comparative evaluation and selection of a method for lipid and fatty acid extraction from macroalgae
TRANSCRIPT

Analytical Biochemistry 415 (2011) 134–144
Contents lists available at ScienceDirect
Analytical Biochemistry
journal homepage: www.elsevier .com/locate /yabio
Comparative evaluation and selection of a method for lipid and fatty acidextraction from macroalgae
Puja Kumari, C.R.K. Reddy ⇑, Bhavanath JhaDiscipline of Marine Biotechnology and Ecology, Central Salt and Marine Chemicals Research Institute, Council of Scientific and Industrial Research (CSIR), Bhavnagar 364021,Gujarat, India
a r t i c l e i n f o
Article history:Received 27 January 2011Received in revised form 16 March 2011Accepted 6 April 2011Available online 16 April 2011
Keywords:LipidsFatty acidsMacroalgaeSonicationBufferExtraction method
0003-2697/$ - see front matter � 2011 Elsevier Inc. Adoi:10.1016/j.ab.2011.04.010
⇑ Corresponding author. Fax: +91 278 2567562/256E-mail address: [email protected] (C.R.K. Reddy).
1 Abbreviations used: BD, Bligh and Dyer; BDB, BligBligh and Dyer with sonication; CM, conventional mmethod; CSB, Cequier-Sánchez method with buffer; Cwith sonication; DT, direct transesterification; FA, fattyesters; FM, Folch method; FMB, Folch method with busonication; GM, Garcia method; LRC, Lepage and RoyPUFAs, polyunsaturated fatty acids; TFA, total fatty afatty acids; SFA, saturated fatty acid.
a b s t r a c t
A comparative evaluation of Bligh and Dyer, Folch, and Cequier-Sánchez methods for quantitative deter-mination of total lipids (TLs) and fatty acids (FAs) was accomplished in selective green (Ulva fasciata), red(Gracilaria corticata), and brown algae (Sargassum tenerrimum) using a full factorial categorical design.Applications of sonication and buffer individually on lipid extraction solvent systems were also evalu-ated. The FA recoveries obtained from the aforementioned methods were compared with those of directtransesterification (DT) methods to identify the best extraction methods. The experimental designshowed that macroalgal matrix, extraction method, and buffer were key determinants for TL and FArecoveries (P 6 0.05), exhibiting significant interactions. But sonication gave erratic results with no inter-action with any of the factors investigated. The buffered solvent system of Folch rendered the highest TLyield in U. fasciata and G. corticata while the buffered system of Bligh and Dyer gave the highest yield in S.tenerrimum. DT methods were more convenient and accurate for FA quantification and rendered 1.5–2times higher yields when compared with the best conventional method, minimizing the use of chlori-nated solvents, their cost of analysis, and disposal. The buffered solvent system was found to be the mostappropriate for lipid research in macroalgae.
� 2011 Elsevier Inc. All rights reserved.
Macroalgae have been reported to contain more than 2400 nat-ural products of commercial importance in pharmaceutical, bio-medical, and nutraceutical industries [1]. They have also beenextensively utilized as ingredients in human and animal food prep-arations owing to their high contents of polyunsaturated fattyacids (PUFAs),1 carbohydrates, vitamins, minerals, and dietary fibers[2,3]. Nowadays, algal resources have been studied with renewedinterest across the world as an alternative source of renewable en-ergy feedstock that circumvents the controversy of ‘‘fuel versusfood’’. The attributes for such choice are their relatively higher pro-duction turnover and amenability for depolymerization of substratein addition to greater carbon sequestration potentials than terrestrialfeedstock [4]. Most recently, Petcavich succeeded in producinghydrocarbon biofuels from transformed kelp Macrocystis pyrifera
ll rights reserved.
6970.
h and Dyer with buffer; BDS,ethods; CS, Cequier-SánchezSS, Cequier-Sánchez method
acid; FAMEs, fatty acid methylffer; FMS, Folch method withmodified by Cohen method;
cid; MUFA, monounsaturated
with high hydrocarbon producing genes of microalgae, Botryococcusbraunii [5].
Fatty acid (FA) analysis has been increasingly gaining impor-tance due to the realization of their beneficial applications in nutri-tional and health products. Further, they have also been used foraddressing various fundamental and pragmatic research problemsin experimental biochemical, physiological, and clinical studies[6,7]. Further, in biodiesel production, clean burn properties ofthe fuel are influenced by FA structural features including chainlength and degree of unsaturation [8]. Thus, a precise quantifica-tion of FA can also be used to predict the quality of biodiesel, whichis reduced considerably with the increase in the amount of satu-rated FAs.
Traditionally, the fatty acid composition of lipid samples isdetermined by assessing the corresponding methyl esters via gaschromatography (GC). A large number of analytical approachesbased on initial lipid extraction by solvents, followed by theirtransmethylation (i.e., conventional methods), are employed andwhere FAs are sought, they are extracted and methylated withone-step procedures wherein methylation reagent is added di-rectly to the samples without previous extraction (direct transeste-rification methods). However, both types of methods have theirown advantages and disadvantages that are well illustrated inthe literature [9–16]. Conventional methods are time consuming

Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144 135
and use toxic solvents such as chloroform and benzene while directtransesterification methods are simplified, rapid, and sensitive,minimize the use of solvents, and give better recoveries of FAs. De-spite several disadvantages, conventional methods of lipid extrac-tion are the only means to study different lipid classes to date [9].
As far as macroalgae are concerned, they have been extensivelyexplored for their novel lipids and fatty acids [17–20]. Althoughboth the conventional and the direct transesterification methodshave been employed by researchers to study their lipids and FAprofiles [3,21–24], Bligh and Dyer [25] and Folch methods [26]based on chloroform/methanol solvent systems have been invari-ably used as standard methods. However, other solvents such asdichloromethane/methanol [27] and diethyl ether [28] have alsobeen used, but there are no established criteria for choosing themost appropriate one. A researcher must be intelligent enough tobe able to choose the best and ignore the rest. Moreover, informa-tion regarding comparison and verification is still required to testwhich methods of extraction and FA derivatization function bestfor these matrices. Since, the determination of FAs in macroalgaeis itself a challenge because of their heterogeneous FAs and highcontents of PUFAs, it becomes imperative to study extraction andderivatization procedures to obtain accurate, qualitative, andquantitative results.
In view of the growing demand of macroalgae in the develop-ment of PUFA-related dietary supplements and lipid-based fuels,a comparison of different lipid and fatty acid extraction and deriv-atization methods was accomplished in three different macroalgalmatrices representing one each from Chlorophyta, Phaeophyta, andRhodophyta. The purpose of the present work was to systemati-cally validate, by a full factorial categorical design, the efficacy ofthree most commonly used solvent systems, viz. chloroform/meth-anol (1/2 and 2/1, v/v) and dichloromethane/methanol (2/1, v/v) onlipid extraction and thereafter the effect of sonication and bufferon the respective solvent systems. Further, the fatty acid recoveriesobtained by the conventional methods were also compared withone-step direct transesterification methods in order to select thebest lipid and FA extraction method for different macroalgalmatrices.
Materials and methods
Chemicals
For the identification and quantification of fatty acids, the fol-lowing analytical grade standards were used: 37-component FAMEmix C4–C24 (Supelco, USA), 7-hexadecenoic acid methyl ester(C16:1, n7), and stearidonic acid methyl ester (C18:4, n3) (CaymanChemicals, USA). The internal standard nonadecanoic acid (C19:0)was purchased from Sigma. All the solvents used (such as chloro-form, methanol, dichloromethane, and hexane) were of HPLC gradeand other reagents of analytical grade.
Algal samples
Three macroalgal samples representing three different phyla,viz. Ulva fasciata (Chlorophyta), Gracilaria corticata (Rhodophyta),and Sargassum tenerrimum (Phaeophyta) were collected in March2009 from Veraval coast, Gujarat, India, and transported to the lab-oratory in wet tissue towels in an ice box. They were immediatelycleaned thoroughly to remove the epiphytes and other undesiredforeign matter from the fronds, frozen in liquid nitrogen, andstored at �40 �C until the analysis commenced.
Conventional methods (CM)
The lipid extraction efficiency of Bligh and Dyer [25], Folch et al.[26], and Cequer-Sànchez method [29] was compared in the pres-ent study. Further, the extraction efficacy of each method by appli-cation of sonication and buffer individually was also investigatedas a part of the optimization. The stepwise experimental proce-dures including modifications are detailed in a flowchart diagramin Fig. 1. All the analyses were done with 500 mg of ground algaltissues. Butylated hydroxyl toluene (BHT) at 0.001% was added toeach solvent system to minimize lipid peroxidation during thesample preparation.
Method 1: Bligh and Dyer method [25]1A (BD). Macroalgal samples were extracted with 3 ml of chloro-form/methanol (1/2, v/v) by vortexing (1 min) and centrifugationat 2057g for 15 min at room temperature (RT). The supernatantswere collected and residues reextracted thrice with 2 ml of chloro-form/methanol (1/1, v/v) by centrifugation as stated above. All thesupernatants were pooled together, filtered with Whatman filterNo. 1 (Whatman, USA), and washed with 2 ml of Milli-Q water, fol-lowed by centrifugation at 2057g for 5 min. The lower organicphases were collected and evaporated to dryness under nitrogenand total lipid contents were determined gravimetrically.
1B (BDS; Bligh and Dyer with sonication). The lipids were extractedas noted above for Method 1A, with slight modification; the tissueswere sonicated for 2 min on ice in a cup sonicator, Branson 2210(USA) for cell disruption in the extraction medium prior tocentrifugation.
1C (BDB; Bligh and Dyer with buffer). The lipids were extracted with3 ml of chloroform/methanol/50 mM phosphate buffer (1/2/0.8, v/v/v) by vortexing (1 min) and centrifugation at 2057g for 15 min atRT. The supernatants were collected and residues reextractedthrice with 2 ml chloroform/methanol/buffer (1/1/0.8, v/v/v) bycentrifugation as stated above. All the supernatants were com-bined, filtered, and washed with 2 ml of 50 mM phosphate buffer,followed by centrifugation at 2057g for 5 min. The lower organicphases were collected and evaporated to dryness under nitrogenand total lipid contents were determined gravimetrically.
