copper toxicity to five parmelia lichens in vitro
TRANSCRIPT

Copper toxicity to five Parmelia lichens in vitro
Joao Paulo Cabral *
Faculty of Sciences, Department of Botany, University of Oporto, Rua do Campo Alegre, 1191, 4150-181 Porto, Portugal
Center of Marine and Environmental Research (CIIMAR), University of Oporto, Rua do Campo Alegre, 823, 4150-180 Porto, Portugal
Received 21 May 2002; received in revised form 15 October 2002; accepted 15 October 2002
Abstract
Treatment of Parmelia caperata , P. perlata , P. subrudecta , P. sulcata and P. tiliacea with CuSO4 resulted in a time-
and copper-concentration-dependent decrease in the total and intracellular potassium concentrations of the thallus,
indicating that copper damaged the cytoplasmic membrane. Treatment with copper also resulted in a time-dependent
increase in the total copper concentration of the thallus. After 4 h of exposure to copper, the process of potassium efflux
was essentially completed but the absorption of copper was still increasing; moreover, the amount of copper bound to
the thallus exceeded twice the amount of potassium released from the thallus, suggesting that cupric ions reached
intracellular sites in the thallus, and K�/Cu2� exchange was not electroneutral. After 5 h of exposure to copper, the
extent of decrease in the total and intracellular potassium concentrations of the thallus was positively correlated with
copper absorption levels, but only at 0.05B/P B/0.10, suggesting that membrane damage was proportional to the
amount of bound copper, but other factors could have been operative, namely binding of copper to the cell wall.
Acetone extracts of untreated thalli contained low concentrations of amino acids, polyols, and sugars, but considerable
amounts of lichen substances: atranorin, caperatic, constictic, lecanoric, menegazziaic, protocetraric, salazinic, stictic,
and usnic acids. Titration of the extracts with copper and assay of the free Cu2� concentration revealed the presence of
copper-binding ligands, and several successive absorption cycles, most probably corresponding to the binding of Cu2�
to each of the lichen substances detected in the extracts. However, no significant correlation (P �/0.10) was found
between the Cu2�-complexing capacity of acetone extracts and copper-induced membrane damage. It was concluded
that in the studied Parmelia species, and in the experimental conditions used in this work, copper toxicity was not a
simple function of the Cu2�-binding properties of the lichen substances present in the thallus. Several hypotheses were
formulated to interpret the results.
# 2002 Elsevier Science B.V. All rights reserved.
Keywords: Cupric ions; Lichen secondary metabolites; Membrane damage; Potassium efflux
1. Introduction
Most borderline (e.g. As, Cd, Co, Cr, Cu, Ni,
Sn, Zn) and soft (class B) (e.g. Ag, Au, Hg, Pb, Tl)
metal ions are toxic (at all concentrations, or
above a certain threshold concentration). Toxicity
* Present address: Departamento de Botanica, Rua do
Campo Alegre, 1191, 4150-181 Porto, Portugal. Tel.: �/351-
22-6002153; fax: �/351-22-6092227
E-mail address: [email protected] (J.P. Cabral).
Environmental and Experimental Botany 49 (2003) 237�/250
www.elsevier.com/locate/envexpbot
S0098-8472/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 0 9 8 - 8 4 7 2 ( 0 2 ) 0 0 0 8 7 - 4

generally increases with increasing class B char-acter (Nieboer and Richardson, 1980). In particu-
lar copper, a borderline metal with a relatively soft
character, is highly toxic to plants, usually in the
micromolar range of exposure concentrations.
Copper inhibits root elongation and branching,
reduces transroot potential, increases lignin synth-
esis, blocks the photosynthetic electron transport,
and induces chlorophyll degradation and leakageof pool metabolites (Sandmann and Boger, 1980;
Arduini et al., 1995; Quartacci et al., 2001;
Patsikka et al., 2002). Although copper can inter-
fere with a number of physiological processes, the
primary site of copper toxicity is probably at the
cell membrane (Demidchik et al., 1997; Chen et al.,
2000). The lipid bilayer can be damaged by
displacement of endogenous-stabilizing cations(mainly Ca2� and Mg2�), by alterations in the
phospholipic composition, and/or by lipid perox-
idation (Avery et al., 1996; Demidchik et al., 1997;
Howlett and Avery, 1997; Murphy et al., 1999;
Quartacci et al., 2001). These result in a disorga-
nization of the packed phospholipidic structure,
with alterations in membrane fluidity and loss of
selective permeability (Quartacci et al., 2001).Copper can substitute essential cofactors and
bind to �/SH groups of proteins (including enzy-
matic systems, channels, and carriers) causing loss
of function (Nieboer and Richardson, 1980; Stau-
ber and Florence, 1987; Meharg, 1994; Demidchik
et al., 1997). Copper-induced membrane damage
usually results in the release of significant amounts
of K� and pool metabolites from the cells(Hassall, 1963; Joho et al., 1984; Ohsumi et al.,
1988).
Lichen mycobionts produce, mainly in the
symbiotic state, a special class of metabolites, the
lichen substances (Lawrey, 1986; Leuckert et al.,
1990; Ahmadjian, 1993; Elix, 1996). Lichen sub-
stances are phenolic secondary metabolites, have
very low or low solubility in water (Iskandar andSyers, 1971), but are highly soluble in organic
solvents (Fahselt, 1994; Elix, 1996). Lichen sub-
stances occur in the cortex or in the medulla of the
thallus, at the surface of the mycobiont hyphae,
and account for 0.1�/5% thallus dry weight (Fah-
selt, 1994). Several hundreds of lichen substances
have been identified in lichens, and most of the
lichen species accumulate these metabolites. Sev-eral roles have assigned to this special class of
molecules, including antimicrobial activity (Hu-
neck, 1968; Lawrey, 1986), antiherbivore roles
(Lawrey, 1986; Fahselt, 1994), antitumor activity
(Huneck, 1968), and light-screening ability (Law-
rey, 1986; Fahselt, 1994 Solhaug and Gausla,
1996).
Most lichen substances are acidic and whenisolated, in vitro, can bind metal ions, including
aluminum, calcium, copper, iron, and magnesium
(Syers, 1969; Iskandar and Syers, 1972; Purvis et
al., 1987). In lichens from cupriferous substrata,
the formation of complexes between lichen sub-
stances (norstictic and psoromic acids) and copper,
in the thallus, has been implicated in the resistance
of these lichens to copper toxicity (Purvis et al.,1985, 1987, 1990). These results suggested that
lichen substances might modulate copper toxicity.
Our initial working hypothesis was that copper
toxicity is a direct function of the Cu2�-binding
properties of the lichen substances present in the
thallus. To test this hypothesis, lichen substances
were extracted from the thallus of five Parmelia
species (growing in unpolluted sites), and thecopper-complexing capacity of these substances
determined by ion-selective electrode potentiome-
try. Copper toxicity was assessed by membrane
damage, and this was evaluated by determining, by
flame emission photometry and atomic absorption
spectrophotometry, the total and intracellular
potassium concentration of the thallus, after
copper treatment. In order to understand thekinetics of membrane damage, the binding of
copper to the thallus was also studied in identical
experimental conditions.
2. Materials and methods
2.1. Lichens and sample preparation
Five Parmelia species were used in this work,
Parmelia caperata (L.) Ach., P. perlata (Huds.)
