comparison with photosynthesis - meps 316 - comparis… · reduction of carbon compounds oxidation...
TRANSCRIPT

Comparison With Photosynthesis
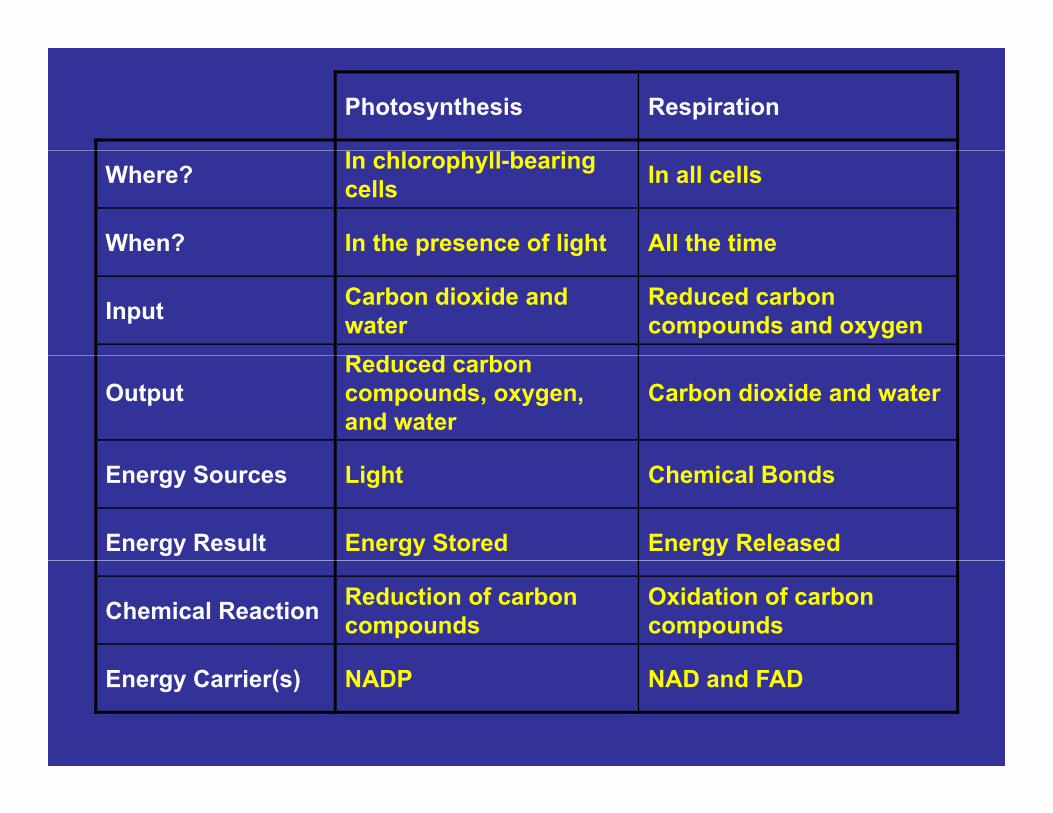
Photosynthesis Respiration
Where? In chlorophyll-bearing cells In all cells
When? In the presence of light All the timep g
Input Carbon dioxide and water
Reduced carbon compounds and oxygen
R d d bOutput
Reduced carbon compounds, oxygen, and water
Carbon dioxide and water
Energy Sources Light Chemical Bonds
Energy Result Energy Stored Energy Released
Chemical Reaction Reduction of carbon compounds
Oxidation of carbon compounds
Energy Carrier(s) NADP NAD and FAD

Cellular Respiration:A O iAn Overview
Cellular respiration consists of a series of pathways by which carbohydrate and other molecules are oxidized for the purpose of retrieving the energy
stored in photosynthesis and to obtain carbon skeletons used in the growth
d i f h lland maintenance of the cell.

Comparison of Photosynthesis d R i iand Respiration
• Photosynthesis:Photosynthesis:6 CO2 + 6 H2O → C6H12O6 + 6 O2
light plantlight, plant
R i ti• Respiration:C6H12O6 + 6 O2 + 6 H2O ↔ 6 CO2 + 12 H2O(Hexose)

Perspective from a Chemical S d iStandpoint
• Respiration represents the oxidation of a 12-p pcarbon molecule sucrose and reduction of 12 molecules of CO2.
C12H22O11 + 13 H2O → 12 CO2+ 48 H+ + 48 e-
12 O2 + 48 H+ + 48 e- → 24 H2O12 O2 48 H 48 e 24 H2O
• Net Reaction
C12H22O11 + 12 O2 → 12 CO2 + 11 H2O
60 ADP + 60 Pi → 60 ATP + 60 H2O60 ADP 60 Pi → 60 ATP 60 H2O

RespirationRespiration• Anabolism: formation of large molecules from small g
molecules; requires an input of energy.
• Catabolism: degradation or breakdown of large• Catabolism: degradation or breakdown of large molecules to small molecules; this process often releases energy.
– Respiration is the major catabolic process that releases energy in all cells.gy
– Respiration involves oxidative breakdown of sugars to CO and H Osugars to CO2 and H2O.

