comparative studies of supraocular lepidosis in squamata (reptilia
TRANSCRIPT

1MULTEQUINA 16: 1-52, 2007
COMPARATIVE STUDIES OF SUPRAOCULARLEPIDOSIS IN SQUAMATA (REPTILIA) AND ITS
RELATIONSHIPS WITH AN EVOLUTIONARYTAXONOMY
ESTUDIOS COMPARATIVOS DE LA LEPIDOSIS SUPRA-OCULAR EN
SQUAMATA (REPTILIA) Y SU RELACIÓN CON LA TAXONOMÍA
EVOLUCIONARIA
JOSÉ M. CEI †
ISSN 0327-9375
RESUMEN
Observaciones morfológicasprevias sobre un gran número deespecies permiten establecer unacorrespondencia entre lapeculiaridad de los patronessistemáticos de las escamassupraoculares de Squamata y laposición evolutiva de cada taxónconsiderado en los cladogramaspropuestos por Estes et al. (1988).Aparte del significado biológicogeneral de estos hallazgos, inclusopara discutidas orientacionestaxonómicas, la lepidosissupraocular llega a refrendar unadecisión sistemática con suevidencia. Así, en Iguania, la familiaLeiosauridae, propuesta por Frostet al. (2001), aparece sostenidahasta en el detalle de su división en
las subfamilias Leiosaurinae yEnyaliinae. Siempre en IguaniaPleurodonta se evidencian ejemploscomo los inconfundibles patrones deescamas supraoculares deOpluridae, Leucocephalidae,Polychrotidae, Tropiduridae. A nivelespecífico la interdependencia enIguanidae de los géneros Iguana,Cercosaura, Brachylophus,Conolophus, puede llevar apostular pretéritos acontecimientospaleogeográficos. También ameritaénfasis la llamativa separación,según este criterio morfológico, en-tre Iguania y Scleroglossa, launiforme lepidosis de centenares deGekkota, o la excepcional fisonomíade Autarchoglossa, en sus ramas tanindividualizadas de Scincomorpha(Lacertoidea; Teiioidea;Scincoidea) o Anguimorpha.

2 José M. Cei
Palabras clave: diagnosis,escamas supraoculares, pa-trón específico lepidosis,Pleurodonta, Squamata,taxonomía
SUMMARY
Former observations oncharacteristic patterns of supraocularscutellation in different specific orsupraspecific taxa of Squamata areextended and analyzed in this generalcritical review. A relationshipbetween a defined morphologicalsupraocular lepidosis in a particulartaxon and its evolutionary positionin a classificatory system, such as thecladograms obtained by Estes et al.(1988), was tested and confirmed.Supraocular scales of Squamata arenot morphological elements atrandom, but can be assumed asmorphological scale characters,useful in systematic comparativeexaminations and able to strengthensome discussed new systematicassessments, such as the family statusof Leiosauridae and its subfamiliesin Pleurodont Iguanians, proposedby Frost et al. (2001), as well as inseveral other cases.
Key words: Squamata,supraocular scutellation,specific lepidosis pattern,diagnosis, Pleurodonta,taxonomy
INTRODUCTION
The selection of significant somaticmorphological characters (shape,size, structure) will play an essentialrole in providing a systematicherpetological arrangement, at leastto species level: furthermore it canapply also to more elevated ranksof the herpetological classification.We are pointing out here someinteresting aspects of the so-calledlepidosis, or cutaneous scalecovering of Reptilia, mainly inLacertilian lizards. More exactly,the differentiated scales bilaterallylocated in the supraocular region ofthe head, bounded inside by theorbital semicircles, and by theciliary eye border outside (Figure1). Given the noticeable position ofthese supraocular scales in the headscutellation of Squamata, theirpresence has generally beenreported in many diagnosticdescriptions of species.Nevertheless, no true emphasis wasever placed on their peculiartaxonomic significance: either asan important generic characteristicstructure of the dorsal headlepidosis, or as distinctivesupraocular patterns in separatedsystematic groups, from differentgenera to families and highercategories. A preliminary paper wasprovided on this matter (Cei, 2003),dealing with supraocular scales inspecies and genera of Iguania, as

3MULTEQUINA 16: 1-52, 2007
Figure 1. Sketched fundamental morphology of supraocular scutellation in Squamata, Pleurodonta: 1.
supraorbital semicircles, 2. supraocular scales, 3. intermediate supraorbital scales and 4.
superimposed superciliaries
Figura 1. Esquema fundamental de la morfología de la escutelación supraocular en Squamata, Pleurodonta:
1. semicírculos supraorbitales, 2. escalas supraoculares, 3. escalas supraorbitales interme-
dias y 4. superciliares superimpuestos
well as in some far-away taxonomiccategories, such as ScleroglossaTeiid lizards. Thus, considering theremarkable number of past andrecent species descriptions, adetailed review and discussion ofsuch a noticeable but stilldisregarded anatomical peculiaritycould be a very interesting job,postulating a distribution patternnot at random but in agreementwith the phylogenetic subdivisionsof well-known modernclassificatory essays.
Some isolated reports aboutspecific supraocular scutellation in
Reptiles, besides a preliminarypaper (Cei, 2003), deal withcircumorbitals in one row or in tworows between supraoculars andmedian head shields, as emphasizedin Frost‘s (1992) taxonomicrevision of Tropidurus groups oflizards that points outdiscrimination among the generaUranoscodon, Microlophus,Plesiomicrolophus and Tropidurus.Likely in Etheridge‘s (1970)comparative research thesystematic differences insupraciliaries were suitably appliedto discriminate Plica andUranoscodon from other members
3
2 1
4

4 José M. Cei
of the Tropidurus groups. However,apart from these interesting butlimited citations, a whole zoologicaldiscussion and critical valuation oflacertilian supraocular scutellationis yet wanting to our knowledge.The present work will attempt toreach better and conclusiveinformation on such a suggestiveand little analyzed subject.
MATERIAL AND METHOD
Given the aim of this work, itsoutcome is a detailed representationof the postulated relationshipsbetween the supraocularscutellation patterns and thetaxonomic categories of Squamata.Our comparative graphicdocumentation, exposed in Figures,was mainly obtained from carefulobservations and drawings ofpreserved specimens, or in somecase from careful adaptations ofcontrolled reproductions by reliableauthors. The available materialsabove all have been the specimensof a personal collection (JMC-DC:José M. Cei Diagnostic Collection),occasionally complemented by paststudies of species from theHerpetological Collections of someMuseums or Scientific Institutionswhose list and acronyms arefollowing. IBA-UNC: Collectionsof the Instituto de Biología Animal,Universidad Nacional de Cuyo,Mendoza, Argentina; MZUSP:
Museo de Zoologia, Universidadede São Paulo, Brasil; MCZ:Museum of Comparative Zoology,Harvard University, Cambridge,USA; NMNH: National Museumof Natural History, SmithsonianInstitution, Washington, USA;FMNH: Field Museum of NaturalHistory, Chicago, USA; MNHNParis: Museum Nationale HistoireNaturelle, Paris; UNNE:Universidad Nacional del Nordeste,Corrientes, Argentina.
Given the Figures including thetotal samples drawn that support ourpresent report and discussion, aseparated list of the examinedspecimens would be prolix andunnecessary. Moreover the generallocalities of samples wereopportunely indicated in the Figures.When it was necessary,morphological details of the lizardswere drawn under a dissectingmicroscope. Peter‘s Dictionary(1964) was a reference formorphological and anatomicalnomenclature, as well as the severalEtheridge‘s papers, i.e. hisCtenoblepharis adspersaredescription (1995). The drawingsof the Figures have been all more orless slightly magnified: we believethat individual magnification ofsymbols in every identified drawingscale is useless to our purposes.Besides the specimens documentedin the Figures, hundreds of specimens

5MULTEQUINA 16: 1-52, 2007
corresponding to the patterns havebeen observed and compared, forimproving the truthfulness andsignificance of our samples.
Among the most importantliterature supporting the harvesting,arranging and discussion of ourrepresentative samples, we wish tohighlight the useful, good oroutstanding papers or reviews byAnderson (1999), Angel (1942),Blarc (1977), Brygoo (1971, 1978),Dixon (1973), Donnelly and Myers(1991), Greer (1970), Grismer etal. (1994), Haas and Werner (1969),Halliday and Adler (2002),Hoogmoed (1973), Kluge (1974),Leviton et al. (1992), Mertens(1958, 1959), Pregill (1992), Ruibal(1964), Savage (1963), Tayer(1956), Van Demburg (1978),Vanzolini et al. (1965, 1980) andWiens (1993).
RESULTS
Analytical review of comparativereports on supraocular lepidosis inSquamata
As pointed out in theIntroduction, the aim of thisresearch was to put in evidence acorrelative relationship between aclassificatory system and asignificant set of data onsupraocular scutellation inSquamata reptiles. The
phylogenetic indented taxonomyexhibited in the valuable conclusivestatements by Estes et al. (1988)was considered as a suitable model(Figure 2), together with thereassessment of Iguania by Frost etal. (2001). Such taxonomy has beenfollowed here for comparison withour detailed morphologicalobservations, far away from anyaprioristic personal opinion. Then,without disregarding otherinteresting systematic contributions,as the recent paper by Schulte et al.(2003), in full agreement with ourfindings, we recognize the taxonPleurodonta and its includedfamilies as in the mentioned Frostet al.’s (2001) arrangement. On thecontrary, the opposite general taxonIguanidae (sensu Boulenger, 1895)postulated in the paper by Schulteet al., based on bio-molecular andmorphological researchs, is fullydiscordant with our results.
In the following 27 Figures,specific supraocular scutellationpatterns belonging to genera andfamilies of the major taxa listed inthe indented taxonomy are reported.Evidently it was impossible tocheck the thousands of livingspecies of lizards, but significantlyrepresentative drawings of enoughgeneric and specific taxa, areprovided.

6 José M. Cei
IguaniaAgamidae
ACRODONTA Chamaeleonidae
PLEURODONTA CorytophanidaeCrotaphytidaeHoplocercidaeIguanidaeLeiocephalidaeLeiosauridae Leiosaurinae
EnyaliinaeLiolaemidaeOpluridaePhrynosomatidaePolychrotidaeTropiduridae
ScleroglossaGEKKOTA Gekkonidae
EublepharidaeDiplodactylidaePygopodidae
AUTARCHOGLOSSA
ScincomorphaLacertoidea
XantusiidaeLacertiformes Lacertidae
Teiioidea Teiidae Gymnophthalmidae
ScincoideaScincidaeCordylidae
AnguimorphaAnguidaeXenosauridae Varanoidea
Helodermatidae Lanthanotidae Varanidae
Figure 2. Squamata: phylogenetic indented taxonomy according to Estes et al. (1988). The taxon Iguania
was adapted in accordance with Frost et al. (2001)
Figura 2. Squamata: taxonomía filogenética indentada según Estes et al. (1988). El taxón Iguania fue
adoptado de acuerdo con Frost et al. (2001)

