafinesque), AMNH 22528, showing condition as inB. D. Sebastes sp., AMNH 36935, showing a more derived condition in which the exoccipital condylesare displaced laterally.
cartilages become one, and apparent evi-dence of a tripartite history may still be de-tected in the structure of some other cteno-squamates (figs. 43B-D). Aulopus (fig. 41C)appears to have a rostral cartilage that in pos-
terior view resembles a somewhat reducedversion ofthe Neoscopelus structure (fig. 43A).
If the foregoing analysis is correct, a newcladogram for the Euteleostei is needed. Finkand Weitzman wished to extend the resolved
1985 33
AMERICAN MUSEUM NOVITATES
AUTNA
EXOF
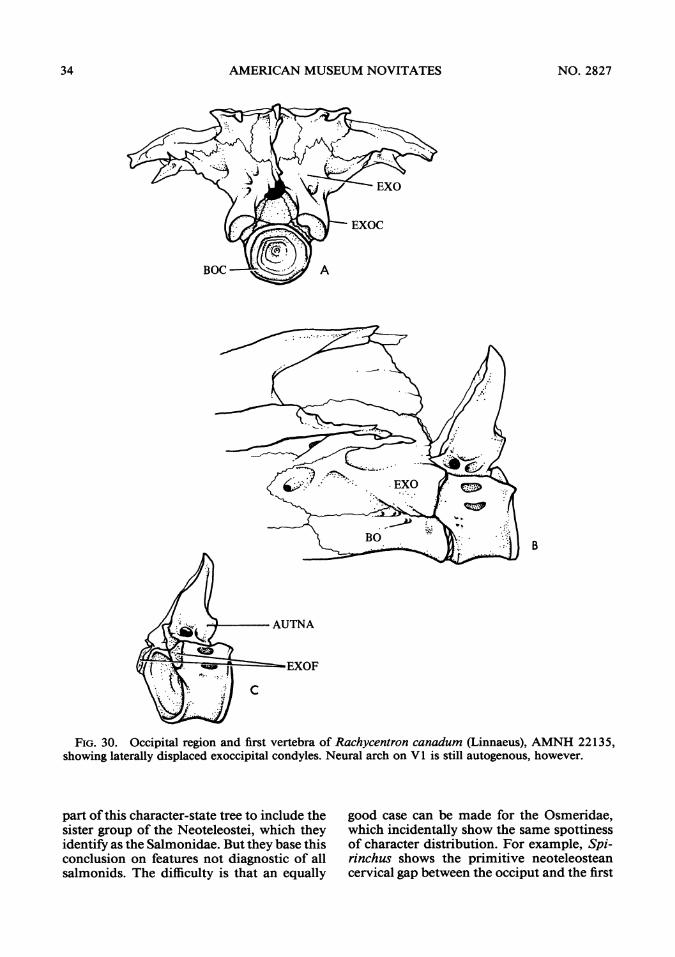
FIG. 30. Occipital region and first vertebra of Rachycentron canadum (Linnaeus), AMNH 22135,showing laterally displaced exoccipital condyles. Neural arch on V1 is still autogenous, however.
part ofthis character-state tree to include thesister group of the Neoteleostei, which theyidentify as the Salmonidae. But they base thisconclusion on features not diagnostic of allsalmonids. The difficulty is that an equally
good case can be made for the Osmeridae,which incidentally show the same spottinessof character distribution. For example, Spi-rinchus shows the primitive neoteleosteancervical gap between the occiput and the first
34 NO. 2827

ROSEN: EUTELEOSTEANS
B
EXOF
BOF
FIG. 31. Occipital region and first two vertebrae in Arripis trutta (Bloch and Schneider), AMNH21632, showing laterally expanded exoccipital condyles just barely in contact in midline. Neural archon Vl autogenous.
vertebra (figs. 1 1A, 12C) and has, as a con-
sequence, exoccipitals that have exposed pos-terior condyle-like faces, and Osmerus mor-dax has the paired premaxillary cartilages (fig.41A) developed to the same extent as theyare in Prosopium williamsoni (the only cor-
egonine I have seen that shows the condi-tion). But if we must make a choice fromamongst the old "salmoniforms" for a neo-teleostean sister group, then the Osmeroidei,or at least Spirinchus, is a better choice thansalmonids on three counts:
351 985

AMERICAN MUSEUM NOVITATES
EXO
BO
NAPZYG
EXOF
D
FIG. 32. Occipital region and first two vertebrae in Coryphaena hippurus Linnaeus, AMNH 21750,showing laterally displaced exoccipital condyles and a still autogenous neural arch on V1.
1. All osmeroids have a modified neuralspine definitely associated with the secondpreural centrum which is reduced in heightin virtually all cases and fitted with laminarbone fore and aft, resembling this spine inaulopids. Salmonines, in contrast, have a full,strong spine on the second preural centrum
(see Vladykov, 1962, figs. 1-3) when the as-sociation ofspine and centrum is ofa definitesort [although the spine is short in corego-nines (fig. 44) and might be primitive for theSalmonidae if that group is monophyletic].IfFink and Weitzman (1982) and Fink (1 984)are correct in aligning galaxioids with os-
36 NO. 2827

