copyright © 2005 pearson education, inc. publishing as benjamin cummings evolutionary biology is...
TRANSCRIPT

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Evolutionary biology is about both process and history.
• The processes of evolution are natural selection and other mechanisms that change the genetic composition of populations and can lead to the evolution of new species.
• A major goal of evolutionary biology is to reconstruct the history of life on earth.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Overview: Investigating the Tree of Life
• Phylogeny is the evolutionary history of a species or group of related species
• Biologists draw on the fossil record, which provides information about ancient organisms

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Systematics is an analytical approach to understanding the diversity and relationships of organisms, both present-day and extinct
• Systematists use morphological, biochemical, and molecular comparisons to infer evolutionary relationships
• Molecular systematics, which uses comparisons of nucleotide sequences in DNA and RNA to help identify evolutionary relationships between individual genes or even entire genomes.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Phylogenies are based on common ancestries inferred from fossil, morphological, and molecular evidence
• To infer phylogenies, systematists gather information about morphologies, development, and biochemistry of living organisms
• They also examine fossils to help establish relationships between living organisms
• Fossils are the preserved remnants or impressions left by organisms that lived in the past. In essence, they are the historical documents of biology.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Sedimentary rocks form from layers of sand and silt that are carried by rivers to seas and swamps, where the minerals settle to the bottom along with the remains of organisms.
• As deposits pile up, they compress older sediments below them into layers called strata.
• The fossil record is the ordered array in which fossils appear within sedimentary rock strata.
• These rocks record the passing of geological time.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Fossils can be used to construct phylogenies only if we can determine their ages.
• The majority of living things were not captured as fossils upon their death.
• Of those that formed fossils, later geological processes destroyed many.
• Only a fraction of existing fossils have been discovered.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Morphological and Molecular Homologies
• Similarities due to shared ancestry are called homologies.
• In addition to fossils, phylogenetic history can be inferred from morphological and molecular similarities in living organisms
• Organisms with very similar morphologies or similar DNA sequences are likely to be more closely related than organisms with vastly different structures or sequences

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Sorting Homology from Analogy
• In constructing a phylogeny, systematists need to distinguish whether a similarity is the result of homology or analogy
• Homology is similarity due to shared ancestry
• Analogy is similarity due to convergent evolution

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Convergent evolution occurs when similar environmental pressures and natural selection produce similar (analogous) adaptations in organisms from different evolutionary lineages
• Similar analogous adaptations may evolve in such organisms.
• Analogies are not due to shared ancestry.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Distinguishing homology from analogy is critical in the reconstruction of phylogeny.
• For example, both birds and bats have adaptations that allow them to fly.
• However, a close examination of a bat’s wing shows a greater similarity to a cat’s forelimb that to a bird’s wing.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Fossil evidence also documents that bat and bird wings arose independently from walking forelimbs of different ancestors.
• Thus a bat’s wing is homologous to other mammalian forelimbs but is analogous in function to a bird’s wing.
• Analogous structures or molecular sequences that evolved independently are also called homoplasies.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In general, the more points of resemblance that two complex structures have, the less likely it is that they evolved independently.
• For example, the skulls of a human and a chimpanzee are formed by the fusion of many bones. The two skulls match almost perfectly, bone for bone.
• It is highly unlikely that such complex structures have separate origins. More likely, the genes involved in the development of both skulls were inherited from a common ancestor.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Systematists compare long stretches of DNA and even entire genomes to assess relationships between species.
• If genes in two organisms have closely similar nucleotide sequences, it is highly likely that the genes are homologous.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Evaluating Molecular Homologies
• Systematists use computer programs and mathematical tools when analyzing comparable DNA segments from different organisms
• Deletions or insertions may shift the remaining sequences, making it difficult to recognize closely matching nucleotide sequences.

LE 25-6
1
2
1
2
Deletion
Insertion
1
2
1
2

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Phylogenetic systematics connectsclassification with evolutionary history
• Taxonomy is the ordered division of organisms into categories based on characteristics used to assess similarities and differences
• In 1748, Carolus Linnaeus published a system of taxonomy based on resemblances.
• Two key features of his system remain useful today: two-part names for species and hierarchical classification

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Binomial Nomenclature
• The two-part scientific name of a species is called a binomial nomenclature
• The first part of the name is the genus
• The second part, called the specific epithet, is unique for each species within the genus
• The first letter of the genus is capitalized, and the entire species name is latinized
• Both parts together name the species (not the specific epithet alone)

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Hierarchical Classification
• Linnaeus introduced a system for grouping species in increasingly broad categories
• Linneaus’s classification was not based on evolutionary relationships but simply on resemblances between organisms.
• Each taxonomic level is more comprehensive than the previous one.
• As an example, all species of cats are mammals, but not all mammals are cats.

