clustering of sodium channels at the neuromuscular junction
TRANSCRIPT

Clustering of Sodium Channelsat the Neuromuscular JunctionJ.H. CALDWELLDepartment of Cellular and Structural Biology and the Department of Physiology and Biophysics, University of Colorado HealthSciences Center, Denver, Colorado 80262
KEY WORDS skeletal muscle; agrin; syntrophin; ankyrin
ABSTRACT Voltage-gated sodium channels (NaChs) are highly concentrated in the postsyn-aptic region of the neuromuscular junction, especially in the depths of postsynaptic folds and in theperijunctional region. The formation of the high NaCh density occurs during synapse maturation,approximately 2 weeks after initial synaptic contact in the rodent. The concentration of NaChs andtheir localization in the troughs of the folds increase the safety factor for neuromuscular transmis-sion by reducing the threshold for initiation of the action potential. There is evidence that agrinplays a role in the formation of NaCh aggregation. Molecules such as ankyrin and syntrophin thatbind NaChs may be important for maintenance of the high channel density at the endplate.Microsc. Res. Tech. 49:84–89, 2000. © 2000 Wiley-Liss, Inc.
INTRODUCTIONThe creation of a synapse requires a complicated or-
chestration of pre- and postsynaptic specializations. Thisis especially obvious for neurons where excitatory andinhibitory synapses can be adjacent to one another; thepostsynaptic neuron manages to concentrate the appro-priate neurotransmitter receptors at high density be-neath the nerve terminals. The neuromuscular junctionhas been utilized as a model synapse, and many labora-tories have focused upon the high density of acetylcholinereceptors (AChRs) in the postsynaptic membrane. Othermolecules are concentrated at the neuromuscular junc-tion, and one of these is the voltage-gated sodium channel(NaCh). The NaCh concentration at the mammalian neu-romuscular junction rivals the node of Ranvier in density(several thousand per mm2). This review will focus uponwhere NaChs are localized in the vicinity of the endplatemembrane, when the high density develops, why NaChsare concentrated in the postsynaptic membrane, and, fi-nally, what molecules are involved in creating and main-taining this high density.
WHERE ARE NaChs CONCENTRATED?The idea that NaChs are enriched at the neuro-
muscular junction was first suggested over two de-cades ago. Microelectrode recordings showed that therate of rise of the action potential was significantlygreater near the endplate than in extrajunctionalmuscle membrane (Nastuk and Alexander, 1973;Thesleff et al., 1974) . The presence of a high densityof NaChs at the endplate was confirmed by electro-physiological recordings (Beam et al., 1985; Betz etal., 1984; Caldwell et al., 1986; Roberts, 1987), im-munocytochemistry (Haimovich et al., 1987), andfluorescently labeled toxin binding (Angelides, 1986).Ultrastructural studies have revealed that NaChsline the postsynaptic troughs and are absent fromthe crests of the folds where AChRs are concentrated(Flucher and Daniels, 1989).
Employment of the loose-patch voltage clamp tech-nique allowed sampling of the NaCh density at severaldifferent points on individual muscle cells. With thistechnique, the density of NaChs in the postsynapticmembrane was estimated to be approximately 2,000NaChs/mm2 (Caldwell et a., 1986; Milton et al., 1992),close to the density measured at the node of Ranvier.Channel density in the extrajunctional membrane was100–200 NaChs/mm
2, implying a 10–20-fold concentra-
tion of NaChs at the endplate. The density of NaChsdecreases away from the endplate as a smooth gradientover 50–200 mm of the perijunctional membrane (Cald-well and Milton, 1988; Lupa et al., 1993) and is de-creased further near the tendon (Fig. 1). Similar re-sults have been reported for mouse, rat, and snaketwitch muscles. A perijunctional increase has also beenshown in human muscle (Ruff and Whittlesey, 1992).Several interesting differences in NaCh density anddistribution have been noted between various muscletypes. The NaCh density at endplates of mammalianslow-twitch muscle is consistently only about half thatfound at synapses of fast-twitch muscle, and the peri-junctional density is also reduced (Milton et al., 1992,Ruff, 1992).
The distribution of NaChs in the postsynaptic mem-brane is nonuniform and is complementary to that ofAChRs. Flucher and Daniels (1989) used immunoelec-tronmicroscopy to show that AChRs (and the associ-ated protein rapsyn) are located on the crests of thepostsynaptic folds while NaChs (and its associated pro-tein ankyrin) are found in the troughs of the folds.Radiolabeled scorpion toxin and electron microscopywere used to confirm that sodium channels are in thetroughs (Boudier et al., 1992). Thus, the postsynapticmembrane is divided into separate and, to a first ap-
*Correspondence to: Dr. John H. Caldwell, Department of Cellular and Struc-tural Biology, Campus Box B-111, UCHSC, 4200 E. Ninth Ave., Denver, CO80262.
