comparative analysis of vector biodistribution, … · nonhuman adenoviruses including bovine...
TRANSCRIPT

Virology 386 (2009) 44–54
Contents lists available at ScienceDirect
Virology
j ourna l homepage: www.e lsev ie r.com/ locate /yv i ro
Comparative analysis of vector biodistribution, persistence and gene expressionfollowing intravenous delivery of bovine, porcine and human adenoviral vectors ina mouse model
Anurag Sharma a,b,1, Dinesh S. Bangari a,b,1, Manish Tandon a,b, Aseem Pandey a,b,Harm HogenEsch a, Suresh K. Mittal a,b,⁎a Department of Comparative Pathobiology, School of Veterinary Medicine, Purdue University, West Lafayette, IN 47907, USAb Bindley Bioscience Center, Purdue University, West Lafayette, IN, USA
⁎ Corresponding author. Department of ComparaVeterinary Medicine, Purdue University, West Lafayette494 9830.
E-mail address: [email protected] (S.K. Mittal).1 Equal contributors.
0042-6822/$ – see front matter © 2009 Elsevier Inc. Adoi:10.1016/j.virol.2009.01.008
a b s t r a c t
a r t i c l e i n f oArticle history:
Nonhuman adenoviruses in Received 11 August 2008Returned to author for revision3 November 2008Accepted 9 January 2009Available online 10 February 2009Keywords:BiodistributionBovine adenovirusGene therapyNonhuman adenoviral vectorsPorcine adenovirus
cluding bovine adenovirus serotype 3 (BAd3) and porcine adenovirus serotype3 (PAd3) can circumvent pre-existing immunity against human adenovirus serotype 5 (HAd5) and arebeing developed as alternative vectors for gene delivery. To assess the usefulness of these vectors for invivo gene delivery, we compared biodistribution, persistence, state of vector genome, and transgene andvector gene expression by replication-defective BAd3 and PAd3 vectors with those of HAd5 vector in aFVB/n mouse model following intravenous inoculation. BAd3 vector efficiently transduced the heart,kidney and lung in addition to the liver and spleen and persisted for a longer duration compared to PAd3or HAd5 vectors. Biodistribution of PAd3 vector was comparable to that of HAd5 vector but showed morerapid vector clearance. Only linear episomal forms of BAd3, PAd3, and HAd5 vector genomes weredetected. All three vectors efficiently expressed the green fluorescent protein (GFP) transgeneproportionate to the vector genome copy number in various tissues. Furthermore, leaky expression ofvector genes, both the early (E4) and the late (hexon) was observed in all three vectors and graduallydeclined with time. These results suggest that BAd3 and PAd3 vectors could serve as an alternative orsupplement to HAd5 for gene delivery applications.
© 2009 Elsevier Inc. All rights reserved.
Introduction
Adenoviral (Ad) vectors have demonstrated great promise astherapeutic and prophylactic gene delivery systems (McConnell andImperiale, 2004; Russell, 2000). Numerous advantages offered by Advectors include the ease of large scale production, ease of geneticmanipulation, the lack of genomic integration, a relatively nonpatho-genic nature, and their ability to transduce both dividing and non-dividing cells. Vectors based on human adenovirus (HAd) serotype 5(HAd5) and HAd serotype 2 (HAd2) are currently most widelystudied (Edelstein et al., 2004; Tatsis and Ertl, 2004). Attachment ofHAd5 and HAd2 to a susceptible cell is mediated by the high-affinitybinding of the Ad fiber knob to a primary receptor, coxsackievirus andAd receptor (CAR) (Bergelson et al., 1997), followed by a secondaryinteraction of the penton base with integrins resulting in virusinternalization into the cell (Wickham et al., 1993). This feature poses
tive Pathobiology, School of, IN 47907, USA. Fax: +1 765
ll rights reserved.
a challenge in targeting of Ad to specific tissues due to the widedistribution of CAR in a variety of cell types. Preferential sequestrationof systemically administered Ad vectors to the liver (hepatotropism)also leads to inefficient transduction of target organs other than liverand at high vector doses, poses serious adverse effects (Raper et al.,2003). Furthermore, due to the endemic nature of HAd5 and HAd2,pre-existing vector immunity may potentially inhibit the levels andduration of transgene expression following inoculation with an Advector. Vector-associated toxicity and induction of strong innateimmunity are some of the other potential concerns for these vectors.
In order to expand the repertoire of Ad vectors, vectors based onless prevalent HAd serotypes such as HAd3, HAd11, and HAd35, ornonhuman adenoviruses such as bovine adenovirus type 3 (BAd3),porcine adenovirus type 3 (PAd3), ovine adenovirus, canine adeno-virus, simian adenoviruses, and fowl adenovirus, are being investi-gated as an alternative or supplement to HAd5 vectors (Bangari andMittal, 2006b; Farina et al., 2001; Hofmann et al., 1999; Kanerva et al.,2002; Kremer et al., 2000; Michou et al., 1999; Reddy et al., 1999a,1999b; Seshidhar Reddy et al., 2003; Stone et al., 2005). Vectors basedon nonhuman Ad or chimeric HAd vectors carrying the knob domainor entire fiber from nonhuman Ad have been developed thatdemonstrated novel and expanded tropism, in addition to evading

45A. Sharma et al. / Virology 386 (2009) 44–54
HAd-specific immunity (Bangari and Mittal, 2006a; Glasgow et al.,2004; Singh et al., 2008).
We have demonstrated that nonhuman Ad vectors based on PAd3and BAd3 efficiently transduce several types of human andmurine cellsin culture (Bangari et al., 2005b). These vectors appeared to utilizedistinct receptors for cell internalization (Bangari and Mittal, 2005;Bangari et al., 2005a) and cell entry was independent of CAR. Mostimportantly, we also showed that there were no pre-existing virusneutralizing antibodies against PAd3 or BAd3 in humans, and HAd5-neutralizing antibodies in human or raised in mice did not cross-neutralize PAd3 or BAd3 (Bangari et al., 2005b; Moffatt et al., 2000).
In the present study, we assessed the usefulness of replication-defective PAd3 and BAd3 vectors carrying the green fluorescentprotein (GFP) gene as a reporter for in vivo gene delivery.Biodistribution, persistence, the state of the vector genome, trans-gene, and vector gene expression of these nonhuman Ad vectors werecompared with those of HAd5 vector in FVB/n mouse modelfollowing intravenous inoculation. The choice of the mouse strainwas based on the fact that we have developed an immunocompetentFVB/n mouse model for breast cancer to investigate the role of EphA2receptor tyrosine kinase in tumorigenesis (Noblitt et al., 2005).Interestingly, the genome copy numbers of BAd3 vector in the heart,kidney, lung, and spleen were significantly (Pb0.05) higher thanthose of PAd3 and HAd5 vectors. BAd3 vector also persisted longer in
Fig. 1. Biodistribution of vector genome at various time points post-inoculation of mice withviral genome copies per 50 ng of total cellular DNA and were quantitated by real-time PCR. V⁎Pb0.05 versus BAd-GFP vector genome level at 0.25 day. †Pb0.05 for BAd-GFP versus HAd
all tissues examined compared to the other two vectors. The majorityof vector genome was detected in the linear form for all three vectors.In addition, expression levels of the viral early region gene 4 (E4) andthe late structural gene (hexon) for all three vectors were similar,indicating leaky vector gene expression and paralleled vectorbiodistribution. The results of this study demonstrate that BAd3 andPAd3 vectors have the potential for in vivo gene delivery applications,as supplements or alternatives to the currently used HAd vectors.
Results
Vector biodistribution and persistence in mice inoculated with HAd-GFP,PAd-GFP, or BAd-GFP
The liver and spleen are the main target organs for E1-deletedAd vector biodistribution following intravenous delivery (Ni et al., 2005;Stone et al., 2007) and the number of vector genomes decline drasticallywith time (Yang et al., 1994a). To investigate whether nonhuman Advectors based on PAd3 and BAd3 have significantly different biodistribu-tion and persistence from that of HAd5, FVB/n mice were inoculatedintravenously with 1010 vector particles (VP) of HAd-GFP (HAd5 withdeleted E1 and the GFP transgene under the cytomegalovirus (CMV)promoter) (Bangari andMittal, 2004), PAd-GFP (PAd3 with deleted E1Aand theGFP transgeneunder CMVpromoter) (Bangari andMittal, 2004),
HAd-GFP, PAd-GFP, or BAd-GFP. Genome concentrations are expressed as the number ofalues are reported as the mean±standard deviation for three mice at each time point.-GFP and PAd-GFP at each time point.