ROSEN: EUTELEOSTEANS
meroids, then a similar problem arises in theosmeroid complex since galaxioids, but notosmeroids, have a full spine on PU2. Whichstate is primitive for such an osmeroid com-plex?6
2. The premaxillary has an alveolar processof sorts (fig. 41A) under which there are nomaxillary teeth (in other words, they show asimple tandem arrangement of these bonesas opposed to their more primitive serialalignment in salmonids (fig. 40A).
3. The osmeroids (as defined by Fink andWeitzman) so far studied (a species of Os-merus and Galaxias) have acellular bone incommon with neoteleosts (Parenti, MS),whereas the salmonids studied have a mix-ture of primitive cellular and some acellularbone. Thus, the relatively smaller, usually def-initely shortened spine in osmeroids, a sim-ple form oftandem upperjaw bone alignmentand acellular bone, are synapomorphies thatosmeroids, but not salmonines, share withthe Neoteleostei.
CHARACTER CONFLICrS
The evidence gathered so far for overalleuteleostean classification involves a number
6 Fink and Weitzman (1982) and Fink (1984) arguedfor a linkage between "osmeroids" and galaxiids mainlyon the grounds that both possess a distinct row of teethdorsomedially on the mesopterygoid. Pterygoid teeth areprimitive for teleosts, and when the mesopterygoid patchis reduced, it usually leaves such a row in the dorso-medial position (e.g., in Pterothrissus gisu and the EocenetDiplomystus dentatus). Such teeth also are present insome primitive neoteleosts such as synodontids (K. Su-lak, personal commun.). Galaxiids show other plesio-morphous features not occurring in "osmeroids." Theseinclude three, and in some cases, four small, accessoryrays in advance of the dorsal fin. And, in the caudalskeleton, neural arches, and often spines, are associatedwith the first ural and first preural centra (cf. Rosen,1974, figs. 18, 19, and illustrations in Patterson and Ro-sen, 1977). Small, but well-ossified, posteriorly cocked,neural arches also occur in some "salmonid" species,especially coregonines, directly over or adjacent to thefirst preural centrum and on both the first ural and firstpreural in a variety of primitive teleosts. These are partof a derived caudal feature of tailed elopomorphs (Pat-terson and Rosen, 1977, cf. figs. 23, 27, 35, 36). Fink's(1984) summary ofthe problems of"lower" euteleosteanclassification is an exemplary statement illustrating theneed for comprehensive character surveys.
of inconsistencies, some more obvious thanothers, involving the rostral cartilage, the oc-cipital region ofthe skull, the caudal skeleton,and the muscle that retracts the dorsal gillarch elements (discussed below).ROSTRAL CARTILAGE: Among primitive
ctenosquamates there is a conflict arising fromthe fact that myctophids, but not neoscope-lids, always have a single median cartilageand a subocular shelf, as in Acanthomorpha.This conflict is resolved by simply admittingthat the group Myctophidae + Neoscopeli-dae = Myctophiformes, is unnatural, andshould be so represented in the cladogram(fig. 45). Stiassny (Ms) treats the group as nat-ural based on several synapomorphies, how-ever.2OCCIPITAL REGION: The capricious manner
in which the tripartite occipital joint is pres-ent or not (except within the Neoteleosteiwhere it is diagnostic) has already been al-luded to, and has several possible taxonomicimplications: (1) the Salmonidae is non-monophyletic; (2) the Osmeridae is nonmon-ophyletic; (3) the Clupeiformes and/or Clu-peomorpha are nonmonophyletic, althoughthe significance of a triple joint in Denticepsis overriden by the congruence ofmany othercharacters (Grande, MS); (4) the presence ofa tripartite joint in Lepidogalaxias (Fink,1984) might be one more example of char-acter capriciousness ifRosen (1974) was cor-rect in linking that genus with the pikes andmudminnows.7THE RETRACTORES ARCUUM BRANCHI-
ALIUM MUSCLE: It has been known for some
7Fink (1984) supports his argument that Lepidoga-laxias is allied to neoteleosts by his discovery ofa dorsalretractor muscle in this fish. And Fink and Weitzman(1982) disagree with my earlier (1974) alignment of itwith esocoids, partly, and I think correctly, on the groundsthat my comparison of anterodorsal outgrowths of itsfirst uroneural with those of esocoids leaves a great dealto the imagination. An alternative placement of Lepi-dogalaxias is suggested by the resemblance ofits cephalicsensory pit-lines with those of Dallia (Nelson, 1972, p.38). A case might also be made for its original placementas a galaxiine (see Rosen, 1974) based on dorsal andcaudal fin anatomy and position, or even with its align-ment to a Novumbra-Dallia esocoid subgroup definedby having but a single epural and uroneural in the caudalskeleton. Fink's (1984) statement that the "position ofLepidogalaxias is controversial" is unarguable.
371985

AMERICAN MUSEUM NOVITATES
ANA
EXOF
D
FIG. 33. Occipital region and first two vertebrae in Echeneis naucrates Linnaeus, AMNH 21844,showing enlarged lateral exoccipital condyles and the neural arch on VI ankylosed.
time that the retractor dorsalis, as this musclehas been called, has a sporadic occurrenceamong halecomorph fishes, which Rosen(1973) commented upon earlier. Most re-cently, Fink (1984) has identified one in Lep-idogalaxias salamandroides. The problem ofthe homology of these different retractor
muscles has been investigated in some detailin relation to the monophyly at the Aulopi-formes by Stiassny (work in progress), whoseconclusions are consistent with my own pres-ent and prior observations. A difficulty incharacter interpretation arises when this fea-ture is used if one considers that Aulopus,
38 NO. 2827

ROSEN: EUTELEOSTEANS
BOC
a_~~~1JANA
BOFFIG. 34. Occipital region and first vertebra of Strongylura marina (Walbaum), AMNH 27805, show-
ing laterally displaced exoccipital condyles and the neural arch on VI ankylosed.
Chlorophthalmus, and neoteleosts, in gener-al, are characterized by a very short retractorthat inserts on the inner faces ofthe third andfourth pharyngobranchials, as opposed to themore posterior insertion in some stomi-iforms and alepisauroids on the fourth pha-ryngobranchial, which often bears a singlelarge toothplate with recurved, fanglike teeth.Stomiiforms have a variety of specialized re-tractor origins, including the shoulder girdleand ribs as in sternoptychids but the condi-tion in Aulopus, Chlorophthalmus, and myc-tophids with the anterior origin and doublepharyngeal insertion appears to be an ad-vanced neoteleostean synapomorphy. Here Isimply defer to Stiassny's work in progresswhich will deal with the analysis of this char-
acter in more detail. It appears possible thatstomiiforms and alepisauroids are linked bya peculiar type of dorsal retractor, althougheach of these groups are individually diag-nosable, the stomiiforms by the peculiar typeof photophores (Fink and Weitzman, 1982)and at least some alepisauroids by the ex-tremely attenuated second pharyngobranchi-al described by Rosen (1973).THE POSITION OF AuLoPus AND IMMEDI-
ATELY ALLIED FoRMs: The title of this sub-section is phrased somewhat ambiguously fora good reason-no one knows with any de-gree of certainty what those immediately al-lied forms might be, but perhaps they arebathysaurids, bathypteroids, and ipnopids.But whoever they are, later collectively termed
1985 39

AMERICAN MUSEUM NOVITATES
F VIANA
N Y XPZYG
/~~~~VI ANA-EXO'
BO
VIANA ~ ViAN
NAPZYG ~ ~ ~ ~ I N
EXO ~ ~ ~ ~ X
BO~ ~ ~ ~ ~ ~ B
E=0
FIG. 35. Comparison ofthe occipital region and anterior vertebrae in a gadiform and some membersof the batrachoid-lophioid clade. A. Gadus morhua Linnaeus, AMNH 21680. B. Porichthys notatusGirard, AMNH 22432. C. Opsanus tau (Linnaeus), AMNH 21564. D. Lophius americanus Valenciennes,AMNH 22129. Note integration of neural arch on Vi with the occipital bones, the sharply angledautocentral prezygapophyses, and their integration with a complex exoccipital condyle. Both the V1prezygapophysis (exoccipital facet) and the exoccipital condyle are formed around cores of cartilage sothat the condyle-facet contact is cartilage-to-cartilage. Compare with figures 36 to 38.
"aulopoids," at least Aulopus may be treated basis of its (1) high-set pectorals, (2) subtho-as a sister group to the Ctenosquamata on the racic pelvics, (3) ctenoid scales, (4) three pre-
40 NO. 2827

ROSEN: EUTELEOSTEANS
ENR
ENR
PZYG
FIG. 36. Occipital region and anterior vertebra in "ophidiiforms." A. Ophidion holbrooki (Putnam),after Rose (1961). B. Ogilbia cayorum Evermann and Kendall, AMNH 26098. A, an ophidioid has arelatively primitive, planar exoccipital facet whereas B, a bithytoid, has paired, codlike facets. Both showthe more posterior position of the basioccipital facet.
dorsal (or supraneural) bones, (5) a reducedspine on NPU2, (6) derived premaxillarymorphology (see Rosen and Patterson, 1969),(7) type of dorsal retractor muscle, (8) atoothplate fused to the third epibranchial(Rosen, 1973), and (9) median rostral carti-lage.STATUS OF THE PARACANTHOPTERYGII: Of
all the proposals for subdividing the oldAcanthopterygii or Percomorpha, none hasbeen more controversial, or, in my presentview, ill-fated, than the Paracanthopterygii.The taxon was proposed partly in an effortto narrow the enormous scope of the taxo-nomic problem posed by 6000 or so species
of spiny-finned euteleosts, and of advancinga specific proposal that could be addressed,or even openly attacked. There was not longto wait after the initial publication ofthe 1966general classification of modem teleosts. Infact, its first reviewer, Carl Hubbs, referredto it as a bizarre collection ofodd bedfellows.Rosen and Patterson (1969) rooted the
problem firmly by producing a monographwhose sole, and now seemingly unfortunatepurpose, was to entrench the problem by col-lecting "confirmatory" evidence of paracan-thopterygian substance. Now, the spirit ofthetimes has changed, and neither Patterson norI would consider that a worthy purpose. In
1985 41