Method 2: Folch method [26]2A (FM). Lipids were extracted with 3 ml of chloroform/methanol(2/1, v/v) by vortexing and centrifugation as described for Method1A. The supernatants were collected, residues were reextractedthrice with 2 ml chloroform/methanol (1/1, v/v) by centrifugation,and combined supernatants were washed with 2 ml of Milli-Qwater. The lower organic phases were collected after centrifuga-tion at 2057g for 5 min at RT; thereafter, evaporated to dryness un-der nitrogen; and total lipid contents were determinedgravimetrically.
2B (FMS; Folch method with sonication). The lipids were extractedas noted above for Method 2A, with the application of sonicationfor 2 min on ice for cell disruption in the extraction medium priorto centrifugation.
2C (FMB; Folch method with buffer). In this method, buffer wasadded to the extraction solvent mixture of Method 2A [26] andthus the tissues were extracted initially with chloroform/metha-nol/50 mM phosphate buffer (2/1/0.8, v/v/v), otherwise all thesteps were the same as in Method 1C.

Fig.1. Flow diagram of different lipid and FA extraction methods in macroalgae. Note: BD, Bligh and Dyer; BDS, Bligh and Dyer with sonication; BDB, Bligh and Dyer withbuffer; FM, Folch method; FMS, Folch method with sonication; FMB, Folch method with buffer; CS, Cequier-Sánchez method; CSS, Cequier-Sánchez method with sonication;CSB, Cequier-Sánchez method with buffer; GM, Garcia method; LRC, Lepage and Roy modified by Cohen method; C, Chloroform; M, Methanol; D, Dichloromethane.
136 Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144
Method 3: Cequier-Sánchez method [29]3A (CS). Lipids were extracted with 3 ml of dichloromethane/methanol (2/1) by vortexing (1 min) and centrifugation at 2057gfor 15 min at RT. Again, the supernatants were collected and resi-dues reextracted thrice with 2 ml dichloromethane/methanol (1/1). All the supernatants were combined, filtered, and washed with2 ml of Milli-Q water and centrifuged at 2057g for 5 min. The lowerorganic phases were collected, evaporated to dryness under nitro-gen, and total lipid contents were determined gravimetrically.
3B (CSS; Cequier-Sánchez with sonication). The lipids and FAs wereextracted as noted above for Method 3A (CS), with the applicationof sonication to tissues for 2 min on ice, for cell disruption in theextraction medium prior to centrifugation.
3C (CSB; Cequier-Sánchez with buffer). In this method, buffer wasadded to the extraction solvent mixture of Method 3A (CS) andthus, the tissues were extracted initially with dichloromethane/methanol/50 mM phosphate buffer (2/1/0.8, v/v/v), otherwise allthe steps were the same as in Method 1C.
Preparation of FAMEs
The fatty acids were converted to their fatty acid methyl esters(FAMEs) by transmethylation of lipid samples thus obtained by sol-vent extraction, with 1 ml of 1% NaOH in MeOH, followed by heat-ing for 15 min at 55 �C, adding 2 ml of 5% methanolic HCl and againheated for 15 min at 55 �C and then adding 1 ml Milli-Q water. Tenmicroliters of 1 mg/ml nonadecanoic acid (C19:0) was spiked intothe lipid samples as internal standard. FAMEs were extracted byhexane (3 � 1 ml) and evaporated to dryness under nitrogen [30].
They were redissolved in 200 ll of hexane and stored in �40 �Cin glass vials until analyzed by GC–MS.
Direct transesterification (DT) methods
Two direct transesterification methods were also applied tomacroalgal samples whereby FAs were directly extracted fromthe tissues without initial lipid extraction as described below.
Method 4: Garcia method (GM) [31]Tissues (500 mg) were treated with 2 ml of toluene and 3 ml of
freshly prepared 5% methanolic HCl, spiked with internal standard,nonadecanoic acid (10 ll of 1 mg/ml), mixed thoroughly by vor-texing in oak ridge tubes, closed under nitrogen, and heated for2 h in a water bath at 70 �C. After cooling to room temperature,4 ml of 6% aqueous K2CO3 and 2 ml of toluene were added andthe mixtures were vortexed and centrifuged for 5 min at 2057g.The organic phases were drawn off and dried with anhydrousNa2SO4 and solvents were removed under nitrogen. FAMEs weresolubilized in 200 ll of toluene and analyzed by GC–MS.
Method 5: Lepage and Roy (LRC) [32] as modified by Cohen et al. [33]Tissues (500 mg) were treated with 5 ml of acetyl chloride/
methanol reagent (1/19; v/v), spiked with internal standard, nona-decanoic acid (10 ll of 1 mg/ml), and esterified at 80 �C for 1 h.After cooling, 1 ml of water and 2 ml of n-hexane were added tothe mixture, vortexed, and centrifuged at 2057g for 5 min. The or-ganic phases were collected, filtered, and dried with anhydrous so-dium sulfate. Solvents were removed under nitrogen and themethyl esters solubilized in 200 ll of n-hexane for GC–MS analysis.

Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144 137
GC–MS analysis
The GC–MS analysis of FAME samples was carried out on aQP2010 gas chromatography mass spectrometer (GC-2010 coupledwith GC–MS QP-2010) equipped with an autosampler (AOC-5000)from Shimadzu (Japan) using a RTX-5 fused silica capillary column,30 m � 0.25 mm � 0.25 lm (Rastek). Helium (99.9% purity) wasused as the carrier gas with a column flow rate of 1 ml/min anda precolumn pressure of 49.7 kPa. The column temperature regimewas 40 �C for 3 min, followed by a 5 �C/min ramp up to 230 �C, fol-lowed by 40 min at 230 �C. The injection volume and temperaturewere 0.2 ll and 240 �C and the split ratio was 1/30. The mass spec-trometer was operated in electron compact mode with an electronenergy of 70 eV. Both the ion source temperature and the interfacetemperature were set at 200 �C. FAME peaks were identified bycomparison of their retention times with authentic standards byGC–MS post run analysis and quantified by area normalization.
Statistical analysis and experimental design
All the analytical determinations were performed in triplicateand the mean values were reported. The yields of fatty acids andtotal lipids obtained by different methods for each macroalgalsamples were compared by analysis of variance (ANOVA), with val-ues significant at P 6 0.05.
Further, the effects of different solvent mixtures and modifica-tions applied on the quantity of TL and FAMEs obtained via con-ventional methods were analyzed by experimental design. Thestudied factors were type of macroalgal matrix (A), with three lev-els (Chlorophyta, Phaeophyta, and Rhodophyta); extraction meth-od (B), with three levels (BD, FM, and CS), sonication (C), withtwo levels 0 (=absent) and 1 (=present) and buffer (D), with twolevels 0 (=absent) and 1 (=present). A full factorial categorical de-sign with {3 � 3 � 2 � 2} 36 runs was selected to study both themain effects and the interactions. This type of design consists ofall levels of combinations of two or more nonquantitative factors,where the user sets the number of levels. The response variablesanalyzed were the amount of total lipids (TLs) and total FAs (TFAs)expressed as the sum of the concentrations (mg/g f.w.) of all iden-tified FAs; three other dependent variables calculated as the sum ofthe contents of the FAs belonging to the same structural class, i.e.,saturated FAs (RSFAs), monounsaturated FAs (RMUFAs), and poly-unsaturated FAs (RPUFAs) (expressed in lg/g f.w.). The results ob-tained were again evaluated by analysis of variance, whichmeasures whether a factor contributes significantly to the varianceof the response. The experimental design and the data analysiswere performed using Unscrambler version 9.8 (Camo, USA).
Results
The TL and TFA contents of the three macroalgae studied in thisstudy are shown in Fig. 2 and the fatty acid profiles are recorded inTables 1–3. The three macroalgae exhibited the typical profiles oftheir respective phyla, Ulva fasciata being rich in C18 PUFAs, G. cor-ticata in C20 PUFAs, and S. tenerrimum in both. As expected, U. fas-ciata reported the highest TL and TFA contents followed by S.tenerrimum and G. corticata.
The efficiency of TL and TFA extraction varied with the macro-algal matrices and extraction methods. The addition of buffer tothe respective solvent systems also rendered significantly differentyields while sonication displayed nonsignificant effects (Table 4).The contents of SFAs, MUFAs, and PUFAs were also greatly affectedby these three factors except extraction method, which was foundnonsignificant for MUFAs. Besides determining the influence ofeach factor on TL and FAs, second-order interactions were also
evaluated (Table 4). A significant interaction between extractionmethod and buffer was found for all the variables studied exceptfor TL which largely depended on the interaction of macroalgal ma-trix and extraction method. Further, all the interaction effects weremore conspicuous for PUFA yield showing significant positiveinteractions except for sonication which showed no significantinteraction with any factor.
The modified Folch method, FMB (Method 2C) gave the highestTL yield in U. fasciata and G. corticata while the modified Bligh andDyer method, BDB (Method 1C) rendered the highest TL yield in S.tenerrimum (Fig. 1). However, the differences between the threeconventional methods were larger and the simple Folch method(Method 2A) gave higher TL contents than the simple Bligh andDyer method (Method1A) and Cequier-Sánchez method (Method3A) in U. fasciata (2.3-fold and 3.3-fold, respectively), G. corticata(4-fold and 1.5-fold, respectively), and S. tenerrimum (2.7-foldand 1.8-fold, respectively). Further, the application of sonicationto three established conventional methods increased the TL yieldsin all the three matrices except FMS, whereby the TL yields de-creased considerably in the three matrices. The addition of bufferalso improved the lipid extraction in macroalgae except in S. ten-errimum where both the FMB and the CSB methods failed to in-crease the efficacy of lipid extraction.
Similarly, the amounts of TFAs and different FA groups werealso affected by the modifications employed in the present study.Method 1C (BDB) demonstrated the highest TFAs, SFAs, MUFAs,and PUFAs yields unanimously for the three different macroalgalmatrices, among the three conventional methods investigatedprior to transmethylation by the Carreau and Dubacq method[30]. The application of sonication and buffer individually to theBD solvent system significantly increased the yields of TFAs, SFAs,MUFAs, and PUFAs in the macroalgal matrices except for MUFAscontent in S. tenerrimum. In contrast, their addition to the FM sol-vent system (Method 2B and Method 2C) considerably decreasedthe recovery of fatty acids while the CS system showed erratic re-sults. The application of sonication with the CS solvent mixture(Method 3B) caused a decrease in FAs yields, whereas buffer(Method 3C) oppositely increased the yield in macroalgae studiedexcept in S. tenerrimum.