7MULTEQUINA 16: 1-52, 2007
Two opposed major systematiccategories stand out in theSquamata indented taxonomy:Iguania and Scleroglossa, sustainedby anatomical, mostly osteologicalcharacters. Acrodonta andPleurodonta are a subdivision ofIguania, likewise countersigned byosteological differences. We cananticipate that a fairly simplesupraocular scutellation belongs toAcrodonta, but a remarkablevariety of differentiatedsupraocular scales is shown by thePleurodonta families.
Agamidae and Chamaeleonidaeare two large and widespreadAcrodont families. With more than50 genera and 400 species,Agamidae are found in the African,Euro-Asiatic and Australiancontinents, with an extraordinarymorpho-ecological diversification.But a relatively uniform coveringof rudely roundish or enlargedscales appears in their supraocularregion, the circumorbitalsemicircles being scarcely evidentand the supraciliaries moderatelyprominent or sharp-pointed, withsome important exceptions like thesmart or spiny supraciliaries ofLyriocephalus, Phrynocephalusand Draco. In Figure 3 severalexamples of these lizards are given,significant enough for the multiplespecies of the Acrodonta generictaxa. Representativeness of these
patterns has been strengthened bytheir direct control in many otheragamid specimens, or by othercareful documentary reports byreliable authors. Agamid patternsof Figure 3 have been comparedand confirmed in other 15 Africanand Indian taxa of the genus Agama,in 15 Asiatic taxa of Draco, in 4Indian taxa of Calotes whoseirregular supraocular scales appearnoticeably bulky, in 70 Australiantaxa of the genera Goniocephalus,Amphibolurus, Tympanocryptis,Chelosania, Dipariphera,Physignathus and Lophognathus.
Chamaeleonidae, a specialized,mostly arboreal family, with morethan 130 species but only 6 generaare distributed in Africa,Madagascar, Arabia, India and SriLanka. The protruding eyes ofChameleons, covered with scalylids, are unique: in Figure 4, thesupraoculars of Chamaeleo andBrookesia from Madagascar areemphasized. A fundamentalanalogy between the Chamaeleopattern and the agamid supraocularscutellation is evident from itsgeneral lining of irregular scales,distinct supraciliary border andalmost indistinguishable innercircumorbital semicircles. Theexamples of Brookesia taxa inFigure 4 point out the peculiarfeature of dermal cranialossifications in these advanced

8 José M. Cei
Figure 3. AGAMIDAE. A. Agama planiceps (Angola SW Africa); B. Laudakia rupta (Iran); C. Trapelus
pallidus (Arabia); D. Uromastyx asmussi (Irán); E. Phrynocephalus maculatus (Arabia Saudita);
F. Draco maculatus (India); G. Japalura tricarinata (Nepal - India); H. Diporiphera reginae;
I. Amphibolurus vadnappa (S Australia)
Figura 3. AGAMIDAE. A. Agama planiceps (Angola SW Africa); B. Laudakia rupta (Iran); C. Trapelus
pallidus (Arabia); D. Uromastyx asmussi (Irán); E. Phrynocephalus maculatus (Arabia Saudita);
F. Draco maculatus (India); G. Japalura tricarinata (Nepal - India); H. Diporiphera reginae;
I. Amphibolurus vadnappa (S Australia)
A B C
D E F
G H I

9MULTEQUINA 16: 1-52, 2007
Figure 4. CHAMAELEONIDAE. A. Chamaeleo bifidus (E Madagascar); B. Chamaeleo cephalolepis (I.
Comore-Madagascar); C. Ch. lateralis (Madagascar); D. Chamaeleo cucullatus (NE Madagascar);
E. Brookesia decaryi (Madagascar); F. Brookesia thieli (Madagascar); G. Brookesia ebenaui
(Madagascar); H. B. perarmatz (Madagascar); I. Brookesia stumpffi (Madagascar)
Figura 4. CHAMAELEONIDAE. A. Chamaeleo bifidus (E Madagascar); B. Chamaeleo cephalolepis (I. Comore-
Madagascar); C. Ch. lateralis (Madagascar); D. Chamaeleo cucullatus (NE Madagascar); E.
Brookesia decaryi (Madagascar); F. Brookesia thieli (Madagascar); G. Brookesia ebenaui
(Madagascar); H. B. perarmatz (Madagascar); I. Brookesia stumpffi (Madagascar)
A B C
D E F
G H I

10 José M. Cei
chamaeleontids, leading to astriking masking of theirsupraocular scale coating, hereillustrated for several species ofuncommon lizards from theisolated Madagascar.
The partition of the Pleurodontaevolutionary stock is remarkable.Also remarkable are our findingsabout a corresponding associationbetween different supraocularscale patterns and the differentfamilies listed in the indentedtaxonomy exposed in Figure 2.These families have beendiscussed in a serial issue ofherpetological contributions: fromthe paper by Frost and Etheridge(1989) to the successive works byFrost (1992) and Frost et al. (2001).Without any intentional attemptof phylogenetic references, everyfamily shall be analyzed now withregard to its supraocular lepidosis.
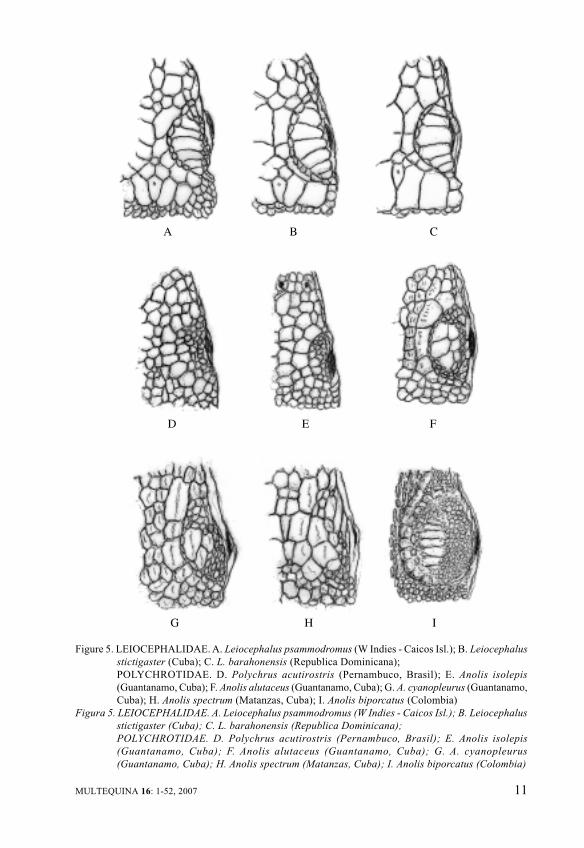
In Figure 5, upper row,Leiocephalidae is only limited tothe genus Leiocephalus from WestIndies. Formerly related toTropiduridae, its family status wasestablished by Frost et al. (2001).In its different insular taxa, the welldifferentiated supraocular lepidosisprevailing in Pleurodonta ismaintained, with minor specificdifferences. A longitudinal centralrow of large supraocular scalestretches is evident between anarrow row of supraciliaries and
moderate, regular circumorbitalsemicircles inside. In the sameFigure 5, midmost and inferiorrows, representative of the familyPolychrotidae, are shown:Polychrus with few neotropicalspecies and Anolis with more than230 species, spread with severalcharacteristic groups from southeastern United States to WestIndies, northern Mexico,southwards through Central andSouth America to about 20ºSlatitude. A true “Polychrotidae”pattern (sensu Frost et al., 2001) isrecognizable in the Figure 5,bearing impressive largecircumorbital semicircles and fewvery variable internal, small shields,surrounded by many minute scales,and very diminished supraciliariesoutside. The foreseeable specificvariation in the multiple Anolis isemphasized in Figure 5 by sometaxa from Cuba and Colombia.
Two families with few generaand species are registered in theupper and inferior rows of Figure6. In Opluridae from Madagascarand Comore islands, the monotypicgenus Chalarodon and somespecies of Oplurus keep a rathersimple supraocular patternreminiscent of Acrodontscutellations. Both very similarexamples of Oplurus in this Figureexhibit irregular supraocular scalesdecreasing from their almost
11MULTEQUINA 16: 1-52, 2007
A B C
D E F
G H I
Figure 5. LEIOCEPHALIDAE. A. Leiocephalus psammodromus (W Indies - Caicos Isl.); B. Leiocephalus
stictigaster (Cuba); C. L. barahonensis (Republica Dominicana);
POLYCHROTIDAE. D. Polychrus acutirostris (Pernambuco, Brasil); E. Anolis isolepis
(Guantanamo, Cuba); F. Anolis alutaceus (Guantanamo, Cuba); G. A. cyanopleurus (Guantanamo,
Cuba); H. Anolis spectrum (Matanzas, Cuba); I. Anolis biporcatus (Colombia)
Figura 5. LEIOCEPHALIDAE. A. Leiocephalus psammodromus (W Indies - Caicos Isl.); B. Leiocephalus
stictigaster (Cuba); C. L. barahonensis (Republica Dominicana);
POLYCHROTIDAE. D. Polychrus acutirostris (Pernambuco, Brasil); E. Anolis isolepis
(Guantanamo, Cuba); F. Anolis alutaceus (Guantanamo, Cuba); G. A. cyanopleurus
(Guantanamo, Cuba); H. Anolis spectrum (Matanzas, Cuba); I. Anolis biporcatus (Colombia)

12 José M. Cei
Figure 6. OPLURIDAE. A. Oplurus cyclurus (Madagascar); B. Oplurus fierinensis (Madagascar); C.
Chalarodon madagascariensis (Madagascar).
PHRYNOSOMATIDAE. D. Prhynosoma modestum (New Mexico, USA); E. Prhynosoma
douglassi (Montana, USA); F. Prhynosoma cornutum (Texas, USA).
CORYTOPHANIDAE. G. Corytophanes cristatus (Costa Rica); H. Basiliscus plumifrons (Costa
Rica); I. Laemanctus serratus (Mexico)
Figura 6. OPLURIDAE. A. Oplurus cyclurus (Madagascar); B. Oplurus fierinensis (Madagascar); C.
Chalarodon madagascariensis (Madagascar).
PHRYNOSOMATIDAE. D. Prhynosoma modestum (New Mexico, USA); E. Prhynosoma
douglassi (Montana, USA); F. Prhynosoma cornutum (Texas, USA).
CORYTOPHANIDAE. G. Corytophanes cristatus (Costa Rica); H. Basiliscus plumifrons (Cos-
ta Rica); I. Laemanctus serratus (Mexico)
A B C
D E F
G H I

13MULTEQUINA 16: 1-52, 2007
indistinct circumorbital semicirclesto their equally indistinguishablesupraciliaries. In Chalarodonmadagascariensis a quite uniformsupraocular layer of small roundedscales is better delimited by narrowcircumorbital semicircles and arecognizable border of very smallsupraciliaries. Corytopharidae fromMexico and Central Americalikewise include only three generawith few species each. Irregularsupraocular scales are broader inCorytophanes and Laemanctus,small in Brasiliseus. Circumorbitalsemicircles and supraciliaries,sharp-pointed in Corytophanes,stand out in this family. Striking incentral row of Figure 6 is theundifferentiated supraocularsurface of the horny skull roof ofsome Phrynosoma, odd genus ofthe family Phrynosomatidae,exposed in the following Figures.Several genera, found from southwestern North America southwardto Panama, were assembled inPhrynosomatidae.
In Figure 7 the mentioned“Pleurodont pattern” isrecognizable for the about fifty taxaof the spiny Sceloporus,distinguished by their expandedcircumorbital semicircles andprominent superciliaries,surrounding 5-6 broad supraocularscales. A somewhat divergingmorphology can be observed, as in
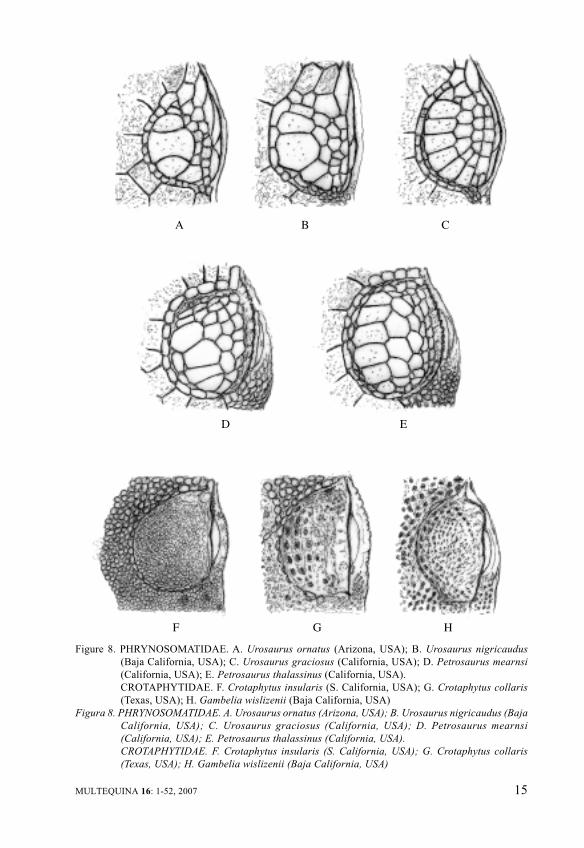
Sceloporus orcutti and S. olivaceus.Interesting to remark is the findingof these kinds of supraocularscutellation here reported for othergenera of such heterogeneousfamily, like Uma, Uta, Holbrookia,with analogous geographicdistribution. Likewise related butpeculiar supraocular lepidosis maybe pointed out for the arborealUrosaurus (Figure 8), so carefullyreviewed by Wiens (1993) for thesame arid environment. Theirvariable six broad longitudinalsupraocular scales, the otheradherent rows of minor scales aswell as the shape and size ofcircumorbital semicircles andsupraciliaries, support anunmistakable generic identity.Similar valuation can be made forthe related Petrosaurus speciesinhabiting mainly the sameCalifornian territories. Even inFigure 8, inferior row, thesupraocular lepidosis inCrotaphytidae can be observed.Collared lizards and Leopardlizards (Crotaphytus andGambelia) also from the subdesertrocky or sandy habitats ofsouthwestern North America areunmistakable for their nearlysmooth supraocular covering byvery minute circumorbitalsemicircles and sprinkled by someinsignificant scales.

