comparative of … ofpepcin c3 plants is regulated by light as in c4 plants. wefound that wheat (c3)...
TRANSCRIPT

Plant Physiol. (1987) 85, 947-9510032-0889/87/85/0947/05/$01.00/0
Comparative Studies of Phosphoenolpyruvate Carboxylase fromC3 and C4 Plants'
Received for publication May 7, 1987 and in revised form August 10, 1987
MAKOTO MATSUOKA* AND SHINGO HATANational Institute ofAgrobiological Resources, Tsukuba Science City, Yatabe, Ibaraki 305 Japan
ABSTRACT
Phosphoenolpyruvate carboxylase (PEPC) from several C3 plants wascompared to maize PEPC by immunoblotting using an antibody againstmaize PEPC and by peptide mapping. In C3 gramineous plants, PEPCsof slightly different monomeric sizes were detected as two bands forwheat and barley leaves, as three bands for etiolated maize leaves and asfour bands for rice leaves by SDS-polyacrylamide gel electrophoresis andimmunoblotting, whereas only one PEPC band was detected for maizeleaves, a C4 plant, or tobacco leaves, a dicotyledonous C3 plant. Thepeptide fragment patterns of the lower molecular weight PEPC (majorband in immunoblotting) in wheat leaves was similar to that of maizePEPC in peptide mapping by protein staining or by immunologicaldetection, but the upper one (minor band) had a different pattern fromthe lower one in peptide mapping by immunological detection and fewpeptide fragments from this were recognized by the anti-(maize) PEPCantibody. These results suggest that there are multiple forms of PEPCsubunits in the gramineous plants tested, and the major PEPC has aprimary structure similar to that of maize PEPC. To obtain informationabout the expression of PEPCs in C3 plants, changes in the amount ofPEPC protein were investigated during the greening of rice and wheatseedlings. Judging from the regulation by light, there were two types ofPEPCs in greening rice seedlings, one induced by light and the otherreduced by it. Greening wheat seedlings also show a PEPC band inducedby light. These findings indicate that some PEPCs in C3 gramineousplants not only have structures similar to that of maize PEPC, but alsoare regulated by light in a similar manner.
Phosphoenolpyruvate carboxylase (EC 4.1.1.31, PEPC) iswidespread among all plants, including C3, C4, and CAM species,and plays a role in the initial carbon fixation reaction in C4 andCAM plants (reviewed in Ref. 17). The enzyme in C4 and CAMplants have been well characterized as to their biochemical andkinetic properties (2, 19), regulation of gene expression (3, 9, 15,18), and molecular structure (7, 11, 12). However, few reportshave appeared on the molecular structure and gene expressionof PEPC in C3 plants (1).We have been conducting biochemical comparisons of C4-
photosynthetic enzymes between C3 and C4 plants because theseenzymes can be used in the study ofchanges in protein propertiesand gene expression in the evolution ofC3- to C4-photosynthesis.We have found a light-inducible pyruvate, Pi dikinase in C3plants which has a similar structure to that in C4 plants and
'Supported by a grant (GEP II- I- 1-2-e) from the Ministry of Agricul-ture, Forestry and Fisheries of Japan.
considered the possibility of similar gene(s) for PPDK2 in C3 andC4 plants (8).
In the present work, we investigated the homology of thePEPC structures in C3 and C4 plants by immunoblotting andpeptide mapping techniques, and also investigated whether theinduction of PEPC in C3 plants is regulated by light as in C4plants. We found that wheat (C3) PEPC and maize (C4) PEPChad similar structures and the changes in some PEPC proteinsduring the greening of etiolated rice and wheat seedlings weresimilar to that of maize PEPC.