AMERICAN MUSEUM NOVITATES
W EXOC
BOC A
C
BOCFIG. 37. Occipital regions of a gadiform and members of the batrachoid-lophiiform clade. A. Gadus
morhua Linnaeus, AMNH 21680. B. Porichthys notatus Girard, AMNH 22432. C. Opsanus tau (Lin-naeus), AMNH 21564. D. Lophius americanus Valenciennes, AMNH 22129. Note the small, laterallydisplaced, cartilage-filled, tubelike exoccipital condyles. In D, the cartilage core is hidden by lateral wingsof bone.
fact, my object now is to show that Hubbs'criticism was well taken, and not only thatthe paracanthopterygians no longer can beaccepted as a natural group, but to point out
that some of its constituents can no longerbe accepted as natural. I refer particularly tothe percopsiforms that have never had as de-fining traits anything but shared primitive
42 NO. 2827
N.

ROSEN: EUTELEOSTEANS
B
BOF
EXOF
C
BOF
FIG. 38. The first vertebra in a gadiform and two batrachoids to illustrate the very derived autocentralprezygapophyses. A. Gadus morhua Linnaeus, AMNH 21680. B. Porichthys notatus Girard, AMNH22432. C. Opsanus tau (Linnaeus), AMNH 21564.
features. Aphredoderids do align themselveswith amblyopsids on the basis ofthe thoracicanus and segmented premaxilla, but nothingof which I am aware properly unites themwith percopsids. In fact, I find it difficult evento state features which link the living and fewfossil percopsids-apart from the broadlyarched alveolar premaxillary process.
Percopsids are, of course, euteleosts be-cause they possess that group's single definingtrait, the adipose fin. And they are, presum-ably, some form of primitive ctenosquamatebecause of their premaxillary processes, ros-tral cartilage, pectoral fin position, and finspines. At least one of them, Percopsis omis-comaycus, is known to have a complex series
431985

AMERICAN MUSEUM NOVITATES
ENR
PZYG
PZYG
C
FIG. 39. Comparison of first two vertebrae in a gadiform and two acanthopterygians. A. Gadusmorhua Linnaeus, AMNH 21680. B. Centropomus undecimalis (Bloch), AMNH 28058. C. Sebastes sp.,AMNH 36935. Note that exoccipital facets are ofautocentral origin, following the angle of a neural archprezygapophysis on either VI or V2.
ofjaw muscles found also in some gadoids,but as Dietz (1914) pointed out, so do someliparids. The one thing they share that is de-rived in relation to current pharacter inter-pretation is, a full spine on PU2, but thatcharacter seems to come and go with such
frequency that investigators would be fool-hardy to base major taxonomic judgmentsupon it unless we could formulate an argu-ment involving a unique ontogeny that doc-uments the redevelopment of a full NPU2.No such empirical ontogenetic data yet exist.
44 NO. 2827

ROSEN: EUTELEOSTEANS
PMX CART
PMX .* B
PMX CARTASC PR
ARTPR
/ PTMX PR
PMX MX
FIG. 40. Euteleostean upper jaw bones in medial view. A. Salmo gairdneri Richardson, AMNH40268, to show small, adherent, premaxillary cartilage. B. Maurolicus muelleri (Gmelin), AMNH 37329.C. Chlorophthalmus agassizi Bonaparte, AMNH 40812. Note the serial (qnd-to-end) alignment of thetwo bones in A and their tandem (overlapping) alignment in B and C. Note also the distribution andshape of premaxillary processes in B and C. Cartilage, showni by the presence of black dots, is inferredin B because of specific alcian blue staining for mucopolysaccharides and the presence ofsome cellulanrtywithin the fibrous tissues engulfing the ascending process. Compare C with figure 4113.
All "percopsiforms" are primitive with re-,spect to the presence of an unconsolidatedsecond ural centrum, an adipose fin, and morethan 15 branched caudal rays. They are de-
rived in 'having enlarged infraorbital canalbones not supporting a subocular shelf, butthe latter might also be primitive or simplyhomoplasious since similar infraorbitals oc-
1985 45

AMERICAN MUSEUM NOVITATES
PMX CARTI- /
MX
A
R CART
PAL
B
C
FIG. 41. Euteleostean upperjaw bones in dorsal view. A. Osmerus mordax (Mitchill), AMNH 40726.B. Chlorophthalmus agassizi Bonaparte, AMNH 40892. C. Aulopusjaponicus Gunther, AMNH 28635.Note paired premaxillary cartilages in A and B; those in A are similar to ones observed in Prosopiumwilliamsoni and P. cylindraceum, except that in the latter two taxa the cartilages are only loosely associatedwith the premaxillae rather than firmly adherent to them as in S. gairdneri (fig. 40A) or 0. mordax (A).The cartilage depicted in C is similar to that shown in figure 43A.
cur in stephanoberycoids. In their dorsal gillarches they lack an interarcual cartilage, orig-inally shown sometime ago to be present inother primitive ctenosquamates such as myc-tophids by Malcolm Jollie (1954) in a sadly
underused thesis ofconsiderable breadth andinsight. The interarcual cartilage has sincebeen found to be absent in other myctophids,however (Stiassny, MS). As for the polymix-iids, the most interesting thing I can think of
46 NO. 2827
ROSEN: EUTELEOSTEANS
PMX
MED CART
B
- MED CART
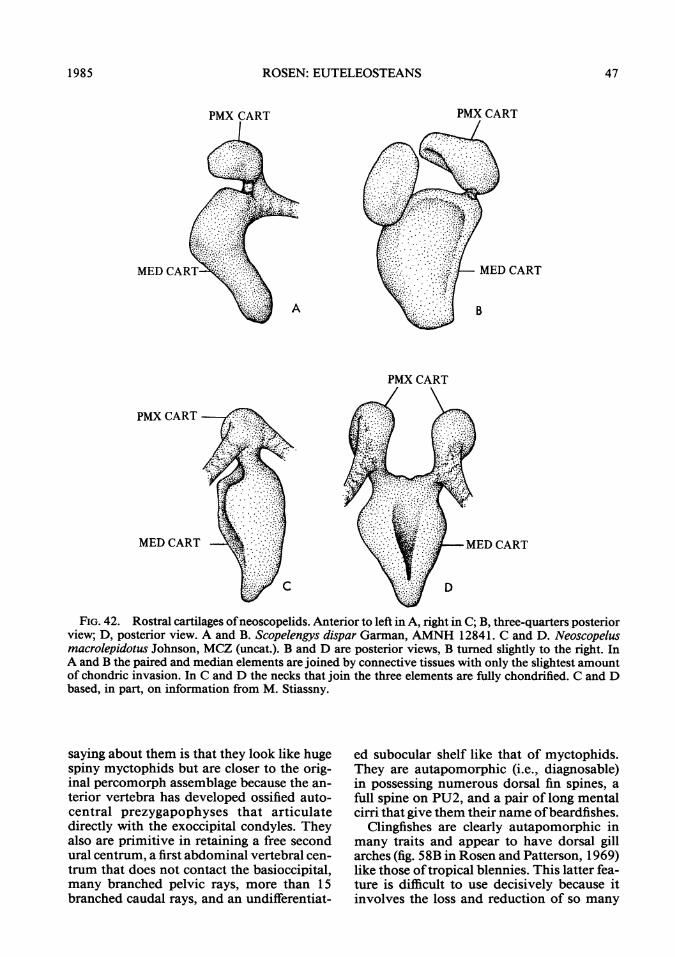
FIG. 42. Rostral cartilages ofneoscopelids. Anterior to left in A, right in C; B, three-quarters posteriorview; D, posterior view. A and B. Scopelengys dispar Garman, AMNH 12841. C and D. Neoscopelusmacrolepidotus Johnson, MCZ (uncat.). B and D are posterior views, B tumed slightly to the right. InA and B the paired and median elements are joined by connective tissues with only the slightest amountof chondric invasion. In C and D the necks that join the three elements are fully chondrified. C and Dbased, in part, on information from M. Stiassny.
saying about them is that they look like hugespiny myctophids but are closer to the orig-inal percomorph assemblage because the an-terior vertebra has developed ossified auto-central prezygapophyses that articulatedirectly with the exoccipital condyles. Theyalso are primitive in retaining a free secondural centrum, a first abdominal vertebral cen-trum that does not contact the basioccipital,many branched pelvic rays, more than 15branched caudal rays, and an undifferentiat-
ed subocular shelf like that of myctophids.They are autapomorphic (i.e., diagnosable)in possessing numerous dorsal fin spines, afull spine on PU2, and a pair of long mentalcirri that give them their name ofbeardfishes.
Clingfishes are clearly autapomorphic inmany traits and appear to have dorsal gillarches (fig. 58B in Rosen and Patterson, 1969)like those oftropical blennies. This latter fea-ture is difficult to use decisively because itinvolves the loss and reduction of so many
A
MED CART-
PMX CART
MED CART
1985 47