14 José M. Cei
Figure 7. PHRYNOSOMATIDAE. A. Sceloporus merriami (Texas, USA); B. Sceloporus poinsetti (Texas,
USA); C. Sceloporus clarki (Arizona, USA); D. S. magíster (S California, USA); E. S. orcutti
(S. California, USA); F. Sceloporus olivaceus (Texas, USA); G. Uma notata (S. California,
USA); H. Uta stamburiana (S. California, USA); I. Holbrookia texana (Texas, USA)
Figura 7. PHRYNOSOMATIDAE. A. Sceloporus merriami (Texas, USA); B. Sceloporus poinsetti (Texas,
USA); C. Sceloporus clarki (Arizona, USA); D. S. magíster (S California, USA); E. S. orcutti
(S. California, USA); F. Sceloporus olivaceus (Texas, USA); G. Uma notata (S. California,
USA); H. Uta stamburiana (S. California, USA); I. Holbrookia texana (Texas, USA)
A B C
D E F
G H I
15MULTEQUINA 16: 1-52, 2007
Figure 8. PHRYNOSOMATIDAE. A. Urosaurus ornatus (Arizona, USA); B. Urosaurus nigricaudus
(Baja California, USA); C. Urosaurus graciosus (California, USA); D. Petrosaurus mearnsi
(California, USA); E. Petrosaurus thalassinus (California, USA).
CROTAPHYTIDAE. F. Crotaphytus insularis (S. California, USA); G. Crotaphytus collaris
(Texas, USA); H. Gambelia wislizenii (Baja California, USA)
Figura 8. PHRYNOSOMATIDAE. A. Urosaurus ornatus (Arizona, USA); B. Urosaurus nigricaudus (Baja
California, USA); C. Urosaurus graciosus (California, USA); D. Petrosaurus mearnsi
(California, USA); E. Petrosaurus thalassinus (California, USA).
CROTAPHYTIDAE. F. Crotaphytus insularis (S. California, USA); G. Crotaphytus collaris
(Texas, USA); H. Gambelia wislizenii (Baja California, USA)
A B C
D E
F G H

16 José M. Cei
Two families of Pleurodontawith a common general supraocularlepidosis are documented in Figure9. Belonging to the neotropicalHoplocercidae, with a dozenspecies, the three queer generaHoplocercus, Morunasaurus andEnyalioides show always verydistinct circumorbital semicirclesand supraciliaries, sharp pointed inEnyahioides. Supraocular scales areundifferentiated, but with threecentral quadrangular rows inMorunasaurus. Similar lepidosis,though with visible tendency topolymorphism is shown by theinteresting taxon Iguanidae (sensuFrost et al., 2001), whose generahold no more than two species each.
Minor differences among theSouth American Iguana,Cercosaura and Dipsosaurusdepend upon the shape and size ofsupraocular scales, whose more orless regular rows are encircled byevident supraciliaries, and behindby circumorbital semicircles, veryenlarged in Iguana. They show thedifferences between the almostundifferentiated probably primitivepatterns of the insular PacificBrachylophus and Conolophus, andthe more differentiated lepidosis inIguana iguana from neotropicalforests. Evolutionary factors ofprimeval distribution in geologicaltimes are likely related with theactual conservative conditions of
the isolated insular taxa as thePolynesian lizards (Brachylophusfasciatus, B. vitiensis) or thepeculiar Conolophus from theGalapagos Archipelago.
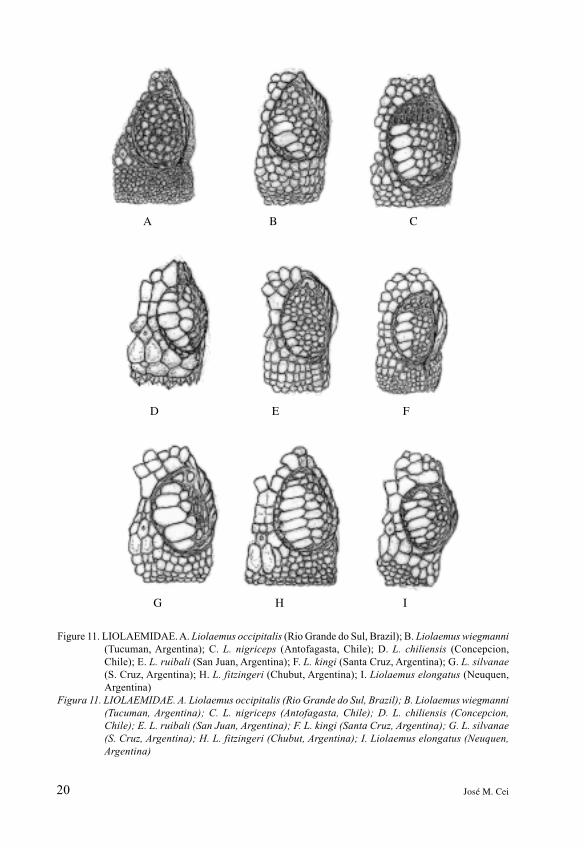
The important familyLiolaemidae, exemplified inFigures 10-11, was also establishedin the cited assessment by Frost etal. (2001): it comprises the generaCtenoplepharis, Phymaturus andLiolaemus from southern SouthAmerica. As stated by Etheridge(1995) in his redescription ofCtenoblepharis adspersa fromdesert landscapes of Peru, theunresolved relationships of suchhighly derived arenicolous reptileswith Phymaturus and Liolaemusare emphasized as differentialcharacters by their small, subequalsupraoculars. That is congruentwith our drawing in Figure 10.Other drawings illustrate severalspecies of the rock-climbing genusPhymaturus, all similar enough forthe scarcely differentiatedsupraocular scutellation.Notwithstanding, the species of thepatagonicus group (those drawnhere are: indistinctus, patagonicus,somuncurensis, payuniae) havemore enlarged supraorbital scalesalong the circumorbital semicircles.Liolaemus assembles about 160taxa, mainly Andean and Sub-Andean in many cases likely ofPleistocene or post-glacial

17MULTEQUINA 16: 1-52, 2007
Figure 9. HOPLOCERCIDAE. A. Hoplocercus spinosus (Brazil); B. Morunasaurus annularis (Rio San-
tiago, Ecuador): C. Enyalioides heterolepis (Ecuador).
IGUANIDAE. D. Iguana iguana (Brazil); E. Cercosaura similis (Central America); F.
Dipsosaurus dorsalis (South California, USA); G. Brachylophus fasciatus (Orchid Isl.,
Polynesia); H. B. vitiensis (Orchid Isl., Polynesia); I. Conolophus subvristatus (Galapagos Isl.)
Figura 9. HOPLOCERCIDAE. A. Hoplocercus spinosus (Brazil); B. Morunasaurus annularis (Rio Santia-
go, Ecuador): C. Enyalioides heterolepis (Ecuador).
IGUANIDAE. D. Iguana iguana (Brazil); E. Cercosaura similis (Central America); F. Dipsosaurus
dorsalis (South California, USA); G. Brachylophus fasciatus (Orchid Isl., Polynesia); H. B. vitiensis
(Orchid Isl., Polynesia); I. Conolophus subvristatus (Galapagos Isl.)
A B C
D E F
G H I

18 José M. Cei
Figure 10. LIOLAEMIDAE. A. Ctenoblepharis adspersa (Peru); B. Phymaturus indistinctus (Chubut,
Argentina); C. Phymaturus patagonicus (Chubut, Argentina); D. Ph. somuncurensis (Rio Ne-
gro, Argentina); E. Ph. payuniae (Payunia, Argentina); F. Ph. punae (San Juan, Argentina); G.
Ph. mallimaccii (La Rioja, Argentina); H. Ph. verdugo (South Mendoza, Argentina); I. Ph.
antofagastensis (Catamarca, Argentina)
Figura 10. LIOLAEMIDAE. A. Ctenoblepharis adspersa (Peru); B. Phymaturus indistinctus (Chubut,
Argentina); C. Phymaturus patagonicus (Chubut, Argentina); D. Ph. somuncurensis (Rio Ne-
gro, Argentina); E. Ph. payuniae (Payunia, Argentina); F. Ph. punae (San Juan, Argentina);
G. Ph. mallimaccii (La Rioja, Argentina); H. Ph. verdugo (South Mendoza, Argentina); I. Ph.
antofagastensis (Catamarca, Argentina)
A B C
D E F
G H I