RespirationRespiration• Respiration is a multi-step process inRespiration is a multi step process in
which carbon is oxidized through a series of reactions, which can be divided into four stages:
1. Glycolysis (Embden-Meyerhoff-Parnas)2. Tricarboxylic Acid Cycle (TCA or
Krebs)Krebs)3. Oxidative pentose phosphate pathway4 Electron Transport Chain (ETC)4. Electron Transport Chain (ETC)

RespirationRespiration• Glucose is commonly cited as the main y
substrate in respiration; however, the reduced carbon may come from many sources such as:sources such as:
• Starch• SucroseSucrose• Fructose• Other sugars• Lipids• Organic acids• ProteinsProteins

Overview of RespirationOverview of Respiration
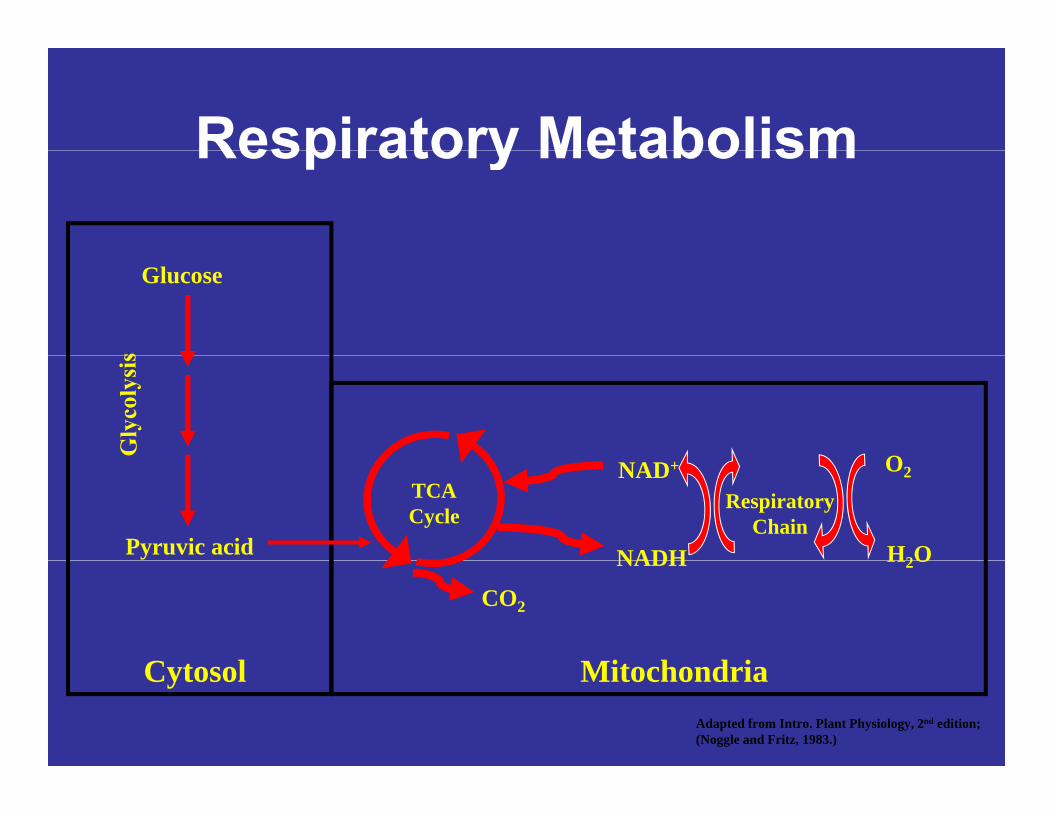
Respiratory MetabolismRespiratory Metabolism
Glucose
ONAD+
NADH
O2
H2O
RespiratoryChain
TCACycle
Pyruvic acid NADH
CO2
H2O
Mit h d iC t l MitochondriaCytosolAdapted from Intro. Plant Physiology, 2nd edition;(Noggle and Fritz, 1983.)

Reactions of Plant Glycolysis and Fermentationand Fermentation

RespirationRespiration• Location of glycolysis: carried out by a group of
l bl l t d i th t l d isoluble enzymes located in the cytosol and in plastids.
• Chemically what happens to sucrose in glycolysis?Chemically what happens to sucrose in glycolysis?
– It undergoes a limited amount of oxidation.Produces 4 molecules of pyruvate (3 C– Produces 4 molecules of pyruvate (3-C compound).
– A little ATP.– Produces stored reducing power in the form of a
reduced pyridine nucleotide, NADH.

Functions of GlycolysisFunctions of Glycolysis1. Converts one hexose molecule into two molecules
f i id d ti l id ti f hof pyruvic acid, and partial oxidation of hexose occurs.
2. Produces a limited amount of ATP.
3. Forms molecules that can be removed from the3. Forms molecules that can be removed from the pathway to synthesize several other constituents of which the plant is made.
4. Produces two molecules of pyruvate that can be oxidized in mitochondria to yield relatively large amounts of ATP, much more than is produced in pglycolysis.

Functions of GlycolysisFunctions of Glycolysis1. Glycolysis converts one hexose molecule y y
into two molecules of pyruvic acid, and partial oxidation of hexose occurs.
– No O2 is used and no CO2 is released.
C i f h h lt i th– Conversion of each hexose results in the reduction of two molecules of NAD+ to NADH (+2H+).
– These NADH can subsequently be oxidized by O2 in the mitochondrion such that NAD+ is regenerated and two molecules of ATP areregenerated and two molecules of ATP are formed.

Functions of GlycolysisFunctions of Glycolysis– Some of these NADH do not enter mitochondria
d d i th t l t d i iand are used in the cytosol to drive various anabolic, reductive processes.
– Example: nitrate reductase an enzyme thatExample: nitrate reductase, an enzyme that transfers two electrons from NADH or, in a few species, from NADPH.
O O ONO3- + NADH + H+ → NO2
- + NAD+ + H2O(Nitrate reductase)
• How many steps in glycolysis result in the formation• How many steps in glycolysis result in the formation of NADH? Only one; during the oxidation of 3-phosphoglyceradehyde to 1,3-bisphosphoglyceric acid, NAD+ is reduced to NADH.acid, NAD is reduced to NADH.

Functions of Glycolysisy y
2 Glycolysis produces ATP2. Glycolysis produces ATP.
• When glucose or fructose enters glycolysis, each is phosphorylated by ATP in reactionseach is phosphorylated by ATP in reactions catalyzed by hexokinase or fructokinase.
• Glucose 6 phosphate and fructose 6 phosphate• Glucose-6-phosphate and fructose-6-phosphate are the products.