Ach., P. subrudecta Nyl., P. sulcata Taylor, and P.
tiliacea (Hoffm.) Ach. These species were chosen
because they are all abundant in the country and
display a considerable diversity of lichen sub-
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250238

stances present in the thallus. Lichens were col-lected in a rural unpolluted area of Oporto district
(northwest Portugal), and kept dry under normal
laboratory conditions for no more than a week.
The day before the experiment, the lichen was
placed on a moist paper filter. Adhering particles
and organisms were carefully removed from the
thallus with the aid of fine stainless steel tweezers
and a binocular microscope. The lichen waswashed twice in Mg2�-PIPES buffer, and placed
on a moist paper filter.
2.2. General layout of the experiments
Mg2�-PIPES buffer (10 mM free acid, 8.5 mM
Mg2�, 26 mM ionic strength; Cabral, 1992), pH
6.50, was used in all experiments since: (i) The use
of buffered solutions is indispensable in studies ofcopper toxicity (Baes and Mesmer, 1976; Buffle,
1990; Sadiq, 1992). (ii) At pH 6.50, most copper
exists as hydrated Cu2� ions (hydroxy and car-
bonate complexes are formed at higher pH), and
these is considered the main toxic species (see
Section 4). (iii) Lichen samples immersed in this
medium showed preserved vitality for several
hours (assessed by the potassium and mannitolconcentrations).
For membrane damage and copper binding to
lichen thallus experiments, samples (40 mg fresh
weight) were placed in small beakers containing 20
ml Mg2�-PIPES buffer. Volumes of a 100 mM
CuSO4 �/5H2O solution were then added, and the
beakers were incubated in a waterbath with
shaking (75 rpm min�1) at 25.0 8C. The use ofrather high copper concentrations (25�/500 mM)
was necessary in order to obtain significant
potassium loss from the cells within 6 h of copper
treatment. At the end of the incubation period, the
samples were removed from the solutions. Total
potassium, intracellular potassium, and total cop-
per in the samples were assayed by flame emission
photometry and atomic absorption spectrometry,respectively.
For copper binding to acetone extracts experi-
ments, the extracts (see below for method of
preparation) were diluted in Mg2�-PIPES buffer,
and transferred to a thermostatized beaker, at
25.0 8C. Small volumes of a concentrated CuSO4 �/
5H2O solution were added to the diluted extracts.After each addition, the free Cu2� concentration
was assayed by ion-selective electrode potentiome-
try. Acetone extracts were titrated immediately
after preparation.
2.3. Membrane damage
Membrane damage was assessed by measuringthe total and intracellular potassium concentra-
tions of samples exposed to copper. For measuring
total potassium concentration, lichen samples were
treated with 500 mM copper for 0, 10, 30, 60, 120,
240, 300, and 360 min. Controls were treated for 0
and 360 min. At the end of the incubation period,
samples were removed from the beakers, blotted
dry, and the total potassium concentration assayedby flame emission photometry, as described below.
For measuring intracellular potassium concentra-
tion, lichen samples were treated with 0, 25, 50,
100, 150, 200, 250, 300, 350, 400, 450, and 500 mM
copper for 300 min. At the end of the incubation
period, samples were removed from the beakers
and blotted dry. For the determination of intra-
cellular potassium, samples were immersed in 10ml distilled water for 30 min, to remove surface
and intercellular potassium, followed by two
treatments with 10 ml NiCl2 (20 mM) for 30 min
each, to remove cell wall bound potassium (Brown
and Beckett, 1984). At the end of the treatments,
samples were removed from the beakers, blotted
dry, and the potassium concentration was assayed
by flame emission photometry, as described below.For each copper concentration, the intracellular
potassium concentration was expressed as a per-
centage of the untreated samples. These percen-
tages were converted into probits, and plotted
against the log of copper dose. The resulting plots
showed a linear relationship. The copper concen-
tration needed to release 75% of intracellular
potassium (in relation to untreated samples),ED75, was calculated using the parameters of the
regression line.
2.4. Copper binding to the thallus
Lichen samples were treated with 500 mM
copper for 0, 10, 30, 60, 120, 240, 300, and 360
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 239

min. At the end of the incubation period, sampleswere removed from the beakers, blotted dry, and
the copper concentration of the samples was
assayed by atomic absorption spectrophotometry,
as described below.
2.5. Acetone extracts of the thallus
Some lichen substances are thermolabile, being
degraded at high temperatures (Culberson et al.,1977; Mirando and Fahselt, 1978). For this reason,
untreated lichen samples (ca. 845 mg fresh weight)
were dried at moderate temperatures (45 8C) for a
restricted period of time (18�/24 h; Culberson et
al., 1977). After drying, samples (ca. 300 mg dry
weight) were cut into pieces with the aid of
stainless steel scissors, and placed in a small
beaker. Samples were not ground since thisprocedure resulted in a significant contamination
of the extracts with intracellular constituents such
as potassium, amino acids, and chlorophylls (un-
published observations). Acetone (1.8 ml) was
then added, and the beaker incubated in a water-
bath with moderate shaking, at 25 8C for 1.5 h. At
the end of the incubation period, the acetone
extract was separated from the lichen fragments,and another volume of acetone was added. After
another period of incubation, the second acetone
extract was added to the first extract. The pooled
extract was stored in a closed tube, at room
temperature. Further treatments of the lichen
fragments with acetone did not result in significant
additional extraction of lichen substances (unpub-
lished observations).Acetone extracts were analyzed for amino acids,
polyols, sugars, and lichen substances. For the
assay of amino acids, polyols, and sugars, acetone
extracts were evaporated to dryness with gentle
heat, and distilled water was added to the residue.
Amino acids were assayed by the ninhydrin
method as described by Rosen (1957), using lysine
as the standard. Polyols were assayed using theperiodate method (Lewis and Smith, 1967; Dudley
and Lechowicz, 1987), with mannitol as the
standard. Sugars were assayed by the cysteine�/
sulphuric acid method (Chaplin, 1986) and by
the phenol�/sulphuric acid method (Dubois et al.,
1956), using glucose as the standard. Lichen
substances were assayed by UV spectrophotome-try and chromatography. For UV spectrophoto-
metry, acetone extracts were evaporated to dryness
with gentle heat, and the residue was dissolved in
methanol. The methanolic extracts were diluted in
buffer, prior to analysis. Spectra were recorded in
a double-beam model spectrophotometer (Jasco;
model V-530) in the 240�/350 nm range. Lichen
substances were identified by one-dimensionalthin-layer chromatography using the three-sol-
vent-system method standardized for lichen pro-
ducts (Culberson and Kristinsson, 1970;
Culberson, 1972; Santesson, 1973; White and
James, 1985; Orange et al., 2001). The acetone
extracts of the studied lichens and solutions of
pure lichen substances (atranorin, norstictic acid,
scrobiculin, stictic acid, and (�/)usnic acid) wereapplied to Merck silica gel 60 F-254 aluminum or
glass plates (20�/20 cm). Plates were developed in
the diagnostic solvents A, B and C. After the
chromatograms had air-dried, they were sprayed
with 10% H2SO4 and heated at 100 8C until
colours developed. Identification of lichen sub-
stances present in the extracts was carried out by
comparison with data reported in the literatureand the behaviour of pure compounds.