19MULTEQUINA 16: 1-52, 2007
speciation. Its more characterizingfeature is, alongside the veryevident circumorbital semicircles,the longitudinal row of 5-6 broadsupraocular scales, not as irregularand confused as in the Phymaturuspatagonicus group (Figure 11). Infew eastern Brazilian arenicoloustaxa, as Liolaemus occipitalis, adiverging pattern ofundifferentiated scales, perhapsprimitive, may be pointed out.
Before moving to the complexfamily Tropiduridae of which sometaxa as Liolaemidae andLeiocephalidae have already beenreferred to (Frost and Etheridge,1989), we must pay attention to thePleurodont lizards Leiosauridae, aswell included in Polychrotidae priorto the remembered reclassificationby Frost et al. (2001).
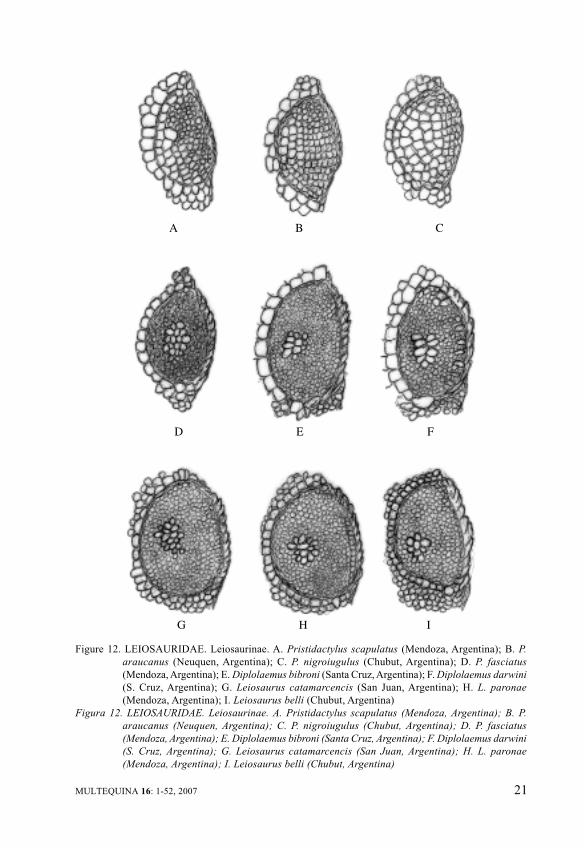
Parted into two subfamilies,Leiosaurinae and Enyaliinae, anumber of genera and a variety ofpatterns pertain to this unusualiguanian systematic entity, to whichFigures 12 and 13 are dedicated. InLeiosauridae, three peculiar generaare known, Pristidactylus,Diplolaemus and Leiosaurus, ofvery ancient lineage: Leiosaurus isfound fossil in Pliocene layers ofArgentina, Pristidactylus downright in the Miocene sediments ofChubut (Argentina).
Pristidactylus, the purpose ofour recent review (Cei et al., 2004),as well as Diplolaemus (Cei et al.,2003), are the most undifferentiatedmembers of the subfamily. All therecognized species ofPristidactylus were analyzed indetail in our revision, adding threeanalogous forms to the fourArgentine entities reported inFigure 12. Pristidactylusnigroiugulus from Chubut exhibitsan acrodont-like pattern, somewhatmodified in P. scapulatus and P.araucanus, but equal in P.achalensis, P. casuhatiensis fromArgentina and P. alvaroi fromChile. However in P. fasciatus,from the arid scrub of centralArgentina, a lepidosis very similarto those of the other two genera isfound, evoking probablyevolutionary genetic relationships.In Figure 12 a unique, justmentioned, iguanian supraocularlepidosis is shown, both inDiplolaemus and Leiosaurus, witha central rounded bouquet of largerscales in a very homogeneous fieldof diminished scales. In neotropicalEnyaliinae the here commentedextant conditions of morphologicalsupraocular uniformity appearaltered enough. In BrazilianEnyalius the homogeneous patternof E. perditus is replaced by acondition almost repeating that ofPristidactylus fasciatus or
20 José M. Cei
Figure 11. LIOLAEMIDAE. A. Liolaemus occipitalis (Rio Grande do Sul, Brazil); B. Liolaemus wiegmanni
(Tucuman, Argentina); C. L. nigriceps (Antofagasta, Chile); D. L. chiliensis (Concepcion,
Chile); E. L. ruibali (San Juan, Argentina); F. L. kingi (Santa Cruz, Argentina); G. L. silvanae
(S. Cruz, Argentina); H. L. fitzingeri (Chubut, Argentina); I. Liolaemus elongatus (Neuquen,
Argentina)
Figura 11. LIOLAEMIDAE. A. Liolaemus occipitalis (Rio Grande do Sul, Brazil); B. Liolaemus wiegmanni
(Tucuman, Argentina); C. L. nigriceps (Antofagasta, Chile); D. L. chiliensis (Concepcion,
Chile); E. L. ruibali (San Juan, Argentina); F. L. kingi (Santa Cruz, Argentina); G. L. silvanae
(S. Cruz, Argentina); H. L. fitzingeri (Chubut, Argentina); I. Liolaemus elongatus (Neuquen,
Argentina)
A B C
D E F
G H I
21MULTEQUINA 16: 1-52, 2007
Figure 12. LEIOSAURIDAE. Leiosaurinae. A. Pristidactylus scapulatus (Mendoza, Argentina); B. P.
araucanus (Neuquen, Argentina); C. P. nigroiugulus (Chubut, Argentina); D. P. fasciatus
(Mendoza, Argentina); E. Diplolaemus bibroni (Santa Cruz, Argentina); F. Diplolaemus darwini
(S. Cruz, Argentina); G. Leiosaurus catamarcencis (San Juan, Argentina); H. L. paronae
(Mendoza, Argentina); I. Leiosaurus belli (Chubut, Argentina)
Figura 12. LEIOSAURIDAE. Leiosaurinae. A. Pristidactylus scapulatus (Mendoza, Argentina); B. P.
araucanus (Neuquen, Argentina); C. P. nigroiugulus (Chubut, Argentina); D. P. fasciatus
(Mendoza, Argentina); E. Diplolaemus bibroni (Santa Cruz, Argentina); F. Diplolaemus darwini
(S. Cruz, Argentina); G. Leiosaurus catamarcencis (San Juan, Argentina); H. L. paronae
(Mendoza, Argentina); I. Leiosaurus belli (Chubut, Argentina)
A B C
D E F
G H I

22 José M. Cei
Diplolaemus (Figure 13), as in E.brasiliensis, E. iheringi andlikewise in Urostrophus. Moreintriguing is the genus Anisolepis,whose specific patterns can lookalike to that of Urostrophus vautieri(i.e. Anisolepis grillii), superficiallyto that of the CorythophanidBasiliscus plumifrons (i.e.Anisolepis undulatus), andstrikingly alike to those of thegenera Urosaurus and Petrosaurusof the family Phrynosomatidae (i.e.Anisolepis longicauda).
Remodeled as subfamily byFrost (1992), Tropiduridae wasconfirmed as a family by Frost etal. (2001) without making clear thedefinitive position (subfamily ortribe) of the diversified Stenocercinitribe. The supraocular lepidosis inTropidurus appears to be the mostsignificant pattern of IguanianPleurodonta (Figures 14, 15, 16).Two strongly diverging types aredistinguishable. The type of thegenus Uranoscodon from tropicalSouth America is present too in theStenocercini tribe, withStenocercus guentheri fromEcuador, Ophryessoides caducusfrom Northern Argentina,Proctotetrus pectinatus and P.doellojuradoi from centralArgentina, the last three lizards
considered as Stenocercus in thecited Frost’s paper (1992). Itsfundamental feature is a completesupraocular lining of imbricatekeeled scales, together with evidentkeeled circumorbital semicircles,and variable supraciliaries, showyand sharp-pointed in Uranoscodon,superimposed in Stenocercini taxa.The other prevailing Tropidurustype shows in the similar generaMicrolophus, Plesiomicrolophusand Tropidurus a general unfoldingof longitudinal rows of 4-6 broadsupraocular scales, of different sizeand flanked by minor, rounded,irregular scales. A remarkablevariation is even observed in theevident circumorbital semicircleswhich are more moderate inStenocercini taxa. Alwaysparticularly distinct in our drawingsis the interparietal scale and its“eye”, more or less developed (crf.Frost et al., 2001: p.21). Theunifying tendency of Frost`sassessment (1992) led to extendthe generic Tropidurus name tosome taxa known as Strobilurusand Plica, now Tropidurusstrobilurus, T. plica and T. umbra.A third Plica, now Tropiduruslumaria from Venezuelan Tepui,was well documented by Donnellyand Myers (1991).

23MULTEQUINA 16: 1-52, 2007
Figure 13. LEIOSAURIDAE. Enyaliinae. A. Enyalius brasiliensis (Espirito Santo, Brazil); B. Enyalius
iheringi (Sao Paulo, Brazil); C. E. perditus (Brazil); D. Urostrophus vautieri (Sao Paulo, Brazil);
E. Urostrophus iheringi (S. Paulo, Brazil); F. U. gallardoi (Cordoba, Argentina); G. Anisolepis
grillii (S. Paulo, Brazil); H. Anisolepis undulatus (S. Paulo, Brazil); I. A. longicauda (Corrien-
tes, Argentina)
Figura 13. LEIOSAURIDAE. Enyaliinae. A. Enyalius brasiliensis (Espirito Santo, Brazil); B. Enyalius
iheringi (Sao Paulo, Brazil); C. E. perditus (Brazil); D. Urostrophus vautieri (Sao Paulo, Brazil);
E. Urostrophus iheringi (S. Paulo, Brazil); F. U. gallardoi (Cordoba, Argentina); G. Anisolepis
grillii (S. Paulo, Brazil); H. Anisolepis undulatus (S. Paulo, Brazil); I. A. longicauda (Corrien-
tes, Argentina)
A B C
D E F
G H I

24 José M. Cei
Figure 14. TROPIDURIDAE. Tropidurini. A. Uranoscodon superciliosus (Northern S. America); B.
Microlophus peruvianus (Arica, Chile); C. Microlophus occipitalis (Ecuador); D. Tropidurus
hispidus (Venezuela); E. Tropidurus torquatus (S. Paulo, Brazil); F. T. etheridgei (La Rioja,
Argentina); G. T. spinulosus (Formosa, Argentina); H. T. spinulosus (Cordoba, Argentina); I.
Tropidurus guarani (Asuncion, Paraguay)
Figura 14. TROPIDURIDAE. Tropidurini. A. Uranoscodon superciliosus (Northern S. America); B.
Microlophus peruvianus (Arica, Chile); C. Microlophus occipitalis (Ecuador); D. Tropidurus
hispidus (Venezuela); E. Tropidurus torquatus (S. Paulo, Brazil); F. T. etheridgei (La Rioja,
Argentina); G. T. spinulosus (Formosa, Argentina); H. T. spinulosus (Cordoba, Argentina); I.
Tropidurus guarani (Asuncion, Paraguay)
A B C
D E F
G H I

25MULTEQUINA 16: 1-52, 2007
Figure 15. TROPIDURIDAE. Tropidurini. A. Tropidurus semitaeniatus (Bahia, Brazil); B. Tropidurus
pictus (male; Salta, Argentina); C. T. pictus (female; Salta, Argentina); D. Tropidurus hygomi
(Sergipe, Brazil); E. Tropidurus bogerti (Bolivar, Venezuela); F. Tropidurus amathites (Bahia,
Brazil); G. Tropidurus strobilurus (NW Brazil); H. Tropidurus plica (Para, Brazil); I. Tropidurus
umbra (French Guyana)
Figura 15. TROPIDURIDAE. Tropidurini. A. Tropidurus semitaeniatus (Bahia, Brazil); B. Tropidurus
pictus (male; Salta, Argentina); C. T. pictus (female; Salta, Argentina); D. Tropidurus hygomi
(Sergipe, Brazil); E. Tropidurus bogerti (Bolivar, Venezuela); F. Tropidurus amathites (Bahia,
Brazil); G. Tropidurus strobilurus (NW Brazil); H. Tropidurus plica (Para, Brazil); I. Tropidurus
umbra (French Guyana)
A B C
D E F
G H I

26 José M. Cei
Figure 16. TROPIDURIDAE. Stenocercini. A. Stenocercus ornatus (Guayaquil, Ecuador); B. Stenocercus
trachycephalus (Bogota, Colombia); C. S. guentheri (Ecuador); D. S. varius (Pichincha, Ecua-
dor); E. Ophryoessoides caducus (Jujuy, Argentina); F. O. iridescens (Esmeralda, Ecuador);
G. Stenocercus roseiventris (Salta, Argentina); H. Stenocercus pectinatus (Rio Negro, Argen-
tina); I. S. doellojuradoi (Santiago del Estero, Argentina)
Figura 16. TROPIDURIDAE. Stenocercini. A. Stenocercus ornatus (Guayaquil, Ecuador); B. Stenocercus
trachycephalus (Bogota, Colombia); C. S. guentheri (Ecuador); D. S. varius (Pichincha, Ecua-
dor); E. Ophryoessoides caducus (Jujuy, Argentina); F. O. iridescens (Esmeralda, Ecuador);
G. Stenocercus roseiventris (Salta, Argentina); H. Stenocercus pectinatus (Rio Negro, Argen-
tina); I. S. doellojuradoi (Santiago del Estero, Argentina)
A B C
D E F
G H I

27MULTEQUINA 16: 1-52, 2007
Critical reports and annotationson so extensive Iguania stem of thefollowed indented taxonomy arefinished. We must comment now onthe next major embranchment,Scleroglossa, then parted into twonatural lineages: Gekkota andAutarchoglossa. The familiesattributed to Gekkota are the like,ecologically versatile Gekkonidae,Diplodactylidae, Eublepharidae andthe unusual snake-like fossorialPygopodidae, reviewed by Kenge(1974) in a prolix fashion. Obviouslyit is impossible to offer hereproportional scutellation samples fortaxa having some 320 species and88 genera, such as the almostcosmopolitan Gekkonidae, or forthe 120 species and 15 genera ofDiplodactylidae from New Zealandand Australia, or the 22 species and6 genera of Eublepharidae, scatteredacross the SW of North America,Central America, Africa andSouthern Asia. However theexamples of Figure 17 areefficacious in expressing the generalsimplicity of supraocularscutellation of all these Gekkota,given the modest specific or genericvariation in their uniform smallscales even reduced to short spots,or almost absent. Also thecircumorbital semicircles have beenrestrained to a nearly concealedstructure. Very different in size andshape are the supraciliaries, mostlylinear and narrow, in some notched
(Rhinogecko misonnei, Homonotafasciata), or sharp-pointed species(Phyllurus platurus, Diplodactylusciliaris). Sometimes growths ortubercles appear on the supraocularsurface, as in Phyllurus cornutus,Ph. caudiannulatus, Nephrurusasper from Queensland andNephrurus wheeleri from WAustralia. Besides Australianexamples, these investigations wereextended to several other samplesfrom Africa, Middle East India, Iranand America. Our research onGekkonidae, Diplodactylidae,Eublepharidae has thus been carriedout on about a hundred and twentyspecimens.
Strongly dissimilar are the morethan 30 species and 8 generabelonging to the snake-likePygopodidae from Australia and NewGuinea. These really unusual lizardsare represented in Figure 18, pointingout their enlarged cephalic shieldsand the extreme reduction of theirsupraocular scales: only one in Aclys,Aprasia and Paradelma, two inDelma, Pygopus and Pletholax,where evident supraciliaries areshown. A most impressive reductionis found in Lialis andOphidiocephalus. Lastly, nothing canbe added concerning the evolutionaryrelationships of Pygopodidae withthe other Gekkota, anatomicalconsiderations being the decisivefactor in this classification.