MATERIALS AND METHODSPlant Materials. Seeds ofrice (Oryza sativa L. cv Nipponbare),
wheat (Triticum aestivum L. cv Norin 61), barley (Hotdeumdistichum L. cv New Golden), and maize (Zea mays L. cvGolden Cross Bantam) were obtained from local fields. Unlessotherwise specified, they were planted in vermiculite and culti-vated in a greenhouse at 25 to 3O°C for 7 d under natural light.Tobacco (Nicotiana tabacum L. cv Samsun) was cultivated inpots under natural light in a greenhouse for 2 to 3 months aftersowing. The tip sections of young leaves (0.05 g fresh weight)were used for protein analyses. Etiolated maize leaves were grownat 30°C for 3 d in the dark in vermiculite.Greening Experiments. Rice or wheat seeds were germinated
and grown at 30°C for 7 d or 25°C for 6 d, respectively, in thedark in vermiculite. The etiolated seedlings (about 15 cm inlength) were irradiated with white light (20 K lux) in a growthchamber at 25°C for the indicated periods. The tip sections ofthe first and second leaves were used for further experiments.
Preparation of Antibody. Maize PEPC was prepared by themethod of Uedan and Sugiyama (19). The final preparation wassubjected to preparative SDS-PAGE in a 5 mm wide running gel(10% polyacrylamide containing 0.1% SDS) at 5 V/cm. Afterelectrophoresis, the gel was stained with 2.5 M Na acetate, thetransparent band of PEPC was cut out, and the gel containingpurified PEPC monomer was mixed with Freund's completeadjuvant. A total of0.5 mg ofPEPC was subcutaneously injectedinto a rabbit (New Zealand White) over 1 month. Ouchterlonydouble-diffusion tests showed that the antiserum obtainedformed a single precipitin line with crude extracts of maize leafand with purified PEPC.Nonimmunized serum did not cross-react with PEPC.Protein Blotting. Each plant tissue (0.05 g fresh weight) was
ground in 2 ml of buffer containing 0.1 M Tris-HCl (pH 7.4), 10mM MgSO4, 2.5 mM pyruvate, 2 mm KH2PO4, 1 mm EDTA,0.5% ascorbate (w/v), and 10 mM 2-mercaptoethanol as de-scribed by Uedan and Sugiyama ( 19), and centrifuged at 12,000gfor 10 min. Aliquots of the resulting supernatant fluid (30 ,g of
'Abbreviations: PPDK, pyruvate, Pi dikinase; PEPC, phosphoenol-pyruvate carboxylase; RuBisCO, ribulose 1,5-bisphosphate carboxylase/oxygenase.
947 www.plantphysiol.orgon June 3, 2018 - Published by Downloaded from Copyright © 1987 American Society of Plant Biologists. All rights reserved.

MATSUOKA AND HATA
protein) were subjected to SDS-PAGE on 10% polyacrylamideslab gels containing 0.2% SDS (13). In the case of maize greenleaves, the extract containing 1 ug of protein was subjected toSDS-PAGE. After electrophoresis, the proteins were transferredfrom the gels to nitrocellulose sheets ( 14). The sheets were probedwith anti-PEPC serum and horseradish peroxidase-conjugatedgoat anti-(rabbit)IgG antibody.
Peptide Mapping. Maize PEPC purified as the method ofUedan and Sugiyama (19) was subjected to SDS-PAGE andstained with Coomassie brilliant blue R. After destaining, theband of PEPC was cut out of the gel. Wheat PEPC was partiallypurified by ammonium sulfate fractionation and DEAE-cellulosecolumn chromatography and subjected to SDS-PAGE. Afterelectrophoresis, gel slices corresponding to the positions of im-munoreactive proteins in blotting were cut out. The gel sliceswere put into sample wells of a second SDS-polyacrylamide geland then overlaid with 10 ,l of 50 ,ug/ml Staphylococcus aureusV8 protease in 0.125 M Tris-HCI buffer (pH 6.8) containing 0.1%SDS. The proteins were digested by the protease for 30 min inthe stacking gel, and the resulting peptides were resolved in theseparating gel (5). The peptides were detected by silver staining(16) or by protein blotting as described above.
Immunoprecipitation of PEPC. The indicated volumes of anti-(maize) PEPC antiserum and nonimmune serum were added to50 ul of the enzyme preparation. The total volume was adjustedto 60 gl with H20, and the mixture was incubated by shakingfor 10 min at room temperature. Next, 30 Ml of protein A-Sepharose CL-4B (Pharmacia) was added to the mixture, whichwas then incubated for another 30 min. After incubation, themixture was centrifuged at 12,000g for 5 min, and the remainingPEPC activity in the supernatant fluid was determined.
