co-conjugating chondroitin-6-sulfate/dermatan sulfate to chitosan scaffold alters chondrocyte gene...
TRANSCRIPT

ARTICLE
Co-Conjugating Chondroitin-6-Sulfate/DermatanSulfate to Chitosan Scaffold Alters ChondrocyteGene Expression and Signaling Profiles
Yen-Lin Chen,1,2 Huang-Chi Chen,1 Hing-Yuen Chan,2 Ching-Kuang Chuang,1
Yu-Han Chang,3 Yu-Chen Hu1
1Department of Chemical Engineering, National Tsing Hua University, Hsinchu, Taiwan,
300; telephone: 886-3-571-8245; fax: 886-3-571-5408; e-mail: [email protected] Industry Research and Development Institute, Hsinchu, Taiwan, 3003Department of Orthopaedic, Chang Gung Memorial Hospital, Kueishan, Taiwan
Received 21 February 2008; revision received 10 April 2008; accepted 18 April 2008
Published online 2 May 2008 in Wiley InterScience (www.interscience.wiley.com). DO
I 10.1002/bit.21953ABSTRACT: Co-conjugating chondroitin-6-sulfate (CSC)and dermatan sulfate (DS) to chitosan scaffolds improveschondrocyte differentiation and extracellular matrix (ECM)production. To further elucidate the cellular responses toCSC/DS conjugation, gene expression profiles for the ratchondrocytes cultured on the CSC/DS/chitosan and chit-osan-only scaffolds were compared by reverse-transcriptionPCR (RT-PCR) and quantitative real-time RT-PCR (qRT-PCR). Our data unraveled that the CSC/DS/chitosan scaf-fold resulted in low-level expression of collagen I, IIA and Xand potentiated the aggrecan, collagen II (including collagenIIB) and TIMP3 expression, but downregulated the decorinexpression. Therefore CSC/DS/chitosan scaffold maintainedthe chondrocyte differentiation while minimized de-differ-entiation and hypertrophy. Furthermore, CSC/DS conjuga-tion affected the expression of 11 genes implicated in 9signaling pathways (as unveiled by cDNA microarray) andupregulated the expression of TGF-b1, Sox9, BMP2, PTHrPand Ihh (as confirmed by qRT-PCR). These data suggestedthat the CSC/DS/chitosan scaffold potentiated the TGF-band Hedgehog pathways, which activated the expression ofPTHrP and its downstream Sox9. The signals were trans-duced to elevate the expression of aggrecan, collagen II andTIMP3, and contributed to the well-differentiated chondro-cyte phenotype. Altogether, this study for the first timeelucidated the roles of GAGs-conjugated biomaterials inmatrix production and breakdown, cellular differentiationand signal transduction at the molecular levels.
Biotechnol. Bioeng. 2008;101: 821–830.
� 2008 Wiley Periodicals, Inc.
KEYWORDS: chondrocyte; chitosan; chondroitin sulfate;dermatan sulfate; gene expression profile; signaling path-way
Correspondence to: Y.-C. Hu
� 2008 Wiley Periodicals, Inc.
Introduction
Healthy articular cartilage consists of chondrocytesembedded in the extracellular matrix (ECM) in whichchondrocytes synthesize matrix molecules such as collagenII and glycosaminoglycans (GAGs). The GAGs, includingchondroitin-4-sulfate (CSA), chondroitin-6-sulfate (CSC)and dermatan sulfate (DS), are present in diverse forms ofproteoglycans. The GAGs not only help maintain thestructural integrity of the cartilage, but also dictate celladhesion, migration, proliferation and differentiation (Plaaset al., 1997). Thus, scaffolds fabricated with GAG moleculeshave been developed to promote cell differentiation andtissue formation (Chang et al., 2003; Chupa et al., 2000;Denuziere et al., 1998; Fan et al., 2006; Seo et al., 2006).
To develop a scaffold biomaterial better mimicking thenatural cartilage ECM, we have employed FractionalFactorial Design approach to rationally fabricate chitosanmembranes co-conjugated with 4 different GAGs (CSA,CSC, DS and heparin), and unveiled that the GAGscomposition profoundly influences the chondrocyte mor-phology, proliferation and ECM production in the 2Dculture (Chen et al., 2006b). Furthermore, we have designed3D chitosan scaffolds co-conjugated with CSC and DS withthe aid of Response Surface Methodology, which reveals theoptimal CSC/DS formulation (2.80 mg CSC and 0.01 mgDS conjugated to each chitosan scaffold) that enhancesECM production and maintains chondrocyte morphologyand chondrocytic gene expression (Chen et al., 2007).Theoretically, conjugating a proper amount of CSC/DS tothe scaffold provides a biomimetic microenvironment thattransmits extracellular stimuli to the cultured chondrocytes,and initiates a cascade of signal transduction pathways thatelevate the gene expression required to maintain cellulardifferentiation. However, how the CSC/DS modulates thecellular gene expression and what signaling pathways and
Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008 821

cellular events are activated by CSC/DS have yet to beidentified.
To answer these questions, in this study the primary ratchondrocytes were cultured on the chitosan scaffolds with orwithout CSC/DS conjugation and the time-course profilesof cell expansion, GAG and collagen production wereinvestigated. Furthermore, the expression profiles of genesassociated with ECM production and degradation wereexamined by reverse transcription-PCR (RT-PCR) andverified by quantitative real-time RT-PCR (qRT-PCR). Theexpression of genes linked to a variety of signaling pathwaysand chondrocyte differentiation was also profiled by cDNAmicroarray and confirmed by qRT-PCR. Our data revealedthat co-conjugating CSC/DS to chitosan scaffold modulatedthe expression of specific signaling molecules and triggered acascade of cellular events in chondrocytes, which playedpivotal roles in chondrocyte differentiation and formationof cartilaginous tissues.
Materials and Methods
Reagents
Chitosan (85% deacetylated, MW� 200,000) from crabshells, CSC (sodium salt, MW� 100,000) from sharkcartilage, DS (sodium salt, MW� 60,000) from porcineintestinal mucosa, EDC (1-ethyl-3-(3-dimethylaminopro-pyl) carbodiimide) and all other reagents were purchasedfrom Sigma unless otherwise noted. N-hydroxysuccinimide(NHS) was purchased from Merck.
Preparation of Scaffolds
The chitosan scaffolds were fabricated by lyophilizationmethod and co-conjugated with CSC/DS by EDC/NHScross-linkage based on the optimal formulation determinedpreviously (Chen et al., 2007). Briefly, 1% chitosan solution(15 ml) in a 60 mm Petri dish was lyophilized to form a sheetof porous scaffold. The scaffolds (5 mm in diameter, 3 mmin thickness, �2.5� 0.5 mg) were cut from the sheet andtransferred to 96-well plates. For each scaffold, 50 mlof HEPES buffer (pH 7.4) containing 2.8 mg of CSC and0.01 mg of DS was added. In parallel, 50 ml HEPES bufferwas added for the preparation of chitosan-only scaffolds.The cross-linking reactions were initiated by adding 150 mlEDC/NHS solution to each scaffold and continued byshaking for 72 h at 48C. The scaffolds were washed 5 timeswith phosphate buffered-saline (PBS, pH 7.4) to removeresidual GAGs and lyophilized. The resultant scaffolds weresterilized by immersing in 70% ethanol for 24 h prior to use.