2.6. Copper binding to acetone extracts
The acetone extracts (180 ml) were diluted in
Mg2�-PIPES buffer (10 ml), and titrated with a
CuSO4 �/5H2O solution. Free Cu2� was assayed
with a copper-selective electrode (Metrohm Ltd.;
model 6.0502.140), together with a double junctionAg/AgCl/saturated KCl reference electrode (Me-
trohm; model 6.0726.100). Binding of Cu2� to P.
caperata , P. perlata , P. sulcata and P. tiliacea
acetone extracts was represented by successive
Langmuirian curves, each corresponding to an
absorption phase. The delimitation of the absorp-
tion phases was carried out by visual inspection of
the absorption isotherms (bound [Cu2�] vs. free(equilibrium) [Cu2�]), and confirmed by examina-
tion of the free [Cu2�]/bound [Cu2�] vs. free
[Cu2�] plots (Van der Berg and Kramer, 1979;
Hart, 1981; Neubecker and Allen, 1983).The
complexation capacity was calculated as the in-
verse of the slope values for each regression line,
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250240
free [Cu2�]/bound [Cu2�] vs. free [Cu2�], corre-sponding to the last absorption phase (Van der
Berg and Kramer, 1979; Hart, 1981; Neubecker
and Allen, 1983).
2.7. Assay of potassium by flame emission
photometry and copper by atomic absorption
spectrophotometry
The assay was carried out in digested subsam-
ples of dried lichen samples (at 60 8C). The
digestion was performed by adding 1 ml ofconcentrated HNO3 to the lichen subsamples,
and heating in a boiling waterbath, until dissolu-
tion. The determination of potassium was carried
out in a flame photometer (Jenway; model PFP7)
and copper in an atomic absorption spectrophot-
ometer (Philips; model PU 9200X). The accuracy
of this procedure was checked by comparison with
determinations carried out on reference lichenmaterial (Evernia prunastri ; reference material
IAEA-336; Stone et al., 1995). Values found for
the potassium and copper concentrations of re-
ference material were within the 95% confidence
interval of the mean certified values, indicating
that the procedure used in the determinations was
accurate.
2.8. Chemical and solutions
Solutions and standards were prepared with
deionised double distilled water or analytical grade
acetone or methanol. Mg2�-PIPES (1,4-piperazi-
nediethanesulfonate) buffer, pH 6.50, was pre-
pared by neutralizing the free acid (Sigma) with
MgO. Reference lichen material (E. prunastri ,
IAEA 336) was bought from Promochem Com-
pany. Atranorin was from Sigma and (�/)usnicacid from Fluka. Norstictic acid, scrobiculin, and
stictic acid were generous gifts from Dr. Thorsten
Lumbsch (Essen University, Germany).
2.9. Statistics
Correlation between variables was assessed by
determining the linear correlation coefficient,
using Microsoft Excel 2000 program.
3. Results
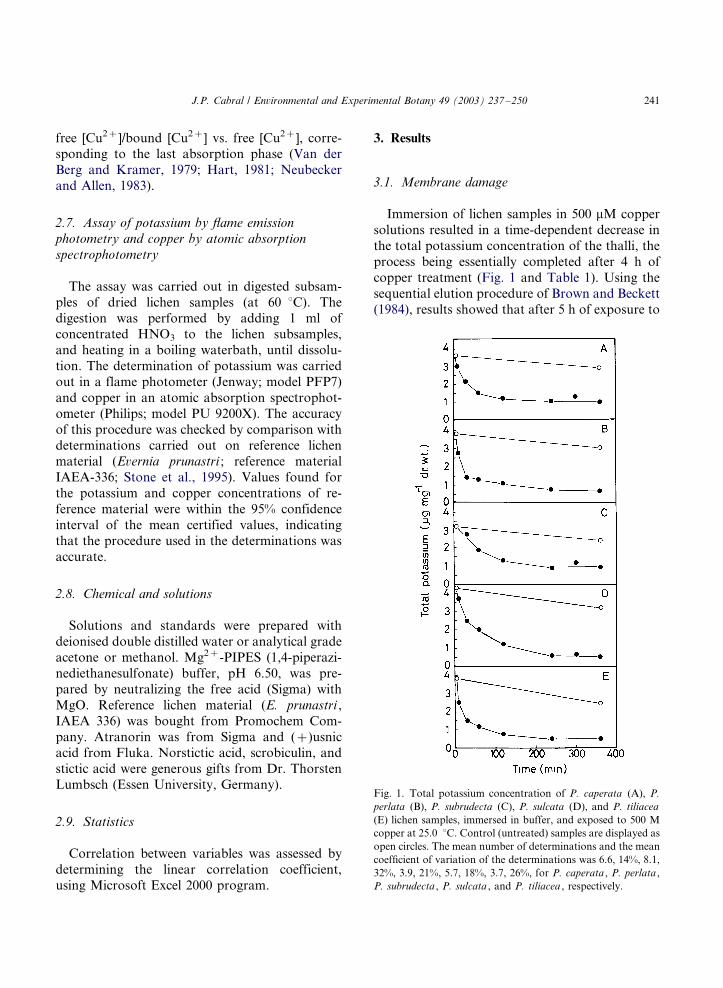
3.1. Membrane damage
Immersion of lichen samples in 500 mM copper
solutions resulted in a time-dependent decrease in
the total potassium concentration of the thalli, the
process being essentially completed after 4 h of
copper treatment (Fig. 1 and Table 1). Using the
sequential elution procedure of Brown and Beckett
(1984), results showed that after 5 h of exposure to
Fig. 1. Total potassium concentration of P. caperata (A), P.
perlata (B), P. subrudecta (C), P. sulcata (D), and P. tiliacea
(E) lichen samples, immersed in buffer, and exposed to 500 M
copper at 25.0 8C. Control (untreated) samples are displayed as
open circles. The mean number of determinations and the mean
coefficient of variation of the determinations was 6.6, 14%, 8.1,
32%, 3.9, 21%, 5.7, 18%, 3.7, 26%, for P. caperata , P. perlata ,
P. subrudecta , P. sulcata , and P. tiliacea , respectively.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 241
500 mM copper, only a small fraction (4%, mean
value for the five species) of the potassium present
in the thallus was extracellular. Potassium leaked
from inside the cells after membrane damage was
therefore released into the suspending medium and
was not bound to cell walls and present in the
intercellular spaces. The observed decrease in total
potassium therefore reflected the extent of copper-
induced membrane damage.