46 A. Sharma et al. / Virology 386 (2009) 44–54
or BAd-GFP (BAd3 with deleted E1A and the GFP transgene under CMVpromoter) (Bangari et al., 2005b). At 0.25, 0.5,1, 2, 4, 8, and 16 days post-inoculation, the mice were sacrificed and the liver, spleen, lung, kidney,and heart were collected. Total cellular DNA extracted from varioustissue samples were analyzed for quantification of vector genomes byreal-time PCR specific to the GFP transgene as it was present in all threevectors. The sensitivity of this assay was 3 copies of the vector genomeper 50 ng of total cellular DNA. The number of vector genomes andpersistence of BAd-GFP in all tissues were significantly (Pb0.05) higherthan those of either PAd-GFP or HAd-GFP except in the liver at 0.25 and1 days post-inoculationwhere the levelswere somewhat comparable inall three vector-inoculated mice (Fig. 1). It is interesting to note thatespecially in the lung, heart, and kidney, significantly high levels of BAd-GFP genomes persisted even at 16 days post-inoculation (the last timepoint examined in the present study). In general, vector genome levelsin PAd-GFP-transduced tissueswere at comparable levels at earlier timepoints and significantly lower (Pb0.05) or even below the detectionlevels at later time points compared to those of HAd-GFP-inoculatedtissues except the heart where the levels were comparable or evenhigher at some later timepoints (Fig.1). No vector genomewas detectedin the mock-inoculated mice at any time point (data not shown).
State of vector genomes in mice inoculated with HAd-GFP, PAd-GFP, orBAd-GFP
The HAd5 vector genome usually persists in episomal form withinthe host cell nucleus (Ehrhardt et al., 2003, 2008). However, the vector
Fig. 2. (A) Southern blot hybridization strategy to determine the state of the various Ad vecvector genome maps with the sites for the restriction enzyme that was used to digest the togenomemaps represent the DNA probes used for Southern blot analyses. The numbers correvector genome at various time points in the liver of mice inoculated with HAd-GFP, PAd-GFP,(n=3) were digested with KpnI, XhoI, or HpaI restriction enzymes, respectively and Southerthe liver of mock-inoculated mouse served as a negative control and purified vector DNA segenome of HAd-GFP, PAd-GFP, or BAd-GFP are presented.
genome may also get integrated into the chromosomal DNA at a verylow frequency (10−3 to 10−5) (Harui et al., 1999). In order to addressthe safety aspect of PAd3 and BAd3 vectors, it was important todetermine the state of their genomes in vivo. To assess the state ofvector DNA in the liver of HAd-GFP, PAd-GFP, or BAd-GFP inoculatedmice, samples of total cellular DNA extracted from the livers collectedat various time points post-inoculation were digested with KpnI (forHAd-GFP), XhoI (for PAd-GFP), or HpaI (for BAd-GFP) and subjected toSouthern blot analyses. The probes for Southern blot assay weredesigned taking into consideration that they differentiate the linearepisomal, circular episomal, and integrated forms of HAd-GFP, PAd-GFP, or BAd-GFP genome based on the fragment size of restrictionendonuclease (RE)-digested DNA. The sensitivity of the Southern blotassay, as determined by the amount of vector genome used as apositive control, was found to be 10 pg of DNA. The expected fragmentsizes for three forms of DNA (linear episomal, circular episomal, andintegrated) were 2.3, 3.4, and N2.3 kb for HAd-GFP, 3, 5, and N3 kb forPAd-GFP, and 2, 6.8, and N2 kb for BAd-GFP (Fig. 2A). In HAd-GFP, PAd-GFP or BAd-GFP-inoculated mice, 2.3, 3, or 2 kb specific bands weredetected up to 2, 1, or 8 days, respectively, suggesting thatpredominantly linear forms of the vector genomes were present inHAd-GFP, PAd-GFP, or BAd-GFP inoculated mice (Fig. 2B). Fragmentsof DNA that would correspond to a circular or integrated form of thevector genomes were not observed even after overexposure of theSouthern blots (data not shown). No signals were detected in samplesfrom mock-inoculated mice (Fig. 2B). The presence of circular form ofgenomes was also investigated by PCR using specific primer sets for
tor genomes in mice liver following systemic inoculation. Schematic diagram showingtal genomic DNA of HAd-GFP, PAd-GFP, or BAd-GFP. The solid bars parallel to the vectorspond to the nucleotide positions of wild type HAd5, PAd3, or BAd3 genome. (B) State ofor BAd-GFP. 20 μg of the liver DNA from HAd-GFP, PAd-GFP, or BAd-GFP-inoculated micen blots were performed with vector-specific probes. Similarly treated DNA sample fromrved as a positive control. The results of Southern blots showing the linear form of the

Fig. 3. Levels of GFP transgene mRNA expression at various time points post-inoculation of mice inoculated with HAd-GFP, PAd-GFP, or BAd-GFP. Real-time RT-PCR for GFP and18S rRNA (as an endogenous control for normalization) was performed using 200 ng of total RNA and comparative quantification of GFP expression levels was done. Values arereported relative to mean expression levels observed with HAd-GFP inoculated mice at 0.25 day post-inoculation, considering it as 100 arbitrary units. Values are reported asthe mean±standard deviation for three mice at each time point. ⁎Pb0.05 versus expression level by BAd-GFP at 0.25 day. †Pb0.05 for BAd-GFP versus HAd-GFP and PAd-GFP ateach time point.
Fig. 4. Immunohistochemistry for GFP expression in the liver sections of mice inoculated with HAd-GFP, PAd-GFP, or BAd-GFP. Immunohistochemistry for GFP was performed asdescribed in the Materials and Methods with the liver sections of mice that were sacrificed at 0.25 day (6 h) and 0.5 day (12 h) post-inoculation. The cells expressing GFP are stainedbrown. Similar results were obtained with tissue sections from three independent mice.
47A. Sharma et al. / Virology 386 (2009) 44–54
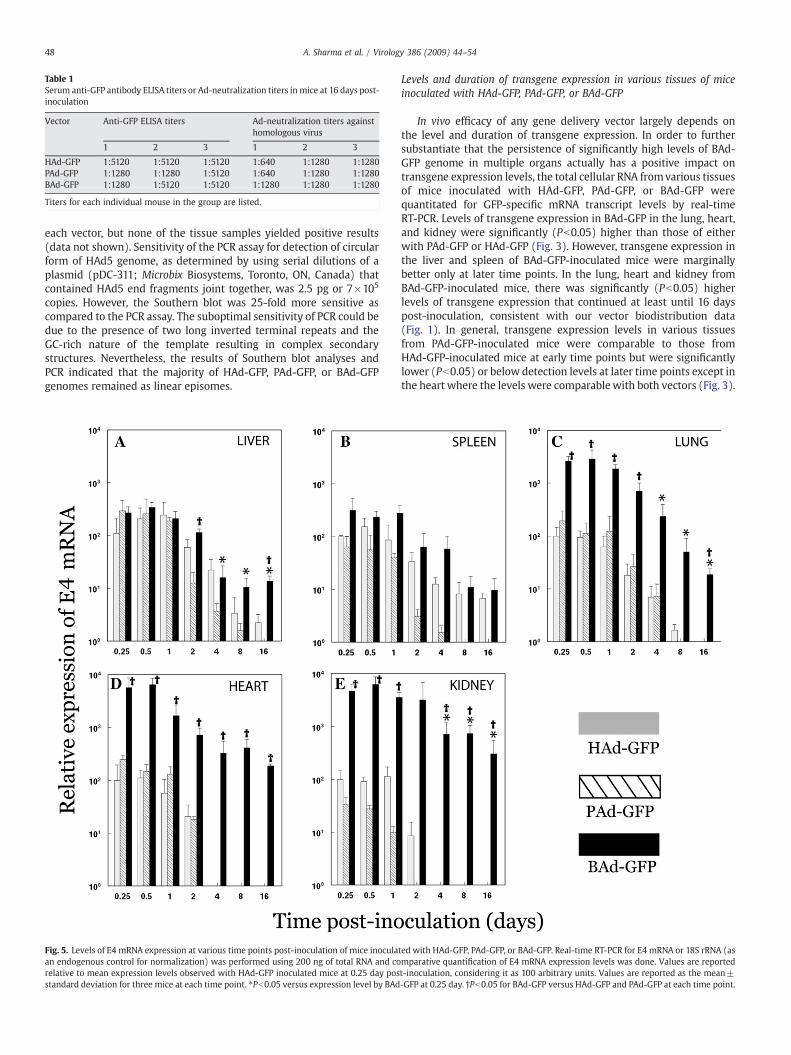
Table 1Serum anti-GFP antibody ELISA titers or Ad-neutralization titers in mice at 16 days post-inoculation
Vector Anti-GFP ELISA titers Ad-neutralization titers againsthomologous virus
1 2 3 1 2 3
HAd-GFP 1:5120 1:5120 1:5120 1:640 1:1280 1:1280PAd-GFP 1:1280 1:1280 1:5120 1:640 1:1280 1:1280BAd-GFP 1:1280 1:5120 1:5120 1:1280 1:1280 1:1280
Titers for each individual mouse in the group are listed.