Isolation and Culture of Articular Chondrocytes
Chondrocytes were isolated from the articular cartilages of7-day old Wistar rats by 0.2% collagenase II digestion (Lee
822 Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008
et al., 2007). The cells were cultured using Dulbecco’sminimal Essential medium (DMEM) containing 10% fetalbovine serum (FBS), 100 U/ml penicillin, 100 mg/mlstreptomycin and 0.25 mg/ml amphotericin B, and passaged2 times (P2) prior to use.
Measurement of Cell Proliferation and ECM Production
The P2 cells were seeded onto the scaffolds (2� 106 cells/scaffold) in 24-well plates and incubated for 1 day. The cell/scaffold constructs were transferred to 6-well plates andcultured with shaking (10 rpm) at 378C. The medium wasexchanged three times a week. At day 0, 1, 10, 14 and 21, thecell/scaffold constructs were collected, washed, lyophilizedand digested with papain for 24 h at 608C for DNA assay anddetermination of corresponding cell numbers as described(Chen et al., 2004). The expansion ratio was calculated bycomparing the cell number at the respective time point withthe cell number at day 0.
Moreover, the aliquots of digested samples were subjectedto acid hydrolysis (6 N HCl for 12 h at 1008C). Totalamounts of collagen were determined in triplicate spectro-photometrically after reaction with chloramines-T andp-dimethylaminobenzaldehyde (Woessner, 1961). Theabsorbance was measured at 550 nm and the collagen con-centrations were calculated using a hydroxyproline tocollagen ratio of 1:9 (Schwartz et al., 1985). The amounts ofGAGs in the constructs were assayed in triplicate by mixingthe papain-digested samples with 1, 9-dimethylmethyleneblue and reading the absorbance at 525 nm (Farndale et al.,1986). In parallel, blank CSC/DS/chitosan and chitosanscaffolds were incubated with DMEM containing 10% FBSfor 1, 10, 14 or 21 days (with 3 medium changes everyweek), followed by GAGs and collagen measurements. Theamounts of GAGs (collagen) that were produced by the cellswere obtained by subtracting the amounts of GAGs(collagen) in the experimental constructs by those in theblank scaffolds. The GAGs (collagen) production levels werecalculated by comparing the GAGs (collagen) amounts ateach time point with those at day 0.
RT-PCR and qRT-PCR Analyses
The constructs were ground to powder in liquid nitrogenand total RNA was extracted using RNeasy Mini Kit(Qiagen). The primer sequences and PCR conditions usedfor RT-PCR and qRT-PCR are summarized in Tables Iand II, respectively. For RT-PCR, approximately 0.5 mgof total RNA was used as the template for cDNA synthesisand subsequent PCR using AccuPower RT/PCR PreMix kit(Bioneer). The PCR products were subjected to 2% agarosegel electrophoresis.
For qRT-PCR, 1 mg of total RNA was used for cDNAsynthesis by ImProm-IITM reverse transcription system(Promega). The qRT-PCR reactions were performed usingABI 7700 real-time PCR system (Applied Biosystems) with
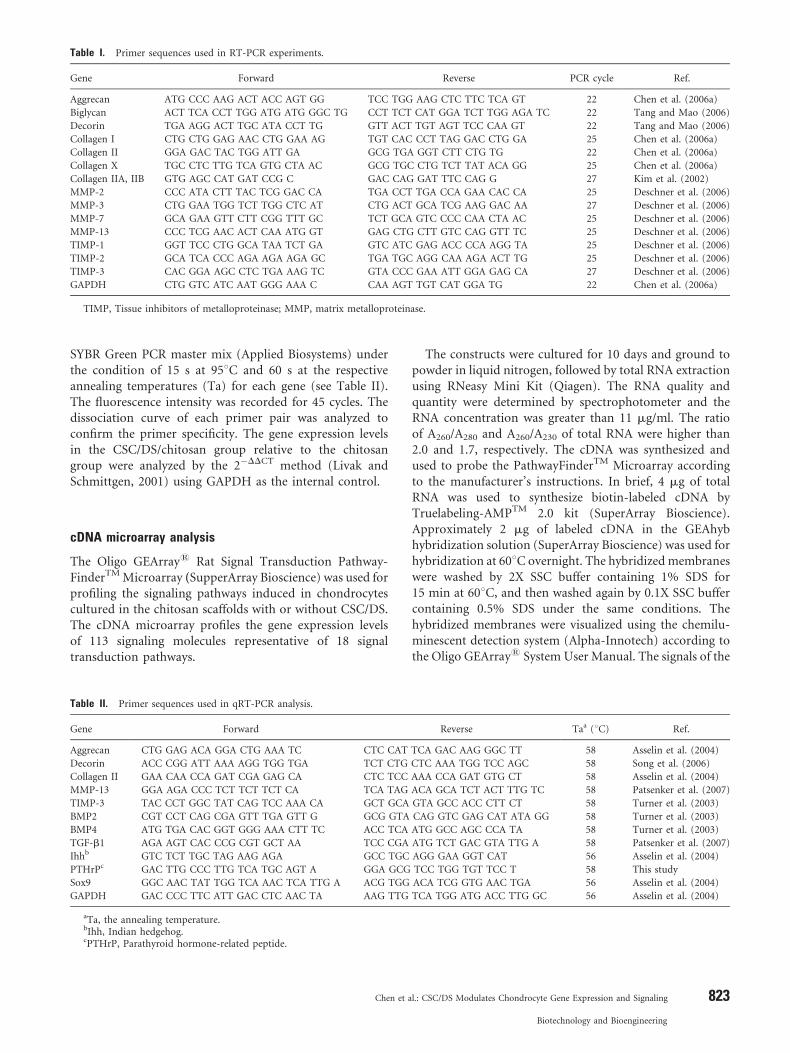
Table I. Primer sequences used in RT-PCR experiments.
Gene Forward Reverse PCR cycle Ref.