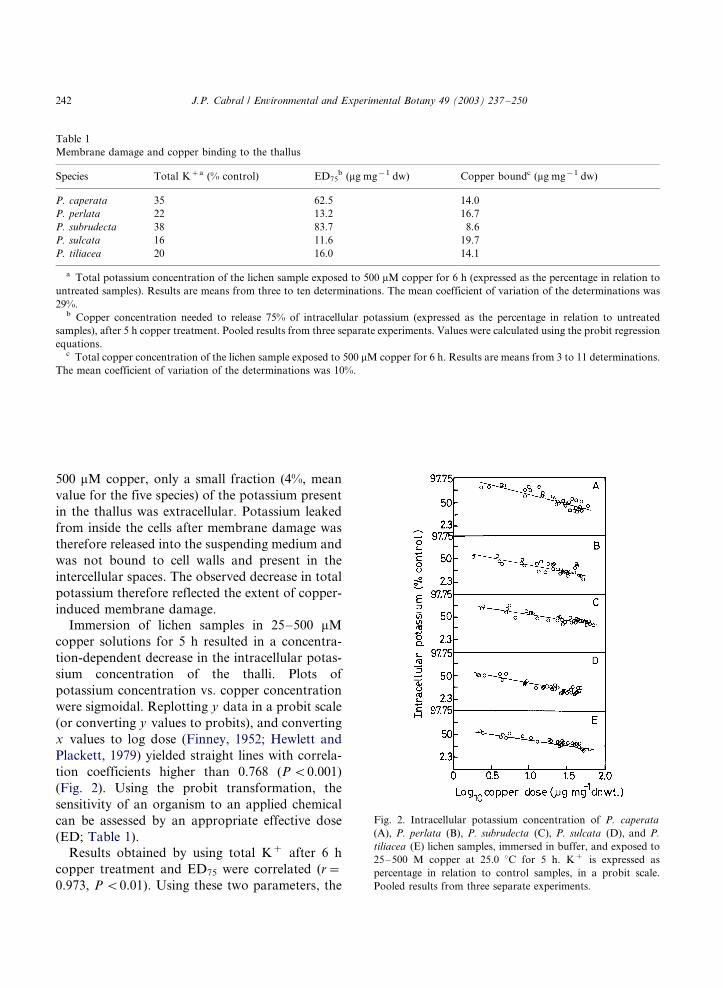
Immersion of lichen samples in 25�/500 mM
copper solutions for 5 h resulted in a concentra-
tion-dependent decrease in the intracellular potas-
sium concentration of the thalli. Plots of
potassium concentration vs. copper concentration
were sigmoidal. Replotting y data in a probit scale
(or converting y values to probits), and converting
x values to log dose (Finney, 1952; Hewlett and
Plackett, 1979) yielded straight lines with correla-
tion coefficients higher than 0.768 (P B/0.001)
(Fig. 2). Using the probit transformation, the
sensitivity of an organism to an applied chemical
can be assessed by an appropriate effective dose
(ED; Table 1).
Results obtained by using total K� after 6 h
copper treatment and ED75 were correlated (r�/
0.973, P B/0.01). Using these two parameters, the
Table 1
Membrane damage and copper binding to the thallus
Species Total K�a (% control) ED75b (mg mg�1 dw) Copper boundc (mg mg�1 dw)
P. caperata 35 62.5 14.0
P. perlata 22 13.2 16.7
P. subrudecta 38 83.7 8.6
P. sulcata 16 11.6 19.7
P. tiliacea 20 16.0 14.1
a Total potassium concentration of the lichen sample exposed to 500 mM copper for 6 h (expressed as the percentage in relation to
untreated samples). Results are means from three to ten determinations. The mean coefficient of variation of the determinations was
29%.b Copper concentration needed to release 75% of intracellular potassium (expressed as the percentage in relation to untreated
samples), after 5 h copper treatment. Pooled results from three separate experiments. Values were calculated using the probit regression
equations.c Total copper concentration of the lichen sample exposed to 500 mM copper for 6 h. Results are means from 3 to 11 determinations.
The mean coefficient of variation of the determinations was 10%.
Fig. 2. Intracellular potassium concentration of P. caperata
(A), P. perlata (B), P. subrudecta (C), P. sulcata (D), and P.
tiliacea (E) lichen samples, immersed in buffer, and exposed to
25�/500 M copper at 25.0 8C for 5 h. K� is expressed as
percentage in relation to control samples, in a probit scale.
Pooled results from three separate experiments.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250242
copper-induced membrane damage increased inthe following order: P. subrudecta , P. caperata , P.
perlata , P. tiliacea , and P. sulcata .
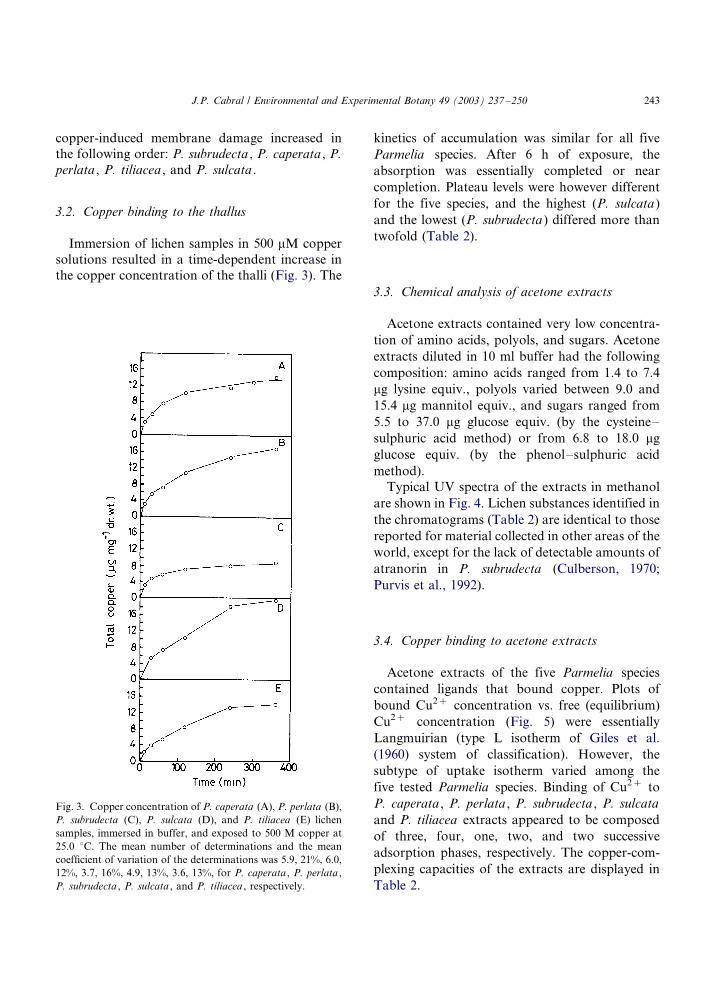
3.2. Copper binding to the thallus
Immersion of lichen samples in 500 mM copper
solutions resulted in a time-dependent increase in
the copper concentration of the thalli (Fig. 3). The
kinetics of accumulation was similar for all five
Parmelia species. After 6 h of exposure, the
absorption was essentially completed or near
completion. Plateau levels were however different
for the five species, and the highest (P. sulcata )
and the lowest (P. subrudecta ) differed more than
twofold (Table 2).
3.3. Chemical analysis of acetone extracts
Acetone extracts contained very low concentra-
tion of amino acids, polyols, and sugars. Acetone
extracts diluted in 10 ml buffer had the following
composition: amino acids ranged from 1.4 to 7.4
mg lysine equiv., polyols varied between 9.0 and
15.4 mg mannitol equiv., and sugars ranged from
5.5 to 37.0 mg glucose equiv. (by the cysteine�/
sulphuric acid method) or from 6.8 to 18.0 mg
glucose equiv. (by the phenol�/sulphuric acid
method).Typical UV spectra of the extracts in methanol
are shown in Fig. 4. Lichen substances identified in
the chromatograms (Table 2) are identical to those
reported for material collected in other areas of the
world, except for the lack of detectable amounts of
atranorin in P. subrudecta (Culberson, 1970;
Purvis et al., 1992).