48 A. Sharma et al. / Virology 386 (2009) 44–54
each vector, but none of the tissue samples yielded positive results(data not shown). Sensitivity of the PCR assay for detection of circularform of HAd5 genome, as determined by using serial dilutions of aplasmid (pDC-311; Microbix Biosystems, Toronto, ON, Canada) thatcontained HAd5 end fragments joint together, was 2.5 pg or 7×105
copies. However, the Southern blot was 25-fold more sensitive ascompared to the PCR assay. The suboptimal sensitivity of PCR could bedue to the presence of two long inverted terminal repeats and theGC-rich nature of the template resulting in complex secondarystructures. Nevertheless, the results of Southern blot analyses andPCR indicated that the majority of HAd-GFP, PAd-GFP, or BAd-GFPgenomes remained as linear episomes.
Fig. 5. Levels of E4 mRNA expression at various time points post-inoculation of mice inoculaan endogenous control for normalization) was performed using 200 ng of total RNA and corelative to mean expression levels observed with HAd-GFP inoculated mice at 0.25 day posstandard deviation for three mice at each time point. ⁎Pb0.05 versus expression level by BAd
Levels and duration of transgene expression in various tissues of miceinoculated with HAd-GFP, PAd-GFP, or BAd-GFP
In vivo efficacy of any gene delivery vector largely depends onthe level and duration of transgene expression. In order to furthersubstantiate that the persistence of significantly high levels of BAd-GFP genome in multiple organs actually has a positive impact ontransgene expression levels, the total cellular RNA fromvarious tissuesof mice inoculated with HAd-GFP, PAd-GFP, or BAd-GFP werequantitated for GFP-specific mRNA transcript levels by real-timeRT-PCR. Levels of transgene expression in BAd-GFP in the lung, heart,and kidney were significantly (Pb0.05) higher than those of eitherwith PAd-GFP or HAd-GFP (Fig. 3). However, transgene expression inthe liver and spleen of BAd-GFP-inoculated mice were marginallybetter only at later time points. In the lung, heart and kidney fromBAd-GFP-inoculated mice, there was significantly (Pb0.05) higherlevels of transgene expression that continued at least until 16 dayspost-inoculation, consistent with our vector biodistribution data(Fig. 1). In general, transgene expression levels in various tissuesfrom PAd-GFP-inoculated mice were comparable to those fromHAd-GFP-inoculated mice at early time points but were significantlylower (Pb0.05) or below detection levels at later time points except inthe heart where the levels were comparablewith both vectors (Fig. 3).
ted with HAd-GFP, PAd-GFP, or BAd-GFP. Real-time RT-PCR for E4 mRNA or 18S rRNA (asmparative quantification of E4 mRNA expression levels was done. Values are reportedt-inoculation, considering it as 100 arbitrary units. Values are reported as the mean±-GFP at 0.25 day. †Pb0.05 for BAd-GFP versus HAd-GFP and PAd-GFP at each time point.
49A. Sharma et al. / Virology 386 (2009) 44–54
No GFP expression was detected in mock-inoculated mice at any timepoint (data not shown). In order to detect the GFP protein in tissuesections, immunohistochemistry was performed. Positive cytoplasmicstaining was observed in hepatocytes of HAd-GFP, PAd-GFP, orBAd-GFP inoculated mice sacrificed at 0.25 and 0.5 day post-inoculation (Fig. 4), while no staining was detected in PBS-inoculatedmice or controls that did not receive the anti-GFP antibody treatment.GFP expression by immunohistochemistry was also observed in othertissues (data not shown).
We were also able to detect substantial levels of anti-GFP ELISAantibodies (1:1280–1:5120) in the serum samples of HAd-GFP, PAd-GFP, or BAd-GFP-inoculated mice at 16 day post-inoculation (Table 1).Similar levels of neutralizing antibodies against HAd5, PAd3, or BAd3vector were also observed in serum samples collected at 16 days post-inoculation (Table 1) suggesting that various adenoviral vectors wereequally capable of inducing vector-specific immunity.
Levels and duration of the early (E4) and late (hexon) gene expression invarious tissues of mice inoculated with HAd-GFP, PAd-GFP, or BAd-GFP
Expression of vector genes at lower levels (“leaky gene expres-sion”) has been reported in E1-deleted replication defective vectors
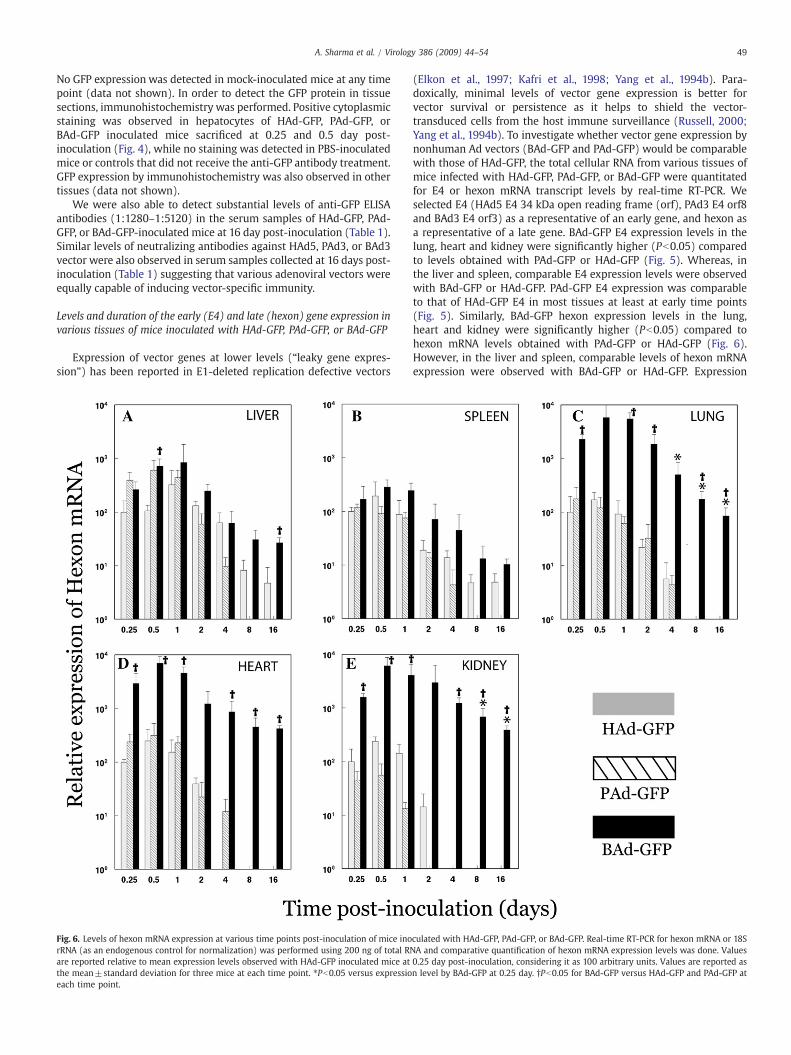
Fig. 6. Levels of hexon mRNA expression at various time points post-inoculation of mice inorRNA (as an endogenous control for normalization) was performed using 200 ng of total Rare reported relative to mean expression levels observed with HAd-GFP inoculated mice atthe mean±standard deviation for three mice at each time point. ⁎Pb0.05 versus expressioeach time point.