Functions of GlycolysisFunctions of Glycolysis– Two ATPs are formed from each triose phosphate p p
derived from the split of fructose-1,6-bisphosphate, making a total of four ATPs for each glucose or fructose respiredeach glucose or fructose respired.
• A yield of four ATPs minus the two (or one)A yield of four ATPs minus the two (or one) required to form fructose-1,6-bisphosphate leaves a net yield of either two or three ATPs for each hexose used in glycolysis (if the PPfor each hexose used in glycolysis (if the PPi-PFK route is used, then three ATPs are formed).

Functions of GlycolysisFunctions of Glycolysis• What two routes exist by whichWhat two routes exist by which
fructose-6-phosphate can be converted to fructose-1,6-bisphosphate?
1. ATP – dependent –phosphofructokinase (ATP-PFK)p p ( )
2. PPi – dependent - PFK2. PPi dependent PFK

Functions of GlycolysisFunctions of Glycolysis
Fructose-6-Pi
ATPPPi ATP-dependent phosphofructokinase
PPi-dependent phosphofructokinase
ADPPi
p p f(ATP-PFK)
phosphofructokinase (PPi-PFK)
Fructose-1,6-bisphosphate, p p

Functions of GlycolysisFunctions of Glycolysis• ATP-phosphofructokinase (ATP-PFK): p p ( )
current evidence suggests that the ATP-PFK route is involved in so-called “maintenance respiration” by cells that are“maintenance respiration” by cells that are not rapidly growing differentiating, or adapting to changing environments (Black p g g g (et al., 1987)
• Reaction occurs mainly in cells that are mature, or nearly so, and that exist for some time in a moderately constant environment.

Functions of GlycolysisFunctions of Glycolysis
• Pyrophosphate phosphofructokinase (PP -PFK): route that is much more(PPi-PFK): route that is much more adaptive and can increase or decrease in importance depending ondecrease in importance depending on developmental processes and environmental conditionsenvironmental conditions.

Functions of GlycolysisFunctions of Glycolysis
• Conversion of glucose or fructose toConversion of glucose or fructose to fructose-1,6-bisphosphate requires:
– 2 ATPs if ATP-PFK route used.
– 1 ATP if PPi-PFK route used.

FermentationFermentation

FermentationFermentation• Glycolysis can function well without O2, but y y 2,without O2 the TCA cycle and electron
transport chain cannot function.
• Why does this present a problem for the continued operation of glycolysis? The cell’s supply of NAD+ is limited and once allcell s supply of NAD is limited, and once all the NAD+ becomes tied up in the reduced state (NADH), the glyceraldehyde-3-phosphate dehydrogenase reaction cannotphosphate dehydrogenase reaction cannot take place.
NADH and pyruvate begin to accumulate when O– NADH and pyruvate begin to accumulate when O2is limiting.

FermentationFermentation• Thus, in the absence of O2, fermentation (anaerobic 2 (
respiration) allows the regeneration of NAD+
needed for glycolysis.
Wh O i l ki l t f th t b li• When O2 is lacking, plants can further metabolize pyruvate by carrying out one or more forms of fermentation metabolism:
– Lactic acid fermentation (does occur in plants and animals): the enzyme lactate dehydrogenase uses NADH to reduce pyruvate to lactate, thus regenerating NAD+.py , g g
– In alcoholic fermentation the two enzymes pyruvate decarboxylase and alcohol dehydrogenase act on pyruvate producing ethanol and CO2; oxidizes NADH.pyruvate producing ethanol and CO2; oxidizes NADH.

Pathways of Fermentation Leading t th R id ti f NADHto the Reoxidation of NADH

Conditions Enhancing F i M b liFermentative Metabolism
• Low (hypoxic) or zero (anoxic)Low (hypoxic) or zero (anoxic) concentrations of ambient oxygen.
• Flooded or water logged soils best example.example.
– Diffusion of O to roots sufficientlyDiffusion of O2 to roots sufficiently reduced to cause root tissues to become hypoxic.

Pentose Phosphate Pathway

Reactions of the Oxidative Pentose Ph h t P th i Hi h Pl tPhosphate Pathway in Higher Plants

Pentose Phosphate Pathway (PPP)(PPP)
• Oxidative pentose pathwayOxidative pentose pathway• Hexose monophosphate shunt• Phosphogluconate pathwayPhosphogluconate pathway
What is the major difference between the jCalvin cycle and the PPP?
C l i l h h t th i d– Calvin cycle – sugar phosphates synthesized.
– PPP – sugar phosphates degraded.g p p g

Pentose Phosphate Pathway (PPP)(PPP)
• Glycolysis and PPP are greatlyGlycolysis and PPP are greatly interwoven.
• What is the one important difference between the PPP and glycolysis?between the PPP and glycolysis?
– PPP electron acceptor: NADP+PPP electron acceptor: NADP
– Glycolysis electron acceptor: NAD+Glycolysis electron acceptor: NAD

Main Functions of the Oxidative P Ph h P hPentose Phosphate Pathway
1. Generates NADPH used as a reductant in biosynthetic processes when NADPH is not being generated by photosynthesis; e.g. fatty acid and several isoprenoids.y p
– Particularly important in non-photosynthetic tissues, e.g. in differentiating tissues, , g g ,germinating seeds and during the hours of darkness.
2 G ib h h i d f2. Generates ribose-5-phosphate required for biosynthesis of nucleotides and nucleic acids (RNA and DNA).( )

Main Functions of the Oxidative P Ph h P hPentose Phosphate Pathway
3. Produces erythrose-4-phosphate required y p p qfor synthesis of shikimic acid, the precursor of aromatic rings.
• Erythrose-4-phosphate + PEP → initial reaction that produces plant phenolic compounds in the shikimic acid pathwayshikimic acid pathway.
4. During the early stages of greening, before leaf tissues become fully photoautotrophic, the oxidative pentose phosphate pathway is thought to be involved in generatingis thought to be involved in generating Calvin cycle intermediates.