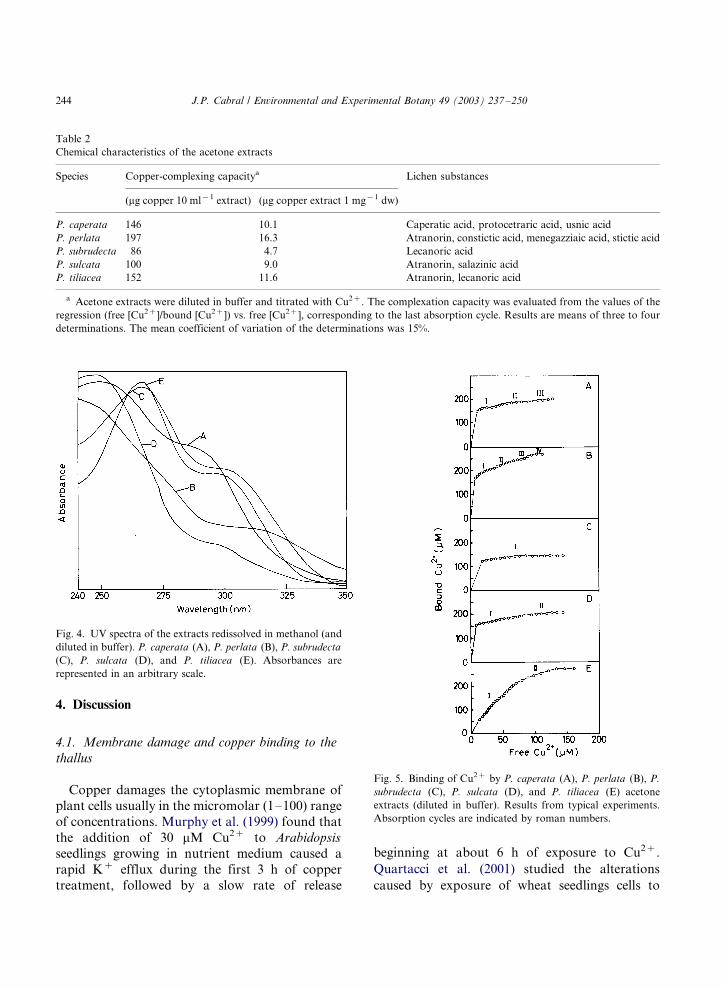
3.4. Copper binding to acetone extracts
Acetone extracts of the five Parmelia species
contained ligands that bound copper. Plots of
bound Cu2� concentration vs. free (equilibrium)
Cu2� concentration (Fig. 5) were essentially
Langmuirian (type L isotherm of Giles et al.
(1960) system of classification). However, the
subtype of uptake isotherm varied among the
five tested Parmelia species. Binding of Cu2� to
P. caperata , P. perlata , P. subrudecta , P. sulcata
and P. tiliacea extracts appeared to be composed
of three, four, one, two, and two successive
adsorption phases, respectively. The copper-com-
plexing capacities of the extracts are displayed in
Table 2.
Fig. 3. Copper concentration of P. caperata (A), P. perlata (B),
P. subrudecta (C), P. sulcata (D), and P. tiliacea (E) lichen
samples, immersed in buffer, and exposed to 500 M copper at
25.0 8C. The mean number of determinations and the mean
coefficient of variation of the determinations was 5.9, 21%, 6.0,
12%, 3.7, 16%, 4.9, 13%, 3.6, 13%, for P. caperata , P. perlata ,
P. subrudecta , P. sulcata , and P. tiliacea , respectively.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 243
4. Discussion
4.1. Membrane damage and copper binding to the
thallus
Copper damages the cytoplasmic membrane ofplant cells usually in the micromolar (1�/100) range
of concentrations. Murphy et al. (1999) found that
the addition of 30 mM Cu2� to Arabidopsis
seedlings growing in nutrient medium caused a
rapid K� efflux during the first 3 h of copper
treatment, followed by a slow rate of release
beginning at about 6 h of exposure to Cu2�.
Quartacci et al. (2001) studied the alterations
caused by exposure of wheat seedlings cells to
Table 2
Chemical characteristics of the acetone extracts
Species Copper-complexing capacitya Lichen substances
(mg copper 10 ml�1 extract) (mg copper extract 1 mg�1 dw)
P. caperata 146 10.1 Caperatic acid, protocetraric acid, usnic acid
P. perlata 197 16.3 Atranorin, constictic acid, menegazziaic acid, stictic acid
P. subrudecta 86 4.7 Lecanoric acid
P. sulcata 100 9.0 Atranorin, salazinic acid
P. tiliacea 152 11.6 Atranorin, lecanoric acid
a Acetone extracts were diluted in buffer and titrated with Cu2�. The complexation capacity was evaluated from the values of the
regression (free [Cu2�]/bound [Cu2�]) vs. free [Cu2�], corresponding to the last absorption cycle. Results are means of three to four
determinations. The mean coefficient of variation of the determinations was 15%.
Fig. 4. UV spectra of the extracts redissolved in methanol (and
diluted in buffer). P. caperata (A), P. perlata (B), P. subrudecta
(C), P. sulcata (D), and P. tiliacea (E). Absorbances are
represented in an arbitrary scale.
Fig. 5. Binding of Cu2� by P. caperata (A), P. perlata (B), P.
subrudecta (C), P. sulcata (D), and P. tiliacea (E) acetone
extracts (diluted in buffer). Results from typical experiments.
Absorption cycles are indicated by roman numbers.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250244

Cu2� for 11 days. After copper treatment, rootswere harvested and incubated in distilled water for
24 h. Whereas seedlings grown with 0 or 0.3 mM
Cu2� and placed in distilled water released 30% of
the total K�, seedlings grown with 50 mM Cu2�
released 60% of cellular potassium, indicating that
this copper concentration damaged the cytoplas-
mic membrane of treated cells. Arduini et al.
(1995) studied the effects of Cu2� on pine seed-lings grown in nutrient solution. After 1, 3, 7, and
10 days of exposure to 0.012, 0.1, 1, or 5 mM
Cu2�, root tips were excised and placed in
aqueous solution of trypan blue. Penetration of
the dye was observed in most of the cells of
seedlings exposed to 1 mM Cu2� for 10 days,
indicating that the cytoplasmic membrane had
been damaged. In this work, it was necessary touse considerably higher (up to 500 mM) copper
concentrations, in order to cause a pronounced
release of potassium from the cells. This can be
due to the use of shorter exposure periods (few
hours instead of several days as often used with
plants) and/or an intrinsic relative resistance of the
studied lichen species to copper.
Immersion of lichen samples in 500 mM coppersolutions resulted in a pronounced release of K�
from the cells and a significant accumulation of
copper in the thalli. The kinetics of these two
processes were however different. Whereas the
decrease in thallus potassium was essentially
completed after 4 h copper treatment, the absorp-
tion of copper increased up to 6 h of incubation.
Moreover, the amount of copper bound to thethallus exceeded twice the amount of potassium
released from the thallus, suggesting that cupric
ions reached intracellular sites in the thallus, and
K�/Cu2� exchange was not electroneutral. In
normal physiological conditions, copper uptake
and trafficking in eukaryotic organisms appears to
follow a general pattern. Cu2� is reduced to Cu�
by a cytoplasmic membrane reductase (Hassettand Kosman, 1995; Hill et al., 1996). Cuprous ions
are then transported to the cytoplasm by specia-
lized transporter proteins (Lin et al., 1997; Fox
and Guerinot, 1998; Harris, 2000). Once inside the
cells, Cu� ions are bound to glutathione or to
specialized proteins (metallochaperones) (Fox and
Guerinot, 1998; Harris, 2000). These chaperones
guide copper to their appropriate biological part-ners (such as the cytochrome c oxidase and super-
oxide dismutase), and prevent intracellular copper
toxicity by decreasing the total cytoplasmic-free
copper to extremely low levels (Pufahl et al., 1997;
Portnoy et al., 1999; O’Halloran and Culotta,
2000). These processes occur in growing condi-
tions, at very low copper concentrations (usually
less than 1 mM Cu2�). In this work, very muchhigher (and stressing) copper concentrations were
used. Therefore, copper could have been accumu-
lated inside the cells as cuprous ions, as reported
for growing cells, and/or, if gross damage occurred
in the cytoplasmic membrane, copper could have
crossed the permeability barriers as unaltered
Cu2� ions.