(Elkon et al., 1997; Kafri et al., 1998; Yang et al., 1994b). Para-doxically, minimal levels of vector gene expression is better forvector survival or persistence as it helps to shield the vector-transduced cells from the host immune surveillance (Russell, 2000;Yang et al., 1994b). To investigate whether vector gene expression bynonhuman Ad vectors (BAd-GFP and PAd-GFP) would be comparablewith those of HAd-GFP, the total cellular RNA from various tissues ofmice infected with HAd-GFP, PAd-GFP, or BAd-GFP were quantitatedfor E4 or hexon mRNA transcript levels by real-time RT-PCR. Weselected E4 (HAd5 E4 34 kDa open reading frame (orf), PAd3 E4 orf8and BAd3 E4 orf3) as a representative of an early gene, and hexon asa representative of a late gene. BAd-GFP E4 expression levels in thelung, heart and kidney were significantly higher (Pb0.05) comparedto levels obtained with PAd-GFP or HAd-GFP (Fig. 5). Whereas, inthe liver and spleen, comparable E4 expression levels were observedwith BAd-GFP or HAd-GFP. PAd-GFP E4 expression was comparableto that of HAd-GFP E4 in most tissues at least at early time points(Fig. 5). Similarly, BAd-GFP hexon expression levels in the lung,heart and kidney were significantly higher (Pb0.05) compared tohexon mRNA levels obtained with PAd-GFP or HAd-GFP (Fig. 6).However, in the liver and spleen, comparable levels of hexon mRNAexpression were observed with BAd-GFP or HAd-GFP. Expression
culated with HAd-GFP, PAd-GFP, or BAd-GFP. Real-time RT-PCR for hexon mRNA or 18SNA and comparative quantification of hexon mRNA expression levels was done. Values0.25 day post-inoculation, considering it as 100 arbitrary units. Values are reported asn level by BAd-GFP at 0.25 day. †Pb0.05 for BAd-GFP versus HAd-GFP and PAd-GFP at

50 A. Sharma et al. / Virology 386 (2009) 44–54
levels of hexon mRNA of PAd-GFP were comparable to those of HAd-GFP E4 in most tissues at least at early time points (Fig. 6). Noexpression of either E4 or hexon was observed in mock-inoculatedmice (data not shown). Overall, it seemed that levels of the earlyand the late gene expression by HAd-GFP, PAd-GFP, or BAd-GFP werecomparable taking into consideration the number of vector genomesin different tissues at various time points post-inoculation.
Discussion
HAd vectors have demonstrated tremendous potential as a genedelivery vehicle for vaccine and gene therapy applications. However,the prevalence of pre-existing vector immunity in the majority ofhuman population and their predominant hepatotropism may limittheir utility. With the anticipation that nonhuman Ad vectors maycircumvent these limitations, we have developed PAd3 and BAd3vectors (Bangari and Mittal, 2004; Mittal et al., 1995b). In this study,we evaluated their biodistribution, persistence, state of vectorgenome, and transgene and vector gene expression to further explorethe potential of these nonhuman Ad vectors for gene delivery.
In our study, biodistribution patterns of HAd-GFP in the liver,spleen, heart, lung, and kidney were similar to the previous findings(Alemany and Curiel, 2001; Ni et al., 2005; Stone et al., 2007; Woodet al., 1999; Zinn et al., 1998). PAd-GFP localized predominantly in theliver and spleen similar to HAd-GFP. BAd-GFP showed distinct tropismas indicated by significantly (Pb0.05) higher levels of vectorlocalization to the heart, kidney, and lung compared to PAd3 andHAd5 vectors, whereas levels of BAd-GFP vector genome in the liverand spleenwere at levels comparable to those of PAd-GFP or HAd-GFPat least at earlier time points. The distinct tissue tropismmay partly bedue to utilization of different receptor(s) by BAd3 (Bangari et al.,2005a). PAd3 and BAd3 vectors internalization is CAR-independentand there are differences in the transduction efficiency of various celllines by these vectors compared to HAd5 (Bangari and Mittal, 2005;Bangari et al., 2005a, 2005b). Recombinant fiber knob of HAd5, PAd3,or BAd3 inhibited cell entry of only homologous virus suggesting thatthese three viruses use distinct receptors for internalization (Bangariet al., 2005a). In addition to CAR, integrins play an important role asco-receptor in HAd5 entry and hence are likely to be a determinant invector biodistribution. Integrin-binding motifs such as Arg–Gly–Asp(RGD) and Leu–Asp–Val (LDV) are also absent in the penton base ofBAd3 and PAd3 (Reddy et al., 1998) and we have demonstrated earlierthat BAd3 or PAd3 internalization was independent of αvβ3 or αvβ5integrins (Bangari et al., 2005a).
Despite advancements in the understanding of Ad interactionswith host cell receptors, the exact mechanism of tissue tropismremains unclear. Various hypotheses have been put forward to explainthe predominant liver tropism exhibited by many Ad vectors. Some ofthe non-CAR binding Ad vectors have also been demonstrated topredominately localize to the liver, suggesting that factors other thanthe fiber knob–CAR interaction are also involved in mediatinghepatotropism (Alemany and Curiel, 2001; Smith et al., 2002). CAR-independent uptake of CAR-binding Ad vectors by Kupffer cells orhepatocytes has been shown (Shayakhmetov et al., 2004). Involve-ment of certain blood factors (such as coagulation factor IX andcomplement components) has been proposed to explain Ad vectorhepatotropism. These factors cross-link Ad vectors with heparansulphate proteoglycans (HSPGs) on hepatocytes and thus leading topreferential hepatotropism. Therefore, modification of certain sol-vent-exposed loops in the fiber knob domain of the Ad vector hasresulted in reduced vector uptake in the liver (Shayakhmetov et al.,2005). Recent studies have demonstrated that HAd5 uptake in theliver is mediated by binding of coagulation factor X (FX) to hexon(Kalyuzhniy et al., 2008; Waddington et al., 2008). Moreover, therewere significant variations among HAd serotypes to bind to FX, thatcorrelated with their ability to transduce hepatocytes. Studies
investigating interaction of FX and/or other blood factors with PAd3or BAd3 capsid proteins would further clarify their role in tropism andbiodistribution of these vectors. HSPG-binding motif (KKTK) in thefiber shaft of HAd5 has also been proposed to drive hepatotropism asdeletion of this motif led to significant decline in hepatic tissuetransduction (Nicol et al., 2004; Smith et al., 2003a, 2003b). Further-more, the size and structure of fiber may play a role in determiningtissue tropism. Ad vectors with short fibers, like HAd35 and HAd40,escape hepatic sequestration in contrast to vector with large fibers(Nakamura et al., 2003; Shayakhmetov et al., 2004). However, achimeric HAd5 vector with ovine Ad-derived longshafted fiberdemonstrated CAR-independence with no particular preference tothe liver (Nakayama et al., 2006). Nicklin et al. (2005) suggested thatthe Ad fiber, rather than knob–receptor interaction may contribute tohepatotropism. BAd3 fiber has a very long shaft with few kinks in itssecondary structure and is devoid of (KKTK) motif (Ruigrok et al.,1994). Though the knob domain has been suggested to be the primaryreceptor-seeking moiety of most Mastadenovirus species (Nicklinet al., 2005), there is a possibility that fiber shaft may also play animportant role in BAd3 or PAd3 internalization.
It has been demonstrated that at low doses, Ad vectors werepredominately sequestered by Kupffer cells resulting in suboptimaltransduction of other cell types (Tao et al., 2001). However, the role ofsequestration of Ad vectors by Kupffer cells on vector biodistributioncan be evaluated effectively only at low vector doses. Since there wasno information available on biodistribution of BAd3 and PAd3 vectorscompared to HAd5, we chose a comparatively lower vector dose (1010
VP/mouse) for this study. Our results suggest that sequestration ofBAd3 vector by Kupffer cells did not significantly influenced vectorbiodistribution. Subsequent studies with higher vector doses shouldfurther clarify the influence of reticuloendothelial system on trans-duction efficiencies of these vectors.
BAd-GFP persisted at significantly (Pb0.05) higher levels in alltissues even at 16 days post-inoculation compared to PAd-GFP orHAd-GFP. It is probably due to better survival of BAd-GFP genome invarious tissues because of slow degradation and/or delayed removalof cells transduced with the vector genome. PAd-GFP had a similarbiodistribution pattern as HAd-GFP but was present at lower levels inall tissues and its persistence in various tissues was for a shorterduration compared to HAd-GFP or BAd-GFP. Whether it is due topaucity of PAd3 receptors in FVB/n mouse tissues is not clear.Experiments to investigate the induction of a host innate and adaptiveimmune response following intravenous administration of thesevectors are underway.
Because integration of the vector genome may lead to insertionalmutagenesis, we examined the state of the vector genome in the liverof mice inoculated with HAd-GFP, PAd-GFP, or BAd-GFP. Vectorgenome could be present within the cell nucleus as a linear episomalform, circular episomal form, or integrated into the host genome(Ehrhardt et al., 2008). It has been reported that the majority of HAd5genome in transduced cells persists in a episomal form (Ehrhardtet al., 2003). However, a fraction of the vector genome can also getintegrated into the host chromosomal DNA (Harui et al., 1999). Inaddition, the genomes of some HAd serotypes such as HAd12frequently integrate into the host cell genome mostly by randomintegration (Green et al., 1977; Knoblauch et al., 1996). Circularizedvector genomemay also replicate to some extent on its own leading toits enhanced persistence (Kreppel and Kochanek, 2004). In this study,only the linear episomal form of HAd-GFP, PAd-GFP, or BAd-GFPgenome was detected suggesting that the majority of nonhuman Adgenome remained as linear episome similar to the HAd-GFP genome.BAd-GFP genome as identified by Southern blot assay persisted athigher levels up to 8-day-post-inoculation, further corroborating ourresults of vector biodistribution and persistence. Additional in vitroexperiments are needed to further explore the state of nonhuman Advector genome.