Oxidative Pentose Phosphate Pathway:Hi h Pl tHigher Plants
• NADPH is generated in the first two reactions gof the pathway.
• The resulting ribulose 5-phosphate is converted to the glycolytic intermediates fructose 6 phosphate and glyceraldyhyde 3-phosphate through a series of metabolic p p ginterconversions.
• The first two reactions are essentially i ibl hil th i t iirreversible, while the interconversions between ribulose 5-phosphate and the glycolytic compounds are freely reversible.g y y p y

The Citric Acid Cycle (Tricarboxylic Acid or Krebs(Tricarboxylic Acid or Krebs
Cycle)
A Mitochondrial Matrix ProcessA Mitochondrial Matrix Process

RespirationRespiration
• Where are the TCA cycle and the electron transport chain located?electron transport chain located?
Both are located within the confines of the– Both are located within the confines of the membrane-bound organelle known as the mitochondrionmitochondrion.

Reactions and Enzymes of the Pl Ci i A id C lPlant Citric Acid Cycle

Conversion of Pyruvate to Acetyl-C A b P i D h dCoA by Pyruvic Dehydrogenase
• Acetyl-CoA is the “fuel” yof the Krebs Cycle.
• The release of CO2 in the Krebs Cycle accounts for theaccounts for the product CO2 in the summary equation for respiration but no O isrespiration, but no O2 is absorbed during any Krebs Cycle reaction.

Primary Functions of the Krebs C lCycle
1. Reduction of NAD+ and ubiquinone to the electron qdonors NADH and ubiquinol, which are subsequently oxidized to yield ATP.
2. Direct synthesis (substrate-level phosphorylation) of a limited amount of ATP (one ATP for each pyruvate oxidized)pyruvate oxidized).
3. Forms carbon skeletons that can be used to synthesize certain amino acids that, in turn, are converted to large molecules.

Primary Functions of the Krebs C lCycle
• The flavin FAD is usually identified as theThe flavin FAD is usually identified as the acceptor of electrons and H+ from succinic acid, with FADH2 as the product.2
• FAD and FADH2 are bound to succinic acid 2dehydrogenase, but they represent transitory intermediate compounds during the overall
d i f b l bl ireduction of membrane-soluble quinone (ubiquinone) to ubiquinol.

Primary Functions of the Krebs C lCycle
• None of the dehydrogenase enzymes of the y g ycycle use NADP+ as an electron acceptor; NADP+ is usually nearly nondetectable in plant mitochondria.p
• The release of CO2 in the Krebs cycle t f th d t CO i thaccounts for the product CO2 in the summary
equation for respiration.
• No O2 is absorbed during any Krebs-cycle reaction.

RespirationRespiration• What purpose does the TCA cycle serve?p p y
– Brings about the complete oxidation of pyruvate to CO2to CO2.
– Generates a considerable amount of reducing power (about 16 NADH + 4 FADH equivalents perpower (about 16 NADH + 4 FADH2 equivalents per sucrose).
These reactions with one exception involve a– These reactions, with one exception, involve a series of soluble enzymes located in the internal aqueous compartment, or matrix, of the mitochondrionmitochondrion.

Glycolysis, The Pentose Phosphate P th d th Cit i A id C lPathway and the Citric Acid Cycle

Glyoxylate CycleGlyoxylate Cycle

Electron Transport and ATP Synthesis at the InnerSynthesis at the Inner
Mitochondrial Membrane
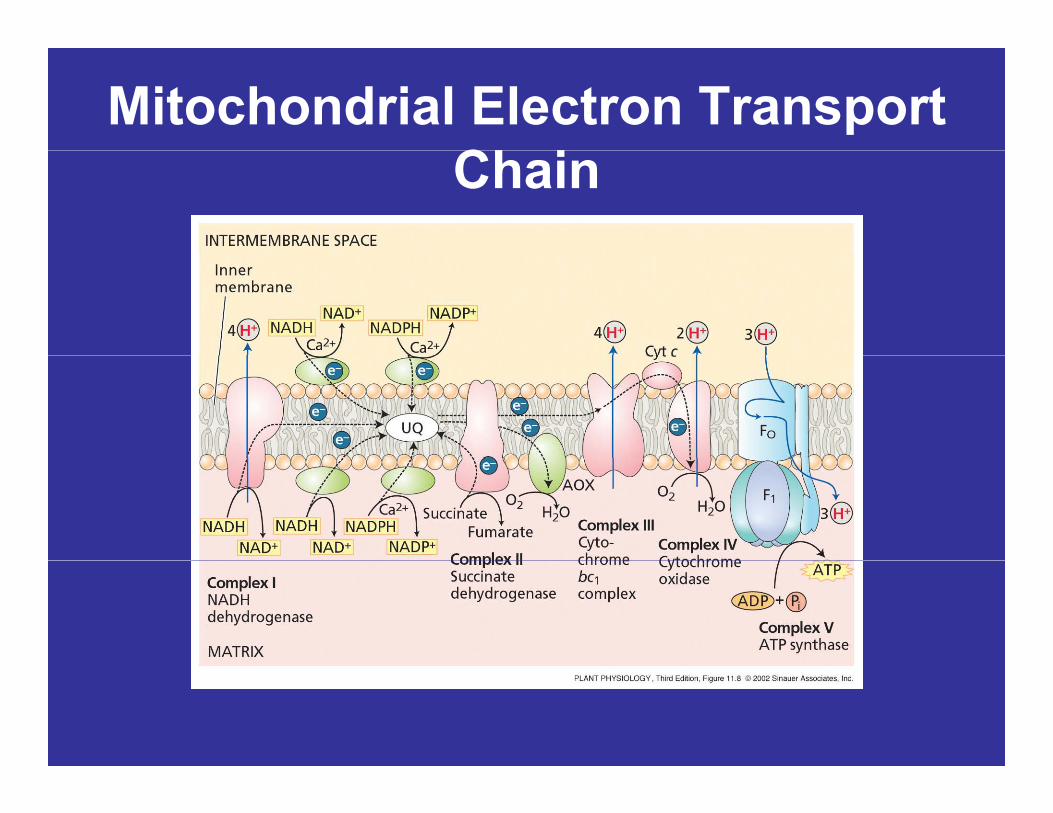
Mitochondrial Electron Transport Ch iChain