28 José M. Cei
Figure 17. GEKKOTA. GEKKONIDAE. A. Cyrtopodion heterocercum (Iran); B. Rhinogecko misonnei
(Pakistan); C. Briba brasiliana (Minas Gerais, Brazil); D. Phyllopezus pollicaris (Pernambuco,
Brazil); E. Hemidactylus turcicus (Mediterranean region); F. Hemidactylus persicus (Saudi Arabia);
G. Gonatodes concinnatus (Napo, Ecuador); H. Stenodactylus grandiceps (Jordan, Middle East);
I. Homonota underwoodi (San Juan, Argentina); J. Homonota whitii (Cordoba, Argentina); K.
Homonota darwini (Payunia, Argentina); L. Homonota fasciata (Cordoba, Argentina).
EUBLEPHARIDAE. M. Coleonyx brevis (Texas, USA); N. Eublepharis macularius (Pakistan);
O. Eublepharis turcmenicus (Iran).
DIPLODACTYLIDAE. P. Phyllurus platurus (South Australia)
Figura 17. GEKKOTA. GEKKONIDAE. A. Cyrtopodion heterocercum (Iran); B. Rhinogecko misonnei
(Pakistan); C. Briba brasiliana (Minas Gerais, Brazil); D. Phyllopezus pollicaris (Pernambuco,
Brazil); E. Hemidactylus turcicus (Mediterranean region); F. Hemidactylus persicus (Saudi Arabia);
G. Gonatodes concinnatus (Napo, Ecuador); H. Stenodactylus grandiceps (Jordan, Middle East);
I. Homonota underwoodi (San Juan, Argentina); J. Homonota whitii (Cordoba, Argentina); K.
Homonota darwini (Payunia, Argentina); L. Homonota fasciata (Cordoba, Argentina).
EUBLEPHARIDAE. M. Coleonyx brevis (Texas, USA); N. Eublepharis macularius (Pakistan);
O. Eublepharis turcmenicus (Iran).
DIPLODACTYLIDAE. P. Phyllurus platurus (South Australia)
A B C D
E F G H
I J K L
M N O P

29MULTEQUINA 16: 1-52, 2007
Figure 18. PYGOPODIDAE. A. Aclys concinna (W Australia); B. Aprasia aurita (SE Australia); C. Aprasia
parapulchella (SE Australia); D. Aprasia repens (SW Australia); E. Delma australis (W Aus-
tralia); F. Delma elegans (W Australia); G. Lialis jicari (New Guinea); H. Ophidiocephalus
taeniatus (N Australia); I. Pygopus nigriceps (Australia); J. Pygopus lepidopodus (SW Aus-
tralia); K. Paradelma orientalis (Queensland, Australia); L. Pletholax gracilis (SW Australia)
Figura 18. PYGOPODIDAE. A. Aclys concinna (W Australia); B. Aprasia aurita (SE Australia); C. Aprasia
parapulchella (SE Australia); D. Aprasia repens (SW Australia); E. Delma australis (W Austra-
lia); F. Delma elegans (W Australia); G. Lialis jicari (New Guinea); H. Ophidiocephalus taeniatus
(N Australia); I. Pygopus nigriceps (Australia); J. Pygopus lepidopodus (SW Australia); K.
Paradelma orientalis (Queensland, Australia); L. Pletholax gracilis (SW Australia)
A B C D
E F G H
I J K L

30 José M. Cei
Drastic morphological changes,facing examined Iguania supraocularpatterns, shall now be commentedupon for the successive branches ofAutarchoglossa, foreseen in ourindented taxonomy, beginning withScincomorpha and its noticeable linesLacertoidea and Scincoidea. Withoutany doubt, observing their cephaliclepidosis, some evolutionary trendsare difficult to understand at first sightbetween the night lizards Xantusidaeand the Lacertiformes Lacertidae andTeiioidea of Lacertoidea, as well asthe families Scincidae and Cordylidae,of Scincoidea. Studied formerly bySavage (1963), who firstly assignedthem to Gekkota, these viviparous,secretive reptiles, with their generaLepidophyma, Xantusia andCricosaura (Figure 19), live increvices or cracks of granite flakes, inforested regions of Mexico, CentralAmerica, Southern and WesternUnited States, and there being onlyone species confined to Cuba, themonotypic Cricosaura (C. typica).From Figure 19 we can note that inXantusia vigilis circumorbitalsemicircles are absent, as in X.henshawi or X. arizonae, besides theother genera, and that the supraocularscales consist of a narrow longitudinalrow near the eye border, with noevident supraciliaries; that in Xantusiariversiana (ancient KlauberinaSavage 1957) and Cricosaura typicatwo narrow adjacent rows on the eye
border are visible, the externalnarrower; almost rudimentary in thetwo adjacent supraocular rows ofCercosaura; that in Lepidophymaflavimaculatum, the reduction of thesupraocular scales is extreme, to “anarrow fleshy ridge above the eye”(Savage, 1963).
Leaving so exceptionally modifiedscale features, the specimens reportedin the inferior drawings of Figure 19introduce us to the great Americanstem of Lacertiformes, the Teiioidea,to whose taxon Teiidae belong thereproduced species of the genusTupinambis, still justifying asystematic definition as subfamily.The specific variation in the patternof Tupinambis anticipates here ageneral distribution of supra-ocularscales dominant in Scincomorpha,with major or minor alteration.Lacking a definition of circumorbitalsemicircles, 4-5 supraocular scales,somewhat different in size, stretchout on the eye border, edged by one ortwo narrower linear supraciliary rows.That simple scheme, of the large-sized, powerful Tupinambis shall becommented upon, more or lessmodified, in the following Figures ofTeiioidea, Lacertidae and Scincoidea.Fossil remains of Tupinambis fromSouth American early Miocene tolate Miocene are very suggestiverecords stating the significantevolutionary lineage of Teiid formsin the continent (Albino et al. 2006).

31MULTEQUINA 16: 1-52, 2007
Figure 19. XANTUSIIDAE. TEIIDAE. XANTUSIIDAE. A. Xantusia vigilis (SW USA); B. Xantusia
vigilis (Lower California, USA); C. X. henshawi (SW USA); D. X. riversiana (SW California,
USA); E. Lepidophyma flavimaculatum (Costa Rica); F. Cricosaura typica (E Cuba).
TEIIDAE. G. Tupinambis nigromaculatus (Barinas, Venezuela); H. T. teguixin (E Brasil); I. T.
merianae (Bs. Aires, Argentina); J. Tupinambis rufescens (Mendoza, Argentina)
Figura 19. XANTUSIIDAE. TEIIDAE. XANTUSIIDAE. A. Xantusia vigilis (SW USA); B. Xantusia vigilis
(Lower California, USA); C. X. henshawi (SW USA); D. X. riversiana (SW California, USA);
E. Lepidophyma flavimaculatum (Costa Rica); F. Cricosaura typica (E Cuba).
TEIIDAE. G. Tupinambis nigromaculatus (Barinas, Venezuela); H. T. teguixin (E Brasil); I. T.
merianae (Bs. Aires, Argentina); J. Tupinambis rufescens (Mendoza, Argentina)
A B C
D E F
G H I J

32 José M. Cei
In Figure 20, upper row, the greatestTeiid species are documented. Fivesupraocular scales with much reducedcircumorbital semicircles and relativelywide supraciliaries are shown byAmeiva and Callopistes: 6-5supraocular scales and a narrowersupraciliary row characterizeDracaena. Taxa of Dicrodon andKentropyx, on the intermediate row,exhibit 3-2 larger broad scales with avery small terminal scale, reduced orabsent circumorbital semicircles, butevident overlapping supraciliaries. Onthe inferior row of the figure, therepeated Teiid lepidosis accounts forthe 3 supraocular scales of Teius, withcircumorbital semicircles andoverlapping supraciliaries. For thenumerous and so variable whiptails(Cnemidophorus) we reported 4-3supraocular scales and developedsupraciliaries: only for Cnemidophorusocellifer the circumorbital semicirclewas drawn here.
Corresponding to the oldsubdivision into Microteiids andMacroteiids, the familyGymnophthalmidae was establishedfor the many small-sized formsdistributed with more than 150species and 35 genera in Central andSouth America. Figures 21-22 dealwith those lizards having 4, 3, 2supraocular scales, mainly lackingcircumorbital semicircles but puttingin evidence a more or less widesupraciliary border. Through the
careful Dixon’s research (1973) itwas possible to remark theevolutionary process leading to lossof the supraocular scales in thegymnophthalmid genus Bachia: aprocess whose progressive steps havebeen already accomplished, i.e., inGymnophthalmus underwoodi fromSurinam or Heterodactylusimbricatus from SE Brazil. In Figure22 we can point out the reduced butstill recognizable two supraocularscales of Bachia heteropa(Venezuela), B. flavescens (Guyana),B. pallidiceps (Panama, Colombia),and also their elimination fromBachia talpa (Colombia) to B.intermedius (Peru), with narrowsupraciliaries remaining in the end.Differences in body, scales,ecological relationships andevolutionary trends within the genus,related to fossorial adaptation, werepreliminarily discussed in the citedpaper.
Now we will go on with theinferior row of Figure 22 and Figure23, illustrating characteristicfeatures of the African and Euro-Asiatic great family Lacertidae,with more than 220 species and 25genera. That is our last informationabout Lacertoidea of theScincomorpha embranchment. Thestriking morphological affinities insupraocular lepidosis betweenLacertidae and their counterpartTeiioidea from the American

33MULTEQUINA 16: 1-52, 2007
Figure 20. TEIIDAE. A. Ameiva ameiva (French Guyana); B. Callopistes palluma (Chile); C. Dracaena
paraguayensis (Mato Grosso, Brazil); D. D. guianensis (Guyana); E. Dicrodon guttulatus (Ecua-
dor); F. Kentropyx viridistriga (Corrientes, Argentina); G. Kentropyx lagartica (Tucuman, Ar-
gentina); H. K. pelviceps (Amazonia, Ecuador); I. Teius oculatus (Cordoba, Argentina); J.
Cnemidophorus lemniscatus (Yaracui, Venezuela); K. C. ocellifer (Chaco, Argentina); L. C.
lacertoides (Jujuy, Argentina)
Figura 20. TEIIDAE. A. Ameiva ameiva (French Guyana); B. Callopistes palluma (Chile); C. Dracaena
paraguayensis (Mato Grosso, Brazil); D. D. guianensis (Guyana); E. Dicrodon guttulatus
(Ecuador); F. Kentropyx viridistriga (Corrientes, Argentina); G. Kentropyx lagartica (Tucuman,
Argentina); H. K. pelviceps (Amazonia, Ecuador); I. Teius oculatus (Cordoba, Argentina); J.
Cnemidophorus lemniscatus (Yaracui, Venezuela); K. C. ocellifer (Chaco, Argentina); L. C.
lacertoides (Jujuy, Argentina)
A B C D
E F G H
I J K L