Table 2Sequence of primers and Taqman® probes for real-time PCR/RT-PCR for quantificationof vector genome and GFP, E4, or hexon mRNA transcripts
Target Primers and probes
GFP Forward primer — 5′-TGGTCCCAATTCTCGTGGAA-3′Reverse primer — 5′-CCTCTCCGCTGACAGAAAATTT-3′Probe — 5′-TGGATGGCGATGTGAATGGGCA-3′
HAd5 hexon Forward primer — 5′-CCTACTCTGGCACTGCCTACAA-3′Reverse primer — 5′-CATCCCATTCGCAAGGATTT-3′Probe — 5′-CCTGGCTCCCAAGGGTGCCC-3′
PAd3 hexon Forward primer — 5′-GAGACCACCATCGATCCTTTG-3′Reverse primer — 5′-TGTCTGACCACGAGCTTGGA-3′Probe — 5′-ATGAGCCCCAACCCCAGCTTG-3′
BAd3 hexon Forward primer —5′-CGGCACGGCCTACAACTC-3′Reverse primer — 5′-GCCTGCCTAAACTGCGTATTG-3′Probe — 5′-TTTGCCCCCAACAGTGCCC-3′
HAd5 E4 (34 kDaorf)
Forward primer — 5′-CCTGCGCTTGTGGTATGATG-3′Reverse primer — 5′-GCGCTGTATCCAAAGCTCATG-3′Probe — 5′-CCACGTGGGTTCTGTGGTCCCC-3′
PAd3 E4 (orf 8) Forward primer — 5′-TTCAGGGAGGCGGTGAAC-3′Reverse primer — 5′-GCGCCTCGAAACATCACACT-3′Probe — 5′-TGCCCAGCGTGAGTCGCGTG-3′
BAd3 E4 (orf 3) Forward primer — 5′-GGGCGAGCAATCAGCTCTTA-3′Reverse primer — 5′-CTAATCCACTGCCCATGTACACA-3′Probe —5′-AGTCCCTGCCCACTTTTGCCTGG-3′
18S rRNA Eukaryotic 18S rRNA Endogenous Control (Catalog number —4352930E, Applied Biosystems, Foster city, CA)
The probes were labeled with 6-carboxyfluorescein (FAM) at the 5′ end and with minorgroove binder (MGB) at the 3′ end.
51A. Sharma et al. / Virology 386 (2009) 44–54
Efficient gene delivery and robust expression of transgene is one ofthe attributes of first generation HAd5 vectors. It was expected fromour vector biodistribution and persistence study that GFP transgeneexpression by BAd-GFP should be better than that of HAd-GFP orPAd-GFP. Comparable transgene expression, proportionate to thelevels of vector genome present in the tissue, was observed.Significantly higher (Pb0.05) levels of transgene expression weredetected in the heart, kidney, and lung from BAd-GFP-inoculated miceand similar levels in the liver and spleen as compared to levelsobtained with HAd-GFP.
Proteins encoded by the E1 region are required for the transactiva-tion of other vector early and late genes (Russell, 2000), therefore, thefirst generation E1-deleted Ad vectors are replication defective.However, leaky expression of the early and late genes of such vectorshas been reported even in the absence of E1 proteins expression(Elkon et al., 1997; Kafri et al., 1998; Yang et al., 1994b). The leakyvector gene expression is strong enough to induce vector-specificcytotoxic T lymphocyte response leading to the removal of vector-transduced cells. Unlike HAd-GFP in which the entire E1 region wasdeleted, only E1A regions were deleted in PAd-GFP and BAd-GFP(Bangari and Mittal, 2004; Bangari et al., 2005a). Partial deletion inthe E1 region of PAd-GFP and BAd-GFP was found to be sufficient torender these vectors replication defective (Bangari and Mittal, 2004;Reddy et al., 1999a; van Olphen et al., 2002). In the present study,expression of both vector E4 (early) and hexon (late) genes wasobserved by all the three vectors at levels that correspond to theamount of vector present in various tissues.
Our results in FVB/n mouse model have strongly suggested thatBAd3-based vectors have considerable potential as gene deliveryvector for vaccine and gene therapy. Our recent study involving aBAd-vector based vaccine in BALB/c mice resulted in significantlyhigher levels of humoral and cell-mediated immune responsescompared to the HAd5-based vaccine (Singh et al., 2008) suggestingthat the results obtained with FVB/n mouse model are applicable toanother mouse strain. The results presented here have prepared thefoundation for additional studies (with higher vector dose and forlonger duration post-inoculation) for evaluating vector toxicity, andinnate and adaptive immune responses following systemic delivery ofnonhuman Ad vectors.
Materials and methods
Adenoviral vectors
Replication-defective HAd-GFP (Bangari and Mittal, 2004), PAd-GFP (Bangari and Mittal, 2004) and BAd-GFP (Bangari et al., 2005b)vectors with deletions in E1 or E1A region and carrying the GFPgene under the control of the cytomegalovirus (CMV) promoterwere propagated as described previously. HAd-GFP, PAd-GFP, andBAd-GFP vectors were grown and titrated in 293 (human embryonickidney cells expressing HAd E1) (Graham et al., 1977), FPRT HE1(fetal porcine retina cells expressing HAd E1) (Bangari and Mittal,2004) and FBRT HE1 (fetal bovine retina cells expressing HAd E1)(van Olphen et al., 2002), respectively. The virus purification wasdone by cesium chloride-density gradient centrifugation as pre-viously described (Bangari and Mittal, 2004). The physical particlecounts of purified HAd-GFP, PAd-GFP and BAd-GFP vectors wereestimated by spectrophotometry and expressed as vector particles(VP) per ml following a previously described protocol (Grahamand Prevec, 1995). The VP/plaque forming units (p.f.u) ratio forHAd-GFP, PAd-GFP and BAd-GFP were 72, 100 and 608 respec-tively. Since plaque assays for these vectors were carried out indifferent cell lines, efficiency of plaques formation varied with thevirus and cell type combination. Therefore, VP was selected as themajor criteria for vector quantification to maintain consistency withvector dosage.
Animal inoculation
Eight-to-ten-week-old female FVB/n mice were obtained fromHarlan Laboratories (Indianapolis IN). FVB/n mice were selected forthe current study because of the availability of an immunocompetenttumor model using this strain (Addison et al., 1995; Noblitt et al.,2005). The use of this strain of mice would allow us to extend ourstudy for investigation of HAd5, PAd3 and BAd3 vectors in cancer genetherapy. All animal inoculations were conducted in accordance withthe guidelines and approval from Institutional Biosafety Committeeand Institutional Animal Care and Use Committee. Mice wereinoculated intravenously through tail vein with HAd-GFP, PAd-GFP,or BAd-GFP at a dose of 1010 VP per mouse in a volume of 100 μl ofPBS++ (Phosphate buffer saline supplemented with 0.01% MgCl2and 0.01% CaCl2). Mock-inoculated mice served as negative controls.Mice (3 animals per group) were sacrificed at various time points(0.25, 0.5, 1, 2, 4, 8 and 16 days) post-inoculation. The liver, spleen,lung, heart, and kidney were collected and stored at −80 °C. Thesetissue samples were used for total cellular DNA or RNA extraction forvarious assays. Serum samples were also collected for the analysis ofAd-specific neutralizing antibody.
Primers and Taqman probes
A set of primer pair and a Taqman® probe specific to the GFP genein HAd-GFP, PAd-GFP, or BAd-GFPwere designed using Primer Express2.0 software (Applied Biosystems, Foster City, CA). These primers andprobe were used for determining the biodistribution of Ad vectors byquantification of vector genomes by real-time PCR, and analyzingtransgene expression by determining expression levels of GFP mRNAby real-time RT-PCR.
Similarly, sets of primer pairs and Taqman® probes specific to thehexon gene or an E4 gene (HAd5 E4 34 kDa orf, PAd3 E4 orf 8 and BAd3E4 orf 3) of each vector were designed for quantification of their hexonor E4 mRNA transcripts by real-time RT-PCR. All primer sets andTaqman® probes were synthesized by Applied Biosystems. Thesequences of various primers and Taqman® probes are in Table 2.All probes were labeledwith 6-carboxyfluorescein (FAM) at the 5′ endand with minor groove binder (MGB) at the 3′ end.