Electron Transport ATP Synthesis at th I Mit h d i l M bthe Inner Mitochondrial Membrane
• ATP is the energy carrier used by cells to gy ydrive living processes.
• The chemical energy conserved during the• The chemical energy conserved during the citric acid cycle in the form of NADH and FADH2 (redox equivalents with high-energy
l t ) t b t d t ATP telectrons) must be converted to ATP to perform useful work in the cell.
• This O2-dependent process occurs in the inner mitochondrial membrane through oxidative phosphorylationoxidative phosphorylation.

Electron Transport Chain Catalyzes Fl f El t f NADH t Oa Flow of Electrons from NADH to O2
• For each molecule of sucrose oxidized through glycolysis and the citric acid cycle pathways:
– 4 molecules of NADH are generated in the cytosol.
16 l l f NADH l ( it h d i )– 16 molecules of NADH plus (mitochondria)
– 4 molecules of FADH22
• These reduced compounds must be reoxidized to keep the respiratory processreoxidized to keep the respiratory process functional.

Electron Transport Chain Catalyzes Fl f El t f NADH t Oa Flow of Electrons from NADH to O2
• The electron transport chain catalyzes an electron fl f NADH ( d FADH ) t th fi lflow from NADH (and FADH2) to oxygen, the final electron acceptor of the respiratory process.
• The role of the electron transport chain is to bring about the oxidation of NADH (and FADH2) and, in the process, utilize some of the free energy released to generate an electrochemical proton gradient Δ ~generate an electrochemical proton gradient Δ H+ , across the inner mitochondrial membrane.
Th i di id l l t t t t i
• The individual electron transport proteins are organized into four multi-protein complexes, all of which are localized in the inner mitochondiral membranemembrane.

Electron Transport Chain:C l IComplex I
• Complex I (NADH dehydrogenase) oxidizes p ( y g )electrons from NADH generated in the mitochondrial matrix during the TCA cycle.
– Electron carriers include FMN and several iron-sulfur centers.
• Complex I then transfers these electrons to ubiquinone.
• Four protons are pumped from the matrix to the intermembrane space for every electronthe intermembrane space for every electron pair passing through the complex.

Electron Transport Chain:Ubi iUbiquinone
• Ubiquinone is a small lipid-solubleUbiquinone is a small lipid soluble electron and proton carrier within the inner membrane.
• The ubiquinone (UQ) pool diffusesThe ubiquinone (UQ) pool diffuses freely within the inner membrane and serves to transfer electrons from the dehydrogenases to either complex III or the alternative oxidase.

Electron Transport Chain:C l II ( i t d h d )Complex II (succinate dehydrogenase)
• Oxidation of succinate in the TCA cycleOxidation of succinate in the TCA cycle is catalyzed by this complex.
• The reducing equivalents are transferred via the FADH2 and a smalltransferred via the FADH2 and a small group of iron-sulfur proteins into the ubiquinone pool.q p
• Complex II does not pump protons.Complex II does not pump protons.

Electron Transport Chain:C l III ( t h b l )Complex III (cytochrome bc1 complex)
• Complex III oxidizes reduced ubiquinone p q(ubiquinol) and transfers the electrons via an iron-sulfur center, two b-type cytochromes (b565 and b560) and a membrane-bound ( 565 560)cytochrome c1 to cytochrome c.
F t l t i d• Four protons per electron pair are pumped by complex III.
• Cytochrome c is a small protein loosely attached to (peripheral protein) that transfers electrons from complex III to complex IVelectrons from complex III to complex IV.

Electron Transport Chain:C l IV ( t h id )Complex IV (cytochrome c oxidase)
• Complex IV contains two copper centers (CuA and C ) d t h dCuB) and cytochrome a and a3.
• Complex IV is the terminal oxidase and brings about p gthe four-electron reduction of O2 to two molecules of H2O.
• Two protons are pumped per electron pair.
• Structurally and functionally ubiquinone and theStructurally and functionally, ubiquinone and the cytochrome bc1 complex are very similar to plastoquinone and the cytochrome b6f complex, respectively, in the photosynthetic electron transport p y p y pchain.

Unique Electron Transport E i Pl Mi h d iEnzymes in Plant Mitochondria
• Two NAD(P)H dehydrogenasesTwo NAD(P)H dehydrogenases
– Ca2+-dependent
– Attach to the outer surface of the inner membrane facing intermembrane space.
– Oxidize cytosolic NADH and NADPH.
– Electrons from these external dehydrogenases enter the main electron transport chain at the level of the ubiquinone pool.level of the ubiquinone pool.

Unique Electron Transport E i Pl Mi h d iEnzymes in Plant Mitochondria
• Two pathways for oxidizing matrix NADHp y g
– Electron flow through Complex I → sensitive to inhibition by several compounds, including rotenone and piericidin.
– In addition, plant mitochondira have a rotenone-resistant dehydrogenase for oxidation of NADH derived from citric acid cycle substrate → thisderived from citric acid cycle substrate → this pathway may be a bypass that is engaged when complex I is overloaded, such as under photorespiratory conditionsphotorespiratory conditions.

