coastal bacterial communities: their potential roles in ... · extend my appreciation to my...
TRANSCRIPT

COASTAL BACTERIAL COMMUNITIES:
THEIR POTENTIAL ROLES IN
DIMETHYLSULPHIDE (DMS)
PRODUCTION AND CORAL DEFENCE
By
FELICITY KUEK WEN IK
A thesis submitted in partial fulfilment of
the requirements for the degree of
Masters of Science (by Research)
Faculty of Engineering, Computing and Science
Swinburne University of Technology (Sarawak campus)
2014

P a g e | ii

P a g e | iii
Abstract
Little is known about the microbial communities in the South China Sea, especially
the eastern region and this study aims to expand our knowledge on the diversity of
culturable bacterial communities in this area. The Talang-Satang region is situated
off the coast of Sematan and is especially important as it is one of the most diverse
ecosystems found off Sarawak. Complex microbial communities are known to have
significant influence over coral reef ecosystems. Through isolation and
identification (16S rDNA) of native microbes from the open ocean, coral surface
mucus layer (SML), as well as the surrounding sediment and waters, we were able
to determine the species composition and abundance of the culturable bacteria in
the South China Sea (Kuching and Kota Kinabalu), the Celebes Sea (Semporna) and
the coral reef ecosystem (Talang-talang reef). Comparisons were made with
regards to physico-chemical parameters and bacterial communities. The diversity
of bacterial communities in these marine environments were analysed through
isolation and identification (16S rDNA) of culturable bacteria, as well as
preparation of clone libraries and subsequent restriction fragment length
polymorphism (RFLP). It was observed that although the majority of bacteria in
Kuching, Kota Kinabalu and Semporna are members of the Proteobacteria group,
the composition of bacterial communities in these three areas did vary
significantly, and the changes were also mirrored in physico-chemical differences.
There is also a clear distinction between the different species found in the different
parts of the reef system. Isolates found attached to the coral were mostly related to
Vibrio spp., presumably attached to the mucus from the water column and
surrounding sediment.
Cultures that were isolated from the SML are found to be closely related to
antibiotic producers with tolerance towards elevated temperatures and heavy
metal contamination. This specialized microbiota may be important for protecting
the corals from pathogens by occupying entry niches and/or through the
production of secondary metabolites (i.e. antibiotics). The role of the mucus-
associated bacteria for the defence of the coral was highlighted by the fact that
isolates related to pathogenic Vibrio spp. and Bacillus spp. were dominant amongst

P a g e | iv
the samples from the water column and sediment, and isolates with closest
matches to the known coral pathogens Vibrio coralliilyticus and Vibrio shiloi were
obtained from the SML and sediment samples respectively. The ability of isolates
living in the SML (associated) to inhibit isolates loosely attached to the SML
(attached) and vice versa was assessed at varying temperatures. All isolates were
also screened (using specific sets of primers) for the presence of type I modular
polyketides synthase (PKS) genes responsible for macrolide polyketides
production and non-ribosomal peptide synthetase (NRPS) genes with the ability to
produce immunosuppressants and other antibiotics. Our results indicate that the
mucus-associated bacteria display maximum efficacy to ward off other bacteria at
28 °C, however the inhibitory abilities of mucus-associated bacteria became less
effective as temperatures increased.
One major and globally important role of surface bacteria is their involvement in
the breakdown or osmoregulation of dimethylsulphoniopropionate (DMSP) to
dimethylsulfide (DMS) or methanethiol (MeSH). Using genomic-based studies,
enzymes responsible for DMSP degradation within the microbial community can
be identified and over 200 culturable bacteria were screened for the existence of
two key genes (dmdA, dddP) which are involved in competing, enzymatically
mediated DMSP degradation pathways. Roseobacter spp. which are mainly
responsible for the degradation of DMSP – a major source of oceans’ organic
sulphur – into MeSH were also successfully isolated from the SML. Bacterial DMSP
degraders may also contribute significantly to DMS production when temperatures
are elevated. This is to our knowledge the first comprehensive study looking at
culturable bacteria in the eastern South China Sea and their potential roles in coral
defence and the DMS(P) cycle.

P a g e | v
Acknowledgements
For since the creation of the world God’s invisible qualities – his eternal power and divine nature –
have been clearly seen, being understood from what has been made, so that people are without excuse.
(Romans 1:20)
Foremost, I would like to express my sincere gratitude to my principal
coordinating supervisor, Dr. Moritz Müller for his continuous support of my MSc
study and research, for his patience, motivation, enthusiasm, and immense
knowledge. Thank you for giving me the chance to explore this field, allowing me
freedom and space to make mistakes and for believing in me. I would also like to
extend my appreciation to my co-supervisors: Dr. Aazani Mujahid, Assoc. Prof. Dr.
Lim Po Teen, and Dr. Leaw Chui Pin, for their encouragements, insightful
comments, hard questions, as well as access to laboratories and facilities in
Universiti Malaysia Sarawak (UNIMAS).
Heartfelt thanks also to the Biotechnology laboratory officers and technicians:
Chua Jia Ni, Dyg. Rafika Atiqah and Nurul Arina, for allowing me to use the labs
past office hours and weekends, and for loaning me apparatus and experiment
materials when I needed them. Without your help, this project may not have been
completed on time.
A big thank you to my fellow lab mates and student helpers: Onn May Ling, Jessica
Fong, Lim Li Fang, and Ngu Lin Hui, for the stimulating discussions, the company
during long hours in the lab, the support during various existential crises and for
all the fun we have had in the last two years.
Last but not least, I would like to thank my family, especially my mother, for
encouraging me to take up this M.Sc. opportunity and for having my back
throughout every circumstance in the past two years.
I am grateful to the Sarawak Foundation for providing me with funding via the
Tunku Abdul Rahman Scholarship which enabled me to pursue this postgraduate
study.

P a g e | vi
Declaration
I hereby declare that this research entitled “Coastal Bacterial Communities: Their
Potential Roles in Dimethylsulphide (DMS) Production and Coral Defence” is
original and contains no material which has been accepted for the award to the
candidate of any other degree or diploma, except where due reference is made in
the text of the examinable outcome; to the best of my knowledge contains no
material previously published or written by another person except where due
reference is made in the text of the examinable outcome; and where work is based
on joint research or publications, discloses the relative contributions of the
respective workers or authors.
(FELICITY KUEK WEN IK)
Date: 9th September 2014
In my capacity as the Principal Coordinating Supervisor of the candidate’s thesis, I
certify that the above statements are true to the best of my knowledge.
(MORITZ MÜLLER)
Date: 9th September 2014.

P a g e | vii
Publications Arising from this Thesis
The work described in this thesis has been submitted as described in the following:
Kuek F.W.I., Mujahid A., Lim P.T., Leaw C.P. & Müller M. ‘Diversity and DMS(P)-
related genes in culturable bacterial communities in Malaysian coastal
waters’. Systematic and Applied Microbiology (Manuscript ID:).
Kuek F.W.I., Lim L.F., Ngu L.H., Mujahid A., Lim P.T., Leaw C.P. & Müller M. ‘The
potential roles of bacterial communities in coral defence: a case study at
Talang-talang reef’. Ocean Science Journal (Manuscript ID: OSJO-D-14-
00062).
Early work has been presented in the following conferences and contributed to the
content presented in Chapters 3 and 4 of this thesis:
Müller M., Kuek F.W.I., Song J.X.P. & Mujahid A. ‘Potential role of microbes in the
local sulphur and nitrogen cycles in Kuching waters’ IOC/WESTPAC 9th
International Scientific Symposium, 22-25 April 2014, Nha Trang, Khanh
Hoa, Vietnam. (Oral presentation)
Kuek F.W.I., Lim L.F., Ngu L.H., Ng C.T., Mujahid A., Lim P.T., Leaw C.P. & Müller M.
‘The potential role of bacterial communities: a case study at Talang-talang
reef’ IOC/WESTPAC 9th International Scientific Symposium, 22-25 April
2014, Nha Trang, Khanh Hoa, Vietnam. (Poster presentation)
Kuek F.W.I., Lim L.F., Ngu L.H., Ng C.T., Mujahid A., Lim P.T., Leaw C.P. & Müller M.
‘Coral mucus bacterial communities of Talang-talang reef and their
potential role in coral defence’ International conference on sustainable
development of tropical coastal zones, 5-6 September 2013, Port Dickson,
Malaysia. (Poster presentation)
Klaus Pfeilsticker and the SHIVA consortium ‘Findings of the SHIVA field campaign
in the South China Sea in Nov.-Dec. 2011’ Geophysical Research Abstracts
Vol. 15, EGU2013-1702, 2013. European Geosciences Union General
Assembly, 7-12 April 2013. Vienna, Austria. (Oral presentation)

P a g e | viii
Mujahid A., Müller M., Ngu E.S.L., Lee S.T.H., Lew Y.L., Kuek F.W.I., Lim H.C., Teng
S.T., Leaw C.P., & Lim P.T. ‘SHIVA local boat deployment in Kuching, and
major findings from Sarawak’ SONNE status seminar, 13-15 February 2013,
Kiel, Germany. (Poster presentation)
Kuek F.W.I., Ngu E.S.L., Lee S.T.H., Mujahid A., Lim P.T., Leaw C.P. & Müller M.
‘Microbial communities of the eastern South China Sea and their possible
role in the DMS(P) cycle’ SONNE status seminar, 13-15 February 2013, Kiel,
Germany. (Poster presentation)
Klaus Pfeilsticker and the SHIVA consortium ‘SHIVA consortium: Overview on the
SHIVA activities and results’ South China Sea Conference, 21-24 October
2012, Kuala Lumpur, Malaysia. (Oral presentation)
Kuek F.W.I., Mujahid A., Lim P.T., Leaw C.P. & Müller M. ‘Diversity of culturable
bacteria from Talang-talang reef and its surrounding waters’ South China
Sea Conference, 21-24 October 2012, Kuala Lumpur, Malaysia. (Poster
presentation)

P a g e | ix
Table of Contents
Page
List of Figures
List of Tables
1 Introduction
1.1 Microbes in the ocean
1.2 South China Sea and the Celebes Sea
1.3 Surface microbes and their roles in the DMS(P) cycle
1.4 Coral reefs
1.4.1 Coral reefs of Malaysia
1.5 Coral bleaching
1.6 Coral Surface Mucus Layer (SML) and associated microbes
1.7 Coral diseases and the role of microbes in the SML
1.8 Polyketide synthase (PKS) and non-ribosomal peptide
synthetase (NRPS)
1.9 Significance and aims of the present study and dissertation
outline
2 Methodology
2.1 Field sampling
2.1.1 Reef samples
2.2 Laboratory procedures
2.2.1 Isolation of bacteria
2.2.2 Molecular characterisation
2.2.3 Clone libraries from water samples
2.2.3.1 Extraction of genomic DNA from sea water
2.2.3.2 DNA cloning and plasmid extraction
2.2.3.3 Bacterial communities based in the
construction of clone libraries
2.2.4 PCR amplification of bacterial DMSP cleavage (dddP)
and demethylation (dmdA) genes
2.2.5 PCR amplification of bacterial antimicrobial PKS and
NRPS genes
xii
xviii
1
1
2
4
8
10
10
11
13
14
18
19
19
20
22
22
23
24
24
25
26
28
30

P a g e | x
2.2.6 Antimicrobial tests
2.2.6.1 Extraction of bioactive compounds
2.2.6.2 Inhibitory interaction tests (well diffusion
assay)
3 Bacterial Communities from the eastern South China Sea and
the Celebes Sea, and Their Potential Role in the DMS(P) Cycle
3.1 Introduction
3.2 Materials and Methods
3.2.1 Study site and sample collection
3.2.2 DNA extraction and purification of cultured bacteria
3.2.3 PCR amplification of bacterial 16S rRNA genes
3.2.4 Sequencing and phylogenetic analysis
3.2.5 Nucleotide sequence accession numbers
3.2.6 PCR amplification of bacterial DMSP cleavage (dddP)
and demethylation (dmdA) genes
3.3 Results and Discussion
3.3.1 Physico-chemical parameters
3.3.2 Diversity of culturable bacterial communities
3.3.3 Variations in the bacterial communities in Kuching,
Kota Kinabalu and Semporna waters
3.3.4 Bacterial strains with potential to metabolise DMS
and/or demethylate DMSP
3.4 Conclusion
3.5 Acknowledgement
4 Bacterial Communities from Talang-Talang Reef and Their
Potential Role in Coral Defence and the DMS(P) Cycle
4.1 Introduction
4.2 Materials and Methods
4.2.1 Study site and sample collection
4.2.2 DNA extraction and purification of cultured bacteria
4.2.3 PCR amplification of bacterial 16S rRNA genes
4.2.4 Sequencing and phylogenetic analysis
4.2.5 Nucleotide sequence accession numbers
31
31
32
34
35
38
38
40
41
41
41
42
42
42
44
59
60
66
67
68
69
71
71
72
72
72
73

P a g e | xi
4.2.6 PCR amplification of bacterial DMSP cleavage (dddP)
and demethylation (dmdA) genes
4.2.7 PCR amplification of bacterial polyketide synthase
(PKS) and non-ribosomal peptide synthetase (NRPS)
genes
4.2.8 Extraction of bioactive compounds
4.2.9 Well diffusion assay
4.3 Results and Discussion
4.3.1 Diversity of culturable bacterial communities
4.3.2 Bacterial strains with PKS and NRPS genes
4.3.3 Role of mucus-associated bacteria in coral defence
4.3.4 Bacterial strains with potential to metabolise DMS
and/or demethylate DMSP
4.4 Conclusion
4.5 Acknowledgement
5 Summary and Future Work
5.1 Future Research
References
Appendix
73
73
74
74
74
74
81
83
88
90
91
92
93
95
121

P a g e | xii
List of Figures
Figure Page
1.1 Map of the South China Sea (source: Morton & Blackmore 2001).
1.2 Overview of DMSP catabolic pathways in marine bacteria and the
fates of carbon and sulphur (taken from Reisch, Moran & Whitman
2011).
1.3 Biochemical pathways of DMSP demethylation. [1] DMSP
demethylase (DmdA); [2] 5,10-methylene-THF reductase (MetF,
E.C.1.5.1.20); [3] methylene-THF dehydrogenase (FolD, E.C.1.5.1.5);
[4] methenyl-THF-cyclohydrolase (E.C.3.5.4.9); [5] methionine
synthase (MetH, E.C. 2.1.1.13); [6] methionine salvage pathway
(multiple enzymes); [7] MMPA-CoA ligase (DmdB); [8] MMPA-CoA
dehydrogenase(DmdC); [9] methylthioacryloyl-CoA hydratase
(DmdD); [10] acetaldehyde dehydrogenase (E.C.1.2.1.10) (source:
Reisch, Moran & Whitman 2011).
1.4 Dimethylsulfoniopropionate cleavage pathways leading to central
carbon metabolism. Reactions 5 and 6 may be coenzyme-A mediated
and would therefore bypass reaction 8. [1] DMSP-cleavage enzyme
(DddD); [2] DMSP lyase (DddL, DddP, DddQ, DddY, DddW, E.C.
4.4.1.3); [3] acrylate hydratase; [4] 3-hydroxypropionate
dehydrogenase; [5] 3-hydroxypropionate reductase; [6] acrylate
reductase (1.3.99.3); [7] malonate semialdehyde
dehydrogenase/decarboxylase (E.C.1.2.1.18); [8] propionate-CoA
ligase (PrpE, E.C.6.2.1.17) (source: Reisch, Moran & Whitman 2011).
1.5 Distribution of coral reefs in the East Asian Seas (source:
http://www.ncdc.noaa.gov/paleo/outreach/coral/sor/sor_asia.html).
1.6 Basic steps during PKS. Each PKS module consists of three core
domains: an acyltransferase (AT) domain, which selects the
appropriate extender unit (usually malonyl-CoA or methylmalonyl-
CoA) and transfers it to the ACP domain where a thioester bond is
formed, and a ketosynthase (KS) domain, responsible for
decarboxylative condensation between the extender unit present on
3
5
6
7
9

P a g e | xiii
the ACP domain of the same module and the polyketide intermediate
bound to the ACP domain of the preceding module. All elongating
modules present these core domains, while the loading module lacks
a functional KS domain and the last module contains an additional
TE domain, for release of the finished polyketide from the PKS. Most
PKS modules contain additional domains for processing the newly
formed b-keto: the b-ketoreductase (KR), the dehydratase (DH) and
the enoylreductase (ER) domains carry out the reactions (source:
Donadio, Monciardini & Sosio 2007).
1.7 Basic steps during NRPS. Each NRPS module consists of three core
domains: an adenylation (A) domain, which selects the cognate
amino acid, activates it as an amino acyl adenylate and transfers it to
the T domain (also known as peptidyl carrier protein, or PCP) where
a thioester bond is formed, a condensation (C) domain, responsible
for peptide bond formation between the amino acid present on the T
domain of the same module and the peptidyl intermediate bound to
the T domain of the preceding module, and the T domain itself.
Usually, all elongation modules present these core domains. A
dedicated loading module (carrying just A and T domains) and a
termination module, containing a thioesterase (TE) domain, usually
complete the NRPS assembly line (source: Donadio, Monciardini &
Sosio 2007).
2.1 Overview of the SHIVA cruise (source: http://shiva.iup.uni-
heidelberg.de/a_activities.html).
2.2 Overview of the Talang-talang Islands just off the shores of Kuching,
Sarawak (source: Yahya, Hassan & Husaini 2012)
2.3 Collection of fresh coral mucus.
2.4 16S rRNA bands of bacterial isolates. Impure bands can be seen at
BSD 16-5, 16-7, 16-11. These isolates were later reisolated to ensure
pure cultures.
2.5 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in clone libraries from Kuching and Kota Kinabalu.
The phylogenetic tree was generated with distance methods, and
15
16
19
21
22
24

P a g e | xiv
sequence distances were estimated with the neighbour-joining
method. Bootstrap values ≥50 are shown and the scale bar
represents a difference of 0.05 substitution per site. Accession
numbers for the reference sequences are indicated.
2.6 PCR-based screening of dmdA genes. Bands highlighted in this figure
indicate presence of the genes.
2.7 PCR-based screening of dddP genes. Bands highlighted in this figure
indicate presence of the genes.
2.8 PCR-based screening of PKS genes. Bands highlighted in this figure
indicate presence of the genes.
2.9 PCR-based screening of NRPS genes. Bands highlighted in this figure
indicate presence of the genes.
2.10 Inhibition zones from bioactive compoinds with antimicrobial
properties.
3.1 The RV Sonne ship track leading from Singapore to Manila between
November 15-29, 2011 during the SHIVA SO 218 cruise.
3.2 Locations of sampling stations in Kuching, Sarawak.
3.3 Locations of sampling stations in Kota Kinabalu, Sabah.
3.4 Locations of sampling stations in Semporna, Sabah.
3.5 Pie charts illustrating the diversity of bacterial groups based on
partial 16S rRNA gene sequences from bacteria isolated from (a)
Kuching, (b) Kota Kinabalu and (c) Semporna.
3.6 Phylogenetic groups of isolates from the waters of Kuching, Kota
Kinabalu and Semporna at depths of 1, 5 and 10 m.
3.7 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kuching 1611. The phylogenetic tree was
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.05
substitution per site. Accession numbers for the reference sequences
are indicated.
3.8 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kuching 1911. The phylogenetic tree was
27
29
30
31
31
33
38
39
40
40
45
45
47

P a g e | xv
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.1
substitution per site. Accession numbers for the reference sequences
are indicated.
3.9 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kota Kinabalu. The phylogenetic tree was
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.05
substitution per site. Accession numbers for the reference sequences
are indicated.
3.10 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Semporna. The phylogenetic tree was generated
with distance methods, and sequence distances were estimated with
the neighbour-joining method. Bootstrap values ≥50 are shown and
the scale bar represents a difference of 0.1 substitution per site.
Accession numbers for the reference sequences are indicated.
3.11 Relative abundance of dmdA and dddP genes in cultured bacterial
communities from the waters of (a) Kuching, (b) Kota Kinabalu and
(c) Semporna.
3.12 Presence of dmdA and/or dddP genes in bacterial isolates from the
waters of Kuching, Kota Kinabalu and Semporna.
3.13 Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Kuching.
3.14 Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Kota Kinabalu.
3.15 Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Semporna.
4.1 Overview of the Talang-talang Islands just off the shores of Kuching,
Sarawak. Enlarged map indicates sampling area.
4.2 Pie charts illustrating the diversity of bacterial groups based on
partial 16S rRNA gene sequences from bacteria isolated from (a)
48
49
51
61
62
63
63
64
71

P a g e | xvi
coral mucus, (b) water column and (c) sediment.
4.3 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in coral mucus The phylogenetic tree was
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.1
substitution per site. Accession numbers for the reference sequences
are indicated.
4.4 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in water column. The phylogenetic tree was
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.05
substitution per site. Accession numbers for the reference sequences
are indicated.
4.5 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in reef sediment. The phylogenetic tree was
generated with distance methods, and sequence distances were
estimated with the neighbour-joining method. Bootstrap values ≥50
are shown and the scale bar represents a difference of 0.05
substitution per site. Accession numbers for the reference sequences
are indicated.
4.6 Percentage of Vibrio isolates in mucus attached and mucus
associated communities.
4.7 Relative abundance of PKS and NRPS genes in cultured bacterial
communities from coral mucus.
4.8 Presence of PKS and/or NRPS genes in bacterial isolates from coral
mucus.
4.9 Total inhibitions of mucus attached bacteria at temperatures of 28,
30 and 32 °C.
4.10 Total inhibitions of mucus associated bacteria at temperatures of 28,
30 and 32 °C.
4.11 Average zone of inhibitions (cm) of mucus attached bacteria at
75
76
77
78
79
82
82
83
83

P a g e | xvii
temperatures of 28, 30 and 32 °C.
4.12 Average zone of inhibitions (cm) of mucus associated bacteria at
temperatures of 28, 30 and 32 °C.
4.13 Relative abundance of dmdA and dddP genes in cultured bacterial
communities from coral mucus.
4.14 Presence of dmdA and/or dddP genes in bacterial isolates from coral
mucus.
84
84
88
89

P a g e | xviii
List of Tables
Table Page
1.1 Regional distribution of coral reefs (source: Veron & Stafford-Smith
2000).
3.1 Locations of sampling stations at Kuching, Kota Kinabalu and
Semporna.
3.2 Physico-chemical parameters measured from Kuching and Kota
Kinabalu at depths of 1 and 5 m.
3.3 Indices used to quantify the diversity of bacterial communities at
Kuching, Kota Kinabalu and Semporna.
A.1 16S rRNA gene sequence analysis of bacterial cultures from Kuching
1611, based on BLAST analysis.
A.2 16S rRNA gene sequence analysis of bacterial cultures from Kuching
1911, based on BLAST analysis.
A.3 16S rRNA gene sequence analysis of bacterial cultures from Kota
Kinabalu, based on BLAST analysis.
A.4 16S rRNA gene sequence analysis of bacterial cultures from
Semporna, based on BLAST analysis.
A.5 16S rRNA gene sequence analysis of bacterial cultures from Talang-
talang reef and its surrounding waters, based on BLAST analysis.
A.6 Screening of antimicrobial (PKS/NRPS) and DMSP-degrading
(dmdA/dddP) genes in coral mucus isolates.
A.7 Total inhibition and inhibition zones of mucus attached isolates at
28, 30 and 32 °C.
A.8 Total inhibition and inhibition zones of mucus associated isolates at
28, 30 and 32 °C.
9
39
42
46
126
130
133
136
139
145
146
147

P a g e | xix

P a g e | 1
CHAPTER 1
Introduction
1.1 Microbes in the oceans
The oceans are made up of a web of different marine organisms that form an
interdependent community. Microbes, a major component of this community, have
been studied by scientists for years in attempts to establish a better understanding
of their diversity, distribution and nature. An estimated total of 3.6×1029 microbial
cells reside in the oceans (Singh 2010). These marine microorganisms have
experienced billions of years’ worth of evolution, forming vast and complex
communities of bacteria, archaea, protists and fungi, within what is said to be the
dominant biome of the Earth (DeLong 2009). The actual number of microbes that
exist in the ocean, however, is thought to surpass published estimates; indicating
that while many have been and are in the process of being identified, an equally
great percentage still remains undiscovered (Karl 2002; Sogin et al. 2006).
These microbes play vital roles in the marine ecosystem by mediating the
geochemical cycles in the ocean (Arrigo 2005) and allowing for rapid nutrient
recycling in an environment that is poor in essential nutrients (Mayer & Wild
2010). Consequently, they are responsible for around 98% of overall primary
production in the ocean, providing short-term sustainability to the marine
ecosystem while a longer term supply of nutrients comes from external sources
(Karl 2002; Sogin et al. 2006). As a result of dominating an ecosystem that
constitutes approximately 40% of the Earth’s surface, these microbes and their
involvement in biogeochemical processes are significant on a global scale (Karl
2002).
For decades, microbiologists have aimed to unravel the mysteries of the microbial
world through culture-based studies. This approach allowed them to discover new
species, as well as to study their biochemical properties. Today, the advances in
molecular biology have brought ecological studies in microbiology to even greater

P a g e | 2
heights. Physiological and biochemical studies, previously hindered by obstacles in
culturing the ‘unculturable’, can now be carried out to establish the identities,
phylogenetic relationships and metabolic processes of both cultured and
uncultured microbial populations via DNA or RNA based methods (Jørgensen
2006).
Characterization of microbes by genera and species, which previously could not be
achieved through biochemical methods alone, can now be executed with relative
ease with the help of sequence-classifier algorithms (Petrosino et al. 2009).
Sequencing studies are conventionally carried out using the Sanger method
(Sanger, Nicklen & Coulson 1977) which is widely used in microbial population
studies. Each metabolic function is encoded by a specific set of genes, thus
scientists today rely on molecular-based protocols for a more rapid and effective
study of the nature of these bacteria (Rappé & Giovannoni 2003). Sequencing will
provide us with an indication of whether these specific genes are present in a
sample, signalling that these special groups of bacteria inhabit the population of
study (Rajendhran & Gunasekaran 2011); a far easier alternative to culture-based
protocols which can eventually lead to more in-depth studies on microbial
metabolism.
1.2 South China Sea and the Celebes Sea
The South China Sea is a marginal sea that is part of the Pacific Ocean,
encompassing an area from the Singapore and Malacca Straits to the Strait of
Taiwan (Morton & Blackmore 2001; see Figure 1.1 for a map of the South China
Sea). The Celebes Sea is connected to the South China Sea through the Sulu Sea
(Yoshida, Nishimura & Kogure 2007).

P a g e | 3
Figure 1.1: Map of the South China Sea (source: Morton & Blackmore 2001).
Not much is known about the diversity and function of the microbial communities
in the South China Sea, especially regarding the eastern region (Kuching and Kota
Kinabalu) and the Celebes Sea. To our knowledge, there is no study on regional
scale or large-scale distribution patterns of microbes in the Malaysian area of the
South China Sea. Most studies about bacterial communities focused on regions
near China, such as those carried out by Li et al. (2006), Jiang et al. (2007) and Tao,
Peng & Pinxian (2008) and a brief mention of communities in the Celebes Sea by
Yoshida, Nishimura & Kogure (2007). All the studies mentioned used culture-
independent techniques to reveal the community structure and diversity of the
predominant bacteria at the sampling environment. No studies on culturable
communities in the region have been made at this time.

P a g e | 4
1.3 Surface microbes and their roles in the DMS(P) cycle
Through recent studies, the biogeochemical processes of marine microbes have
been discovered not only to regulate marine ecosystems but to potentially have an
indirect influence on the Earth’s climate as well. In 1983 it was first hypothesized
that the sulphur gases released from biotic ecosystems, upon their oxidation in the
atmosphere, would also affect the Earth’s climate (Shaw 1983) and marine
planktonic algae have since been found to contribute largely to the exchange of
sulphur gases across the ocean-atmosphere boundary. They are known to produce
dimethylsufoniopropionate (DMSP), a metabolite precursor to dimethylsulfide
(DMS) which is the primary source of sulphur released into the atmosphere (Strom
2008; Reisch, Moran & Whitman 2011). Once in the atmosphere, DMS is oxidized
into sulphate aerosols that will act as cloud condensation nuclei (CCN) which will
promote cloud formation (Charlson et al. 1987). The protective layer of clouds
formed will serve to reduce the amount of solar radiation that reaches the Earth’s
surface (Welsh 2000; Vallina & Simó 2007) thus, possibly reducing the Earth’s
overall temperature by several degrees. Certain species of marine microbes have
been found to be mediators to the reaction by producing the enzymes necessary to
cleave DMSP into the desired gas product (Vila-Costa et al. 2010) and are therefore
being put under extensive study as a model for climate change studies.
In research today, scientists use modern molecular technology to locate the genes
responsible for the production of these enzymes that will determine the fate of
DMSP. Two possible pathways exist for DMSP (see Figure 1.2 for an overview),
catabolism into DMS or demethylation into methanethiol (MeSH), the former of
which – while quantitatively less important than demethylation (Kiene 1996a) – is
the largest biogenic source of sulphur to the atmosphere (Andreae & Raemdonck
1983). The cleavage pathway is important in mediating organic sulphur emission
into the atmosphere by splitting of the DMSP molecule into acrylate and DMS
(Reisch, Moran & Whitman 2011). The demethylation pathway involves the
removal of a methyl group from DMSP to produce 3-methiolpropionate, which is
then cleaved to methanethiol and probably acrylate or propionate (González, Kiene
& Moran 1999). The potential of bacterial strains to use more than one DMSP

P a g e | 5
catabolic pathway has been previously noted, especially among Roseobacter spp. by
González et al. (1999).
Figure 1.2: Overview of DMSP catabolic pathways in marine bacteria and the fates
of carbon and sulphur (taken from Reisch, Moran & Whitman 2011).
To date, six different enzymes responsible for DMSP cleavage have been identified,
each encoded by different genes and known to catalyse different reactions that
ultimately lead down different pathways. Using genomic-based studies, specific
metabolic processes within a microbial population can be identified more easily
which will contribute to studies on the different biochemical pathways and
regulatory factors involved in DMSP metabolism – something that still remains
very poorly understood (Reisch, Moran & Whitman 2011). Vila-Costa et al. (2010)
carried out a transcriptomic analysis on the marine microbial population in the
Sargasso Sea to study gene expression of the microbes in the presence of low
amounts of DMSP. They were able to identify several genes known to be directly
involved in DMSP degradation and could classify them according to the taxonomic
groups.

P a g e | 6
Figure 1.3: Biochemical pathways of DMSP demethylation. [1] DMSP demethylase
(DmdA); [2] 5,10-methylene-THF reductase (MetF, E.C.1.5.1.20); [3] methylene-
THF dehydrogenase (FolD, E.C.1.5.1.5); [4] methenyl-THF-cyclohydrolase
(E.C.3.5.4.9); [5] methionine synthase (MetH, E.C. 2.1.1.13); [6] methionine salvage
pathway (multiple enzymes); [7] MMPA-CoA ligase (DmdB); [8] MMPA-CoA
dehydrogenase(DmdC); [9] methylthioacryloyl-CoA hydratase (DmdD); [10]
acetaldehyde dehydrogenase (E.C.1.2.1.10) (source: Reisch, Moran & Whitman
2011).

P a g e | 7
Figure 1.4: Dimethylsulfoniopropionate cleavage pathways leading to central
carbon metabolism. Reactions 5 and 6 may be coenzyme-A mediated and would
therefore bypass reaction 8. [1] DMSP-cleavage enzyme (DddD); [2] DMSP lyase
(DddL, DddP, DddQ, DddY, DddW, E.C. 4.4.1.3); [3] acrylate hydratase; [4] 3-
hydroxypropionate dehydrogenase; [5] 3-hydroxypropionate reductase; [6]
acrylate reductase (1.3.99.3); [7] malonate semialdehyde
dehydrogenase/decarboxylase (E.C.1.2.1.18); [8] propionate-CoA ligase (PrpE,
E.C.6.2.1.17) (source: Reisch, Moran & Whitman 2011).
The identification of the DMSP-demethylase gene (dmdA), which encodes the first
step in the demethylation pathway (see Figure 1.3 for a complete overview of the
demethylation pathway), has enabled quantification of the gene in marine
metagenomic surveys and revealed it to be taxonomically diverse and highly
abundant (del Valle, Kiene & Karl 2012). In comparison to dmdA, the genes
involved in DMS production (dddD, dddL, dddP dddQ, dddY, and dddW, all of
which mediate the same step of DMSP cleavage; see Figure 1.4) are present in less
than 10% of bacteria based on marine metagenomic surveys (Todd et al. 2007;

P a g e | 8
Curson et al. 2008; Howard et al. 2008; Todd et al. 2009; Curson, Sullivan, et al.
2011; Todd et al. 2011; Todd, Curson, et al. 2012). The most abundant ddd genes in
bacterial taxa are dddP and dddQ, occurring in genomes of some Roseobacter spp.
(Howard et al. 2008; Todd et al. 2009, 2011) and SAR116.
As can be seen from the lack of studies mentioned above, there is very limited
information available on bacteria in the South China Sea (SCS) and even less on
their potential role in the DMS(P) cycle. One aim of this thesis is to provide data on
both. Besides surface waters, bacteria also play major roles in other oceanic
ecosystems for example coral reefs. In the following, coral reefs will be introduced
as well as the role that bacteria play in them
1.4 Coral reefs
Coral reefs are among the most diverse and productive ecosystems on this planet.
Millions of people rely on harvests derived from coral reefs as their major source
of protein and income (Wilkinson & Buddemeier 1994). In addition, revenue
earned from tourism, recreation, education and research are of major importance
to our national economy (Wilson et al. 2012). Coral reefs also act as a natural
protection between the open seas and coastlines by acting as wave breaks, thus
effectively preventing coastal erosion (Buddemeier, Kleypas & Aronson 2004;
McLeod et al. 2010; Eghtesadi-Araghi 2011). They perform a vital role in
protecting coastal areas from the consequences of rising sea levels such as storm
flooding (Wilkinson 1999). There is also increasing evidence of the potential of
reefs to act as bio-indicators for climate change, as they are sensitive to rising sea
levels and increasing sea temperature (Awang, Moshidi & Muda 2003). In addition,
reefs are good indicators of coastal pollution, as they are sensitive to changes in
their ambient environment (Moberg & Folke 1999). Coral reefs in the South Pacific
cover the highest amount of space (116,200 km2; see Table 1.1), closely followed
by Southeast Asia (87,760 km2, see Table 1.1.), indicating their important role for
the local communities.

P a g e | 9
Table 1.1: Regional distribution of coral reefs (source: Veron & Stafford-Smith
2000).
Region Reef area (km2)
South Pacific 116,200 Southeast Asia 87,760 Indian Ocean 31,930 Middle East 21,450 Caribbean 20,360 Western Atlantic 2.820
Reefs are widely distributed in the area (see Figure 1.5 for an overview of reef
distribution in the East Asian Seas) and Southeast Asia’s coral reefs have the
highest biodiversity of all the world’s reefs (Veron & Stafford-Smith 2000). This
region contains more than 600 of the nearly 800 reef building coral species found
worldwide (Veron & Stafford-Smith 2000).
Figure 1.5: Distribution of coral reefs in the East Asian Seas (source:
http://www.ncdc.noaa.gov/paleo/outreach/coral/sor/sor_asia.html).