52 A. Sharma et al. / Virology 386 (2009) 44–54
Quantification of vector genome
Total genomic DNA from 50mg of tissue samples was isolated usingDNeasy kit (Qiagen Inc., Valencia, CA) as per the manufacturer'sguidelines. Initially, standard curves were generated using 10-folddilutions (from3 to 3×108 copies) of purifiedgenomicDNAofHAd-GFP,PAd-GFP, or BAd-GFP as a template for real-time PCR using GFP primersand probe. The copy number of each vector genome was calculatedbased on spectrophotometric quantification and molecular mass ofvector genomic DNA. Subsequently, these dilutions of each vector wererun as standardswith each set of real-timePCR assay. DNA samples frommock-inoculated mice tissues served as negative controls. For quanti-fication of the vector genome by real-time PCR using the GFP primersand probe, 50 ng of the total cellular DNA was used in a 25 μl reactionusing Taqman® PCR core reagents (Applied Biosystems). The reactionmixture contained 10×Taqman® buffer, 250 nM each of forward andreverse primers, and 100 nM of Taqman® probe along with otherstandard kit components. Each reactionwas carriedout induplicate. Thereal-time PCR was performed using the Mx3000 Thermocycler(Stratagene, Cedar Creek, TX). The reaction conditions includedincubation at 50 °C for 2 min, followed by polymerase activation(95 °C for 10 min), and 45 cycles of denaturation (95 °C for 15 s) andannealing/extension (60 °C for 1min). The threshold cycle (Ct) value forindividual reactions was determined and data were analyzed withMxPro software (Stratagene) to obtain the absolute copy number ofviral genome per 50 ng of total cellular DNA.
Quantification of GFP, E4, or hexon mRNA transcripts
Total cellular RNA was isolated from 50 mg of each tissue sampleusing RNA miniprep kit (Stratagene). RNA samples were treated withDNase I to remove the residual DNA. 200 ng of total cellular RNA wasprocessed for real-time RT-PCR using GFP-, E4-, or hexon-specificprimers and probe and One-step Brilliant QRT-PCR Master Mix Kit(Stratagene). For normalization of the target gene expression, similarreal-time RT-PCR reactions targeting the endogenous 18S rRNA weresimultaneously carried out in separate tubes. Quantification of expres-sion levels of GFP, E4, or hexon mRNA transcripts in various tissuesampleswasdonebyΔΔCtmethod (Winer et al.,1999) andexpressed inrelation to the mean expression levels of respective genes observed byHAd-GFP at 0.25 day post-inoculation (referred to as calibrator). Folddifference in expression levels in relation to calibrator was calculated as2−ΔΔCt (where ΔΔCt=[Cttarget gene(unknown)−Ct18S rRNA(unknown)]− [Cttarget gene(calibrator)−Ct18S rRNA (calibrator)], Ct is the cyclenumber at which fluorescence signal crosses the threshold). Forcalculation of arbitrary units, the mean expression levels of mRNAtranscripts in HAd-GFP-inoculated mice tissues at 6 h post-inoculationwere considered as 100 arbitrary units. Reaction mixture consisted of2×QRT-PCRmastermix, 250 nMeach of respective forward and reverseprimers, and 100 nM of Taqman® probe along with other standard kitcomponents. Each reaction was carried out in duplicate. The real-timeRT-PCR was performed using the Mx3000 Thermocycler. The reactionconditions included cDNA synthesis step at 50 °C for 30min, followed bypolymerase activation (95 °C for10 min), and 45 cycles of denaturation(95 °C for 15 s) and annealing/extension (60 °C for 1min). The Ct valuesfor individual reactions were determined and data were analyzed withMxPro software to obtain the relative expression levels of GFP, E4, orhexon mRNA transcripts. All RNA samples were also used for PCR assayusing GFP, E4, or hexon primers and probe to ensure the absence of theresidual DNA.
Southern blot analysis
Total cellular DNA from the livers of mice inoculated with HAd-GFP, PAd-GFP, BAd-GFP, or PBS was isolated by proteinase-K digestionand isopropanol precipitation as per previously described method
(Sambrook and Russell, 2001). 20 μg of the total cellular DNA wasdigested with KpnI (for HAd-GFP inoculated mice), XhoI (for PAd-GFPinoculated mice), or HpaI (for BAd-GFP inoculated mice) and loadedon to a 1% agarose gel. DNA from the liver of mock-inoculated miceand purified vector DNA were digested similarly and used as negativeand positive controls, respectively. After agarose gel electrophoresis,the separated DNA fragments were transferred on to a positivelycharged nylon membrane (Hybond-N+, GE Healthcare Ltd., LittleChalfront, UK) by upward capillary transfer method (Sambrook andRussell, 2001). Following the transfer the DNA fragments were cross-linked to the membrane by baking at 80 °C for 2 h. DNA hybridizationwas performed using HAd5 (nt 34,524–35,537), PAd3 (nt 32,353–33,582) or BAd3 (nt 32,520–33,615) specific probes which werechosen specifically from the area near the right terminus of the vectorgenome to aid in identification of the linear, circular, or integratedform of the vector genome. The probes were prepared by PCRamplification and radiolabeled using [alpha32P]dCTP (MP Biomedi-cals, Solon, OH) by random priming (DECAprime II kit, AppliedBiosystems). The probes were column purified (NucAway spincolumns, Applied Biosystems) and used for DNA hybridization at65 °C overnight in Danhardt's buffer (Sambrook and Russell, 2001).The membrane was washed twice each with 2×SSC (0.3 M sodiumchloride, 0.03M sodium citrate), 0.1% SDS (Sodium dodecyl sulphate);1×SSC, 0.1% SDS and 0.1×SSC, 0.1% SDS, wrapped with saran wrapand exposed to Cyclone Storage Phosphor Screen (Packard InstrumentCompany, Meridin, CT). After 2 h of exposure, signals were visualizedby Cyclone Storage Phosphor System (Packard Instrument Company).
Immunohistochemistry
Formalin-fixed and paraffin-embedded tissues were deparaffi-nized in xylene and rehydrated using graded dilutions of ethanolaccording to standard immunohistochemistry procedures. The slideswere immersed in target retrieval solution (DakoCytomation, Carpin-teria, CA) at 95 °C for 15 min to retrieve antigens. Endogenousperoxidase activity was quenched by immersing slides in 3% hydrogenperoxide solution for 5 min. Endogenous biotin and avidin wereblocked by incubating sections in avidin/biotin blocking reagents(Vector Laboratories, Burlingame, CA) for 15 min. The sections wereincubated in blocking solution (MOM immunodetection kit, VectorLaboratories) according to manufacturer's instruction. The sectionswere then incubated with a monoclonal anti-GFP antibody (MilliporeCorporation, Billerica, MA) at 1:500 dilution for 30 min at roomtemperature followed by a 1:250 dilution of biotinylated anti-mousesecondary antibody (Vector Laboratories) for 10 min. This step wasfollowed by incubation with streptavidin–horseradish peroxidaseconjugate (DakoCytomation) for 30 min. Signal amplification wasperformed by Tyramide Signal Amplification Kit (PerkinElmer,Waltham, MA). The color development was performed by AminoethylCarbazole (Red) Substrate Kit (Zymed Laboratories Inc., San Francisco,CA). The sections were thoroughly rinsed with tris-buffered saline(100 mM Tris–HCl, pH 7.5 and 150 mM sodium chloride) in betweeneach of the above mentioned steps. Counter staining was performedby dipping slides in hematoxylin solution. The slides were thenthoroughly rinsed with water, mounted with Clearmount solution(Zymed Laboratories Inc.) and left to drying. The slides were thencover slipped with Permount (Fisher Scientific, Pittsburgh, PA).
Detection of Ad-neutralizing antibodies and anti-GFP ELISA antibodies
Serum samples from vector- or PBS-inoculated mice sacrificed at16 days post-inoculationwere collected and tested for the presence ofHAd5-, PAd3-, or BAd3-neutralizing antibodies, as described pre-viously (Moffatt et al., 2000). Similarly, the serum samples were alsoused to detect GFP-specific antibodies by ELISA as previouslydescribed (Mittal et al., 1995a). Briefly, 96-well ELISA plates (Immulon

53A. Sharma et al. / Virology 386 (2009) 44–54
2HB, Thermo Scientific, Asheville, NC) were coated with a purifiedpreparation of GFP (1 μg/ml) in carbonate bicarbonate buffer and thenreacted with serial 2-fold dilutions of each of the test serum samples.Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Bio-Rad, Hercules, CA) was used as secondary antibody. The opticaldensity (OD) was measured at 450 nm or 490 nm using an ELISAreader (Molecular Devices, Sunnyvale, CA). The reciprocal of thehighest serum dilution with an OD reading of at least the mean+2standard deviation (SD) above the PBS-inoculated mouse serumsample was taken as the ELISA antibody titer.