Compared with DT methods, the conventional methods for FAextraction prior to transmethylation yielded significantly lessFAs. The LRC (Method 6) rendered the highest yields in U. fasciataabout 2 times higher TFA, SFA, and PUFA and 1.1 times MUFA con-tents than the most efficient BDB (Method 3) while GM (Method 5)was more efficient for G. corticata and S. tenerrimum, both showing1.5–2 times higher yields. However, LRC (Method 6) was the onlyexception in G. corticata and S. tenerrimum to give much lowerFAs contents than BDB (Method 1C).
Discussion
Reliable methods for quantitative extraction of lipids and FAsare of paramount importance owing to their biochemical, physio-logical, clinical, and nutritional applications. The accuracy of differ-ent lipid extraction methods depends on the solubility of theirconstituent lipid classes in the solvents employed and the natureof sample matrix as both could influence the extent of lipid extrac-tion. According to Christie [34], extraction solvents/mixturesshould be polar enough to remove lipids from their associating cellconstituents, but not too polar that the solvents do not readily sol-ubilize all the triacylglycerols (TAGs) and other nonpolar lipids.
The present study also displayed significant variations in lipidand FA contents, owing to the matrix effect and solubilizing abilityof different solvent systems employed to completely disrupt thecellular membranes and dissolve the entrapped lipids from macro-
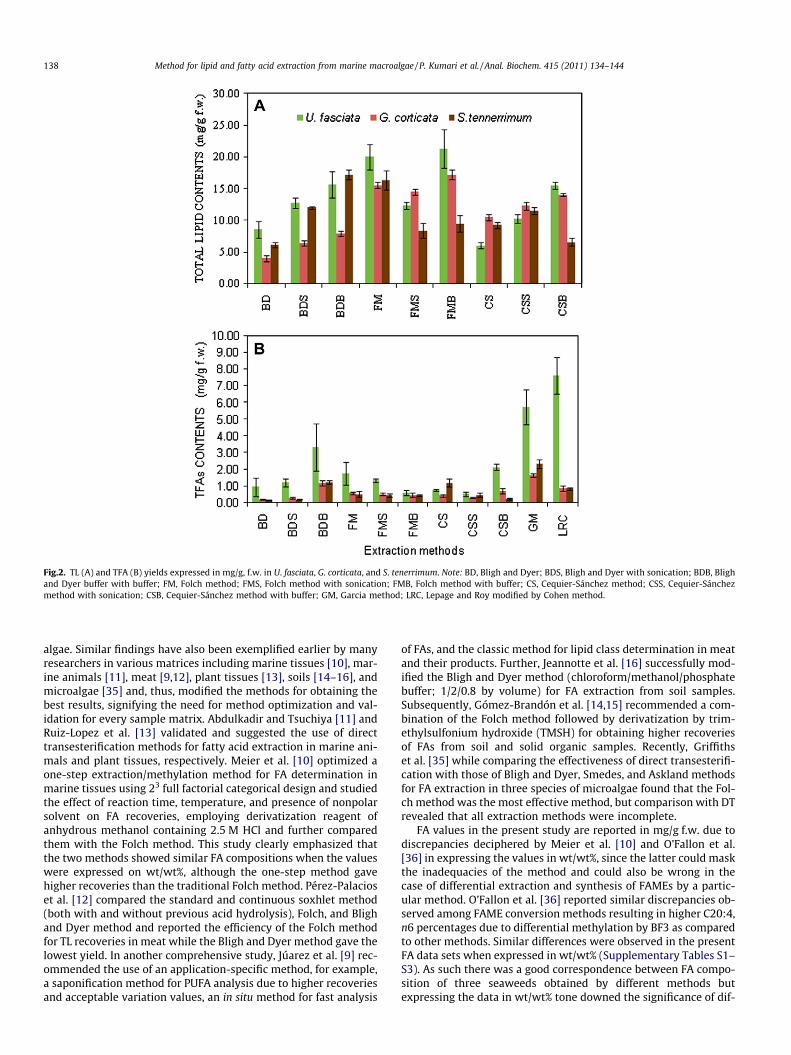
Fig.2. TL (A) and TFA (B) yields expressed in mg/g, f.w. in U. fasciata, G. corticata, and S. tenerrimum. Note: BD, Bligh and Dyer; BDS, Bligh and Dyer with sonication; BDB, Blighand Dyer buffer with buffer; FM, Folch method; FMS, Folch method with sonication; FMB, Folch method with buffer; CS, Cequier-Sánchez method; CSS, Cequier-Sánchezmethod with sonication; CSB, Cequier-Sánchez method with buffer; GM, Garcia method; LRC, Lepage and Roy modified by Cohen method.
138 Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144
algae. Similar findings have also been exemplified earlier by manyresearchers in various matrices including marine tissues [10], mar-ine animals [11], meat [9,12], plant tissues [13], soils [14–16], andmicroalgae [35] and, thus, modified the methods for obtaining thebest results, signifying the need for method optimization and val-idation for every sample matrix. Abdulkadir and Tsuchiya [11] andRuiz-Lopez et al. [13] validated and suggested the use of directtransesterification methods for fatty acid extraction in marine ani-mals and plant tissues, respectively. Meier et al. [10] optimized aone-step extraction/methylation method for FA determination inmarine tissues using 23 full factorial categorical design and studiedthe effect of reaction time, temperature, and presence of nonpolarsolvent on FA recoveries, employing derivatization reagent ofanhydrous methanol containing 2.5 M HCl and further comparedthem with the Folch method. This study clearly emphasized thatthe two methods showed similar FA compositions when the valueswere expressed on wt/wt%, although the one-step method gavehigher recoveries than the traditional Folch method. Pérez-Palacioset al. [12] compared the standard and continuous soxhlet method(both with and without previous acid hydrolysis), Folch, and Blighand Dyer method and reported the efficiency of the Folch methodfor TL recoveries in meat while the Bligh and Dyer method gave thelowest yield. In another comprehensive study, Júarez et al. [9] rec-ommended the use of an application-specific method, for example,a saponification method for PUFA analysis due to higher recoveriesand acceptable variation values, an in situ method for fast analysis
of FAs, and the classic method for lipid class determination in meatand their products. Further, Jeannotte et al. [16] successfully mod-ified the Bligh and Dyer method (chloroform/methanol/phosphatebuffer; 1/2/0.8 by volume) for FA extraction from soil samples.Subsequently, Gómez-Brandón et al. [14,15] recommended a com-bination of the Folch method followed by derivatization by trim-ethylsulfonium hydroxide (TMSH) for obtaining higher recoveriesof FAs from soil and solid organic samples. Recently, Griffithset al. [35] while comparing the effectiveness of direct transesterifi-cation with those of Bligh and Dyer, Smedes, and Askland methodsfor FA extraction in three species of microalgae found that the Fol-ch method was the most effective method, but comparison with DTrevealed that all extraction methods were incomplete.
FA values in the present study are reported in mg/g f.w. due todiscrepancies deciphered by Meier et al. [10] and O’Fallon et al.[36] in expressing the values in wt/wt%, since the latter could maskthe inadequacies of the method and could also be wrong in thecase of differential extraction and synthesis of FAMEs by a partic-ular method. O’Fallon et al. [36] reported similar discrepancies ob-served among FAME conversion methods resulting in higher C20:4,n6 percentages due to differential methylation by BF3 as comparedto other methods. Similar differences were observed in the presentFA data sets when expressed in wt/wt% (Supplementary Tables S1–S3). As such there was a good correspondence between FA compo-sition of three seaweeds obtained by different methods butexpressing the data in wt/wt% tone downed the significance of dif-
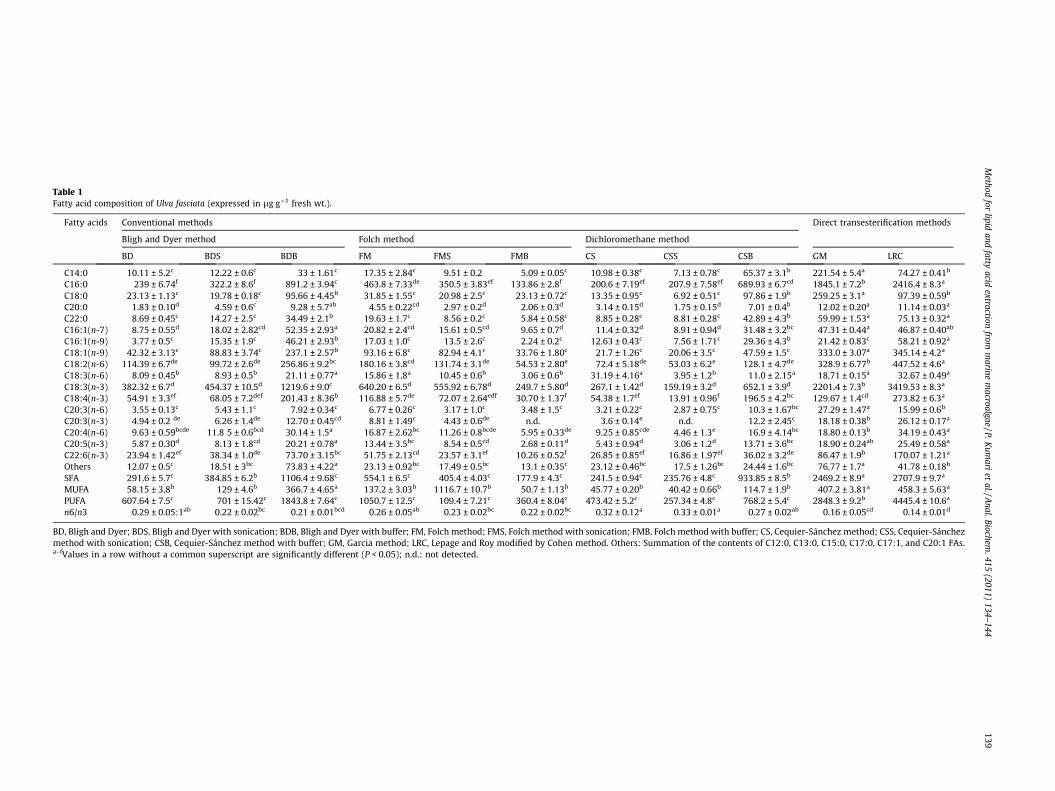
Table 1Fatty acid composition of Ulva fasciata (expressed in lg g�1 fresh wt.).