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The named taxonomic unit at any level is called a taxon.
• Example: Panthera is a taxon at the genus level, and Mammalia is a taxon at the class level that includes all of the many orders of mammals.

LE 25-8
Species
Pantherapardus
PantheraGenus
FamilyFelidae
CarnivoraOrder
MammaliaClass
PhylumChordata
KingdomAnimalia
EukaryaDomain

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Linking Classification and Phylogeny
• Systematists depict evolutionary relationships in branching phylogenetic trees
• The branching of the tree reflects the hierarchical classification of groups nested within more inclusive groups.
• Methods for tracing phylogeny began with Darwin, who realized the evolutionary implications of Linnaean hierarchy.

LE 25-9
Carnivora
Pantherapardus
(leopard)
Mephitismephitis
(striped skunk)
Lutra lutra(European
otter)
Canisfamiliaris
(domestic dog)
Canislupus(wolf)S
pec
ies
Gen
us
Fam
ilyO
rder
Felidae Mustelidae Canidae
Panthera Mephitis Lutra Canis

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Each branch point represents the divergence of two species
• “Deeper” branch points represent progressively greater amounts of divergence

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Concept 25.3: Phylogenetic systematics informs the construction of phylogenetic trees based on shared characteristics
• If shared characteristics are homologous and, thus, explained by common ancestry, then the cladogram forms the basis of a phylogenetic tree.
• A cladogram depicts patterns of shared characteristics among taxa
• A clade is a group of species that includes an ancestral species and all its descendants
• Cladistics studies resemblances among clades

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• A shared derived character is an evolutionary novelty unique to a particular clade
• The presence of a backbone can qualify as a shared derived character, but at a deeper branch point that distinguishes all vertebrates from other mammals.
• Among vertebrates, the backbone is a shared primitive character because it evolved in the ancestor common to all vertebrates.