The extent of released potassium (assessed byresidual K� and ED75) and the amount of copper
bound to the thallus were correlated but only at
0.05B/P B/0.10, suggesting that membrane da-
mage was proportional to the amount of bound
copper, but other factors were operative. Among
these, it is likely that a fraction of copper bound to
the thallus was extracellular, and therefore was not
directly involved in membrane damage.
4.2. Chemical composition of the acetone extracts
and copper-binding capacity
As expected from the method of preparation of
the extracts, acetone extracts contained very low
concentrations of pool and intracellular metabo-
lites such as amino acids, polyols, and sugars, but
significant concentration of lichen substances.Analysis of the extracts by thin-layer chromato-
graphy confirmed UV spectra (Hall, 1956; Rao et
al., 1967; Huneck, 1973). P. caperata extract
displayed maxima at 249�/254 and 283�/286 nm,
in accordance with the presence of protocetraric
acid (peaks at 239 and 314 nm) and usnic acid
(peaks at 232 and 282 nm). P. perlata extract
showed an inflexion at 242 nm and a smooth peakat 304�/310 nm, in accordance with the presence of
atranorin (peak at 252 nm; inflexion at 290 nm),
and b-orcinol depsidones*/constictic, menegaz-
ziaic, and stictic acids (peaks at 238 and 312
nm). P. subrudecta and P. tiliacea showed maxima
at 266�/267 and 296�/299 nm, close to the peaks of
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 245

orcinol depsides�/lecanoric acid, at 270 and 307
nm. P. sulcata extract displayed a maximum at
247�/250 nm and an inflexion at 294�/299 nm, in
accordance with the atranorin and salazinic acid
(maxima at 236.5 and 313 nm).
Titration of the extracts with copper and assay
of free copper by ion-selective electrode potentio-
metry revealed the presence of copper-binding
ligands. Although Cu2� coordinate more strongly
with nitrogen- and sulphur-containing ligands
than with oxygen-type ligands, cupric ions can
form stable chelates with ligands containing only
oxygen (Irving and Williams, 1953; Smith and
Martell, 1977). Data reported in the literature
show that Cu2� can form stable chelate com-
pounds with neighbouring hydroxyl, and/or car-
bonyl groups such as aldehyde, carboxylate, ester,
or ketone (Calvin and Wilson, 1945; Syers, 1969;
Iskandar and Syers, 1972; Casellato et al., 1977;
Smith and Martell, 1977). An analysis of the
molecular structure of the lichen substances pre-
sent in the extracts (Fig. 6) shows that these
molecules have neighbouring hydroxyl and/or
carbonyl groups, and therefore it is likely that
the copper-binding ligands detected in the extracts
were, partially or totally, the identified lichen
substances. This conclusion is further supported
by the presence of several lichen acids in the
extracts, and the existence of successive phases ofcopper binding.
4.3. Copper binding to acetone extracts and
membrane damage
Data reported in the literature indicate thatcopper-complexing compounds usually reduce
copper toxicity, since the main toxic species is
the free hydrated cupric ions (hydroxy complexes
are also toxic, but other inorganic copper com-
plexes are much less toxic), and most organic
copper complexes are non-toxic (however, some
lipophilic organocopper complexes can be very
toxic; see below) (Florence, 1983; Florence andStauber, 1986; Sadiq, 1992; De Filippis and
Pallaghy, 1994). However, in this work, no sig-
nificant correlation (r B/0.72, P �/0.10) was found
between the Cu2�-binding capacity of acetone
extracts and copper-induced membrane damage
(Table 2; Table 3), and therefore our initial
hypothesis was rejected. Whereas membrane da-
mage increased in the following order: P. subru-
decta , P. caperata , P. perlata , P. tiliacea , and P.
sulcata ; and Cu2�-complexing capacity decreased
in the following order: P. perlata , P. tiliacea , P.
caperata , P. sulcata , and P. subrudecta . It can,
therefore, be concluded that in the studied Parme-
Fig. 6. Molecular formula of selected lichen substances identified in the acetone extracts. One representative substance of each class is
shown.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250246

lia species, and in the experimental conditions
used in this work, copper toxicity was not a simple
function of the Cu2�-binding properties of the
lichen substances present in the thallus, others
factors being operative.
The following hypotheses can be formulated to
interpret these results:
i) Lichen substances, isolated, in vitro could
bind considerable amounts of copper, but
when present in the living thallus, the binding
was not significant. In the experimental con-
ditions used-lichen samples immersed in water
for a restricted period of time, lichen sub-
stances present in the thallus could have
ineffective in binding considerable amounts
of copper. This interpretation is supported by
data reported by Brown (1976) showing that
the pretreatment of lichens with acetone prior
to metal exposure did not resulted in de-
creased metal binding (it should be stressed,
however, that in these experiments much
higher metal concentrations were used than
in the present work). According to Syers and
Iskandar (1973), lichen substances, although
having a low solubility in water, do form, in
natural conditions, soluble metal complexes,
which are directly involved in the biogeochem-
ical weathering of rocks and minerals. How-
ever, these are long-term processes and not
directly comparable with the time scale of the
experiments reported in this work, with in-
cubation periods only up to 6 h. Rapid
formation of soluble coloured complexes
with water suspensions of minerals and
rocks was described, but this occurred with
isolated lichen compounds or ground lichen
thalli.
ii) Lichen substances present in the thallus did
bind copper, but saturation was exceeded, and
most of the absorbed cupric ions remained
free, and reached target sites on the cytoplas-
mic membrane. According to this interpreta-
tion, at low copper concentrations, lichen
substances should have prevented copper
toxicity. This interpretation, however, is not
supported by the following results: (1) Probits
were linear functions of copper dose in theentire range of copper concentrations, suggest-
ing a similar mechanism throughout. (2)
Intracellular potassium after treatment with
a low copper dose (25 mM Cu2� correspond-
ing to 2 mg copper mg�1 dw) for 5 h was not
correlated with the copper-complexing capa-
city of the extracts (Fig. 2 and Table 2). The
existence of complexes between copper and
lichen substances in the thallus has indeed
been demonstrated, but only for species
adapted to high metal concentrations. Purvis
et al. (1985, 1987, 1990) studied several lichen
species occurring on copper-rich rocks, and
demonstrated the existence of copper�/norstic-
tic acid and copper�/psoromic acid complexes,
resulting in a green colouration of the thallus.
The Parmelia lichens used in this work were
not collected in copper-rich substrata. There-
fore, the concentration of lichen acids in the
thallus could have been insufficiently high to
bind most of the adsorbed copper in the
experiments.
iii) Some of copper�/lichen substance complexes
were toxic, and therefore increased, and not
decreased, copper availability. Data reported
in the literature do indicate that several
copper-complexing compounds can increasecopper toxicity by the formation of lipid
soluble complexes, which can rapidly pene-
trate the membranes. In addition, lipid-soluble
complexes carry into the membrane and
cytoplasm both copper and the ligand, which
may exert its own toxicity (Florence and
Stauber, 1986). Florence and Stauber (1986)
studied the toxicity of several copper com-plexes to the bacillariophyta Nitzschia closter-
ium and to the chlorophyta Chlorella
pyrenoidosa . It was found that, in general,
ligands which produced water-soluble com-
plexes with copper reduced its toxicity,
whereas ligands which strongly chelated cop-
per to yield lipid-soluble complexes, greatly
increased toxicity. Toxicity increased withincreasing lipophilicity of copper complexes.