Fatty acids Conventional methods Direct transesterification methods
Bligh and Dyer method Folch method Dichloromethane method
BD BDS BDB FM FMS FMB CS CSS CSB GM LRC
C14:0 10.11 ± 5.2c 12.22 ± 0.6c 33 ± 1.61c 17.35 ± 2.84c 9.51 ± 0.2 5.09 ± 0.05c 10.98 ± 0.38c 7.13 ± 0.78c 65.37 ± 3.1b 221.54 ± 5.4a 74.27 ± 0.41b
C16:0 239 ± 6.74f 322.2 ± 8.6f 891.2 ± 3.94c 463.8 ± 7.33de 350.5 ± 3.83ef 133.86 ± 2.8f 200.6 ± 7.19ef 207.9 ± 7.58ef 689.93 ± 6.7cd 1845.1 ± 7.2b 2416.4 ± 8.3a
C18:0 23.13 ± 1.13c 19.78 ± 0.18c 95.66 ± 4.45b 31.85 ± 1.55c 20.98 ± 2.5c 23.13 ± 0.72c 13.35 ± 0.95c 6.92 ± 0.51c 97.86 ± 1.9b 259.25 ± 3.1a 97.39 ± 0.59b
C20:0 1.83 ± 0.10d 4.59 ± 0.6c 9.28 ± 5.7ab 4.55 ± 0.22cd 2.97 ± 0.2d 2.06 ± 0.3d 3.14 ± 0.15d 1.75 ± 0.15d 7.01 ± 0.4b 12.02 ± 0.20a 11.14 ± 0.03a
C22:0 8.69 ± 0.45c 14.27 ± 2.5c 34.49 ± 2.1b 19.63 ± 1.7c 8.56 ± 0.2c 5.84 ± 0.58c 8.85 ± 0.28c 8.81 ± 0.28c 42.89 ± 4.3b 59.99 ± 1.53a 75.13 ± 0.32a
C16:1(n-7) 8.75 ± 0.55d 18.02 ± 2.82cd 52.35 ± 2.93a 20.82 ± 2.4cd 15.61 ± 0.5cd 9.65 ± 0.7d 11.4 ± 0.32d 8.91 ± 0.94d 31.48 ± 3.2bc 47.31 ± 0.44a 46.87 ± 0.40ab
C16:1(n-9) 3.77 ± 0.5c 15.35 ± 1.9c 46.21 ± 2.93b 17.03 ± 1.0c 13.5 ± 2.6c 2.24 ± 0.2c 12.63 ± 0.43c 7.56 ± 1.71c 29.36 ± 4.3b 21.42 ± 0.83c 58.21 ± 0.92a
C18:1(n-9) 42.32 ± 3.13c 88.83 ± 3.74c 237.1 ± 2.57b 93.16 ± 6.8c 82.94 ± 4.1c 33.76 ± 1.80c 21.7 ± 1.26c 20.06 ± 3.5c 47.59 ± 1.5c 333.0 ± 3.07a 345.14 ± 4.2a
C18:2(n-6) 114.39 ± 6.7de 99.72 ± 2.6de 256.86 ± 9.2bc 180.16 ± 3.8cd 131.74 ± 3.1de 54.53 ± 2.80e 72.4 ± 5.18de 53.03 ± 6.2e 128.1 ± 4.7de 328.9 ± 6.77b 447.52 ± 4.6a
C18:3(n-6) 8.09 ± 0.45b 8.93 ± 0.5b 21.11 ± 0.77a 15.86 ± 1.8a 10.45 ± 0.6b 3.06 ± 0.6b 31.19 ± 4.16a 3.95 ± 1.2b 11.0 ± 2.15a 18.71 ± 0.15a 32.67 ± 0.49a
C18:3(n-3) 382.32 ± 6.7d 454.37 ± 10.5d 1219.6 ± 9.0c 640.20 ± 6.5d 555.92 ± 6.78d 249.7 ± 5.80d 267.1 ± 1.42d 159.19 ± 3.2d 652.1 ± 3.9d 2201.4 ± 7.3b 3419.53 ± 8.3a
C18:4(n-3) 54.91 ± 3.3ef 68.05 ± 7.2def 201.43 ± 8.36b 116.88 ± 5.7de 72.07 ± 2.64edf 30.70 ± 1.37f 54.38 ± 1.7ef 13.91 ± 0.96f 196.5 ± 4.2bc 129.67 ± 1.4cd 273.82 ± 6.3a
C20:3(n-6) 3.55 ± 0.13c 5.43 ± 1.1c 7.92 ± 0.34c 6.77 ± 0.26c 3.17 ± 1.0c 3.48 ± 1.5c 3.21 ± 0.22c 2.87 ± 0.75c 10.3 ± 1.67bc 27.29 ± 1.47a 15.99 ± 0.6b
C20:3(n-3) 4.94 ± 0.2 de 6.26 ± 1.4de 12.70 ± 0.45cd 8.81 ± 1.49c 4.43 ± 0.6de n.d. 3.6 ± 0.14e n.d. 12.2 ± 2.45c 18.18 ± 0.38b 26.12 ± 0.17a
C20:4(n-6) 9.63 ± 0.59bcde 11.8 5 ± 0.6bcd 30.14 ± 1.5a 16.87 ± 2.62bc 11.26 ± 0.8bcde 5.95 ± 0.33de 9.25 ± 0.85cde 4.46 ± 1.3e 16.9 ± 4.14bc 18.80 ± 0.13b 34.19 ± 0.43a
C20:5(n-3) 5.87 ± 0.30d 8.13 ± 1.8cd 20.21 ± 0.78a 13.44 ± 3.5bc 8.54 ± 0.5cd 2.68 ± 0.11d 5.43 ± 0.94d 3.06 ± 1.2d 13.71 ± 3.6bc 18.90 ± 0.24ab 25.49 ± 0.58a
C22:6(n-3) 23.94 ± 1.42ef 38.34 ± 1.0de 73.70 ± 3.15bc 51.75 ± 2.13cd 23.57 ± 3.1ef 10.26 ± 0.52f 26.85 ± 0.85ef 16.86 ± 1.97ef 36.02 ± 3.2de 86.47 ± 1.9b 170.07 ± 1.21a
Others 12.07 ± 0.5c 18.51 ± 3bc 73.83 ± 4.22a 23.13 ± 0.92bc 17.49 ± 0.5bc 13.1 ± 0.35c 23.12 ± 0.46bc 17.5 ± 1.26bc 24.44 ± 1.6bc 76.77 ± 1.7a 41.78 ± 0.18b
SFA 291.6 ± 5.7c 384.85 ± 6.2b 1106.4 ± 9.68c 554.1 ± 6.5c 405.4 ± 4.03c 177.9 ± 4.3c 241.5 ± 0.94c 235.76 ± 4.8c 933.85 ± 8.5b 2469.2 ± 8.9a 2707.9 ± 9.7a
MUFA 58.15 ± 3.8b 129 ± 4.6b 366.7 ± 4.65a 137.2 ± 3.03b 1116.7 ± 10.7b 50.7 ± 1.13b 45.77 ± 0.20b 40.42 ± 0.66b 114.7 ± 1.9b 407.2 ± 3.81a 458.3 ± 5.63a
PUFA 607.64 ± 7.5c 701 ± 15.42c 1843.8 ± 7.64c 1050.7 ± 12.5c 109.4 ± 7.21c 360.4 ± 8.04c 473.42 ± 5.2c 257.34 ± 4.8c 768.2 ± 5.4c 2848.3 ± 9.2b 4445.4 ± 10.6a
n6/n3 0.29 ± 0.05:1ab 0.22 ± 0.02bc 0.21 ± 0.01bcd 0.26 ± 0.05ab 0.23 ± 0.02bc 0.22 ± 0.02bc 0.32 ± 0.12a 0.33 ± 0.01a 0.27 ± 0.02ab 0.16 ± 0.05cd 0.14 ± 0.01d
BD, Bligh and Dyer; BDS, Bligh and Dyer with sonication; BDB, Bligh and Dyer with buffer; FM, Folch method; FMS, Folch method with sonication; FMB, Folch method with buffer; CS, Cequier-Sánchez method; CSS, Cequier-Sánchezmethod with sonication; CSB, Cequier-Sánchez method with buffer; GM, Garcia method; LRC, Lepage and Roy modified by Cohen method. Others: Summation of the contents of C12:0, C13:0, C15:0, C17:0, C17:1, and C20:1 FAs.a–fValues in a row without a common superscript are significantly different (P < 0.05); n.d.: not detected.
Method
forlipid
andfatty
acidextraction
fromm
arinem
acroalgae/P.K
umari
etal./A
nal.Biochem.415
(2011)134–
144139
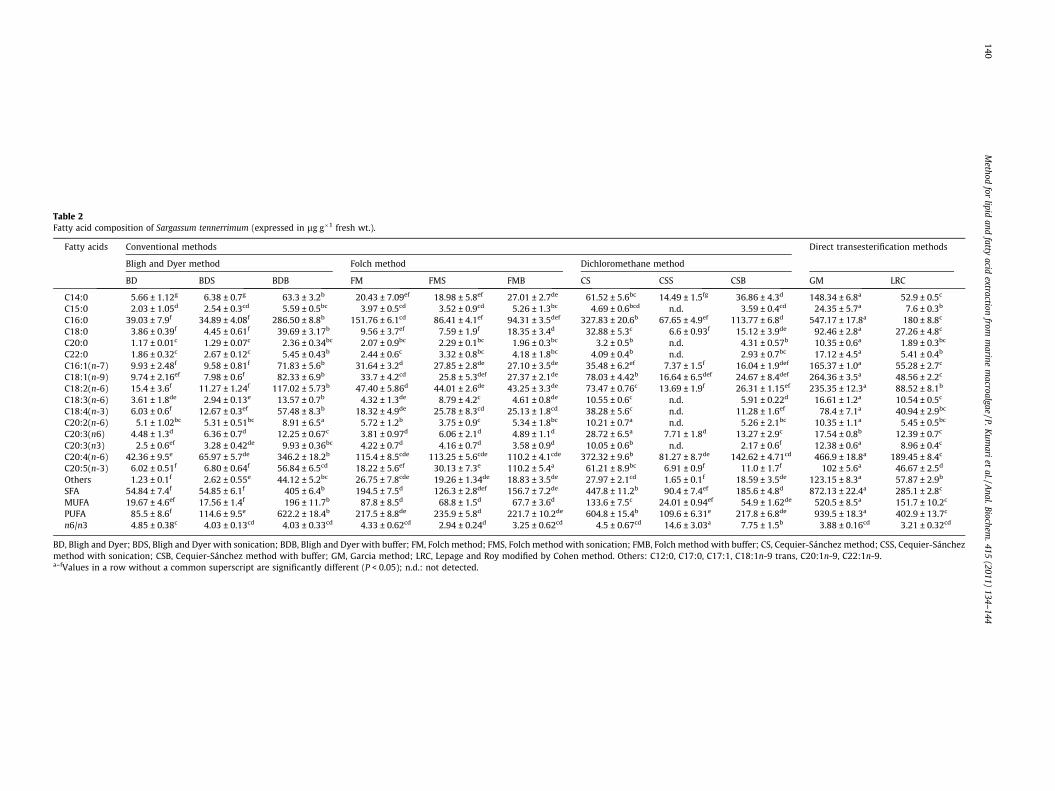
Table 2Fatty acid composition of Sargassum tennerrimum (expressed in lg g�1 fresh wt.).