Received 3 September 1999; Accepted in revised form 15 November 1999
MICROSCOPY RESEARCH AND TECHNIQUE 49:84–89 (2000)
© 2000 WILEY-LISS, INC.

proximation, non-overlapping domains. This is illus-trated schematically in Figure 2. The difference in lo-calization is one indication that some of the moleculesimportant for creating and maintaining the high AChRdensity will be different from the molecules requiredfor NaChs.
WHEN DOES THE HIGH CONCENTRATIONDEVELOP AT THE NEUROMUSCULAR
JUNCTION?Neuromuscular junctions begin to form at embryonic
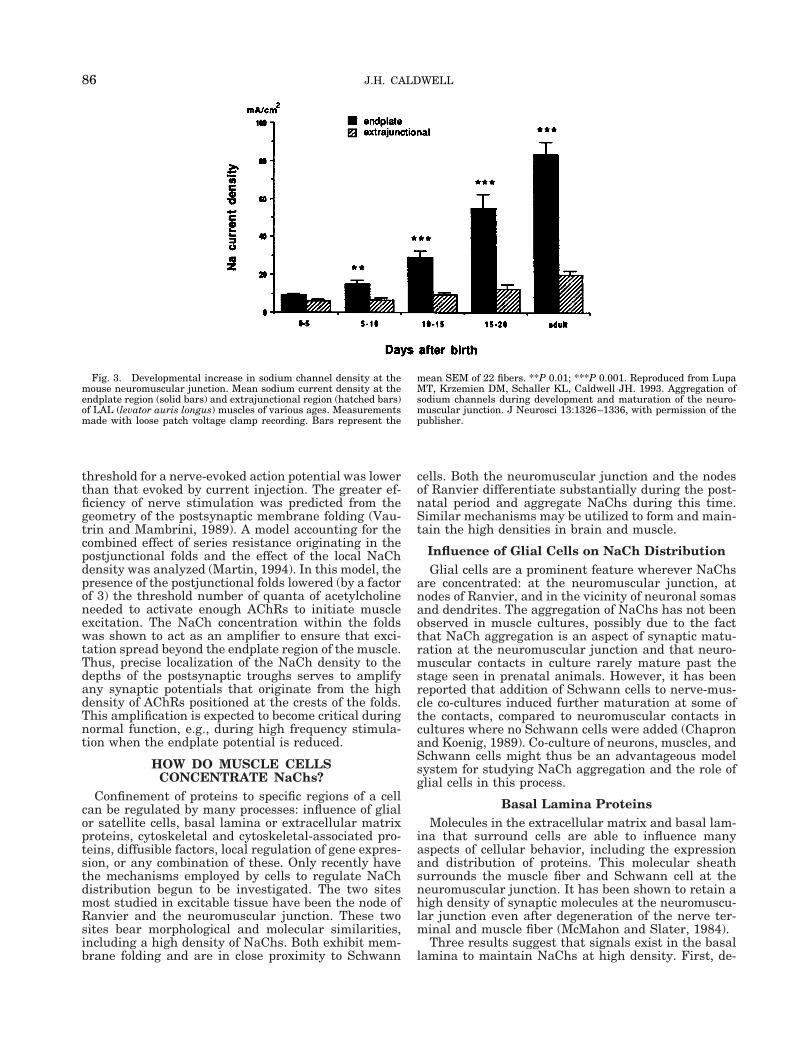
day 15 in the rodent, which is about a week beforebirth. Acetylcholine receptors begin to aggregate al-most immediately following nerve contact. In contrast,electrophysiological recordings (Lupa et al., 1993) indi-cated that NaChs start aggregating at the endplateabout 1–2 weeks after birth (Fig. 3). Immunocytochem-ical measurements (Wood and Slater, 1998) showedthat NaChs begin to be concentrated in the perijunc-tional region about the time of birth, with the mainaccumulation 1–2 weeks after birth. The period ofNaCh concentration coincides with the formation ofpostsynaptic junctional folds. Thus, the clustering ofNaChs begins about 2 weeks after the aggregation ofAChRs and is associated with synapse maturationrather than synapse formation. This difference in tim-ing reinforces the concept (described above) that somefactors regulating the concentration of AChRs andNaChs are different.
WHY ARE NaChs CONCENTRATED IN THEJUNCTIONAL AND PERIJUNCTIONAL
MEMBRANE?It seems reasonable to assume that there is a func-
tion subserved by clustering NaChs in the subsynapticand perijunctional membrane. The most likely physio-logical role for sodium channel aggregation is to supplysufficient current to initiate an action potential. Al-though the safety factor for neuromuscular transmis-sion is large for a single stimulus (a “twitch”), thesafety factor will be reduced under normal conditions ofsustained tetanic contraction and also in neuromuscu-
lar junctions that are compromised (e.g., in patientswith myasthenia gravis).