AMERICAN MUSEUM NOVITATES
R CART
(A
R CART
A D
FIG. 43. Ctenosquamate rostral cartilages. A. Neoscopelus macrolepidotus Johnson, MCZ (uncat.)(from a sketch by M. Stiassny). B. Notoscopelus resplendens Richardson, AMNH 29528. C. Polymixialowei Gunther, AMNH 49674. D. Scopeloberyx sp., AMNH 40268. Note that indications of a tripartiteorigin (as per fig. 42) are present in all. A, B, C (left), and D (right) are posterior views. C (right), a dorsalview, anterior to left. D (left), a lateral view, anterior to left. Fink (1984) has reported that in a cichlidspecies (Acanthopterygii), the rostral cartilage has only a dual origin ontogenetically from paired pre-maxillary cartilages.
gill arch elements; their relationship to dra- gested by Gosline (1971). The latter (i.e., cal-conettids and callionymids has been sug- lionymids) have similar dorsal gill arches. The
48 NO. 2827

ROSEN: EUTELEOSTEANS
UN 1 EP
HYP 6
,P HYP 5
FIG. 44. Caudal skeleton of Prosopium williamsoni (Girard), AMNH 37967. The small, recurved,neural arch and spine element on PU2 appears to be a derived feature that occurs in coregonines, andsome salmonines and is variably associated with the first or second preural centrum (PU1 or PU2).Within the Salmoninae a usual condition is to have a full spine associated with PU2, whether or not areduced, recurved element joins it on that centrum. The primitive eurypterygian neoteleost condition,however, is to have a spatulate or broadly lance-shaped, somewhat or greatly reduced spine on PU2(Rosen, 1973, figs. 46 to 48) and this is true also of osmeroids (excluding salangids) (e.g., Rosen, 1974,figs. 25, 26C, 27 and Greenwood and Rosen, 1971, fig. 16). More primitive neoteleosts such as stomi-iforms have usually either a full spine on PU2 (based on a random survey of eight gonostomatids andsternoptychids) or, if somewhat shorter than that on PU3 (e.g., Diplophos, as figured by Fink andWeitzman, 1982), it is a narrow spine without platelike expansions. Spines like that of Diplophos occuralso in some of the more derived paralepidids and evermanellids and full spines on PU2 occur in avariety ofacanthomorphs (e.g., in the "paracanthopterygii" and some acanthopterygians). The conditionshown here for Prosopium might, therefore, serve as a synapomorphy for salmonids.
similarity does not, however, include theclosely united third and fourth epibranchials,the converging ventral gill arches, and unos-sified copula that clingfishes share with lo-phiiforms or, at least, lophiids and anten-nariids (fig. 58C in Rosen and Patterson,1969), but not with callionymids.What appear to unite the batrachoid clade
with the gadiforms are derived features ofthe
occipital region discussed above, and somefeatures of the dorsal gill arches to be pre-sented in a forthcoming paper.
Features that appear to unite batrachoids,lophiiformes, gadiforms, and bythitoids arethe relation between the occipital region andthe first vertebra. The contact between theexoccipital facets and the prezygapophyses ofthe first vertebra is between the cartilage cores
1985 49

AMERICAN MUSEUM NOVITATES
37 - 38
12 -14
11-5
FIG. 45. Proposed interrelationships of the main groups of clupeocephalans, based on the synapo-morphies as numbered in the text. Uncertainties about the placement of a number of groups are rep-resented by unresolved polychotomies. This scheme differs from that proposed by Fink and Weitzman(1982) and Fink (1984) mainly in the position of salmonids and "osmeroids" and in excluding Lepi-dogalaxias.
or tips of these elements (figs. 35, 36B, 37,38, 39A), as compared with the bone-to-bonecontacts in acanthopterygians and the bery-coid-like arrangement in ophidioids (cf. figs.19, 20, and 29A with 36A). The other char-acter which also includes ophidioids as wellas bythitoids, is the articulation of pleuralribs with ventrolateral cavities in the verte-brae normally occupied by parapophyses.The feature that might be synapomor-
phous for gadiforms and at least some ophid-iiform subgroups is the position of the ex-occipital facets anterior to the basioccipitaland the corresponding anterior extension ofthe first vertebral prezygapophyses onto theback of the occiput to meet the exoccipitals.Neural arches are carried forward along withthe prezygapophyses, in some cases firmlyincorporated into the exoccipitals and the su-praoccipital (figs. 47-49A). These vertebralfeatures, alone, exclude zoarcids and gobie-
socids from the assemblage since they haveno derived chondrification associated withthe exoccipitals and vertebral prezygapoph-yses.As mentioned earlier, it is primitive for
neoteleosts to lack a direct vertebral contactwith the occiput, and it is this spatial sepa-ration that seems most closely correlated withthe presence of the exoccipitals in the occip-ital joint region. What characterizes thectenosquamates is a gradual closing of thisspace so that acanthomorph fishes, excludingPolymixia, have the first vertebra firmlyunited with the occiput by one of two means(see, e.g., an atherinomorph, fig. 34). One isunique to acanthomorphs and this is the for-mation of autocentral prezygapophyses thatgrow forward to contact the exoccipital con-dyles. The other is typical ofseveral primitiveacanthomorphs (holocentrids, berycids, andophidioids, for example). Here, the articu-
50 NO. 2827