Fatty acids Conventional methods Direct transesterification methods
Bligh and Dyer method Folch method Dichloromethane method
BD BDS BDB FM FMS FMB CS CSS CSB GM LRC
C14:0 5.66 ± 1.12g 6.38 ± 0.7g 63.3 ± 3.2b 20.43 ± 7.09ef 18.98 ± 5.8ef 27.01 ± 2.7de 61.52 ± 5.6bc 14.49 ± 1.5fg 36.86 ± 4.3d 148.34 ± 6.8a 52.9 ± 0.5c
C15:0 2.03 ± 1.05d 2.54 ± 0.3cd 5.59 ± 0.5bc 3.97 ± 0.5cd 3.52 ± 0.9cd 5.26 ± 1.3bc 4.69 ± 0.6bcd n.d. 3.59 ± 0.4cd 24.35 ± 5.7a 7.6 ± 0.3b
C16:0 39.03 ± 7.9f 34.89 ± 4.08f 286.50 ± 8.8b 151.76 ± 6.1cd 86.41 ± 4.1ef 94.31 ± 3.5def 327.83 ± 20.6b 67.65 ± 4.9ef 113.77 ± 6.8d 547.17 ± 17.8a 180 ± 8.8c
C18:0 3.86 ± 0.39f 4.45 ± 0.61f 39.69 ± 3.17b 9.56 ± 3.7ef 7.59 ± 1.9f 18.35 ± 3.4d 32.88 ± 5.3c 6.6 ± 0.93f 15.12 ± 3.9de 92.46 ± 2.8a 27.26 ± 4.8c
C20:0 1.17 ± 0.01c 1.29 ± 0.07c 2.36 ± 0.34bc 2.07 ± 0.9bc 2.29 ± 0.1bc 1.96 ± 0.3bc 3.2 ± 0.5b n.d. 4.31 ± 0.57b 10.35 ± 0.6a 1.89 ± 0.3bc
C22:0 1.86 ± 0.32c 2.67 ± 0.12c 5.45 ± 0.43b 2.44 ± 0.6c 3.32 ± 0.8bc 4.18 ± 1.8bc 4.09 ± 0.4b n.d. 2.93 ± 0.7bc 17.12 ± 4.5a 5.41 ± 0.4b
C16:1(n-7) 9.93 ± 2.48f 9.58 ± 0.81f 71.83 ± 5.6b 31.64 ± 3.2d 27.85 ± 2.8de 27.10 ± 3.5de 35.48 ± 6.2ef 7.37 ± 1.5f 16.04 ± 1.9def 165.37 ± 1.0a 55.28 ± 2.7c
C18:1(n-9) 9.74 ± 2.16ef 7.98 ± 0.6f 82.33 ± 6.9b 33.7 ± 4.2cd 25.8 ± 5.3def 27.37 ± 2.1de 78.03 ± 4.42b 16.64 ± 6.5def 24.67 ± 8.4def 264.36 ± 3.5a 48.56 ± 2.2c
C18:2(n-6) 15.4 ± 3.6f 11.27 ± 1.24f 117.02 ± 5.73b 47.40 ± 5.86d 44.01 ± 2.6de 43.25 ± 3.3de 73.47 ± 0.76c 13.69 ± 1.9f 26.31 ± 1.15ef 235.35 ± 12.3a 88.52 ± 8.1b
C18:3(n-6) 3.61 ± 1.8de 2.94 ± 0.13e 13.57 ± 0.7b 4.32 ± 1.3de 8.79 ± 4.2c 4.61 ± 0.8de 10.55 ± 0.6c n.d. 5.91 ± 0.22d 16.61 ± 1.2a 10.54 ± 0.5c
C18:4(n-3) 6.03 ± 0.6f 12.67 ± 0.3ef 57.48 ± 8.3b 18.32 ± 4.9de 25.78 ± 8.3cd 25.13 ± 1.8cd 38.28 ± 5.6c n.d. 11.28 ± 1.6ef 78.4 ± 7.1a 40.94 ± 2.9bc
C20:2(n-6) 5.1 ± 1.02bc 5.31 ± 0.51bc 8.91 ± 6.5a 5.72 ± 1.2b 3.75 ± 0.9c 5.34 ± 1.8bc 10.21 ± 0.7a n.d. 5.26 ± 2.1bc 10.35 ± 1.1a 5.45 ± 0.5bc
C20:3(n6) 4.48 ± 1.3d 6.36 ± 0.7d 12.25 ± 0.67c 3.81 ± 0.97d 6.06 ± 2.1d 4.89 ± 1.1d 28.72 ± 6.5a 7.71 ± 1.8d 13.27 ± 2.9c 17.54 ± 0.8b 12.39 ± 0.7c
C20:3(n3) 2.5 ± 0.6ef 3.28 ± 0.42de 9.93 ± 0.36bc 4.22 ± 0.7d 4.16 ± 0.7d 3.58 ± 0.9d 10.05 ± 0.6b n.d. 2.17 ± 0.6f 12.38 ± 0.6a 8.96 ± 0.4c
C20:4(n-6) 42.36 ± 9.5e 65.97 ± 5.7de 346.2 ± 18.2b 115.4 ± 8.5cde 113.25 ± 5.6cde 110.2 ± 4.1cde 372.32 ± 9.6b 81.27 ± 8.7de 142.62 ± 4.71cd 466.9 ± 18.8a 189.45 ± 8.4c
C20:5(n-3) 6.02 ± 0.51f 6.80 ± 0.64f 56.84 ± 6.5cd 18.22 ± 5.6ef 30.13 ± 7.3e 110.2 ± 5.4a 61.21 ± 8.9bc 6.91 ± 0.9f 11.0 ± 1.7f 102 ± 5.6a 46.67 ± 2.5d
Others 1.23 ± 0.1f 2.62 ± 0.55e 44.12 ± 5.2bc 26.75 ± 7.8cde 19.26 ± 1.34de 18.83 ± 3.5de 27.97 ± 2.1cd 1.65 ± 0.1f 18.59 ± 3.5de 123.15 ± 8.3a 57.87 ± 2.9b
SFA 54.84 ± 7.4f 54.85 ± 6.1f 405 ± 6.4b 194.5 ± 7.5d 126.3 ± 2.8def 156.7 ± 7.2de 447.8 ± 11.2b 90.4 ± 7.4ef 185.6 ± 4.8d 872.13 ± 22.4a 285.1 ± 2.8c
MUFA 19.67 ± 4.6ef 17.56 ± 1.4f 196 ± 11.7b 87.8 ± 8.5d 68.8 ± 1.5d 67.7 ± 3.6d 133.6 ± 7.5c 24.01 ± 0.94ef 54.9 ± 1.62de 520.5 ± 8.5a 151.7 ± 10.2c
PUFA 85.5 ± 8.6f 114.6 ± 9.5e 622.2 ± 18.4b 217.5 ± 8.8de 235.9 ± 5.8d 221.7 ± 10.2de 604.8 ± 15.4b 109.6 ± 6.31e 217.8 ± 6.8de 939.5 ± 18.3a 402.9 ± 13.7c
n6/n3 4.85 ± 0.38c 4.03 ± 0.13cd 4.03 ± 0.33cd 4.33 ± 0.62cd 2.94 ± 0.24d 3.25 ± 0.62cd 4.5 ± 0.67cd 14.6 ± 3.03a 7.75 ± 1.5b 3.88 ± 0.16cd 3.21 ± 0.32cd
BD, Bligh and Dyer; BDS, Bligh and Dyer with sonication; BDB, Bligh and Dyer with buffer; FM, Folch method; FMS, Folch method with sonication; FMB, Folch method with buffer; CS, Cequier-Sánchez method; CSS, Cequier-Sánchezmethod with sonication; CSB, Cequier-Sánchez method with buffer; GM, Garcia method; LRC, Lepage and Roy modified by Cohen method. Others: C12:0, C17:0, C17:1, C18:1n-9 trans, C20:1n-9, C22:1n-9.a–fValues in a row without a common superscript are significantly different (P < 0.05); n.d.: not detected.