LE 25-10a
Grouping 1
Monophyletic
A valid clade is monophyletic, signifying
that it consists of the ancestor species and all
its descendants
LE 25-10b
Paraphyletic
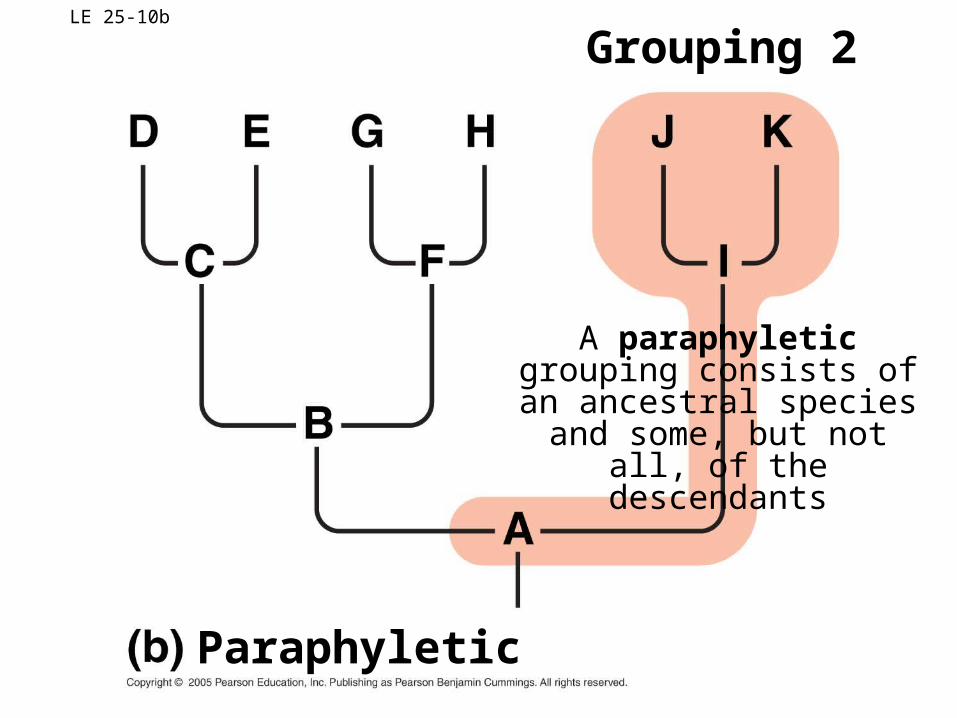
Grouping 2
A paraphyletic grouping consists of an ancestral species
and some, but not all, of the descendants
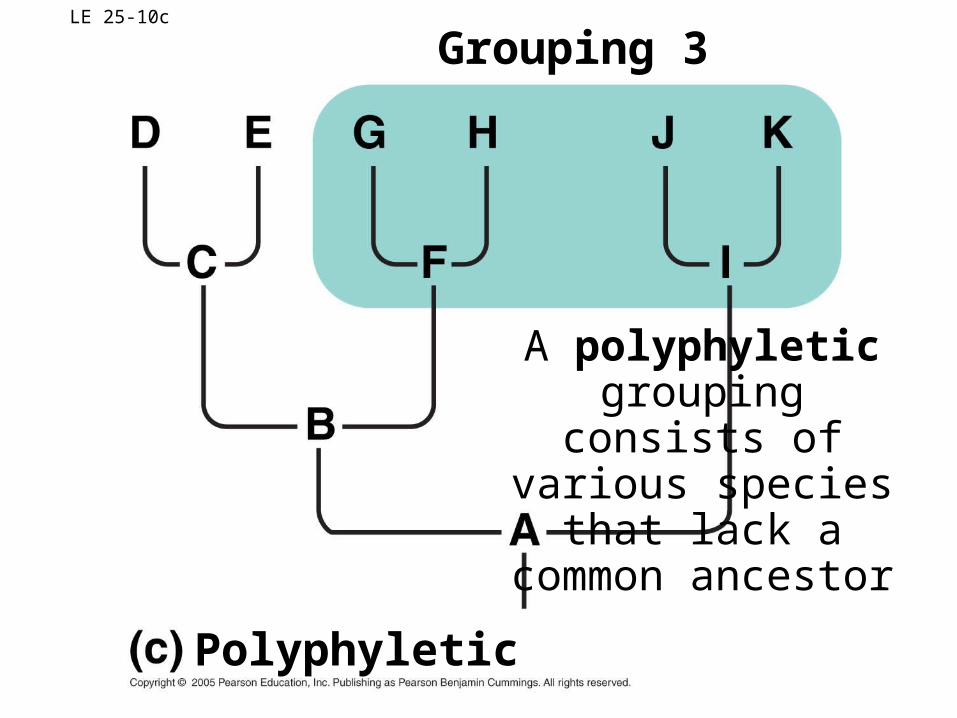
LE 25-10c
Polyphyletic
Grouping 3
A polyphyletic grouping consists of various species that lack a common ancestor

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Outgroups
• An outgroup is a species or group of species that is closely related to the ingroup, the various species being studied
• Systematists compare each ingroup species with the outgroup to differentiate between shared derived and shared primitive characteristics

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• Outgroup comparison assumes that homologies shared by the outgroup and ingroup must be primitive characters that predate the divergence of both groups from a common ancestor
• It enables us to focus on characters derived at various branch points in the evolution of a clade

LE 25-11
Hair
Amniotic (shelled) egg
Four walking legs
Hinged jaws
Vertebral column(backbone)
Character table
CH
AR
AC
TE
RS
TAXA
La
nc
ele
t(o
utg
rou
p)
La
mp
rey
Tu
na
Sa
lam
an
de
r
Tu
rtle
Le
op
ard
Turtle Leopard
Hair
Amniotic egg
Four walking legs
Hinged jaws
Vertebral column
Salamander
Tuna
Lamprey
Lancelet (outgroup)
Cladogram

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Phylogenetic Trees and Timing
• Any chronology represented by the branching of a phylogenetic tree is relative rather than absolute in representing timing of divergences
• In a phylogram, the length of a branch in a cladogram reflects the number of genetic changes that have taken place in a particular DNA or RNA sequence in that lineage

LE 25-12
Droso
phila
Lanc
elet
Fish
Amph
ibia
nBird
Human
Rat
Mou
se

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Ultrametric Trees
• Branching in an ultrametric tree is the same as in a phylogram, but all branches traceable from the common ancestor to the present are equal length