RESULTSImmunodetection of PEPC in C3 Plants. Soluble protein ex-
tracts from green leaves of the C3 plants, wheat, barley, rice andtobacco, and green and etiolated leaves of maize (C4 plant), weresubjected to SDS-PAGE and analyzed by immunoblotting usinganti (maize) PEPC antibody (Fig. 1). While green leaves of maizeshowed one PEPC band (C4-type PEPC), two immunoreactiveproteins against the antibody were detected in wheat and barley,the upper one was relatively faint and migrated slightly moreslowly than maize PEPC while the lower one was stronger andmigrated as fast as maize PEPC. The monomeric mol wt ofmaize PEPC and the lower wheat one were calculated to be 100kD and that of the upper one to be 110 kD. In the case of rice,four bands were detected, the largest band was 1 5 kD and thesmallest 95 kD. With tobacco, only one band, with the samemol wt as maize PEPC (about 100 kD), was detected. In etiolatedmaize leaves, three immunoreactive proteins were detected, theupper one was very faint and migrated as fast as wheat upperone, and the lower bands were doublet and migrated as fast asC4-type PEPC. These proteins were considered to be a C3-typePEPC in maize (9). These results show that every gramineousplant tested has some C3-type PEPCs with different subunit sizes,but the dicotyledonous plant, tobacco, does not.
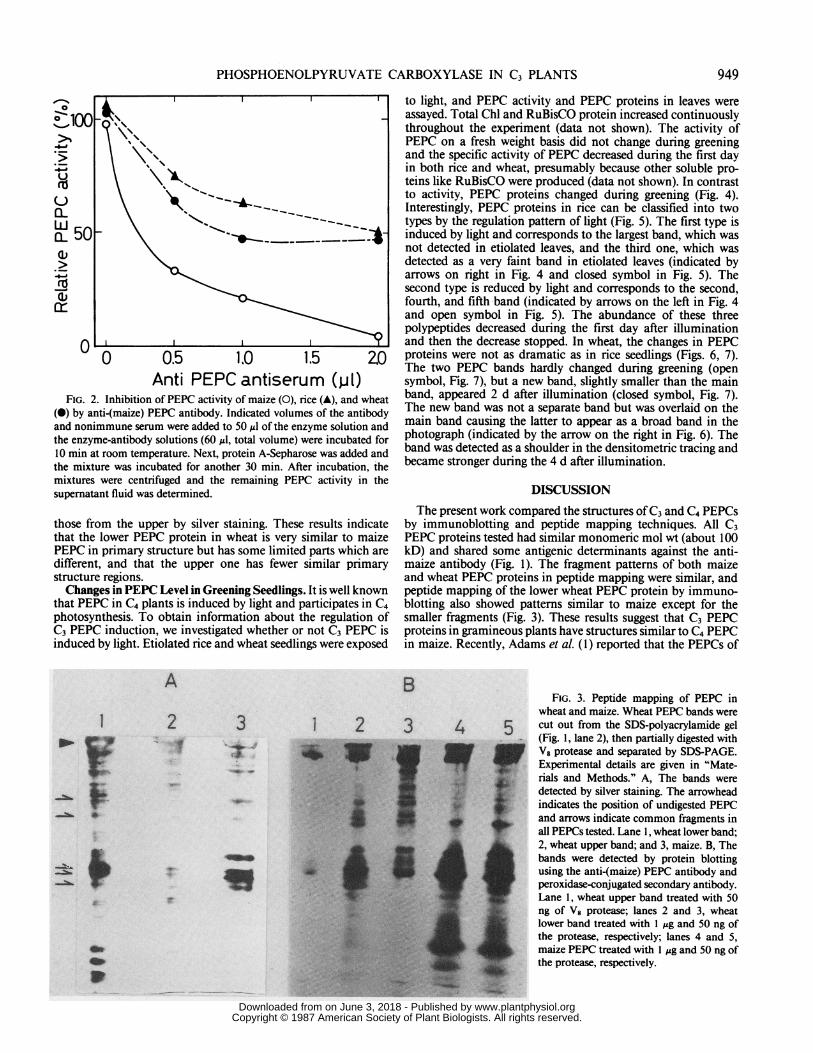
Immunoprecipitation of C3 PEPCs. To confirm the presenceof PEPC activity in the immunoreactive proteins in C3 plants,we used the antibody to check for precipitation of enzymeactivity. Increasing amounts of the antibody quantitatively pre-cipitated the maize enzyme and about half of the proteins withPEPC activities in rice and wheat (Fig. 2). These results showthat the immunoreactive proteins in C3 plants are active PEPCenzymes. It is not clear why half ofthe enzyme activity remainedin supernatant fluid; perhaps other PEPC(s) not recognized bythe antibody exist in C3 plants or PEPC-activities in the antibody.Comparison of C3 and C4 PEPC Structures. Peptide mapping
was carried out to compare the primary structures of the two
1 2 3 4 5 6__-FIG. 1. Immunodetection ofPEPC in C3 and C4 plants. Extracts from
plant leaves were subjected to SDS-PAGE, then transferred onto anitrocellulose sheet, and stained with anti-(maize) PEPC antibody andperoxidase-conjugated secondary antibody. Lane 1, extract from maizegreen leaves (1 ug of protein); 2, etiolated maize leaves (30 Mg); 3, wheat(30 ,g); 4, barley (30 Mg); 5, rice (30 Mg); and 6, tobacco (30 Ag).