140M
ethodfor
lipidand
fattyacid
extractionfrom
marine
macroalgae
/P.Kum
ariet
al./Anal.Biochem
.415(2011)
134–144
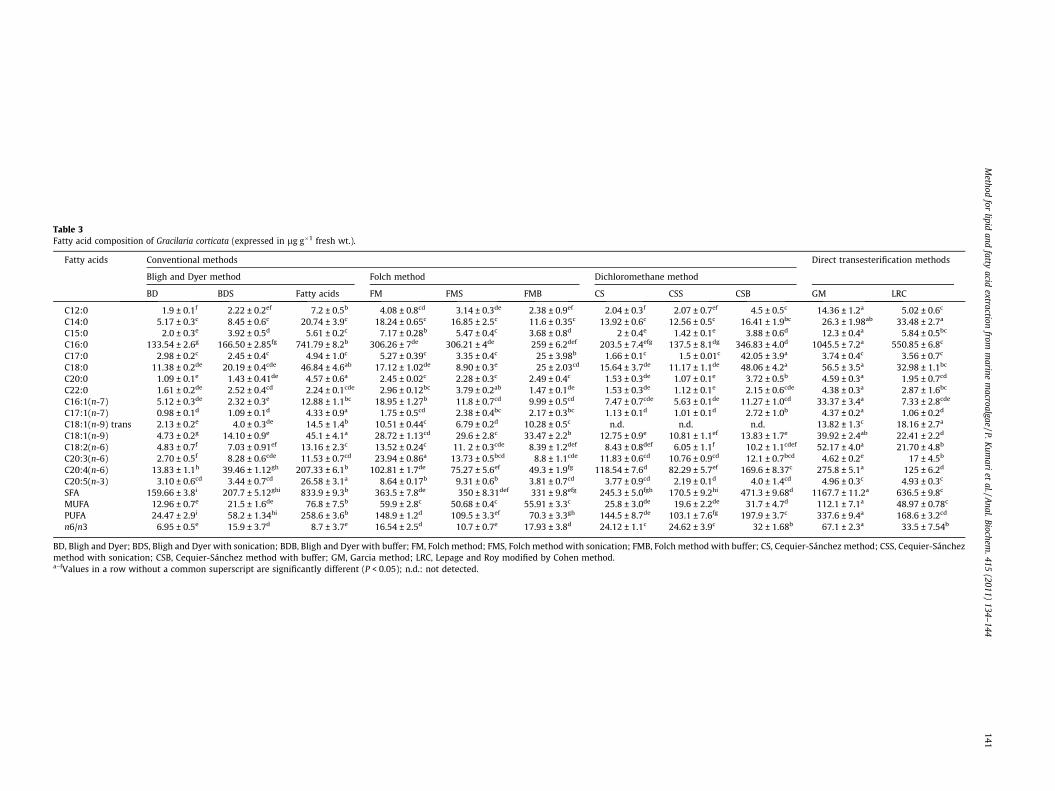
Table 3Fatty acid composition of Gracilaria corticata (expressed in lg g�1 fresh wt.).
Fatty acids Conventional methods Direct transesterification methods
Bligh and Dyer method Folch method Dichloromethane method
BD BDS Fatty acids FM FMS FMB CS CSS CSB GM LRC
C12:0 1.9 ± 0.1f 2.22 ± 0.2ef 7.2 ± 0.5b 4.08 ± 0.8cd 3.14 ± 0.3de 2.38 ± 0.9ef 2.04 ± 0.3f 2.07 ± 0.7ef 4.5 ± 0.5c 14.36 ± 1.2a 5.02 ± 0.6c
C14:0 5.17 ± 0.3c 8.45 ± 0.6c 20.74 ± 3.9c 18.24 ± 0.65c 16.85 ± 2.5c 11.6 ± 0.35c 13.92 ± 0.6c 12.56 ± 0.5c 16.41 ± 1.9bc 26.3 ± 1.98ab 33.48 ± 2.7a
C15:0 2.0 ± 0.3e 3.92 ± 0.5d 5.61 ± 0.2c 7.17 ± 0.28b 5.47 ± 0.4c 3.68 ± 0.8d 2 ± 0.4e 1.42 ± 0.1e 3.88 ± 0.6d 12.3 ± 0.4a 5.84 ± 0.5bc
C16:0 133.54 ± 2.6g 166.50 ± 2.85fg 741.79 ± 8.2b 306.26 ± 7de 306.21 ± 4de 259 ± 6.2def 203.5 ± 7.4efg 137.5 ± 8.1dg 346.83 ± 4.0d 1045.5 ± 7.2a 550.85 ± 6.8c
C17:0 2.98 ± 0.2c 2.45 ± 0.4c 4.94 ± 1.0c 5.27 ± 0.39c 3.35 ± 0.4c 25 ± 3.98b 1.66 ± 0.1c 1.5 ± 0.01c 42.05 ± 3.9a 3.74 ± 0.4c 3.56 ± 0.7c
C18:0 11.38 ± 0.2de 20.19 ± 0.4cde 46.84 ± 4.6ab 17.12 ± 1.02de 8.90 ± 0.3e 25 ± 2.03cd 15.64 ± 3.7de 11.17 ± 1.1de 48.06 ± 4.2a 56.5 ± 3.5a 32.98 ± 1.1bc
C20:0 1.09 ± 0.1e 1.43 ± 0.41de 4.57 ± 0.6a 2.45 ± 0.02c 2.28 ± 0.3c 2.49 ± 0.4c 1.53 ± 0.3de 1.07 ± 0.1e 3.72 ± 0.5b 4.59 ± 0.3a 1.95 ± 0.7cd
C22:0 1.61 ± 0.2de 2.52 ± 0.4cd 2.24 ± 0.1cde 2.96 ± 0.12bc 3.79 ± 0.2ab 1.47 ± 0.1de 1.53 ± 0.3de 1.12 ± 0.1e 2.15 ± 0.6cde 4.38 ± 0.3a 2.87 ± 1.6bc
C16:1(n-7) 5.12 ± 0.3de 2.32 ± 0.3e 12.88 ± 1.1bc 18.95 ± 1.27b 11.8 ± 0.7cd 9.99 ± 0.5cd 7.47 ± 0.7cde 5.63 ± 0.1de 11.27 ± 1.0cd 33.37 ± 3.4a 7.33 ± 2.8cde
C17:1(n-7) 0.98 ± 0.1d 1.09 ± 0.1d 4.33 ± 0.9a 1.75 ± 0.5cd 2.38 ± 0.4bc 2.17 ± 0.3bc 1.13 ± 0.1d 1.01 ± 0.1d 2.72 ± 1.0b 4.37 ± 0.2a 1.06 ± 0.2d
C18:1(n-9) trans 2.13 ± 0.2e 4.0 ± 0.3de 14.5 ± 1.4b 10.51 ± 0.44c 6.79 ± 0.2d 10.28 ± 0.5c n.d. n.d. n.d. 13.82 ± 1.3c 18.16 ± 2.7a
C18:1(n-9) 4.73 ± 0.2g 14.10 ± 0.9e 45.1 ± 4.1a 28.72 ± 1.13cd 29.6 ± 2.8c 33.47 ± 2.2b 12.75 ± 0.9e 10.81 ± 1.1ef 13.83 ± 1.7e 39.92 ± 2.4ab 22.41 ± 2.2d
C18:2(n-6) 4.83 ± 0.7f 7.03 ± 0.91ef 13.16 ± 2.3c 13.52 ± 0.24c 11. 2 ± 0.3cde 8.39 ± 1.2def 8.43 ± 0.8def 6.05 ± 1.1f 10.2 ± 1.1cdef 52.17 ± 4.0a 21.70 ± 4.8b
C20:3(n-6) 2.70 ± 0.5f 8.28 ± 0.6cde 11.53 ± 0.7cd 23.94 ± 0.86a 13.73 ± 0.5bcd 8.8 ± 1.1cde 11.83 ± 0.6cd 10.76 ± 0.9cd 12.1 ± 0.7bcd 4.62 ± 0.2e 17 ± 4.5b
C20:4(n-6) 13.83 ± 1.1h 39.46 ± 1.12gh 207.33 ± 6.1b 102.81 ± 1.7de 75.27 ± 5.6ef 49.3 ± 1.9fg 118.54 ± 7.6d 82.29 ± 5.7ef 169.6 ± 8.37c 275.8 ± 5.1a 125 ± 6.2d
C20:5(n-3) 3.10 ± 0.6cd 3.44 ± 0.7cd 26.58 ± 3.1a 8.64 ± 0.17b 9.31 ± 0.6b 3.81 ± 0.7cd 3.77 ± 0.9cd 2.19 ± 0.1d 4.0 ± 1.4cd 4.96 ± 0.3c 4.93 ± 0.3c
SFA 159.66 ± 3.8i 207.7 ± 5.12ghi 833.9 ± 9.3b 363.5 ± 7.8de 350 ± 8.31def 331 ± 9.8efg 245.3 ± 5.0fgh 170.5 ± 9.2hi 471.3 ± 9.68d 1167.7 ± 11.2a 636.5 ± 9.8c
MUFA 12.96 ± 0.7e 21.5 ± 1.6de 76.8 ± 7.5b 59.9 ± 2.8c 50.68 ± 0.4c 55.91 ± 3.3c 25.8 ± 3.0de 19.6 ± 2.2de 31.7 ± 4.7d 112.1 ± 7.1a 48.97 ± 0.78c
PUFA 24.47 ± 2.9i 58.2 ± 1.34hi 258.6 ± 3.6b 148.9 ± 1.2d 109.5 ± 3.3ef 70.3 ± 3.3gh 144.5 ± 8.7de 103.1 ± 7.6fg 197.9 ± 3.7c 337.6 ± 9.4a 168.6 ± 3.2cd
n6/n3 6.95 ± 0.5e 15.9 ± 3.7d 8.7 ± 3.7e 16.54 ± 2.5d 10.7 ± 0.7e 17.93 ± 3.8d 24.12 ± 1.1c 24.62 ± 3.9c 32 ± 1.68b 67.1 ± 2.3a 33.5 ± 7.54b
BD, Bligh and Dyer; BDS, Bligh and Dyer with sonication; BDB, Bligh and Dyer with buffer; FM, Folch method; FMS, Folch method with sonication; FMB, Folch method with buffer; CS, Cequier-Sánchez method; CSS, Cequier-Sánchezmethod with sonication; CSB, Cequier-Sánchez method with buffer; GM, Garcia method; LRC, Lepage and Roy modified by Cohen method.a–fValues in a row without a common superscript are significantly different (P < 0.05); n.d.: not detected.