34 José M. Cei
Figure 21. GYMNOPHTHALMIDAE. A. Alopoglossus angulatus (Northern South America); B.
Neusticurus bicarinatus (Brasil, Guyana); C. Pantodactylus schreibersi (Cordoba, Argenti-
na); D. Arthrosaura kockii (Surinam, N Brazil); E. Anadia bipunctata (Merida, Venezuela); F.
Tretioscincus agilis (Guyana, N Brazil); G. Leposoma guianense (Guyana, N Brazil); H. Iphisa
elegans (Northern South America); I. Microblepharus maximiliani (NE Brazil, Paraguay); J.
Prionodactylus argulus (Ecuador, Guyana); K. Gymnophthalmus multiscutatus (NE Brazil);
L. Gymnophthalmus underwoodi (Surinam, Tobago Is.)
Figura 21. GYMNOPHTHALMIDAE. A. Alopoglossus angulatus (Northern South America); B.
Neusticurus bicarinatus (Brasil, Guyana); C. Pantodactylus schreibersi (Cordoba, Argenti-
na); D. Arthrosaura kockii (Surinam, N Brazil); E. Anadia bipunctata (Merida, Venezuela); F.
Tretioscincus agilis (Guyana, N Brazil); G. Leposoma guianense (Guyana, N Brazil); H. Iphisa
elegans (Northern South America); I. Microblepharus maximiliani (NE Brazil, Paraguay); J.
Prionodactylus argulus (Ecuador, Guyana); K. Gymnophthalmus multiscutatus (NE Brazil);
L. Gymnophthalmus underwoodi (Surinam, Tobago Is.)
A B C D
I F G H
I J K L

35MULTEQUINA 16: 1-52, 2007
Figure 22. GYMNOPHTHALMIDAE. A. Heterodactylus imbricatus (SE Brazil); B. Bachia heteropa (Ve-
nezuela); C. Bachia flavescens (Guyana); D. B. pellidiceps (Panama, Colombia); E. B. talpa
(Colombia); F. B. trisanale (Peru, Ecuador); G. B. huallagana (Peru); H. B. intermedia (Peru).
LACERTIDAE. I. Psammodromus algirus (SW Europe, NW Africa); J. Acanthodactylus
erytrurus (SW Europe); K. A. pardalis (Maroc, Marruecos); L. A. boskianus (NW Africa)
Figura 22. GYMNOPHTHALMIDAE. A. Heterodactylus imbricatus (SE Brazil); B. Bachia heteropa (Vene-
zuela); C. Bachia flavescens (Guyana); D. B. pellidiceps (Panama, Colombia); E. B. talpa (Co-
lombia); F. B. trisanale (Peru, Ecuador); G. B. huallagana (Peru); H. B. intermedia (Peru).
LACERTIDAE. I. Psammodromus algirus (SW Europe, NW Africa); J. Acanthodactylus erytrurus
(SW Europe); K. A. pardalis (Maroc, Marruecos); L. A. boskianus (NW Africa)
A B C D
E F G H
I J K L
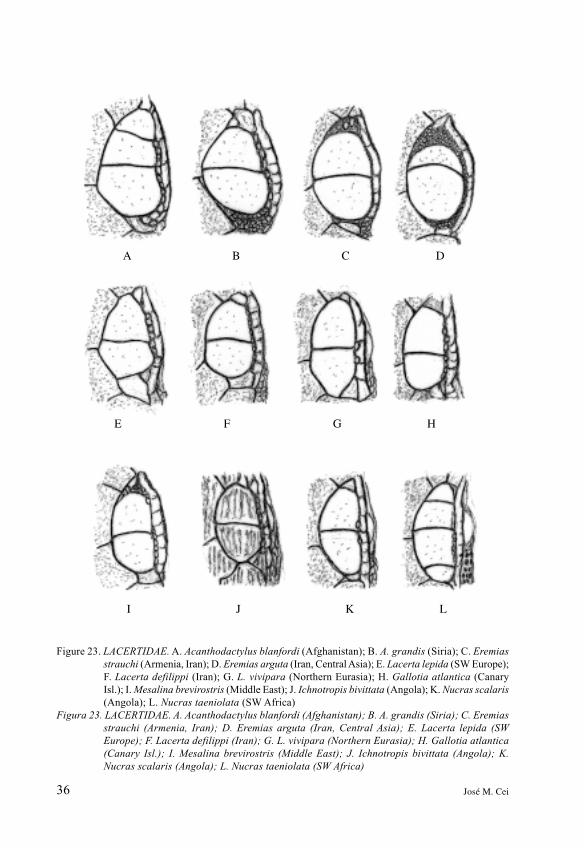
36 José M. Cei
Figure 23. LACERTIDAE. A. Acanthodactylus blanfordi (Afghanistan); B. A. grandis (Siria); C. Eremias
strauchi (Armenia, Iran); D. Eremias arguta (Iran, Central Asia); E. Lacerta lepida (SW Europe);
F. Lacerta defilippi (Iran); G. L. vivipara (Northern Eurasia); H. Gallotia atlantica (Canary
Isl.); I. Mesalina brevirostris (Middle East); J. Ichnotropis bivittata (Angola); K. Nucras scalaris
(Angola); L. Nucras taeniolata (SW Africa)
Figura 23. LACERTIDAE. A. Acanthodactylus blanfordi (Afghanistan); B. A. grandis (Siria); C. Eremias
strauchi (Armenia, Iran); D. Eremias arguta (Iran, Central Asia); E. Lacerta lepida (SW
Europe); F. Lacerta defilippi (Iran); G. L. vivipara (Northern Eurasia); H. Gallotia atlantica
(Canary Isl.); I. Mesalina brevirostris (Middle East); J. Ichnotropis bivittata (Angola); K.
Nucras scalaris (Angola); L. Nucras taeniolata (SW Africa)
A B C D
E F G H
I J K L

37MULTEQUINA 16: 1-52, 2007
Figure 24. SCINCIDAE. A. Leiolopisma coventryi (S Australia); B. Emoia nativitatis (Christmas Is., W
Australia); C. Egernia striolata (E Australia); D. Egernia coventryi (S Australia); E. Ctenotus
inornatus (N Australia); F. C. helenae (W Central Australia); G. Ctenotus dux (Central Austra-
lia); H. C. allotropis (SE Australia); I. Mabuya dorsivittata (Cordoba, Argentina); J. Mabuya
frenata (Brazil); K. Mabuya aurata (Lebanon, Syria); L. Mabuya maculilabris (Ghana, W
Africa)
Figura 24. SCINCIDAE. A. Leiolopisma coventryi (S Australia); B. Emoia nativitatis (Christmas Is., W Aus-
tralia); C. Egernia striolata (E Australia); D. Egernia coventryi (S Australia); E. Ctenotus inornatus
(N Australia); F. C. helenae (W Central Australia); G. Ctenotus dux (Central Australia); H. C.
allotropis (SE Australia); I. Mabuya dorsivittata (Cordoba, Argentina); J. Mabuya frenata (Brazil);
K. Mabuya aurata (Lebanon, Syria); L. Mabuya maculilabris (Ghana, W Africa)
A B C D
E F G H
I J K L

38 José M. Cei
continent sound suggestive. Thesixteen species and eight generarepresented may be a sufficientsample, given our multiple controlscarried out on a remarkable numberof additional specimens. In the wellknown genus Acanthodactylus 3-2broad supraocular scales are therule, without apparentcircumorbital semicircles,pronounced supraciliaries beinginstead separated or not fromsupraoculars by a narrow row ofdiminutive scales. Comparing thispattern with the other drawnsupraocular patterns of Lacertidae,such as Psammodromus, Eremias,Lacerta, Mesalina, Ichnotropis,Nucras or Gallotia, theirfundamental trends of similarityare recognizable at first sight.However, a diligent comparison ofthe supraocular lepidosis inLacertidae with the above reportedsupraocular lepidosis in Teiioidea(Figures 19, 20 and 21) may pointout the independent systematicidentity of both these major taxa,surely in accordance with differentevolutionary lines, in spite of anyindubitable morphologicalconvergence of their supraocularscutellar patterns.
It is now the turn of ourcomments on supraocular lepidosisin Scincoidea families, Scincidaeand Cordylidae - Gerrhosauridae,thus completing our analytical
relation as regards (regarding) theScincomorpha branch ofAutarchoglossa. The generalhomogeneity and shape uniformityof supraorbital scales, beforerevised, are again almost the samein Scincoidea, such as an authentic“Autarchoglossa band”.
In all subfamilies of Scincidae(Greer, 1970) the supraocularpattern exposed in Figures 24-25reveals no noticeable variations foreither the 6, 5, 4, 3 broadlongitudinal scales, or the reducedsupraciliaries known forLeiolapisma, Emoia, Egernia,Ctenotus from Australia, Mabuyafrom middle East Africa and SouthAmerica, Eumeces taeniolatusfrom Pakistan, Eumeces obsoletusfrom Kansas, USA, Scincus scincusfrom Irak, or Scincus mitranus fromSaudi Arabia.
The scarce exceptions,registered in Figure 25, were drawnfor Sepsina (Angola), Chalcides(South Europe, Canary Islands),Malaconthas holomelas,Typhlosaurus lineatus (S. Africa),Tiliqua occipitalis (Australia),Ablepharus kitaibelii (SE Europe).All the evolutionarydiversifications (scale reduction,shape, etc.) likely originated fromsome primitive common strainunder adaptive environmental push(ecological, climate changes,fossorial life, etc.).

39MULTEQUINA 16: 1-52, 2007
Figure 25. SCINCIDAE. A. Eumeces taeniolatus (Pakistan); B. Eumeces obsoletus (Kansas, USA); C.
Scincus scincus (Irak, Arabia); D. Scincus mitranus (Saudi Arabia); E. Sepsina copei (Angola,
W Africa); F. Sepsina bayonii (Angola, W Africa); G. Chalcides chalcides (S Italy); H. Chalcides
viridens (Canary Isl.); I. Malacontias holomelas (Madagascar); J. Typhlosaurus lineatus (S
Africa); K. Tiliqua occipitalis (Australia); L. Ablepharus kitaibelii (SE Europe, Turkey)
Figura 25. SCINCIDAE. A. Eumeces taeniolatus (Pakistan); B. Eumeces obsoletus (Kansas, USA); C.
Scincus scincus (Irak, Arabia); D. Scincus mitranus (Saudi Arabia); E. Sepsina copei (Angola,
W Africa); F. Sepsina bayonii (Angola, W Africa); G. Chalcides chalcides (S Italy); H. Chalcides
viridens (Canary Isl.); I. Malacontias holomelas (Madagascar); J. Typhlosaurus lineatus (S
Africa); K. Tiliqua occipitalis (Australia); L. Ablepharus kitaibelii (SE Europe, Turkey)
A B C D
E F G H
I J K L

40 José M. Cei
Figure 26. CORDYLIDAE. A. Cordylus vittifer (Angola); B. Cordylus giganteus (S Africa) ; C. Cordylus
polyzonus (S Africa); D. Pseudocordylus microlepidotus (Cape Prov., S Africa); E. Platysaurus
guttatus (S Africa); F. Platysaurus capensis (S Africa); G. Chamaesaura aenea (Transvaal, S
Africa); H. Chamaesaura anguina (Cape Prov., S Africa).
GERRHOSAURIDAE. I. Gerrhosaurus major (S Africa); J. G. nigrolineatus (S Africa); K.
Zonosaurus maximus (Madagascar); L. Tetradactylus tetradactylus (S Africa)
Figura 26. CORDYLIDAE. A. Cordylus vittifer (Angola); B. Cordylus giganteus (S Africa); C. Cordylus
polyzonus (S Africa); D. Pseudocordylus microlepidotus (Cape Prov., S Africa); E. Platysaurus
guttatus (S Africa); F. Platysaurus capensis (S Africa); G. Chamaesaura aenea (Transvaal, S
Africa); H. Chamaesaura anguina (Cape Prov., S Africa).
GERRHOSAURIDAE. I. Gerrhosaurus major (S Africa); J. G. nigrolineatus (S Africa); K.
Zonosaurus maximus (Madagascar); L. Tetradactylus tetradactylus (S Africa)
A B C D
E F G H
I J K L