PEPC proteins in wheat and maize PEPC. As shown in Figure3A, there were several common bands indicated by arrows in allthe proteins tested, but the three PEPC proteins had differentfragment patterns. This result suggests that the two immuno-reactive PEPC proteins in wheat have similar primary structuresto that of maize PEPC, but the wheat proteins have somedifferent regions from the maize one and different structuresbetween themselves. To further examine the similarities amongthese proteins, we examined the reactivities of the digested pep-tide fragments and the antibody (Fig. 3B). Almost all peptidefragments from maize PEPC were recognized by the antibody.In the case of the lower PEPC in wheat, many peptide fragmentswere also recognized and the pattern of immunoreactive frag-ments was similar to that of maize, although some small peptidefragments were less immunoreactive than those from the maizePEPC. On the other hand, few peptide fragments of the upperPEPC were recognized by the antibody and some, includinglarger ones, were not recognized. It is difficult to compare theimmunoreactivities of the upper and the lower PEPC because amuch larger amount of the lower PEPC was used in this experi-ment. However, we did find that some fragments digested fromthe upper PEPC were not recognized by the antibody whichrecognized fragments from the lower one stained as strongly as
948 Plant Physiol. Vol. 85, 1987
www.plantphysiol.orgon June 3, 2018 - Published by Downloaded from Copyright © 1987 American Society of Plant Biologists. All rights reserved.
PHOSPHOENOLPYRUVATE CARBOXYLASE IN C3 PLANTS
nv 0 0.5 1.0 1.5 ZO
Anti PEPC antiserum (p1)FIG. 2. Inhibition of PEPC activity of maize (0), rice (A), and wheat
(i) by anti-(maize) PEPC antibody. Indicated volumes of the antibodyand nonimmune serum were added to 50 ul of the enzyme solution andthe enzyme-antibody solutions (60 ,ul, total volume) were incubated for10 min at room temperature. Next, protein A-Sepharose was added andthe mixture was incubated for another 30 min. After incubation, themixtures were centrifuged and the remaining PEPC activity in thesupernatant fluid was determined.
those from the upper by silver staining. These results indicatethat the lower PEPC protein in wheat is very similar to maizePEPC in primary structure but has some limited parts which aredifferent, and that the upper one has fewer similar primarystructure regions.Changes in PEPC Level in Greening Seedlings. It is well known
that PEPC in C4 plants is induced by light and participates in C4photosynthesis. To obtain information about the regulation ofC3 PEPC induction, we investigated whether or not C3 PEPC isinduced by light. Etiolated rice and wheat seedlings were exposed
A
I
go..
I.
&s_
2 3It s
:
C
e*..