LE 25-13
Droso
phila
Lanc
elet
Fish
Amph
ibia
nBird
Human
Rat Mou
se
Ce
no
zoic
Me
so
zoic
Pa
leo
zoic
65
.52
51
54
2
Ne
op
rote
rozo
ic
Mil
lio
ns
of
ye
ars
ag
o

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Maximum Parsimony and Maximum Likelihood
• Systematists can never be sure of finding the best tree in a large data set
• They narrow possibilities by applying the principles of maximum parsimony and maximum likelihood

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• The most parsimonious tree requires the fewest evolutionary events to have occurred in the form of shared derived characters
• The principle of maximum likelihood states that, given certain rules about how DNA changes over time, a tree can be found that reflects the most likely sequence of evolutionary events
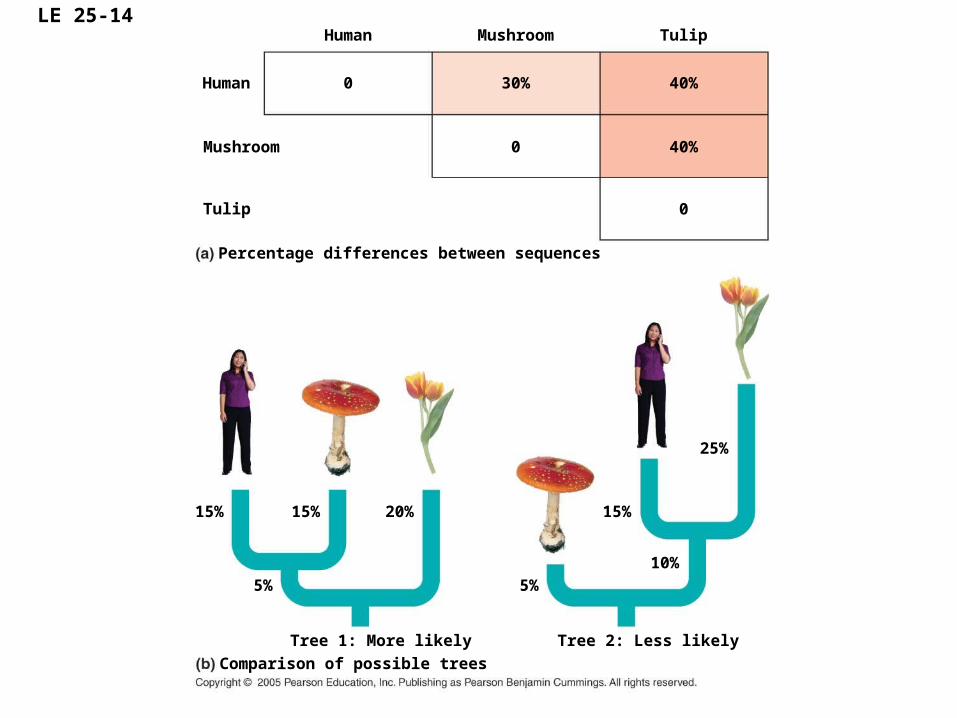
LE 25-14Human
0
Mushroom
30%
0
Tulip
40%
40%
Human
Mushroom
0Tulip
Percentage differences between sequences
Comparison of possible trees
15% 15% 20%
5% 5%
10%
15%
25%
Tree 1: More likely Tree 2: Less likely