Aggrecan ATG CCC AAG ACT ACC AGT GG TCC TGG AAG CTC TTC TCA GT 22 Chen et al. (2006a)
Biglycan ACT TCA CCT TGG ATG ATG GGC TG CCT TCT CAT GGA TCT TGG AGA TC 22 Tang and Mao (2006)
Decorin TGA AGG ACT TGC ATA CCT TG GTT ACT TGT AGT TCC CAA GT 22 Tang and Mao (2006)
Collagen I CTG CTG GAG AAC CTG GAA AG TGT CAC CCT TAG GAC CTG GA 25 Chen et al. (2006a)
Collagen II GGA GAC TAC TGG ATT GA GCG TGA GGT CTT CTG TG 22 Chen et al. (2006a)
Collagen X TGC CTC TTG TCA GTG CTA AC GCG TGC CTG TCT TAT ACA GG 25 Chen et al. (2006a)
Collagen IIA, IIB GTG AGC CAT GAT CCG C GAC CAG GAT TTC CAG G 27 Kim et al. (2002)
MMP-2 CCC ATA CTT TAC TCG GAC CA TGA CCT TGA CCA GAA CAC CA 25 Deschner et al. (2006)
MMP-3 CTG GAA TGG TCT TGG CTC AT CTG ACT GCA TCG AAG GAC AA 27 Deschner et al. (2006)
MMP-7 GCA GAA GTT CTT CGG TTT GC TCT GCA GTC CCC CAA CTA AC 25 Deschner et al. (2006)
MMP-13 CCC TCG AAC ACT CAA ATG GT GAG CTG CTT GTC CAG GTT TC 25 Deschner et al. (2006)
TIMP-1 GGT TCC CTG GCA TAA TCT GA GTC ATC GAG ACC CCA AGG TA 25 Deschner et al. (2006)
TIMP-2 GCA TCA CCC AGA AGA AGA GC TGA TGC AGG CAA AGA ACT TG 25 Deschner et al. (2006)
TIMP-3 CAC GGA AGC CTC TGA AAG TC GTA CCC GAA ATT GGA GAG CA 27 Deschner et al. (2006)
GAPDH CTG GTC ATC AAT GGG AAA C CAA AGT TGT CAT GGA TG 22 Chen et al. (2006a)
TIMP, Tissue inhibitors of metalloproteinase; MMP, matrix metalloproteinase.
SYBR Green PCR master mix (Applied Biosystems) underthe condition of 15 s at 958C and 60 s at the respectiveannealing temperatures (Ta) for each gene (see Table II).The fluorescence intensity was recorded for 45 cycles. Thedissociation curve of each primer pair was analyzed toconfirm the primer specificity. The gene expression levelsin the CSC/DS/chitosan group relative to the chitosangroup were analyzed by the 2�DDCT method (Livak andSchmittgen, 2001) using GAPDH as the internal control.
cDNA microarray analysis
The Oligo GEArray1 Rat Signal Transduction Pathway-FinderTM Microarray (SupperArray Bioscience) was used forprofiling the signaling pathways induced in chondrocytescultured in the chitosan scaffolds with or without CSC/DS.The cDNA microarray profiles the gene expression levelsof 113 signaling molecules representative of 18 signaltransduction pathways.
Table II. Primer sequences used in qRT-PCR analysis.
Gene Forward
Aggrecan CTG GAG ACA GGA CTG AAA TC CTC CAT
Decorin ACC CGG ATT AAA AGG TGG TGA TCT CTG
Collagen II GAA CAA CCA GAT CGA GAG CA CTC TCC
MMP-13 GGA AGA CCC TCT TCT TCT CA TCA TAG
TIMP-3 TAC CCT GGC TAT CAG TCC AAA CA GCT GCA
BMP2 CGT CCT CAG CGA GTT TGA GTT G GCG GTA
BMP4 ATG TGA CAC GGT GGG AAA CTT TC ACC TCA
TGF-b1 AGA AGT CAC CCG CGT GCT AA TCC CGA
Ihhb GTC TCT TGC TAG AAG AGA GCC TGC
PTHrPc GAC TTG CCC TTG TCA TGC AGT A GGA GCG
Sox9 GGC AAC TAT TGG TCA AAC TCA TTG A ACG TGG
GAPDH GAC CCC TTC ATT GAC CTC AAC TA AAG TTG
aTa, the annealing temperature.bIhh, Indian hedgehog.cPTHrP, Parathyroid hormone-related peptide.
Chen et a
The constructs were cultured for 10 days and ground topowder in liquid nitrogen, followed by total RNA extractionusing RNeasy Mini Kit (Qiagen). The RNA quality andquantity were determined by spectrophotometer and theRNA concentration was greater than 11 mg/ml. The ratioof A260/A280 and A260/A230 of total RNA were higher than2.0 and 1.7, respectively. The cDNA was synthesized andused to probe the PathwayFinderTM Microarray accordingto the manufacturer’s instructions. In brief, 4 mg of totalRNA was used to synthesize biotin-labeled cDNA byTruelabeling-AMPTM 2.0 kit (SuperArray Bioscience).Approximately 2 mg of labeled cDNA in the GEAhybhybridization solution (SuperArray Bioscience) was used forhybridization at 608C overnight. The hybridized membraneswere washed by 2X SSC buffer containing 1% SDS for15 min at 608C, and then washed again by 0.1X SSC buffercontaining 0.5% SDS under the same conditions. Thehybridized membranes were visualized using the chemilu-minescent detection system (Alpha-Innotech) according tothe Oligo GEArray1 System User Manual. The signals of the
Reverse Taa (8C) Ref.
TCA GAC AAG GGC TT 58 Asselin et al. (2004)
CTC AAA TGG TCC AGC 58 Song et al. (2006)
AAA CCA GAT GTG CT 58 Asselin et al. (2004)
ACA GCA TCT ACT TTG TC 58 Patsenker et al. (2007)
GTA GCC ACC CTT CT 58 Turner et al. (2003)
CAG GTC GAG CAT ATA GG 58 Turner et al. (2003)
ATG GCC AGC CCA TA 58 Turner et al. (2003)
ATG TCT GAC GTA TTG A 58 Patsenker et al. (2007)
AGG GAA GGT CAT 56 Asselin et al. (2004)
TCC TGG TGT TCC T 58 This study
ACA TCG GTG AAC TGA 56 Asselin et al. (2004)
TCA TGG ATG ACC TTG GC 56 Asselin et al. (2004)
l.: CSC/DS Modulates Chondrocyte Gene Expression and Signaling 823
Biotechnology and Bioengineering

hybridized spots were recorded, analyzed by GEArrayExpression Analysis Suite (SuperArray Bioscience) andnormalized against those of GAPDH.
Statistical analysis
Data were analyzed using Student’s t-test. All dataare expressed as means� standard deviations (SD) of 3 or4 independent experiments, and P< 0.05 was consideredsignificant.
Figure 1. Time-course profile of (a) cell expansion ratio, (b) GAGs production
and (c) collagen production. The P2 chondrocytes (2� 106 cells/scaffold) were seeded
to and cultured in the CSC/DS/chitosan or chitosan scaffolds and the constructs (n¼ 3
in each experiment) were harvested at the indicated times for analysis. The data
represent the mean� SD of 4 independent experiments and the mean values are
shown above the bars. �, P< 0.05.
Results
The Effect of CSC/DS on Cell Proliferation and ECMProduction Over Time
We have previously shown that culturing chondrocytes onchitosan scaffolds co-conjugated with CSC/DS resulted inheightened production of GAGs and collagen after 21-dayculture (Chen et al., 2007), yet whether the scaffoldcomposition continued to influence the chondrocytebehavior (e.g. cell proliferation and GAG production)throughout the culture period was unknown. To gain moreinsight on how the CSC/DS-associated stimulation variedwith time, the chondrocytes (2� 106 cells/scaffold) wereseeded to and cultured in the chitosan scaffolds with orwithout CSC/DS. The cell expansion ratio as well as GAGsand collagen production were quantified after 1-, 10-, 14-and 21-day culture. Figure 1a depicts that the expansionratios for the cells cultured in the chitosan and CSC/DS/chitosan scaffolds increased slowly, but steadily, with timeand at day 21 (the endpoint of the experiment) reached�1.6- and 2.0-fold, respectively. No significant differencebetween the two groups arose until day 21.