The testing of these hypotheses will require the
study of the copper-binding characteristics of pure
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 247

lichen substances, and the assay of the toxicity ofcomplex copper�/lichen substances.
Acknowledgements
I am specially indebted to Dr. Olafsdottir
(University of Iceland, Iceland) for helpful discus-sions, and to Dr. Thorsten Lumbsch (University of
Essen, Germany) for giving samples of pure lichen
substances. The excellent technical assistance of
Mr. Sousa (Faculty of Sciences, Oporto Univer-
sity) on the atomic absorption spectrophotometer
was greatly appreciated.
References
Ahmadjian, V., 1993. The Lichen Symbiosis. Wiley, New York.
Arduini, I., Godbold, D.L., Onnis, A., 1995. Influence of
copper on root growth and morphology of Pinus pinea L.
and Pinus pinaster Ait. seedlings. Tree Physiol. 15, 411�/415.
Avery, S.V., Howlett, N.G., Radice, S., 1996. Copper toxicity
towards Saccharomyces cerevisiae : dependence on plasma
membrane fatty acid composition. Appl. Environ. Micro-
biol. 62, 3960�/3966.
Baes, C.F., Mesmer, R.E., 1976. The Hydrolysis of Cations.
Wiley, New York.
Brown, D.H., 1976. Mineral uptake by lichens. In: Brown, H.,
Hawksworth, D.D.L., Bailey, R.H. (Eds.), Lichenology:
Progress and Problems. Academic Press, London, pp. 419�/
439.
Brown, D.H., Beckett, R.P., 1984. Uptake and effect of cations
on lichen metabolism. Lichenologist 16, 173�/188.
Buffle, J., 1990. Complexation Reactions in Aquatic Systems:
An Analytical Approach. Ellis Horwood, New York.
Cabral, J.P.S., 1992. The assay of free potassium in biological
buffers with a K�-selective glass electrode. Chem. Spec.
Bioavail. 4, 69�/74.
Calvin, M., Wilson, K.W., 1945. Stability of chelate com-
pounds. J. Am. Chem. Soc. 67, 2003�/2007.
Casellato, U., Vigato, P.A., Vidali, M., 1977. Transition metal
complexes with binucleating ligands. Coord. Chem. Rev. 23,
31�/117.
Chaplin, M.F., 1986. Monosaccharides. In: Chaplin, M.F.,
Kennedy, J.F. (Eds.), Carbohydrate Analysis. A Practical
Approach. IRL Press, Oxford, pp. 1�/36.
Chen, L.-M., Lin, C.C., Kao, C.H., 2000. Copper toxicity in
rice seedlings: changes in antioxidative enzyme activities,
H2O2 level, and cell wall peroxidase activity in roots. Bot.
Bull. Acad. Sin. 41, 99�/103.
Culberson, C.F., 1970. Supplement to �/Chemical and Bota-
nical Guide to Lichen Products�/. Bryologist 73, 177�/377.
Culberson, C.F., 1972. Improved conditions and new data for
the identification of lichen products by a standardized thin-
layer chromatographic method. J. Chromatogr. 72, 113�/
125.
Culberson, C.F., Kristinsson, H.D., 1970. A standardized
method for the identification of lichen products. J. Chro-
matogr. 46, 85�/93.
Culberson, C.F., Culberson, W.L., Johnson, A., 1977. Ther-
mally induced chemical artefacts in lichens. Phytochemistry
16, 127�/130.
De Filippis, L.F., Pallaghy, C.K., 1994. Heavy metals: sources
and biological effects. Adv. Limnol. 42, 31�/77.
Demidchik, V., Sokolik, A., Yurin, V., 1997. The effect of
Cu2� on ion transport systems of the plant cell plasma-
lemma. Plant Physiol. 114, 1313�/1325.
Dubois, M., Gilles, K.A., Hamilton, J.K., Rebers, P.A., Smith,
F., 1956. Colorimetric method for the determination of
sugars and related substances. Anal. Chem. 28, 350�/356.
Dudley, S.A., Lechowicz, M.J., 1987. Losses of polyol through
leaching in subartic lichens. Plant Physiol. 83, 813�/815.
Elix, J.A., 1996. Biochemistry and secondary products. In:
Nash, T.H. (Ed.), Lichen Biology. Cambridge University
Press, Cambridge, pp. 154�/180.
Fahselt, D., 1994. Secondary biochemistry of lichens. Symbiosis
16, 117�/165.
Finney, D.J., 1952. Probit Analysis: A Statistical Treatment of
the Sigmoid Response Curve. Cambridge University Press,
Cambridge.
Florence, T.M., 1983. Trace element speciation and aquatic
toxicology. Trends Anal. Chem. 2, 162�/166.
Florence, T.M., Stauber, J.L., 1986. Toxicity of copper com-
plexes to the marine diatom Nitzschia closterium . Aquat.
Toxicol. 8, 11�/26.
Fox, T.C., Guerinot, M.L., 1998. Molecular biology of cation
transport in plants. Annu. Rev. Plant Physiol. Plant Mol.
Biol. 49, 669�/696.
Giles, C.H., MacEwan, T.H., Nakhawa, S.N., Smith, D., 1960.
Studies in adsorption. XI. A system of classification of
solution adsorption isotherms, and its use in diagnosis of
adsorption mechanisms and in measurement of specific
surface areas of solids. J. Chem. Soc. 1960, 3973�/3993.
Hall, M.E., 1956. Ultraviolet absorption spectra of lichen
depsides and depsidones. Science 123, 671.
Harris, E., 2000. Cellular copper transport and metabolism.
Annu. Rev. Nutr. 20, 291�/310.
Hart, B.T., 1981. Trace metal complexing capacity of natural
waters: a review. Environ. Technol. Lett. 2, 95�/110.
Hassall, K.A., 1963. Uptake of copper and its physiological
effects on Chlorella vulgaris . Physiol. Plant. 16, 323�/332.
Hassett, R., Kosman, D.J., 1995. Evidence for Cu(II) reduction
as a component of copper uptake by Saccharomyces
cerevisiae . J. Biol. Chem. 270, 128�/134.
Hewlett, P.S., Plackett, R.L., 1979. The Interpretation of
Quantal Responses in Biology. Edward Arnold, London.
Hill, K.L., Hassett, R., Kosman, D., Merchant, S., 1996.
Regulated copper uptake in Chlamydomonas reinhardtii in
response to copper availability. Plant Physiol. 112, 697�/704.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250248

Howlett, N.G., Avery, S.V., 1997. Induction of lipid peroxida-
tion during heavy metal stress in Saccharomyces cerevisiae
and influence of plasma membrane fatty acid unsaturation.
Appl. Environ. Microbiol. 63, 2971�/2976.
Huneck, S., 1968. Lichen substances. Prog. Phytochem. 1, 223�/
346.
Huneck, S., 1973. Nature of lichen substances. In: Ahmadjian,
V., Hale, M.E. (Eds.), The Lichens. Academic Press, New
York, pp. 495�/546.