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
• In considering possible phylogenies for a group of species, systematists compare molecular data for the species.
• The most efficient way to study hypotheses is to consider the most parsimonious hypothesis, the one requiring the fewest evolutionary events (molecular changes)
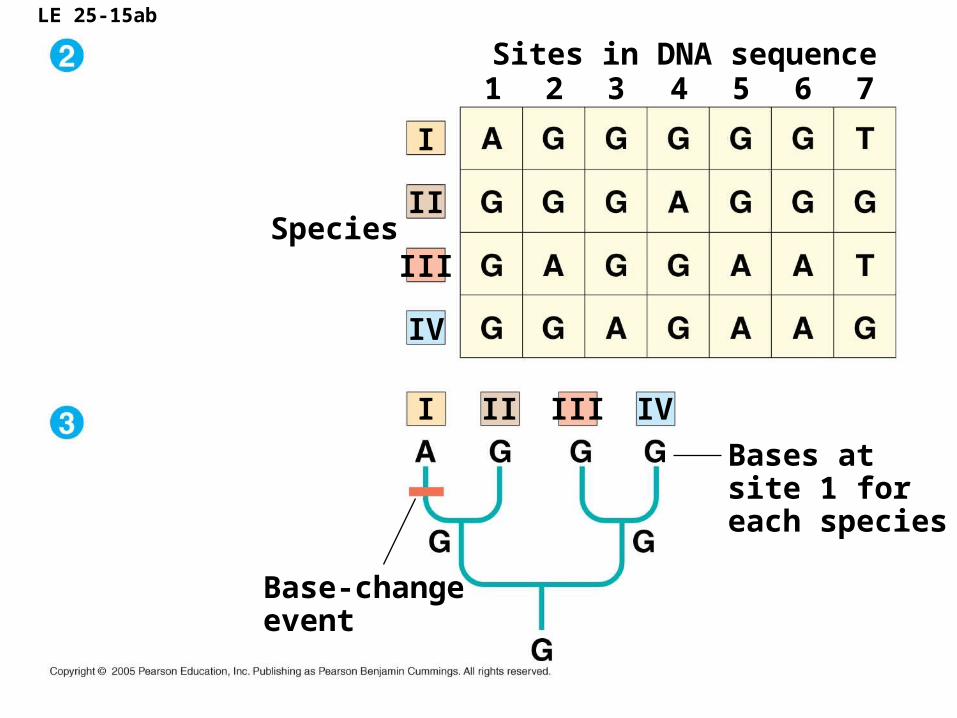
LE 25-15ab
Sites in DNA sequence
I
Species
1
Base-changeevent
Bases atsite 1 foreach species
2 3 4 5 6 7
II
III
IV
I II III IV

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Phylogenetic Trees as Hypotheses
• The best hypotheses for phylogenetic trees fit the most data: morphological, molecular, and fossil
• Sometimes the best hypothesis is not the most parsimonious

LE 25-16
Lizard Bird Mammal
Four-chamberedheart
Mammal-bird clade
Lizard Bird Mammal
Four-chamberedheart
Four-chamberedheart
Lizard-bird clade

• Orthologous genes are genes found in a single copy in the genome
• They can diverge only after speciation occurs
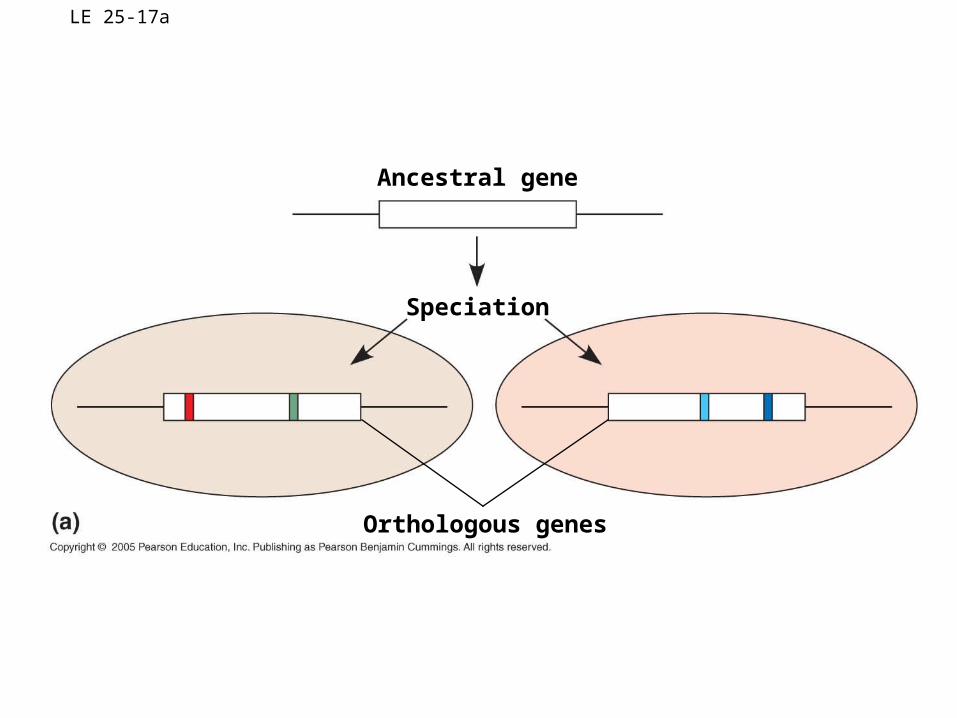
LE 25-17a
Ancestral gene
Speciation
Orthologous genes