For both groups, GAGs were rapidly synthesized fromday 1 to day 10 and continued to accumulate until day 21(Fig. 1b). Notably, the CSC/DS/chitosan group producedsignificantly ( P< 0.05) more GAGs (91.2� 21.7 mg/106
cells) than the chitosan group (51.0� 17.3 mg/106 cells) at asearly as day 10. At day 21, the CSC/DS/chitosan group(126.6� 25.3 mg/106 cells) resulted in a �62% increase inGAGs production compared with the chitosan group(78.0� 27.3 mg/106 cells). Concomitantly, the collagenproduction in both groups (Fig. 1c) occurred prior to day 10and continued throughout the culture, hence at day 21the CSC/DS/chitosan group (76.0� 5.7 mg/106 cells)produced �17% more collagen than the chitosan group(64.6� 3.9 mg/106 cells). However, the difference in collagenproduction between both groups was not significant( P> 0.05) until day 14 ( P< 0.05). These data revealedthat chondrocytes cultured on the CSC/DS/chitosanscaffolds remained capable of proliferation and ECMproduction throughout the culture period, and confirmedour previous findings that the co-conjugated CSC/DSelevated the GAGs and collagen production.
824 Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008
The Effect of CSC/DS on ECM-related Gene Expression
How the co-conjugated CSC/DS impacted the ECMsynthesis and degradation was explored by analyzingthe expression of relevant genes by RT-PCR at days 10and 21 (Fig. 2). Three genes involved in proteoglycans(aggrecan, biglycan and decorin) synthesis and 3 genesresponsible for collagen (collagen I, II and X) productionwere detected. The genes involved in regulating the ECMdegradation encoded 4 matrix metalloproteinases (MMP-2,

Figure 2. Comparison of ECM-related gene expression by RT-PCR. The chondrocytes were cultured in the CSC/DS/chitosan or chitosan scaffolds (n¼ 3 for each time point in
each experiment) as in Figure 1 and the gene expression was analyzed at days 10 and 21 by RT-PCR. Representative data from 3 independent experiments are shown. All
experiments were performed using GAPDH as the control.
Figure 3. Quantitative comparison of ECM-related gene expression by qRT-PCR.
The chondrocytes were cultured in the CSC/DS/chitosan or chitosan scaffolds for 10
days (n¼ 3 for each experiment). The gene expression levels in the CSC/DS/chitosan
group relative to the chitosan group were analyzed by the 2�DDCT method using
GAPDH as the internal control. The numbers above the bar represent the percent
change of gene expression (%). The data represent the mean�SD of 3 independent
culture experiments.�, P< 0.05.
-3, -7 and -13) and 3 tissue inhibitors of metalloproteinase(TIMP-1, -2 and -3). To evaluate the chondrocytedifferentiation, collagen IIA and IIB, the two spliced formsof collagen II predominantly expressed in the chondropro-genitor cells and differentiated chondrocytes, respectively(Kim et al., 2002), were analyzed.
Figure 2 shows that the gene expression patterns wereconsistent at days 10 and 21. Active expression of ECMsynthesis genes (e.g. aggrecan and collagen II) and ECMdegradation genes (e.g. MMP-2, -3 and -13) were observed,suggesting that the cartilaginous constructs in both groupswere undergoing matrix remodeling. Also cell de-differ-entiation and hypertrophy were not prominent in bothgroups as judged by the barely detectable collagen I and X.When comparing both groups, the expression levels ofaggrecan, collagen II (including collagen IIB), MMP-13and TIMP-3 were higher in the CSC/DS/chitosan groupthan in the chitosan group. In contrast, the expression ofdecorin was suppressed in the CSC/DS/chitosan group whencompared with the chitosan group. These data indicatedthat co-conjugating CSC/DS to chitosan scaffold resulted indifferential expression of certain genes responsible for ECMsynthesis and degradation. However, the conjugation ofCSC/DS to chitosan scaffold did not further affect theexpression of other genes (biglycan, collagen I, collagen X,MMP-2, -3, -7 and TIMP-1, -2) during the culture.
To further confirm and quantify the altered geneexpression induced by co-conjugating CSC/DS to chitosan
Chen et a
scaffold, the expression levels of collagen II, aggrecan,decorin, TIMP-3 and MMP-13 were measured by qRT-PCR(Fig. 3). Since the gene expression patterns were identical atboth day 10 and day 21 (Fig. 2), only the samples harvestedat day 10 were analyzed. Figure 3 depicts that the CSC/DS/
l.: CSC/DS Modulates Chondrocyte Gene Expression and Signaling 825
Biotechnology and Bioengineering

chitosan group led to significantly ( p<0.05) higher collagenII (�91% increase) and aggrecan (�312% increase)expression, but significantly lower (�55% decrease) decorinexpression when compared with the chitosan group.Conversely, the CSC/DS/chitosan group gave rise to a46% increase ( P< 0.05) in TIMP-3 expression as opposedto the chitosan group, implying that CSC/DS conjugationcould inhibit the ECM degradation. On the other hand,the CSC/DS/chitosan group only resulted in a 33% increase( P> 0.05) in MMP-13 expression in comparison with thechitosan group, which indicated that CSC/DS conjugationdid not significantly alter the MMP-13 activity.
The Effect of CSC/DS on Signal Transduction Pathways(cDNA Microarray Analysis)
How the co-conjugation of CSC/DS to chitosan scaffoldaffected the signal transduction in rat chondrocytes wasexamined by PathwayFinderTM cDNA microarray, whichprofiles the expression of 113 signaling molecules repre-sentative of 18 signal transduction pathways. The cells werecultured as described above for 10 days and the mRNA wasextracted for microarray analysis to compare the geneexpression in the CSC/DS/chitosan group and the chitosangroup. Only expression levels increasing or decreasing morethan 20% were considered significant.
Figure 4 illustrates that augmenting CSC/DS to chitosanscaffolds altered the expression levels of 11 signaling
Figure 4. Differential gene expression in the CSC/DS/chitosan and chitosan
groups. The chondrocytes were cultured as described in Figure 3 for 10 days and the
gene expression was analyzed by PathwayFinderTM cDNA microarray. The genes with
expression levels increasing or decreasing more than 20% are indicated by the
arrows.
826 Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008
molecules (as indicated by arrows) that are implicated in9 different pathways including Hedgehog, Wnt, PI3 kinase/AKT, TGF-b, p53, Androgen, Estrogen, Retinoic acid andStress pathways (Table III). Among the 11 signalingmolecules, 7 genes (BMP2, Cdkn1a, Ctsd, Egfr, Fn1, Hspb1and Hspca) were upregulated in the CSC/DS/chitosangroup, while the other 4 (BMP4, Ccdn 1, Cdkn2a and Wsb1)were downregulated (Table III).
The Effect of CSC/DS on BMP2, TGF-b1, Ihh, PTHrPand Sox9 Expression
The microarray data unveiled the altered expression levels ofgenes involved in the Hedgehog (BMP2, BMP4 and Wsb1)and TGF-b (Cdkn1a and Cdkn2a) pathways. Since bothpathways are associated with chondrocyte proliferation anddifferentiation (Kaps et al., 2006; Minina et al., 2001), weinferred that the Hedgehog and TGF-b pathways playedimportant roles in modulating the ECM accumulation in theCSC/DS/chitosan group. To attest this hypothesis, theexpression of selected genes implicated in or activated bythese pathways was measured by qRT-PCR.