P a g e | 10
1.4.1 Coral reefs of Malaysia
The wide geographic range that Malaysia covers means that coral reefs can be
found in varied conditions across the country. Malaysia is included in the Coral
Triangle, a global centre of marine biodiversity (Lee & Mohamed 2009). The
waters of the Coral Triangle hold the highest diversity of iridescent corals, fish,
crustaceans, mollusks and marine plant species in the world (Veron et al. 2009).
The area sustains over 120 million people and garners more than $12 billion a year
from nature-based tourism (Cabral et al. 2013).
Malaysia has an estimated 4,000 km2 of coral reefs (Yaman n.d.). Little reef
development occurs along the west coast of Peninsular Malaysia, but the east coast
has some fringing reefs along the coast and many reefs around the offshore islands
(Harborne et al., 2000; see Figure 1.6). In East Malaysia, reef development around
Sarawak is limited due to high sedimentation (Pilcher & Cabanban 2000a) (Yaman
n.d.; see Figure 1.5). However, Sabah contains more than 75% of all Malaysian
reefs and has high levels of coral diversity (Pilcher & Cabanban 2000b). Overall,
more than 350 coral species have been recorded in Peninsular Malaysia and over
500 in East Malaysia (Praveena, Siraj & Aris 2012). Unfortunately, there are
several threats to coral reefs such as sedimentation, eutrophication, temperature
rise and ocean acidification (Praveena, Siraj & Aris 2012) and Malaysia is no
exception to the rule. Corals that are under stress for long time will eventually die,
mainly due to a process called "coral bleaching".
1.5 Coral bleaching
Bleaching is defined as the disruption of the symbiosis between the coral host and
its endosymbiotic zooxanthellae, resulting in the loss of the algal symbiont and/or
of the algal pigments, thus making the coral tissue transparent and exposing the
underlying white calcium carbonate skeleton (Rosenberg et al. 2009). If symbiont
populations are not restored within weeks or months of a bleaching event, then
whole or partial coral mortality is likely (Hoegh-Guldberg 2004). Coral bleaching
has increased in frequency, intensity and geographical extent over the last few
decades (Huppert & Stone 1998) and has been correlated with increased seawater
temperatures as well as high levels of solar irradiance (Jokiel & Brown 2004).

P a g e | 11
Predicted ocean warming in the current century is expected to result in higher
frequency of mass coral bleaching events and associated mortality (Hoegh-
Guldberg 1999).
Temperature-related bleaching is the most widely reported cause of coral reef
stress (Buddemeier, Kleypas & Aronson 2004). The range of temperatures
tolerated by reef-building corals worldwide is relatively narrow, and studies have
shown that a temperature of only 1-2 °C above the normal threshold temperature
for a few weeks is sufficient to cause a bleaching event (Fitt et al. 2001). Sea-
surface temperatures have now moved so close to coral thermal limits that the
fluctuations of temperatures within natural climatic events such as the El Niño
Southern Oscillation (ENSO) can cause massive coral bleaching (Praveena, Siraj &
Aris 2012). The most severe ENSO event since statistics have been recorded
occurred in 1997-1998. Although the effects from the 1997-1998 event were most
severe in the central Indian Ocean, major bleaching was also reported across
Southeast Asia, where an estimated 18% of reefs were damaged (Praveena, Siraj &
Aris 2012).
Two mechanisms for causing bleaching have been discovered: photoinhibition
leading to the damage of photosystem II (Jones et al. 1998); and infection by a
pathogen that targets the zooxanthellae (Rosenberg 2004). As discussed in the
following, bacteria play a significant role in the latter and other coral diseases.
1.6 Coral Surface Mucus Layer (SML) and associated microbes
All corals have a surface mucus layer (SML) that is generated by secretion of a
polysaccharide-protein complex by mucocytes (Sharon & Rosenberg 2008). The
SML serves as an ecological niche rich in nutrients and diverse in bacterial
populations (Shnit-Orland & Kushmaro 2008). It plays an important role in
structuring microbial communities on the coral surface by providing a hostile
environment for some bacteria and a nurturing environment for others (Ritchie
2006). Various functions have been ascribed to the SML including defence against
disease-causing pathogens, desiccation resistance, shedding of sediments and
protection against radiation (Sharon & Rosenberg 2008). On average, 20-30 % of

P a g e | 12
bacterial isolates originating from coral SML possess antibacterial properties
(Ritchie 2006) that may assist the coral holobiont as a first line of defence against
pathogens and fouling organisms (Shnit-Orland & Kushmaro 2009). It has been
suggested that these antimicrobial compounds are temperature sensitive (Ritchie
2006). Antibacterial activity was found to be optimal at 26 °C and slightly
decreased at 30 °C, with partial inactivation occurring at 60 °C and complete loss of
activity occurring at 80 °C (Shnit-Orland & Kushmaro 2009).
High temperatures act on the coral microorganisms as well as on the coral host,
causing a change in microbial community (Rosenberg et al. 2009). There is
evidence that a community shift in the coral SML from beneficial bacteria to Vibrio-
dominance occurs prior to zooxanthellae loss (Ritchie 2006). Studies have shown
that Vibrio may be normal constituents of the coral microbial assemblages and can
opportunistically proliferate if holobiont health is compromised (Bourne & Munn
2005). Previous studies have implicated Vibrio spp. as the principal causative agent
in seasonal and species-specific episodes of coral bleaching (Kushmaro et al. 1996,
1997; Ben-Haim & Rosenberg 2002; Ben-Haim et al. 2003). It was speculated that
the endosymbiotic zooxanthellae (Symbiodinium spp.) play a significant role in
restricting Vibrio growth in the coral SML by producing free radicals (Sharon &
Rosenberg 2008). However, three separate studies (Ritchie et al. 1994; Kushmaro
et al. 1996; Ben-Haim, Zicherman-Keren & Rosenberg 2003) showed that the
number of Vibrio in coral SML did increase with increasing temperatures. In
elevated temperatures, Vibrio spp. will produce a photosynthesis inhibitor (Ben-
Haim et al. 1999), thereby allowing them to multiply, leading to overgrowth and in
turn, causing the loss of antibiotic properties of the SML inhabiting
microorganisms (Ritchie 2006). During bleaching, coral mucus production changes
in quality and can decrease in quantity (Ritchie 2006). Elevated sea water
temperatures can also induce pathogens to produce adhesions that allow it to
adhere to the coral surface and subsequently establish infections in the pathogenic
systems of the coral (Banin, Ben-Haim, et al. 2000). The production of toxins and
lytic enzymes which cause bleaching and lysis of zooxanthellae were also found to
be temperature-regulated (Banin, Ben-Haim, et al. 2000). Although temperature
may affect the metabolism and diversity of the microbial community, the loss of

P a g e | 13
zooxanthellae is potentially the fundamental driving parameter changing the
microbiota of the corals (Bourne et al. 2008).
1.7 Coral diseases and the role of microbes in the SML
Emerging diseases have been responsible for the death of about 30% of corals
worldwide in the last few decades and it is predicted that by 2050, most of the
world’s coral reefs will be destroyed (Reshef et al. 2006). Many disease outbreaks
involve opportunistic infections by endemic microbes following periods of stress
(Lesser et al. 2007; Rosenberg et al. 2009; Bourne et al. 2009). Bleached corals are
additionally vulnerable because the loss of algae reduces the concentration of
oxygen and the resulting radicals that protect the coral animal (Banin et al. 2003).
Disease susceptibility is positively correlated with a change in coral SML
composition, loss of antibiotic activity and an increase in pathogenic microbes
(Reshef et al. 2006). The bacterial communities of diseased corals are different
from healthy ones, both qualitatively and quantitatively (Reshef et al. 2006). The
bacterial population of apparently healthy corals undergo changes within a period
of a few months, probably as a result of temperature changes (Koren & Rosenberg
2006). Previous studies have shown a sudden shift to pathogen dominance
occurring in the coral SML prior to a bleaching event (Lipp, Huq & Colwell 2002;
Rosenberg & Ben-Haim 2002; Ritchie 2006) and it has been demonstrated that
antibiotic activity and antibiotic-producing bacteria in the SML decline in times of
increased water temperature when bleaching is most likely to occur (Ritchie
2006). One possible explanation for an increased incidence of coral diseases is
stress-induced susceptibility to opportunistic microbes trapped in the coral SML
(Ritchie 2006). Indigenous bacteria may help prevent infection by pathogens by
producing antibacterial materials (Koh 1997).
Vibrio shiloi is a known bacterial pathogen to the coral Oculina patagonica found in
the Mediterranean sea (Kushmaro et al. 1996, 1997, 2001). It induces bleaching by
reducing the amount of viable zooxanthellae available for symbiosis with the coral.
This is achieved by the secretion of a toxin (a proline-rich, 12 amino acid peptide)
(Banin, Israely, et al. 2000) that inhibits photosynthesis, and bleaches and lyses

P a g e | 14
zooxanthellae (Ben-Haim et al. 1999). Vibrio shiloi is only actively pathogenic at
temperatures of 20-32°C and displays maximum efficacy around 29-30°C
(Kushmaro et al. 2001).
A more recently discovered temperature-dependent agent of bleaching is Vibrio
coralliilyticus which infects the coral Pocillopora damicornis (Ben-Haim et al.
2003). A patchy pattern of bleaching of Pocillopora damicornis has been observed
at 24 °C, suggesting that bacterial bleaching results from an attack on the
zooxanthellae, followed by bacterium-induced coral lysis and death caused by
bacterial extracellular proteases which were produced at temperatures of 24 to 28
°C (Ben-Haim, Zicherman-Keren & Rosenberg 2003; Rosenberg 2004).
There has been only one published report about coral reefs in Sarawak (Awang,
Moshidi & Muda 2003) and this report does not contain any information about
coral-associated microbial communities. Since associated bacteria play a major
role as a first line of defence against pathogens (Shnit-Orland & Kushmaro 2009),
and are of significance to the survival of coral reefs in the area, the second aim of
the thesis was therefore to take a closer look at the bacteria in a local reef and their
potential role in coral defence as well as their response to changing temperatures.
1.8 Polyketide synthase (PKS) and non-ribosomal peptide synthetase
(NRPS)
Polyketides and non-ribosomal peptides comprise two large families of secondary
metabolites and numerous natural products belonging to these groups are widely
used as pharmaceuticals, industrial agents or agrochemicals (Silakowski, Kunze &
Müller 2001). Both types are biosynthesized by extremely large polyfunctional
enzyme systems within the protein. The responsible biosynthetic proteins are
known as polyketide synthases (PKS) and nonribosomal polypeptide sythetases
(NRPS) (Cane 1997).

P a g e | 15
Figure 1.6: Basic steps during PKS. Each PKS module consists of three core
domains: an acyltransferase (AT) domain, which selects the appropriate extender
unit (usually malonyl-CoA or methylmalonyl-CoA) and transfers it to the ACP
domain where a thioester bond is formed, and a ketosynthase (KS) domain,
responsible for decarboxylative condensation between the extender unit present
on the ACP domain of the same module and the polyketide intermediate bound to
theACP domain of the preceding module. All elongating modules present these
core domains, while the loading module lacks a functional KS domain and the last
module contains an additional TE domain, for release of the finished polyketide
from the PKS. Most PKS modules contain additional domains for processing the
newly formed b-keto: the b-ketoreductase (KR), the dehydratase (DH) and the
enoylreductase (ER) domains carry out the reactions (source: Donadio,
Monciardini & Sosio 2007).
PKS is known from both the systems of eukaryotes and prokaryotes. This enzyme
catalyses the fusion of carbon chains into long polymers via Claisen condensation
reaction (Heath & Rock 2002). PKS is related to fatty acid synthase structurally and
functionally as both of the enzymes catalyse the condensation of activated primary
metabolites to produce β-ketoacetyl polymers attached to the enzyme via thioester
bonds (Donadio, Monciardini & Sosio 2007). In synthesis of polyketides, these
reduction steps are eliminated partly or completely in a controlled way and thus
results in polyketides chain with respect to the production of β-hydroxyl, β-ketone
and alkyl groups (Fujii et al. 2001; see Figure 1.6 for an overview of PKS).

P a g e | 16
PKS has been characterized in terms of their subunits number and the synthesis
mode, such as type I modular PKS, type I iterative PKS, type II PKS and type III PKS
(Ansari et al. 2004). Type I modular PKS that can be found in bacteria is the best
categorized class, but the functional information derived from these generally
apply to other categories (Watanabe & Ebizuka 2004). Type I PKS are categorized
by being multi enzymes, carrying out enzymatic domains that are needed for
polyketides formation, in particular, clinical and economical macrolide polyketides
production, for instance rifamycin and erythromycin A (Ansari et al. 2004). For
type II PKS, the catalytic domains are located on individual proteins which interact
to produce a functional PKS enzyme complex (Ansari et al. 2004). The type III PKS
is different from the types I and II as it does not rely on acyl carrier protein
domains (Meier & Burkart 2009).
Figure 1.7: Basic steps during NRPS. Each NRPS module consists of three core
domains: an adenylation (A) domain, which selects the cognate amino acid,
activates it as an amino acyl adenylate and transfers it to the T domain (also
known as peptidyl carrier protein, or PCP) where a thioester bond is formed, a
condensation (C) domain, responsible for peptide bond formation between the
amino acid present on the T domain of the same module and the peptidyl
intermediate bound to the T domain of the preceding module, and the T domain
itself. Usually, all elongation modules present these core domains. A dedicated
loading module (carrying just A and T domains) and a termination module,

P a g e | 17
containing a thioesterase (TE) domain, usually complete the NRPS assembly line
(source: Donadio, Monciardini & Sosio 2007).
NRPS is a group of enzymes that typically found in most of the bacteria and fungi
which synthesizes non-ribosomal peptides, a family of complex natural products
synthesized from amino acid monomers (see Figure 1.7 for an overview of NRPS).
NRPS is achieved by the thiotemplate function of modular enzyme complexes
known collectively as peptide synthetases (Radjasa & Sabdono 2003). It has been
established that the specific combination of modules and various functional
domains within the peptide synthetase determines the structure and the activity of
peptide product (Neilan et al. 1999). Most non-ribosomal peptides from
microorganisms are classified as secondary metabolites, rarely having a role in
primary metabolism, growth or reproduction, but instead having evolved to
benefiting the producing organisms (Neilan et al. 1999). The products of microbial
NRPS include the immunosuppressant cyclosporine and antibiotics such as
erythromycin, gramicin S, lovastatin, rapamycin, surfactins, and tyrocin A
(Kleinkauf & Von Döhren 1996; Du, Sánchez & Shen 2001). NRPS usually works in
conjunction with PKS to give hybrid products which are significant pharmaceutical
products (Ansari et al. 2004).
NRPS is organized based on modules, where each of the modules is responsible to
catalyse a single cycle of product length elongation and modification of that
functional group. The minimum set of domains necessary for a single elongation
cycle consists of a module with Thiolation (T), Adenylation (A) or Peptidyl Carrier
Protein (PCP) and a Condensation (C) domain. The structural variation of the
peptide product is determined by the number and order of the module as well as
the type of domains present in a module of NRPS (Ansari et al. 2004). Thus, with
advanced techniques such as polymerase chain reaction (PCR), the screening for
the presence of PKS and NRPS genes is possible by using specific primers of PKS
and NRPS.

P a g e | 18
1.9 Significance and aims of the present study and dissertation outline
To our knowledge, microbial communities in the eastern South China Sea and the
Celebes Sea are practically unknown and we are therefore missing vital data to
understand these ecosystems. Understanding will help to predict their reaction to
changes in the global climate and other factors such as anthropogenic pollution.
One major and globally important role of surface bacteria is their involvement in
the breakdown or osmoregulation of DMSP to DMS or MeSH and this will be the
first comprehensive study looking at culturable bacteria in the eastern South China
Sea and their potential roles in the DMS(P) cycle.
In chapter 3, we look at samples from different regions to: (a) distinguish
differences in species distributions and (b) discuss their potential involvement in
the DMS(P) cycle.
Another globally important role of bacteria is their involvement in the coral
defence. The biodiversity and natural diversity of coral reefs in our region are
under threat from various anthropogenic and natural impacts, causing major
changes in their structure and function. Current research suggests that coral reefs
support an unknown number of organisms that may prove to be of major benefit in
the treatment of critical human diseases. In chapter 4, we have isolated bacteria in
a local reef and looked at their potential involvement in coral defence. The
isolation of native microbes from the coral SML allowed us to determine the
species composition and abundance of various bacteria in the SML. Results from
this study will update our current understanding of the basic ecology of coral-
associated microbial communities. This will help improve monitoring and
management efforts and aid in related issues of coral health.
The objectives of this study are:
i. Isolation and identification of native microbes in the South China Sea and
the coral SML.
ii. Testing of bacterial isolates for potential DMSP-degrading and/or antibiotic
properties.
iii. Determination of the effects of elevated temperatures on the antibiotic
properties of bacterial isolates.

P a g e | 19
CHAPTER 2
Methodology
2.1 Field sampling
In November 2011, a core field campaign took place in the South China Sea, and
along the coastline of Peninsula Malaysia and Borneo using the Sonne Research
Vessel, the DLR Falcon aircraft, satellites, and land-based investigation teams (see
Figure 2.1 for a schematic overview of activities and cruise track). The project was
supported by the 7th Framework programme of the European Union (call
ENV.2008.1.1.2.1) and is called Stratospheric Ozone: Halogen Impacts in a Varying
Atmosphere (SHIVA). By combining measurements from land, ship, aircraft, and
space-based platforms, with sophisticated numerical models, SHIVA aims to better
predict the rate, timing and climate-sensitivity of ozone-layer recovery, and
identify potential risks to that recovery.
Figure 2.1: Overview of the SHIVA cruise (source: http://shiva.iup.uni-
heidelberg.de/a_activities.html).

P a g e | 20
The sampling of the RV Sonne was supported by local cruises to provide
complimentary data from the coast. As part of the complementary three-part
series of local boat deployments, two transects were organised in Kuching on the
16th and 19th November 2011, one in Kota Kinabalu on the 23rd November 2011,
and one in Semporna on the 26th November 2011, respectively. Air and water
samples were collected at (at least) 5 stations with intervals of 5 km, along
transects at 1 to 20 km off the coasts.
The main aims of the local boat deployments were two-fold:
i. To obtain coastal samples to complement open ocean samples made by RV
Sonne, Falcon aircraft and satellites;
ii. To enable the exchange of samples collected at the coasts (for example VSLS
and nutrients) to be taken onboard RV Sonne for further analyses, and
those collected onboard RV Sonne (sensitive biological samples) to be taken
back for storage and further analyses.
The rendezvous stations were at the RV Sonne diurnal stations on the 19th
(Kuching) and 23rd (Kota Kinabalu) November 2011. Samples used in this thesis
were collected from 10 stations in Kuching (16th and 19th November 2011), five
stations in Kota Kinabalu (23rd November 2011) and eight stations in Semporna
(26th November 2011).
The water samples were collected using a Ruttner water sampler up to 10 m depth
and stored in sterile water bottles placed in cooling boxes to be transported back
to the laboratory for further analysis.
2.1.1 Reef samples
Samples of coral mucus from corals of different colonies, sediment and water
samples (up to 5 m depth) were collected from the reefs of Talang-talang (see
Figure 2.2) and its surrounding waters in July 2011.

P a g e | 21
Figure 2.2: Overview of the Talang-talang Islands just off the shores of Kuching,
Sarawak (source: Yahya, Hassan & Husaini 2012).
Loose coral fragments were collected and brought to the surface. The corals were
held upside down, allowing excess water to drip off and fresh mucus to form at the
surface of the coral. Coral mucus were dripped into sterile falcon tubes (see Figure
2.3) and stored in in cooling boxes maintained at 4 °C to be transported back to the
laboratory for further analysis.

P a g e | 22
Figure 2.3: Collection of fresh coral mucus.
Sediment samples were collected at areas adjacent to coral colonies in sterile falcon
tubes. Water samples were collected in sterile bottles at the surface and 5 m depth
while diving. These samples were also stored in cooling boxes maintained at 4 °C to be
transported back to the laboratory for further analysis.
2.2 Laboratory procedures
2.2.1 Isolation of bacteria
Most marine bacteria face an oligotrophic environment with diverse needs for growth
(Schut, Prins & Gottschal 1997) so to avoid a ‘nutrient shock’, the sea water samples
and coral mucus were streaked on marine agar at half strength. Bacterial colonies
were isolated based on their morphological differences. Colonies were picked and
purified by repeated streaking on plates. Pure cultures were preserved as a glycerol
suspension (20%, w/v) at -70 °C (Feltham et al. 1978).

P a g e | 23
2.2.2 Molecular characterisation
Small subunit ribosomal RNA (16S rRNA) has been proven to be most useful for
establishing evolutionary relationships because of their high information content,
conservative nature, and universal distribution (Lane et al. 1985). The 16S
sequence analysis is used in two major applications: (a) identification and
classification of isolated pure cultures and, (b) estimation of bacterial diversity in
environmental samples without culturing through metagenomic approaches. New
bacterial isolates are identified based on the 16S sequence homology analysis with
existing sequences in the databases (Rajendhran & Gunasekaran 2011).
Bacterial isolates were grown overnight in half strength marine broth at 30 °C and
pelleted by centrifugation at 13,000 x g for 5 min. The pellet was resuspended in
50 µl of TE buffer (pH 8.0). Three cycles of freezing in a -80 °C freezer and thawing
in a 85°C water bath were conducted to release DNA from the microbial cells.
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of bacterial
16S rRNA genes was performed with broad-specificity primers 8F (Eden et al.
1991) and 519R (Lane et al. 1985).
Amplification was performed by using REDTaq® ReadyMix™ PCR Reaction Mix
(Sigma Aldrich) using instructions provided by the Sigma Aldrich with the
following cycling conditions:
Initial denaturation at 96 °C for 4 min.
40 cycles of:
- 96 °C for 1 min.
- 55 °C for 2 min.
- 72 °C for 3 min.
Final elongation at 72 °C for 4 min.
Samples of extracted DNA were analyzed on a 1% agarose gel containing 1 µg of
ethidium bromide per ml (see Figure 2.4).

P a g e | 24
Figure 2.4: 16S rRNA bands of bacterial isolates. Impure bands can be seen at BSD
16-5, 16-7, 16-11. These isolates were later reisolated to ensure pure cultures.
Nucleotide sequences were determined by the dideoxynucleotide method by cycle
sequencing of the purified PCR products. An ABI Prism BigDye Terminator Cycle
Sequencing Kit was used in combination with an ABI Prism 877 Integrated
Thermal Cycler and ABI Prism 377 DNA Sequencer (Perkin Elmer Applied
Biosystems).
Sequences (typically 500 bp) were analysed against the NCBI (USA) database
(Zhang et al. 2000) using BLAST program packages and matched to known 16S
rRNA gene sequences. Gene sequences were corrected manually. Results are
shown in the Appendix. Sequences were aligned and phylogenetic trees were
created with MEGA 5 (Tamura et al. 2011) using the neighbor-joining method.
2.2.3 Clone libraries from water samples
2.2.3.1 Extraction of genomic DNA from sea water
Total DNA from a few selected water samples from the local SHIVA cruises were
cloned (see chapter 3 for further information). Seawater from different depths (1,
5 and 10 m) was collected using a Ruttner water sampler, filtered onto a 0.22 µm
membrane filter (Milipore). The filters were immersed in saline ethanol (70%
EtOH, 0.9% NaCl) and kept at -22 °C until further processed in the laboratory.
The filtrate samples were sonicated for 20 seconds to dislodge bacterial cells from
the filter and a total of 10 ml of each sample centrifuged at 10,000 rpm for 10 min

P a g e | 25
to concentrate the samples. The I-genomic BYF DNA extraction mini kit and the I-
genomic CTB DNA extraction mini kit (iNTRON Biotechnology, Korea) were used
on the Kuching samples, while three freeze and thaw cycles followed by ethanol
washing were carried out on the Kota Kinabalu samples. No samples were
processed for Semporna.
Prior to the freeze and thaw cycles, 10 ml of each sample were pelleted by
centrifugation at 5,000 rpm and 4 °C for 40 min. The pellets were resuspended in
50 µL of TE buffer (10 mM Tris-HC pH 8.0, 1 mM EDTA). Three cycles of freezing in
a -80 °C freezer for 3 min and thawing in a 85 °C water bath for 3 min were
conducted to release DNA from the microbial cells.
The bacterial DNA were amplified by polymerase chain reaction (PCR) using
broad-specificity primers 8F (Eden et al. 1991) and 519R (Lane et al. 1985).
Amplification was performed by using REDTaq® ReadyMix™ PCR Reaction Mix
(Sigma Aldrich) using instructions provided by the Sigma Aldrich with the
following cycling conditions:
Initial denaturation at 96 °C for 4 min.
40 cycles of:
- 96 °C for 1 min.
- 55 °C for 2 min.
- 72 °C for 3 min.
Final elongation at 72 °C for 4 min.
2.2.3.2 DNA cloning and plasmid extraction
The replicated DNA was inserted into vectors using the p-GEMT Easy Vector
Systems (Promega, USA) cloning kit and cloned with Escherichia coli JM109
competent cells as the host cell. The white colonies on the cloning agar plate which
contain species DNA were selected.
Plasmid extraction by alkaline Lysis method (Birnboim & Doly 1979) was carried
out on the selected white colonies. Each colony was cultured in 5 ml Luria Broth
(Conda Laboratories, Spain) and incubated overnight in an incubator shaker (37

P a g e | 26
°C, 250 rpm). After incubation, an Eppendorf tube (1.5 ml) was filled with bacterial
culture and centrifuged at 13,000 rpm for 1 min. The supernatant was removed
and more bacterial culture was added and centrifuged to obtain more plasmid. To
resuspend the pellets, 200 µl of Solution 1 (50 mM glucose, 25 mM Tris-HCL pH 8.0
and 10 mM EDTA pH 8.0) were added before adding 400 µl of Solution 2 (1%
Sodium Dodecyl Sulphate, 0.2 N NaOH). The tubes were then inverted 5 times and
incubated at room temperature for 5 min. 300 µl of Solution 3 (3 M K+, 5 M
acetate) were added and the tubes were inverted five times. After inverting the
tubes, they were placed on ice for 10 minutes. After the incubation, the tubes were
centrifuged for 5 min at 13,000 rpm. The supernatant was transferred into a new
tube and filled with isopropanol before incubating the tubes at room temperature
for 2 min. The tubes were centrifuged again at 13,000 rpm for 5 min, the
supernatant was removed and 1 ml of ice-cold 70% ethanol was added. Lastly, the
tubes were subjected to quick spin (1 min) and the supernatant was removed. The
tubes were dried and 50 µl of TE buffer (10 mM Tris-HC pH 8.0, 1 mM EDTA) was
added before storage.
2.2.3.3 Bacterial communities based in the construction of clone
libraries
The following is a brief discussion of the clone libraries obtained in this study. The
data available for the bacterial communities based on clone libraries is not
significant as the sample size is too small; therefore the results were not included
in chapter 3.

P a g e | 27
Figure 2.5: 16S rRNA gene-based phylogenetic tree representing bacterial sequences
found in clone libraries from Kuching and Kota Kinabalu. The phylogenetic tree was
generated with distance methods, and sequence distances were estimated with the
neighbour-joining method. Bootstrap values ≥50 are shown and the scale bar
represents a difference of 0.05 substitution per site. Accession numbers for the
reference sequences are indicated.
The phylogenetic tree (see Figure 2.5) shows the evolutionary relationships
between the 12 bacterial clones from the samples with nine species obtained from
the NCBI BLAST program, based on similarities in the DNA sequences. The tree
showed only the Proteobacteria were eligible for comparison with the sample
species because of the repeat of the species in a majority of BLAST results, proving
that the bacteria species extracted from the samples are distantly related to this
group. The BLAST results also showed the highest query coverage of less than
75%, raising the possibility that the clones may be novel, yet-to-be-described
species. The species selected from the BLAST results showed a diversity of bacteria
from various parts of the world; ranging from a fish pathogen causing fish
pasteurellosis (Juíz-Río et al. 2005), to a bacteria found in the North Atlantic Ocean
(Muhling et al. 2008) and a bacteria found in the hot springs of Tunisia (Sayeh et al.
2010). The Kota Kinabalu species were grouped on a separate branch from the
Kuching species indicating that the two locations clearly contain two separate
bacterial communities. However, the identities of the sampled sequences could not
be identified due to limited overlap in sequences.

P a g e | 28
The species composition of the clone libraries differed dramatically from that of
the cultured bacterial community. It is important to note that the samples used
were collected at the same time from the same stations. This finding supports the
idea that a majority of microorganisms are not easily cultured using standard
microbiological techniques (Rappé & Giovannoni 2003).
The influence of riverine input with regards to species composition was further
supported by KCH (1) 1 metre which was obtained from Kuching 1911 Station 1 at
1 m depth, and which was grouped away from the other samples. The lower pH
and salinity of the surface water at the station allowed what may be a different
community to thrive. The other Kuching 1911 bacterium which was obtained from
the same station but at 10 m depth is grouped together with a bacterium from Kota
Kinabalu which is representative of ocean waters, showing that the denser sea
water at the location provides a slightly different environment, influencing the
community at that depth. It can also be observed that a most of our samples are
grouped together on a separate branch, raising the possibility that the bacteria
obtained are undescribed novel species.
2.2.4 PCR amplification of bacterial DMSP cleavage (dddP) and
demethylation (dmdA) genes
dmdA is a key gene in the catabolism of DMSP which involves demethylation
where DMSP is degraded to MeSH (Curson et al. 2008) while dddP is the one of the
most common gene which mediates DMSP cleavage in DMS production (Varaljay et
al. 2010; Curson, Fowler, et al. 2011). Based on gene abundance, we can measure
the distribution of DMSP degrading genes and hypothesize the involvement of
members of the bacterial community in the local sulphur cycle (Varaljay et al.
2012). The presence of DMSP degrading genes in our cultured bacterial isolates
were screened using degenerate dddP primers and universal dmdA primers.
Bacterial DNA was amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of dddP
genes was performed with degenerate dddP primers dddP_874F and dddP_971R

P a g e | 29
(Levine et al. 2012) while amplification of dmdA genes was performed with
universal dmdA primers dmdAUF160 and dmdAUR697 (Varaljay et al. 2010).
Amplification was performed by using REDTaq® ReadyMix™ PCR Reaction Mix (Sigma
Aldrich) using instructions provided by the Sigma Aldrich with the following cycling
conditions:
Initial denaturation at 95 °C for 30 s.
40 cycles of:
- 95 °C for 30 s.
- 41 °C for 30 s.
- 72 °C for 30 s.
Final elongation at 72 °C for 4 min.
Samples of extracted DNA were analyzed on a 1% agarose gel containing 1 µg of
ethidium bromide per ml (see Figures 2.6 and 2.7).
Figure 2.6: PCR-based screening of dmdA genes. Bands highlighted in this figure
indicate presence of the genes.

P a g e | 30
Figure 2.7: PCR-based screening of dddP genes. Bands highlighted in this figure
indicate presence of the genes.
2.2.5 PCR amplification of bacterial antimicrobial PKS and NRPS genes
Small subunit ribosomal DNA (16S rDNA) based approach has been very successful
in the search of secondary metabolites particularly by using specific degenerated
primers (Radjasa & Sabdono 2003). The application of PCR-based approach on
screening of secondary metabolites-producing parts is used to identify PKS and
NRPS genes within our cultured bacterial isolates.
Bacterial DNA was amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of PKS genes
was performed with PKS degenerated primers KSDPQQF and KSHGTGR (Piel
2002) while amplification of NRPS genes was performed with NRPS degenerated
primers A2gamF and A3gamR (Marahiel, Stachelhaus & Mootz 1997).
Amplification was performed by using REDTaq® ReadyMix™ PCR Reaction Mix
(Sigma Aldrich) using instructions provided by the Sigma Aldrich with the
following cycling conditions:
Initial denaturation at 94 °C for 2 min.
45 cycles of:
- 94 °C for 1 min.
- 55 °C for 1 min.
- 72 °C for 2 min.
Final elongation at 72 °C for 4 min.

P a g e | 31
Samples of extracted DNA were analyzed on a 1% agarose gel containing 1 µg of
ethidium bromide per ml (see Figures 2.8 and 2.9).
Figure 2.8: PCR-based screening of PKS genes. Bands highlighted in this figure
indicate presence of the genes.
Figure 2.9: PCR-based screening of NRPS genes. Bands highlighted in this figure
indicate presence of the genes.
2.2.6 Antimicrobial tests
2.2.6.1 Extraction of bioactive compounds
Crude extract from each mucus associated isolate was extracted using ethyl acetate
solvent. This extraction method is particularly useful for extraction of both
extracellular (excreted by bacteria into the medium) and intracellular bioactive
compounds.

P a g e | 32
The bacterial isolates were grown in half strength marine broth at 28, 30 and 32 °C
for three days. 20 ml of ethyl acetate was added into 20 ml of bacterial broth and
shaken on a rotary shaker overnight. The mixtures were poured into separating
funnels and the broth layer was discarded while the layer containing the ethyl
acetate phase was collected in pre-weighed beakers. Another 20 ml of ethyl acetate
were added into the funnel and the extraction was repeated to rinse out any
residue extract. The ethyl acetate extract was then dried in the fume hood to give a
solid and oily residue. The dried extract was kept at -20 °C until further use.
2.2.6.2 Inhibitory interaction tests (well diffusion assay)
Modified well diffusion assays were used to detect the antimicrobial activities of
samples (Ndyetabura, Lyantagaye & Mshandete 2010). The well diffusion assay is
an alternative approach to the Kirby-Bauer disk diffusion method (Boyle, Fancher
& Ross 1973) for testing antimicrobial activities of the isolate microbes.
The dried extract was weighed and the extracted metabolite was diluted to 500
ppm using dimethyl sulfoxide (DMSO) (Matu et al. 2012). Mucus associated
bacterial isolates (test organisms) were grown overnight in half strength marine
broth at 28, 30 and 32 °C. Wells with a diameter of 5 mm were punched into half
strength marine agar and the test organisms were swabbed onto the agar plates.
50 μl of extract from each bacterial culture were loaded each well.
Chloramphenicol and DMSO adjusted to concentrations of 500 ppm were used as
positive and negative controls. Chloramphenicol is a broad-spectrum antibiotic
and is effective against a wide variety of Gram-positive and Gram-negative bacteria
(Neu & Gootz 1996).
The agar plates were incubated at 28, 30 and 32 °C for three days. The agar plates
were observed for any zone of inhibitions and recorded (see Figure 2.10).

P a g e | 33
Figure 2.10: Inhibition zones from bioactive compounds with antimicrobial
properties.