• Paralogous genes result from gene duplication, so are found in more than one copy in the genome
• They can diverge within the clade that carries them, often adding new functions
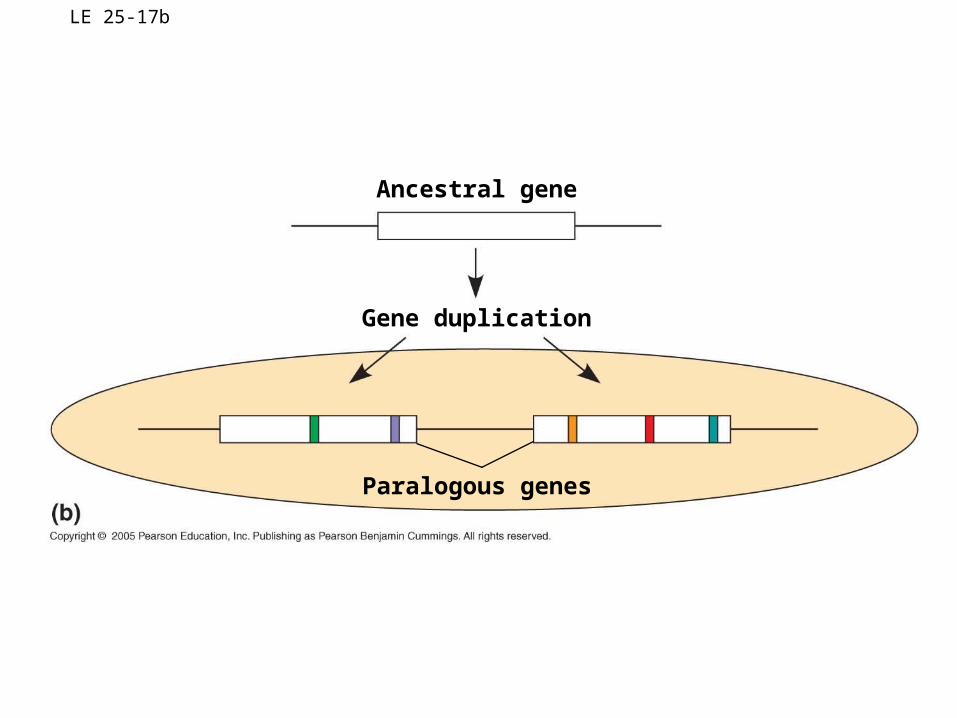
LE 25-17b
Ancestral gene
Gene duplication
Paralogous genes

Genome Evolution
• Orthologous genes are widespread and extend across many widely varied species
• The widespread consistency in total gene number in organisms indicates genes in complex organisms are very versatile and that each gene can perform many functions

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Much of an organism’s evolutionary history is documented in its genome
• Comparing nucleic acids or other molecules to infer relatedness is a valuable tool for tracing organisms’ evolutionary history
• Gene duplication increases the number of genes in the genome, providing more opportunities for evolutionary changes

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Concept 25.5: Molecular clocks help track evolutionary time
• To extend molecular phylogenies beyond the fossil record, we must make an assumption about how change occurs over time
• The molecular clock is a yardstick for measuring absolute time of evolutionary change based on the observation that some genes and other regions of genomes seem to evolve at constant rates

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Neutral Theory
• Neutral theory states that much evolutionary change in genes and proteins has no effect on fitness and therefore is not influenced by Darwinian selection
• It states that the rate of molecular change in these genes and proteins should be regular like a clock

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Difficulties with Molecular Clocks
• The molecular clock does not run as smoothly as neutral theory predicts
• Irregularities result from natural selection in which some DNA changes are favored over others
• Estimates of evolutionary divergences older than the fossil record have a high degree of uncertainty

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
Applying a Molecular Clock: The Origin of HIV
• Phylogenetic analysis shows that HIV is descended from viruses that infect chimpanzees and other primates
• Comparison of HIV samples throughout the epidemic shows that the virus evolved in a very clocklike way

Copyright © 2005 Pearson Education, Inc. publishing as Benjamin Cummings
The Universal Tree of Life
• The tree of life is divided into three great clades called domains: Bacteria, Archaea, and Eukarya
• The early history of these domains is not yet clear