Statistical analyses
Student's paired t-tests for multiple comparisons with adjust-ments in the P-value using Tukey or Bonferroni methods were usedfor determining the statistical significance. For all tests, Pb0.05 wasconsidered significant.
Acknowledgments
This work was supported by Public Health Service grant CA110176from the National Cancer Institute. We are thankful to Jane Kovach forher excellent secretarial assistance and Brian Denton for help withstatistical analyses.
References
Addison, C.L., Braciak, T., Ralston, R., Muller, W.J., Gauldie, J., Graham, F.L., 1995.Intratumoral injection of an adenovirus expressing interleukin 2 induces regressionand immunity in a murine breast cancer model. Proc. Natl. Acad. Sci. U.S.A. 92,8522–8526.
Alemany, R., Curiel, D.T., 2001. CAR-binding ablation does not change biodistributionand toxicity of adenoviral vectors. Gene Ther. 8, 1347–1353.
Bangari, D.S., Mittal, S.K., 2004. Porcine adenoviral vectors evade preexisting humoralimmunity to adenoviruses and efficiently infect both human and murine cells inculture. Virus Res. 105, 127–136.
Bangari, D.S., Mittal, S.K., 2005. Porcine adenovirus serotype 3 internalization isindependent of CAR and alphavbeta3 or alphavbeta5 integrin. Virology 332, 157–166.
Bangari, D.S., Mittal, S.K., 2006a. Current strategies and future directions for eludingadenoviral vector immunity. Curr. Gene Ther. 6, 215–226.
Bangari, D.S., Mittal, S.K., 2006b. Development of nonhuman adenoviruses as vaccinevectors. Vaccine 24, 849–862.
Bangari, D.S., Sharma, A., Mittal, S.K., 2005a. Bovine adenovirus type 3 internalization isindependent of primary receptors of human adenovirus type 5 and porcineadenovirus type 3. Biochem. Biophys. Res. Commun. 331, 1478–1484.
Bangari, D.S., Shukla, S., Mittal, S.K., 2005b. Comparative transduction efficiencies ofhuman and nonhuman adenoviral vectors in human, murine, bovine, and porcinecells in culture. Biochem. Biophys. Res. Commun. 327, 960–966.
Bergelson, J.M., Cunningham, J.A., Droguett, G., Kurt-Jones, E.A., Krithivas, A., Hong, J.S.,Horwitz, M.S., Crowell, R.L., Finberg, R.W., 1997. Isolation of a common receptorfor Coxsackie B viruses and adenoviruses 2 and 5. Science 275, 1320–1323.
Edelstein, M.L., Abedi, M.R., Wixon, J., Edelstein, R.M., 2004. Gene therapy clinical trialsworldwide 1989–2004—an overview. J. Gene Med. 6, 597–602.
Ehrhardt, A., Xu, H., Kay, M.A., 2003. Episomal persistence of recombinant adenoviralvector genomes during the cell cycle in vivo. J. Virol. 77, 7689–7695.
Ehrhardt, A., Haase, R., Schepers, A., Deutsch, M.J., Lipps, H.J., Baiker, A., 2008. Episomalvectors for gene therapy. Curr. Gene Ther. 8, 147–161.
Elkon, K.B., Liu, C.C., Gall, J.G., Trevejo, J., Marino, M.W., Abrahamsen, K.A., Song, X., Zhou,J.L., Old, L.J., Crystal, R.G., Falck-Pedersen, E., 1997. Tumor necrosis factor alphaplays a central role in immune-mediated clearance of adenoviral vectors. Proc. Natl.Acad. Sci. U.S.A. 94, 9814–9819.
Farina, S.F., Gao, G.P., Xiang, Z.Q., Rux, J.J., Burnett, R.M., Alvira, M.R., Marsh, J., Ertl, H.C.,Wilson, J.M., 2001. Replication-defective vector based on a chimpanzee adenovirus.J. Virol. 75, 11603–11613.
Glasgow, J.N., Kremer, E.J., Hemminki, A., Siegal, G.P., Douglas, J.T., Curiel, D.T., 2004.An adenovirus vector with a chimeric fiber derived from canine adenovirus type 2displays novel tropism. Virology 324, 103–116.
Graham, F.L., Prevec, L., 1995. Methods for construction of adenovirus vectors. Mol.Biotechnol. 3, 207–220.
Graham, F.L., Smiley, J., Russell, W.C., Nairn, R., 1977. Characteristics of a human cell linetransformed by DNA from human adenovirus type 5. J. Gen. Virol. 36, 59–74.
Green, M.R., Mackey, J.K., Green, M., 1977. Multiple copies of human adenovirus 12genomes are integrated in virus-induced hamster tumors. J. Virol. 22, 238–242.
Harui, A., Suzuki, S., Kochanek, S., Mitani, K., 1999. Frequency and stability ofchromosomal integration of adenovirus vectors. J. Virol. 73, 6141–6146.
Hofmann, C., Loser, P., Cichon, G., Arnold, W., Both, G.W., Strauss, M., 1999. Ovineadenovirus vectors overcome preexisting humoral immunity against humanadenoviruses in vivo. J. Virol. 73, 6930–6936.
Kafri, T., Morgan, D., Krahl, T., Sarvetnick, N., Sherman, L., Verma, I., 1998. Cellularimmune response to adenoviral vector infected cells does not require de novo viralgene expression: implications for gene therapy. Proc. Natl. Acad. Sci. U.S.A. 95,11377–11382.
Kalyuzhniy, O., Di Paolo, N.C., Silvestry, M., Hofherr, S.E., Barry, M.A., Stewart, P.L.,Shayakhmetov, D.M., 2008. Adenovirus serotype 5 hexon is critical for virusinfection of hepatocytes in vivo. Proc. Natl. Acad. Sci. U.S.A. 105, 5483–5488.
Kanerva, A., Mikheeva, G.V., Krasnykh, V., Coolidge, C.J., Lam, J.T., Mahasreshti, P.J.,Barker, S.D., Straughn, M., Barnes, M.N., Alvarez, R.D., Hemminki, A., Curiel, D.T.,2002. Targeting adenovirus to the serotype 3 receptor increases gene transferefficiency to ovarian cancer cells. Clin. Cancer Res. 8, 275–280.
Knoblauch, M., Schroer, J., Schmitz, B., Doerfler, W., 1996. The structure of adenovirustype 12 DNA integration sites in the hamster cell genome. J. Virol. 70, 3788–3796.
Kremer, E.J., Boutin, S., Chillon, M., Danos, O., 2000. Canine adenovirus vectors: analternative for adenovirus-mediated gene transfer. J. Virol. 74, 505–512.
Kreppel, F., Kochanek, S., 2004. Long-term transgene expression in proliferatingcells mediated by episomally maintained high-capacity adenovirus vectors. J. Virol.78, 9–22.
McConnell, M.J., Imperiale, M.J., 2004. Biology of adenovirus and its use as a vectorfor gene therapy. Hum. Gene Ther. 15, 1022–1033.
Michou, A.I., Lehrmann, H., Saltik, M., Cotten, M., 1999. Mutational analysis of theavian adenovirus CELO, which provides a basis for gene delivery vectors. J. Virol.73, 1399–1410.
Mittal, S.K., Middleton, D.M., Tikoo, S.K., Babiuk, L.A., 1995a. Pathogenesis andimmunogenicity of bovine adenovirus type 3 in cotton rats (Sigmodon hispidus).Virology 213, 131–139.
Mittal, S.K., Prevec, L., Graham, F.L., Babiuk, L.A., 1995b. Development of a bovineadenovirus type 3-based expression vector. J. Gen. Virol. 76, 93–102.
Moffatt, S., Hays, J., HogenEsch, H., Mittal, S.K., 2000. Circumvention of vector-specificneutralizing antibody response by alternating use of human and non-humanadenoviruses: implications in gene therapy. Virology 272, 159–167.
Nakamura, T., Sato, K., Hamada, H., 2003. Reduction of natural adenovirus tropism tothe liver by both ablation of fiber-coxsackievirus and adenovirus receptorinteraction and use of replaceable short fiber. J. Virol. 77, 2512–2521.