P a g e | 34
CHAPTER 3
Bacterial Communities from the
eastern South China Sea and the
Celebes Sea, and Their Potential Role
in the DMS(P) Cycle
ABSTRACT
Little is known about the microbial communities in the South China Sea, especially
the eastern region and this study aims to expand our knowledge on the diversity of
culturable bacterial communities in Malaysian waters which include parts of the
eastern South China Sea (Kuching and Kota Kinabalu) and the Celebes Sea
(Semporna). Comparisons were made with regards to physico-chemical
parameters and bacterial communities. The diversity of bacterial communities in
these waters were analysed through isolation and identification (16S rDNA) of
culturable bacteria. It was observed that although the majority of bacteria in
Kuching, Kota Kinabalu and Semporna are members of the Proteobacteria group,
the composition of bacterial communities in these three areas did vary
significantly , and the changes were also mirrored in physico-chemical differences.
Riverine input was the highest in Kuching which was mirrored by dominance of
potentially pathogenic Vibrio, whereas the Kota Kinabalu community is more
indicative of an open ocean environment. Isolates obtained from Kota Kinabalu
and Semporna show that the communities in these areas have potential roles in
bioremediation, nitrogen fixing and sulphate reduction. One major and globally
important role of surface bacteria is their involvement in the breakdown or
osmoregulation of DMSP to DMS or MeSH. The cultured bacteria were screened for
the existence of two key genes (dmdA, dddP) which are involved in competing,
enzymatically mediated DMSP degradation pathways. This is to our knowledge the

P a g e | 35
first study looking at the diversity of culturable bacteria in coastal waters of
Malaysia and also their potential roles in the DMS(P) cycle.
Keywords: culturable bacterial communities, diversity, dimethylsulphide,
dimethylsulphoniopropionate
3.1 INTRODUCTION
The South China Sea is a marginal sea that is part of the Pacific Ocean,
encompassing an area from the Singapore and Malacca Straits to the Strait of
Taiwan (Morton & Blackmore 2001). The Celebes Sea is connected to the South
China Sea through the Sulu Sea (Yoshida, Nishimura & Kogure 2007). While the
bacterial community structure in these two regions have been previously reported
to display some similarities when compared (Yoshida, Nishimura & Kogure 2007)
not much is known about the diversity and function of the microbial communities
in the South China Sea, especially regarding the eastern region (Kuching and Kota
Kinabalu) and the Celebes Sea and no studies on culturable communities in the
region have been made at this time.
Studies indicate that Alphaproteobacteria, together with SAR 11 and SAR 86
phylotypes, dominate bacterial communities of the surface ocean waters (up to
50% of rDNA analyses; (González et al. 2000). Members of the
Gammaproteobacteria, and Bacteroidetes also often make up a significant portion
of marine microbial communities (Abell et al., 2005; Alonso et al., 2007). Other
common bacteria found in marine sediment surfaces include
Gammaproteobacteria, Bacteroidetes and sulphate reducing bacteria (Stevens &
Brinkhoff 2005). However, information about the diversity and function of the
microbial communities in the South China Sea is still sparse; especially regarding
the eastern region (Kuching and Kota Kinabalu) and the Celebes Sea. To our
knowledge, there is no study on regional scale or large-scale distribution patterns
of microbes in the Malaysian area of the South China Sea. Most studies about
bacterial communities focused on regions near China, such as those carried out by
Li et al. (2006), Jiang et al. (2007) and Tao, Peng & Pinxian (2008). All three studies
used culture-independent techniques to reveal the community structure and

P a g e | 36
diversity of the predominant bacteria at the sampling environment. Their results
showed no significant difference in community structure with depth at each
location, but significant difference between geographical sites and hosts. The latter
two studies revealed that most lineages within the Proteobacteria represented
uncultured microorganisms, suggesting that a vast amount of microbial resource in
the South China Sea is unknown and unexplored.
Studies of microbial communities in rivers and estuarine communities have shown
that the Alpha-, Beta- and Gammaproteobacteria, and gram-positive bacteria are
the dominant groups. This is true for different regions of the world such as the
Columbia River (Crump, Armbrust & Baross 1999) and Changjiang River in China
(Sekiguchi et al. 2002).
In an interesting study, Nakase et al. (2007) found that a predominance of
Alphaproteobacteria and the Cytophaga–Flavobacterium cluster appear to be
beneficial for successful fish larval production, whereas Gammaproteobacteria
seem to increase their mortality. A comprehensive study of the bacterial
communities in the Malaysian area of the South China Sea could therefore
potentially be beneficial to fisheries as well.
The ocean is the major source of sulphur (Andreae 1986) and microorganisms
residing in the ocean have the ability to metabolise organic and inorganic sulphur
(Sievert, Kiene & Schultz-Vogt 2007). Dimethylsulphoniopropionate (DMSP) does
represent a major carrier for sulphur transfer through microbial food webs and
organic sulphur cycling in the pelagic ocean as it is an abundant component in
many phytoplankton taxa and is very prone to microbial degradation, making it
very appetizing for bacteria and grazers (Kiene, Linn & Bruton 2000). The
Roseobacter which are part of the Alphaproteobacteria lineage are mainly
responsible for the degradation of DMSP – a major source of oceans’ organic
sulphur – into methanethiol (MeSH) (González, Kiene & Moran 1999; González et
al. 2000). Hence, these bacteria play an important role in organic sulphur cycling
and are found in different regions ranging from the Sargasso Sea to the Black Sea
(González, Kiene & Moran 1999).

P a g e | 37
A competing metabolic pathway results in the production of dimethylsulfide (DMS)
from DMSP (González, Kiene & Moran 1999; Johnston et al. 2008). Due to the
highly efficient bacterial DMSP demethylation and DMS consumption processes,
only a small percentage (1-2 %) of DMSP produced by marine phytoplankton is
ventilated to the atmosphere as DMS (Levine et al. 2012). Despite the low
percentage, DMS does, however, represent a major source of biogenic sulphur to
the atmosphere, where oxidation products form cloud condensation nuclei and
ultimately influence radiative backscatter (Lovelock, Maggs & Rasmussen 1972;
Andreae & Crutzen 1997; Simó 2001).
Using genomic-based studies, enzymes responsible for DMSP degradation within
the microbial community can be identified more easily, which will contribute to
studies on the different biochemical pathways and regulatory factors involved in
DMSP metabolism. The DMSP demethylase gene (dmdA), encodes the first step in
the demethylation pathway. It has been revealed to be taxonomically diverse and
highly abundant (present in >50 % of marine bacterioplankton) (Howard et al.
2008). In comparison to dmdA, the genes involved in DMS production (dddD, dddL,
dddP dddQ, dddY, and dddW; all of which mediate the same step of DMSP
cleavage) are present in less than 10% of bacteria based on marine metagenomic
surveys (Todd et al. 2007; Curson et al. 2008; Howard et al. 2008; Curson, Sullivan,
et al. 2011; Todd et al. 2011; Todd, Kirkwood, et al. 2012). We focus on dddP as
past studies indicate that this gene, is one of most abundant occurring ddd genes
(Todd et al. 2009; Varaljay et al. 2012; Levine et al. 2012).
The present study tries to expand our knowledge on microbial communities in the
South China Sea by assessing the surface bacterial communities along the eastern
region of the South China Sea as well as the Celebes Sea. Isolates obtained were
also screened for the existence of key genes involved in the competing,
enzymatically mediated DMSP degradation pathways (dmdA, dddP) to identify
potential key players in the local DMS(P) cycle.
3.2 MATERIALS AND METHODS
3.2.1 Study site and sample collection

P a g e | 38
In conjunction with European and Malaysian research partners, the SHIVA
(Stratospheric ozone: Halogen Impacts in a Varying Atmosphere, EU call
ENV.2008.1.1.2.1) Western Pacific field campaign was performed in the fall of
2011. The core field campaign took place in the South China Sea and along the
coastline of Peninsula Malaysia and Borneo using the German Research Vessel
(RV) Sonne during a cruise leading from Singapore to Manila, Philippines (see
Figure 3.1 for cruise track).
Figure 3.1: The RV Sonne ship track leading from Singapore to Manila between
November 15-29, 2011 during the SHIVA SO 218 cruise.
Local cruises took place in Kuching on both November 16 and 19, 2011 (see Figure
3.2 for sampling stations), Kota Kinabalu (November 23, 2011; see Figure 3.3 for
sampling stations) and Semporna (November 26, 2011; see Figure 3.4 for sampling
stations) to provide additional data for coastal input. Samples for this study were
collected during the local cruises. Table 3.1 provides an overview of the sampling
stations and their respective GPS coordinates.
Raw sea water samples were streaked on marine agar at half strength and
bacterial colonies were isolated based on their morphological differences. Colonies

P a g e | 39
were picked and purified by repeated streaking on plates. Pure cultures were
preserved as a glycerol suspension (20%, w/v) at -70 °C.
Table 3.1: Locations of sampling stations at Kuching, Kota Kinabalu and Semporna.
Sampling stations
GPS coordinates
Kuching (1611) Kuching (1911) Kota Kinabalu Semporna
Station 1 1°38'36.24"N, 110°30'5.28"E
1°39'28.81"N, 110°31'24.42"E
6° 3'4.56"N, 116° 5'54.60"E
4°35'15.96"N, 118°32'58.14"E
Station 2 1°39'44.82"N, 110°32'7.26"E
1°42'44.24"N, 110°33'23.46"E
6° 3'5.82"N, 116° 4'1.45"E
4°38'37.86"N, 118°20'25.44"E
Station 3 1°42'2.80"N, 110°37'12.36"E
1°45'32.93"N, 110°35'16.86"E
6° 3'4.02"N, 116° 0'2.77"E
4°42'31.68"N, 118°23'19.38"E
Station 4 1°42'46.62"N, 110°39'17.40"E
1°48'2.16"N, 110°37'51.53"E
6° 2'49.85"N, 115°57'38.26"E
4°40'42.48"N, 118°32'11.34"E
Station 5 1°45'49.07"N, 110°41'27.77"E
1°50'54.15"N, 110°40'11.26"E
6° 4'23.64"N, 115°54'36.42"E
4°37'31.26"N, 118°41'5.99"E
Station 6 – – – 4°35'56.76"N, 118°43'19.14"E
Station 7 – – – 4°35'30.66"N, 118°42'17.10"E
Station 8 – – – 4°33'17.83"N, 118°39'22.57"E
*1611 and 1911 denotes November 16 and 19, 2011 respectively.
Figure 3.2: Locations of sampling stations in Kuching, Sarawak.

P a g e | 40
Figure 3.3: Locations of sampling stations in Kota Kinabalu, Sabah.
Figure 3.4: Locations of sampling stations in Semporna, Sabah.
3.2.2 DNA extraction and purification of cultured bacteria
The isolates were grown in marine broth at half strength and pelleted by
centrifugation at 13,000 rpm for 5 min. The pellets were then suspended in 50 µl of
TE buffer (10 mM Tris-HC pH 8.0, 1 mM EDTA). Three cycles of freezing in a -80 °C

P a g e | 41
freezer for 3 min and thawing in an 85 °C water bath for 3 min were conducted to
release DNA from the microbial cells.
3.2.3 PCR amplification of bacterial 16S rRNA genes
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of bacterial
16S rRNA genes was performed with broad-specificity primers 8F (Eden et al.
1991) and 519R (Lane et al. 1985). Amplification was performed by using
RedTaqMix (Sigma Aldrich) using instructions provided by the Sigma Aldrich with
the following cycling conditions: initial denaturation at 96 °C for 4 min, 40 cycles of
96 °C for 1 min, 55 °C for 1 min, extension at 72 °C for 2 min, and then a final
elongation at 72 °C for 4 min. Samples of extracted DNA were analysed on a 1%
agarose gel containing 1 µg of ethidium bromide per mL.
3.2.4 Sequencing and phylogenetic analysis
Sequences were analysed against the NCBI (USA) database (Zhang et al. 2000)
using BLAST program packages and matched to known 16S rRNA gene sequences.
Gene sequences were corrected manually. Results are shown in Appendix (see
Tables A.1 to A.4). Sequences were aligned and phylogenetic trees created with
MEGA 5 (Tamura et al. 2011) using the maximum likelihood method, and are
presented in Figures 3.7, 3.8, 3.9 and 3.10.
3.2.5 Nucleotide sequence accession numbers
The nucleotide sequences obtained in the present study have been deposited in
GenBank database (http://www.ncbi.nlm.nih.gov) under accession numbers
KF373266 to KF373440.
3.2.6 PCR amplification of bacterial DMSP cleavage (dddP) and
demethylation (dmdA) genes
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of dddP

P a g e | 42
genes was performed with degenerate dddP primers dddP_874F and dddP_971R
(Levine et al. 2012) while amplification of dmdA genes was performed with
universal dmdA primers dmdAUF160 and dmdAUR697 (Varaljay et al. 2010).
Amplification was performed by using RedTaqMix (Sigma Aldrich) with the
following cycling conditions: initial denaturation at 95 °C for 5 mins, 40 cycles of
95 °C for 30 s, 41 °C for 30 s, extension at 72 °C for 30 s, and then a final
denaturation and annealing for 1 min each. Samples of extracted DNA were
analyzed on a 1% agarose gel containing 1 µg of ethidium bromide per mL.
3.3 RESULTS AND DISCUSSION
3.3.1 Physico-chemical parameters
Basic physico-chemical parameters were recorded during sampling in Kuching
(November 19, 2011) and Kota Kinabalu (see Table 3.2). Values for Kuching
(November 16, 2011) and Semporna are not reported as the measuring
instruments were inconsistent and not functioning properly at the time of
sampling.
Table 3.2: Physico-chemical parameters measured from Kuching and Kota
Kinabalu at depths of 1 and 5 m.
Station Depth
(m) Temp
(°C) pH
Salinity (ppt)
Nitrate (ppm)
Phosphate (ppm)
Nitrite (ppm)
Silicate (ppm)
KCH-1 1 29.06 7.90 28.48 9.13 0.60 1.46 23.47 5 29.34 8.10 30.59 BD BD BD BD
KCH-2 1 28.98 8.25 30.65 2.02 0.33 0.60 7.48 5 29.11 8.25 30.89 BD BD BD BD
KCH-3 1 29.05 8.33 31.18 0.85 0.15 0.03 2.97 5 29.16 8.30 30.53 BD BD BD BD
KCH-4 1 29.00 8.33 31.07 0.49 0.10 0.00 4.52 5 29.10 8.29 30.52 BD BD BD BD
KCH-5 1 29.27 8.31 31.61 0.15 0.06 0.00 5.86 5 29.40 8.29 31.85 0.15 0.05 0.00 2.21
KCH mean
29.15 8.24 30.74 2.13 0.22 0.35 7.75
KK-1 1 29.80 8.44 31.85 1.04 0.15 BD 3.47 5 29.90 8.37 32.04 BD BD BD BD
KK-2 1 29.73 8.36 31.44 0.25 0.17 BD 3.21 5 29.78 8.33 31.95 BD BD BD BD
KK-3 1 29.55 8.34 31.88 0.23 0.11 BD 2.67 5 29.54 8.33 31.87 BD BD BD BD
KK-4 1 29.52 8.36 31.93 BD BD BD BD 5 29.45 8.34 31.91 BD BD BD BD
KK-5 1 29.68 8.38 32.03 0.13 0.03 BD 2.74 5 29.50 8.37 31.92 0.15 0.02 BD 2.79

P a g e | 43
KK mean
29.65 8.36 31.88 0.36 0.10 BD 2.98
*KCH denotes Kuching; KK denotes Kota Kinabalu.
**BD denotes values that are below detection limit.
Sampling stations at Kota Kinabalu stretched further away from the coastline and
displayed average values of salinity at 31.88 ppt, pH of 8.36 and temperature of
29.65 °C (see Table 3.2), all indicative of a typical ocean environment (Raven et al.
2005). The first sampling station at Kuching (KCH-1) was closer to the river mouth
of the Sarawak river and displayed a visible influence by riverine water with its
surface water displaying a salinity of 28.48 ppt and pH of 7.90 (see Table 3.2). The
salt and minerals in sea water made it denser than fresh water thus at the mouth of
the river where it meets the sea, the fresh water flows downstream across the
surface and the sea flows upstream at the bottom, resulting in an increasing trend
from low to high salinity as it goes deeper (salinity at depth is 2 units higher; see
Table 3.2). The pH values at KCH-1 also showed an increase from pH 7.90 at 1 m
depth to pH 8.10 at 5 m depth. However, the pH is still lower than the average pH
of seawater, indicating mixing of fresh- and marine water and/or supporting the
theory that not only the salt content of the sea water contributed to its pH but
other compounds present in sea water as well (Millero, Lee & Roche 1998).
Subsequent stations were however more representative of ocean waters with pH
around 8.3 and salinity around 31ppt (see Table 3.2). Temperatures at Kuching
and Kota Kinabalu are consistent at both 1 and 5 m depth (mean temperature of
29.15 and 29.65 °C respectively; see Table 3.2). The majority of the South China
Sea is in the tropical region where warm temperatures are constant and deviations
in temperature are not significant (McKnight & Hess 2000).
The riverine input at Kuching was also visible with significantly higher nitrate,
phosphate, nitrite, and silicate values closer to the river mouth (KCH-1 and KCH-2;
see Table 3.2). Nutrient levels in Kuching were also generally higher than in Kota
Kinabalu. To assess differences in distribution in the upper surface layers, samples
were also taken from 5 m depth (KCH-5 and KK-5; see Table 3.2). Interestingly, the
samples for Kota Kinabalu show consistent values, however, for Kuching, the

P a g e | 44
silicate drops from 5.86 ppm to 2.21 ppm within the first 5 metres (see Table 3.2),
indicative of an active biological pump (Dugdale, Wilkerson & Minas 1995).
3.3.2 Diversity of culturable bacterial communities
The present study provides what we believe to be the first analysis of cultured
bacterial communities from the eastern South China Sea and the Celebes Sea. The
bacterial communities from the sampling sites in Kuching, Kota Kinabalu and
Semporna were found to be diverse with representatives from several groups. The
total bacterial assemblage had representatives within the Alpha-, Beta- and
Gammaproteobacteria, as well as Firmicutes (see Figure 3.5). The general
similarity in groups can be explained by the use of a singular isolation media
(marine agar at half strength). However, the total number of bacterial isolates
obtained and assemblages at the three sampling sites were different as discussed
in the following.
From Kuching waters, 89 isolates were obtained over two sampling periods
(November 16 and 19, 2011). The diversity of bacterial phylotypes is presented in
Figures 3.7 and 3.8, and Tables 3.3 and 3.4. Overall, 89% of the cultured bacteria
were clustered within the Gammaproteobacteria, 8% within the
Alphaproteobacteria and 3% within the Firmicutes (see Figure 3.5). In Kota
Kinabalu waters, 39 isolates were obtained and the majority (72% of the cultured
bacteria) were clustered within the Gammaproteobacteria. The remaining isolates
were members of the Firmicutes (18%) and Alphaproteobacteria (10%) (see
Figures 3.5, 3.9 and Table 3.5). In Semporna waters, 48 isolates were obtained
from four phylogenetic groups. In total, 88% of the cultured bacteria were
members of the Gammaproteobacteria, 6% of the Firmicutes, 4% of the
Alphaproteobacteria and 2% Betaproteobacteria (see Figures 3.5, 3.10 and Table
3.6).

P a g e | 45
Figure 3.5: Pie charts illustrating the diversity of bacterial groups based on partial
16S rRNA gene sequences from bacteria isolated from (a) Kuching, (b) Kota
Kinabalu and (c) Semporna.
Figure 3.6: Phylogenetic groups of isolates from the waters of Kuching, Kota
Kinabalu and Semporna at depths of 1, 5 and 10 m.
Several ecological diversity indices frequently applied to microbial community
profile data were used in order to compare diversity among microbial
communities, enabling us to quantify diversity within the communities and
describe their numerical structure (see Table 3.3). Taxonomic classification up to
genus was used as some BLAST results could only relate the isolates to strains
which have been identified up to genus level.
0
5
10
15
20
25
30
35
40
1m 5m ≥10m 1m 5m ≥10m 1m 5m ≥10m
Kuching Kota Kinabalu Semporna
Nu
mb
er
of
iso
late
s
α-proteobacteria β-proteobacteria γ-proteobacteria Firmicutes

P a g e | 46
Table 3.3: Indices used to quantify the diversity of bacterial communities at
Kuching, Kota Kinabalu and Semporna.
Genus Kuching Kota Kinabalu Semporna
Total isolates (N) 89 39 48 Total genus (S) 14 15 14 Margalef index (DMg) 2.90 3.82 3.36 Shannon index (H’) 1.60 2.42 2.18 Shannon evenness (J’) 0.61 0.89 0.83 Smith and Wilson evenness (Evar) 0.49 0.69 0.59
*Formulae of diversity indices are from Margalef (1958), Shannon & Weaver (1963)
and Smith & Wilson (1996).
The Margalef index (DMg) measures species richness and is highly sensitive to
sample size (Magurran 2004). DMg is a more accurate index when it comes to
sample richness as it utilises absolute numbers compared to a density data matrix
(Gamito 2010). The commonly used Shanon index (H’) considers proportions,
ensuring no differences when using either data set (Gamito 2010). However,
calculated H’ values can be underestimations due to incomplete coverage as it
gives more weight to rare than to common species (S), making it more sensitive to
absolute (but not relative) changes in their abundance (Hill et al. 2003). Values for
both indices indicate that the bacterial community in Kota Kinabalu is the most
diverse with a greater number of genuses within the community, followed by the
communities in Semporna and Kuching.
The Shannon evenness index (J’) is derived from H’ which therefore makes it
sensitive to changes in evenness of rare species, thereby possibly overestimating
its true value (Hill et al. 2003). The Smith and Wilson evenness index (Evar),
however, is known to show greater resolution in reflecting true values (Blackwood
et al. 2007). The evenness values from both J’ and Evar show that not only does the
bacterial community in Kota Kinabalu have a greater amount of genuses present,
but the individuals in the community are distributed most equitably among these
genuses, and this corelation is replicated in the results from Semporna and
Kuching.

P a g e | 47
Figure 3.7: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kuching 1611. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-
joining method. Bootstrap values ≥50 are shown and the scale bar represents a

P a g e | 48
difference of 0.05 substitution per site. Accession numbers for the reference
sequences are indicated.
Figure 3.8: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kuching 1911. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-
joining method. Bootstrap values ≥50 are shown and the scale bar represents a

P a g e | 49
difference of 0.1 substitution per site. Accession numbers for the reference
sequences are indicated.
Figure 3.9: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Kota Kinabalu. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-

P a g e | 50
joining method. Bootstrap values ≥50 are shown and the scale bar represents a
difference of 0.05 substitution per site. Accession numbers for the reference
sequences are indicated.

P a g e | 51
Figure 3.10: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in Semporna. The phylogenetic tree was generated with distance
methods, and sequence distances were estimated with the neighbour-joining

P a g e | 52
method. Bootstrap values ≥50 are shown and the scale bar represents a difference
of 0.1 substitution per site. Accession numbers for the reference sequences are
indicated.
Gammaproteobacteria are the dominant phylogenetic group at all three locations
and at all sampling depths, followed by Alphaproteobacteria (see Figure 3.5).
Betaproteobacteria were only found at Semporna at 1m depth (see Figure 3.6).
These results correlate with existing records of microbial communities found in
coastal and open-ocean environments (Bernard et al. 2000) although samples from
Kuching have some riverine influence. The percentage of bacterial culturability is
2% (Button et al. 1993), thus, giving the possibility that although some groups may
be present in low numbers in cultures, they may still occupy a significant portion
of the bacterial community. However, to better understand their physiology and
ecology, the isolation of bacteria in pure culture remains an essential step in
microbial ecology (Bernard et al. 2000).
In the following, we discuss some highlights of the diversity found within the major
bacterial groups and also try and establish differences between the three different
sampling sites.
The cultured Alphaproteobacteria group consisted of representatives from the
Caulobacteraceae, Phyllobacteriaceae, Rhodobacteraceae and Rhodospirillaceae.
Caulobacteraceae were only found in Kota Kinabalu at 1 m depth with two isolates
related to Brevundimonas diminuta (see Figure 3.9). They are aerobic, non-
photosynthetic organisms which are widespread in natural bodies of water (Stove
Poindexter & Cohen-Bazire 1964). The closest related strains were Brevundimonas
diminuta strain c138 (GenBank accession number FJ950570; 99% similarity) and
Brevundimonas diminuta strain KSC_AK3a (GenBank accession number EF191247;
100% similarity), both of which have shown antibiotic resistance under extreme
conditions (La Duc et al. 2007; Li et al. 2010). Members of the family
Phyllobacteriaceae are part of a large variety of bacteria able to reduce nitrate to
nitrite or to molecular nitrogen (Zumft 1997; Labbé, Parent & Villemur 2004).
Isolates related to this family were grouped with Nitratireductor spp. and were

P a g e | 53
found in Kota Kinabalu and Semporna, at depths of 5 and 10 m (see Figures 3.9 and
3.10). Isolates belonging to Rhodobacteraceae were related to Roseovarius
pacificus strain 81-2 (GenBank accession number NR_043564) and Rhodobacter
capsulatus strain PSB-06 (GenBank accession number FJ866784), with overlaps
across Kuching and Semporna at 1 and 5 m depth (see Figures 3.7, 3.8 and 3.10).
The property to reduce nitric oxide is not restricted to denitrifiers within
Phyllobacteriaceae as strains of Rhodobacter capsulatus have been shown to be
able to transform nitric oxide to nitrous oxide at a significant rate (Bell, Richardson
& Ferguson 1992) and are also able to convert nitrous oxide to nitrogen through
the involvement of cytochrome bc1 complex (Itoh, Matsuura & Satoh 1989;
Richardson et al. 1989). Roseovarius pacificus was previously isolated from deep-
sea sediment of the Western Pacific Ocean and displayed the ability to degrade
polycyclic aromatic hydrocarbons (Wang, Tan & Shao 2009). Rhodospirillaceae are
typically purple non-sulphur photosynthetic bacteria, possessing the adaptive
capacity to grow photosynthetically and by oxidative phosphorylation (Saunders
1978). Cultures from this family were related to Thalassospira spp. which generally
form opaque, unpigmented or slightly yellow colonies on agar (López-López et al.
2002) and are potential bioremediation agents as they have the ability to degrade
polycyclic aromatic hydrocarbons and diesel fuel (Liu et al. 2007; Kim & Kwon
2010; Lai & Shao 2012). Isolates related to the Alphaproteobacteria at all three
areas seem to be involved in the nitrogen cycle and possibly in the degradation of
hydrocarbons.
The sole Betaproteobacteria that was cultured was related to Alcaligenes faecalis
(GenBank accession number JF264463; 88% similarity) which was previously
isolated from a coastal aquaculture environment. The isolate was obtained from
Semporna (see Figure 3.10), an area that is surrounded with aquaculture and
seaweed farms. Alcaligenes faecalis have also been found in salt marsh and
estuarine waters (Ansede, Friedman & Yoch 2001). It has the potential to degrade
DMSP to DMS via acrylate metabolism through the induction of β-
hydroxypropionate (Yoch, Ansede & Rabinowitz 1997; Ansede, Pellechia & Yoch
1999). It is easily recognizable by its diffusable yellow pigment on agar plates, a
characteristic not produced by the non-DMS-producing terrestrial Alcaligenes

P a g e | 54
faecalis strains (Ansede, Friedman & Yoch 2001) and our isolate displayed yellow
pigmentation. Other non-DMSP degrading strains of Alcaligenes faecalis have been
found to contribute towards coral defence by exhibiting anti-nematode activity
(Kooperman et al. 2007).
Within the Gammaproteobacteria group, isolates from Aeromonadaceae,
Pseudoalteromonadaceae, Shewanellaceae, Pseudomonadaceae and Vibronaceae
can be found across all three sampling sites (see Figures 3.7, 3.8, 3.9 and 3.10).
Uncommon groups of bacteria from Gammaproteobacteria that are related to
isolates in this study include Burzelia and Stenotrophomonas from Kuching (see
Figure 3.7); and Bowmanella, Idiomarina and Allomonas from Semporna (see
Figure 3.10).
Aeromonadaceae are primarily found in freshwater and associated with aquatic
animals and sewage, with the ability to reduce nitrate (Colwell, Macdonell & De
Ley 1986). An isolate related to Aeromonas enteropelogenes strain KADR14
(GenBank accession number JX136699; 99% similarity) was successfully cultured
from surface waters of Kota Kinabalu (see Figure 3.9). Aeromonas enteropelogenes
was previously found to be synonymous with Aeromonas trota on the basis of 16S
rRNA sequences (Collins, Martinez-Murcia & Cai 1993), and is a clinically relevant
species (Figueras, Guarro & Martínez-Murcia 2000). Oceanimonas spp. were
isolated from all three sampling locations (see Figures 3.7, 3.9 and 3.10). Isolates
were closely matched with Oceanimonas smirnovii strain 31-13 (GenBank
accession number NR_042963), which was previously isolated from the Black Sea
(Ivanova et al. 2005), had overlaps across Kota Kinabalu and Semporna.
Oceanimonas spp. are one of the earliest Gammaproteobacteria to have been
studied biochemically for multiple DMSP degrading genes including dddP (Yoch
2002). Studies have indicated that Oceanimonas spp. have multiple DMSP
degrading genes, allowing them to play a role in the sulphur cycle (Curson, Fowler,
et al. 2011). The availability of different ddd genes in Oceanimonas spp. implies
that DMSP may be a key substrate for this bacteria genus, enabling them to
produce DMS from DMSP (Ledyard, DeLong & Dacey 1993). They also have a
cytoplasmic DMSP lyase (de Souza & Yoch 1995; Yoch, Ansede & Rabinowitz 1997)

P a g e | 55
resembling the periplasmic dddY of Alcaligenes faecalis (de Souza, Yoch & Souza
1996). Our results displayed a slightly more diverse picture with isolates related to
Oceanimonas spp. from Kuching possessing dddP genes, whereas isolates from KK
and Semporna possessed dmdA genes (see Figures 3.14, 3.15, 3.16).
Members of the Pseudoalteromonadaceae have been previously reported to be
found in marine environments, invertebrates, fish and algae, generally without the
ability to denitrify (Ivanova, Flavier & Christen 2004). Isolates from Kuching
collected on November 16 and 19, 2011 were closely related to Pseudoalteromonas
maricaloris strain KMM636 (GenBank accession number NR_025009; 100%
similarity) and Pseudoalteromonas ganghwensis (GenBank accession number
DQ011614; 99% similarity) respectively, from surface water (1 m depth).
Pseudoalteromonas maricaloris strain KMM636 was isolated from an Australian
sponge (Fascaplysinopsis reticulate) which was collected at 10 m depth from the
Coral Sea, and exhibits antibacterial properties which possibly play a role in the
chemical defence of the sponge (Ivanova et al. 2002). Our cultures displayed
similar growth characteristics as Pseudoalteromonas maricaloris strain KMM636,
such as, translucent colonies which turn lemon yellow after 48 h of incubation
(Ivanova et al. 2002). Different strains of Pseudoalteromonas ganghwensis have
been shown to possess the ability to form biofilms and contribute in part to the
removal of excess proteineous matters from the sediment sludge of fish farms
(Iijima et al. 2009). Kota Kinabalu had isolates that were closely related to
Pseudoalteromonas lipolytica strain ZR064 (GenBank accession number
JX173567). Pseudoalteromonas lipolytica has only been recently characterised (Xu
et al. 2010) and has the ability to hydrolyse lipids and reduce nitrate to nitrite.
Members of the Shewanellaceae family generally have the ability to reduce nitrate
to nitrite and can be isolated from marine invertebrates and marine environments
(Ivanova, Flavier & Christen 2004). They are also known to be opportunistic
pathogens (Aguirre et al. 1994; Brink, van Straten & van Rensburg 1995). An
isolate from Kuching (collected on November 16, 2011) was related to Shewanella
chilikensis strain JC5 (GenBank accession number HM016088; 99% similarity). Not
much is known about Shewanella chilikensis as it has only been recently described

P a g e | 56
by Sucharita et al. in 2009. They were previously isolated from lagoon sediment
and do not share the family’s ability to reduce nitrate (Sucharita et al. 2009). Our
isolate was obtained near the river mouth at 10 m depth (Station 1; see Figure 3.7).
Isolates related to various Shewanella haliotis strains were cultured from the
waters of Kuching and Kota Kinabalu (see Figures 3.7 and 3.9) at depths of 1 and 5
m. Shewanella haliotis has been described to be sensitive to antibiotics (Kim et al.
2007). In Kota Kinabalu, an isolate related to Shewanella putrefaciens strain R1418
(GenBank accession number AB208055; 99% similarity) was found at 1 m depth. S.
putrefaciens is a hydrogen sulphide producing bacteria (Satomi et al. 2006)
frequently isolated from marine water and spoiling fish (Ziemke et al. 1998) and in
rare cases can be a human pathogen (Brink, van Straten & van Rensburg 1995).
The family Pseudomonadaceae is an extremely diverse group of bacteria.
Pseudomonas spp. are found at all three sites and all three depths. Isolates obtained
from Kuching were related to Pseudomonas aeruginosa strain 11.2 (GenBank
accession number JX286673; 100% similarity) and Pseudomonas oleovorans strain
HNS030 (GenBank accession number JN128264; 99% similarity). Both were
isolated from 1 and 7 m respectively at stations near the river mouth. Pseudomonas
aeruginosa is a clinically relevant opportunistic pathogen, ubiquitous in the
environment due to its resistance to the antibiotics and disinfectants, and
environmental adaptability (Stover et al. 2000). Pseudomonas oleovorans was first
isolated from oil-water emulsions used as lubricants and cooling agents in the
cutting and grinding of metals (Lee & Chandler 1941). The species is classified part
of the Pseudomonas aeruginosa group (Anzai et al. 2000). An isolate from Kota
Kinabalu is also found to be closely related to the same Pseudomonas oleovorans
strain mentioned (99% similarity) at 1 m depth, indicating an overlap between the
locations. Other Pseudomonas spp. related isolates from Kota Kinabalu include
Pseudomonas plecoglossicida strain AIMST Aie20 (GenBank accession number
JQ312025; 100% similarity) and Pseudomonas stutzeri strain UP-1 (GenBank
accession number AY364327; 99% similarity) at 5 and 1 m depth respectively.
Pseudomonas stutzeri is distributed widely in the environment with denitrifying
abilities (Lalucat et al. 2006). It has also been isolated as an opportunistic pathogen
from humans although infections are rare (Noble & Overman 1994), and are

P a g e | 57
involved in the degradation of biogenic and xenobiotic compounds (oil derivatives
– aromatic and nonaromatic hydrocarbons – and biocides) (Lalucat et al. 2006).
Pseudomonas plecoglossicida was first isolated by Nishimori, Kita-Tsukamoto &
Wakabayashi (2000) from ayu fish (Plecoglossus altivelis). It is well known as a fish
pathogen causing bacterial haemorrhagic ascites and has the ability to reduce
nitrate to nitrite (Nishimori, Kita-Tsukamoto & Wakabayashi 2000). This species is
also related to an isolate from Semporna, although of a different strain
(Pseudomonas plecoglossicida strain R8-591-1; GenBank accession number
JQ659971; 100% similarity) at 10 m depth. The occurence of these bacteria at Kota
Kinabalu and Semporna is possibly due to the numerous aquaculture farms
observed near or surrounding the two locations. Isolates closely related to
Pseudomonas pseudoalcaligenes strain K29411 (GenBank accession number
DQ298030) were also obtained from the surface waters (1 m depth) of Semporna.
This species was found to be synonymous with Pseudomonas oleovorans (Saha et
al. 2010) as they share the same morphological, biochemical, and molecular
characteristics. An isolate related to Pseudomonas fulva strain SMA24 (GenBank
accession number JQ618288; 100% similarity) was also cultured from the waters
of Semporna at 1 m depth. Pseudomonas fulva was originally isolated from
Japanese rice paddy and are commonly associated with rice plants, grains and
paddy fields (Uchino et al. 2001). Our study indicates that they can also be in other
environments, potentially involved in hydrocarbon degradation (Ni et al. 2010).
Members of Vibrionaceae are common in the marine environment, with species
found in hydrothermal vents, deep sea, open water, estuaries, and marine
sediments (Lee & Ruby 1994; Raguénès et al. 1997; Eilers et al. 2000; Maruyama et
al. 2000) and is the most heavily represented within the group, with several
overlaps between the sites displaying the same GenBank hits (e.g. Vibrio splendidus
strain AP625, GenBank ascension number GQ254509). Studies have suggested that
Vibrios degrade some ecologically hazardous compounds, such as polycyclic
aromatic hydrocarbons (Ramaiah et al. 2000), and are major decomposers of chitin
in the ocean (Nagasawa & Terazaki 1987; Hedlund & Staley 2001). Members of
Vibrionaceae have on the other hand also been shown to cause potentially lethal
diseases in humans and fish (Kusuda & Kawai 1998; McCarter 1999). More

P a g e | 58
recently, studies have shown Vibrio shiloi to be a coral pathogen, producing toxins
that inhibit photosynthesis and lyse zooxanthellae resulting in bleaching (Banin,
Ben-Haim, et al. 2000; Banin, Israely, et al. 2000). Species such as Vibrio
parahaemolyticus and Vibrio vulnificus have been shown to express virulence-
related properties such as production of the toxR gene (Lin et al. 1993; Okuda et al.
2001) and production of phenolate siderophore (Stelma et al. 1992). Vibrio harveyi
and Photobacterium spp. are luminous bacteria which often cause disease in
aquaculture (Baticados et al. 1990; Prayitno & Latchford 1995). While most Vibrio
spp. isolated from Kuching appear to be related to pathogenic strains, many of the
isolates from Kota Kinabalu and Semporna have potential roles in bioremediation,
nitrogen fixing and sulphate reduction.
Members of the cultured Firmicutes group consisted of members of the
Bacillaceae, Bacillaceae Family XII. incertae sedis and Paenibacillaceae. Isolates
from Bacillaceae were mostly related to Bacillus spp. and Lysinibacillus spp. with no
overlaps across the sampling sites. Bacillaceae are able to form endospores that
allow them to survive for extended periods under adverse environmental
conditions and have been shown to fix nitrogen (Jordan, McNicol & Marshall 1978),
synthesise antifungal peptides (Kajimura 1995) and plant growth promoting
substances, including gibberellin and indoleacetic acid (Broadbent, Baker &
Waterworth 1977; Turner & Backman 1991). As such, members of this group have
been used for agricultural crop enhancement (Wipat & Harwood 1999). Related
strains were obtained from agricultural soil and compost with the exception of
Bacillus sphaericus isolate BS11 (GenBank accession number AM269451; 100%
similarity) which was isolated from the East China Sea. Isolates from the
Bacillaceae Family XII. incertae sedis were matched with Exiguobacterium spp.,
which have previously been isolated from, or molecularly detected in, a wide range
of habitats including cold and hot environments with temperature range from -12
to 55 °C (Vishnivetskaya, Kathariou & Tiedje 2009). Interestingly, members of this
family were only isolated from Kota Kinabalu and Semporna where recent
temperature spikes resulted in mass coral bleaching in the region (Tan & Heron
2011) and of the three sampling sites, Sarawak was the only area with no reported
bleaching events (Tun et al. 2010). The different strains of Exiguobacterium spp.