Method
forlipid
andfatty
acidextraction
fromm
arinem
acroalgae/P.K
umari
etal./A
nal.Biochem.415
(2011)134–
144141
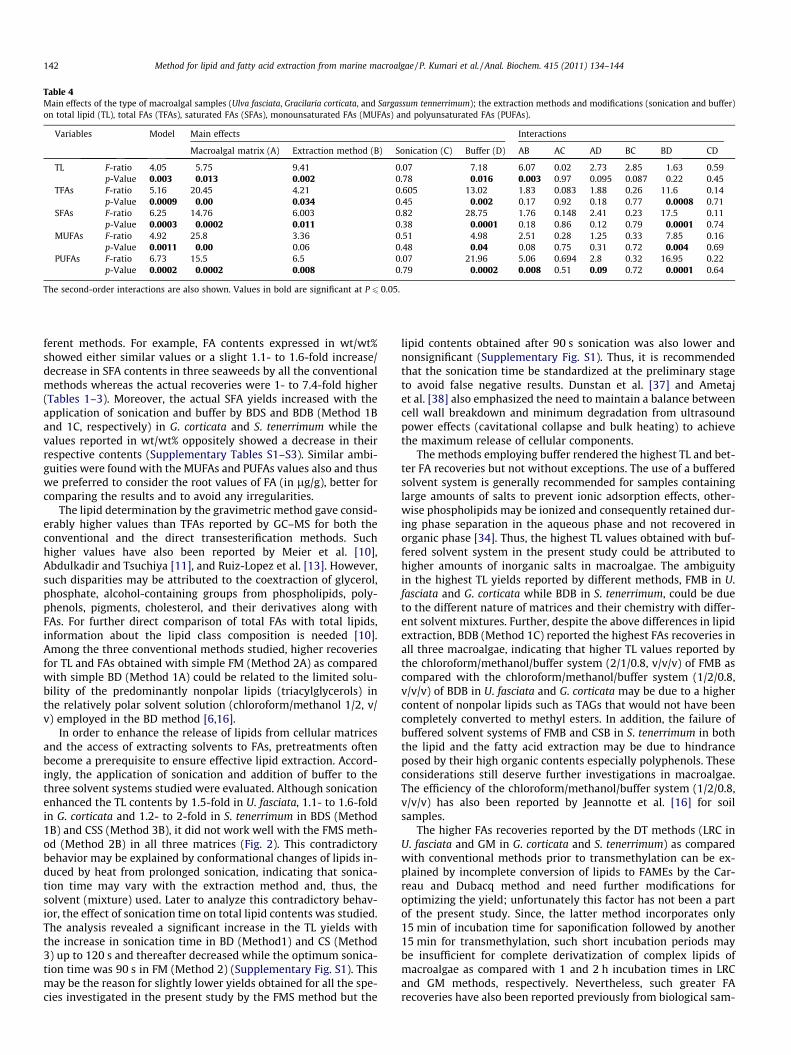
Table 4Main effects of the type of macroalgal samples (Ulva fasciata, Gracilaria corticata, and Sargassum tennerrimum); the extraction methods and modifications (sonication and buffer)on total lipid (TL), total FAs (TFAs), saturated FAs (SFAs), monounsaturated FAs (MUFAs) and polyunsaturated FAs (PUFAs).
Variables Model Main effects Interactions
Macroalgal matrix (A) Extraction method (B) Sonication (C) Buffer (D) AB AC AD BC BD CD
TL F-ratio 4.05 5.75 9.41 0.07 7.18 6.07 0.02 2.73 2.85 1.63 0.59p-Value 0.003 0.013 0.002 0.78 0.016 0.003 0.97 0.095 0.087 0.22 0.45
TFAs F-ratio 5.16 20.45 4.21 0.605 13.02 1.83 0.083 1.88 0.26 11.6 0.14p-Value 0.0009 0.00 0.034 0.45 0.002 0.17 0.92 0.18 0.77 0.0008 0.71
SFAs F-ratio 6.25 14.76 6.003 0.82 28.75 1.76 0.148 2.41 0.23 17.5 0.11p-Value 0.0003 0.0002 0.011 0.38 0.0001 0.18 0.86 0.12 0.79 0.0001 0.74
MUFAs F-ratio 4.92 25.8 3.36 0.51 4.98 2.51 0.28 1.25 0.33 7.85 0.16p-Value 0.0011 0.00 0.06 0.48 0.04 0.08 0.75 0.31 0.72 0.004 0.69
PUFAs F-ratio 6.73 15.5 6.5 0.07 21.96 5.06 0.694 2.8 0.32 16.95 0.22p-Value 0.0002 0.0002 0.008 0.79 0.0002 0.008 0.51 0.09 0.72 0.0001 0.64
The second-order interactions are also shown. Values in bold are significant at P 6 0.05.
142 Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144
ferent methods. For example, FA contents expressed in wt/wt%showed either similar values or a slight 1.1- to 1.6-fold increase/decrease in SFA contents in three seaweeds by all the conventionalmethods whereas the actual recoveries were 1- to 7.4-fold higher(Tables 1–3). Moreover, the actual SFA yields increased with theapplication of sonication and buffer by BDS and BDB (Method 1Band 1C, respectively) in G. corticata and S. tenerrimum while thevalues reported in wt/wt% oppositely showed a decrease in theirrespective contents (Supplementary Tables S1–S3). Similar ambi-guities were found with the MUFAs and PUFAs values also and thuswe preferred to consider the root values of FA (in lg/g), better forcomparing the results and to avoid any irregularities.
The lipid determination by the gravimetric method gave consid-erably higher values than TFAs reported by GC–MS for both theconventional and the direct transesterification methods. Suchhigher values have also been reported by Meier et al. [10],Abdulkadir and Tsuchiya [11], and Ruiz-Lopez et al. [13]. However,such disparities may be attributed to the coextraction of glycerol,phosphate, alcohol-containing groups from phospholipids, poly-phenols, pigments, cholesterol, and their derivatives along withFAs. For further direct comparison of total FAs with total lipids,information about the lipid class composition is needed [10].Among the three conventional methods studied, higher recoveriesfor TL and FAs obtained with simple FM (Method 2A) as comparedwith simple BD (Method 1A) could be related to the limited solu-bility of the predominantly nonpolar lipids (triacylglycerols) inthe relatively polar solvent solution (chloroform/methanol 1/2, v/v) employed in the BD method [6,16].
In order to enhance the release of lipids from cellular matricesand the access of extracting solvents to FAs, pretreatments oftenbecome a prerequisite to ensure effective lipid extraction. Accord-ingly, the application of sonication and addition of buffer to thethree solvent systems studied were evaluated. Although sonicationenhanced the TL contents by 1.5-fold in U. fasciata, 1.1- to 1.6-foldin G. corticata and 1.2- to 2-fold in S. tenerrimum in BDS (Method1B) and CSS (Method 3B), it did not work well with the FMS meth-od (Method 2B) in all three matrices (Fig. 2). This contradictorybehavior may be explained by conformational changes of lipids in-duced by heat from prolonged sonication, indicating that sonica-tion time may vary with the extraction method and, thus, thesolvent (mixture) used. Later to analyze this contradictory behav-ior, the effect of sonication time on total lipid contents was studied.The analysis revealed a significant increase in the TL yields withthe increase in sonication time in BD (Method1) and CS (Method3) up to 120 s and thereafter decreased while the optimum sonica-tion time was 90 s in FM (Method 2) (Supplementary Fig. S1). Thismay be the reason for slightly lower yields obtained for all the spe-cies investigated in the present study by the FMS method but the
lipid contents obtained after 90 s sonication was also lower andnonsignificant (Supplementary Fig. S1). Thus, it is recommendedthat the sonication time be standardized at the preliminary stageto avoid false negative results. Dunstan et al. [37] and Ametajet al. [38] also emphasized the need to maintain a balance betweencell wall breakdown and minimum degradation from ultrasoundpower effects (cavitational collapse and bulk heating) to achievethe maximum release of cellular components.
The methods employing buffer rendered the highest TL and bet-ter FA recoveries but not without exceptions. The use of a bufferedsolvent system is generally recommended for samples containinglarge amounts of salts to prevent ionic adsorption effects, other-wise phospholipids may be ionized and consequently retained dur-ing phase separation in the aqueous phase and not recovered inorganic phase [34]. Thus, the highest TL values obtained with buf-fered solvent system in the present study could be attributed tohigher amounts of inorganic salts in macroalgae. The ambiguityin the highest TL yields reported by different methods, FMB in U.fasciata and G. corticata while BDB in S. tenerrimum, could be dueto the different nature of matrices and their chemistry with differ-ent solvent mixtures. Further, despite the above differences in lipidextraction, BDB (Method 1C) reported the highest FAs recoveries inall three macroalgae, indicating that higher TL values reported bythe chloroform/methanol/buffer system (2/1/0.8, v/v/v) of FMB ascompared with the chloroform/methanol/buffer system (1/2/0.8,v/v/v) of BDB in U. fasciata and G. corticata may be due to a highercontent of nonpolar lipids such as TAGs that would not have beencompletely converted to methyl esters. In addition, the failure ofbuffered solvent systems of FMB and CSB in S. tenerrimum in boththe lipid and the fatty acid extraction may be due to hindranceposed by their high organic contents especially polyphenols. Theseconsiderations still deserve further investigations in macroalgae.The efficiency of the chloroform/methanol/buffer system (1/2/0.8,v/v/v) has also been reported by Jeannotte et al. [16] for soilsamples.
The higher FAs recoveries reported by the DT methods (LRC inU. fasciata and GM in G. corticata and S. tenerrimum) as comparedwith conventional methods prior to transmethylation can be ex-plained by incomplete conversion of lipids to FAMEs by the Car-reau and Dubacq method and need further modifications foroptimizing the yield; unfortunately this factor has not been a partof the present study. Since, the latter method incorporates only15 min of incubation time for saponification followed by another15 min for transmethylation, such short incubation periods maybe insufficient for complete derivatization of complex lipids ofmacroalgae as compared with 1 and 2 h incubation times in LRCand GM methods, respectively. Nevertheless, such greater FArecoveries have also been reported previously from biological sam-

Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144 143
ples with the direct transesterification method against classicalmultistep techniques [10,11,35].
Apart from this, to avoid the use of hazardous and toxic solventssuch as chloroform, dichloromethane was tested as an alternativeoption but the dichloromethane–methanol system (Method 3)adapted from Cequier-Sánchez et al., along with their modifica-tions (Method 3B and 3C) failed to compete with the potential ofthe chloroform/methanol/buffer system. Actually, the chloroformand methanol in combination exhibits strong dissolving powerfor the entire range of polarity found in lipids, as well as the abilityto break up membrane and denature (-lipo)proteins [39] and buf-fer added in the present study helped to overcome the ionicadsorption effects of salt that may hinder lipid extraction inmacroalgae.
Despite the variations among the FA yields of each method, thethree macroalgae showed typical profiles corresponding to theirrespective phyla, i.e., U. fasciata being a green alga was rich inC18 PUFAs while G. corticata being a red alga was rich with C20 PU-FAs, and S. tenerrimum being brown alga was rich with both (Tables1–3). Such trends have already been established earlier in severalstudies [3,22,24].
Further, standard FAME mix (Supelco) was analyzed on inter/intraday basis to ensure instrument response and data accuracyand a five-point calibration was carried out to ensure the linearity(r2 = 0.995), data not shown. Lastly, regardless of the method used,quantitative estimation of lipids and FAs requires researchers totake precautions at every step involved, to avoid variations in theresults.