ROSEN: EUTELEOSTEANS
latory surface on the first vertebra for theexoccipitals is a more-or-less continuous,planar surface (figs. 19B, 36A) that fits neatlyagainst an opposing exoccipital surface clear-ly divided by suture into right and left halves(fig. 29A).
In fishes that have been previously calledacanthopterygians or percomorphs, this typeof contact has been altered by the lateral dis-placement of the two exoccipital facets andtheir growth backward over the basioccipital(as in Centropomus and Sebastes, fig. 29B,D). What characterizes the batrachoids, gad-iforms, and some ophidiiforms is that (1) theexoccipital facets have moved laterally to anexceptional degree, as noted by Rosen andPatterson (1969), and usually have a deepcore ofcartilage (fig. 37); and (2) the vertebralprezygapophyses extend well forward onto thebraincase, in many cases, carrying a neuralarch and spine component with them to meetthe epioccipitals, supraoccipital, and the dis-placed exoccipitals (fig. 35).Even in the ophidiiforms that retain the
primitive, continuous, planar surface for ex-occipital contact, these prezygapophyses ex-tend well forward over the centrum as clearlyillustrated by Rose (1961) in Ophidion hol-brooki (fig. 36A). Some of the other ophidi-iforms (the bythitoids) are much more cod-like (see fig. 36B, which shows separate rightand left vertebral facets). And, since ophid-iiforms fall readily into two classes based oncaudal anatomy, gill arches, viviparity and,to some degree, fin structure (Cohen andNielsen, 1978, and Patterson and Rosen, workin progress), there is an implication that theophidiiforms might be nonmonophyletic. Onegroup, the more derived in anatomy (bythi-toids) might be linked to cods and batra-choids, and the other (ophidioids) could bethe sister group to the whole lot.
SUMMARY AND CONCLUSIONSAccording to evidence presented here, the
significance of the paired cartilages and tri-partite occipital condyles found in some sal-monids is ambiguous because of the unevendistribution of the characters, the lack of agood theory of relationships among the taxa,the occurrence of both features in some os-meroids, and a parsimony argument that fa-vors osmeroids as a neoteleost sister group.
The significance of this conclusion is that themonophyly of both the Salmonidae and Os-meroidei should be reevaluated, and thatmuch more detailed character surveys areneeded-undertakings that lie outside thescope of this study. Paired rostral cartilagesin their transformed or untransformed statesare present consistently in members of Ro-sen's (1973) Aulopiformes, even though thattaxon must now be abandoned as monophy-letic since Aulopus appears to have a rostralcartilage synapomorphous with that ofcteno-squamates that is lacking in Chlorophthal-mus. However, I can find no obstacle to mak-ing a transformation sequence between thesalmonine occipital region and that of"higher" euteleosteans-unless the Osme-roidei is nonmonophyletic and Spirinchus isa more appropriate immediate sister groupto the neoteleosts. The latter possibility willdepend on two kinds ofevidence: (1) that thegap between the occiput and the first cervicalvertebra is truly diagnostic for primitive neo-teleosts, including stomiiforms, and (2) thatthe postoccipital gap in Spirinchus is synapo-morphous, rather than homoplasious, withthat of neoteleosts.
If the occipital anatomy of Spirinchus isprimitive for osmeroids, as Fink and Weitz-man (1982) claim the anatomy of Diplophosis for stomiiforms, then I would be forced toplace osmeroids, stomiiforms, and neote-leosts in an unresolved trichotomy and toexclude the salmonids on two grounds (cau-dal skeleton and premaxillary anatomy), ex-cept perhaps as the sister group to those threeif the Salmonidae is monophyletic and thesalmonine premaxillary cartilages and occip-ital anatomy are synapomorphies. Mean-while, however, I conclude that the type ofjaw, caudal skeleton, and bone histology earnthe Osmeroidei a closer linkage with stomi-iforms and eurypterygians than do these samefeatures in salmonines.Below is a synapomorphy scheme of the
Clupeocephali representing the data pre-sented here and by other workers which sum-marize anatomical findings relevant to thectenosquamates and the included acantho-morphs.Clupeomorphs share the following derived
features with euteleosts (Patterson and Ro-sen, 1977):
1985 51

AMERICAN MUSEUM NOVITATES
1. Articular bone in the lower jaw co-os-sified with the angular.
2. Retroarticular in lowerjaw excluded fromthe joint surface.
3. A median (in clupeomorphs) or paired(in euteleosts) anteriorly directed mem-branous outgrowth on the anterodorsalmargin of the first uroneural (the stegu-ral).
4. Neural arch over U1, when present, re-duced, rudimentary, lying free over U1or nestled against the posterior face ofthe neural arch ofPU 1, or ankylosed withother neural arch or uroneural com-plexes, or absent.
5. Toothplates, when present, fused withfirst three pharyngobranchials and fifthceratobranchial.
Euteleosts, primitively, have but a single,unambiguous, derived feature:
6. An adipose dorsal fin (not present in eso-coids). The stegural feature (character 3)of the first uroneural has been used as aeuteleost synapomorphy, but this featureseems properly to define only a subgroupof euteleosts. It does not allow decisiveinclusion of the esocoids [which, at best,have an outgrowth of the first uroneuralof doubtful homology with the steguralof other euteleosts (see illustrations inRosen, 1974)].
Within the Euteleostei, the Salmonidae (butnot argentinoids or ostariophysans) share withall other groups only:
7. Paired stegural outgrowths of the firsturoneural.
8. At least some parts of the endoskeletonwith acellular bone (Parenti, MS).
Osmeroids share with stomiiforms andprimitive neoteleosts:
9. Acellular endoskeletal bone (but thisstatement is based only on study of Os-mems mordax. However, K6lliker (185 9)reported acellular bone in a galaxioid,which is consistent with Fink and Weitz-man's (1982) realignment of galaxioidswith osmeroids (and see Parenti, MS).
10. A toothed alveolar process on the pre-maxilla which lies under the maxilla,
completely so in galaxioids (see characterconflicts above).
11. In the caudal skeleton, NPU2 is shorterthan NPU3 and bladelike. [NPU2 is alsoshorter than NPU3 in the stomiiform,Diplophos, but is not bladelike accordingto Fink and Weitzman (1982, fig. 16).]
Stomiiforms and alepisauroids share withother neoteleosts:
12. The exoccipitals and basioccipital ex-posed posteriorly and joined by an in-verted Y-shaped suture. Presence of acervical gap between the occiput and firstvertebra. [When the first vertebra is re-mote from the occipital region, it is cor-related with the exposure ofthe exoccip-ital surfaces as parts of the posterioroccipital outline-the basioccipital hav-ing dorsolateral depressions to accom-modate the exoccipitals in juvenile Os-merus (prevertebral space being occupiedby the notochord and its connective tis-sue sheath and by a small, spineless neu-ral arch).] The "osmeroid" Spirinchusalso has the exoccipitals entering the oc-cipital region posteriorly and future studymay demonstrate that osmeroids arenonmonophyletic.
13. An interoperculohyoid ligament, presentbut feebly developed in stomiiforms (seeLauder and Liem, 1983, p. 34).
14. A retractores dorsalis branchialiummuscle [in most stomiiforms and alepi-sauroids originating far back in the ab-dominal region, with fibers insertingprincipally or only on the fourth gill arch(Rosen, 1973), and perhaps nonho-mologous with that in eurypterygians,according to work in progress by M.Stiassny].2
Neoscopelids share with primitive cteno-squamates (e.g., myctophids):
15. Ctenoid scales and median fin spines(well developed in Cretaceous forms; seeRosen, 1973, pp. 456-459).
16. Premaxilla with ascending, articular, andpostmaxillary processes.
17. Pectoral fins arising laterally rather thanventrally.
18. Pelvic fins arising anteriorly in a sub-
52 NO. 2827