P a g e | 59
did not overlap between sites (see Figures 3.9 and 3.10). The only Paenibacillaceae
isolated (from Kota Kinabalu at 1 m depth) was related to Brevibacillus
laterosporus strain GZUB11 (GenBank accession number FJ434663; 100%
similarity). Brevibacillus laterosporus are aerobic spore-forming bacteria that have
demonstrated pathogenicity towards insects and nematodes, with a potential to be
used as a biological control agent (Zahner et al. 1999; de Oliveira et al. 2004; Tian
et al. 2007). It is also reported to have the ability to produce lignin peroxidase
which can be used to degrade sulfonated azo dyes (Gomare, Jadhav & Govindwar
2008).
3.3.3 Variations in the bacterial communities in Kuching, Kota Kinabalu and
Semporna waters
The bacterial communities in the waters of Kuching, Kota Kinabalu and Semporna
are almost entirely unknown and have not been sampled by either culture or
culture-independent techniques. Previous studies have shown that microbial
community composition is influenced by physico-chemical variables such as
salinity, pH and temperature among others (Lamberti & Resh 1983; Nold & Zwart
1998; Arnon et al. 2005; Fierer & Jackson 2006). Our isolates are also mostly
related to species that are highly adaptable environmentally, indicating that the
communities in these waters employ various mechanisms that regulate the activity
of cells in natural communities (Bernard et al. 2000).
It is not surprising that cultured bacterial communities differ from clone libraries
which lacks culturable species (Ward, Weller & Bateson 1990; Bidle & Fletcher
1995; Suzuki et al. 1997; Bernard et al. 2000). Only 2% of bacteria grow in culture
(Button et al. 1993) as they can be affected by nutrients in growth mediums (Schut
et al. 1993), viral infection (Rehnstam et al. 1993) or lack of bacterial
commensalism (Saville Waid 1999; Grover 2000). Our cultures were isolated from
diluted marine agar, so these results may differ if the growth medium were used at
full strength.

P a g e | 60
3.3.4 Bacterial strains with potential to metabolise DMS and/or
demethylate DMSP
Since there are no published reports on the microbial biodiversity in the eastern
region of the South China Sea, their role in local biogeochemical cycles is also
unclear. To date, there are no available reports on the sulphur cycle in the region,
or of DMSP catabolism from bacterial communities of Kuching, Kota Kinabalu and
Semporna; neither are any bioinformatics data available on the prevalence of
dmdA and dddP genes in bacteria from these regions. As part of our effort to
understand the importance of bacteria in the region for the local sulphur cycle, we
screened our isolates for the existence of dmdA and dddP genes. Since our isolates
have been cultured in a very general way using a method that does not involve
selection for DMSP utilisation, any presence of these genes in our isolates is most
likely a fundamental trait of these bacteria.
Previously reported bacteria with the ability to demethylate DMSP and/or
metabolise DMS which we also managed to isolate and culture include Rhodobacter
and Roseovarius within the Alphaproteobacteria (González et al. 2003; Moran et al.
2007; Curson et al. 2008; Johnston et al. 2008; Todd et al. 2009; Kirkwood et al.
2010); the aforementioned Alcaligenes faecalis within the Betaproteobacteria;
Oceanimonas, Pseudomonas, Shewanella and Vibro within the
Gammaproteobacteria (de Souza & Yoch 1995; Yoch, Ansede & Rabinowitz 1997;
Ansede, Pellechia & Yoch 1999; Yoch 2002; Moran et al. 2007; Sievert, Kiene &
Schultz-Vogt 2007; Johnston et al. 2008; Raina et al. 2009, 2010); and Bacillus
within the Firmicutes (Todd et al. 2009).
DMSP lyase enzymes are present in diverse bacteria (Taylor 1993). Past studies
have revealed that DMS is a relatively minor product of DMSP metabolism under
normal circumstances in the marine water column (Kiene 1996b; Ledyard, Dacey
& Dacey 1996; van Duyl et al. 1998). Past studies found that the demethylation
pathway is the major fate of DMSP in seawater (Kiene 1996a). There are two
schools of thought regarding the regulation of the two competing pathways: Kiene,
Linn & Bruton (2000) and Simó (2001) hypothesized that the ‘bacterial switch’ is
influenced by bacterial carbon and sulphur demands and by DMSP availability;

P a g e | 61
while Slezak & Brugger (2001), Sunda et al. (2002), Toole et al. (2006), Archer et
al. (2010) and Levine et al. (2012) suggest that phytoplankton DMS production is
enhanced by UV-A radiation while bacterial DMSP consumption may be UV-A
intolerant.
Figure 3.11: Relative abundance of dmdA and dddP genes in cultured bacterial
communities from the waters of (a) Kuching, (b) Kota Kinabalu and (c) Semporna.
Bacteria isolated from Kuching displayed the highest abundance of both DMSP
degrading genes (36%) compared to communities isolated from Kota Kinabalu and
Semporna with 13 and 19 %, respectively. The bacterial community in Kota
Kinabalu has the highest percentage of dmdA gene occurrence (28%) while the
dddP gene responsible for DMS production appears to be most abundant (29%)
within the bacterial community Semporna (see Figure 3.11).

P a g e | 62
Figure 3.12: Presence of dmdA and/or dddP genes in bacterial isolates from the
waters of Kuching, Kota Kinabalu and Semporna.
The Gammaproteobacteria group is the largest identified fraction within the
communities at all three sampling sites with the potential for DMSP-assimilation.
Interestingly, the composition of the DMSP-assimilating community generally
mirrored the composition of the total bacterial community at each sampling site
(see Figures 3.6 and 3.12). This is unlike previous studies at the Gulf of Maine and
the Sargasso sea where the dominating group are the Alphaproteobacteria
(Malmstrom, Kiene & Kirchman 2004). Our findings indicate that the community
structure of Gammaproteobacteria in the area could be tightly linked to the local
sulphur and also possibly the nitrogen cycle.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
α-p
rote
ob
acte
ria
β-p
rote
ob
acte
ria
γ-p
rote
ob
acte
ria
Fir
mic
ute
s
α-p
rote
ob
acte
ria
β-p
rote
ob
acte
ria
γ-p
rote
ob
acte
ria
Fir
mic
ute
s
α-p
rote
ob
acte
ria
β-p
rote
ob
acte
ria
γ-p
rote
ob
acte
ria
Fir
mic
ute
s
Kuching Kota Kinabalu Semporna
Pe
rce
nta
ge
(%
)
Only dmdA Only dddP Both None

P a g e | 63
Figure 3.13: Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Kuching.
Figure 3.14: Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Kota Kinabalu.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
dmdA dddP Both
Vibrio sp.
Stenotrophomonas sp.
Shewanella sp.
Pseudomonas sp.
Pseudoalteromonas sp.
Oceanimonas sp.
Citrobacter sp
Burzellia sp.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
dmdA dddP Both
Vibrio sp.
Shewanella sp.
Pseudomonas sp.
Oceanimonas sp.
Enterobacter sp.

P a g e | 64
Figure 3.15: Relative abundance of dmdA and dddP genes in isolated
Gammaproteobacteria from Semporna.
Vibrio appear to be the dominant group within Gammaproteobacteria with dmdA
and dddP genes at all three stations. At Kuching, they appear well adapted with the
potential ability to undergo both competing pathways as a majority of them (88%;
see Figure 3.13) have both DMSP degrading genes; in Kota Kinabalu, Vibrio are the
only genus possessing dddP genes (see Figure 3.14); and in Semporna, Vibrio
appear to have even number of isolates with either one or both genes (see Figure
3.15).
It was previously hypothesized that DMSP production is an overflow mechanism
for when growth is unbalanced by lack of nutrients and the need to release excess
energy and excess reduced sulphur (Stefels 2000). These carbon-energy overflow
substances might evolve through natural selection to be useful in the cell (e.g.
through auxiliary structures or defence mechanisms) (Hill, White & Cottrell 1998).
Based on our findings, it seems likely that at low nutrient conditions, the
distribution of dmdA and dddP genes within the bacterial community become
more specific (e.g. more dmdA in KK and more dddP in Semporna; see Figure 3.11)
to adapt to a preferred pathway to degrade DMSP. This is discussed in the
following.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
dmdA dddP Both
Vibrio sp.
Shewanella sp.
Pseudomonas sp.
Photobacterium sp.
Oceanimonas sp.
Bowmanella sp.
Allomonas sp

P a g e | 65
The sampling locations at Kuching and Kota Kinabalu were observed to have heavy
shipping traffic which may influence the sulphur concentration in the area. Ship
plumes emit large amounts of anthropogenic nitrogen and sulphur into the
atmosphere, particularly within potential transport distance of land regions
(Corbett, Fischbeck & Pandis 1999) which may influence the algal production of
DMSP (Malin & Erst 1997).
The waters of Kota Kinabalu are known for having seasonal phytoplankton blooms
(Adam et al. 2011). The relative production of DMSP was suggested to depend on
nitrogen availability (Andreae 1986). Small haptophytes (e.g. coccolithophorids)
and many small dinoflagellates are typical of more nitrogen-deficient conditions,
so they have evolved to produce more DMSP, implying the probability of finding
higher levels of DMSP is greater under conditions of nitrogen depletion during
phytoplankton blooms (Simó 2001).
Nitrate and nitrite concentrations at Kota Kinabalu are low (0.36 ppm and not
detectable respectively; see Table 3.2), especially in comparison with Kuching,
indicating a low nutrient environment and suggesting the likelihood of high
concentration of DMSP in the area especially in the event of phytoplankton blooms.
The bacterial community in the area have possibly evolved to adapt to these
conditions and preferred the demethylation pathway as the occurrence of dmdA
genes in the community is the highest (see Figure 3.11). Due to riverine input, the
waters of Kuching have significantly higher in nutrients compared to Kota
Kinabalu (see Table 3.2). It is possible that the high nutrient environment at
Kuching forces the bacterial community in the area to be more flexible, hence the
diverse occurrence of dmdA and/or dddP genes in the community (see Figure
3.11) which allows them to use different pathways in DMSP degradation.
Sampling stations at Semporna were surrounded by seaweed farms. Micro- and
macroalgae and halophytic plants are abundant sources of DMSP in the marine
environment (Yoch 2002) and past studies (Scarratt et al. 2000) suggested that
bacteria growing near algal cells might be exposed to high local levels of DMSP,
which would lead to DMS yields that are higher than those inferred from bulk

P a g e | 66
seawater measurements. Our results support this as the dddP gene which is
responsible for DMS production is most abundant in the bacterial community at
Semporna.
Most studies show that bacteria are a major sink for DMS. Therefore, because
bacterioplankton are involved in both DMSP and DMS utilization, factors
controlling bacterial activity (e.g. UV radiation, temperature, nutrients and
dissolved organic matter) (Kirchmann 2000) ultimately play a role in controlling
DMS concentration.
Based on our preliminary observations, we believe that these isolates have the
ability to undergo both DMSP-degradation processes depending on current
environmental conditions. Considering the observed conditions of the sampling
sites, our data supports the hypothesis of a ‘bacterial switch’. However, UV
radiation measurements at the sampling locations were not taken at this time and
may play a role in the local sulphur cycle.
3.4 CONCLUSION
The bacterial communities that could be cultured from water samples taken in
Kuching, Kota Kinabalu and Semporna vary significantly. Differences between the
three areas can partly be explained by differences in physico-chemical parameters.
The Kuching community is influenced by higher nutrients and riverine input, and
is dominated by potentially pathogenic Vibrio spp., while the Kota Kinabalu
community is more indicative of an open ocean environment. The bacterial
community in Kota Kinabalu were found to be the most diverse, followed by
communities in Semporna and Kuching. This correlates with community evenness
from each site. Isolates obtained from Kota Kinabalu and Semporna show that the
communities in these areas have potential roles in bioremediation, nitrogen fixing
and sulphate reduction.
The preliminary study on the potential role of the bacterial communities in the
local sulphur cycle indicates a major role for Gammaproteobacteria, and in
particular Vibrio spp.. Occurrence of dddP, dmdA in Gammaproteobacteria mirrors

P a g e | 67
total Gammaproteobacteria community structure at all three sampling sites. Kota
Kinabalu and Semporna are dmdA- and dddP-dominant respectively, indicating
DMSP degrading pathway preferences of the communities in these areas. The
majority of isolates from Kuching have almost equal abundance both genes,
showing high adaptability in DMS(P) utilisation. Considering how the majority of
isolates within the three communities have DMSP-degrading genes, we believe that
they are influenced by bacterial carbon and sulphur demands and by DMSP
availability, giving some of them the ability to ‘switch’ pathways according to
necessity.
Further in-depth characterization of these communities through a combination of
physical, chemical and molecular biological studies will improve our
understanding of the role of bacteria in DMS(P) cycling in the eastern South China
Sea and the Celebes Sea and their impacts on climate change.
3.5 ACKNOWLEDGEMENTS
The authors would like to thank N.M. Levine for her assistance with the
identification of dddP and dmdA genes. We also thank the Sarawak Biodiversity
Centre for their kind permission to conduct research in Sarawak waters (Permit
No. SBC-RA-0094-MM). F.W.I. Kuek is funded by the Sarawak Foundation’s Tunku
Abdul Rahman scholarship. The research leading to these results has received
funding from the European Union's Seventh Framework Programme FP7/2007-
2013 under grant agreement no. 226224 - SHIVA.

P a g e | 68
CHAPTER 4
Bacterial Communities from Talang-
Talang Reef and Their Potential Role
in Coral Defence and the DMS(P) Cycle
ABSTRACT
The Talang-Satang region is situated off the coast of Sematan and is especially
important as it is one of the most diverse ecosystems found off Sarawak, including
a healthy coral reef. Complex microbial communities are known to have significant
influence over coral reef ecosystems. Through isolation and identification (16S
rDNA) of native microbes from corals, their surface mucus layer (SML), as well as
the surrounding sediment and waters, we were able to determine the species
composition and abundance of culturable bacteria in the coral reef ecosystem.
There was a clear distinction between the species found in the different parts of
the reef system. Isolates found attached to the coral were mostly related to Vibrio
spp., presumably attached to the mucus from the water column and surrounding
sediment. Cultures that were isolated from the SML were found to be closely
related to antibiotic producers with tolerance towards elevated temperatures and
heavy metal contamination. This specialized microbiota may be important for
protecting the corals from pathogens by occupying entry niches and/or through
the production of secondary metabolites (i.e. antibiotics). The role of the mucus-
associated bacteria for the defence of the coral was highlighted by the fact that
isolates related to pathogenic Vibrio spp. and Bacillus spp. were dominant amongst
the samples from the water column and sediment. Isolates with closest matches to
the known coral pathogens Vibrio coralliilyticus and Vibrio shiloi were obtained
from the SML and sediment samples respectively. The ability of isolates living in
the SML (associated) to inhibit isolates loosely attached to the SML (attached) and
vice versa was assessed at varying temperatures. All isolates were also screened
(using specific sets of primers) for the presence of type I modular polyketides

P a g e | 69
synthase (PKS) genes responsible for macrolide polyketides production and non-
ribosomal peptide synthetase (NRPS) genes with the ability to produce
immunosuppressants and other antibiotics. Our results indicate that the mucus-
associated bacterial microbes display maximum efficacy to ward off other bacteria
at 28 °C, however the inhibitory abilities of mucus-associated bacteria became less
effective as temperatures increased. Roseobacter spp. which are mainly responsible
for the degradation of dimethylsulphoniopropionate (DMSP) – a major source of
oceans’ organic sulphur – into methanethiol (MeSH) were also successfully
isolated from the SML. Bacterial DMSP degraders may also contribute significantly
to dimethylsulfide (DMS) production when temperatures are elevated.
Keywords: culturable bacterial communities; coral mucus; antimicrobial;
increasing temperatures; coral reefs
4.1 INTRODUCTION
Coral reefs are a rare feature in Sarawak due to its shallow sea shelf extending a
long way into the ocean. The reefs of Sarawak are limited to the areas off the
shores of Bintulu, Miri and offshore islands including the Talang-Satang region in
Kuching. The Talang-Satang region is situated off the coast of Sematan and is
especially important as it is one of the most diverse ecosystems found off Sarawak.
Reef-building corals have a narrow range of thermal tolerance, making them
extremely susceptible to temperature stress and outbreaks of coral diseases,
whereby the immunity of corals decrease (Baker, Glynn & Riegl 2008). This makes
the corals more vulnerable towards pathogens that are more virulent, especially at
higher temperatures (Goreau & Hayes 2008). The coral surface mucus layer (SML)
contains a complex microbial community that respond to such changes in the
environment (Ritchie & Smith 2004). The normal microbial flora within the SML
can protect the coral against pathogen invasion and disturbances which may have
led to coral diseases (Sutherland, Porter & Torres 2004). On average, 20-30 % of
bacterial isolates originating from coral SML possess antibacterial properties
(Ritchie 2006) that may assist the coral holobiont as a first line of defence against
pathogens (Shnit-Orland & Kushmaro 2009). It has been suggested that these

P a g e | 70
antimicrobial compounds are temperature sensitive (Ritchie 2006). Antibacterial
activity was found to be optimal at 26 °C and slightly decreased at 30 °C, with
partial inactivation occurring at 60 °C and complete loss of activity occurring at 80
°C (Shnit-Orland & Kushmaro 2009).
Polyketide synthase (PKS) and non-ribosomal peptide synthetase (NRPS) are
multiple enzymes that catalyse the oligomerization of small building blocks into
complex structure such as active compounds (Donadio, Monciardini & Sosio 2007).
NRPS usually works in conjunction with PKS to produce hybrid compounds which
are significant in pharmaceutical products (Ansari et al. 2004). Both biosynthetic
systems are responsible for synthesizing numerous biologically vital active
metabolites from microorganisms (Ayuso-Sacido & Genilloud 2005).
Coral reefs are potentially significant sources of dissolved DMS and DMSP,
particularly when corals are stressed or exposed during low tides (Broadbent &
Jones 2004). The concentrations of DMS and DMSP measured in mucus are the
highest reported in the marine environment, with values exceeding those reported
from highly productive polar waters (Fogelqvist 1991; Trevena et al. 2000, 2003)
and sea algal communities (Kirst et al. 1991; Levasseur, Gosselin & Michaud 1994;
Trevena et al. 2003).
DMSP in the marine environment is degraded by bacteria via two possible
pathways (González, Kiene & Moran 1999): the cleavage pathway which involves
splitting of the DMSP molecule into acrylate and DMS (important in mediating
organic sulfur emission into the atmosphere), and the demethylation pathway
which involves the removal of a methyl group from DMSP to produce 3-
methiolpropionate, which is then cleaved to methanethiol (MeSH). The high levels
of DMSP and DMS produced by corals, coupled with involvement of bacteria in
DMSP and DMS conversion, suggest that corals are likely to harbor bacterial
species involved in the cycling of these compounds (Raina et al. 2009, 2010).

P a g e | 71
4.2 MATERIALS AND METHODS
4.2.1 Study site and sample collection
Field sampling took place at the Talang-talang reef in July 2011. Samples for this
study were collected during the sampling. Figure 4.1 shows an overview of the
sampling region. Recorded temperatures at Talang-talang reef indicates that it is
exposed to temperatures between 28 to 31 °C throughout the year (Ng et al., pers.
comm.), however coral assemblages are healthy and diverse (Kaie et al., pers.
comm.) .
Figure 4.1: Overview of the Talang-talang Islands just off the shores of Kuching,
Sarawak. Enlarged map indicates sampling area.
Sea water, sediment and coral mucus samples were streaked on marine agar at half
strength and bacterial colonies were isolated based on their morphological
differences.
The coral mucus samples were isolated via two different methods. Mucus-
associated bacteria were isolated using ultraviolet (UV) light exposure for 15 min
as a form of sterilisation for the first layer of mucus to remove any possible surface
microbes that may attach to it during the transfer (Chang et al. 1985). A second
layer of coral mucus was streaked on top of the UV-exposed mucus, allowing only
mucus-associated bacteria to grow on the mucus-regulated surface (Ritchie 2006).
Mucus-attached bacteria were isolated without the UV exposure, allowing any
bacteria that happened to be attached to the mucus at the time of collection to be

P a g e | 72
grown. This approach is based on the hypothesis that true coral-associated
bacteria will be impervious to the antibiotic properties of the mucus, while many
attached bacteria may be sensitive to the bactericidal properties of the mucus.
Colonies were picked and purified by repeated streaking on plates. Pure cultures
were preserved as a glycerol suspension (20%, w/v) at -70 °C.
4.2.2 DNA extraction and purification of cultured bacteria
The isolates were grown in marine broth at half strength and pelleted by
centrifugation at 13,000 rpm for 5 min. The pellets were then suspended in 50 µL
of TE buffer (10 mM Tris-HC pH 8.0, 1 mM EDTA). Three cycles of freezing in a -80
°C freezer for 3 min and thawing in a 85 °C water bath for 3 min were conducted to
release DNA from the microbial cells.
4.2.3 PCR amplification of bacterial 16S rRNA genes
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of bacterial
16S rRNA genes was performed with broad-specificity primers 8F (Eden et al.
1991) and 519R (Lane et al. 1985). Amplification was performed by using
RedTaqMix (Sigma Aldrich) using instructions provided by the Sigma Aldrich with
the following cycling conditions: initial denaturation at 96 °C for 4 min, 40 cycles of
96 °C for 1 min, 55 °C for 1 min, extension at 72 °C for 2 min, and then a final
elongation at 72 °C for 4 min. Samples of extracted DNA were analyzed on a 1%
agarose gel containing 1 µg of ethidium bromide per mL.
4.2.4 Sequencing and phylogenetic analysis
Sequences were analysed against the NCBI (USA) database (Zhang et al. 2000)
using BLAST program packages and matched to known 16S rRNA gene sequences.
Gene sequences were corrected manually. Results are shown in Appendix (see
Table A.5). Sequences were aligned and phylogenetic trees were created with
MEGA 6 (Tamura et al. 2013) using the maximum likelihood method, and are
presented in Figures 4.3, 4.4 and 4.5.

P a g e | 73
4.2.5 Nucleotide sequence accession numbers
The nucleotide sequences obtained in the present study have been deposited in
GenBank database (http://www.ncbi.nlm.nih.gov) under accession numbers
KF373441 to KF373533.
4.2.6 PCR amplification of bacterial DMSP cleavage (dddP) and
demethylation (dmdA) genes
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of dddP
genes was performed with degenerate dddP primers dddP_874F and dddP_971R
(Levine et al. 2012) while amplification of dmdA genes was performed with
universal dmdA primers dmdAUF160 and dmdAUR697 (Varaljay et al. 2010).
Amplification was performed by using RedTaqMix (Sigma Aldrich) with the
following cycling conditions: initial denaturation at 95 °C for 5 mins, 40 cycles of
95 °C for 30 s, 41 °C for 30 s, extension at 72 °C for 30 s, and then a final elongation
at 72 °C for 4 min. Samples of extracted DNA were analyzed on a 1% agarose gel
containing 1 µg of ethidium bromide per mL.
4.2.7 PCR amplification of bacterial polyketide synthase (PKS) and non-
ribosomal peptide synthetase (NRPS) genes
The bacterial DNA were amplified by polymerase chain reaction (PCR) and PCR
products were purified using PureLink® PCR Purification Kit following the
manufacturer’s protocol (Invitrogen Life Technologies). Amplification of PKS genes
was performed with PKS degenerated primers KSDPQQF and KSHGTGR (Piel
2002) while amplification of NRPS genes was performed with NRPS degenerated
primers A2gamF and A3gamR (Marahiel, Stachelhaus & Mootz 1997).
Amplification was performed by using RedTaqMix (Sigma Aldrich) with the
following cycling conditions for PKS: initial denaturation at 94 °C for 2 min,
followed by 45 cycles of 94 °C for 1 min, annealing at 55°C for 1 min and extension
at 72°C for 2 min, and then a final elongation at 72 °C for 4 min. The following are
the cycling conditions for NRPS: initial denaturation at 94 °C for 2 min, followed by

P a g e | 74
40 cycles of denaturation at 95 °C for 1 min, annealing at 70°C for 1 min and
extension at 72 °C for 2 min and then a final elongation at 72 °C for 4 min.
4.2.8 Extraction of bioactive compounds
The coral mucus bacterial isolates were grown in 20 ml of half strength marine
broth at 28, 30 and 32 °C for three days. 20 ml of ethyl acetate was added into the
bacterial broth and shaken on a rotary shaker overnight. The mixtures were
poured into separating funnels and the broth layer was discarded while the layer
containing the ethyl acetate phase was collected in pre-weighed beakers. Another
20 ml of ethyl acetate were added into the funnel and the extraction was repeated
to rinse out any residue extract. The ethyl acetate extract was then dried in the
fume hood to give a solid and oily residue. The dried extract was then kept in -20
°C until further use.
4.2.9 Well diffusion assay
The dried extract was weighed and the extracted metabolite was diluted to 500
ppm using dimethyl sulfoxide (DMSO) (Matu et al. 2012). Coral mucus bacterial
isolates (test organisms) were grown overnight in half strength marine broth at
28, 30 and 32 °C. Wells with a diameter of 5 mm were punched into half strength
marine agar and the test organisms were swabbed onto the agar plates. 50 μl of
extract from each bacterial culture were loaded each well. Chloramphenicol and
DMSO adjusted to concentrations of 500 ppm were used as positive and negative
controls. Chloramphenicol is a broad-spectrum antibiotic and is effective against a
wide variety of Gram-positive and Gram-negative bacteria (Neu & Gootz 1996).
The agar plates were incubated at 28, 30 and 32 °C for three days. The agar plates
were observed for any zone of inhibitions and recorded.
4.3 RESULTS AND DISCUSSION
4.3.1 Diversity of culturable bacterial communities
The present study provides what we believe is the first analysis of cultured
bacterial communities from the reefs of Talang- Talang. The bacterial communities
from coral mucus, reef sediment and water column were found to be diverse with
representatives from several bacterial groups. The total bacterial assemblage had

P a g e | 75
representatives within the Actinobacteria, Proteobacteria (Alpha- and
Gammaproteobacteria), as well as Firmicutes (see Figure 4.2 for an overview of the
major groups). The total number of bacterial isolates obtained and assemblages
from the three reef environments are discussed in the following.
Figure 4.2: Pie charts illustrating the diversity of bacterial groups based on partial
16S rRNA gene sequences from bacteria isolated from (a) coral mucus, (b) water
column and (c) sediment.
A total of 93 isolates were cultured from coral mucus, water column and reef
sediment of the Talang-talang reef. Overall, 3% of the cultured bacteria were
clustered within the Actinobacteria, 76% within the Gammaproteobacteria, 6%
within the Alphaproteobacteria and 13% within the Firmicutes. From the coral
mucus, 39 isolates were obtained with the majority clustered within the
Gammaproteobacteria (64%), followed by Alphaproteobacteria (13%), Firmicutes
(13%) and Actinobacteria (8%). There is an unknown isolate that was cultured
from coral mucus. Its closest related sequence is unidentified (see Figure 4.3).
Within the water column, 82% of the isolates were Gammaproteobacteria, 9%
Actinobacteria and 9% Firmicutes. Isolates from reef sediment were less diverse
with cultures from only two bacterial groups: the Gammaproteobacteria (86%)
and Firmicutes (14%).

P a g e | 76
Figure 4.3: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in coral mucus. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-
joining method. Bootstrap values ≥50 are shown and the scale bar represents a
difference of 0.1 substitution per site. Accession numbers for the reference
sequences are indicated.

P a g e | 77
Figure 4.4: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in water column. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-
joining method. Bootstrap values ≥50 are shown and the scale bar represents a
difference of 0.05 substitution per site. Accession numbers for the reference
sequences are indicated.

P a g e | 78
Figure 4.5: 16S rRNA gene-based phylogenetic tree representing bacterial
sequences found in reef sediment. The phylogenetic tree was generated with
distance methods, and sequence distances were estimated with the neighbour-
joining method. Bootstrap values ≥50 are shown and the scale bar represents a

P a g e | 79
difference of 0.05 substitution per site. Accession numbers for the reference
sequences are indicated.
It has been established that mucus presents a specific environment which contains
vast microbial communities (Sharon & Rosenberg 2008). Similar coral associated
bacteria can be present in different species of corals that are also geographically
distinct (Shnit-Orland & Kushmaro 2009). The coral mucus layer is in constant
association with the surrounding water column, and bacteria may shift from the
water column to the mucus and vice versa (Kooperman et al. 2007). Therefore, it is
not surprising that there are overlaps between the mucus and its surrounding
environment.
Figure 4.6: Percentage of Vibrio isolates in mucus attached and mucus associated
communities.
The mucus associated isolates are related to representatives of bacteria
documented in earlier studies, including a subset of Vibrio spp. consistently found
in association with healthy corals (Ritchie & Smith 1995a, 1995b, 2004). Figure 4.6
shows that there is a higher percentage of Vibrios (91%) when comparing mucus
attached isolates to mucus associated isolates (29%). This illustrates the defensive
qualities of coral mucus, and how a potential composition shift from beneficial
bacteria to Vibrio dominance (which are known to be opportunistic) under
conditions of increased temperature can occur.

P a g e | 80
The isolates linked to the phylum Gammaproteobacteria consist of members of the
families Alteromonadaceae, Enterobacteriaceae, Moraxellaceae, Halomonadaceae
and Vibrionaceae. Vibrionaceae members are present in high numbers in all three
environments, where the predominant bacteria are related to Vibrio communis,
Vibrio harveyi and Vibrio parahaemolyticus. Vibrio spp. are often associated with
diseases in corals and other marine organisms (Rosenberg et al. 2007). Isolates
related to known coral pathogens, Vibrio coralliilyticus strain LMG 21349
(GenBank accession number AJ440004) and Vibrio shiloi (GenBank accession
number AF007115; 99% similarity) were cultured from mucus and sediment
samples respectively. These findings indicate that the reef environment harbour
potentially pathogenic bacteria which can cause disease under the right conditions.
However, some Vibrios establish mutualistic partnerships with corals by providing
nutrients and secondary metabolites to their hosts (Ritchie 2006). Vibrio spp.
associated with the coral mucus are known to produce antibacterial compounds
against several pathogens, thereby protecting the coral host against pathogens
(Shnit-Orland & Kushmaro 2009).
Firmicutes are the second largest bacterial group in all three environments and are
dominated by members of the Bacillus genera. Bacillus spp. present in the mucus of
corals have been reported to exhibit antibacterial activity against pathogens
(Shnit-Orland & Kushmaro 2009) and those of marine origin have been reported to
produce unusual metabolites (Jensen & Fenical 1994) including peptide antibiotics
such as bacitracin, gramicidin and polymyxin B (Wiese et al. 2009).
The Actinobacteria are known for their production of many bioactive compounds
(Magarvey et al. 2004; Fiedler et al. 2005; Jensen et al. 2005) and may influence the
susceptibility of corals to pathogens (Rohwer et al. 2002). Isolates related to this
phylum are only found within the coral mucus. While the number of Actinobacteria
in this study may not be high, other studies have found that they are generally
found in corals (Nithyanand & Pandian 2009), and bacterial clone libraries of the
coral species included a significant proportion of Actinobacteria (Lampert et al.
2008). The distribution of Actinomycetes in the sea remains largely undescribed
and only a few of which were culturable (Webster et al. 2001). Only recently were

P a g e | 81
novel marine Actinomycetes discovered in sponges (Webster et al. 2001) and
ocean sediment (Mincer et al. 2002; Mincer, Fenical & Jensen 2005), and cultured
from corals (Lampert et al. 2008; Nithyanand & Pandian 2009).
Isolates related to the Alphaproteobacteria are only found in coral mucus samples.
A couple of the cultures are related to uncultured sequences (BCM 33 and 35-2; see
Figure 4.3) suggesting a possible novel species. Isolates related to Roseobacter spp.
and Sphingobium amiense strain D3AT58 (GenBank accession number JF459959;
97% similarity) were also obtained. Roseobacter spp. are widely associated with
corals (Frias-Lopez et al. 2002; Rohwer et al. 2002; Bourne & Munn 2005;
Kooperman et al. 2007; Bourne et al. 2008) and are potentially central to the
health of corals. Antibacterial activities of Roseobacter have been observed against
a wide range of marine pathogens (Hjelm et al. 2004). Coral associated bacteria
from this genus have also been previously implicated in the degradation of DMSP
(Raina et al. 2009, 2010). Thiotropocin, an antibiotic produced by Roseobacter, is a
sulfur compound that might be derived from DMSP metabolism (Wagner-Döbler et
al. 2004). These bacteria are suspected to be involved in a symbiotic relationship
with coral-cultured zooxanthellae (Raina et al. 2009) which produces high
concentrations of DMSP (Hill, Dacey & Krupp 1995; Broadbent, Jones & Jones
2002; Van Alstyne, Schupp & Slattery 2006). It is likely that the occurrence of
Roseobacter spp. within the coral mucus may be due to the availability of DMSP
produced by the zooxanthellae.
4.3.2 Bacterial strains with PKS and NRPS genes
Not all isolates from the coral mucus were capable of amplifying PKS and NRPS
gene fragments (see Figure 4.7). It appears that most of the isolates have the
potential to produce NRPS compounds (51%) and only 8% have the ability to
undergo PKS. Some isolates have the capability to form hybrids (18%) and may
play a more important role in coral defence.