41MULTEQUINA 16: 1-52, 2007
Cordylidae and Gerrhosauridae,considered as separate families inrecent classifications, are lizardsliving in African and Madagascarterritories. Exemplified in Figure26, we remark the impressivesimilarity between theirsupraocular lepidosis and the aboveanalysed scincid pattern. Thesupraciliary borders of theseScincomorpha are wider and moredeveloped than supraciliaries ofScincidae. In the large, armouredCordylus species, such as C.giganteus, C. cataphractus, C.warreni, C. polyzonus, thesupraocular region is subject tosourrounding quills, horns, sharp-points of a strong dermal sclerosis.The broad longitudinal scales of sopeculiar reptiles differ from 6 to 4in number (mostly 4). An analogousremark can be made for other generaof Cordylidae and Gerrhosauridae,such as Platysaurus, Gerrhosaurus,Zonosaurus, Tetradactylus or theunusual snake-like Chamaesaurawhose residual legged speciesexhibit deeply keeled scales.
To all appearance the “scincoid”model of supraocular scutellation isa prevalent one in the Scleroglossataxon. Indeed, if we examine asample of Squamata Anguimorpha,the great family Anguidae, i.e., withits more than 100 species and 13genera, we get back to the kind ofsupraocular pattern we were
reporting before for scincids as someEumeces, Scincus or Chalcides, alllizards showing a tendency tominimization of legs and to havingsnake-like features. Of course, someadaptive Anguid characters arerecognizable in drawings of Figure27, as the strengthening ofsupraciliaries or the 7, 6, 5, 4 broadsupraocular scales. A snake-likeform is besides visible in the Euro-Asiatic Anguis, in Ophisaurusgracilis or O. apodus from SE Asia,in the several Diploglossus andOphiodes from South America andWest Indies.
In Estes et al.’s indentedtaxonomy, Xenosauridae,Anguidae and Varanoidea areincluded under the taxonAnguimorpha. But, as these authorsstated, “the Xenosauridae andAnguidae cannot be demonstratedto be more closely related to eachother than either is to Varanoids”.Therefore, the authors placedxenosaurs, anguids and varanoidsin an unresolved triatomy, and theymentioned that a metataxonAnguioidea wants sufficient reasonfor being formally recognized.
The impoverished familyXenosauridae adds to its strange,disjoint distribution (China,Mexico) some peculiarities of itssupraocular scutellation, being thusin accordance with the sentencenot long ago stated here on the

42 José M. Cei
Figure 27. ANGUIDAE. A. Anguis fragilis (SE Europe); B. A. fragilis (N Europe); C. Ophisaurus gracilis
(Hymalayas, Burma); D. Elgaria coerulea (Pacific coast USA, Costa Rica); E. Diploglossus
lessonae (NE Brazil); F. D. hewardii (Jamaica, West Indies); G. D. monotropis (Costa Rica);
H. D. warreni (Hispaniola, West Indies); I. Ophiodes yacupoi (Misiones, Argentina); J. O.
striatus (S. Paulo, Brazil); K. Ophiodes intermedius (Entre Ríos, Argentina); L. O. vertebralis
(Buenos Aires, Argentina)
Figura 27. ANGUIDAE. A. Anguis fragilis (SE Europe); B. A. fragilis (N Europe); C. Ophisaurus gracilis
(Hymalayas, Burma); D. Elgaria coerulea (Pacific coast USA, Costa Rica); E. Diploglossus
lessonae (NE Brasil); F. D. hewardii (Jamaica, West Indies); G. D. monotropis (Costa Rica);
H. D. warreni (Hispaniola, West Indies); I. Ophiodes yacupoi (Misiones, Argentina); J. O.
striatus (S. Paulo, Brasil); K. Ophiodes intermedius (Entre Ríos, Argentina); L. O. vertebralis
(Buenos Aires, Argentina)
A B C D
E F G H
I J K L

43MULTEQUINA 16: 1-52, 2007
precariousness of systematiccategories such as Anguimorphaor Anguioidea. In fact, the Chinesemonotypic Shinisaurus (S.crocodilurus, Figure 28) displays asupraocular bare covering withparticular circumorbital semicirclesand supraciliaries, round asomewhat eccentric relief morerelated to a Chinese decoration thanto cephalic lepidosis. The fewXenosaurus species from Mexico,on the contrary, exhibit a veryregular Squamata supraocularpattern with some Pleurodontidreminiscences, affording complete,evident circumorbital semicircles,opposed to moderate supraciliariesand, within their boundaries, alongitudinal row of broader scales,4-5 in number, having some rowsof irregular smaller scales on bothsides.
In this same Figure 28, taxabelonging to the heterogeneousVaranoidea assemblage arepresented. It is about two small,probably primitive families, asLanthanotidae with only one speciesfrom Borneo, or Helodermatidaewith two species from Mexico andSW United States, besides theremarkable Varanidae, whose morethan 55 species of the unique genusVaranus spread from Africa and SEAsia to Australia and New Guinea.In Lanthanotus borneensis, thesupraocular region is uniquely
covered by dense, irregular smallscales, without circumorbitalsemicircles but bordered byoverlapping ciliaries. Somewhatsimilar is the pattern of Helodermasuspectum and H. horridum, therobust venomous lizards from sandyAmerican deserts, even though sizeand distribution of their prominentround scales are distinct. In Figure28, inferior row, the supraocularcovering of the African Varanusniloticus and V. exanthematicus isdrawn, together with that of theSouth Asiatic Varanus monitor. Theuncertain boundaries of reducedsupraocular semicircles, the absenceof broad scales and the very narrowciliaries of the African forms can bepointed out, contrasting with themore regular small supraocularscales of Varanus monitor, whosesupraciliaries are reduced in theextreme. In Figure 29, a selection ofsupraocular coverings in severalspecies from the extensivedistribution of the genus is provided,from the gigantic Varanuskomodensis or the differentiated scalesof Varanus grayi from Philippines,where circumorbital semicircles areidentifiable, to the very variablepatterns of Australian taxa, where onlythe minimized scales of supraocularcoverings and the utmost restrictedspace of supraciliaries are commoncharacters.

44 José M. Cei
Figure 28. XENOSAURIDAE. A. Shinisaurus crocodilurus (China); B. Xenosaurus rackhani (Mexico);
C. Xenosaurus penai (Mexico). HELODERMATIDAE. D. Heloderma suspectus (SW USA,
N Mexico); E. Heloderma horridum (Tehuantepec, Mexico). LANTHANOTIDAE. F.
Lanthanotus borneensis (Borneo). VARANIDAE. G. Varanus niloticus (E Africa); H. V.
exanthematicus (S Africa); I. V. monitor (S Asia)
Figura 28. XENOSAURIDAE. A. Shinisaurus crocodilurus (China); B. Xenosaurus rackhani (Mexico);
C. Xenosaurus penai (Mexico). HELODERMATIDAE. D. Heloderma suspectus (SW USA, N
Mexico); E. Heloderma horridum (Tehuantepec, Mexico). LANTHANOTIDAE. F. Lanthanotus
borneensis (Borneo). VARANIDAE. G. Varanus niloticus (E Africa); H. V. exanthematicus (S
Africa); I. V. monitor (S Asia)
A B C
D E F
G H I

45MULTEQUINA 16: 1-52, 2007
Figure 29. VARANIDAE. A. Varanus komodoensis (Komodo Isl. Indonesia); B. Varanus grayi (Philippine
Isls.); C. V. nebulorus (Burma, Malasia); D. V. mitchelli (N Australia); E. V. giganteus (N
Australia); F. V. gouldii (N Australia); G. V. spenceri (N Australia); H. V. mertensi (SW Austra-
lia); I. V. timorensis (Timor Isls., Australasia)
Figura 29. VARANIDAE. A. Varanus komodoensis (Komodo Isl. Indonesia); B. Varanus grayi (Philippine
Isls.); C. V. nebulorus (Burma, Malasia); D. V. mitchelli (N Australia); E. V. giganteus (N
Australia); F. V. gouldii (N Australia); G. V. spenceri (N Australia); H. V. mertensi (SW Aus-
tralia); I. V. timorensis (Timor Isls., Australasia)
A B C
D E F
G H I

46 José A. Scolaro, F. Videla , S. Puig y A. Marcus
DISCUSSION
We must apologize for so long,often monotonous descriptivereports despite their unavoidabledocumentary essence. As firstsentence, we can now state that apostulated relationship between themany morphological trends ofLacertilian supraocular lepidosisand a taxonomic evolutionarysystem is a reality. That is supportedby drawings of our 27 Figures andtheir joined analytical comparativecomments. We would like to addthat the chosen indented taxonomyfrom the thought over treatise byEstes et al. (1988) was a verycongruent one with the peculiarfeatures and the systematicdistribution of the many taxaexamined in this paper. A still moreconvincing observation can beemphasized here with theundoubted affinity of our resultswith the reclassification of theIguania by Frost et al. (2001). Thefamilies proposed in such areclassification are wellindividualized by their supraocularscale patterns, thus a first distinctionbetween Acrodonta andPleurodonta is possible on theselepidosis characters. The almostuniform supraocular lepidosis ofAgamid or Chamaeleonid taxa,with their more or less rounded orsquared moderate scales, is in factat first sight dissimilar from the
well differentiated supraocularscales in Pleurodontid families asLeiocephalidae, Phrynosomatidae,Polychrotidae and Tropiduridae.Impressive is the familyLeiosauridae, whose supraocularscutellation is unmistakable, ofcourse taking generic and specificvariations into account. But evenmore remarkable is its partition intotwo subfamilies, Leiosaurinae andEnyaliinae, proposed by Frost etal. (2001) and reproduced in thedifferentiation of supraocular scalesin their respective taxa. Genericand specific variations occur inother families such asHoplocercidae, Corytophanidae,Iguanidae, Liolaemidae, alreadywith a recognizable Pleurodontbrand, however with someprimitive characters as inOpluridae, and exceptionaldivergence perhaps adaptive as inCrotaphytidae.
The great Squamata division intoIguania and Scleroglossa appearsfully in accordance with theevolutionary morphological statusof the supraocular lepidosis inlizards. A first separation betweenGekkota and Autarchoglossa is alsojustified. In the cosmopolitanGekkota stem, the familiesGekkonidae, Diplodactylidae,Eublepharidae are distinguished bya variable expression of theirscantily differentiated supraocular

47MULTEQUINA 16: 1-52, 2007
lepidosis, wanting recognizablecircumorbital semicircles,showing moderate supraciliariesexceptionally prominent or sharp-pointed, and having a veryheterogeneous layer of moderateroundish scales, frequentlyrarefied. Striking is a comparisonwith the joined snake-like family,the Australian Pygopodidae,whose adaptive modifications ofthe cephalic lepidosis, formerlystudied by Kluge (1974), reaffirmthe importance of supraocularscales as a discriminatingtaxonomic character.
Autarchoglossa and itssubdivisions exhibit the most clear-cut difference from Iguania becauseof the supraocular lepidosis of allits numerous taxa, eitherScincomorpha or Anguimorpha. Ageneral pattern of some wide scales(2 to 7) longitudinally disposedalong the ciliary border is repeatedeither in Lacertoidea or Scincoidea,as well as in Anguidae. A somewhatdiscordant supraocular scalereduction of the family Xantusiidaeis unusual and takes us back to itsprobably uncertain taxonomicposition invoked in a past carefulSavage’s research (1963).
The two major branches ofLacertiformes, Lacertidae fromAfrica and Eurasia, Teiioidea fromthe American continents, put in
evidence a clear-cut example ofparallel evolution, with their almostsimilar supraocular lepidosispatterns illustrated in our Figures19-23. Some minor features attainto highlight the naturalcharacteristics of an African orEuropean lacertilid as well as thoseof a South American Teiid orGimnophthalmid.
Gymnophthalmidae andTeiidae, formerly considered asmicroteiids or macroteiids aremoreover distinguished now atfamily level also by their significantsupraocular morphology. Thepeculiar case of the evolutionarymodification of supraocular scalesuntil their disappearance in theGymnophthalmid genus Bachia hasbeen reported. Scincoidea is stillmore respectful of the mentionedAutarchoglossa model ofsupraocular scutellation, either inits cosmopolitan family Scincidaeor in its related African familyCordylidae, now divided by thecreation of the related taxonGerrhosauridae. With unavoidablydistinct morphological features,also Anguidae, South American andEuro-Asiatic lizards appear not todeviate from the just commentedmorphological expression ofsupraocular lepidosis in Scincoidea.But the other taxon placed inAnguimorpha in our chosenindented taxonomy, the family