ROSEN: EUTELEOSTEANS
abdominal, rather than in the more pos-terior abdominal, position.
19. A hyoid bar and branchiostegals ofacan-thomorph type (see McAllister, 1968).
20. Clamshell-shaped saccular otolith withdefinitive cauda and ostium in sulcus(Nolf, MS).
21. A pair of small, lateral cartilages joinedto the premaxillary ascending processesand to a larger median cartilage.
22. Retractores dorsalis muscle short, orig-inating anteriorly on basioccipital or cer-vical vertebrae and inserting on connec-tive tissues on dorsomedial edges ofthirdand fourth pharyngobranchials. This as-sumes character 22 to be a more derived(or simply nonhomologous) condition ofcharacter 14.
"Aulopoids" share with ctenosquamates:
23. A median rostral cartilage without lat-eral components (Stiassny, personalcommun., informs me that some myc-tophids retain small, lateral components,so that this character would be statedmore accurately as: distinct lateral com-ponents greatly reduced or absent.)
24. Pelvic fins more anterior in a subthoracicposition. (Fin spines are unknown inmodem "aulopoids," which could be in-terpreted as an autapomorphic loss in"aulopoids" or gain in neoscopelids.)
25. Three or fewer predorsal bones (four inneoscopelids).
Chlorophthalmids share with ctenosqua-mates:
26. A subocular shelf (said by Stiassny, per-sonal commun., to be very narrow andthus interpreted here as primitive rela-tive to the much larger, subocular shelvesin ctenosquamates).
Chlorophthalmids exhibit an additional con-flicting character in this alignment because,in the upper jaw, they lack a median rostralcartilage, but have well-developed paired oneson the premaxillary ascending processes.Myctophids share with acanthomorphs:
27. A subocular shelfthat extends well underthe eyeball (Rosen and Patterson, 1969,fig. 11).
28. Rostral cartilage-a single median struc-
ture of apparently tripartite origin insome species (and with a relic pair oflateral cartilages in a few species).
29. An interarcual cartilage present betweenthe first epibranchial and second pha-ryngobranchial (small, when present, andabsent (according to N. Stiassny, person-al commun.) in some taxa.
30. No more than three predorsal bones.31. Partial closure of the cervical gap be-
tween the occiput and first vertebra.32. Absence of an accessory neural arch in
the cervical region.33. Presence ofneural arch prezygapophyses
on the anterior vertebrae.34. Direct connection (via ligaments) of the
autocentrum ofthe first vertebra with theexoccipital condyles.
Polymixiids share with other acantho-morphs:
35. Autocentral prezygapophyses betweenfirst vertebra and exoccipital condyles.
36. A pelvic fin spine.
Other acanthomorphs share:
37. Complete closure of the notochordal-connective tissue space between the ba-sioccipital and the centrum of the firstvertebra.
38. Primitively, a rodlike interarcual carti-lage between the first and second gillarches [in some beryciforms (e.g. an-omalopids) and in most "percoids"].
Within the above scheme I recognize threemain areas ofuncertainty involving the align-ment of alepisauroids and other members ofthe Aulopiformes recognized by Rosen(1973). Unexpectedly, the neoscopelids seemto me now of uncertain status, although theyand the myctophids are usually closely linked.Lauder (1983) described a character, theurohyal-third hypobranchial ligament, thathe thought was present uniquely in membersof these two families, but the neoscopelidligamentous connection is more like that ofPolymixia.2 I have not searched elsewhere,and I reserve judgment on precise realign-ment ofneoscopelids until all relevant groupshave been more thoroughly reviewed.The various character conflicts noted above
extend to other components of the Aulopi-
1985 53

AMERICAN MUSEUM NOVITATES
formes, and each ofthese is represented eitherby an unresolved tri- or tetrachotomy. Per-haps others (Fink and Weitzman, 1982; Fink,1984) would prefer an additional unresolvedpoint involving salmonids and osmeroids, orsimply to transpose the two. I cannot, how-ever, accept their evidence of the salmonineoccipital joint for the principal reason thatthe salmonine occiput most closely resemblesan advanced acanthomorph occiput ratherthan the basic inverted Y-junction of prim-itive neoteleosts and thus appears to be anindependent specialization of salmonines,which is nonhomologous with the basic neo-teleostean arrangement.
Chlorophthalmids and aulopids appear notto form parts of a monophyletic group be-cause the former have two features (charac-ters 21 and 22) linking them with cteno-squamates, and at least one alepisauroid, asynodontid, also lacks character 21, and alsocharacter 22 according to McAllister (1968,plate 12). Furthermore, some alepisauroidshave ethmoid cartilages resembling those ofstomiiforms (Stiassny, MS) and aulopids havea rostral cartilage ofacanthomorph, or at leastctenosquamate, type. Such data effectivelydissolve the Aulopiformes of Rosen (1973),and Rosen's (1973) more inclusive taxon, theEurypterygii, will come to include the Cteno-squamata and perhaps the Chlorophthalmi-dae and some "aulopoid." I have not restud-ied the alepisauroid fishes, however, and sothese are represented as unresolved in thesynapomorphy scheme and summary clado-gram.
There are numerous derived anatomicalfeatures of euteleostean fishes that appear ina confusing array of taxa which suggest theneed for much additional study and the re-definition oftaxonomic boundaries. Much ofthe confusion has resulted from the assign-ment of "groups," such as "osmeroids" and"aulopiforms" that never were defined prop-erly to begin with. Within the Acanthomor-pha, improperly defined taxa are probablythe rule rather than the exception. At leastten ambiguous features come immediately tomind among fishes that have been associatedat some time or another with the Neoteleos-tei.
1. Interarcual cartilage present in some
myctophids (Travers, 1 98 1), but not oth-ers and absent in neoscopelids and po-lymixiids.
2. Acanthomorph type of hyoid bar andbranchiostegals (see McAllister, 1968) inchlorophthalmids but not in aulopids orsynodontids.
3. Presence of three occipital condyles inmyctophids and neoscopelids, and,among synodontids, in Synodus, but notin Saurida.
4. A cervical gap in primitive cteno-squamates, but not in synodontids.
5. Acanthomorph type of rostral cartilage(fig. 43) in some myctophids, but notothers, and not in neoscopelids (fig. 42).
6. Paired premaxillary components of ros-tral cartilage present in some salmoninesand coregonines, and not others; presentin at least one osmerid.
7. Cervical gap present, and exoccipitalsentering occipital joint surface in someosmerids.
8. In the caudal skeleton, a full spine on thesecond preural centrum in salmonines,but not coregonines.
9. Large, marginal basihyal fangs in sal-monines and galaxioids, but not in thy-mallines or coregonines.
10. Row ofdorsomedial mesopterygoid teethin osmeroids, galaxioids, and also inPterothrissus, in the primitive clupeo-morph tDiplomystus, and in the cteno-squamate, Synodus.
11. Neural spine on second preural centrumreduced to a low crest except in Poly-mixia and fishes formerly united as par-acanthopterygians and a few subgroupsof acanthopterygians.
In this extended essay on the taxonomy ofthe single largest recognized group ofteleosts,the Euteleostei, I have adopted a scheme ofrelationships ofits subgroups differing some-what from my own and those of other recentworkers. My proposal raises the general ques-tion of why taxonomists periodically adoptnovel taxonomies. Many years ago it was be-cause taxonomists used symplesiomorphiesto describe groups, e.g., the Clupeiformes,Malacopterygii, Isospondyli, and Mesich-thys. But the recent flux of fish classificationshas other causes, since most ichthyologistshave abandoned symplesiomorphy in favor
54 NO. 2827