Compared with the chitosan group, Figure 5 delineatesthat the CSC/DS/chitosan group led to significantly( P< 0.05) higher expression of TGF-b1, Sox9, BMP2,PTHrP (parathyroid hormone-related peptide) and Ihh(Indian hedgehog) (�77%, 113%, 174%, 33% and 1139%increases, respectively) but lower expression of BMP4(�55% decrease). These data not only confirmed thefindings of cDNA microarray (Table III), but furtherindicated that the CSC/DS conjugation regulated theexpression of TGF-b1, Ihh, PTHrP and Sox9.
Discussion
Although it is well recognized that scaffold componentsgovern the cell proliferation, migration and differentiation,and are crucial for tissue engineering, to date most studiesstriving to develop cartilage tissue engineering scaffoldsmainly focus on assessing the effects of materials onchondrocyte phenotype (e.g. cell proliferation, morphologyand ECM production) and the expression of a few cartilage-specific genes (Chang et al., 2003; Chen et al., 2006b; Donatiet al., 2005; Holland et al., 2007; Mouw et al., 2005).However, the underlying molecular mechanisms dictatingthe cell-biomaterial interactions remain poorly understoodand little is known about how the scaffold materials triggerthe signaling cascades that ultimately guide the chondrocytedifferentiation and contribute to enhanced ECM produc-tion. To address these questions, a few recent studies havestarted to explore the molecular basis of cell-biomaterialinteractions by transcriptomic or proteomic methods (forreview see (Gallagher et al., 2006)). In the context of cartilageengineering, however, only Kaps et al. have examined thegene expression profiling of (human) articular chondrocytes
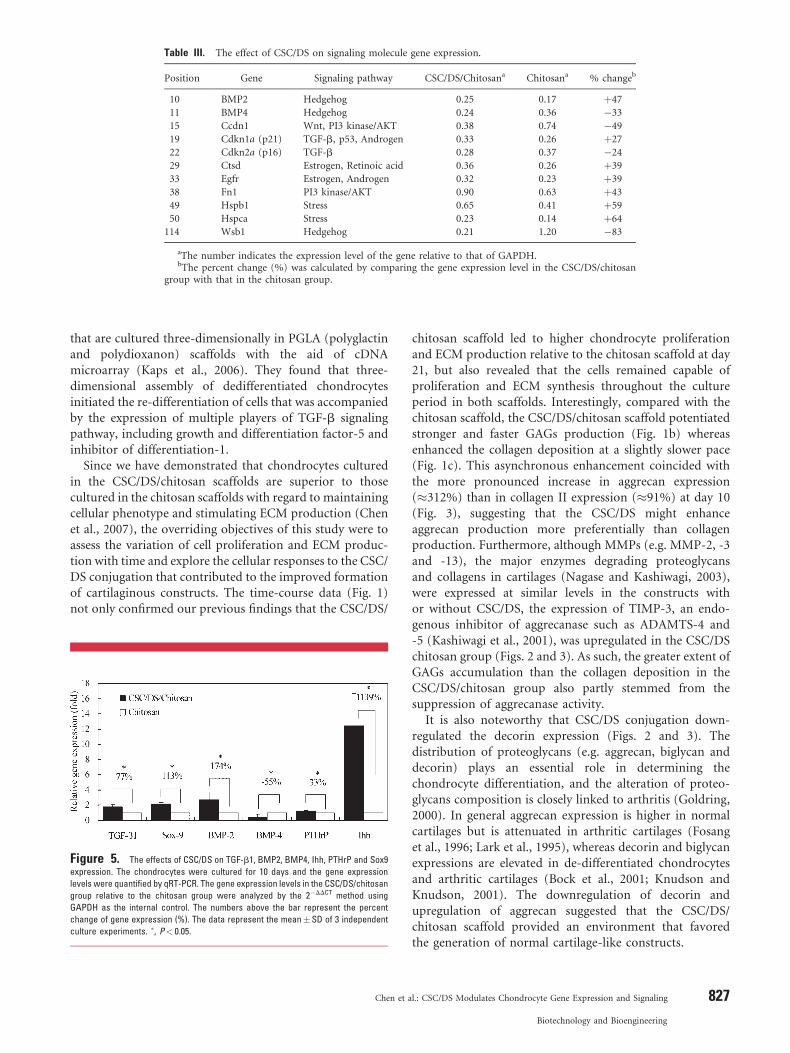
Table III. The effect of CSC/DS on signaling molecule gene expression.
Position Gene Signaling pathway CSC/DS/Chitosana Chitosana % changeb
10 BMP2 Hedgehog 0.25 0.17 þ47
11 BMP4 Hedgehog 0.24 0.36 �33
15 Ccdn1 Wnt, PI3 kinase/AKT 0.38 0.74 �49
19 Cdkn1a (p21) TGF-b, p53, Androgen 0.33 0.26 þ27
22 Cdkn2a (p16) TGF-b 0.28 0.37 �24
29 Ctsd Estrogen, Retinoic acid 0.36 0.26 þ39
33 Egfr Estrogen, Androgen 0.32 0.23 þ39
38 Fn1 PI3 kinase/AKT 0.90 0.63 þ43
49 Hspb1 Stress 0.65 0.41 þ59
50 Hspca Stress 0.23 0.14 þ64
114 Wsb1 Hedgehog 0.21 1.20 �83
aThe number indicates the expression level of the gene relative to that of GAPDH.bThe percent change (%) was calculated by comparing the gene expression level in the CSC/DS/chitosan
group with that in the chitosan group.
that are cultured three-dimensionally in PGLA (polyglactinand polydioxanon) scaffolds with the aid of cDNAmicroarray (Kaps et al., 2006). They found that three-dimensional assembly of dedifferentiated chondrocytesinitiated the re-differentiation of cells that was accompaniedby the expression of multiple players of TGF-b signalingpathway, including growth and differentiation factor-5 andinhibitor of differentiation-1.
Since we have demonstrated that chondrocytes culturedin the CSC/DS/chitosan scaffolds are superior to thosecultured in the chitosan scaffolds with regard to maintainingcellular phenotype and stimulating ECM production (Chenet al., 2007), the overriding objectives of this study were toassess the variation of cell proliferation and ECM produc-tion with time and explore the cellular responses to the CSC/DS conjugation that contributed to the improved formationof cartilaginous constructs. The time-course data (Fig. 1)not only confirmed our previous findings that the CSC/DS/
Figure 5. The effects of CSC/DS on TGF-b1, BMP2, BMP4, Ihh, PTHrP and Sox9
expression. The chondrocytes were cultured for 10 days and the gene expression
levels were quantified by qRT-PCR. The gene expression levels in the CSC/DS/chitosan
group relative to the chitosan group were analyzed by the 2�DDCT method using
GAPDH as the internal control. The numbers above the bar represent the percent
change of gene expression (%). The data represent the mean�SD of 3 independent
culture experiments. �, P< 0.05.