P a g e | 82
Figure 4.7: Relative abundance of PKS and NRPS genes in cultured bacterial
communities from coral mucus.
The Gammaproteobacteria, the largest faction within the coral mucus community
is the only group with the potential ability to form PKS-NRPS hybrids (see Figure
4.8). The Alphaproteobacteria and Actinobacteria can only produce PKS
compounds while Firmicutes appear to be more dominant in NRPS.
Figure 4.8: Presence of PKS and/or NRPS genes in bacterial isolates from coral
mucus.
To estimate the ecological role of positive strains as well as their biotechnological
potency, inhibitory tests were carried out against other coral associated bacteria.
8%
51% 18%
23%
Abundance of PKS and NRPS genes in bacteria from coral mucus
PKS NRPS Both None
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
α-proteobacteria γ-proteobacteria Actinobacteria Firmicutes Unknown
PKS NRPS Both None

P a g e | 83
4.3.3 Role of mucus-associated bacteria in coral defence
The mucus isolates were separated into two groups: mucus attached (11 isolates)
and mucus associated (28 isolates).
Figure 4.9: Total inhibitions of mucus attached bacteria at temperatures of 28, 30
and 32 °C.
Figure 4.10: Total inhibitions of mucus associated bacteria at temperatures of 28,
30 and 32 °C.
The total inhibitions of mucus attached bacteria against all 39 coral mucus isolates
increased with temperature (see Figure 4.9). A total of 44 inhibitions occurred at
0
2
4
6
8
10
12
14
BCM22-1
BCM22-2
BCM 23 BCM24-1
BCM24-2
BCM 25 BCM26-1
BCM26-2
BCM 27BCM 28BCM 29
To
tal
inh
ibit
ion
s
Samples
28 °C 30 °C 32 °C
0
2
4
6
8
10
12
14
BC
M 3
1
BC
M 3
2
BC
M 3
3
BC
M 3
4
BC
M 3
5-1
BC
M 3
5-2
BC
M 3
6
BC
M 3
7
BC
M 3
8
BC
M 3
9
BC
M 4
0
BC
M 4
1
BC
M 4
2
BC
M 4
3
BC
M 4
4
BC
M 4
5
BC
M 4
6
BC
M 4
8
BC
M 4
9
BC
M 5
0
BC
M 5
1
BC
M 5
2
BC
M 5
3
BC
M 5
4
BC
M 5
6
BC
M 5
7
BC
M 5
8
BC
M 5
9
To
tal
inh
ibit
ion
s
Samples
28 °C 30 °C 32 °C

P a g e | 84
28 °C, 67 inhibitions at 30 °C and 69 inhibitions at 32 °C. For coral associated
bacteria, inhibition activity is highest at 30 °C (163 inhibitions). This is followed by
141 inhibitions at 28 °C and 107 inhibitions at 30 °C. A few isolates did not show
any inhibition at temperatures of 28 °C and/or 32 °C even though there was
activity at 30 °C (see Figure 4.10).
Figure 4.11: Average zone of inhibitions (cm) of mucus attached bacteria at
temperatures of 28, 30 and 32 °C.
Figure 4.12: Average zone of inhibitions (cm) of mucus associated bacteria at
temperatures of 28, 30 and 32 °C.
0.0
0.2
0.4
0.6
0.8
1.0
1.2
BCM22-1
BCM22-2
BCM 23 BCM24-1
BCM24-2
BCM 25 BCM26-1
BCM26-2
BCM 27BCM 28BCM 29Av
era
ge
zo
ne
of
inh
ibit
ion
(cm
)
Samples
28 °C 30 °C 32 °C
0.0
0.2
0.4
0.6
0.8
1.0
1.2
BC
M 3
1
BC
M 3
2
BC
M 3
3
BC
M 3
4
BC
M 3
5-1
BC
M 3
5-2
BC
M 3
6
BC
M 3
7
BC
M 3
8
BC
M 3
9
BC
M 4
0
BC
M 4
1
BC
M 4
2
BC
M 4
3
BC
M 4
4
BC
M 4
5
BC
M 4
6
BC
M 4
8
BC
M 4
9
BC
M 5
0
BC
M 5
1
BC
M 5
2
BC
M 5
3
BC
M 5
4
BC
M 5
6
BC
M 5
7
BC
M 5
8
BC
M 5
9
Av
era
ge
zo
ne
of
inh
ibit
ion
(cm
)
Samples
28 °C 30 °C 32 °C

P a g e | 85
Overall, inhibitory activities of the mucus attached bacteria appear to be most
effective at 30 °C (see Figure 4.11). The largest inhibitory zone is 1.0 cm at 30 °C by
isolate BCM 22-1. The second largest zone (0.9 cm) came from the extracts of BCM
25 (at 28 and 30 °C) and BCM 26-1 (at 32 °C). The inhibitory abilities of coral
associated bacteria were observed to become less effective as temperatures
increase (see Figure 4.12). Inhibition zones at 28 °C stay between 0.6 to 0.9 cm.
Activity were erratic at 30 °C, with the largest zone at 1.0 cm (isolate BCM 35-2)
and the smallest at 0.4 cm (isolates BCM 39, 44 and 58). At 32 °C, the zones were
between 0.3 to 0.7 cm, with a couple of isolates (BCM 32 and 50) exhibiting larger
zones at 0.9 cm.
Mucus attached isolates BCM 22-1 and 26-1 are closely matched with Vibrio
parahaemolyticus strain DHC22 (GenBank accession number JQ904733; 99-100 %
similarities) while isolate BCM 25 is closely matched with Vibrio harveyi isolate
VHJR19 (GenBank accession number DQ995251; 99% similarity). Strains of Vibrio
parahaemolyticus and Vibrio harveyi have previously been isolated through a
similar method (Ritchie 2006) and were reported to have PKS and NRPS genes and
exhibit inhibition activity against other coral-associated and pathogenic bacteria
(Radjasa & Sabdono 2003). Other strains Vibrio parahaemolyticus have also been
observed to be capable of producing a temperature regulated enzyme, superoxide
dismutase (SOD) that detoxifies oxygen radicals which has been suggested to be a
key virulence factor in the infection of corals (Banin et al. 2003). Vibrio harveyi
strains have also been reported to be antibiotic-resistant (Sussman et al. 2009;
Vizcaino et al. 2010) and implicated as part of bacterial consortiums that caused
yellow band and black band diseases (Barneah et al. 2007; Cervino et al. 2008),
and white plague (Sunagawa et al. 2009) in corals.
Mucus associated isolate BCM 35-2 is closely matched with an uncultured alpha
proteobacterium clone FF-20 (GenBank accession number AY682051; 99%
similarity). The isolate also showed a positive for NRPS genes. As not much is
known about uncultured bacteria, this shows that the mucus regulated medium
can be used to isolate previously unculturable bacteria. BCM 32 is closely matched
with Bacillus arsenicus strain HLSB44 (GenBank accession number FJ999563; 95%

P a g e | 86
similarity) and BCM 50 is closely matched with Brachybacterium
paraconglomeratum (GenBank accession number AB362255; 100% similarity). B.
arsenicus is an arsenic-resistant bacterium (Shivaji et al. 2005) which can possibly
aid the coral against heavy metal pollution. Brachybacterium paraconglomeratum
have previously been found in coral tissue (Nithyanand & Pandian 2009) and
mucus (Wilson et al. 2012).
The surface of living corals is covered by a mucoid material. This
mucopolysacchride layer provides a matrix for bacterial colonization, allowing
establishment of a ‘normal bacterial community’ (Ducklow & Mitchell 1979;
Ritchie et al. 1994). The normal bacterial flora may produce antimicrobial
compounds that help the coral avoid infection by pathogens (Jensen & Fenical
1994). Worldwide coral decline have been attributed to the increase of sea surface
temperatures, coastal degradation, pollution, diseases, ecosystem imbalance
caused by anthropogenic influences, and the synergistic effect of multiple stressors
(Harvell et al. 2002; Rosenberg & Ben-Haim 2002; Sutherland, Porter & Torres
2004). An explanation for the increased incidence of diseases in corals is its stress-
induced susceptibility to opportunistic microbes trapped within the SML. It is
acknowledged that stress conditions, particularly temperature, can cause certain
bacteria to become virulent, by ‘turning on’ virulence genes (Colwell 1996; Patz et
al. 1996). The mucus attached bacteria do not undergo mucus regulated selection
and are therefore potentially invasive under the right conditions (Ritchie 2006).
In this study, we can conclude that the antibacterial properties of mucus attached
and mucus associated bacteria have different optimum temperatures. Mucus
associated bacteria work best at 28 °C while the mucus attached bacteria has the
potential to take over at 30 °C. The attached bacteria also have more potential to
produce bioactive compounds as 27% of them have NRPS genes while 64% of
them have both PKS and NRPS genes (see Table 4.3). In contrast, the mucus
associated isolates only has PKS (11%) or NRPS genes (61%), making them unable
to form hybrids. This indicates the potential strength of the attached community to
overcome resident mucus bacteria at elevated temperatures when they turn on
their virulence genes.

P a g e | 87
It is noteworthy that cultures related to the Vibrio coralliilyticus strain we isolated
(98-99 % similarity; see Figure 4.3) is known to cause rapid tissue lysis in the
stony coral Pocillopora damicornis by metalloproteinase at elevated temperatures
(Ben-Haim et al. 2003; Ben-Haim, Zicherman-Keren & Rosenberg 2003; Rosenberg
& Falkovitz 2004). Vibrio shiloi, is the causative agent of bacterial bleaching in the
coral Occulina patagonica (Kushmaro et al. 2001). Similar to Vibrio coralliilyticus, it
produces a proline-rich peptide that inhibits photosynthesis and a protease that
lyses zooxanthellae (Ben-Haim et al. 1999; Banin, Israely, et al. 2000; Rosenberg &
Falkovitz 2004).
Two of our Vibrio coralliilyticus -related isolates (BCM 38 and 39) has PKS genes
while a third related isolate (BCM 45) has NRPS genes (see Table 4.3). These
isolated were isolated from mucus regulated media, indicating that these bacteria
are part of the mucus associated environment. Interestingly, these isolates also
showed a slight decrease in inhibition activity at 30 and 32 °C (see Figure 4.10). It
is possible that these isolates are not virulent strains and are part of the coral
defence system at optimum temperatures.
Vibrio shiloi was isolated from reef sediment (BSD 16-11) and was used as a test
organism along with the 39 coral mucus isolates. However, no inhibition zones
were observed at the three temperatures we tested on, indicating that the isolate is
resistant towards the antimicrobial properties of the SML bacteria and may pose a
problem at elevated temperatures.
Laboratory studies had previously revealed that coral bleaching occurs when
water temperature is increased roughly 1 °C above normal optimum temperatures
of 26 to 27 °C during the warmest part of the year (Goreau & Hayes 2008).
However, this was not the case for corals in the Talang-talang reef and our results
indicate that these corals – and by extension, the bacterial community in the
surrounding environment – may have a higher temperature threshold. It is
possible that a more obvious demonstration of the antimicrobial properties of
coral mucus isolates can be observed at lower and higher temperatures (i.e. ~26
and 34 °C).

P a g e | 88
This study shows that the different groups of coral mucus isolates can dominate
the SML environment at different periods depending on temperature, and that
mucus attached isolates has a high chance of turning virulent against the mucus
associated isolates and cause diseases which may lead to bleaching at elevated
temperatures.
4.3.4 Bacterial strains with potential to metabolise DMS and/or
demethylate DMSP
To our knowledge, screening of dmdA and dddP genes in coral SML bacterial
communities has not been done before. This preliminary study is part of our effort
to understand the importance of bacteria in the region for the local sulphur cycle.
Our isolates were not cultured in a method that involves specific selection for
DMSP utilisation, therefore any presence of these genes in our isolates is most
likely fundamental.
The dddP gene which is responsible for DMS production appears to be most
abundant (26%) within the coral mucus bacterial community (see Figure 4.13).
Many of our isolates also show potential in undergoing both DMSP degrading
pathways as 20% of them have both dmdA and dddP genes.
Figure 4.13: Relative abundance of dmdA and dddP genes in cultured bacterial
communities from coral mucus.
18%
26%
20%
36%
Abundance of DMSP-degrading genes in bacteria from coral mucus
dmdA dddP Both None

P a g e | 89
Figure 4.14: Presence of dmdA and/or dddP genes in bacterial isolates from coral
mucus.
The presence of DMSP degrading genes in the coral mucus bacterial groups is
similar to their occurrence in bacterial communities in the Kuching area of the
South China Sea (see Chapter 3) where their composition generally mirrored the
bacterial community. The Gammaproteobacteria group is the largest identified
fraction within the community with the potential for DMSP-assimilation, followed
by the Alphaproteobacteria and Firmicutes.
Within the coral mucus, bacteria are extremely dependent on photosynthetic
products produced by zooxanthellae which play a role in regulating microbial
communities present in corals (Ritchie & Smith 2004). Studies into coral-
associated bacteria capable of metabolizing DMSP and DMS have emerged only
recently (Raina et al. 2009, 2010). Little information is available and the nature of
their interactions with the coral host remains an important research question.
Roseobacter-related strains (BCM 37 and 56; 100% similarity) were isolated from
the coral mucus. Both isolates may play a possible role in the the biogeochemical
cycling of sulphur within the mucus as they appear to have both DMSP degrading
genes. The Roseobacter genus is potentially central to the health of corals. The
Roseobacter spp. are widely associated with corals (Frias-Lopez et al. 2002;
Rohwer et al. 2002; Bourne & Munn 2005; Kooperman et al. 2007; Bourne et al.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
α-proteobacteria γ-proteobacteria Actinobacteria Firmicutes Unknown
Pe
rce
nta
ge
(%
)
dmdA dddP Both None

P a g e | 90
2008) and suspected to be involved in a symbiotic relationship with zooxanthellae
(Raina et al. 2009). An antibiotic produced by Roseobacter, thiotropocin, is a
sulphur compound derived from DMSP metabolism (Wagner-Döbler et al. 2004).
4.4 CONCLUSION
The bacterial communities at the Talang-talang reef were different according to
the environment (coral SML, water column and reef sediment). The coral mucus
community is the most diverse with isolates playing potential roles in coral
defence, while the community from reef sediment is dominated by potentially
pathogenic Vibrio spp.. Two known coral pathogens, Vibrio coralliilyticus and Vibrio
shiloi were successfully cultured from the coral reef environment. While the corals
are healthy at the time of isolation, these opportunistic pathogens may pose a
problem at elevated temperatures.
The coral mucus community also showed high potential in the production of PKS
and NRPS compounds. The inhibitory results support the efficiency of PCR
screening using specific PKS and NRPS primers, whereby PKS and/or NRPS strains
exhibit substantial inhibition activity. Antimicrobial activities of mucus associated
bacteria decrease as temperature increase while mucus attached bacteria are most
effective at 30 °C. This study also confirms the coral mucus as a regulating media
capable of choosing associated communities exhibiting antibacterial properties
under optimum conditions.
The preliminary study on the potential role of coral SML bacterial communities in
the local sulphur cycle revealed that the presence of DMSP degrading genes in the
coral mucus bacterial groups mirrors the general bacterial community where the
majority of gene abundance are within the Gammaproteobacteria, indicating a
major role for the group. The majority of the SML isolates were observed to have
both dmdA and/or dddP genes, showing potential in undergoing both DMSP
degrading pathways depending on DMSP availability. Members of the Roseobacter
genus which is widely associated with corals and DMSP degrading capabilities
were successfully isolated from the coral SML, indicating possible roles (such as?)
in the biogeochemical cycling of sulphur within the mucus.

P a g e | 91
Further in-depth characterization of these communities through a combination of
physical, chemical and molecular biological studies is however needed to improve
our understanding of the role of bacteria in coral defence and especially in DMS(P)
cycling.
4.5 ACKNOWLEDGEMENTS
The authors would like to thank the Sarawak Forestry Department for their kind
permission to conduct research at the Talang-Satang National Park (Permit No.
NCCD.907.4.4 (Jld.VI)-104 and Park Permit No. 54/2011). Kuek FWI is funded by
the Sarawak Foundation’s Tunku Abdul Rahman scholarship. The research leading
to these results has received funding from the European Union's Seventh
Framework Programme FP7/2007-2013 under grant agreement no. 226224 -
SHIVA.

P a g e | 92
CHAPTER 5
Summary and Future Work
This study has presented (i) an overview of culturable bacterial communities in
waters of the South China Sea, Celebes Sea and a coral reef environment (Talang-
talang reef), (ii) the potential roles of these communities in the marine DMS(P)
cycle and (iii) the antimicrobial properties of cultured isolates from coral SML at
elevated temperatures and their potential role in coral defence.
The bacterial communities in the waters of Kuching and Kota Kinabalu (within the
South China Sea) and Semporna (within the Celebes Sea) are almost entirely
unknown and have not been sampled by either culture or culture-independent
techniques. Members of the Alphaproteobacteria, Gammaproteobacteria and
Firmicutes were successfully cultured from all three sampling locations while
isolates from Betaproteobacteria were only found in Semporna. Differences in
bacterial communities between the three areas can partly be explained by
differences in physico-chemical parameters. Kuching is dominated by potentially
pathogenic Vibrio spp. possibly due to higher nutrients and riverine input at the
sampling locations, while the community at Kota Kinabalu is more indicative of an
open ocean environment. Bacterial communities from Kota Kinabalu and
Semporna also show potential roles in bioremediation, nitrogen fixing and
sulphate reduction.
The bacterial communities at the Talang-talang reef also show variations between
environments (coral SML, water column and reef sediment). The isolated
community from coral mucus is the most diverse of the three, with members from
Actinobacteria, Alphaproteobacteria, Gammaproteobacteria and Firmicutes.
Isolates from the SML also indicate potential roles in coral defence with strains
related to antibiotic producers with tolerance towards elevated temperatures and
heavy metal contamination, while the community from reef sediment is dominated
by potentially pathogenic Vibrio spp..

P a g e | 93
Isolates from the SML isolates also displayed a high potential in the production of
PKS and NRPS compounds. Strains that contained PKS and/or NRPS genes did
exhibit substantial inhibition activity in the well diffusion assay. Antimicrobial
properties of mucus associated bacteria were observed to decrease as temperature
increase while mucus attached bacteria were most effective at 30 °C. This is an
indication that different groups of coral mucus bacteria can dominate the SML
environment at different periods depending on temperature, and that
opportunistic pathogens can cause diseases which may lead to bleaching at
elevated temperatures.
Two known coral pathogens, Vibrio coralliilyticus and Vibrio shiloi were
successfully cultured from the coral reef environment, the latter showing
resistance against the antimicrobial properties of the mucus associated bacterial
community. While the corals are healthy at the time of isolation, these
opportunistic pathogens may pose a problem at elevated temperatures.
In both open water and coral reef environments studied, the cultured bacterial
communities displayed an abundance of DMSP degrading genes. Communities in
this study have either dmdA or dddP or both genes when screened, showing high
adaptability in DMS(P) utilisation which we believe is influenced by bacterial
carbon and sulphur demands and by DMSP availability.
5.1 Future research
Culturing and isolation of bacteria is necessary for detailed studies of physiology
and ecological function. Culture-based methods used in this study enables us to
further biochemically classify and analyse the bacterial portion of marine
environment. Further in-depth characterization of these communities through a
combination of physical, chemical and molecular biological studies is needed and
will improve our understanding of the role of bacteria in DMS(P), coral defence
and their impacts on climate change. Initial clone library from Kuching and Kota
Kinabalu showed that culture-independent and cultured bacterial communities are
very different, so further molecular-based studies are essential for a more
complete assessment of their diversity.

P a g e | 94
The use of an assortment of media types and growth condition variables can aid in
increasing the diversity of microorganisms recovered by culturing and discovery of
other specific properties fundamental to the species. Studies by Vila-Costa et al.
2010 have successfully utilised DMSP enriched media to select for bacteria capable
of degrading DMSP into DMS from the natural environment. The approach used in
this study did uncover the existence of dmdA and dddP genes in species that were
previously involved in DMSP degradation (i.e. Alcaligenes faecalis), confirming
their potential role in our waters. However, our understanding of the role of the
genes in the various isolates (i.e. gene activity, conditions for ‘bacterial switch’) is
limited and further studies are needed to reveal their role in the sulphur cycle.
Partial sequencing of the 16S gene is insufficient for a thorough identification of
the bacterial isolates; therefore these isolates will require further genetic
delineation using gene specific primers.
After final identification it would also be of interest to see if the isolates that are
related to Vibrio coralliilyticus and Vibrio shiloi actually do cause diseases on
corals; if the disease symptoms differ or even why the corals in our reef are healthy
despite enhanced temperatures and existence of potentially pathogenic strains.
Furthermore, some of the isolates that have displayed enhanced antibiotic activity
at higher temperatures could be tested on corals and see if they develop diseases.

P a g e | 95
References
Adam, A, Mohammad-Noor, N, Anton, A, Saleh, E, Saad, S & Muhd Shaleh, SR 2011, “Temporal and spatial distribution of harmful algal bloom (HAB) species in coastal waters of Kota Kinabalu, Sabah, Malaysia,” Harmful Algae, vol. 10, no. 5, pp. 495–502.
Aguirre, AA, Balazs, GH, Zimmerman, B & Spraker, TR 1994, “Evaluation of Hawaiian green turtles (Chelonia mydas) for potential pathogens associated with fibropapillomas,” Journal of Wildlife Diseases, vol. 30, no. 1, pp. 8–15.
Van Alstyne, K, Schupp, P & Slattery, M 2006, “The distribution of dimethylsulfoniopropionate in tropical Pacific coral reef invertebrates,” Coral Reefs, vol. 25, no. 3, Springer-Verlag, pp. 321–327.
Andreae, MO 1986, “The Ocean as a Source of Atmospheric Sulfur Compounds,” in P Buat-Ménard (ed.), The Role of Air-Sea Exchange in Geochemical Cycling SE - 14, Springer Netherlands, pp. 331–362.
Andreae, MO & Crutzen, PJ 1997, “Atmospheric Aerosols: Biogeochemical Sources and Role in Atmospheric Chemistry,” Science, vol. 276, no. 5315, pp. 1052–1058.
Andreae, MO & Raemdonck, H 1983, “Dimethyl Sulfide in the Surface Ocean and the Marine Atmosphere: A Global View,” Science, vol. 221, no. 4612, pp. 744–747.
Ansari, MZ, Yadav, G, Gokhale, RS & Mohanty, D 2004, “NRPS-PKS: a knowledge-based resource for analysis of NRPS/PKS megasynthases,” Nucleic Acids Research, vol. 32, no. suppl 2, pp. W405–W413.
Ansede, JH, Friedman, R & Yoch, DC 2001, “Phylogenetic Analysis of Culturable Dimethyl Sulfide-Producing Bacteria from a Spartina-Dominated Salt Marsh and Estuarine Water,” Applied and Environmental Microbiology, vol. 67, no. 3, pp. 1210–1217.
Ansede, JH, Pellechia, PJ & Yoch, DC 1999, “Metabolism of Acrylate to β-Hydroxypropionate and Its Role in Dimethylsulfoniopropionate Lyase Induction by a Salt Marsh Sediment Bacterium, Alcaligenes faecalis M3A,” Applied and Environmental Microbiology, vol. 65, no. 11, pp. 5075–5081.
Anzai, Y, Kim, H, Park, JY, Wakabayashi, H & Oyaizu, H 2000, “Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence.,” International Journal of Systematic and Evolutionary Microbiology, vol. 50, no. 4, pp. 1563–1589.
Archer, SD, Ragni, M, Webster, R, Airs, RL & Geider, RJ 2010, “Dimethyl sulfoniopropionate and dimethyl sulfide production in response to

P a g e | 96
photoinhibition in Emiliania huxleyi,” Limnology and oceanography, vol. 55, no. 4, American Society of Limnology and Oceanography, pp. 1579–1589.
Arnon, S, Ronen, Z, Adar, E, Yakirevich, A & Nativ, R 2005, “Two-dimensional distribution of microbial activity and flow patterns within naturally fractured chalk,” Journal of Contaminant Hydrology, vol. 79, no. 3–4, pp. 165–186.
Arrigo, KR 2005, “Marine microorganisms and global nutrient cycles,” Nature, vol. 437, no. 7057, pp. 349–355.
Awang, D, Moshidi, MZ & Muda, AA 2003, “Living coral reef resources of Sarawak, with special reference Kuching area,” fri.gov.my, pp. 1–13.
Ayuso-Sacido, a & Genilloud, O 2005, “New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: detection and distribution of these biosynthetic gene sequences in major taxonomic groups.,” Microbial ecology, vol. 49, no. 1, pp. 10–24.
Baker, AC, Glynn, PW & Riegl, B 2008, “Climate change and coral reef bleaching: An ecological assessment of long-term impacts, recovery trends and future outlook,” Estuarine, Coastal and Shelf Science, vol. 80, no. 4, Elsevier Ltd, pp. 435–471.
Banin, E, Ben-Haim, Y, Israely, T, Loya, Y & Rosenberg, E 2000, “Effect of the Environment on the Bacterial Bleaching of Corals,” Water, Air, and Soil Pollution, vol. 123, no. 1-4, Kluwer Academic Publishers, pp. 337–352.
Banin, E, Israely, T, Kushmaro, A, Loya, Y, Orr, E & Rosenberg, E 2000, “Penetration of the Coral-Bleaching Bacterium Vibrio shiloi into Oculina patagonica,” Applied and Environmental Microbiology, vol. 66, no. 7, pp. 3031–3036.
Banin, E, Vassilakos, D, Orr, E, Martinez, RJ & Rosenberg, E 2003, “Superoxide dismutase is a virulence factor produced by the coral bleaching pathogen Vibrio shiloi.,” Current microbiology, vol. 46, no. 6, pp. 418–22.
Barneah, O, Ben-Dov, E, Kramarsky-Winter, E & Kushmaro, A 2007, “Characterization of black band disease in Red Sea stony corals.,” Environmental microbiology, vol. 9, no. 8, pp. 1995–2006.
Baticados, MCL, Lavilla-Pitogo, CR, Cruz-Lacierda, ER, de la Peña, LD & Suñaz, NA 1990, “Studies on the chemical control of luminous bacteria Vibrio harveyi and V splendidus solated from diseased Penaeus monodon arvae and rearing water.,” Diseases of Aquatic Organisms, vol. 9, no. 2, Inter Research, pp. 133–139.
Bell, LC, Richardson, DJ & Ferguson, SJ 1992, “Identification of nitric oxide reductase activity in Rhodobacter capsulatus: the electron transport pathway can either use or bypass both cytochrome c2 and the cytochrome bc1 complex,” Journal of General Microbiology, vol. 138, no. 3, pp. 437–443.

P a g e | 97
Ben-Haim, Y, Banim, E, Kushmaro, a, Loya, Y & Rosenberg, E 1999, “Inhibition of photosynthesis and bleaching of zooxanthellae by the coral pathogen Vibrio shiloi.,” Environmental microbiology, vol. 1, no. 3, pp. 223–9.
Ben-Haim, Y & Rosenberg, E 2002, “A novel Vibrio sp. pathogen of the coral Pocillopora damicornis,” Marine Biology, vol. 141, no. 1, Springer-Verlag, pp. 47–55.
Ben-Haim, Y, Thompson, FL, Thompson, CC, Cnockaert, MC, Hoste, B, Swings, J & Rosenberg, E 2003, “Vibrio coralliilyticus sp. nov., a temperature-dependent pathogen of the coral Pocillopora damicornis,” International Journal of Systematic and Evolutionary Microbiology, vol. 53, no. 1, pp. 309–315.
Ben-Haim, Y, Zicherman-Keren, M & Rosenberg, E 2003, “Temperature-Regulated Bleaching and Lysis of the Coral Pocillopora damicornis by the Novel Pathogen Vibrio coralliilyticus,” Applied and Environmental Microbiology, vol. 69, no. 7, pp. 4236–4242.
Bernard, L, Schäfer, H, Joux, F, Courties, C, Muyzer, G & Lebaron, P 2000, “Genetic diversity of total, active and culturable marine bacteria in coastal seawater,” Aquatic Microbial Ecology, vol. 23, no. 1, pp. 1–11.
Bidle, KD & Fletcher, M 1995, “Comparison of free-living and particle-associated bacterial communities in the chesapeake bay by stable low-molecular-weight RNA analysis.,” Applied and Environmental Microbiology, vol. 61, no. 3, pp. 944–952.
Birnboim, HC & Doly, J 1979, “A rapid alkaline extraction procedure for screening recombinant plasmid DNA,” Nucleic Acids Research, vol. 7, no. 6, pp. 1513–1523.
Blackwood, CB, Hudleston, D, Zak, DR & Buyer, JS 2007, “Interpreting Ecological Diversity Indices Applied to Terminal Restriction Fragment Length Polymorphism Data: Insights from Simulated Microbial Communities,” Applied and Environmental Microbiology, vol. 73, no. 16, pp. 5276–5283.
Bourne, D, Iida, Y, Uthicke, S & Smith-Keune, C 2008, “Changes in coral-associated microbial communities during a bleaching event.,” The ISME journal, vol. 2, no. 4, pp. 350–63.
Bourne, DG, Garren, M, Work, TM, Rosenberg, E, Smith, GW & Harvell, CD 2009, “Microbial disease and the coral holobiont.,” Trends in microbiology, vol. 17, no. 12, pp. 554–62.
Bourne, DG & Munn, CB 2005, “Diversity of bacteria associated with the coral Pocillopora damicornis from the Great Barrier Reef,” Environmental Microbiology, vol. 7, no. 8, Blackwell Science Ltd, pp. 1162–1174.

P a g e | 98
Boyle, VJ, Fancher, ME & Ross, RW 1973, “Rapid, Modified Kirby-Bauer Susceptibility Test with Single, High-Concentration Antimicrobial Disks,” Antimicrobial Agents and Chemotherapy, vol. 3, no. 3, pp. 418–424.
Brink, A, van Straten, A & van Rensburg, A 1995, “Shewanella (Pseudomonas) putrefaciens Bacteremia,” Clinical Infectious Diseases, vol. 20, no. 5, pp. 1327–1332.
Broadbent, AD & Jones, GB 2004, “DMS and DMSP in mucus ropes, coral mucus, surface films and sediment pore waters from coral reefs in the Great Barrier Reef,” Marine and Freshwater Research, pp. 849–855.
Broadbent, AD, Jones, GB & Jones, RJ 2002, “DMSP in Corals and Benthic Algae from the Great Barrier Reef,” Estuarine, Coastal and Shelf Science, vol. 55, no. 4, pp. 547–555.
Broadbent, P, Baker, KF & Waterworth, Y 1977, “Effect of Bacillus spp. on increased growth of seedlings in steamed and non-treated soil,” Phytopathology, vol. 67, pp. 1027–1034.
Buddemeier, RW, Kleypas, JA & Aronson, RB 2004, Coral reefs & global climate change: potential contributions of climate change to stresses on coral reef ecosystems, Pew Center on Global Climate Change.
Button, DK, Schut, F, Quang, P, Martin, R & Robertson, BR 1993, “Viability and Isolation of Marine Bacteria by Dilution Culture: Theory, Procedures, and Initial Results,” Applied and Environmental Microbiology, vol. 59, no. 3, pp. 881–891.
Cabral, R, Cruz-Trinidad, A, Geronimo, R, Napitupulu, L, Lokani, P, Boso, D, Casal, CM, Ahmad Fatan, N & Aliño, P 2013, “Crisis sentinel indicators: Averting a potential meltdown in the Coral Triangle,” Marine Policy, vol. 39, no. 0, pp. 241–247.
Cane, DE 1997, “Introduction: Polyketide and nonribosomal polypeptide biosynthesis. From collie to coli,” Chemical Reviews, vol. 97, no. 7, ACS Publications, pp. 2463–2464.
Cervino, JM, Thompson, FL, Gomez-Gil, B, Lorence, E a, Goreau, TJ, Hayes, RL, Winiarski-Cervino, KB, Smith, GW, Hughen, K & Bartels, E 2008, “The Vibrio core group induces yellow band disease in Caribbean and Indo-Pacific reef-building corals.,” Journal of applied microbiology, vol. 105, no. 5, pp. 1658–71.
Chang, JC, Ossoff, SF, Lobe, DC, Dorfman, MH, Dumais, CM, Qualls, RG & Johnson, JD 1985, “UV inactivation of pathogenic and indicator microorganisms.,” Applied and Environmental Microbiology, vol. 49, no. 6, pp. 1361–1365.