Unique Electron Transport E i Pl Mi h d iEnzymes in Plant Mitochondria
• An NADPH dehydrogenase is present on the y g pmatrix surface; little is known about this enzyme.
• Most, if not all, plants have an “alternate” respiratory pathway for reduction of oxygenrespiratory pathway for reduction of oxygen.
– This pathway involves the so-called alternativeThis pathway involves the so called alternative oxidase; unlike cytochrome c oxidase, it is insensitive to inhibition by cyanide, azide, or carbon monoxide.carbon monoxide.

Transporters Exchange S b d P dSubstrates and Products
• Although ATP is synthesized in theAlthough ATP is synthesized in the mitochondrial matrix, most of it is used outside the mitochondrion.
– An efficient mechanism for moving ATP in and out of the organelle is neededand out of the organelle is needed.
• Adenylate transport involves the ADP/ATP ( d i l tid )ADP/ATP (adenine nucleotide) transporter, another inner-membrane transporter.p
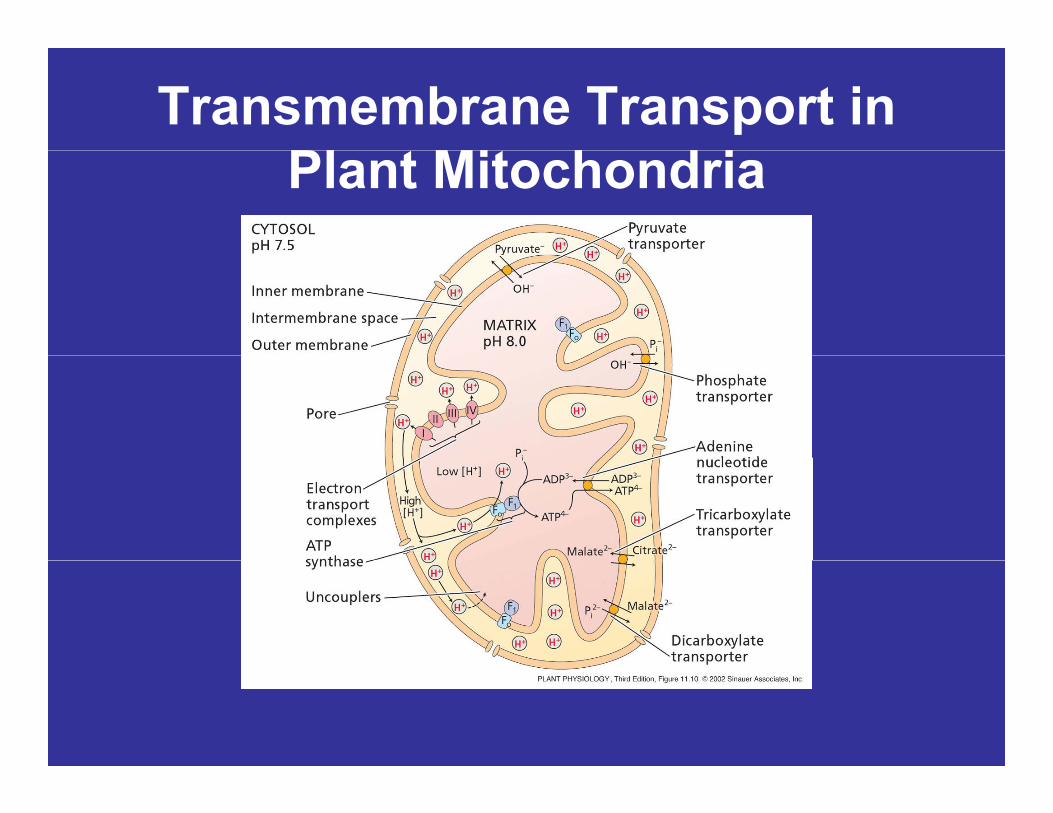
Transmembrane Transport in Pl Mi h d iPlant Mitochondria

Electron Transport ChainElectron Transport Chain

Chemiosmotic Theory• As electron pairs pass down the ETC, H+
ions are pumped across the innerions are pumped across the inner mitochondrial membrane from the matrix to the intermembrane space.to the intermembrane space.
• The electrochemical gradient of H+ ionsThe electrochemical gradient of H ions across the inner membrane constitutes the driving force for the ATP-ase (ATP g (synthase) catalyzed phosphorylation of ADP.