_*
_ _ol
a*
.0
to light, and PEPC activity and PEPC proteins in leaves wereassayed. Total Chl and RuBisCO protein increased continuouslythroughout the experiment (data not shown). The activity ofPEPC on a fresh weight basis did not change during greeningand the specific activity of PEPC decreased during the first dayin both rice and wheat, presumably because other soluble pro-teins like RuBisCO were produced (data not shown). In contrastto activity, PEPC proteins changed during greening (Fig. 4).Interestingly, PEPC proteins in rice can be classified into twotypes by the regulation pattern of light (Fig. 5). The first type isinduced by light and corresponds to the largest band, which wasnot detected in etiolated leaves, and the third one, which wasdetected as a very faint band in etiolated leaves (indicated byarrows on right in Fig. 4 and closed symbol in Fig. 5). Thesecond type is reduced by light and corresponds to the second,fourth, and fifth band (indicated by arrows on the left in Fig. 4and open symbol in Fig. 5). The abundance of these threepolypeptides decreased during the first day after illuminationand then the decrease stopped. In wheat, the changes in PEPCproteins were not as dramatic as in rice seedlings (Figs. 6, 7).The two PEPC bands hardly changed during greening (opensymbol, Fig. 7), but a new band, slightly smaller than the mainband, appeared 2 d after illumination (closed symbol, Fig. 7).The new band was not a separate band but was overlaid on themain band causing the latter to appear as a broad band in thephotograph (indicated by the arrow on the right in Fig. 6). Theband was detected as a shoulder in the densitometric tracing andbecame stronger during the 4 d after illumination.
DISCUSSIONThe present work compared the structures ofC3 and C4 PEPCs
by immunoblotting and peptide mapping techniques. All C3PEPC proteins tested had similar monomeric mol wt (about 100kD) and shared some antigenic determinants against the anti-maize antibody (Fig. 1). The fragment patterns of both maizeand wheat PEPC proteins in peptide mapping were similar, andpeptide mapping of the lower wheat PEPC protein by immuno-blotting also showed patterns similar to maize except for thesmaller fragments (Fig. 3). These results suggest that C3 PEPCproteins in gramineous plants have structures similar to C4 PEPCin maize. Recently, Adams et al. (1) reported that the PEPCs of
BFIG. 3. Peptide mapping of PEPC in
wheat and maize. Wheat PEPC bands were12 3 4 5 cut out from the SDS-polyacrylamide gel
(Fig. 1, lane 2), then partially digested with
rials and Methods." A, The bands weredetected by silver staining. The arrowhead
if s jF g indicates the position of undigested PEPC;
g ^ ~~~~~~~~and arrows indicate common fragments inall PEPCs tested. Lane 1, wheat lower band;2, wheat upper band; and 3, maize. B, The
fl̂_ _ bands were detected by protein blottingusing the anti-(maize) PEPC antibody andperoxidase-conjugated secondary antibody.Lane 1, wheat upper band treated with 50ng of V8 protease; lanes 2 and 3, wheatlower band treated with 1 gg and 50 ng ofthe protease, respectively; lanes 4 and 5,maize PEPC treated with I sg and 50 ng ofthe protease, respectively.
949
www.plantphysiol.orgon June 3, 2018 - Published by Downloaded from Copyright © 1987 American Society of Plant Biologists. All rights reserved.

MATSUOKA AND HATA
Illumination time (h)
O N @3s';1- CD " 'S
Illumination time (h)
(N N$IIQ cor- C~~~~NJ CCy) -
o CN .4(.0 tQOD
3.~.1
i£AC-FF -i
_ ew umo w Ol - _ _ a-.L-
FIG. 4. Changes in PEPC polypeptides during the greening of etiol-ated rice seedlings. Leaves (0.05 g fresh weight) were collected at theindicated times after the initiation of greening. Their extracts (30 jg ofprotein) were subjected to SDS-PAGE, transferred to a nitrocellulosesheet, and stained with the anti-maize) PEPC antibody and the peroxi-dase-conjugated secondary antibody. The arrows on the right or leftindicate bands induced or reduced by light, respectively.