Chen et a
chitosan scaffold led to higher chondrocyte proliferationand ECM production relative to the chitosan scaffold at day21, but also revealed that the cells remained capable ofproliferation and ECM synthesis throughout the cultureperiod in both scaffolds. Interestingly, compared with thechitosan scaffold, the CSC/DS/chitosan scaffold potentiatedstronger and faster GAGs production (Fig. 1b) whereasenhanced the collagen deposition at a slightly slower pace(Fig. 1c). This asynchronous enhancement coincided withthe more pronounced increase in aggrecan expression(�312%) than in collagen II expression (�91%) at day 10(Fig. 3), suggesting that the CSC/DS might enhanceaggrecan production more preferentially than collagenproduction. Furthermore, although MMPs (e.g. MMP-2, -3and -13), the major enzymes degrading proteoglycansand collagens in cartilages (Nagase and Kashiwagi, 2003),were expressed at similar levels in the constructs withor without CSC/DS, the expression of TIMP-3, an endo-genous inhibitor of aggrecanase such as ADAMTS-4 and-5 (Kashiwagi et al., 2001), was upregulated in the CSC/DSchitosan group (Figs. 2 and 3). As such, the greater extent ofGAGs accumulation than the collagen deposition in theCSC/DS/chitosan group also partly stemmed from thesuppression of aggrecanase activity.
It is also noteworthy that CSC/DS conjugation down-regulated the decorin expression (Figs. 2 and 3). Thedistribution of proteoglycans (e.g. aggrecan, biglycan anddecorin) plays an essential role in determining thechondrocyte differentiation, and the alteration of proteo-glycans composition is closely linked to arthritis (Goldring,2000). In general aggrecan expression is higher in normalcartilages but is attenuated in arthritic cartilages (Fosanget al., 1996; Lark et al., 1995), whereas decorin and biglycanexpressions are elevated in de-differentiated chondrocytesand arthritic cartilages (Bock et al., 2001; Knudson andKnudson, 2001). The downregulation of decorin andupregulation of aggrecan suggested that the CSC/DS/chitosan scaffold provided an environment that favoredthe generation of normal cartilage-like constructs.
l.: CSC/DS Modulates Chondrocyte Gene Expression and Signaling 827
Biotechnology and Bioengineering

Figure 6. The putative model summarizing how CSC/DS conjugation modulated
the expression of signaling molecules and genes involved in ECM production. ": up-
regulation; #: down-regulation.
Augmenting CSC/DS to chitosan scaffolds also alteredthe expression levels of signaling molecules implicated in9 signal transduction pathways (Table III and Fig. 4), amongwhich TGF-b and Hedgehog pathways are known toprofoundly influence chondrogenesis, chondrocyte differ-entiation and extracellular matrix (Furumatsu et al., 2005;Grimaud et al., 2002; Minina et al., 2001). TGF-b super-family signaling can be classified into the TGF-b branch(represented by ligands such as TGF-b that activates Smad2and 3) and the BMP branch (represented by ligands such asBMP2 that activates Smad1, 5 and 8) (Herpin andCunningham, 2007). Of note, TGF-b signaling results inthe upregulation of PTHrP (Serra et al., 1999), TIMP-3 (Suet al., 1996), Sox9 (Huang et al., 2001), and subsequentstimulation of downstream aggrecan and collagen IIexpression. TGF-b signaling also leads to the down-regulation of decorin expression (Grimaud et al., 2002).The outcome of TGF-b signaling concurred with theenhanced expression of TGF-b1, Sox9 (Fig. 5), aggrecan,collagen II and TIMP-3, and suppressed expression ofdecorin (Figs. 2 and 3) by the chondrocytes cultured in theCSC/DS/chitosan scaffolds. The correlation suggests thatCSC/DS conjugation enhanced the expression of TGF-b1,which potentiated the TGF-b pathway and upregulated theexpression of secondary signaling molecule PTHrP. Thesignal was transduced to upregulate the master transcriptionfactor Sox9 (Huang et al., 2001), and subsequently relayed toboost the expression of aggrecan, collagen II and TIMP3.
In addition, the CSC/DS conjugation elevated theexpression of BMP2 and Ihh (Fig. 5). BMP and Ihh caninduce each other’s expression (Minina et al., 2002; Seki andHata, 2004) and both can interact to regulate chondrocytedevelopment, proliferation (Minina et al., 2002) and thenpromote the aggrecan and collagen II expression in thechondrocytes (Grunder et al., 2004; Li et al., 2004). Ihh is animportant signaling molecule in the Hedgehog pathway andupregulates PTHrP expression (Long et al., 2001). PTHrP,in turn, elevates the expression of Sox9 (Huang et al., 2001)which enhances the aggrecan and collagen production (Tewet al., 2005; Tsuchiya et al., 2003). Therefore, CSC/DS alsoactivated the Hedgehog pathway and potentiated down-stream signaling cascade.
Intriguingly, although the CSC/DS conjugation upregu-lated the expression of BMP2, a growth factor that couldprovoke chondrocyte hypertrophy as marked by collagen Xexpression, the collagen X expression remained barelydetectable in the CSC/DS/chitosan group (Fig. 2). The lowdegree of hypertrophy was probably due to the co-activationof not only BMP2, but also TGF-b1, Ihh and PTHrP byCSC/DS conjugation. TGF-b1 mitigates the function ofBMP2 in activating collagen X expression (Hanada et al.,2001), while Ihh upregulates PTHrP expression which alsodelays the onset of hypertrophy (Long et al., 2001).
On the other hand, the CSC/DS conjugation inhibitedBMP4 expression (Table III and Fig. 5). BMP2 and BMP4belong to the BMPs family and their relative expression levelis closely associated with chondrogenesis (Shum et al.,
828 Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008
2003). Contradictory to BMP2 expression which is elevatedin mature chondrocytes, BMP4 expression is attenuated asthe progenitor chondrocytes differentiate towards matureor pre-hypertrophic chondrocytes (Goldring et al., 2006).Additionally, the chondrocytes cultured in the CSC/DS/chitosan scaffolds expressed predominantly collagen IIB(the collagen II highly expressed in mature chondrocytes)as opposed to collagen IIA (the collagen II expressed inchondroprogenitor cells). In combination with the elevatedexpression of PTHrP and Sox9, which suppress chondro-cyte differentiation into hypertrophy (Goldring et al.,2006), these data collectively suggest that chondrocytescultured in the CSC/DS/chitosan scaffold maintained a well-differentiated status with minimal hypertrophy and terminaldifferentiation. The signaling molecules that are activated byCSC/DS conjugation are summarized in Figure 6.