P a g e | 99
Charlson, RJ, Lovelock, JE, Andreae, MO & Warren, SG 1987, “Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate,” Nature, vol. 326, no. 6114, pp. 655–661.
Collins, MD, Martinez-Murcia, AJ & Cai, J 1993, “Aeromonas enteropelogenes and Aeromonas ichthiosmia Are Identical to Aeromonas trota and Aeromonas veronii, Respectively, as Revealed by Small-Subunit rRNA Sequence Analysis,” International Journal of Systematic Bacteriology, vol. 43, no. 4, pp. 855–856.
Colwell, RR 1996, “Global Climate and Infectious Disease: The Cholera Paradigm*,” Science, vol. 274, no. 5295, pp. 2025–2031.
Colwell, RR, Macdonell, MT & De Ley, J 1986, “Proposal to Recognize the Family Aeromonadaceae fam. nov.,” International Journal of Systematic Bacteriology, vol. 36, no. 3, pp. 473–477.
Corbett, JJ, Fischbeck, PS & Pandis, SN 1999, “Global nitrogen and sulfur inventories for oceangoing ships,” Journal of Geophysical Research: Atmospheres, vol. 104, no. D3, pp. 3457–3470.
Crump, BC, Armbrust, EV & Baross, JA 1999, “Phylogenetic Analysis of Particle-Attached and Free-Living Bacterial Communities in the Columbia River, Its Estuary, and the Adjacent Coastal Ocean,” Applied and Environmental Microbiology, vol. 65, no. 7, pp. 3192–3204.
Curson, ARJ, Fowler, EK, Dickens, S, Johnston, AWB & Todd, JD 2011, “Multiple DMSP lyases in the γ-proteobacterium Oceanimonas doudoroffii,” Biogeochemistry, vol. 110, no. 1-3, pp. 109–119.
Curson, ARJ, Rogers, R, Todd, JD, Brearley, CA & Johnston, AWB 2008, “Molecular genetic analysis of a dimethylsulfoniopropionate lyase that liberates the climate-changing gas dimethylsulfide in several marine alpha-proteobacteria and Rhodobacter sphaeroides.,” Environmental microbiology, vol. 10, no. 3, pp. 757–67.
Curson, ARJ, Sullivan, MJ, Todd, JD & Johnston, AWB 2011, “DddY, a periplasmic dimethylsulfoniopropionate lyase found in taxonomically diverse species of Proteobacteria,” ISME J, vol. 5, no. 7, International Society for Microbial Ecology, pp. 1191–1200.
DeLong, EF 2009, “The microbial ocean from genomes to biomes,” Nature, vol. 459, no. 7244, Nature Publishing Group, pp. 200–206.
Donadio, S, Monciardini, P & Sosio, M 2007, “Polyketide synthases and nonribosomal peptide synthetases: the emerging view from bacterial genomics.,” Natural product reports, vol. 24, no. 5, pp. 1073–109.

P a g e | 100
Du, L, Sánchez, C & Shen, B 2001, “Hybrid Peptide–Polyketide Natural Products: Biosynthesis and Prospects toward Engineering Novel Molecules,” Metabolic Engineering, vol. 3, no. 1, pp. 78–95.
La Duc, MT, Dekas, A, Osman, S, Moissl, C, Newcombe, D & Venkateswaran, K 2007, “Isolation and Characterization of Bacteria Capable of Tolerating the Extreme Conditions of Clean Room Environments,” Applied and Environmental Microbiology, vol. 73, no. 8, pp. 2600–2611.
Ducklow, H & Mitchell, R 1979, “Composition of mucus released by coral reef coelenterates,” Limnol. Oceanogr, vol. 24, no. 4, pp. 706–714.
Dugdale, RC, Wilkerson, FP & Minas, HJ 1995, “The role of a silicate pump in driving new production,” Deep Sea Research Part I: Oceanographic Research Papers, vol. 42, no. 5, pp. 697–719.
Van Duyl, FC, Gieskes, WWC, Kop, AJ & Lewis, WE 1998, “Biological control of short-term variations in the concentration of DMSP and DMS during a Phaeocystis spring bloom,” Journal of Sea Research, vol. 40, no. 3–4, pp. 221–231.
Eden, PA, Schmidt, TM, Blakemore, RP & Pace, NR 1991, “Phylogenetic Analysis of Aquaspirillum magnetotacticum Using Polymerase Chain Reaction-Amplified 16S rRNA-Specific DNA,” International Journal of Systematic Bacteriology, vol. 41, no. 2, pp. 324–325.
Eghtesadi-Araghi, P 2011, “Coral reefs in the Persian Gulf and Oman Sea: an integrated perspective on some important stressors,” J Fish Aquat Sci, vol. 6, pp. 48–56.
Eilers, H, Pernthaler, J, Glöckner, FO & Amann, R 2000, “Culturability and In Situ Abundance of Pelagic Bacteria from the North Sea,” Applied and Environmental Microbiology, vol. 66, no. 7, pp. 3044–3051.
Feltham, RKA, Power, AK, Pell, PA & Sneath, PHA 1978, “A Simple Method for Storage of Bacteria at — 76°C,” Journal of Applied Microbiology, vol. 44, no. 2, Blackwell Publishing Ltd, pp. 313–316.
Fiedler, H-P, Bruntner, C, Bull, AT, Ward, A, Goodfellow, M, Potterat, O, Puder, C & Mihm, G 2005, “Marine actinomycetes as a source of novel secondary metabolites,” Antonie van Leeuwenhoek, vol. 87, no. 1, Kluwer Academic Publishers, pp. 37–42.
Fierer, N & Jackson, RB 2006, “The diversity and biogeography of soil bacterial communities,” Proceedings of the National Academy of Sciences of the United States of America, vol. 103, no. 3, pp. 626–631.
Figueras, MJ, Guarro, J & Martínez-Murcia, A 2000, “Clinically Relevant Aeromonas Species,” Clinical Infectious Diseases, vol. 30, no. 6, pp. 988–989.

P a g e | 101
Fitt, W, Brown, B, Warner, M & Dunne, R 2001, “Coral bleaching: interpretation of thermal tolerance limits and thermal thresholds in tropical corals,” Coral Reefs, vol. 20, no. 1, Springer-Verlag, pp. 51–65.
Fogelqvist, E 1991, “Dimethylsulphide (DMS) in the Weddell Sea surface and bottom water,” Marine Chemistry, vol. 35, no. 1–4, pp. 169–177.
Frias-Lopez, J, Zerkle, AL, Bonheyo, GT & Fouke, BW 2002, “Partitioning of Bacterial Communities between Seawater and Healthy, Black Band Diseased, and Dead Coral Surfaces,” Applied and Environmental Microbiology, vol. 68, no. 5, pp. 2214–2228.
Fujii, I, Watanabe, A, Sankawa, U & Ebizuka, Y 2001, “Identification of Claisen cyclase domain in fungal polyketide synthase WA, a naphthopyrone synthase of Aspergillus nidulans,” Chemistry & Biology, vol. 8, no. 2, pp. 189–197.
Gamito, S 2010, “Caution is needed when applying Margalef diversity index,” Ecological Indicators, vol. 10, no. 2, pp. 550–551.
Gomare, S, Jadhav, J & Govindwar, S 2008, “Degradation of sulfonated azo dyes by the purified lignin peroxidase from Brevibacillus laterosporus MTCC 2298,” Biotechnology and Bioprocess Engineering, vol. 13, no. 2, The Korean Society for Biotechnology and Bioengineering, pp. 136–143.
González, JM, Covert, JS, Whitman, WB, Henriksen, JR, Mayer, F, Scharf, B, Schmitt, R, Buchan, A, Fuhrman, JA, Kiene, RP & Moran, MA 2003, “Silicibacter pomeroyi sp. nov. and Roseovarius nubinhibens sp. nov., dimethylsulfoniopropionate-demethylating bacteria from marine environments,” International Journal of Systematic and Evolutionary Microbiology, vol. 53, no. 5, pp. 1261–1269.
González, JM, Kiene, RP & Moran, MA 1999, “Transformation of Sulfur Compounds by an Abundant Lineage of Marine Bacteria in the α-Subclass of the ClassProteobacteria,” Applied and Environmental Microbiology, vol. 65, no. 9, pp. 3810–3819.
González, JM, Simó, R, Massana, R, Covert, JS, Casamayor, EO, Pedrós-Alió, C & Moran, MA 2000, “Bacterial Community Structure Associated with a Dimethylsulfoniopropionate-Producing North Atlantic Algal Bloom,” Applied and Environmental Microbiology, vol. 66, no. 10, pp. 4237–4246.
Goreau, TJ & Hayes, RL 2008, “Effects of rising seawater temperature on coral reefs,” Encyclopedia of Life Support Systems (EOLSS), Eolss Publishers, Oxford, UK.
Grover, JP 2000, “Resource competition and community structure in aquatic micro-organisms: experimental studies of algae and bacteria along a gradient of organic carbon to inorganic phosphorus supply,” Journal of Plankton Research, vol. 22, no. 8, pp. 1591–1610.

P a g e | 102
Harborne, A, Fenner, D, Barnes, A, Beger, M, Harding, S & Roxburgh, T 2000, “Status report on the coral reefs of the east coast of Peninsula Malaysia,” Report Prepared to Department of Fisheries Malaysia, Kuala Lumpur, Malaysia.
Harvell, CD, Mitchell, CE, Ward, JR, Altizer, S, Dobson, AP, Ostfeld, RS & Samuel, MD 2002, “Climate Warming and Disease Risks for Terrestrial and Marine Biota,” Science, vol. 296, no. 5576, pp. 2158–2162.
Heath, RJ & Rock, CO 2002, “The Claisen condensation in biology,” Natural Product Reports, vol. 19, no. 5, The Royal Society of Chemistry, pp. 581–596.
Hedlund, BP & Staley, JT 2001, “Vibrio cyclotrophicus sp. nov., a polycyclic aromatic hydrocarbon (PAH)-degrading marine bacterium.,” International Journal of Systematic and Evolutionary Microbiology, vol. 51, no. 1, pp. 61–66.
Hill, RW, Dacey, JWH & Krupp, DA 1995, “Dimethylsulfoniopropionate in Reef Corals,” Bulletin of Marine Science, pp. 489–494.
Hill, RW, White, BA & Cottrell, MT 1998, “Virus-mediated total release of dimethylsulfoniopropionate from marine phytoplankton: a potential climate process,” Aquatic Microbial Ecology, vol. 14, no. 1, pp. 1–6.
Hill, TCJ, Walsh, KA, Harris, JA & Moffett, BF 2003, “Using ecological diversity measures with bacterial communities,” FEMS Microbiology Ecology, vol. 43, no. 1, Blackwell Publishing Ltd, pp. 1–11.
Hjelm, M, Bergh, Ø, Riaza, A, Nielsen, J, Melchiorsen, J, Jensen, S, Duncan, H, Ahrens, P, Birkbeck, H & Gram, L 2004, “Selection and Identification of Autochthonous Potential Probiotic Bacteria from Turbot Larvae (Scophthalmus maximus) Rearing Units,” Systematic and Applied Microbiology, vol. 27, no. 3, pp. 360–371.
Hoegh-Guldberg, O 1999, “Climate change, coral bleaching and the future of the world’s coral reefs,” Marine and freshwater research.
Hoegh-Guldberg, O 2004, “Coral Reefs and Projections of Future Change,” in E Rosenberg & Y Loya (eds), Coral Health and Disease SE - 26, Springer Berlin Heidelberg, pp. 463–484.
Howard, EC, Sun, S, Biers, EJ & Moran, MA 2008, “Abundant and diverse bacteria involved in DMSP degradation in marine surface waters.,” Environmental microbiology, vol. 10, no. 9, pp. 2397–410.
Huppert, A & Stone, L 1998, “Chaos in the Pacific’s coral reef bleaching cycle,” The American Naturalist, vol. 152, no. 3, pp. 447–459.
Iijima, S, Washio, K, Okahara, R & Morikawa, M 2009, “Biofilm formation and proteolytic activities of Pseudoalteromonas bacteria that were isolated from

P a g e | 103
fish farm sediments,” Microbial Biotechnology, vol. 2, no. 3, Blackwell Publishing Ltd, pp. 361–369.
Itoh, M, Matsuura, K & Satoh, T 1989, “Involvement of cytochrome bc1 complex in the electron transfer pathway for N2O reduction in a photodenitrifier, Rhodobacter sphaeroides f. s. denitrificans,” FEBS Letters, vol. 251, no. 1–2, pp. 104–108.
Ivanova, EP, Flavier, S & Christen, R 2004, “Phylogenetic relationships among marine Alteromonas-like proteobacteria: emended description of the family Alteromonadaceae and proposal of Pseudoalteromonadaceae fam. nov., Colwelliaceae fam. nov., Shewanellaceae fam. nov., Moritellaceae fam. nov., Ferri,” International Journal of Systematic and Evolutionary Microbiology, vol. 54, no. 5, pp. 1773–1788.
Ivanova, EP, Onyshchenko, OM, Christen, R, Zhukova, N V, Lysenko, AM, Shevchenko, LS, Buljan, V, Hambly, B & Kiprianova, EA 2005, “Oceanimonas smirnovii sp. nov., a novel organism isolated from the Black Sea,” Systematic and Applied Microbiology, vol. 28, no. 2, pp. 131–136.
Ivanova, EP, Shevchenko, LS, Sawabe, T, Lysenko, AM, Svetashev, VI, Gorshkova, NM, Satomi, M, Christen, R & Mikhailov, V V 2002, “Pseudoalteromonas maricaloris sp. nov., isolated from an Australian sponge, and reclassification of [Pseudoalteromonas aurantia] NCIMB 2033 as Pseudoalteromonas flavipulchra sp. nov.,” International Journal of Systematic and Evolutionary Microbiology, vol. 52, no. 1, pp. 263–271.
Jensen, PR & Fenical, W 1994, “Strategies for the discovery of secondary metabolites from marine bacteria: ecological perspectives.,” Annual review of microbiology, vol. 48, Annual Reviews 4139 El Camino Way, P.O. Box 10139, Palo Alto, CA 94303-0139, USA, pp. 559–84.
Jensen, PR, Mincer, TJ, Williams, PG & Fenical, W 2005, “Marine actinomycete diversity and natural product discovery,” Antonie van Leeuwenhoek, vol. 87, no. 1, Kluwer Academic Publishers, pp. 43–48.
Jiang, H, Dong, H, Ji, S, Ye, Y & Wu, N 2007, “Microbial Diversity in the Deep Marine Sediments from the Qiongdongnan Basin in South China Sea,” Geomicrobiology Journal, vol. 24, no. 6, Taylor & Francis, pp. 505–517.
Johnston, AWB, Todd, JD, Sun, L, Nikolaidou-Katsaridou, MN, Curson, ARJ & Rogers, R 2008, “Molecular diversity of bacterial production of the climate-changing gas, dimethyl sulphide, a molecule that impinges on local and global symbioses.,” Journal of experimental botany, vol. 59, no. 5, pp. 1059–67.
Jokiel, PL & Brown, EK 2004, “Global warming, regional trends and inshore environmental conditions influence coral bleaching in Hawaii,” Global Change Biology, vol. 10, no. 10, Blackwell Science Ltd, pp. 1627–1641.

P a g e | 104
Jones, RJ, Hoegh-Guldberg, O, Larkum, AWD & Schreiber, U 1998, “Temperature-induced bleaching of corals begins with impairment of the CO2 fixation mechanism in zooxanthellae,” Plant, Cell & Environment, vol. 21, no. 12, Blackwell Publishing Ltd, pp. 1219–1230.
Jordan, DC, McNicol, PJ & Marshall, MR 1978, “Biological nitrogen fixation in the terrestrial environment of a high Arctic ecosystem (Truelove Lowland, Devon Island, N.W.T.),” Canadian Journal of Microbiology, vol. 24, no. 6, NRC Research Press, pp. 643–649.
Jørgensen, B 2006, “Bacteria and Marine Biogeochemistry,” in H Schulz & M Zabel (eds), Marine Geochemistry SE - 5, Springer Berlin Heidelberg, pp. 169–206.
Juíz-Río, S, Osorio, CR, de Lorenzo, V & Lemos, ML 2005, “Subtractive hybridization reveals a high genetic diversity in the fish pathogen Photobacterium damselae subsp. piscicida: evidence of a SXT-like element,” Microbiology, vol. 151, no. 8, pp. 2659–2669.
Kajimura, Y 1995, “Bacillopeptins, new cyclic lipopeptide antibiotics from Bacillus subtilis FR-2,” J. Antibiotics, vol. 48, pp. 1095–1103.
Karl, DM 2002, “Nutrient dynamics in the deep blue sea,” Trends in Microbiology, vol. 10, no. 9, pp. 410–418.
Kiene, RP 1996a, “Production of methanethiol from dimethylsulfoniopropionate in marine surface waters,” Marine Chemistry, vol. 54, no. 1–2, pp. 69–83.
Kiene, RP 1996b, “Turnover of Dissolved DMSP in Estuarine and Shelf Waters of the Northern Gulf of Mexico,” in R Kiene, P Visscher, M Keller & G Kirst (eds), Biological and Environmental Chemistry of DMSP and Related Sulfonium Compounds SE - 29, Springer US, pp. 337–349.
Kiene, RP, Linn, LJ & Bruton, J a. 2000, “New and important roles for DMSP in marine microbial communities,” Journal of Sea Research, vol. 43, no. 3-4, pp. 209–224.
Kim, D, Baik, KS, Kim, MS, Jung, B-M, Shin, T-S, Chung, G-H, Rhee, MS & Seong, CN 2007, “Shewanella haliotis sp. nov., isolated from the gut microflora of abalone, Haliotis discus hannai,” International Journal of Systematic and Evolutionary Microbiology, vol. 57, no. 12, pp. 2926–2931.
Kim, S-J & Kwon, KK 2010, “Marine, Hydrocarbon-Degrading Alphaproteobacteria,” in K Timmis (ed.), Handbook of Hydrocarbon and Lipid Microbiology SE - 120, Springer Berlin Heidelberg, pp. 1707–1714.
Kirchmann, DL 2000, “Microbial ecology of the oceans,” Wiley Series in Ecological and Applied Microbiology.

P a g e | 105
Kirkwood, M, Le Brun, NE, Todd, JD & Johnston, AWB 2010, “The dddP gene of Roseovarius nubinhibens encodes a novel lyase that cleaves dimethylsulfoniopropionate into acrylate plus dimethyl sulfide,” Microbiology, vol. 156, no. 6, pp. 1900–1906.
Kirst, GO, Thiel, C, Wolff, H, Nothnagel, J, Wanzek, M & Ulmke, R 1991, “Dimethylsulfoniopropionate (DMSP) in icealgae and its possible biological role,” Marine Chemistry, vol. 35, no. 1–4, pp. 381–388.
Kleinkauf, H & Von Döhren, H 1996, “A Nonribosomal System of Peptide Biosynthesis,” European Journal of Biochemistry, vol. 236, no. 2, Blackwell Science Ltd, pp. 335–351.
Koh, EGL 1997, “Do Scleractinian Corals Engage in Chemical Warfare Against Microbes?,” Journal of Chemical Ecology, vol. 23, no. 2, pp. 379–398.
Kooperman, N, Ben-Dov, E, Kramarsky-Winter, E, Barak, Z & Kushmaro, A 2007, “Coral mucus-associated bacterial communities from natural and aquarium environments,” FEMS Microbiology Letters, vol. 276, no. 1, Blackwell Publishing Ltd, pp. 106–113.
Koren, O & Rosenberg, E 2006, “Bacteria associated with mucus and tissues of the coral Oculina patagonica in summer and winter.,” Applied and environmental microbiology, vol. 72, no. 8, pp. 5254–9.
Kushmaro, A, Banin, E, Loya, Y, Stackebrandt, E & Rosenberg, E 2001, “Vibrio shiloi sp. nov., the causative agent of bleaching of the coral Oculina patagonica.,” International journal of systematic and evolutionary microbiology, vol. 51, no. Pt 4, pp. 1383–8.
Kushmaro, A, Loya, Y, Fine, M & Rosenberg, E 1996, “Bacterial infection and coral bleaching,” Nature, vol. 380, no. 6573, p. 396.
Kushmaro, A, Rosenberg, E, Fine, M & Loya, Y 1997, “Bleaching of the coral Oculina patagonica by Vibrio AK-1,” Marine ecology progress …, vol. 147, pp. 159–165.
Kusuda, R & Kawai, K 1998, “Bacterial diseases of cultured marine fish in Japan,” Fish Pathology, vol. 33, no. 4, pp. 221–227.
Labbé, N, Parent, S & Villemur, R 2004, “Nitratireductor aquibiodomus gen. nov., sp. nov., a novel α-proteobacterium from the marine denitrification system of the Montreal Biodome (Canada),” International Journal of Systematic and Evolutionary Microbiology, vol. 54, no. 1, pp. 269–273.
Lai, Q & Shao, Z 2012, “Genome Sequence of Thalassospira xiamenensis Type Strain M-5,” Journal of Bacteriology, vol. 194, no. 24, p. 6957.

P a g e | 106
Lalucat, J, Bennasar, A, Bosch, R, García-Valdés, E & Palleroni, NJ 2006, “Biology of Pseudomonas stutzeri,” Microbiology and Molecular Biology Reviews, vol. 70, no. 2, pp. 510–547.
Lamberti, GA & Resh, VH 1983, “Geothermal Effects on Stream Benthos: Separate Influences of Thermal and Chemical Components on Periphyton and Macroinvertebrates,” Canadian Journal of Fisheries and Aquatic Sciences, vol. 40, no. 11, NRC Research Press, pp. 1995–2009.
Lampert, Y, Kelman, D, Nitzan, Y, Dubinsky, Z, Behar, A & Hill, RT 2008, “Phylogenetic diversity of bacteria associated with the mucus of Red Sea corals,” FEMS Microbiology Ecology, vol. 64, no. 2, Blackwell Publishing Ltd, pp. 187–198.
Lane, DJ, Pace, B, Olsen, GJ, Stahl, DA, Sogin, ML & Pace, NR 1985, “Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses,” Proceedings of the National Academy of Sciences, vol. 82, no. 20, pp. 6955–6959.
Ledyard, K, DeLong, E & Dacey, JH 1993, “Characterization of a DMSP-degrading bacterial isolate from the Sargasso Sea,” Archives of Microbiology, vol. 160, no. 4, Springer-Verlag, pp. 312–318.
Ledyard, KM, Dacey, JWH & Dacey, JWH 1996, “Microbial cycling of DMSP and DMS in coastal and oligotrophic seawater,” Limnology and oceanography, vol. 41, no. 1, pp. 33–40.
Lee, JN & Mohamed, CAR 2009, “Trace Metal Contents in the Porites Corals of PeninsularMalaysia,” International Journal of Environmental Research, vol. 3, no. 1, pp. 85–94.
Lee, K-H & Ruby, EG 1994, “Effect of the Squid Host on the Abundance and Distribution of Symbiotic Vibrio fischeri in Nature,” Applied and Environmental Microbiology, vol. 60, no. 5, pp. 1565–1571.
Lee, M & Chandler, AC 1941, “A Study of the Nature, Growth and Control of Bacteria in Cutting Compounds,” Journal of Bacteriology, vol. 41, no. 3, pp. 373–386.
Lesser, MP, Bythell, JC, Gates, RD, Johnstone, RW & Hoegh-Guldberg, O 2007, “Are infectious diseases really killing corals? Alternative interpretations of the experimental and ecological data,” Journal of Experimental Marine Biology and Ecology, vol. 346, no. 1-2, pp. 36–44.
Levasseur, M, Gosselin, M & Michaud, S 1994, “A new source of dimethylsulfide (DMS) for the arctic atmosphere: ice diatoms,” Marine Biology, vol. 121, no. 2, Springer-Verlag, pp. 381–387.

P a g e | 107
Levine, NM, Varaljay, V a, Toole, D a, Dacey, JWH, Doney, SC & Moran, MA 2012, “Environmental, biochemical and genetic drivers of DMSP degradation and DMS production in the Sargasso Sea.,” Environmental microbiology, vol. 14, no. 5, pp. 1210–23.
Li, D, Yu, T, Zhang, Y, Yang, M, Li, Z, Liu, M & Qi, R 2010, “Antibiotic Resistance Characteristics of Environmental Bacteria from an Oxytetracycline Production Wastewater Treatment Plant and the Receiving River,” Applied and Environmental Microbiology, vol. 76, no. 11, pp. 3444–3451.
Li, Z-Y, He, L-M, Wu, J & Jiang, Q 2006, “Bacterial community diversity associated with four marine sponges from the South China Sea based on 16S rDNA-DGGE fingerprinting,” Journal of Experimental Marine Biology and Ecology, vol. 329, no. 1, pp. 75–85.
Lin, Z, Kumagai, K, Baba, K, Mekalanos, JJ & Nishibuchi, M 1993, “Vibrio parahaemolyticus has a homolog of the Vibrio cholerae toxRS operon that mediates environmentally induced regulation of the thermostable direct hemolysin gene.,” Journal of Bacteriology, vol. 175, no. 12, pp. 3844–3855.
Lipp, EK, Huq, A & Colwell, RR 2002, “Effects of global climate on infectious disease: the cholera model,” Clinical microbiology reviews, vol. 15, no. 4.
Liu, C, Wu, Y, Li, L, Ma, Y & Shao, Z 2007, “Thalassospira xiamenensis sp. nov. and Thalassospira profundimaris sp. nov.,” International Journal of Systematic and Evolutionary Microbiology, vol. 57, no. 2, pp. 316–320.
López-López, A, Pujalte, MJ, Benlloch, S, Mata-Roig, M, Rosselló-Mora, R, Garay, E & Rodríguez-Valera, F 2002, “Thalassospira lucentensis gen. nov., sp. nov., a new marine member of the alpha-Proteobacteria.,” International Journal of Systematic and Evolutionary Microbiology, vol. 52, no. 4, pp. 1277–1283.
Lovelock, JE, Maggs, RJ & Rasmussen, RA 1972, “Atmospheric Dimethyl Sulphide and the Natural Sulphur Cycle,” Nature, vol. 237, no. 5356, pp. 452–453.
Magarvey, NA, Keller, JM, Bernan, V, Dworkin, M & Sherman, DH 2004, “Isolation and Characterization of Novel Marine-Derived Actinomycete Taxa Rich in Bioactive Metabolites,” Applied and Environmental Microbiology, vol. 70, no. 12, pp. 7520–7529.
Magurran, AE 2004, Measuring biological diversity, Blackwell Publishing, London.
Malin, G & Erst, GO 1997, “Algal Production of Dimethyl Sulfide and its Atmospheric Role,” Journal of Phycology, vol. 33, no. 6, Blackwell Publishing Ltd, pp. 889–896.
Malmstrom, RR, Kiene, RP & Kirchman, DL 2004, “Identification and enumeration of bacteria assimilating dimethylsulfoniopropionate (DMSP) in the North

P a g e | 108
Atlantic and Gulf of Mexico,” Limnology and Oceanography, vol. 49, no. 2, pp. 597–606.
Marahiel, MA, Stachelhaus, T & Mootz, HD 1997, “Modular Peptide Synthetases Involved in Nonribosomal Peptide Synthesis,” Chemical Reviews, vol. 97, no. 7, American Chemical Society, pp. 2651–2674.
Margalef, DR 1958, “Information theory in ecology,” General Systems, vol. 3, Society for General Systems Research, pp. 36–71.
Maruyama, A, Honda, D, Yamamoto, H, Kitamura, K & Higashihara, T 2000, “Phylogenetic analysis of psychrophilic bacteria isolated from the Japan Trench, including a description of the deep-sea species Psychrobacter pacificensis sp. nov.,” International Journal of Systematic and Evolutionary Microbiology, vol. 50, no. 2, pp. 835–846.
Matu, EN, Kirira, PG, Kigondu, EVM, Moindi, EM & Amugune, BA 2012, “Antimicrobial activity of organic total extracts of three Kenyan medicinal plants,” African Journal of Pharmacology and Therapeutics.
Mayer, FW & Wild, C 2010, “Coral mucus release and following particle trapping contribute to rapid nutrient recycling in a Northern Red Sea fringing reef,” Marine and Freshwater Research, pp. 1006–1014.
McCarter, L 1999, “The multiple identities of Vibrio parahaemolyticus,” Journal of molecular microbiology and biotechnology, vol. 1, no. 1, p. 51.
McKnight, TL & Hess, D 2000, “Climate zones and types: the Köppen system,” Physical Geography: A Landscape Appreciation. Upper Saddle River, NJ: Prentice Hall.
McLeod, E, Moffitt, R, Timmermann, A, Salm, R, Menviel, L, Palmer, MJ, Selig, ER, Casey, KS & Bruno, JF 2010, “Warming Seas in the Coral Triangle: Coral Reef Vulnerability and Management Implications,” Coastal Management, vol. 38, no. 5, Taylor & Francis, pp. 518–539.
Meier, JL & Burkart, MD 2009, “The chemical biology of modular biosynthetic enzymes,” Chemical Society Reviews, vol. 38, no. 7, The Royal Society of Chemistry, pp. 2012–2045.
Millero, FJ, Lee, K & Roche, M 1998, “Distribution of alkalinity in the surface waters of the major oceans,” Marine Chemistry, vol. 60, no. 1–2, pp. 111–130.
Mincer, TJ, Fenical, W & Jensen, PR 2005, “Culture-Dependent and Culture-Independent Diversity within the Obligate Marine Actinomycete Genus Salinispora,” Applied and Environmental Microbiology, vol. 71, no. 11, pp. 7019–7028.

P a g e | 109
Mincer, TJ, Jensen, PR, Kauffman, CA & Fenical, W 2002, “Widespread and Persistent Populations of a Major New Marine Actinomycete Taxon in Ocean Sediments,” Applied and Environmental Microbiology, vol. 68, no. 10, pp. 5005–5011.
Moberg, F & Folke, C 1999, “Ecological goods and services of coral reef ecosystems,” Ecological Economics, vol. 29, no. 2, pp. 215–233.
Moran, M, Belas, R, Schell, M, González, J, Sun, F, Sun, S, Binder, B, Edmonds, J, Ye, W, Orcutt, B, Howard, E, Meile, C, Palefsky, W, Goesmann, A, Ren, Q, Paulsen, I, Ulrich, L, Thompson, L, Saunders, E & Buchan, A 2007, “Ecological genomics of marine Roseobacters.,” Applied and environmental microbiology, vol. 73, no. 14, pp. 4559–69.
Morton, B & Blackmore, G 2001, “South China Sea,” Marine Pollution Bulletin, vol. 42, no. 12, pp. 1236–1263.
Muhling, M, Woolven-Allen, J, Murrell, JC & Joint, I 2008, “Improved group-specific PCR primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of complex microbial communities,” ISME J, vol. 2, no. 4, International Society for Microbial Ecology, pp. 379–392.
Nagasawa, S & Terazaki, M 1987, “Bacterial epibionts of the deep-sea copepod calanus-cristatus kroyer,” Oceanologica acta, vol. 10, no. 4, Gauthier-Villars, pp. 475–479.
Ndyetabura, T, Lyantagaye, SL & Mshandete, AM 2010, “Antimicrobial activity of ethyl acetate extracts from edible tanzanian Coprinus cinereus (schaeff) s. Gray s. lat. Cultivated on grasses supplemented with cow dung manure,” Arpn. J. Agric. Biol. Sci, vol. 5, no. 5.
Neilan, BA, Dittmann, E, Rouhiainen, L, Bass, RA, Schaub, V, Sivonen, K & Börner, T 1999, “Nonribosomal Peptide Synthesis and Toxigenicity of Cyanobacteria,” Journal of Bacteriology, vol. 181, no. 13, pp. 4089–4097.
Neu, HC & Gootz, TD 1996, “Antimicrobial chemotherapy,” Medical Microbiology. 4th ed.
Ni, Y-Y, Kim, DY, Chung, MG, Lee, SH, Park, H-Y & Rhee, YH 2010, “Biosynthesis of medium-chain-length poly(3-hydroxyalkanoates) by volatile aromatic hydrocarbons-degrading Pseudomonas fulva TY16,” Bioresource Technology, vol. 101, no. 21, pp. 8485–8488.
Nishimori, E, Kita-Tsukamoto, K & Wakabayashi, H 2000, “Pseudomonas plecoglossicida sp. nov., the causative agent of bacterial haemorrhagic ascites of ayu, Plecoglossus altivelis.,” International Journal of Systematic and Evolutionary Microbiology, vol. 50, no. 1, pp. 83–89.

P a g e | 110
Nithyanand, P & Pandian, SK 2009, “Phylogenetic characterization of culturable bacterial diversity associated with the mucus and tissue of the coral Acropora digitifera from the Gulf of Mannar,” FEMS Microbiology Ecology, vol. 69, no. 3, Blackwell Publishing Ltd, pp. 384–394.
Noble, RC & Overman, SB 1994, “Pseudomonas stutzeri infection a review of hospital isolates and a review of the literature,” Diagnostic Microbiology and Infectious Disease, vol. 19, no. 1, pp. 51–56.
Nold, S & Zwart, G 1998, “Patterns and governing forces in aquatic microbial communities,” Aquatic Ecology, vol. 32, no. 1, Kluwer Academic Publishers, pp. 17–35.
Okuda, J, Nakai, T, Chang, PS, Oh, T, Nishino, T, Koitabashi, T & Nishibuchi, M 2001, “The toxR Gene of Vibrio(Listonella) anguillarum Controls Expression of the Major Outer Membrane Proteins but Not Virulence in a Natural Host Model,” Infection and Immunity, vol. 69, no. 10, pp. 6091–6101.
De Oliveira, EJ, Rabinovitch, L, Monnerat, RG, Passos, LKJ & Zahner, V 2004, “Molecular Characterization of Brevibacillus laterosporus and Its Potential Use in Biological Control,” Applied and Environmental Microbiology, vol. 70, no. 11, pp. 6657–6664.
Patz, J, Epstein, P, Burke, T & Balbus, J 1996, “Global climate change and emerging infectious diseases,” JAMA, vol. 275, no. 3, pp. 217–223.
Petrosino, JF, Highlander, S, Luna, RA, Gibbs, R a & Versalovic, J 2009, “Metagenomic pyrosequencing and microbial identification.,” Clinical chemistry, vol. 55, no. 5, pp. 856–66.
Piel, J 2002, “A polyketide synthase-peptide synthetase gene cluster from an uncultured bacterial symbiont of Paederus beetles.,” Proceedings of the National Academy of Sciences of the United States of America, vol. 99, no. 22, pp. 14002–7.
Pilcher, N & Cabanban, A 2000a, “The status of coral reefs in Eastern Malaysia,” Global Coral Reef Monitoring Network (GCRMN) Report. Australia Institute of Marine Science, Townsville.
Pilcher, N & Cabanban, A 2000b, “The status of coral reefs in Sabah, Labuan and Sarawak, East Malaysia,” Australian Institute of Marine Sciences, Townsville.
Praveena, SM, Siraj, SS & Aris, AZ 2012, “Coral reefs studies and threats in Malaysia: a mini review,” Reviews in Environmental Science and Biotechnology, vol. 11, no. 1, pp. 27–39.
Prayitno, SB & Latchford, JW 1995, “Experimental infections of crustaceans with luminous bacteria related to Photobacterium and Vibrio. Effect of salinity and pH on infectiosity,” Aquaculture, vol. 132, no. 1–2, pp. 105–112.