Experimental and theoretical studies support thispremise. Experiments utilizing m-conotoxin, whichpreferentially blocks muscle NaChs, revealed thatNaCh activation could account for 15–20% of the end-plate potential (Hong and Chang, 1989). This effectmay be most important in man and other large mam-mals. A study of neuromuscular junctions in vastuslateralis muscle of man has revealed a lower quantalcontent and more elaborate postsynaptic membranefolding than that found in smaller mammals (Slater etal., 1992). It was proposed that the high concentrationof NaChs found in these extensive postsynaptic foldswould reduce the threshold voltage for action potentialgeneration at the neuromuscular junction. This pro-posal was recently examined experimentally (Woodand Slater, 1995) and theoretically. Using intracellularelectrodes to inject depolarizing current, Wood andSlater (1995) showed that the threshold for generatingan action potential was several millivolts lower at theneuromuscular junction than in extrajunctional re-gions. Ruff and Lennon (1998) showed a decreasedthreshold in the vicinity of human neuromuscular junc-tions. Wood and Slater (1995) also found that the
Fig. 2. Distribution of AChRs and NaChs at the neuromuscularjunction. AChRs are clustered at the tops of the postsynaptic folds.NaChs are found in the troughs of the postsynaptic folds and in theperijunctional membrane.
Fig. 1. Schematic representation of NaCh distribution in adult,mammalian skeletal muscle. Dots indicate the relative density ofNaChs. Density is highest at the endplate and decreases with dis-tance (both circumferentially and longitudinally) from the endplate.Average channel density does not change in the extrajunctional mem-
brane except near the tendon where the density decreases even fur-ther. Reproduced from Caldwell JH, Milton RL. 1988. Sodium channeldistribution in normal and denervated rodent and snake skeletalmuscle. J Physiol (Lond ) 401:145–161, with permission of the pub-lisher.
85ENDPLATE SODIUM CHANNELS
threshold for a nerve-evoked action potential was lowerthan that evoked by current injection. The greater ef-ficiency of nerve stimulation was predicted from thegeometry of the postsynaptic membrane folding (Vau-trin and Mambrini, 1989). A model accounting for thecombined effect of series resistance originating in thepostjunctional folds and the effect of the local NaChdensity was analyzed (Martin, 1994). In this model, thepresence of the postjunctional folds lowered (by a factorof 3) the threshold number of quanta of acetylcholineneeded to activate enough AChRs to initiate muscleexcitation. The NaCh concentration within the foldswas shown to act as an amplifier to ensure that exci-tation spread beyond the endplate region of the muscle.Thus, precise localization of the NaCh density to thedepths of the postsynaptic troughs serves to amplifyany synaptic potentials that originate from the highdensity of AChRs positioned at the crests of the folds.This amplification is expected to become critical duringnormal function, e.g., during high frequency stimula-tion when the endplate potential is reduced.
HOW DO MUSCLE CELLSCONCENTRATE NaChs?
Confinement of proteins to specific regions of a cellcan be regulated by many processes: influence of glialor satellite cells, basal lamina or extracellular matrixproteins, cytoskeletal and cytoskeletal-associated pro-teins, diffusible factors, local regulation of gene expres-sion, or any combination of these. Only recently havethe mechanisms employed by cells to regulate NaChdistribution begun to be investigated. The two sitesmost studied in excitable tissue have been the node ofRanvier and the neuromuscular junction. These twosites bear morphological and molecular similarities,including a high density of NaChs. Both exhibit mem-brane folding and are in close proximity to Schwann
cells. Both the neuromuscular junction and the nodesof Ranvier differentiate substantially during the post-natal period and aggregate NaChs during this time.Similar mechanisms may be utilized to form and main-tain the high densities in brain and muscle.
Influence of Glial Cells on NaCh DistributionGlial cells are a prominent feature wherever NaChs
are concentrated: at the neuromuscular junction, atnodes of Ranvier, and in the vicinity of neuronal somasand dendrites. The aggregation of NaChs has not beenobserved in muscle cultures, possibly due to the factthat NaCh aggregation is an aspect of synaptic matu-ration at the neuromuscular junction and that neuro-muscular contacts in culture rarely mature past thestage seen in prenatal animals. However, it has beenreported that addition of Schwann cells to nerve-mus-cle co-cultures induced further maturation at some ofthe contacts, compared to neuromuscular contacts incultures where no Schwann cells were added (Chapronand Koenig, 1989). Co-culture of neurons, muscles, andSchwann cells might thus be an advantageous modelsystem for studying NaCh aggregation and the role ofglial cells in this process.