In summary, hereby we confirmed that co-conjugatingCSC/DS to chitosan scaffolds for chondrocyte culture gaverise to simultaneous regulation of ECM synthesis anddegradation as well as maintenance of chondrocyte differ-entiation with minimal hypertrophy and de-differentiation,which rendered the resultant cartilage-like constructs morefavorably resembling normal cartilages. Intriguingly, CSC/DS conjugation upregulated the endogenous expression ofBMP2 and TGF-b1. Since supplementing these two factorsto chondrocytes favors the chondrocytic differentiationand ECM production (Hanada et al., 2001; Lee et al., 2004;Sung et al., 2007), conjugating CSC/DS to chitosanscaffolds offers an attractive approach to enhancing co-expression of endogenous BMP2 and TGF-b1, which mayenable sustained stimulation of chondrocytic differentia-tion. Furthermore, CSC/DS conjugation potentiated theactivation of at least TGF-b and Hedgehog pathways.Although it remains unknown whether these two signaling

pathways act sequentially or in parallel, and whether cross-talk between the pathways exists, they can orchestrate thetransduction of signals to activate PTHrP and Sox9 andsubsequently modulate the downstream genes that con-tribute to chondrocyte differentiation and matrix remodel-ing. To our best knowledge, this is the first report that shedlight on the roles of GAGs conjugated to biomaterials inECM production, cellular differentiation and signal trans-duction at the molecular levels. The data further confirmedthat CSC/DS/chitosan scaffolds are suitable for chondrocyteculture in the context of cartilage tissue engineering. Thedetailed information concerning chondrocytic gene expres-sion influenced by the scaffold materials will pave the way tounderstanding the molecular mechanisms contributing tothe chondroinductive effects imparted by the scaffoldmaterials, and will eventually lead to improved design ofbiomimetic scaffolds.
The author gratefully acknowledges the financial support from the
National Tsing Hua University Booster Program (96N2511E1) and
CGMH-NTHU Joint Research Program (CGTH96-T8 and
CMRPG361041), Taiwan.
References
Asselin A, Hattar S, Oboeuf M, Greenspan D, Berdal A, Sautier JM. 2004.
The modulation of tissue-specific gene expression in rat nasal chon-
drocyte cultures by bioactive glasses. Biomaterials 25:5621–5630.
Bock HC, Michaeli P, Bode C, Schultz W, Kresse H, Herken R, Miosge N.
2001. The small proteoglycans decorin and biglycan in human articular
cartilage of late-stage osteoarthritis. Osteoarthritis Cartilage 9:654–663.
Chang CH, Liu HC, Lin CC, Chou CH, Lin FH. 2003. Gelatin-chondroitin-
hyaluronan tri-copolymer scaffold for cartilage tissue engineering.
Biomaterials 24:4853–4858.
Chen H-C, Lee H-P, Sung M-L, Liao C-J, Hu Y-C. 2004. A novel rotating-
shaft bioreactor for two-phase cultivation of tissue-engineered carti-
lage. Biotechnol Prog 20:1802–1809.
Chen H-C, Lee H-P, Ho Y-C, Sung M-L, Hu Y-C. 2006a. Combination of
baculovirus-mediated gene transfer and rotating-shaft bioreactor for
cartilage tissue engineering. Biomaterials 27:3154–3162.
Chen Y-L, Chen H-C, Lee H-P, Chan H-Y, Hu Y-C. 2006b. Rational
development of GAG-augmented chitosan membranes by fractional
factorial design methodology. Biomaterials 27:2222–2232.
Chen Y-L, Lee H-P, Chan H-Y, Sung L-Y, Chen H-C, Hu Y-C. 2007.
Composite chondroitin-6-sulfate/dermatan sulfate/chitosan scaffolds
for cartilage tissue engineering. Biomaterials 28:2294–2305.
Chupa JM, Foster AM, Sumner SR, Madihally SV, Matthew HW. 2000.
Vascular cell responses to polysaccharide materials: in vitro and in vivo
evaluations. Biomaterials 21:2315–2322.
Denuziere A, Ferrier D, Damour O, Domard A. 1998. Chitosan-chondroitin
sulfate and chitosan-hyaluronate polyelectrolyte complexes: biological
properties. Biomaterials 19:1275–1285.
Deschner J, Rath-Deschner B, Agarwal S. 2006. Regulation of matrix
metalloproteinase expression by dynamic tensile strain in rat fibro-
chondrocytes. Osteoarthritis Cartilage 14:264–272.
Donati I, Stredanska S, Silvestrini G, Vetere A, Marcon P, Marsich E,
Mozetic P, Gamini A, Paoletti S, Vittur F. 2005. The aggregation of pig
articular chondrocyte and synthesis of extracellular matrix by a lactose-
modified chitosan. Biomaterials 26:987–998.
Fan H, Hu Y, Qin L, Li X, Wu H, Lv R. 2006. Porous gelatin-chondroitin-
hyaluronate tri-copolymer scaffold containing microspheres loaded
with TGF-beta1 induces differentiation of mesenchymal stem cells
Chen et a
in vivo for enhancing cartilage repair. J Biomed Mater Res A
77:785–794.
Farndale RW, Buttle DJ, Barrett AJ. 1986. Improved quantitation and
discrimination of sulphated glycosaminoglycans by use of dimethyl-
methylene blue. Biochim Biophys Acta 883:173–177.
Fosang AJ, Last K, Maciewicz RA. 1996. Aggrecan is degraded by matrix
metalloproteinases in human arthritis. Evidence that matrix metallo-
proteinase and aggrecanase activities can be independent. J Clin Invest
98:2292–2299.
Furumatsu T, Tsuda M, Taniguchi N, Tajima Y, Asahara H. 2005. Smad3
induces chondrogenesis through the activation of SOX9 via CREB-
binding protein/p300 recruitment. J Biol Chem 280:8343–8350.
Gallagher WM, Lynch I, Allen LT, Miller I, Penney SC, O’Connor DP,
Pennington S, Keenan AK, Dawson KA. 2006. Molecular basis of cell-
biomaterial interaction: Insights gained from transcriptomic and pro-
teomic studies. Biomaterials 27:5871–5882.
Goldring MB. 2000. The role of the chondrocyte in osteoarthritis. Arthritis
Rheum 43:1916–1926.
Goldring MB, Tsuchimochi K, Ijiri K. 2006. The control of chondrogenesis.
J Cell Biochem 97:33–44.
Grimaud E, Heymann D, Redini F. 2002. Recent advances in TGF-b effects
on chondrocyte metabolism: Potential therapeutic roles of TGF-b in
cartilage disorders. Cytokine Growth Factor Rev 13:241–257.
Grunder T, Gaissmaier C, Fritz J, Stoop R, Hortschansky P, Mollenhauer J,
Aicher WK. 2004. Bone morphogenetic protein (BMP)-2 enhances the
expression of type II collagen and aggrecan in chondrocytes embedded
in alginate beads. Osteoarthritis Cartilage 12:559–567.
Hanada K, Solchaga LA, Caplan AI, Hering TM, Goldberg VM, Yoo JU,
Johnstone B. 2001. BMP-2 induction and TGF-b1 modulation of rat
periosteal cell chondrogenesis. J Cell Biochem 81:284–294.
Herpin A, Cunningham C. 2007. Cross-talk between the bone morphoge-
netic protein pathway and other major signaling pathways results in
tightly regulated cell-specific outcomes. FEBS J 274:2977–2985.
Holland TA, Bodde EW, Cuijpers VM, Baggett LS, Tabata Y, Mikos AG,
Jansen JA. 2007. Degradable hydrogel scaffolds for in vivo delivery of
single and dual growth factors in cartilage repair. Osteoarthritis Car-
tilage 15:187–197.