48 José M. Cei
Xenosauridae, is the rarest findingin the whole Scleroglossaembranchment, either for itsdisjoint distribution (China,Mexico) or for the unusualsupraocular scales. The probableart if icial i ty of the taxonAnguimorpha was considered bythe same authors of the indentedtaxonomy.
We reach the end of a simple,objective discussion of data fromour patient comparative checkingof hundreds of Squamata lizards.The last scanty familiesHelodermatidae and Lanthanotidaeshow a quite uniform, likelyprimitive, supraocular lepidosis,with roundish scales, evidentsupraciliaries but indistinctcircumorbital semicircles. Theextensive family Varanidae,unknown in the Americancontinents, assembles a veryvariable set of supraocularcoverings, from scales distinctlyarranged to almost naked surfaces,mottled or variegated. A typicalVaranid supraocular pattern is thusindefinable.
CONCLUSIONS
Conclusions of a scientific paperare the summarized exposition ofevery confirmed novelty orimprovement concerning theoriginal status or condition of
morphological or biologicalsystems that are the subject ofresearch. Thus, in a syntheticsequence, the first conclusion ofthe present study is that thesupraocular lepidosis in theSquamata taxa is not a structure atrandom, but a constant, definedscale arrangement, different inshape and size, in accordance withthe species or other taxa consideredand their evolutionary systematicposition. Second conclusion is thesignificant agreement observedbetween a classificatory system, asthe indented taxonomy elicited byEstes et al. (1988), and the resultsof our comparative research,leading to establishing definedsupraocular lepidosis patterns ascomparable morphologicalcharacters of every taxon. Thirdconclusion is that a reassessmentof the Iguanian Pleurodonta taxonof Estes et al.’s indented taxonomy,as in the reclassification proposedby Frost et al. (2001), is also inagreement with morphologicalresults on the supraocularscutellation of Squamata, shown inour comparative review.
A final conclusive remark isthe repeated application of thepointed out relationships betweenindented phylogenetic taxonomyand differentiated supraocularlepidosis, such as in the case ofseveral systematic new proposals.

49MULTEQUINA 16: 1-52, 2007
The new Pleurodont familyLeiosauridae and its twosubfamilies must be hereremembered, according to Frost etal. (2001). Other significantsystematic agreements in relationto supraocular lepidosis can be citedfor the families Scincoidea,Scincidae and Cordylidae and forthe families Teiioidea, Teiidae andGymnophthalmidae (macroteiidsand microteiids).
ACKNOWLEDGEMENTS
For their valuable aid in theselection of documentaryspecimens I thank very much thefriendly support of Kraig Adler(Dep. Neurobiology, CornellUniversity, Ithaca, N.Y., USA), andof William E. Duelman (Nat. Hist.Mus. Univ, of Kansas, Lawrence,Kansas, USA). My acknowledgementmust be here emphasized for the heavyand careful work of Valeria Corbalán(IADIZA-CONICET) in the editorialphase of my paper, given theunfortunate conditions of my personalactivity during this period.
His wife Giudita and his disciples J.Alejandro Scolaro and FernandoVidela considered the publication ofthis paper to be a very importantcontribution to science, andacknowledge the editorial revisionmade by Nelly Horak, Silvia Puigand Nora Ibarguengoytía.
NOTE: This manuscript was still on hisdesk when unfortunately Dr. Cei died. He
was internationally recognized and his
work will remain indelible for futuregenerations of herpetologists, because of
his profuse and invaluable scientific
contribution and proposal of new ideasfor the biological sciences and evolution.
REFERENCES
ALBINO, A.M., S. BRIZUELA &C.I. MONTALVO, 2006. NewTupinambis remains from thelate Miocene of Argentina anda review of the SouthAmerican Miocene Teiids. J.Herpetol. 40(2): 206-213.
ANDERSON, S.T., 1999. TheLizards of Iran. SSARContributions to Herpetology,Vol. 15. 442 pp.
ANGEL, F., 1942. Les Lézards deMadagascar. Mem. Acad.Malgache 36: 1-193.
BLARC, Ch.P., 1977. ReptilesSauriens Iguanidae. Faune deMadagascar. ORSTOM -CNRS, París, 45. 195 pp.
BRYGOO, E.R., 1971. ReptilesSauriens Chamaelonidae,genre Chamaeleo. Faune deMadagascar. ORSTOM -CNRS, París, 33. 318 pp.
BRYGOO, E.R., 1978. ReptilesSauriens Chamaelonidae,genre Brookesia etcomplements pour la genreChamaeleo. Faune deMadagascar. ORSTOM -CNRS, París, 47. 173 pp.

50 José M. Cei
CEI, J.M., 2003. Specific supraocularscutellation patterns assignificant diagnostic characters:a taxonomic inter andintrageneric “Finger-Print” inLacertilia. Facena 19: 155-161.
CEI, J.M., J.A. SCOLARO & F.VIDELA, 2003. A taxonomicrevision of recognized Argentinespecies of the Leiosaurid genusDiplolaemus (Reptilia,Squamata, Leiosauridae).Facena 19: 87-106.
CEI, J.M., J.A. SCOLARO & F.VIDELA, 2004. An updatedbiosystematic approach to theLeiosaurid genus Pristidactylus.Boll. Mus. Reg. ScienzeNaturali, Torino (Italy) 21(1):159-192.
DIXON, J.R., 1973. A systematicreview of the Teiid lizards,genus Bachia, with remarkson Heterodactylus andAnotosaura. MiscellaneousPublications 57, Mus. Nat.Hist., Univ. of Kansas. 47 pp.
DONNELLY, M.A. & C.W.MYERS, 1991. Herpetologi-cal results of the 1990 Ven-ezuelan expedition to the sum-mit of Cerro Guaiquirima, withnew tepui reptiles. AmericanMus. Novitates 3017. 54 pp.
ESTES, R., K. DE QUEIROZ & J.GAUTHIER, 1988. Phyloge-netic relationships withinSquamata. Pp. 119-281, In:Estes R. and G. Pregill (Eds.),Phylogenetic relationships ofthe lizard families. Essays com-memorating Charles L. Camp.Stanford University Press,California, USA, 631 Pp.
ETHERIDGE, R., 1970. A reviewof the South American iguanidgenus Plica. Bull. Br. Mus.Nat. Hist. (Zoology) 19: 237-256.
ETHERIDGE, R., 1995. Redescrip-tion of Ctenoblepharisadspersa Tschudi, 1845, andthe taxonomy of Liolaeminae(Reptilia, Squamata,Tropiduridae). Amer. Mus.Novitates 3142: 24 pp.
ETHERIDGE, R. & K. DEQUEIROZ, 1988. Phylogenyof Iguanidae. Pp. 283-367, In:Estes, R and G. Pregill (Eds.),Phylogenetic relationships ofthe lizard families. Essays com-memorating Charles L. Camp.Stanford University Press,California, USA, 631 Pp.
FROST, D.R., 1992. Phylogeneticanalysis and taxonomy of theTropidurus group of Lizards(Iguania, Tropiduridae). Amer.Mus. Novitates 3033: 1-68.
FROST, D.R. & R. ETHERIDGE,1989. A phylogenetic analysisand taxonomy of IguaniaLizards (Reptilia, Squamata).Misc. Publ. Mus. Nat. Hist.Univ. Kansas USA 81: 1-65.

51MULTEQUINA 16: 1-52, 2007
FROST, D.R., A.G. KLUGE & D.M.HILLIS, 1992. Species incontemporary herpetology:comments on phylogeneticinferences and taxonomy.Herpetological Review 23(2):46-54.
FROST, D.R., R. ETHERIDGE, D.JANIES & T.A. TITOS, 2001.Total evidence, sequencealignment, evolution ofPolychrotid Lizards, and areclassification of the Iguania(Squamata, Iguania). Amer.Mus. Novitates 3344: 1-38.
GREER, A.E., 1970. A subfamilialclassification of Scincidlizards. Bull. Mus. Comp. Zool.139(3): 151-184.
GRISMER, L.L., J.A. McGUIRE &B.D. HOLLINGSWORTH,1994. A report on theHerpetofauna of the VizcainoPeninsula, Baja California,Mexico, with a Discussion ofits biogeographic andtaxonomic implications. Bull.Southern California Acad. Sci.93(2): 45-80.
HAAS, G. & Y. L. WERNER, 1969.Lizards and Snakes fromSouthwestern Asia, collectedby Henry Field. Bull. Mus.Comp. Zool. Harvard Univ.138(6): 327-406.
HALLIDAY, T. & K. ADLER, 2002.The Firefly encyclopedia ofreptiles and amphibians.Firefly Books Ltd., Ontario,Canada: 240 pp.
HOOGMOED, M.S., 1973. Noteson the Herpetofauna ofSurinam. IV. The Lizards andAmphisbaenians of Surinam.W. Junk, The Hague. 417 pp.
KLUGE, A.G., 1974. A taxonomicrevision of the lizards familyPygopodidae. Miscell.Publications, Mus. Zoology,University of Michigan, 1-227 pp.
LEVITON, A.E., S.C. ANDERSON,K. ADLER & S.A. MINTON,1992. Handbook to middle eastamphibians and reptiles. SSARContributions to Herpetology8. Oxford, Ohio. 252 pp.
MERTENS, R., 1958. Bemerkungenüber die Varane Australiens.Senck Biol. 39(5-6): 229-264.
MERTENS, R., 1959. Liste derVarane Asiens under der Indo-australischen Inselwelt mits y s t e m a t i s c h e nBemerkungen. Senck Biol.40(5-6): 221-240.
PETERS, J.A., 1964. Dictionary ofHerpetology. Hafner Publ. Co.New York. 392 pp.
PREGILL, G.K., 1992. Systematics ofthe West Indian lizard genusLeiocephalus (Squamata,Iguania, Tropiduridae).Miscellaneous Publ. 84. Mus.Nat. Hist. Univ. of Kansas. 69 pp.
RUIBAL, R., 1964. An annotatedchecklist and key to theAnoline lizards of Cuba. Bull.of the Museum ComparativeZoology 130 (8)�: 476-520.

52 José M. Cei
SAVAGE, J.M., 1963. Studies on thelizard family Xantusiidae IV.The genera. Contrib. Sci. .LosAngeles County Mus. 71: 1-38.
SCHULTE II, J.A., J.P.VALLADARES & A.LARSON, 2003. Phylogeneticrelationships within Iguanidaeinferred using molecular andmorphological data and aphylogenetic taxonomy ofiguanian lizards. Herpetologica59: 399-419.
TAYLOR, E.H., 1956. A review oflizards of Costa Rica. Univ.Kansas Sci. Bull. 38: 1-320.
VAN DEMBURG, J., 1978.Herpetology of lowerCalifornia�: Collected Papers.Facsimile reprints inherpetology. SSAR. 85 pp.
VANZOLINI, P.E. & J. VALENCIA,1965. The genus Dracaena,with a brief consideration ofmacroteiid relationships(Sauria, Teiidae). Arquivos deZoologia, 13: 1-35.
VANZOLINI, P., A. M. RAMOS-COSTA & L.J. VITT., 1980.Repteis das Caatingas. AcademiaBrasileira de Ciencias, Río deJaneiro. 161 pp.
WIENS, J.J., 1993. Phylogeneticsystematics of the tree lizards(genus Urosaurus).Herpetologica 49(4): 399-420.
Recibido: 04/2007
Aceptado: 07/2007