P a g e | 111
Radjasa, OK & Sabdono, A 2003, “Screening of secondary metabolite-producing bacteria associated with corals using 16S rDNA-based approach,” Journal of Coastal Development, vol. 7, no. 1, pp. 11–19.
Raguénès, G, Christen, R, Guezennec, J, Pignet, P & Barbier, G 1997, “Vibrio diabolicus sp. nov., a New Polysaccharide-Secreting Organism Isolated from a Deep-Sea Hydrothermal Vent Polychaete Annelid, Alvinella pompejana,” International Journal of Systematic Bacteriology, vol. 47, no. 4, pp. 989–995.
Raina, J-B, Dinsdale, EA, Willis, BL & Bourne, DG 2010, “Do the organic sulfur compounds DMSP and DMS drive coral microbial associations?,” Trends in microbiology, vol. 18, no. 3, Elsevier Ltd, pp. 101–8.
Raina, J-B, Tapiolas, D, Willis, BL & Bourne, DG 2009, “Coral-associated bacteria and their role in the biogeochemical cycling of sulfur.,” Applied and environmental microbiology, vol. 75, no. 11, pp. 3492–501.
Rajendhran, J & Gunasekaran, P 2011, “Microbial phylogeny and diversity: Small subunit ribosomal RNA sequence analysis and beyond,” Microbiological Research, vol. 166, no. 2, pp. 99–110.
Ramaiah, N, Hill, RT, Chun, J, Ravel, J, Matte, MH, Straube, WL & Colwell, RR 2000, “Use of a chiA probe for detection of chitinase genes in bacteria from the Chesapeake Bay1,” FEMS Microbiology Ecology, vol. 34, no. 1, Blackwell Publishing Ltd, pp. 63–71.
Rappé, MS & Giovannoni, SJ 2003, “The uncultured microbial majority.,” Annual review of microbiology, vol. 57, Annual Reviews 4139 El Camino Way, P.O. Box 10139, Palo Alto, CA 94303-0139, USA, pp. 369–94.
Raven, J, Caldeira, K, Elderfield, H, Hoegh-Guldberg, O, Liss, P, Riebesell, U, Shepherd, J, Turley, C & Watson, A 2005, “Ocean acidification due to increasing atmospheric carbon dioxide,” The Royal Society.
Rehnstam, A-S, Bäckman, S, Smith, DC, Azam, F & Hagström, Å 1993, “Blooms of sequence-specific culturable bacteria in the sea,” FEMS Microbiology Letters, vol. 102, no. 3–4, pp. 161–166.
Reisch, CR, Moran, MA & Whitman, WB 2011, “Bacterial catabolism of dimethylsulfoniopropionate (DMSP),” Frontiers in microbiology, vol. 2, Frontiers Media SA.
Reshef, L, Koren, O, Loya, Y, Zilber-Rosenberg, I & Rosenberg, E 2006, “The coral probiotic hypothesis.,” Environmental microbiology, vol. 8, no. 12, pp. 2068–73.
Richardson, DJ, McEwan, AG, Jackson, JB & Ferguson, SJ 1989, “Electron transport pathways to nitrous oxide in Rhodobacter species,” European Journal of Biochemistry, vol. 185, no. 3, Blackwell Publishing Ltd, pp. 659–669.

P a g e | 112
Ritchie, K, Dennis, J, McGrath, T & Smith, G 1994, “Bacterial Asociated with Bleached and Nonbleached Areas of Montastrea Annularis,” Proceedings of the 5th Symposium on the Natural History of The Bahamas, vol. 5, pp. 75–80.
Ritchie, K & Smith, G 2004, “Microbial Communities of Coral Surface Mucopolysaccharide Layers,” in E Rosenberg & Y Loya (eds), Coral Health and Disease SE - 13, Springer Berlin Heidelberg, pp. 259–264.
Ritchie, KB 2006, “Regulation of microbial populations by coral surface mucus and mucus-associated bacteria,” Marine Ecology Progress Series, vol. 322, pp. 1–14.
Ritchie, KB & Smith, GW 1995a, “Carbon-source utilization patterns of coral-associated marine heterotrophs,” Journal of marine biotechnology, vol. 3, no. 1, pp. 105–107.
Ritchie, KB & Smith, GW 1995b, “Preferential carbon utilization by surface bacterial communities from water mass, normal, and white-band diseased Acropora cervicornis,” Molecular Marine Biology and Biotechnology, vol. 4, no. 4, pp. 345–352.
Rohwer, F, Seguritan, V, Azam, F & Knowlton, N 2002, “Diversity and distribution of coral-associated bacteria,” Marine Ecology Progress Series, vol. 243, pp. 1–10.
Rosenberg, E 2004, “The Bacterial Disease Hypothesis of Coral Bleaching,” in E Rosenberg & Y Loya (eds), Coral Health and Disease SE - 25, Springer Berlin Heidelberg, pp. 445–461.
Rosenberg, E & Ben-Haim, Y 2002, “Microbial diseases of corals and global warming,” Environmental Microbiology, vol. 4, no. 6, Blackwell Science Ltd, pp. 318–326.
Rosenberg, E & Falkovitz, L 2004, “The Vibrio shiloi/Oculina patogonica Model System of Coral Bleaching,” Annual Review of Microbiology, vol. 58, no. 1, Annual Reviews, pp. 143–159.
Rosenberg, E, Koren, O, Reshef, L, Efrony, R & Zilber-Rosenberg, I 2007, “The role of microorganisms in coral health, disease and evolution,” Nature reviews. Microbiology, vol. 5, no. 5, pp. 355–62.
Rosenberg, E, Kushmaro, A, Kramarsky-Winter, E, Banin, E & Yossi, L 2009, “The role of microorganisms in coral bleaching,” The ISME journal, vol. 3, no. 2, pp. 139–46.
Saha, R, Spröer, C, Beck, B & Bagley, S 2010, “Pseudomonas oleovorans subsp. lubricantis subsp. nov., and Reclassification of Pseudomonas pseudoalcaligenes ATCC 17440T as Later Synonym of Pseudomonas oleovorans ATCC 8062T,” Current Microbiology, vol. 60, no. 4, Springer-Verlag, pp. 294–300.

P a g e | 113
Sanger, F, Nicklen, S & Coulson, AR 1977, “DNA sequencing with chain-terminating inhibitors,” Proceedings of the National Academy of Sciences, vol. 74, no. 12, pp. 5463–5467.
Satomi, M, Vogel, BF, Gram, L & Venkateswaran, K 2006, “Shewanella hafniensis sp. nov. and Shewanella morhuae sp. nov., isolated from marine fish of the Baltic Sea,” International Journal of Systematic and Evolutionary Microbiology, vol. 56, no. 1, pp. 243–249.
Saunders, VA 1978, “Genetics of Rhodospirillaceae.,” Microbiological reviews, vol. 42, no. 2, pp. 357–84.
Saville Waid, J 1999, “Does soil biodiversity depend upon metabiotic activity and influences?,” Applied Soil Ecology, vol. 13, no. 2, pp. 151–158.
Sayeh, R, Birrien, J, Alain, K, Barbier, G, Hamdi, M & Prieur, D 2010, “Microbial diversity in Tunisian geothermal springs as detected by molecular and culture-based approaches,” Extremophiles, vol. 14, no. 6, Springer Japan, pp. 501–514.
Scarratt, M, Cantin, G, Levasseur, M & Michaud, S 2000, “Particle size-fractionated kinetics of DMS production: where does DMSP cleavage occur at the microscale?,” Journal of Sea Research, vol. 43, no. 3–4, pp. 245–252.
Schut, F, Prins, R & Gottschal, J 1997, “Oligotrophy and pelagic marine bacteria: facts and fiction,” Aquatic Microbial Ecology, vol. 12, no. 2, pp. 177–202.
Schut, F, de Vries, EJ, Gottschal, JC, Robertson, BR, Harder, W, Prins, RA & Button, DK 1993, “Isolation of Typical Marine Bacteria by Dilution Culture: Growth, Maintenance, and Characteristics of Isolates under Laboratory Conditions,” Applied and Environmental Microbiology, vol. 59, no. 7, pp. 2150–2160.
Sekiguchi, H, Watanabe, M, Nakahara, T, Xu, B & Uchiyama, H 2002, “Succession of Bacterial Community Structure along the Changjiang River Determined by Denaturing Gradient Gel Electrophoresis and Clone Library Analysis,” Applied and Environmental Microbiology, vol. 68, no. 10, pp. 5142–5150.
Shannon, CE & Weaver, W 1963, Mathematical theory of communication, University Illinois Press, p. 117.
Sharon, G & Rosenberg, E 2008, “Bacterial growth on coral mucus,” Current microbiology, vol. 56, no. 5, pp. 481–8.
Shaw, G 1983, “Bio-controlled thermostasis involving the sulfur cycle,” Climatic Change, vol. 5, no. 3, Kluwer Academic Publishers, pp. 297–303.
Shivaji, S, Suresh, K, Chaturvedi, P, Dube, S & Sengupta, S 2005, “Bacillus arsenicus sp. nov., an arsenic-resistant bacterium isolated from a siderite concretion in

P a g e | 114
West Bengal, India,” International Journal of Systematic and Evolutionary Microbiology, vol. 55, no. 3, pp. 1123–1127.
Shnit-Orland, M & Kushmaro, A 2008, “Coral mucus bacteria as a source for antibacterial activity,” Proceedings of the 11th International Coral Reef Symposium, no. 8, pp. 257–259.
Shnit-Orland, M & Kushmaro, A 2009, “Coral mucus-associated bacteria: a possible first line of defense,” FEMS microbiology ecology, vol. 67, no. 3, pp. 371–80.
Sievert, SM, Kiene, RP & Schultz-Vogt, HN 2007, “The sulfur cycle,” Oceanography, vol. 20, no. 2, Oceanography Society, pp. 117–123.
Silakowski, B, Kunze, B & Müller, R 2001, “Multiple hybrid polyketide synthase/non-ribosomal peptide synthetase gene clusters in the myxobacterium Stigmatella aurantiaca,” Gene, vol. 275, no. 2, pp. 233–240.
Simó, R 2001, “Production of atmospheric sulfur by oceanic plankton: biogeochemical, ecological and evolutionary links,” Trends in Ecology & Evolution, vol. 16, no. 6, pp. 287–294.
Singh, BK 2010, “Exploring microbial diversity for biotechnology: the way forward,” Trends in Biotechnology, vol. 28, no. 3, pp. 111–116.
Slezak, D & Brugger, A 2001, “Impact of solar radiation on the biological removal of dimethylsulfoniopropionate and dimethylsulfide in marine surface waters,” Aquatic Microbial Ecology, vol. 25, no. 1, pp. 87–97.
Smith, B & Wilson, JB 1996, “A consumer’s guide to evenness indices,” Oikos, JSTOR, pp. 70–82.
Sogin, ML, Morrison, HG, Huber, JA, Welch, DM, Huse, SM, Neal, PR, Arrieta, JM & Herndl, GJ 2006, “Microbial diversity in the deep sea and the underexplored ‘rare biosphere,’” Proceedings of the National Academy of Sciences, vol. 103, no. 32, pp. 12115–12120.
De Souza, MP & Yoch, DC 1995, “Comparative Physiology of Dimethyl Sulfide Production by Dimethylsulfoniopropionate Lyase in Pseudomonas doudoroffii and Alcaligenes sp. Strain M3A.,” Applied and Environmental Microbiology, vol. 61, no. 11, pp. 3986–3991.
De Souza, MP, Yoch, DC & Souza, M 1996, “N-Terminal Amino Acid Sequences and Comparison of DMSP Lyases from Pseudomonas Doudoroffii and Alcagenes Strain M3A,” in R Kiene, P Visscher, M Keller & G Kirst (eds), Biological and environmental chemistry of DMSP and related sulfonium compounds, Springer, pp. 293–304.

P a g e | 115
Stefels, J 2000, “Physiological aspects of the production and conversion of DMSP in marine algae and higher plants,” Journal of Sea Research, vol. 43, no. 3–4, pp. 183–197.
Stelma, GN, Reyes, AL, Peeler, JT, Johnson, CH & Spaulding, PL 1992, “Virulence characteristics of clinical and environmental isolates of Vibrio vulnificus.,” Applied and Environmental Microbiology, vol. 58, no. 9, pp. 2776–2782.
Stevens, H & Brinkhoff, T 2005, “Composition of free-living, aggregate-associated and sediment surface-associated bacterial communities in the German Wadden Sea,” Aquatic Microbial Ecology, vol. 38, no. 1, pp. 15–30.
Stove Poindexter, JL & Cohen-Bazire, G 1964, “The Fine Structure of Stalked Bacteria Belonging to the Family Caulobacteraceae,” The Journal of Cell Biology, vol. 23, no. 3, pp. 587–607.
Stover, CK, Pham, XQ, Erwin, AL, Mizoguchi, SD, Warrener, P, Hickey, MJ, Brinkman, FSL, Hufnagle, WO, Kowalik, DJ, Lagrou, M, Garber, RL, Goltry, L, Tolentino, E, Westbrock-Wadman, S, Yuan, Y, Brody, LL, Coulter, SN, Folger, KR, Kas, A, Larbig, K, Lim, R, Smith, K, Spencer, D, Wong, GK-S, Wu, Z, Paulsen, IT, Reizer, J, Saier, MH, Hancock, REW, Lory, S & Olson, M V 2000, “Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen,” Nature, vol. 406, no. 6799, pp. 959–964.
Strom, SL 2008, “Microbial Ecology of Ocean Biogeochemistry: A Community Perspective,” Science, vol. 320, no. 5879, pp. 1043–1045.
Sucharita, K, Sasikala, C, Park, SC, Baik, KS, Seong, CN & Ramana, C V 2009, “Shewanella chilikensis sp. nov., a moderately alkaliphilic gammaproteobacterium isolated from a lagoon,” International Journal of Systematic and Evolutionary Microbiology, vol. 59, no. 12, pp. 3111–3115.
Sunagawa, S, DeSantis, TZ, Piceno, YM, Brodie, EL, DeSalvo, MK, Voolstra, CR, Weil, E, Andersen, GL & Medina, M 2009, “Bacterial diversity and White Plague Disease-associated community changes in the Caribbean coral Montastraea faveolata,” ISME J, vol. 3, no. 5, International Society for Microbial Ecology, pp. 512–521.
Sunda, W, Kieber, DJ, Kiene, RP & Huntsman, S 2002, “An antioxidant function for DMSP and DMS in marine algae,” Nature, vol. 418, no. 6895, Macmillian Magazines Ltd., pp. 317–320.
Sussman, M, Mieog, JC, Doyle, J, Victor, S, Willis, BL & Bourne, DG 2009, “Vibrio zinc-metalloprotease causes photoinactivation of coral endosymbionts and coral tissue lesions.,” PloS one, vol. 4, no. 2, p. e4511.
Sutherland, KP, Porter, JW & Torres, C 2004, “Disease and immunity in Caribbean and Indo-Pacific zooxanthellate corals,” Marine ecology. Progress series, vol. 266, Inter-Research, pp. 273–302.

P a g e | 116
Suzuki, MT, Rappé, MS, Haimberger, ZW, Winfield, H, Adair, N, Ströbel, J & Giovannoni, SJ 1997, “Bacterial diversity among small-subunit rRNA gene clones and cellular isolates from the same seawater sample.,” Applied and Environmental Microbiology, vol. 63, no. 3, pp. 983–989.
Tamura, K, Peterson, D, Peterson, N, Stecher, G, Nei, M & Kumar, S 2011, “MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods.,” Molecular biology and evolution, vol. 28, no. 10, pp. 2731–9.
Tamura, K, Stecher, G, Peterson, D, Filipski, A & Kumar, S 2013, “MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0,” Molecular Biology and Evolution, vol. 30, no. 12, pp. 2725–2729.
Tan, CH & Heron, SF 2011, “First observed severe mass bleaching in Malaysia, Greater Coral Triangle,” Galaxea, Journal of Coral Reef Studies, vol. 13, no. 1, pp. 27–28.
Tao, L, Peng, W & Pinxian, W 2008, “Microbial diversity in surface sediments of the Xisha Trough, the South China Sea,” Acta Ecologica Sinica, vol. 28, no. 3, pp. 1166–1173.
Taylor, BF 1993, “Bacterial Transformations of Organic Sulfur Compounds in Marine Environments,” in R Oremland (ed.), Biogeochemistry of Global Change SE - 40, Springer US, pp. 745–781.
Tian, B, Yang, J, Lian, L, Wang, C, Li, N & Zhang, K-Q 2007, “Role of an extracellular neutral protease in infection against nematodes by Brevibacillus laterosporus strain G4,” Applied Microbiology and Biotechnology, vol. 74, no. 2, Springer-Verlag, pp. 372–380.
Todd, JD, Curson, ARJ, Dupont, CL, Nicholson, P & Johnston, AWB 2009, “The dddP gene, encoding a novel enzyme that converts dimethylsulfoniopropionate into dimethyl sulfide, is widespread in ocean metagenomes and marine bacteria and also occurs in some Ascomycete fungi.,” Environmental microbiology, vol. 11, no. 6, pp. 1376–85.
Todd, JD, Curson, ARJ, Kirkwood, M, Sullivan, MJ, Green, RT & Johnston, AWB 2011, “DddQ, a novel, cupin-containing, dimethylsulfoniopropionate lyase in marine roseobacters and in uncultured marine bacteria,” Environmental Microbiology, vol. 13, no. 2, Blackwell Publishing Ltd, pp. 427–438.
Todd, JD, Curson, ARJ, Sullivan, MJ, Kirkwood, M & Johnston, AWB 2012, “The Ruegeria pomeroyi acuI gene has a role in DMSP catabolism and resembles yhdH of E. coli and other bacteria in conferring resistance to acrylate.,” PloS one, vol. 7, no. 4, p. e35947.
Todd, JD, Kirkwood, M, Newton-Payne, S & Johnston, AWB 2012, “DddW, a third DMSP lyase in a model Roseobacter marine bacterium, Ruegeria pomeroyi

P a g e | 117
DSS-3,” ISME J, vol. 6, no. 1, International Society for Microbial Ecology, pp. 223–226.
Todd, JD, Rogers, R, Li, YG, Wexler, M, Bond, PL, Sun, L, Curson, ARJ, Malin, G, Steinke, M & Johnston, AWB 2007, “Structural and Regulatory Genes Required to Make the Gas Dimethyl Sulfide in Bacteria,” Science, vol. 315, no. 5812, pp. 666–669.
Toole, DA, Slezak, D, Kiene, RP, Kieber, DJ & Siegel, DA 2006, “Effects of solar radiation on dimethylsulfide cycling in the western Atlantic Ocean,” Deep Sea Research Part I: Oceanographic Research Papers, vol. 53, no. 1, pp. 136–153.
Trevena, AJ, Jones, GB, Wright, SW & van den Enden, RL 2003, “Profiles of dimethylsulphoniopropionate (DMSP), algal pigments, nutrients, and salinity in the fast ice of Prydz Bay, Antarctica,” Journal of Geophysical Research: Oceans, vol. 108, no. C5, p. 3145.
Trevena, AJ, Jones, GB, Wright, SW & van den Enden, RL 2000, “Profiles of DMSP, algal pigments, nutrients and salinity in pack ice from eastern Antarctica,” Journal of Sea Research, vol. 43, no. 3–4, pp. 265–273.
Tun, K, Chou, LM, Low, J, Yeemin, T, Phongsuwan, N, Setiasih, N, Wilson, J, Amri, AY, Adzis, KAA, Lane, D, van Bochove, JW, Kluskens, B, Long, N Van, Tuan, VS & Gomez, E 2010, “A Regional overview on the 2010 coral bleaching event in Southeast Asia,” Status of Coral Reefs in East Asian Seas Region: 2010, Ministry of the Environment, Japan, pp. 1–27.
Turner, JT & Backman, PA 1991, “Factors relating to peanut yield increases after seed treatment with Bacillus subtilis,” Plant disease, vol. 75, no. 4, American Phytopathological Society, pp. 347–353.
Uchino, M, Shida, O, Uchimura, T & Komagata, K 2001, “Recharacterization of Pseudomonas fulva Iizuka and Komagata 1963, and proposals of Pseudomonas parafulva sp. nov. and Pseudomonas cremoricolorata sp. nov.,” The Journal of General and Applied Microbiology, vol. 47, no. 5, pp. 247–261.
Del Valle, D, Kiene, R & Karl, D 2012, “Effect of visible light on dimethylsulfoniopropionate assimilation and conversion to dimethylsulfide in the North Pacific Subtropical Gyre,” Aquatic Microbial Ecology, vol. 66, no. 1, pp. 47–62.
Vallina, SM & Simó, R 2007, “Strong Relationship Between DMS and the Solar Radiation Dose over the Global Surface Ocean,” Science, vol. 315, no. 5811, pp. 506–508.
Varaljay, VA, Gifford, SM, Wilson, ST, Sharma, S, Karl, DM & Moran, MA 2012, “Bacterial dimethylsulfoniopropionate degradation genes in the oligotrophic north pacific subtropical gyre.,” Applied and environmental microbiology, vol. 78, no. 8, pp. 2775–82.

P a g e | 118
Varaljay, VA, Howard, EC, Sun, S & Moran, MA 2010, “Deep sequencing of a dimethylsulfoniopropionate-degrading gene (dmdA) by using PCR primer pairs designed on the basis of marine metagenomic data.,” Applied and environmental microbiology, vol. 76, no. 2, pp. 609–17.
Veron, J, Devantier, LM, Turak, E, Green, AL, Kininmonth, S, Stafford-Smith, M & Peterson, N 2009, “Delineating the Coral Triangle,” Galaxea, Journal of Coral Reef Studies, vol. 11, no. 2, pp. 91–100.
Veron, JEN & Stafford-Smith, M 2000, Corals of the World.
Vila-Costa, M, Rinta-Kanto, JM, Sun, S, Sharma, S, Poretsky, R & Moran, MA 2010, “Transcriptomic analysis of a marine bacterial community enriched with dimethylsulfoniopropionate,” ISME J, vol. 4, no. 11, International Society for Microbial Ecology, pp. 1410–1420.
Vishnivetskaya, T, Kathariou, S & Tiedje, J 2009, “The Exiguobacterium genus: biodiversity and biogeography,” Extremophiles, vol. 13, no. 3, Springer Japan, pp. 541–555.
Vizcaino, MI, Johnson, WR, Kimes, NE, Williams, K, Torralba, M, Nelson, KE, Smith, GW, Weil, E, Moeller, PDR & Morris, PJ 2010, “Antimicrobial resistance of the coral pathogen Vibrio coralliilyticus and Caribbean sister phylotypes isolated from a diseased octocoral.,” Microbial ecology, vol. 59, no. 4, pp. 646–57.
Wagner-Döbler, I, Rheims, H, Felske, A, El-Ghezal, A, Flade-Schröder, D, Laatsch, H, Lang, S, Pukall, R & Tindall, BJ 2004, “Oceanibulbus indolifex gen. nov., sp. nov., a North Sea alphaproteobacterium that produces bioactive metabolites,” International Journal of Systematic and Evolutionary Microbiology, vol. 54, no. 4, pp. 1177–1184.
Wang, B, Tan, T & Shao, Z 2009, “Roseovarius pacificus sp. nov., isolated from deep-sea sediment,” International Journal of Systematic and Evolutionary Microbiology, vol. 59, no. 5, pp. 1116–1121.
Ward, DM, Weller, R & Bateson, MM 1990, “16S rRNA sequences reveal numerous uncultured microorganisms in a natural community,” Nature, vol. 345, no. 6270, pp. 63–65.
Watanabe, A & Ebizuka, Y 2004, “Unprecedented Mechanism of Chain Length Determination in Fungal Aromatic Polyketide Synthases,” Chemistry & Biology, vol. 11, no. 8, pp. 1101–1106.
Webster, NS, Wilson, KJ, Blackall, LL & Hill, RT 2001, “Phylogenetic Diversity of Bacteria Associated with the Marine Sponge Rhopaloeides odorabile,” Applied and Environmental Microbiology, vol. 67, no. 1, pp. 434–444.

P a g e | 119
Welsh, DT 2000, “Ecological significance of compatible solute accumulation by micro-organisms: from single cells to global climate,” FEMS Microbiology Reviews, vol. 24, no. 3, Blackwell Publishing Ltd, pp. 263–290.
Wiese, J, Thiel, V, Nagel, K, Staufenberger, T & Imhoff, J 2009, “Diversity of Antibiotic-Active Bacteria Associated with the Brown Alga Laminaria saccharina from the Baltic Sea,” Marine Biotechnology, vol. 11, no. 2, Springer-Verlag, pp. 287–300.
Wilkinson, CR 1999, “Global and local threats to coral reef functioning and existence: review and predictions,” Marine and Freshwater Research, vol. 50, no. 8, pp. 867–878.
Wilkinson, CR & Buddemeier, RW 1994, Global Climate Change and Coral Reefs: Implications for People and Reefs: Report of the UNEP-IOC-ASPEI-IUCN Global Task Team on the Implications of Climate Change on Coral Reefs, IUCN.
Wilson, B, Aeby, GS, Work, TM & Bourne, DG 2012, “Bacterial communities associated with healthy and Acropora white syndrome-affected corals from American Samoa,” FEMS Microbiology Ecology, vol. 80, no. 2, pp. 509–520.
Wipat, A & Harwood, CR 1999, “The Bacillus subtilis genome sequence: the molecular blueprint of a soil bacterium,” FEMS Microbiology Ecology, vol. 28, no. 1, Blackwell Publishing Ltd, pp. 1–9.
Xu, X-W, Wu, Y-H, Wang, C-S, Gao, X-H, Wang, X-G & Wu, M 2010, “Pseudoalteromonas lipolytica sp. nov., isolated from the Yangtze River estuary,” International Journal of Systematic and Evolutionary Microbiology, vol. 60, no. 9, pp. 2176–2181.
Yahya, NK, Hassan, R & Husaini, AASA 2012, “Molecular Phylogeny of Sarawak Green Sea Turtle (Chelonia mydas) inferred by the D-loop region and 16S rRNA gene.,” Borneo Journal of Resource Science and Technology, vol. 2, no. 1, pp. 20–27.
Yaman, ARBG n.d., “Coral Reefs in the Coastal Waters of the South China Sea.”
Yoch, DC 2002, “Dimethylsulfoniopropionate: Its Sources, Role in the Marine Food Web, and Biological Degradation to Dimethylsulfide,” Applied and Environmental Microbiology, vol. 68, no. 12, pp. 5804–5815.
Yoch, DC, Ansede, JH & Rabinowitz, KS 1997, “Evidence for Intracellular and Extracellular Dimethylsulfoniopropionate (DMSP) Lyases and DMSP Uptake Sites in Two Species of Marine Bacteria.,” Applied and Environmental Microbiology, vol. 63, no. 8, pp. 3182–3188.
Yoshida, A, Nishimura, M & Kogure, K 2007, “Bacterial community structure in the Sulu Sea and adjacent areas,” Deep Sea Research Part II: Topical Studies in Oceanography, vol. 54, no. 1–2, pp. 103–113.

P a g e | 120
Zahner, V, Rabinovitch, L, Suffys, P & Momen, H 1999, “Genotypic Diversity among Brevibacillus laterosporus Strains,” Applied and Environmental Microbiology, vol. 65, no. 11, pp. 5182–5185.
Zhang, Z, Schwartz, S, Wagner, L & Miller, W 2000, “A greedy algorithm for aligning DNA sequences,” Journal of Computational biology, vol. 7, no. 1-2, Mary Ann Liebert, Inc., pp. 203–214.
Ziemke, F, Höfle, MG, Lalucat, J & Rossellö-Mora, R 1998, “Reclassification of Shewanella putrefaciens Owen’s genomic group II as Shewanella baltica sp. nov.,” International Journal of Systematic Bacteriology, vol. 48, no. 1, pp. 179–186.
Zumft, WG 1997, “Cell biology and molecular basis of denitrification.,” Microbiology and Molecular Biology Reviews, vol. 61, no. 4, pp. 533–616.

P a g e | 121
APPENDIX
Table A.1: 16S rRNA gene sequence analysis of bacterial cultures from Kuching
1611, based on BLAST analysis.
Sequence GenBank accession number
Closest match Identities Phylogenetic division
1611-S1-01-1
KF373266 Pseudomonas aeruginosa strain 11.2 [JX286673]
460/460 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
1611-S1-01-1.3
KF373267 Vibrio harveyi strain E385 [JX290081]
462/463 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-01-2
KF373268 Pseudomonas sp. Mexd38 [JX436405]
462/462 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
1611-S1-05-1
KF373269
Vibrio parahaemolyticus strain 448 [JN188417]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-05-2.1
KF373270
Vibrio parahaemolyticus strain 448 [JN188417]
476/476 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-05-2.2
KF373271
Vibrio parahaemolyticus strain 448 [JN188417]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-05-3
KF373272
Vibrio parahaemolyticus strain 448 [JN188417]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-10-1.1.1
KF373273 Vibrio alginolyticus strain XSBZ14 [JX221045]
397/464 (86%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S1-10-1.1.2.1
KF373274 Shewanella chilikensis strain JC5 [HM016088]
461/463 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S1-10-1.1.2.2
KF373275 Photobacterium sp. TKY4 [AB583193]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Photobacterium
1611-S1-10-1.2.1
KF373276 Bacillus sphaericus isolate BS11 [AM269451]
475/475 (100%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Lysinibacillus
1611-S1-10-2.1
KF373277 Oceanimonas sp. D6083 [FJ161317]
425/462 (92%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
1611-S2-01-1.1.2
KF373278 Shewanella haliotis strain Z4 [JX286502]
425/426 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S2-01-2.1
KF373279 Shewanella haliotis strain Z4 [JX286502]
467/467 (100%)
Gammaproteobacteria; Alteromonadales;

P a g e | 122
Shewanellaceae; Shewanella
1611-S2-01-2.2
KF373280 Shewanella haliotis strain MS41 [FN997635]
463/466 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S2-01-2.3
KF373281 Shewanella haliotis strain MS41 [FN997635]
469/469 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S2-01-3.2
KF373282 Pseudomonas sp. Mexd38 [JX436405]
461/461 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
1611-S2-05-1.1
KF373283 Shewanella haliotis strain MS41 [FN997635]
468/468 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S2-05-1.2
KF373284
Vibrio parahaemolyticus strain Aj2010072802A90 [JF432066]
474/474 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S2-05-2
KF373285 Shewanella haliotis strain Z4 [JX286502]
467/467 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
1611-S2-10-1
KF373286 Oceanimonas sp. D6083 [FJ161317]
461/461 (100%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
1611-S2-10-2
KF373287 Bacillus sp. 1-1(2012) [JN942108]
461/461 (100%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Bacillus
1611-S4-01-1
KF373288
Vibrio parahaemolyticus isolate Mm004 [FR686998]
460/464 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-01-1.1
KF373289 Oceanimonas sp. D6083 [FJ161317]
463/463 (100%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
1611-S4-01-2.1.1
KF373290
Vibrio parahaemolyticus strain 448 [JN188417]
471/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-01-2.2
KF373291 Vibrio natriegens strain AUCASVE5 [JQ277719]
458/465 (98%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-01-3.1
KF373292
Vibrio parahaemolyticus strain 448 [JN188417]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-01-3.1.2
KF373293
Vibrio parahaemolyticus strain 448 [JN188417]
471/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-01-3.2
KF373294
Vibrio parahaemolyticus strain 448 [JN188417]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-05-1
KF373295 Roseovarius pacificus strain 81-2 [NR_043564]
405/405 (100%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Roseovarius
1611-S4-05-2
KF373296 Roseovarius pacificus strain 81-2
408/408 (100%)
Alphaproteobacteria; Rhodobacterales;

P a g e | 123
[NR_043564] Rhodobacteraceae; Roseovarius 1611-S4-05-3
KF373297 Vibrio harveyi isolate VHJR6 [DQ995240]
475/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-10-1.2
KF373298 Vibrio azureus strain M2-164 [JQ810832]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-10-2
KF373299
Vibrio parahaemolyticus strain RW1 [FJ172044]
477/479 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-10-3.1
KF373300 Vibrio rotiferianus strain 5S [JF792070]
469/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S4-10-3.2
KF373301 Vibrio natriegens strain AUCASVE5 [JQ277719]
470/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S5-01-1
KF373302 Bacillus subtilis strain y86-7 [FJ460478]
453/475 (95%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Bacillus
1611-S5-05-1
KF373303 Vibrio natriegens strain CM3 [EU660320]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S5-05-3.1
KF373304
Vibrio parahaemolyticus isolate Mm004 [FR686998]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S5-05-3.2
KF373305 Burzellia piscidermidis strain P6-6 [EU127296]
464/464 (100%)
Gammaproteobacteria; Burzellia
1611-S5-10-1
KF373306 Burzellia piscidermidis strain P6-6 [EU127296]
463/464 (99%)
Gammaproteobacteria; Burzellia
1611-S5-10-2
KF373307 Vibrio azureus strain F77118 [HQ908716]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-01-1.1
KF373308 Vibrio sinaloensis strain CAIM 1068 [HM584056]
469/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-01-1.2
KF373309
Pseudoalteromonas maricaloris strain KMM636 [NR_025009]
458/458 (100%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
1611-S6-01-2
KF373310 Rhodobacteraceae bacterium SCSWE04 [FJ461471]
360/375 (96%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae
1611-S6-05-1.1
KF373311
Stenotrophomonas maltophilia strain BQAPs-03d [FJ217200]
471/471 (100%)
Gammaproteobacteria; Xanthomonadales; Xanthomonadaceae; Stenotrophomonas; Stenotrophomonas maltophilia group
1611-S6-05-1.2
KF373312 Vibrio harveyi isolate VHJR6 [DQ995240]
472/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-05-2.1
KF373313 Vibrio harveyi isolate VHJR14 [EF011651]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-05-2.2
KF373314 Vibrio harveyi strain HL19 [JQ948038]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-05-3.2
KF373315 Vibrio harveyi isolate VHJR6 [DQ995240]
468/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6-10-1.1
KF373316 Vibrio harveyi isolate VHJR6 [DQ995240]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1611-S6- KF373317 Vibrio harveyi isolate 477/477 Gammaproteobacteria;

P a g e | 124
10-1.2 VHJR12 [DQ995245] (100%) Vibrionales; Vibrionaceae; Vibrio
1611-S6-10-2
KF373318 Vibrio campbellii strain CAIM 886 [HM584033]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio

P a g e | 125
Table A.2: 16S rRNA gene sequence analysis of bacterial cultures from Kuching
1911, based on BLAST analysis.
Sequence GenBank accession number
Closest match Identities Phylogenetic division
1911-S1-01-1.2.1
KF373319 Vibrio orientalis strain JC97, isolate Pkl-17 [FR837599]
465/468 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S1-01-1.2.2
KF373320 Rhodobacter capsulatus strain PSB-06 [FJ866784]
434/440 (99%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Rhodobacter
1911-S1-01-2
KF373321 Rhodobacter capsulatus strain PSB-06 [FJ866784]
440/455 (97%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Rhodobacter
1911-S1-01-3
KF373322 Rhodobacter capsulatus strain PSB-06 [FJ866784]
433/440 (98%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Rhodobacter
1911-S1-05-2
KF373323 Pseudomonas oleovorans strain HNS030 [JN128264]
456/457 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
1911-S1-07-1
KF373324 Pseudomonas oleovorans strain HNS030 [JN128264]
459/460 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
1911-S2-01-1
KF373325 Vibrio alginolyticus isolate Va150 [EU155497]
476/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S2-05-1
KF373326 Vibrio alginolyticus strain HZBC71 [JN188402]
471/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S2-07-1
KF373327 Vibrio alginolyticus strain HZBC71 [JN188402]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S2-07-2
KF373328
Vibrio parahaemolyticus isolate Vp481 [EU155540]
471/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-01-1.1.1
KF373329 Pseudoalteromonas ganghwensis [DQ011614]
464/465 (99%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
1911-S3-01-1.1.2
KF373330
Vibrio parahaemolyticus strain VPMP55 [JQ663925]
319/402 (79%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-01-1.2
KF373331 Vibrio alginolyticus strain P61224 [AJ704375]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-01-2
KF373332 Vibrio diabolicus strain KM30-12-3 [JQ670740]
475/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-05-1
KF373333
Vibrio parahaemolyticus strain 93A-5807 [DQ497398]
474/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3- KF373334 Vibrio 470/473 Gammaproteobacteria;