Huang W, Chung UI, Kronenberg HM, de Crombrugghe B. 2001. The
chondrogenic transcription factor Sox9 is a target of signaling by the
parathyroid hormone-related peptide in the growth plate of endochon-
dral bones. Proc Natl Acad Sci U S A 98:160–165.
Kaps C, Frauenschuh S, Endres M, Ringe J, Haisch A, Lauber J, Buer J,
Krenn V, Haupl T, Burmester GR, Sittinger M. 2006. Gene expression
profiling of human articular cartilage grafts generated by tissue engi-
neering. Biomaterials 27:3617–3630.
Kashiwagi M, Tortorella M, Nagase H, Brew K. 2001. TIMP-3 is a potent
inhibitor of aggrecanase 1 (ADAM-TS4) and aggrecanase 2 (ADAM-
TS5). J Biol Chem 276:12501–12504.
Kim TK, Park JS, Lee MC, Seong SC, Kim HJ. 2002. Alternative splicing of type
II procollagen gene in the dedifferentiation of rat epiphyseal chondrocytes
serially cultured in monolayer. Connect Tissue Res 43:56–62.
Knudson CB, Knudson W. 2001. Cartilage proteoglycans. Semin Cell Dev
Biol 12:69–78.
Lark MW, Bayne EK, Lohmander LS. 1995. Aggrecan degradation in osteoar-
thritis and rheumatoid arthritis. Acta Orthop Scand Suppl 266:92–97.
Lee JE, Kim KE, Kwon IC, Ahn HJ, Lee SH, Cho H, Kim HJ, Seong SC, Lee
MC. 2004. Effects of the controlled-released TGF-beta 1 from chitosan
microspheres on chondrocytes cultured in a collagen/chitosan/glyco-
saminoglycan scaffold. Biomaterials 25:4163–4173.
Lee H-P, Chen Y-L, Shen H-C, Lo W-H, Ho Y-C, Hu Y-C. 2007. Bacu-
lovirus transduction of rat articular chondrocytes: Roles of cell cycle.
J Gene Med 9:33–43.
Li J, Yoon ST, Hutton WC. 2004. Effect of bone morphogenetic protein-2
(BMP-2) on matrix production, other BMPs, and BMP receptors in rat
intervertebral disc cells. J Spinal Disord Tech 17:423–428.
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data
using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.
Methods (San Diego, Calif) 25:402–408.
l.: CSC/DS Modulates Chondrocyte Gene Expression and Signaling 829
Biotechnology and Bioengineering

Long F, Zhang XM, Karp S, Yang Y, McMahon AP. 2001. Genetic
manipulation of hedgehog signaling in the endochondral skeleton
reveals a direct role in the regulation of chondrocyte proliferation.
Development 128:5099–5108.
Minina E, Wenzel HM, Kreschel C, Karp S, Gaffield W, McMahon AP,
Vortkamp A. 2001. BMP and Ihh/PTHrP signaling interact to coordi-
nate chondrocyte proliferation and differentiation. Development
128:4523–4534.
Minina E, Kreschel C, Naski MC, Ornitz DM, Vortkamp A. 2002. Inter-
action of FGF, lhh/Pthlh, and BMP signaling integrates chondrocyte
proliferation and hypertrophic differentiation. Developmental Cell
3:439–449.
Mouw JK, Case ND, Guldberg RE, Plaas AH, Levenston ME. 2005. Varia-
tions in matrix composition and GAG fine structure among scaffolds
for cartilage tissue engineering. Osteoarthritis Cartilage 13:828–836.
Nagase H, Kashiwagi M. 2003. Aggrecanases and cartilage matrix degrada-
tion. Arthritis Res Ther 5:94–103.
Patsenker E, Popov Y, Wiesner M, Goodman SL, Schuppan D. 2007.
Pharmacological inhibition of the vitronectin receptor abrogates
PDGF-BB-induced hepatic stellate cell migration and activation in
vitro. J Hepatol 46:878–887.
Plaas AH, Wong-Palms S, Roughley PJ, Midura RJ, Hascall VC. 1997.
Chemical and immunological assay of the nonreducing terminal
residues of chondroitin sulfate from human aggrecan. J Biol Chem
272:20603–20610.
Schwartz DE, Choi Y, Sandell LJ, Hanson WR. 1985. Quantitative analysis
of collagen, protein and DNA in fixed, paraffin-embedded and sec-
tioned tissue. Histochem J 17:655–663.
Seki K, Hata A. 2004. Indian hedgehog gene is a target of the bone
morphogenetic protein signaling pathway. J Biol Chem 279:18544–
18549.
Seo SJ, Choi YJ, Akaike T, Higuchi A, Cho CS. 2006. Alginate/galactosylated
chitosan/heparin scaffold as a new synthetic extracellular matrix for
hepatocytes. Tissue Eng 12:33–44.
830 Biotechnology and Bioengineering, Vol. 101, No. 4, November 1, 2008
Serra R, Karaplis A, Sohn P. 1999. Parathyroid hormone-related peptide
(PTHrP)-dependent and -independent effects of transforming growth
factor beta (TGF-beta) on endochondral bone formation. J Cell Biol
145:783–794.
Shum L, Wang X, Kane AA, Nuckolls GH. 2003. BMP4 promotes chon-
drocyte proliferation and hypertrophy in the endochondral cranial
base. Int J Dev Biol 47:423–431.
Song SJ, Hutmacher D, Nurcombe V, Cool SM. 2006. Temporal expression
of proteoglycans in the rat limb during bone healing. Gene 379:92–
100.
Su S, Dehnade F, Zafarullah M. 1996. Regulation of tissue inhibitor of
metalloproteinases-3 gene expression by transforming growth factor-b
and dexamethasone in bovine and human articular chondrocytes. DNA
Cell Biol 15:1039–1048.
Sung L-Y, Lo W-H, Chiu H-Y, Chen H-C, Chuang C-K, Lee H-P, Hu Y-C.
2007. Modulation of chondrocyte phenotype via baculovirus-mediated
growth factor expression. Biomaterials 28:3437–3447.
Tang M, Mao JJ. 2006. Matrix and gene expression in the rat cranial base
growth plate. Cell Tissue Res 324:467–474.
Tew SR, Li Y, Pothacharoen P, Tweats LM, Hawkins RE, Hardingham TE.
2005. Retroviral transduction with SOX9 enhances re-expression of the
chondrocyte phenotype in passaged osteoarthritic human articular
chondrocytes. Osteoarthritis Cartilage 13:80–89.
Tsuchiya H, Kitoh H, Sugiura F, Ishiguro N. 2003. Chondrogenesis
enhanced by overexpression of sox9 gene in mouse bone marrow-
derived mesenchymal stem cells. Biochem Biophys Res Commun
301:338–343.
Turner KJ, McIntyre BS, Phillips SL, Barlow NJ, Bowman CJ, Foster PM.
2003. Altered gene expression during rat Wolffian duct development in
response to in utero exposure to the antiandrogen linuron. Toxicol Sci
74:114–128.
Woessner JF Jr. 1961. The determination of hydroxyproline in tissue and
protein samples containing small proportions of this imino acid. Arch
Biochem Biophys 93:440–447.