ROSEN: EUTELEOSTEANS
of synapomorphy as a basis for defininggroups.To me, at least, there appear to be three
major reasons for this flux. One is that dif-ferent taxonomists perceive characters dif-ferently. Since our experiences do not coin-cide, neither do our perceptions. This is aphenomenological problem of what each ofus is able to see and how we interpret whatwe perceive. And it is aggravated by a lin-guistic problem ofhow we describe those per-ceptions. A second main reason for the steadyflow of new taxonomic proposals has to dowith the willingness of some taxonomists toalign groups based on a trait peculiar to oneor only a few ofits members, a trait that mustbe suspected ofbeing homoplasious unless itcan be argued successfully to be primitive forthe entire group. The tripartite occipital con-dyle and paired rostral cartilages of salmon-ines would fall in this category (vide Fink andWeitzman, 1982, and Fink, 1984) as wouldthe basihyal dentition of salmonines (videRosen, 1974). And evaluation of an ambig-uous trait, too, is aggravated by a linguisticproblem ofhow the trait is described. A thirdmain reason is a genuinely new discovery ofsuch a major character that it seems obviousand significant to one and all. An example ofthis type would be the Weberian apparatusof otophysans or the recessus lateralis of clu-peiforms. The retractor dorsalis muscleseemed like one of these characters when itwas first reported and described in acantho-morphs by Dietz in a series of papers (seeRosen, 1973) and by Holtzvoogd (1965), butthe retractor muscle appears widely amongsthalecomorph neopterygians and its homo-plasious nature made application ofthe char-acter ambiguous at various hierarchical levels(Nelson, 1967). There appear to be only twoways of dealing with this ambiguity. One isto conduct an empirical study of the featurein different taxa showing that its ontogeny isunique in some ofthose taxa and that its finalexpression is, therefore, homoplasious (i.e.,irrelevant because of nonhomology). Theother is to study many other features thatyield congruent hierarchical arrangements-a congruence which the ambiguous featuredoes not share (i.e., again, because it is anexpression of a different ontogeny and is,therefore, nonhomologous with a trait that it
resembles). But ambiguous or inconsistentlydistributed characters still plague fish system-atics and are used again and again to supportsome proposed relationship. Having beenguilty of the crime myself, I am naturallytolerant of others who commit it, allowingthat it is a far better thing to have proposedand been rejected on grounds of nonhomol-ogy than never to have proposed at all.
LITERATURE CITED
Agassiz, J. L. R.1858. The classification of fishes. Proc. Amer.
Acad. Sci., vol. 4, p. 108.Berg, L. S.
1940. Classification offishes, both Recent andfossil. Russian and English, Ann Arbor,Michigan, Edwards Bros.
Boulenger, G. A.1904. A synopsis ofthe suborders and families
of the teleostean fishes. Ann. Mag. Nat.Hist., vol. 7, pp. 161-190.
Cavender, T. M., and R. R. Miller1972. Smilodonichthys rastrosus, a new Plio-
cene salmonid fish from western UnitedStates. Mus. Nat. Hist., Univ. Oregon,Bull. 18, 44 pp.
Cohen, D. M., and J. G. Nielsen1978. Guide to the identification of genera of
the fish order Ophidiiformes with a ten-tative classification ofthe order. NOAATech. Rept. NMFS Circular 417,72 pp.
Dietz, P. A.1914. Beitrage zur Kenntnis der Kiefer- und
Kiemen-bogenmuskulatur der Teleos-tier. I. Die Kiefer- und Kiemen-bogen-muskeln der Acanthopterygier. Mitt.Zool. Staz. Neapel, vol. 22, pp. 99-162.
Fink, S. V., and W. L. Fink1981. Interrelationships of the osteriophysan
fishes (Teleostei). Jour. Linn. Soc.(Zool.), vol. 72, no. 4, pp. 297-353.
Fink, W. L.1984. Basal euteleosts: relationships. In Spe-
cial Publ. no. 1, Amer. Soc. Ichthyol.and Herp. Ontogeny and Systematics ofFishes, pp. 202-206.
Fink, W. L., and S. H. Weitzman1982. Relationships of the stomiiform fishes
(Teleostei), with a description of Diplo-phos. Bull. Mus. Comp. Zool., vol. 150,no. 2, pp. 31-93.
Forey, P. L.1973. A revision ofthe elopiform fishes, fossil
and recent. Bull. British Mus. (Nat.Hist.), Geol., Suppl. 10, 222 pp.
1985 55