%
0 24 48 72IUumination time (h)
FIG. 5. Time course of the amount of PEPC polypeptides during thegreening of etiolated rice seedlings. Data are taken from densitometrictracing ofthe immunoblotting in Figure 4. Ordinate, relative peak heightof the largest (A), second (v), third (0), fourth (0), and fifth band (A);abscissa, time after illumination.
FIG. 6. Changes in PEPC polypeptides during the greening of etiol-ated wheat seedlings. Experimental conditions are as given in Figure 4.
2.O
0 24 48 72Illumination time (h)
FIG. 7. Time course of the amount of PEPC polypeptides during thegreening of etiolated wheat seedlings. Data are taken from densitometrictracing ofthe immunoblotting in Figure 6. Ordinate, relative peak heightof the upper band (A), lower band (0), and the band induced by lightindicated by the arrow in Figure 6 (0).
C3 and C3-C4 intermediate species of Flaveria were markedlydifferent from C4 Flaveria PEPC in peptide mapping, while allthree PEPCs reacted with an anti-maize PEPC antibody. Al-though this observation conflicts our findings, it is possible thatthe homology among C3 and C4 PEPCs in Flaveria is less thanthat in gramineous plants.We also showed that PEPC proteins of slightly different sizes
exist in wheat, barley, rice, and etiolated maize, but not intobacco. The presence of multiple bands in the immunoblotting
950 Plant Physiol. Vol. 85, 1987
www.plantphysiol.orgon June 3, 2018 - Published by Downloaded from Copyright © 1987 American Society of Plant Biologists. All rights reserved.

PHOSPHOENOLPYRUVATE CARBOXYLASE IN C3 PLANTS
does not itself mean that there are PEPCs of different sizes in C3plants, because PEPC is often degraded by endogenous proteasesduring extraction, and the digested protein band shows slightlygreater mobility than the intact monomer (1). However, thePEPC proteins of different sizes that we observed were not likelycaused by partial degradation from the largest one because of thefollowing findings. First, the smaller bands in SDS-PAGE werenot eliminated by the addition of a protease inhibitor, like 1 mMphenylmethylsulfonyl fluoride or 5 mM EGTA, and a proteindenaturant, like 10% SDS, to the extraction buffer (data notshown). Second, the peptide maps of the upper and lower bandsfrom wheat were not identical (Fig. 3A) and peptide mapping byimmunoblotting also showed differences (Fig. 3B). Finally, thesePEPC proteins changed individually in greening leaves of riceand wheat (Figs. 4-7). There have been few papers reporting theexistence of PEPCs of different sizes in maize green leaves (6)and in C3 and C3-C4 intermediate Flaveria species (1). However,we found only one PEPC band in maize green leaves and in theC3 dicotyledonous plant, tobacco, by immunoblotting. Recently,Budde and Chollet (4) detected only one PEPC band in maizegreen leaves by two-dimensional gel and immunoblotting. Thisfinding corresponds to our result in maize green leaves. On theother hand, Huber et aL (10) detected only one PEPC band inetiolated maize leaves by immunoblotting. This finding does notcorrespond to our result in etiolated maize leaves. It is possiblethat they loaded small amount of PEPC protein on a gel andcould not detect multiple PEPC bands. Actually, more recently,they found different sizes of PEPCs in etiolated maize leaves (TSugiyama, personal communication).
In order to compare the regulation of PEPC expression, wedetermined whether C3 PEPCs are induced by light as in C4plants. During the greening of etiolated rice and wheat seedlings,PEPC activity remained constant and the specific activity de-creased rapidly, presumably due to the induction ofother solubleleaf proteins. On the other hand, PEPC proteins detected byimmunoblotting changes during the greening of rice and wheatseedlings. The PEPC proteins in rice could be classified into twotypes, one induced and the other reduced by light (Fig. 5). Theseresults show that there are two types of PEPC expression in rice,one resembling that of PEPC in C4 plants, which is induced bylight (9, 14). Greening wheat seedlings also had one band inducedby light (Fig. 6). Similar results were obtained for another C4-photosynthetic enzyme, PPDK. This enzyme in rice has a struc-ture similar to that in maize and is also induced by light (8).These results suggest that there may exist similar genes for C4
photosynthetic enzymes for not only structural regions but alsoregulatory regions in C3 gramineous plants.