Chemiosmotic TheoryChemiosmotic Theory• This ‘driving force’ has been termedThis driving force has been termed
‘proton motive force’ and is composed of two elements:
– pH difference or gradient across the inner membrane of about 1 5 pH units (7 0 inmembrane of about 1.5 pH units (7.0 in intermembrane space and 8.5 in matrix).
A l t i l t ti l diff– An electrical potential difference or gradient of about 0.15 V (H+ ions on outer surface of inner membrane and OH- on the inner surface of the membrane).
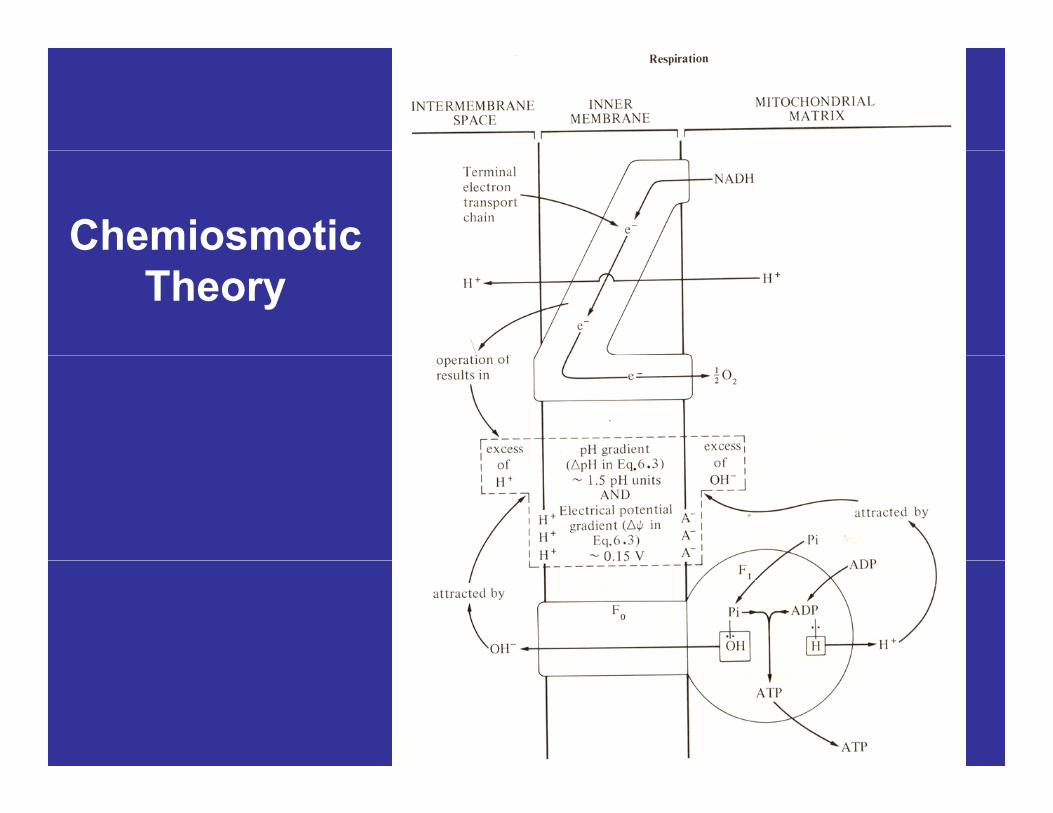
Chemiosmotic Theory

Five Complexes of Integral Membrane Proteins in InnerMembrane Proteins in Inner
Mitochondrial MembraneComplex NameComplex Name• *I NADH dehydrogenase• II Succinate dehydrogenaseII Succinate dehydrogenase • *III Cytochrome c reductase
(complex cytochrome bc )(complex cytochrome bc1)• *IV Cytochrome c oxidase• V ATP synthase• V ATP synthase
* Sit t hi h t d* Sites at which protons are pumped

Electron Pathways in PlantsElectron Pathways in Plants

ATP / Pi Transporter in Mitochondria
• The one-for-one exchange of mitochondrial ATP and cytosolic ADP across the inner membrane is driven by the membrane potentialby the membrane potential.
• Pi is returned to the matrix in exchange for OH-.Pi is returned to the matrix in exchange for OH .

Alternate Electron Pathways in Plants
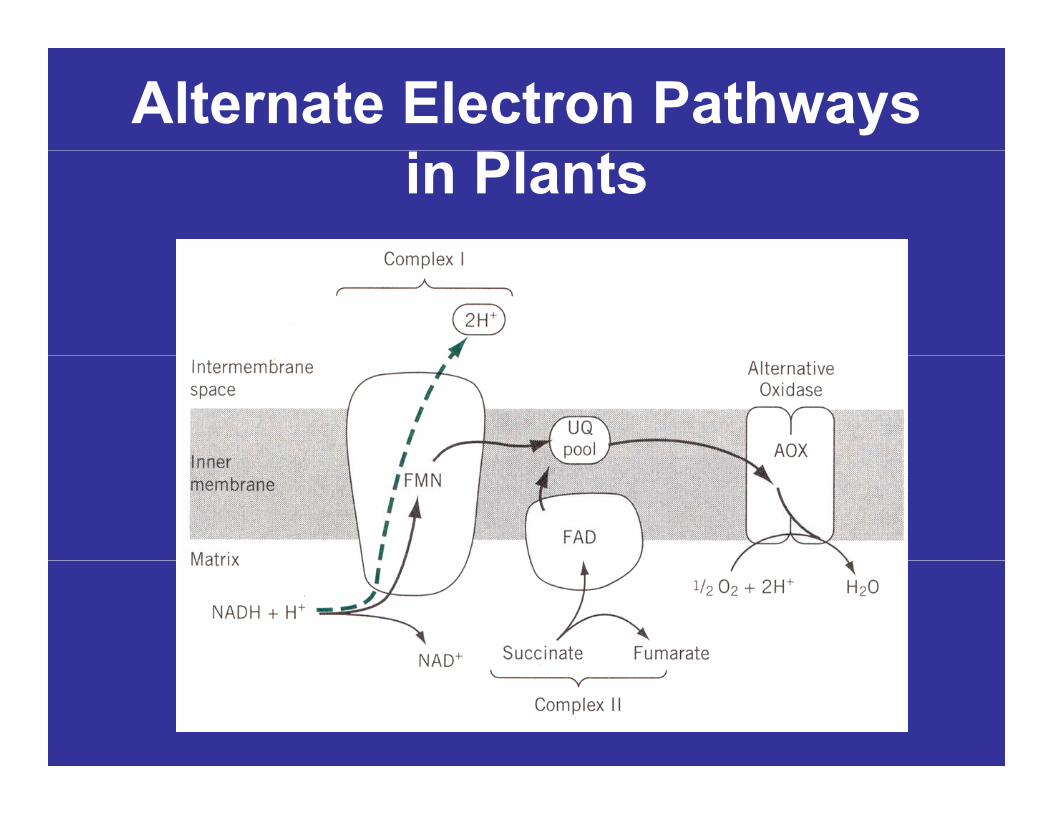
Alternate Electron Pathways in Plants

CYANIDE-RESISTANT RESPIRATIONRESPIRATION
• The way the plant controls the flow of electrons through the two electron transport chains is notthrough the two electron transport chains is not clear but there is evidence to indicate that the intensity of the electron flux through the cytochrome system of the conventional chain is acytochrome system of the conventional chain is a regulating factor.
It ti it i hi h t i ll i h i (– Its activity is highest in cells rich in sugars (as after rapid photosynthesis) when glycolysis and the Krebs cycle occur unusually rapidly, because then the normal electron-transport pathway cannot handle all the electrons provided to it.