Irving, H., Williams, R.J.P., 1953. The stability of transition-
metal complexes. J. Chem. Soc. 1953, 3192�/3210.
Iskandar, I.K., Syers, J.K., 1971. Solubility of lichen com-
pounds in water: pedogenetic implications. Lichenologist 5,
45�/50.
Iskandar, I.K., Syers, J.K., 1972. Metal-complex formation by
lichen compounds. J. Soil Sci. 23, 255�/265.
Joho, M., Ishibe, A., Murayama, T., 1984. The injurious effect
of heavy metal ions on the cell membrane in Saccharomyces
cerevisiae . Trans. Mycol. Soc. Jpn. 25, 485�/488.
Lawrey, J.D., 1986. Biological roles of lichen substances.
Bryologist 89, 111�/122.
Leuckert, C., Ahmadjian, V., Culberson, C.F., Johnson, A.,
1990. Xanthones and depsidones of the lichen Lecanora
dispersa in nature and of its mycobiont in culture. Myco-
logia 82, 370�/378.
Lewis, D.H., Smith, D.C., 1967. Sugar alcohols (polyols) in
fungi and green plants. II. Methods of detection and
quantitative estimation in plant extracts. New Phytol. 66,
185�/204.
Lin, S.-J., Pufahl, R.A., Dancis, A., O’Halloran, T.V., Culotta,
V.C., 1997. A role for the Saccharomyces cerevisiae ATX1
gene in copper trafficking and iron transport. J. Biol. Chem.
272, 9215�/9220.
Meharg, A.A., 1994. Integrated tolerance mechanisms: consti-
tutive and adaptative plant responses to elevated metal
concentrations in the environment. Plant Cell Environ. 17,
989�/993.
Mirando, M., Fahselt, D., 1978. The effect of thallus age and
drying procedure on extractable lichen substances. Can. J.
Bot. 56, 1499�/1504.
Murphy, A.S., Eisinger, W.R., Shaff, J.E., Kochian, L.V., Taiz,
L., 1999. Early copper-induced leakage of K� from
Arabidopsis seedlings is mediated by ion channels and
coupled to citrate efflux. Plant Physiol. 121, 1375�/1382.
Neubecker, T.A., Allen, H.E., 1983. The measurement of
complexation capacity and conditional stability constants
for ligands in natural waters. Water Res. 17, 1�/14.
Nieboer, E., Richardson, D.H.S., 1980. The replacement of the
nondescript term �/heavy metals�/ by a biologically and
chemically significant classification of metal ions. Environ.
Pollut. Ser. B 1, 3�/26.
O’Halloran, T.V., Culotta, V.C., 2000. Metallochaperones, an
intracellular shuttle service for metal ions. J. Biol. Chem.
275, 25057�/25060.
Ohsumi, Y., Kitamoto, K., Anaraku, Y., 1988. Changes in the
permeability barrier of the yeast plasma membrane by
cupric ion. J. Bacteriol. 170, 2676�/2682.
Orange, A., James, P.W., White, F.J., 2001. Microchemical
Methods for the Identification of Lichens. British Lichen
Society, London.
Patsikka, E., Kairavuo, M., .Sersen, F., Aro, E.-M., Tyystjarvi,
E., 2002. Excess copper predisposes photosystem II
to photoinhibition in vivo by outcompeting iron and
causing decrease in leaf chlorophyll. Plant Physiol. 129,
1359�/1367.
Portnoy, M.E., Rosenzweig, A.C., Rae, T., Huffman, D.L.,
O’Hallorn, T.V., Culotta, V.C., 1999. Structure�/function
analyses of the ATX1 metallochaperone. J. Biol. Chem. 274,
15041�/15045.
Pufahl, R.A., Singer, C.P., Peariso, K.L., Lin, S.-G., Schmidt,
P.J., Fahrni, C.J., Culotta, V.C., Penner-Hahn, J.E.,
O’Halloran, T.V., 1997. Metal ion chaperone function
of the soluble Cu(I) receptor Atx1. Science 278, 853�/
856.
Purvis, O.W., Gilbert, O.L., James, P.W., 1985. The influence
of copper mineralization on Acarospora smaragdula . Liche-
nologist 17, 111�/116.
Purvis, O.W., Elix, J.A., Broomhead, J.A., Jones, G.C., 1987.
The occurrence of copper�/norstictic acid in lichens from
cupriferous substrata. Lichenologist 19, 193�/203.
Purvis, O.W., Elix, J.A., Gaul, K.L., 1990. The occurrence of
copper�/psoromic acid in lichens from cupriferous sub-
strata. Lichenologist 22, 345�/354.
Purvis, O.W., Coppins, B.J., Hawksworth, D.L., James, P.W.,
Moore, D.M. (Eds.), 1992. The Lichen Flora of Great
Britain and Ireland. The British Lichen Society, London.
Quartacci, M.F., Cosi, E., Navari-Izzo, F., 2001. Lipids and
NADPH-dependent superoxide production in plasma mem-
brane vesicles from roots of wheat grown under copper
deficiency or excess. J. Exp. Bot. 52, 77�/84.
Rao, P.S., Sarma, K.G., Seshadri, T.R., 1967. The ultraviolet
and infrared spectra of some lichen depsides and depsi-
dones. Proc. Ind. Acad. Sci. A 66, 1�/14.
Rosen, H., 1957. A modified ninhydrin colorimetric analysis for
amino acids. Arch. Biochem. Biophys. 67, 10�/15.
Sadiq, M., 1992. Toxic Metal Chemistry in Marine Environ-
ments. Marcel Dekker, New York.
Sandmann, G., Boger, P., 1980. Copper-mediated lipid perox-
idation processes in photosynthetic membranes. Plant
Physiol. 66, 797�/800.
Santesson, J., 1973. Identification and isolation of lichen
substances. In: Ahmadjian, V., Hale, M.E. (Eds.), The
Lichens. Academic Press, New York, pp. 633�/652.
Smith, R.M., Martell, A.E., Critical Stability Constants.
Volume 3: Other Organic ligands. Plenum Press, New York.
Solhaug, K.A., Gausla, Y., 1996. Parietin, a photoprotective
secondary product of the lichen Xanthoria parietina . Oeco-
logia 108, 412�/418.
Stauber, J.L., Florence, T.M., 1987. Mechanism of toxicity of
ionic copper and copper complexes to algae. Mar. Biol. 94,
511�/519.
Stone, S.F., Freitas, M.C., Parr, R.M., Zeisler, R., 1995.
Elemental characterization of a candidate lichen re-
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250 249

search material*/IAEA-336. Fres. J. Anal. Chem. 352,
227�/231.
Syers, J.K., 1969. Chelating ability of fumarprotocetraric
acid and Parmelia conspersa . Plant Soil 31, 205�/
208.
Syers, J.K., Iskandar, I.K., 1973. Pedogenetic significance of
lichens. In: Ahmadjian, V., Hale, M.E. (Eds.), The Lichens.
Academic Press, New York, pp. 225�/248.
Van der Berg, C.M.G., Kramer, J.R., 1979. Determination of
complexing capacities of ligands in natural waters and
conditional stability constants of the copper complexes by
means of manganese dioxide. Anal. Chim. Acta 106, 113�/
120.
White, F.J., James, P.W., 1985. A new guide to microchemical
techniques for the identification of lichen substances. Br.
Lichen Soc. Bull. 57 (Suppl.), 1�/41.
J.P. Cabral / Environmental and Experimental Botany 49 (2003) 237�/250250