LITERATURE CITED
1. ADAMS CA, F LEUNG, SSM SUN 1986 Molecular properties of phosphoenol-pyruvate carboxylase from C3, C3-C4 intermediate, and C4 Flaveria species.Plant 167: 218-225
2. AMAGASA T 1984 Enzymatic properties of phosphoenolpyruvate carboxylaseisoforms in the CAM plant Kalanchoe daigremontiana. Plant Cell Physiol25: 625-633
3. BROGLIE R, G CORUZZ7, B KEITH, N-H CHUA 1984 Molecular biology of C4photosynthesis in Zea mays: differential localization ofproteins and mRNAsin the two leaf cell types. Plant Mol Biol 3: 431-444
4. BUDDE RJA, R CHOLLET 1986 In vitro phosphorylation of maize leaf phos-phoenolpyruvate carboxylase. Plant Physiol 82: 1107-1114
5. CLEVELAND DW, SG FISHER, MW KIRSCHNER, UK LAEMMLI 1977 Peptidemapping by limited proteolysis in sodium dodecyl sulfate and analysis bygel electrophoresis. J Biol Chem 252: 1102-1106
6. HAGUE DR, TL SIMS 1980 Evidence for light-stimulated synthesis of phos-phoenolpyruvate carboxylase in leaves of maize. Plant Physiol 66: 505-509
7. HARPSTER MH, WC TAYLOR 1986 Maize phosphoenolpyruvate carboxylase:cloning and characterization of mRNAs encoding isozymic forms. J BiolChem 261: 6132-6136
8. HATA S, M MATSUOKA 1987 Immunological studies on pyruvate orthophos-phate dikinase in C3 plants. Plant Cell Physiol 28: 635-641
9. HAYAKAWA S, K MATSUNAGA, T SUGIYAMA 1981 Light induction of phos-phoenolpyruvate carboxylase in etiolated maize leaf tissue. Plant Physiol 67:133-138
10. HUBER SC, T SUGIYAMA, T AKAZAWA 1986 Light modulation of maize leafphosphoenolpyruvate carboxylase. Plant Physiol 82: 550-554
1 1. HUDSPETH R, CA GLACKIN, J BONNER, JW GRULA 1986 Genomic and cDNAclones for maize phosphoenolpyruvate carboxylase and pyruvate, ortho-phosphate dikinase: expression of different gene-family members in leavesand roots. Proc Natl Acad Sci USA 83: 2884-2888
12. IzUl K, S ISHUIMA, Y YAMAGUCHI, F KATAGIRI, T MURATA, K SHIGESADA, TSUGIYAMA, H KATSUKI 1986 Cloning and sequence analysis of cDNAencoding active phosphoenolpyruvate carboxylase of the C4-pathway frommaize. Nucleic Acids Res 14: 1615-1628
13. LAEMMLI UK 1970 Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227: 680-685
14. MATSUOKA M, T ASAHI 1983 Mechanism of the increase in cytochrome coxidase activity in pea cotyledons during seed hydration. Eur J Biochem134: 223-229
15. NELSON T, MH HARPSTER, SP MAYFIELD, WC TAYLOR 1984 Light-regulatedgene expression during maize leaf development. J Cell Biol 98: 558-564
16. OAKLEY BR, DR KIRSCH, NR MORRIS 1980 A simplified ultrasensitive silverstain for detecting proteins in polyacrylamide gels. Anal Biochem 105: 361-363
17. O'LEARY MH 1982 Phosphoenolpyruvate carboxylase: an enzymologist's view.Annu Rev Plant Physiol 33: 279-315
18. SIMs TL, DR HAGUE 1981 Light-stimulated increase of translatable mRNAfor phosphoenolpyruvate carboxylase in leaves of maize. J Biol Chem 256:8252-8255
19. UEDAN K, T SUGIYAMA 1976 Purification and characterization of phosphoen-olpyruvate carboxylase from maize leaves. Plant Physiol 57: 906-910
951
www.plantphysiol.orgon June 3, 2018 - Published by Downloaded from Copyright © 1987 American Society of Plant Biologists. All rights reserved.