CYANIDE-RESISTANTCYANIDE RESISTANT RESPIRATION
– Several experts have concluded that the alternative pathway operates largely as an overflow mechanism to remove
l t h th t helectrons when the cytochrome pathway becomes saturated by rapid glycolysis and Krebs-cycle activitiesglycolysis and Krebs-cycle activities.

CYANIDE-RESISTANT RESPIRATION
• Physiological significance?y g g
• The physiological significance of cyanide-resistant respiration is not clear.resistant respiration is not clear.
– Goodwin and Mercer (1981) indicate that it is doubtful that its function is to allow plants todoubtful that its function is to allow plants to survive in the presence of cyanide even though many plants release HCN from
i l id h th i j dcyanogenic glucosides when they are injured. Why? Because those plant tissues with the most active cyanide-resistant respiration have y pno cyanogenic glucosides.

CYANIDE-RESISTANT RESPIRATION
C id i t t i ti i th ht t b– Cyanide-resistant respiration is thought to be responsible for the climacteric in fruits (i.e., the marked increase in respiration during and p gjust prior to ripening). The climacteric is induced by ethylene and it is thought that ethylene brings this about by stimulatingethylene brings this about by stimulating cyanide-resistant respiration.
– Certain germinating seeds exhibit cyanide-resistant respiration during the early stages of water imbibitionwater imbibition.

CYANIDE-RESISTANT RESPIRATION– The best understood role of cyanide-resistant
respiration is, however, in generating heat in thermogenic tissues:thermogenic tissues:
• Thermogenicity is seen in the flowers of inflorescences of certain plants such as water plily, Victoria, and the arum lilies, Arummaculatum.
• In skunk cabbage (Symplocarpus foetidus) the• In skunk cabbage (Symplocarpus foetidus), the inflorescence is a spadix covered with tiny hermaphroditic flowers which produce heat and a foul smell. The heat is produced in these ou s e e eat s p oduced t eseinflorescences to volatize the odiferous compounds formed in them. The latter are frequently amines or indoles and serve to attract
lli ti i tpollinating insects.

CYANIDE-RESISTANT RESPIRATIONRESPIRATION
• Energy production of cyanide-insensitive respiration?
– Since the P/O ratio of cyanide-resistant respiration is 1 in contrast to 3 for conventional respiration, the cells have to oxidize three times as much fuel to get the required amount ofoxidize three times as much fuel to get the required amount of ATP.
– Assuming that oxidation is via NADH, this will produce aboutAssuming that oxidation is via NADH, this will produce about 4.5 times as much heat (i.e., energy that is not conserved as ATP) than normal.
Air temperature InflorescenceArum italicum 15 C 51 CSkunk cabbage 5 C 30 C

CYANIDE-RESISTANT RESPIRATIONRESPIRATION

Does Respiration Reduce Crop Yields?
• Plant respiration can consume an• Plant respiration can consume an appreciable amount of carbon fixed each day during photosynthesis overeach day during photosynthesis over and above losses due to photorespiration
• To what extent can changes in a plant’sTo what extent can changes in a plant s respiratory metabolism affect crop yields?

Does Respiration Reduce Crop Yields?
• Two components of respiration:Two components of respiration:
– Growth respiration - involves the processing of reduced carbon to bring about the growth of g gnew plant matter
– Maintenance respiration - component of respiration needed to keep existing, mature cells in a viable state
Utilization of energy by maintenance• Utilization of energy by maintenance respiration is not well understood
• Estimates indicate that it can representEstimates indicate that it can represent more than 50% of the total respiratory flux

Whole-Plant Respiration• Many factors affect the respiration rates of
a plant– Species
Growth habit of the plant– Growth habit of the plant
– Type and age of specific organ
– Environmental variables
• External oxygen concentration• Temperature• Plant water status

Whole-Plant Respiration• Even though plants generally have low
respiration rates, the contribution of respiration to the overall carbon economyrespiration to the overall carbon economy of the plant can be substantial
– Several herbaceous species surveyed indicated that 30-60% of the daily gain in photosynthetic carbon was lost to dark p yrespiration
• These values tended to decrease with ageg
• Young trees lose roughly a third of their daily photosynthate to respirationdaily photosynthate to respiration

Whole-Plant Respiration
• Loss can double in older trees as theLoss can double in older trees as the ratio of photosynthetic to nonphotosynthetic tissue decreases.
• In tropical areas, 70-80% of the daily p , yphotosynthetic gain can be lost to respiration owing to the high nighttime
i i i d i hrespiration rates associated with elevated night temperatures.

Whole-Plant Respiration
• Empirical relations between plant respiration rates and crop yieldrates and crop yield
– Yield increases of 10-20% were correlated with a 20% decrease in the leaf respiration rate of pthe forage crop perennial ryegrass (Lolium perenne) (Wilson and Jones, 1982)Similar correlations have been found for other– Similar correlations have been found for other plants, including corn and tall fescue (Lambers, 1985)
• Appears that a potential for increasing crop yields through reduction of respiration rates exists

Whole-Plant Respiration• What about the cyanide-resistant
alternative pathway?alternative pathway?
– Cyanide-resistant pathway has the t ti l f tili i id blpotential for utilizing considerable
amounts of the cell’s reduced carbon to no apparent useful endno apparent useful end
– Estimates of the alternative pathway in wheat roots alone suggest a loss ofwheat roots alone suggest a loss of carbon equivalent to 6% of the final grain yield