P a g e | 126
05-2 parahaemolyticus strain 93A-5807 [DQ497398]
(99%) Vibrionales; Vibrionaceae; Vibrio
1911-S3-10-1.1
KF373335 Vibrio harveyi strain IS01 [GU974342]
473/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-10-1.2
KF373336 Vibrio campbellii strain CAIM 886 [HM584033]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S3-10-2.1
KF373337 Vibrio rotiferianus strain BV1 [JN391272]
475/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S4-01-1
KF373338 Pseudoalteromonas ganghwensis [DQ011614]
462/463 (99%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
1911-S4-01-1.1
KF373339 Vibrio alginolyticus strain H050815-1 [EF219054]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S4-01-2.2
KF373340 Thalassospira xiamenensis strain PTG4-18 [EU603449]
411/416 (99%)
Alphaproteobacteria; Rhodospirillales; Rhodospirillaceae; Thalassospira
1911-S4-05-1.1
KF373341 Citrobacter freundii strain AIMST Ehe5 [JQ312038]
461/462 (99%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Citrobacter
1911-S4-05-1.2
KF373342
Leclercia adecarboxylata strain AIMST Ehe6 [JQ312039]
461/462 (99%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Leclercia
1911-S4-05-2
KF373343 Vibrio azureus strain 41113 [HM032787]
452/468 (97%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S4-10-2.1
KF373344 Vibrio alginolyticus strain H050815-1 [EF219054]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-01-1
KF373345 Vibrio natriegens strain AUCASVE1 [JQ043186]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-01-2.1
KF373346 Vibrio natriegens strain AUCASVE1 [JQ043186]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-01-2.2
KF373347 Vibrio natriegens strain AUCASVE1 [JQ043186]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-05-1.1.2
KF373348 Citrobacter freundii strain AIMST Ehe5 [JQ312038]
462/463 (99%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Citrobacter
1911-S5-05-1.2
KF373349 Vibrio natriegens strain AUCASVE1 [JQ043186]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-05-1.2.1
KF373350 Vibrio azureus strain F77118 [HQ908716]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-05-2
KF373351 Vibrio parahaemolyticus strain 448 [JN188417]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5- KF373352 Vibrio natriegens 471/472 Gammaproteobacteria;

P a g e | 127
05-3 strain AUCASVE1 [JQ043186]
(99%) Vibrionales; Vibrionaceae; Vibrio
1911-S5-10-1
KF373353 Vibrio azureus strain 41113 [HM032787]
471/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
1911-S5-10-2
KF373354 Vibrio splendidus strain AP625 [GQ254509]
469/471 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio

P a g e | 128
Table A.3: 16S rRNA gene sequence analysis of bacterial cultures from Kota
Kinabalu, based on BLAST analysis.
Sequence GenBank accession number
Closest match Identities Phylogenetic division
2311-S1-01-1.1
KF373355 Pseudomonas oleovorans strain HNS030 [JN128264]
452/453 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
2311-S1-01-1.2
KF373356 Shewanella haliotis strain MS41 [FN997635]
461/461 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S1-01-2.1
KF373357 Shewanella haliotis strain MS41 [FN997635]
469/469 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S1-01-2.2
KF373358 Shewanella haliotis strain MS41 [FN997635]
467/467 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S1-01-3.1
KF373359 Shewanella haliotis strain MS41 [FN997635]
466/466 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S1-05-1
KF373360 Exiguobacterium aurantiacum var. Colo. Road [AY047481]
485/485 (100%)
Firmicutes; Bacilli; Bacillales; Bacillales Family XII. Incertae Sedis; Exiguobacterium
2311-S1-05-2
KF373361 Oceanimonas smirnovii strain 31-13 [NR_042963]
442/461 (96%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2311-S1-10-1
KF373362 Vibrio rotiferianus strain 5S [JF792070]
466/470 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S2-01-1
KF373363 Brevibacillus laterosporus strain GZUB11 [FJ434663]
472/472 (100%)
Firmicutes; Bacilli; Bacillales; Paenibacillaceae; Brevibacillus
2311-S2-10-1
KF373364 Vibrio splendidus strain AP625 [GQ254509]
414/453 (91%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S3-01-1.1
KF373365 Bacillus sphaericus clone 7-16 [DQ364585]
431/456 (95%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Lysinibacillus
2311-S3-01-1.2
KF373366 Shewanella putrefaciens strain R1418 [AB208055]
455/461 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S3-01-2
KF373367 Shewanella putrefaciens strain R1418 [AB208055]
459/462 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2311-S3-01-3
KF373368 Vibrio vulnificus strain W045 [EF114147]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S3-05-1
KF373369 Enterobacter ludwigii strain KW 93 [JX262395]
463/463 (100%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Enterobacter
2311-S3-05-2.1
KF373370
Pseudomonas plecoglossicida strain AIMST Aie20 [JQ312025]
459/459 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
2311-S3- KF373371 Thalassospira sp. SKUK 417/417 Alphaproteobacteria;

P a g e | 129
10-1 MB1005 [EU907920] (100%) Rhodospirillales; Rhodospirillaceae; Thalassospira
2311-S3-10-2.1
KF373372 Bacillus malacitensis strain TP12 [FJ887890]
404/408 (99%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Bacillus
2311-S3-10-2.2
KF373373 Vibrio natriegens strain AUCASVE5 [JQ277719]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S4-01-1
KF373374 Providencia sp. Sam130-9A [FJ418577]
456/460 (99%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Providencia
2311-S4-05-1
KF373375 Nitratireductor basaltis strain J3 [NR_044414]
409/409 (100%)
Alphaproteobacteria; Rhizobiales; Phyllobacteriaceae; Nitratireductor
2311-S4-10-1
KF373376 Oceanimonas smirnovii strain 31-13 [NR_042963]
463/468 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2311-S4-10-2.1.1
KF373377 Oceanimonas smirnovii strain 31-13 [NR_042963]
463/468 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2311-S4-10-2.1.3
KF373378 Lysinibacillus fusiformis strain R3 [JQ991002]
476/476 (100%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Lysinibacillus
2311-S4-10-2.2
KF373379 Exiguobacterium aurantiacum var. Colo. Road [AY047481]
489/490 (99%)
Firmicutes; Bacilli; Bacillales; Bacillales Family XII. Incertae Sedis; Exiguobacterium
2311-S4-10-2.3
KF373380 Oceanimonas smirnovii strain 31-13 [NR_042963]
460/465 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2311-S4-18-1.1
KF373381 Vibrio vulnificus strain W045 [EF114147]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S4-18-1.2
KF373382 Oceanimonas smirnovii strain 31-13 [NR_042963]
436/447 (98%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2311-S5-01-1.2
KF373383 Pseudoalteromonas lipolytica strain ZR064 [JX173567]
464/465 (99%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
2311-S5-01-2.1
KF373384 Pseudoalteromonas lipolytica strain ZR064 [JX173567]
463/463 (100%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
2311-S5-01-2.2
KF373385 Pseudomonas stutzeri strain UP-1 [AY364327]
453/454 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
2311-S5-01-2.3
KF373386 Pseudomonas stutzeri strain UP-1 [AY364327]
458/459 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
2311-S5-01-3.1.1
KF373387 Brevundimonas diminuta strain c138 [FJ950570]
405/406 (99%)
Alphaproteobacteria; Caulobacterales; Caulobacteraceae; Brevundimonas
2311-S5-01-3.1.2
KF373388 Exiguobacterium arabatum [JF758868]
438/479 (91%)
Firmicutes; Bacilli; Bacillales; Bacillales Family XII. Incertae

P a g e | 130
Sedis; Exiguobacterium
2311-S5-01-3.2
KF373389 Brevundimonas diminuta strain KSC_AK3a [EF191247]
407/407 (100%)
Alphaproteobacteria; Caulobacterales; Caulobacteraceae; Brevundimonas
2311-S5-01B-1
KF373390 Vibrio natriegens strain AUCASVE5 [JQ277719]
472/472 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S5-05-1
KF373391 Vibrio splendidus strain AP625 [GQ254509]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2311-S5-05-2
KF373392 Vibrio splendidus strain AP625 [GQ254509]
470/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio

P a g e | 131
Table A.4: 16S rRNA gene sequence analysis of bacterial cultures from Semporna,
based on BLAST analysis.
Sequence GenBank accession number
Closest match Identities Phylogenetic division
2611-S1-01-1.1
KF373393 Alcaligenes faecalis strain OCEN2DBT [JF264463]
410/465 (88%)
Betaproteobacteria; Burkholderiales; Alcaligenaceae; Alcaligenes
2611-S1-01-1.2
KF373394 Vibrio communis strain F75216 [HQ161743]
472/472 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S1-05-1.1
KF373395
Exiguobacterium lactigenes strain: HYS0503-MK66 [AB259161]
483/483 (100%)
Firmicutes; Bacilli; Bacillales; Bacillales Family XII. Incertae Sedis; Exiguobacterium
2611-S1-05-1.2
KF373396 Oceanimonas smirnovii strain 31-13 [NR_042963]
463/468 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2611-S2-01-1
KF373397 Vibrio natriegens strain CM3 [EU660320]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S2-01-3
KF373398 Vibrio furnissii strain MM5 [FJ906812]
451/473 (95%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S2-05-1.1
KF373399 Allomonas enterica strain JC74, isolate R2A [FR837595]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S2-05-1.2
KF373400 Allomonas enterica strain JC74, isolate R2A [FR837595]
476/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S2-05-2.2
KF373401 Bowmanella denitrificans strain BD1 [NR_043738]
448/459 (98%)
Gammaproteobacteria; Alteromonadales; Alteromonadaceae; Bowmanella
2611-S2-05-3
KF373402 Allomonas enterica strain JC74, isolate R2A [FR837595]
470/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S2-10-2
KF373403 Pseudomonas plecoglossicida strain R8-591-1 [JQ659971]
459/459 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas
2611-S3-01-1
KF373404 Vibrio splendidus strain AP625 [GQ254509]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S3-01-2.2
KF373405 Vibrio alginolyticus strain XHS1-3 [JN188407]
472/472 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S3-05-1
KF373406 Oceanimonas smirnovii strain 31-13 [NR_042963]
449/464 (97%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2611-S4-01-1
KF373407 Vibrio harveyi isolate VHJR19 [DQ995251]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S4-01-2
KF373408 Vibrio rotiferianus strain BV1 [JN391272]
471/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S4- KF373409 Pseudomonas fulva 458/458 Gammaproteobacteria;

P a g e | 132
01-2.1 strain SMA24 [JQ618288]
(100%) Pseudomonadales; Pseudomonadaceae; Pseudomonas
2611-S4-01-2.2
KF373410 Vibrio harveyi isolate VHJR19 [DQ995251]
470/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S4-01-4
KF373411 Pseudidiomarina sediminum strain c121 [NR_044176]
440/461 (95%)
Gammaproteobacteria; Alteromonadales; Idiomarinaceae; Idiomarina
2611-S4-01A-2
KF373412
Pseudomonas pseudoalcaligenes strain K29411 [DQ298030]
460/461 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
2611-S4-01B-1
KF373413 Allomonas enterica strain JC74, isolate R2A [FR837595]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S4-01B-2.1
KF373414 Shewanella sp. UMS11/10 [JQ231163]
428/428 (100%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2611-S4-01B-2.2
KF373415 Oceanimonas smirnovii strain 31-13 [NR_042963]
454/459 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2611-S4-01B-3
KF373416 Exiguobacterium profundum strain SigaKolEp3 [JX987048]
474/476 (99%)
Firmicutes; Bacilli; Bacillales; Bacillales Family XII. Incertae Sedis; Exiguobacterium
2611-S4-01C-1
KF373417 Allomonas enterica strain JC74, isolate R2A [FR837595]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S4-01C-2
KF373418 Allomonas enterica strain JC74, isolate R2A [FR837595]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Allomonas
2611-S4-05-2
KF373419 Bacillus cereus strain 14B [JX901104]
329/433 (76%)
Firmicutes; Bacilli; Bacillales; Bacillaceae; Bacillus
2611-S4-06A-1
KF373420 Shewanella sp. UMS11/10 [JQ231163]
460/468 (98%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2611-S5-01-1
KF373421 Pseudidiomarina sediminum strain c121 [NR_044176]
423/463 (91%)
Gammaproteobacteria; Alteromonadales; Idiomarinaceae; Idiomarina
2611-S5-05A-1
KF373422
Pseudomonas pseudoalcaligenes strain K29411 [DQ298030]
437/438 (99%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
2611-S5-05B-1.1
KF373423 Pseudoalteromonas sp. S187 [FJ457123]
465/466 (99%)
Gammaproteobacteria; Alteromonadales; Pseudoalteromonadaceae; Pseudoalteromonas
2611-S5-05B-1.2
KF373424 Photobacterium sp. MM14 [JN791371]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Photobacterium
2611-S5-05B-3.2.1
KF373425 Shewanella sp. UMS11/10 [JQ231163]
460/465 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2611-S5-05B-3.2.2
KF373426 Shewanella sp. UMS11/10 [JQ231163]
464/465 (99%)
Gammaproteobacteria; Alteromonadales; Shewanellaceae; Shewanella
2611-S5-05C-2
KF373427 Photobacterium sp. MM14 [JN791371]
477/477 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae;

P a g e | 133
Photobacterium
2611-S5-10-2
KF373428 Nitratireductor aquimarinus CL-SC21 [HQ176467]
404/406 (99%)
Alphaproteobacteria; Rhizobiales; Phyllobacteriaceae; Nitratireductor
2611-S6-01-1
KF373429
Pseudomonas pseudoalcaligenes strain K29411 [DQ298030]
347/410 (85%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
2611-S6-01-1.1
KF373430
Pseudomonas pseudoalcaligenes strain K29411 [DQ298030]
450/450 (100%)
Gammaproteobacteria; Pseudomonadales; Pseudomonadaceae; Pseudomonas; Pseudomonas
2611-S6-01-1.2
KF373431 Vibrio campbellii strain CAIM 886 [HM584033]
463/467 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S6-01-3
KF373432 Vibrio alginolyticus strain 486 [JN188409]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S6-05-1.1
KF373433 Oceanimonas smirnovii strain 31-13 [NR_042963]
445/449 (99%)
Gammaproteobacteria; Aeromonadales; Aeromonadaceae; Oceanimonas
2611-S6-05-2
KF373434 Rhodobacter capsulatus strain PSB-06 [FJ866784]
468/468 (100%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Rhodobacter
2611-S6-09-1
KF373435 Vibrio parahaemolyticus strain 448 [JN188417]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S6-09-2
KF373436 Vibrio parahaemolyticus strain 448 [JN188417]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S7-01-1
KF373437 Vibrio parahaemolyticus strain 448 [JN188417]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S7-01-2
KF373438
Vibrio parahaemolyticus strain S9-891-B0919354-5-8F [KC520577]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S8-01-1.1
KF373439 Vibrio alginolyticus strain 486 [JN188409]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
2611-S8-01-3
KF373440 Vibrio communis strain F75216 [HQ161743]
474/474 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio

P a g e | 134
Table A.5: 16S rRNA gene sequence analysis of bacterial cultures from Talang-
talang reef and its surrounding waters, based on BLAST analysis.
Sequence GenBank accession number
Closest match Identities Phylogenetic division
BCM 22-1 KF373441 Vibrio parahaemolyticus strain DHC22 [JQ904733]
451/451 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 22-2 KF373442 Vibrio harveyi strain F75032 [HQ161747]
459/467 (98%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 23 KF373443 Vibrio parahaemolyticus strain DHC22 [JQ904733]
418/418 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 24-1 KF373444 Vibrio harveyi isolate VHJR19 [DQ995251]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 24-2 KF373445 Vibrio harveyi isolate VHJR19 [DQ995251]
472/472 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 25 KF373446 Vibrio harveyi isolate VHJR19 [DQ995251]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 26-1 KF373447 Vibrio parahaemolyticus strain DHC22 [JQ904733]
472/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 26-2 KF373448 Vibrio harveyi isolate VHJR19 [DQ995251]
472/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 27 KF373449 Vibrio harveyi isolate VHJR12 [DQ995245]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 28 KF373450 Vibrio azureus strain M2-164 [JQ810832]
472/473 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 29 KF373451 Halomonas aquamarina strain Ve1-10-83 [EU684464]
460/460 (100%)
Gammaproteobacteria; Oceanospirillales; Halomonadaceae; Halomonas
BCM 31 KF373452 Bacillus arsenicus strain HLSB44 [FJ999563]
455/466 (98%)
Firmicutes; Bacillales; Bacillaceae; Bacillus
BCM 32 KF373453 Bacillus arsenicus strain HLSB44 [FJ999563]
444/468 (95%)
Firmicutes; Bacillales; Bacillaceae; Bacillus
BCM 33 KF373454 Uncultured alpha proteobacterium clone FF-20 [AY682051]
382/384 (99%)
Alphaproteobacteria; environmental samples
BCM 34 KF373455 Psychrobacter celer strain K-W15 [JQ799068]
453/454 (99%)
Gammaproteobacteria; Pseudomonadales; Moraxellaceae; Psychrobacter
BCM 35-1 KF373456 Mucus bacterium 108 [AY654761]
377/406 (93%)
Bacteria
BCM 35-2 KF373457 Uncultured alpha proteobacterium clone FF-20 [AY682051]
382/385 (99%)
Alphaproteobacteria; environmental samples
BCM 36 KF373458 Staphylococcus 474/474 Firmicutes; Bacilli;

P a g e | 135
lugdunensis strain NBL01 [JX629460]
(100%) Bacillales; Staphylococcus
BCM 37 KF373459 Roseobacter sp. H454 [AY368572]
406/406 (100%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Roseobacter
BCM 38 KF373460 Vibrio coralliilyticus strain LMG 21349 [AJ440004]
467/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 39 KF373461 Vibrio coralliilyticus strain LMG 21349 [AJ440004]
472/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 40 KF373462 Vibrio brasiliensis strain HQSB7 [JF721971]
472/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 41 KF373463 Vibrio parahaemolyticus strain DHC22 [JQ904733]
470/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 42 KF373464 Vibrio parahaemolyticus strain DHC22 [JQ904733]
478/480 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 43 KF373465 Klebsiella oxytoca strain AIMST 10.Pl.3 [HQ683968]
458/460 (99%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Klebsiella
BCM 44 KF373466 Microbulbifer sp. 2ta13 [FJ952779]
455/455 (100%)
Gammaproteobacteria; Alteromonadales; Alteromonadaceae; Microbulbifer
BCM 45 KF373467 Vibrio coralliilyticus strain LMG 21349 [AJ440004]
470/479 (98%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 46 KF373468 Vibrio harveyi strain F75087 [HQ161750]
469/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 48 KF373469 Photobacterium jeanii strain R-21419 [GU065212]
471/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Photobacterium
BCM 49 KF373470 Kocuria rosea strain CT22 [EU660350]
442/446 (99%)
Actinobacteria; Actinobacteridae; Actinomycetales; Micrococcineae; Micrococcaceae; Kocuria
BCM 50 KF373471 Brachybacterium paraconglomeratum [AB362255]
441/441 (100%)
Actinobacteria; Actinobacteridae; Actinomycetales; Micrococcineae; Dermabacteraceae; Brachybacterium
BCM 51 KF373472 Kocuria rosea strain CT22 [EU660350]
441/445 (99%)
Actinobacteria; Actinobacteridae; Actinomycetales; Micrococcineae; Micrococcaceae; Kocuria
BCM 52 KF373473 Alteromonadales bacterium fav-2-10-05 [FJ041083]
465/465 (100%)
Gammaproteobacteria; Alteromonadales
BCM 53 KF373474 Sphingobium amiense 397/408 Alphaproteobacteria;

P a g e | 136
strain D3AT58 [JF459959]
(97%) Sphingomonadales; Sphingomonadaceae; Sphingobium
BCM 54 KF373475 Vibrio harveyi isolate VHJR12 [DQ995245]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BCM 56 KF373476 Roseobacter sp. NY93C [EU660505]
407/407 (100%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Roseobacter
BCM 57 KF373477 Klebsiella oxytoca strain AIMST 10.Pl.3 [HQ683968]
463/463 (100%)
Gammaproteobacteria; Enterobacteriales; Enterobacteriaceae; Klebsiella
BCM 58 KF373478 Bacillus firmus strain AIR-NUS-07 [JQ413263]
471/471 (100%)
Firmicutes; Bacillales; Bacillaceae; Bacillus
BCM 59 KF373479 Bacillus sp. WRB-4 [EF636891]
459/473 (97%)
Firmicutes; Bacillales; Bacillaceae; Bacillus
BSD 128-4-1 L
KF373480 Vibrio harveyi [EU373091]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-4-2
KF373481 Vibrio harveyi strain 090212 [GU262992]
477/477 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-5 KF373482 Vibrio harveyi strain A3 [JN391271]
424/428 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-6 KF373483 Vibrio parahaemolyticus isolate Mm007 [FR686999]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-7 L
KF373484 Vibrio communis strain F75216 [HQ161743]
474/474 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-8-1-1
KF373485 Vibrio ponticus strain AN62 [JQ409384]
478/478 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 128-8-1-2 L
KF373486 Vibrio campbellii strain VSD807 [KC534398]
475/476(99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 13 KF373487 Vibrio communis strain P274 [JF836181]
477/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 14 KF373488 Vibrio azureus strain F77118 [HQ908716]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 15 KF373489 Vibrio parahaemolyticus strain 448 [JN188417]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 16-10 KF373490 Bacillus cereus strain B3 [JN252053]
475/475 (100%)
Firmicutes; Bacillales; Bacillaceae; Bacillus; Bacillus cereus group
BSD 16-11 KF373491 Vibrio shiloi [AF007115] 472/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 16-2-1 KF373492 Vibrio communis strain F75214 [HQ161741]
476/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 16-2-2 KF373493 Lysinibacillus fusiformis 476/476 Firmicutes; Bacillales;

P a g e | 137
[JQ897408] (100%) Bacillaceae; Lysinibacillus
BSD 16-3 KF373494 Vibrio ponticus strain AN62 [JQ409384]
476/476 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 16-5 KF373495 Alteromonadales bacterium fav-2-10-05 [FJ041083]
460/460 (100%)
Gammaproteobacteria; Alteromonadales
BSD 16-7 KF373496 Alteromonadales bacterium fav-2-10-05 [FJ041083]
459/459 (100%)
Gammaproteobacteria; Alteromonadales
BSD 16-8 L KF373497 Vibrio communis strain F75214 [HQ161741]
476/477 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 2-10 L KF373498 Vibrio rotiferianus strain HT110622 [JQ792238]
476/476 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 2-6 L KF373499 Vibrio communis strain F75214 [HQ161741]
473/474 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 2-7-1 KF373500 Lysinibacillus fusiformis [JQ897408]
474/474 (100%)
Firmicutes; Bacillales; Bacillaceae; Lysinibacillus
BSD 2-7-2 KF373501 Vibrionaceae bacterium PaD2.06 [GQ406614]
431/433 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae
BSD 2-8 L KF373502 Vibrio parahaemolyticus isolate Mm007 [FR686999]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 2-9-1 KF373503 Vibrio azureus strain HNS029 [JN128263]
472/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 2-9-2 KF373504 Vibrio natriegens strain AUCASVE1 [JQ043186]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 256-5 KF373505 Bacillus anthracis strain: PD7-4 [AB506122]
474/474 (100%)
Firmicutes; Bacillales; Bacillaceae; Bacillus; Bacillus cereus group
BSD 32-5 L KF373506 Vibrio natriegens strain AUCASVE1 [JQ043186]
474/475(99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 32-6-1 KF373507 Vibrio harveyi [EU373091]
476/476 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 32-6-2 KF373508 Ferrimonas sp. A3B-58 [AB193755]
466/469 (99%)
Gammaproteobacteria; Alteromonadales; Ferrimonadaceae; Ferrimonas
BSD 4-4 KF373509 Vibrio harveyi isolate VHJR19 [DQ995251]
477/477 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 4-5 KF373510 Vibrio harveyi isolate VHJR19 [DQ995251]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 4-7 KF373511 Vibrionaceae bacterium PaD2.06 [GQ406614]
433/433 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae
BSD 4-8 L KF373512 Vibrio parahaemolyticus isolate Mm007 [FR686999]
475/475 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 4-9 KF373513 Vibrio harveyi strain S090801 [HM236045]
474/474 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae;

P a g e | 138
Vibrio
BSD 64-1-1 KF373514 Bacillus cereus strain 2 [JX439638]
467/467 (100%)
Firmicutes; Bacillales; Bacillaceae; Bacillus; Bacillus cereus group
BSD 64-1-2 KF373515 Vibrio harveyi strain S090801 [HM236045]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 64-2-1 KF373516 Bacillus anthracis strain: PD7-4 [AB506122]
477/477 (100%)
Firmicutes; Bacillales; Bacillaceae; Bacillus; Bacillus cereus group
BSD 64-2-2 KF373517 Vibrio fortis strain VPMP50 [JQ663920]
477/478 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 8-2 L KF373518 Vibrio harveyi strain BK2 [HM355956]
473/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 8-3 KF373519 Vibrio azureus strain HNS022 [JN128256]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 8-4 KF373520 Vibrio harveyi strain S090801 [HM236045]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSD 8-5 KF373521 Vibrionaceae bacterium PaD2.06 [GQ406614]
432/433 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae
BSD 8-6 L KF373522 Vibrio communis strain F75214 [HQ161741]
471/472 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BSF 11 KF373523 Lysinibacillus boronitolerans [FJ237498]
473/473 (100%)
Firmicutes; Bacillales; Bacillaceae; Lysinibacillus
BSF 12 KF373524 Halomonas sp. 612M-23 [GU371676]
451/452 (99%)
Gammaproteobacteria; Oceanospirillales; Halomonadaceae; Halomonas
BSF 14 KF373525 Halomonas sp. 612M-23 [GU371676]
449/450 (99%)
Gammaproteobacteria; Oceanospirillales; Halomonadaceae; Halomonas
BWC 04-1 KF373526 Rhodobacter capsulatus strain PSB-06 [FJ866784]
463/463 (100%)
Alphaproteobacteria; Rhodobacterales; Rhodobacteraceae; Rhodobacter
BWC 13 KF373527 Halomonas sp. 612M-23 [GU371676]
449/450 (99%)
Gammaproteobacteria; Oceanospirillales; Halomonadaceae; Halomonas
BWC 14 KF373528 Vibrio harveyi strain S090801 [HM236045]
473/473 (100%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BWC 15 KF373529 Alteromonas macleodii [AB238950]
457/457 (100%)
Gammaproteobacteria; Alteromonadales; Alteromonadaceae; Alteromonas
BWC 16 L KF373530 Vibrio harveyi strain HL19 [JQ948038]
474/475 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BWC 17 KF373531 Vibrionaceae bacterium PaD2.06 [GQ406614]
423/424 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae

P a g e | 139
BWC 18 KF373532 Vibrio harveyi strain HL19 [JQ948038]
473/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio
BWC 19 L KF373533 Vibrio harveyi strain HL19 [JQ948038]
475/476 (99%)
Gammaproteobacteria; Vibrionales; Vibrionaceae; Vibrio

P a g e | 140
Table A.6: Screening of antimicrobial (PKS/NRPS) and DMSP-degrading (dmdA/dddP) genes in coral mucus isolates.
Samples Presence of genes
PKS NRPS dmdA dddP
BCM 22-1 + + - - BCM 22-2 - + + - BCM 23 + + - + BCM 24-1 + + - + BCM 24-2 + + + + BCM 25 + + + + BCM 26-1 - + - + BCM 26-2 - + - + BCM 27 + + - - BCM 28 - - + + BCM 29 + + + - BCM 31 + - - + BCM 32 - - + - BCM 33 - - - + BCM 34 - + - - BCM 35-1 - + + - BCM 35-2 - + - - BCM 36 - - - - BCM 37 - + - + BCM 38 + - - + BCM 39 + - - - BCM 40 - + - - BCM 41 - + + - BCM 42 - + - + BCM 43 - + - + BCM 44 - + - - BCM 45 - + - - BCM 46 - + + - BCM 48 - - + + BCM 49 - + - - BCM 50 - - - - BCM 51 - - - - BCM 52 - + - - BCM 53 - + + + BCM 54 - - - - BCM 56 - + + + BCM 57 - + + + BCM 58 - + + - BCM 59 - - + +
“+” indicates positive presence of the genes, “-“ indicates negative presence of genes.
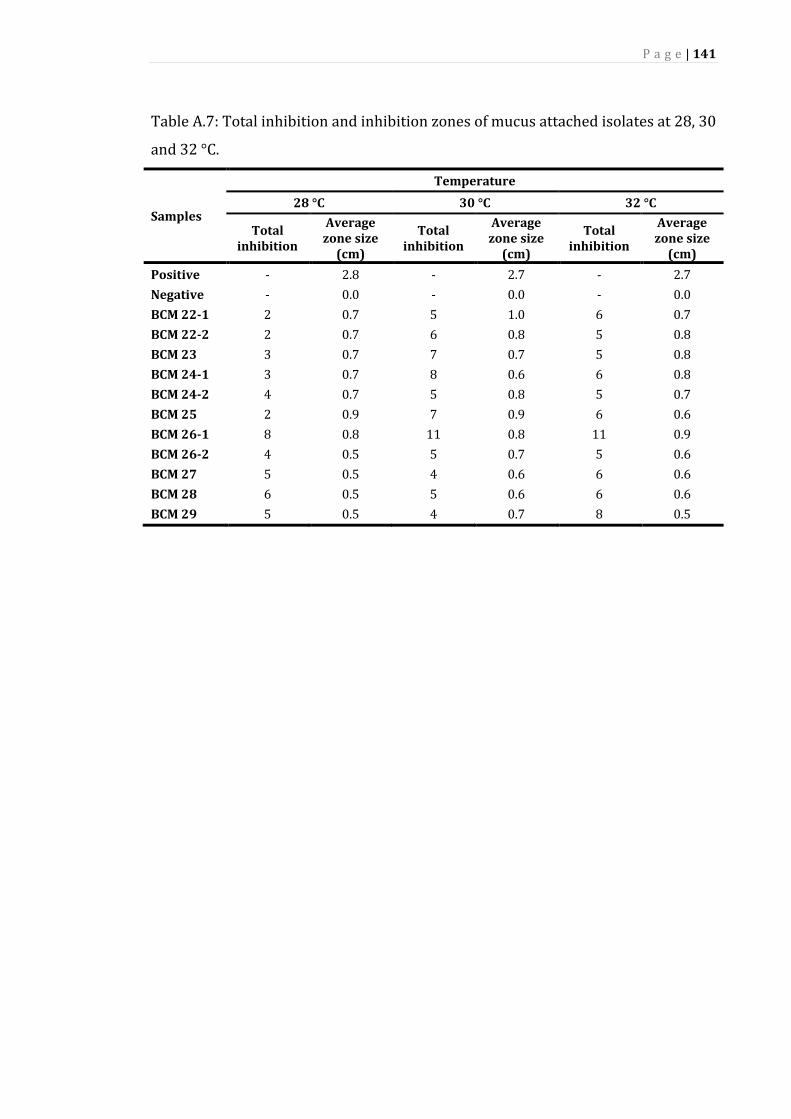
P a g e | 141
Table A.7: Total inhibition and inhibition zones of mucus attached isolates at 28, 30
and 32 °C.
Samples
Temperature
28 °C 30 °C 32 °C
Total inhibition
Average zone size
(cm)
Total inhibition
Average zone size
(cm)
Total inhibition
Average zone size
(cm)
Positive - 2.8 - 2.7 - 2.7
Negative - 0.0 - 0.0 - 0.0
BCM 22-1 2 0.7 5 1.0 6 0.7
BCM 22-2 2 0.7 6 0.8 5 0.8
BCM 23 3 0.7 7 0.7 5 0.8
BCM 24-1 3 0.7 8 0.6 6 0.8
BCM 24-2 4 0.7 5 0.8 5 0.7
BCM 25 2 0.9 7 0.9 6 0.6
BCM 26-1 8 0.8 11 0.8 11 0.9
BCM 26-2 4 0.5 5 0.7 5 0.6
BCM 27 5 0.5 4 0.6 6 0.6
BCM 28 6 0.5 5 0.6 6 0.6
BCM 29 5 0.5 4 0.7 8 0.5

P a g e | 142
Table A.8: Total inhibition and inhibition zones of mucus associated isolates at 28,
30 and 32 °C.
Samples
Temperature
28 °C 30 °C 32 °C
Total inhibition
Average zone size
(cm)
Total inhibition
Average zone size
(cm)
Total inhibition
Average zone size
(cm)
Positive - 2.7 - 2.6 - 2.7
Negative - 0.0 - 0.0 - 0.0
BCM 31 5 0.5 3 0.9 6 0.6
BCM 32 9 0.8 9 0.8 11 0.9
BCM 33 7 0.7 5 0.9 4 0.4
BCM 34 8 0.7 5 0.5 4 0.4
BCM 35-1 5 0.7 8 0.7 3 0.3
BCM 35-2 5 0.7 5 1.0 4 0.6
BCM 36 6 0.7 6 0.9 3 0.5
BCM 37 6 0.8 8 0.8 4 0.4
BCM 38 10 0.9 11 0.9 9 0.8
BCM 39 4 1.0 5 0.4 3 0.7
BCM 40 5 0.8 5 0.5 3 0.4
BCM 41 5 0.9 6 0.6 4 0.6
BCM 42 4 0.6 6 0.5 2 0.3
BCM 43 5 0.9 7 0.5 4 0.5
BCM 44 6 0.8 5 0.4 2 0.4
BCM 45 6 0.9 9 0.8 9 0.8
BCM 46 4 0.7 6 0.7 4 0.6
BCM 48 5 0.6 7 0.6 4 0.5
BCM 49 4 0.8 6 0.7 3 0.7
BCM 50 7 0.6 7 0.6 2 0.9
BCM 51 7 0.6 8 0.6 2 0.7
BCM 52 6 0.7 7 0.7 4 0.5
BCM 53 10 0.9 10 0.8 13 0.9
BCM 54 0 0.0 2 0.9 0 0.0
BCM 56 1 0.7 1 0.6 0 0.0
BCM 57 1 0.8 1 0.9 0 0.0
BCM 58 0 0.0 3 0.4 0 0.0
BCM 59 0 0.0 2 0.9 0 0.0