AMERICAN MUSEUM NOVITATES
Gill, T. N.1872. Arrangement of the families of fishes,
or classes Pisces, Marsipobranchii, andLeptocardii. Smithsonian Misc. Coll.,vol. 11, no. 247, pp. i-1-49.
Goodrich, E. S.1909. Studies on the structure and develop-
ment of vertebrates. Dover Publ. re-print 1958, 2 vols.
Gosline, W. A.1969. The morphology and systematic posi-
tion of the alepocephaloid fishes. Bull.British Mus. (Nat. Hist.), vol. 18, no. 6,pp.183-218.
1971. Functional morphology and classifica-tion of teleostean fishes. Univ. HawaiiPress, 208 pp.
Grande, L.[in press] Recent and fossil clupeomorph fishes
with materials for a revision of thesubgroup ofclupeoids. Bull. Amer. Mus.Nat. Hist.
Greenwood, P. H.1968. The osteology and relationships of the
Denticipitidae, a family of clupeo-morph fishes. Bull. Brit. Mus. (Nat.Hist.), Zool., vol. 16, pp. 13-273.
1970. Skull and swimbladder connections infishes of the family Megalopidae. Bull.Brit. Mus. (Nat. Hist.), Zool., vol. 19,no. 1, pp. 121-135.
Greenwood, P. H., G. S. Myers, D. E. Rosen, andS. H. Weitzman1967. Named main divisions of teleostean
fishes. Proc. Biol. Soc. Washington, vol.80, pp. 227-228.
Greenwood, P. H., and D. E. Rosen1971. Notes on the structure and relationships
ofthe alepocephaloid fishes. Amer. Mus.Novitates 2473, pp. 1-41.
Greenwood, P. H., D. E. Rosen, S. H. Weitzman,and G. S. Myers1966. Phyletic studies ofteleostean fishes, with
a provisional classification of livingforms. Bull. Amer. Mus. Nat. Hist., vol.131, pp.339-456.
Gunther, A. C. L. G.1859-1870. Catalogue of fishes of the British
Museum, vols. 1-8, Brit. Mus. Nat. Hist.London.
Holtzvoogd, C.1965. The pharyngeal bones and muscles in
Teleostei, a taxonomic study. Proc. K.ned. Akad. Wet. (Zool.) (Ser. C), vol.68, no. 3, pp. 209-218.
Jollie, M. T.1954. The general anatomy of Lampanyctus
leucopsarus (Eigenmann and Eigen-mann). Univ. Microfilms, 239 pp.
1 984a. Development of the head and pectoralskeleton of Amia with a note on thescales. Gegen baurs morph. Jahrb.,Leipzig, vol. 130, no. 2, pp. 315-351.
1984b. Development of the head skeleton andpectoral girdle of salmons, with a noteon the scales. Canadian Jour. Zool., vol.62, pp. 1757-1778.
Jordan, D. S.1923. A classification offishes. Stanford Univ.
Publ., Univ. Ser., Biol. Sci., vol. 3, no.2, pp. 1-243 + x.
K6lliker, A.1859. On the different types in the microscop-
ic structure of the skeleton of osseousfishes. Proc. Roy. Soc. London, vol. 9,pp. 1-13.
Lauder, G. V.1983. Functional design and evolution of the
pharyngeal jaw apparatus in euteleos-tean fishes. Zool. Jour. Linn. Soc., vol.77, no. 1, pp. 1-38.
Lauder, G. V., and K. F. Liem1983. The evolution and interrelationships of
the actinopterygian fishes. Bull. Mus.Comp. Zool., vol. 150, no. 3, pp. 95-197.
McAllister, D. E.1968. Evolution of branchiostegals and clas-
sification ofteleostome fishes. Natl. Mus.Canada, Bull. 221, Biol. Ser., no. 77,239 pp.
Mead, G. W.1966. Family Chlorophthalmidae. In Fishes
of the Western North Atlantic. Mem.Sears Found. Mar. Res., no. 1, pt. 5, pp.162-189.
Muller, J.1844. Uber den Ban und die Grenzen der
Ganoiden und uiber das naturliche sys-tem der Fische. Abhandl. Akad. Wiss.Berlin, phys.-math., pp. 201-204.
Myers, G. S.1958. Nomenclator of certain terms used for
higher categories offishes. Stanford Ich-thyol. Bull., vol. 7, no. 3, pp. 31-40.
Nelson, G.1967. Branchial muscles in some generalized
teleostean fishes. Acta Zool., Stock-holm, vol. 48, pp. 277-288.
Nelson, J.1984. Fishes of the world. New York, John
Wiley, 2nd ed.Nolf, D.
1974. De Teleostei-otolieten uit het Eoceenvan het Belgisch Bekken-reconstructievan de fauna en biostratigrafische toe-passing. Doctoral dissertation, Rijks-universiteit, Gent.
56 NO. 2827

ROSEN: EUTELEOSTEANS
Norden, C. R.1961. Comparative osteology of representa-
tive salmonid fishes, with particular ref-erence to the grayling (Thymallus arc-ticus) and its phylogeny. Jour. Fish. Res.Bd. Canada, vol. 18, no.5, pp. 679-791.
Norman, J. R.1934. A draft synopsis of the orders, families
and genera of Recent fishes and fishlikevertebrates. 1957 photo offset by BritishMus. Nat. Hist.
Parenti, L. R.[MS] The phylogenetic significance of bone
types in euteleost fishes. Submitted toLinn. Soc. London.
Patterson, C.1964. A review ofMesozoic acanthopterygian
fishes, with special reference to those ofthe English Chalk. Phil. Trans. Roy. Soc.London, ser. B, vol. 247, pp. 213-482.
1975. The braincase of pholidophorid andleptolepid fishes, with a review of theactinopterygian braincase. Ibid., Biol.Sci., vol. 269, no. 899, pp. 275-579.
Patterson, C., and D. E. Rosen1977. Review of ichthyodectiform and other
Mesozoic teleost fishes and the theoryand practice of classifying fossils. Bull.Amer. Mus. Nat. Hist., vol. 158, art. 2,pp. 81-172.
Peiiaz, M.1983. Ecomorphological laws and saltation in
the early ontogeny of Salmonoidei. Fo-lia Zool., vol. 32, no. 4, pp. 365-373.
Regan, C. T.1909. The classification of teleostean fishes.
Ann. Mag. Nat. Hist., ser. 7, vol. 11,pp. 459-466.
1929. Fishes. In Encyclopedia Britannica, 14thed., London and New York, vol. 9, pp.305-329.
Ridewood, W. G.1904. On the cranial osteology of the fishes of
the families Elopidae and Albulidae,with remarks on the morphology of theskull in the lower teleostean fishes gen-erally. Proc. Zool. Soc. London, pp. 35-81.
1905. On the cranial osteology ofthe clupeoidfishes. Ibid., pt. 1, pp. 448-493.
Rose, J. A.1961. Anatomy and sexual dimorphism ofthe
swimbladder and vertebral column inOphidion holbrooki (Pisces: Ophidi-idae). Bull. Mar. Sci. Gulf and Carib.,vol. 11, no. 2, pp. 280-308.
Rosen, D. E.1973. Interrelationships of higher euteleos-
tean fishes. In Greenwood, P. H., R. S.Miles, and C. Patterson (eds.), Interre-lationships infishes. London, AcademicPress, pp. 397-513.
1974. Phylogeny and zoogeography of sal-moniform fishes and relationships ofLepidogalaxias salamandroides. Bull.Amer. Mus. Nat. Hist., vol. 153, art. 2,pp. 265-326.
1982. Teleostean interrelationships, morpho-logical function and evolutionary infer-ence. Amer. Zool., vol. 22, no. 2, pp.261-273.
Rosen, D. E., and C. Patterson1969. The structure and relationships of the
paracanthopterygian fishes. Bull. Amer.Mus. Nat. Hist., vol. 141, art. 3, pp.357-474.
Smith, C. L., and R. M. Bailey1961. Evolution of the dorsal-fin supports of
percoid fishes. Papers Mich. Acad. Sci.,Arts and Letters, vol. 46, pp. 345-363.
Smith-Woodward, A.1932. Textbook ofPalaeontology. 2nd English
ed. revised with addition of C. East-man's English translation of vol. 2 byKarl A. von Zittel.
Stiassny, M. L. J.[MS] The limits and relationships ofthe acan-
thomorph teleosts.Travers, R. A.
1981. The interarcual cartilage: a review of itsdevelopment, distribution and value asan indicator of phyletic relationships ineuteleostean fishes. Jour. Nat. Hist., vol.15, pp. 853-871.
Vladykov, V. D.1962. Osteological studies on Pacific salmon
of the genus Oncorhynchus. Bull. Fish.Res. Bd. Canada, no. 136, pp. 1-172.
Weitzman, S. H.1974. Osteology and evolutionary relation-
ships ofthe Sternoptychidae, with a newclassification of stomatiatoid families.Bull. Amer. Mus. Nat. Hist., vol. 153,art. 3, pp. 327-478.
Whitehead, P. J. P., and G. G. Teugels[in press] The West African pygmy herring
Sierrathrissa leonensis: general fea-tures, visceral anatomy and osteology.Amer. Mus. Novitates.
Woods, L. P., and R. F. Inger1957. The cave, spring, and swampfishes of
the family Amblyopsidae of central andeastern United States. Amer. Midl. Nat.,vol. 58, no. 1, pp. 232-256.
1985 57