In conclusion, this comparative study enabled us to test the effi-ciency of different methods for extraction of lipids and FAs in dif-ferent macroalgal matrices and experimental design furtherproved to be an efficient strategy for identifying its key factors.The macroalgal matrix, extraction method, and buffer were thekey determinants of optimum lipid and FA recoveries. FMB (Meth-od 2C) was the most efficient method of lipid extraction in U. fas-ciata (Chlorophyta) and G. corticata (Rhodophyta), while BDB(Method 1C) in S. tenerrimum (Phaeophyta) and DT methods gaveoptimum FA yields in all the three macroalgae investigated: LRC(Method 6) in U. fasciata and GM (Method 5) in G. corticata andS. tenerrimum. Thus, it is ideal to choose an extraction methodaccording to the desired purpose: DT methods for FA researchand conventional methods for the extensive study of lipid classesas the former does not separate the lipid fractions with extractionand methylation being accomplished in a single step. Further, careshould be taken while selecting the method for macroalgae,according to the group to which they belong, otherwise therewould be a risk of obtaining erratic and inaccurate results.
Acknowledgments
The financial assistance received from Council of Scientific andIndustrial Research (NWP-018) is gratefully acknowledged. We ex-press our sincere gratitude to Dr. Parimal Paul, Head, AnalyticalSciences, for generously allowing us to use the GC facility andMr. Harshad Brahmbhatt for technical help. The first author (P.K.)thanks CSIR for the award of Junior Research Fellowship (JRF).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ab.2011.04.010.
References
[1] M.H.G. Munro, J.W. Blunt, Marinlit, Version 10.4, Marine Chemical Group,University of Canterbury, Christchurch, New Zealand, 1999.
[2] S.K. Chandini, P. Ganesan, P.V. Suresh, N. Bhaskar, Seaweeds as source ofnutritionally beneficial compounds – a review, J. Food Sci. Technol. 45 (2008)1–13.
[3] P. Kumari, M. Kumar, V. Gupta, C.R.K. Reddy, B. Jha, Tropical marinemacroalgae as potential sources of nutritionally important PUFAs, FoodChem. 120 (2010) 749–757.
[4] K. Lee, M. Hong, S. Jung, S. Ha, B.J. Yu, H.M. Koo, S.M. Park, J. Seo, D. Kweon, J.C.Park, Y. Jin, Improved galactose fermentation of Saccharomyces cerevisiaethrough inverse metabolic engineering, Biotechnol. Bioeng., 108 (2010) 621–631.
[5] R. Petcavich, Method of producing hydrocarbon biofuels using geneticallymodified seaweed, US Patent Publication No. 2010/0120111 A1, 2010.
[6] S.J. Iverson, S.L.C. Lang, M.H. Cooper, Comparison of the Bligh and Dyer andFolch methods for total lipid determination in a broad range of marine tissue,Lipids 36 (2001) 1283–1287.
[7] A. Masood, K.D. Stark, N. Salem Jr., A simplified and efficient method for theanalysis of fatty acid methyl esters suitable for large clinical studies, J. LipidRes. 46 (2005) 2299–2305.
[8] G. Knothe, Dependence of biodiesel fuel properties on the structure of fattyacid alkyl esters, Fuel Process. Technol. 86 (2005) 1059–1070.
[9] M. Júarez, O. Polvillo, M. Conto, A. Ficco, S. Ballico, S. Failla, Comparison of fourextraction/methylation analytical methods to measure fatty acid compositionby gas chromatography in meat, J. Chromatogr. A 1190 (2008) 327–332.
[10] S. Meier, S.A. Mjøs, H. Joensen, O. Grahl-Nielsen, Validation of a one-stepextraction/methylation method for determination of fatty acids andcholesterol in marine tissues, J. Chromatogr. A 1104 (2006) 291–298.
[11] S. Abdulkadir, M. Tsuchiya, One-step method for quantitative and qualitativeanalysis of fatty acids in marine animal samples, J. Exp. Mar. Biol. Ecol. 354(2008) 1–8.
[12] T. Pérez-Palacios, J. Ruiz, D. Martín, E. Muriel, T. Antequera, Comparison ofdifferent methods for total lipid quantification in meat and meat products,Food Chem. 110 (2008) 1025–1029.
[13] N. Ruiz-Lopez, E. Martinez-Force, R. Garcés, Sequential one-step extraction andanalysis of triacylglycerols and fatty acids in plant tissues, Anal. Biochem. 317(2003) 247–254.
[14] M. Gómez-Brandón, M. Lores, J. Domínguez, A new combination of extractionand derivatization methods that reduces the complexity and preparation timein determining phospholipid fatty acids in solid environmental samples,Biores. Technol. 101 (2010) 1348–1354.
[15] M. Gómez-Brandón, M. Lores, J. Domínguez, Comparison of extraction andderivatization methods for fatty acid analysis in solid environmental matrixes,Anal. Bioanal. Chem. 392 (2008) 505–514.
[16] R. Jeannotte, C. Hamela, S. Jabaji, J.K. Whalen, Comparison of solvent mixturesfor pressurized solvent extraction of soil fatty acid biomarkers, Talanta 77(2008) 195–199.
[17] H.W. Chang, K.H. Jang, D. Lee, H.R. Kang, T.Y. Kim, B.H. Lee, B.W. Choi, S. Kim, J.Shin, Monoglycerides from the brown alga Sargassum sagamianum: isolation,synthesis, and biological activity, Bioorg. Med. Chem. Lett. 18 (2008) 3589–3592.
[18] Y.H. Kim, E.-H. Kim, C. Lee, M.-H. Kim, J.-R. Rho, Two new monogalactosyldiacylglycerols from brown alga Sargassum thunbergii, Lipids 42 (2007) 395–399.
[19] S.V. Khotimchenko, V.E. Vas’kovsky, An inositol-containing sphingolipid fromthe red alga Gracilaria verrucosa, Russ, J. Bioorg. Chem. 30 (2004) 168–171.
[20] A. Al-Fadhli, S. Wahidulla, L. D’Souza, Glycolipids from the red alga Chondriaarmata (Kütz.) Okamura, Glycobiology 16 (2006) 902–915.
[21] P. Matanjun, S. Mohamed, N.M. Mustapha, K. Muhammad, Nutrient content oftropical edible seaweeds, Euchema cottonii, Caulerpa lentillifera and Sargassumpolycystum, J. Appl. Phycol. 21 (2008) 75–80.
[22] S.V. Khotimchenko, V.E. Vaskovsky, T.V. Titlyanova, Fatty acids from the Pacificcoast of North California, Bot. Mar. 45 (2002) 17–22.
[23] D.I. Sanchez-Machado, J. Lopez-Cervantes, J. Lopez-Hernandez, P. Paseiro-Losada, Fatty acids, total lipid, protein and ash contents of processed edibleseaweeds, Food Chem. 85 (2004) 439–444.
[24] X. Li, X. Fan, L. Han, Q. Lou, Fatty acids of some algae from the Bohai Sea,Phytochemistry 59 (2002) 157–161.
[25] E.G. Bligh, W.J. Dyer, A rapid method of total lipid extraction and purification,Can. J. Biochem. Biophysiol. 37 (1959) 911–915.
[26] J. Folch, M. Lees, G.H. Sloane-Stanley, A simple method for the isolation andpurification of total lipids from animal tissues, J. Biol. Chem. 226 (1957) 497–509.
[27] M. Graeve, G. Kattner, C. Wiencke, U. Karsten, Fatty acid composition of Arcticand Antarctic macroalgae: indicator of phylogenetic and trophic relationships,Mar. Ecol. Prog. Ser. 231 (2002) 67–74.
[28] G.A. El-Shoubaky, A.M.Y. Moustafa, Essam A.E. Salem, Comparativephytochemical investigation of beneficial essential fatty acids on a variety ofmarine seaweeds algae, Res. J. Phytochem. 2 (2008) 18–26.
[29] E. Cequier-Sànchez, C. Rodriguez, À.G. Ravelo, R. Zàrate, Dichloromethane as asolvent for lipid extraction and assessment of lipid classes and fatty acids fromsamples of different natures, J. Agric. Food Chem. 56 (2008) 4297–4303.
[30] J.P. Carreau, J.P. Dubacq, Adaptation of macroscale method to the microscalefor fatty acid methyl transesterification of biological lipid extracts, J.Chromatogr. 151 (1978) 384–390.
[31] C. de la Cruz-Garcia, J. Lopez-Hernandez, J. Simal-Lozano, Gas chromatographicdetermination of the fatty-acid content of heat-treated green beans, J.Chromatogr. A 891 (2000) 367–370.

144 Method for lipid and fatty acid extraction from marine macroalgae / P. Kumari et al. / Anal. Biochem. 415 (2011) 134–144
[32] G. Lepage, C.C. Roy, Direct transesterification of all classes of lipids in a one-step reaction, J. Lipid Res. 27 (1986) 114–119.
[33] Z. Cohen, A. Vonshak, A. Richmond, Effect of environmental conditions on fattyacid composition of the red alga Porphyridium cruentum: correlation to growthrate, J. Phycol. 24 (1988) 328–332.
[34] W.W. Christie, Preparation of ester derivates of fatty acids for chromatographicanalysis, in: W.W. Christie (Ed.), Advances in Lipid Methodology, Oily Press,Dundee, 1993.
[35] M.J. Griffiths, R.P. van Hille, S.T.L. Harrison, Selection of directtransesterification as the preferred method for assay of fatty acid content ofmicroalgae, Lipids 45 (2010) 1053–1060.
[36] J.V. O’Fallon, J.R. Busboom, M.L. Nelson, C.T. Gaskins, A direct method for fattyacid methyl ester synthesis: application to wet meat tissues, oils andfeedstuffs, J. Anim. Sci. 85 (2007) 1511–1521.
[37] G.A. Dunstan, J.K. Volkman, S.M. Barrett, The effect of lyophilization on thesolvent extraction of lipid classes, fatty acids and sterols from the oysterCrassostrea gigas, Lipids 28 (1993) 937–944.
[38] B.N. Ametaj, G. Bobe, Y. Lu, J.W. Young, D.C. Beitz, Effect of sample preparation,length of time, and sample size on quantification of total lipids from bovineliver, J. Agric. Food Chem. 51 (2003) 2105–2110.
[39] M. Schreiner, Optimization of solvent extraction and direct transmethylationmethods for the analysis of egg yolk lipids, Int. J. Food Prop. 9 (2006) 573–581.