Nakayama, M., Both, G.W., Banizs, B., Tsuruta, Y., Yamamoto, S., Kawakami, Y., Douglas,J.T., Tani, K., Curiel, D.T., Glasgow, J.N., 2006. An adenovirus serotype 5 vectorwith fibers derived from ovine atadenovirus demonstrates CAR-independenttropism and unique biodistribution in mice. Virology 350, 103–115.
Ni, S., Bernt, K., Gaggar, A., Li, Z.Y., Kiem, H.P., Lieber, A., 2005. Evaluation ofbiodistribution and safety of adenovirus vectors containing group B fibers afterintravenous injection into baboons. Hum. Gene Ther. 16, 664–677.
Nicklin, S.A., Wu, E., Nemerow, G.R., Baker, A.H., 2005. The influence of adenovirus fiberstructure and function on vector development for gene therapy. Mol. Ther. 12,384–393.
Nicol, C.G., Graham, D., Miller, W.H., White, S.J., Smith, T.A., Nicklin, S.A., Stevenson, S.C.,Baker, A.H., 2004. Effect of adenovirus serotype 5 fiber and pentonmodifications onin vivo tropism in rats. Mol. Ther. 10, 344–354.
Noblitt, L.W., Bangari, D.S., Shukla, S., Mohammed, S., Mittal, S.K., 2005. Immunocom-petent mouse model of breast cancer for preclinical testing of EphA2-targetedtherapy. Cancer Gene Ther. 12, 46–53.
Raper, S.E., Chirmule, N., Lee, F.S., Wivel, N.A., Bagg, A., Gao, G.P., Wilson, J.M., Batshaw,M.L., 2003. Fatal systemic inflammatory response syndrome in a ornithinetranscarbamylase deficient patient following adenoviral gene transfer. Mol.Genet. Metab. 80, 148–158.
Reddy, P.S., Idamakanti, N., Song, J.Y., Lee, J.B., Hyun, B.H., Park, J.H., Cha, S.H., Bae, Y.T.,Tikoo, S.K., Babiuk, L.A.,1998. Nucleotide sequence and transcriptionmap of porcineadenovirus type 3. Virology 251, 414–426.
Reddy, P.S., Idamakanti, N., Babiuk, L.A., Mehtali, M., Tikoo, S.K., 1999a. Porcineadenovirus-3 as a helper-dependent expression vector. J. Gen. Virol. 80, 2909–2916.
Reddy, P.S., Idamakanti, N., Chen, Y., Whale, T., Babiuk, L.A., Mehtali, M., Tikoo, S.K.,1999b. Replication-defective bovine adenovirus type 3 as an expression vector.J. Virol. 73, 9137–9144.
Ruigrok, R.W., Barge, A., Mittal, S.K., Jacrot, B., 1994. The fibre of bovine adenovirus type3 is very long but bent. J. Gen. Virol. 75, 2069–2073.
Russell, W.C., 2000. Update on adenovirus and its vectors. J. Gen. Virol. 81, 2573–2604.Sambrook, J., Russell, D.W., 2001. Molecular Cloning: A Laboratory Manual, 3rd ed. Cold
Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.Seshidhar Reddy, P., Ganesh, S., Limbach, M.P., Brann, T., Pinkstaff, A., Kaloss, M., Kaleko,
M., Connelly, S., 2003. Development of adenovirus serotype 35 as a gene transfervector. Virology 311, 384–393.
Shayakhmetov, D.M., Li, Z.Y., Ni, S., Lieber, A., 2004. Analysis of adenovirus sequestrationin the liver, transduction of hepatic cells, and innate toxicity after injection of fiber-modified vectors. J. Virol. 78, 5368–5381.
Shayakhmetov, D.M., Gaggar, A., Ni, S., Li, Z.Y., Lieber, A., 2005. Adenovirus binding toblood factors results in liver cell infection and hepatotoxicity. J. Virol. 79, 7478–7491.
Singh, N., Pandey, A., Jayashankar, L., Mittal, S.K., 2008. Bovine adenoviral vector-basedH5N1 influenza vaccine overcomes exceptionally high levels of pre-existingimmunity against human adenovirus. Mol. Ther. 16, 965–971.
Smith, T., Idamakanti, N., Kylefjord, H., Rollence, M., King, L., Kaloss, M., Kaleko, M.,Stevenson, S.C., 2002. In vivo hepatic adenoviral gene delivery occurs indepen-dently of the coxsackievirus-adenovirus receptor. Mol. Ther. 5, 770–779.
Smith, T.A., Idamakanti, N., Marshall-Neff, J., Rollence, M.L., Wright, P., Kaloss, M., King,L., Mech, C., Dinges, L., Iverson, W.O., Sherer, A.D., Markovits, J.E., Lyons, R.M.,Kaleko, M., Stevenson, S.C., 2003a. Receptor interactions involved in adenoviral-mediated gene delivery after systemic administration in non-human primates.Hum. Gene Ther. 14, 1595–1604.

54 A. Sharma et al. / Virology 386 (2009) 44–54
Smith, T.A., Idamakanti, N., Rollence, M.L., Marshall-Neff, J., Kim, J., Mulgrew, K.,Nemerow, G.R., Kaleko, M., Stevenson, S.C., 2003b. Adenovirus serotype 5 fiber shaftinfluences in vivo gene transfer in mice. Hum. Gene Ther. 14, 777–787.
Stone, D., Ni, S., Li, Z.Y., Gaggar, A., DiPaolo, N., Feng, Q., Sandig, V., Lieber, A., 2005.Development and assessment of human adenovirus type 11 as a gene transfervector. J. Virol. 79, 5090–5104.
Stone, D., Liu, Y., Li, Z.Y., Tuve, S., Strauss, R., Lieber, A., 2007. Comparison of adenovirusesfrom species B, C, e, and f after intravenous delivery. Mol. Ther. 15, 2146–2153.
Tao, N., Gao, G.P., Parr, M., Johnston, J., Baradet, T., Wilson, J.M., Barsoum, J., Fawell, S.E.,2001. Sequestration of adenoviral vector by Kupffer cells leads to a nonlinear doseresponse of transduction in liver. Mol. Ther. 3, 28–35.
Tatsis, N., Ertl, H.C., 2004. Adenoviruses as vaccine vectors. Mol. Ther. 10, 616–629.van Olphen, A.L., Tikoo, S.K., Mittal, S.K., 2002. Characterization of bovine adenovirus
type 3 E1 proteins and isolation of E1-expressing cell lines. Virology 295, 108–118.Waddington, S.N., McVey, J.H., Bhella, D., Parker, A.L., Barker, K., Atoda, H., Pink, R.,
Buckley, S.M., Greig, J.A., Denby, L., Custers, J., Morita, T., Francischetti, I.M.,Monteiro, R.Q., Barouch, D.H., van Rooijen, N., Napoli, C., Havenga, M.J., Nicklin, S.A.,Baker, A.H., 2008. Adenovirus serotype 5 hexon mediates liver gene transfer. Cell132, 397–409.
Wickham, T.J., Mathias, P., Cheresh, D.A., Nemerow, G.R., 1993. Integrins alpha v beta 3and alpha v beta 5 promote adenovirus internalization but not virus attachment.Cell 73, 309–319.
Winer, J., Jung, C.K., Shackel, I., Williams, P.M., 1999. Development and validation of real-time quantitative reverse transcriptase-polymerase chain reaction for monitoringgene expression in cardiac myocytes in vitro. Anal. Biochem. 270, 41–49.
Wood, M., Perrotte, P., Onishi, E., Harper, M.E., Dinney, C., Pagliaro, L., Wilson, D.R., 1999.Biodistribution of an adenoviral vector carrying the luciferase reporter genefollowing intravesical or intravenous administration to a mouse. Cancer Gene Ther.6, 367–372.
Yang, Y., Ertl, H.C., Wilson, J.M., 1994a. MHC class I-restricted cytotoxic T lymphocytes toviral antigens destroy hepatocytes in mice infected with E1-deleted recombinantadenoviruses. Immunity 1, 433–442.
Yang, Y., Nunes, F.A., Berencsi, K., Furth, E.E., Gonczol, E., Wilson, J.M., 1994b. Cellularimmunity to viral antigens limits E1-deleted adenoviruses for gene therapy. Proc.Natl. Acad. Sci. U.S.A. 91, 4407–4411.
Zinn, K.R., Douglas, J.T., Smyth, C.A., Liu, H.G., Wu, Q., Krasnykh, V.N., Mountz, J.D., Curiel,D.T., Mountz, J.M., 1998. Imaging and tissue biodistribution of 99mTc-labeledadenovirus knob (serotype 5). Gene Ther. 5, 798–808.