Basal Lamina ProteinsMolecules in the extracellular matrix and basal lam-
ina that surround cells are able to influence manyaspects of cellular behavior, including the expressionand distribution of proteins. This molecular sheathsurrounds the muscle fiber and Schwann cell at theneuromuscular junction. It has been shown to retain ahigh density of synaptic molecules at the neuromuscu-lar junction even after degeneration of the nerve ter-minal and muscle fiber (McMahon and Slater, 1984).
Three results suggest that signals exist in the basallamina to maintain NaChs at high density. First, de-
Fig. 3. Developmental increase in sodium channel density at themouse neuromuscular junction. Mean sodium current density at theendplate region (solid bars) and extrajunctional region (hatched bars)of LAL (levator auris longus) muscles of various ages. Measurementsmade with loose patch voltage clamp recording. Bars represent the
mean SEM of 22 fibers. **P 0.01; ***P 0.001. Reproduced from LupaMT, Krzemien DM, Schaller KL, Caldwell JH. 1993. Aggregation ofsodium channels during development and maturation of the neuro-muscular junction. J Neurosci 13:1326–1336, with permission of thepublisher.
86 J.H. CALDWELL

nervation of muscle leads to degeneration of the nerveterminal but NaChs remain concentrated at the neu-romuscular junction (Caldwell and Milton, 1988: Lupaet al., 1995). Second, NaChs were shown to aggregateat synaptic sites on muscle fibers that were damagedand allowed to regenerate in the absence of nerve ter-minals; NaChs became concentrated either at the sametime or soon after formation of AChR clusters at theoriginal endplate region (Lupa and Caldwell, 1994).The conclusion that a signal persists in the basal lam-ina is supported by a third observation: treatment ofadult muscle fibers with collagenase induces a gradualfragmentation and dispersal of both AChR and NaChaggregates from the endplate region (Bloch et al., 1986;Lupa and Caldwell, 1991). Assuming that collagenasetreatment does not directly affect intracellular struc-tures such as the cytoskeleton, these results imply thatan extracellular matrix or basal lamina molecule isresponsible for maintaining the high concentration ofNaChs at adult endplates.
Agrin is a basal lamina protein that initiates theformation of AChR densities at developing and regen-erating neuromuscular junctions (McMahan et al.,1992; Ruegg and Bixby, 1998). cDNAs that code for rat,chick, and Torpedo agrin have been isolated and char-acterized. These molecular studies revealed multipleisoforms that are generated by alternative RNA splic-ing of a single gene. The isoforms are functionallysignificant since they differ both in their ability tocluster AChRs and in their developmental expression.The distinction between different agrin isoforms raisedthe possibility that NaCh clustering in rat muscle re-quires a specific rat agrin isoform that is expressedpostnatally. Cocultures of rat muscle fibers with CHOcells transfected with specific isoforms of rat agrinshowed that a neural isoform (denoted Y4Z8) will clus-ter NaChs within 1–2 days (Sharp and Caldwell, 1996).A second set of studies, utilizing an in vivo preparation(Cohen et al., 1997; Meier et al., 1997) tested the long-term effects of agrin and suggested that the Y4Z8 iso-form of agrin is capable of aggregating NaChs. Ratagrin cDNA for the Y4Z8 isoform was injected into theextrajunctional region of the soleus muscle. Some mus-cle fibers took up the cDNA and locally synthesizedthat agrin isoform at the injection site. AChR clustersand membrane folding resembling postsynaptic foldsdeveloped at the areas of agrin synthesis. Localizationusing anti-NaCh antibodies on these muscles showedthat NaChs were aggregated not only at sites of AChRclusters but also in the adjacent membrane, similar tothe distribution at the neuromuscular junction (Rimeret al., 1998). Thus, agrin is a likely candidate for abasal lamina molecule that triggers the clustering ofNaChs. However, since both the spatial and temporalaspects of AChR clustering are so different from thosefor NaCh clustering, it is likely that additional mole-cules, specific for NaChs, act to mediate events inNaCh clustering.
Brain NaChs are a complex consisting of the pore-forming alpha subunit and two beta subunits (beta1and beta2). Muscle NaChs are associated only with thebeta1 subunit (beta2 is not expressed in muscle). Te-nascin is an extracellular matrix molecule that hasbeen shown to bind to the beta2 subunit of brain so-dium channels (Srninvasan et al., 1998). Binding of the
brain NaChs by tenascin could participate in localizingthe channels or in regulating their activity. Moleculesthat bind similarly to muscle NaChs have not beenidentified.
Role of Cytoskeletal-Associated ProteinsThe function of the intracellular cytoskeleton is to
provide a rigid, modifiable structure for cell shape, aswell as scaffolding to which various organelles andmolecules can attach and interact. NaChs have beenshown by antibody precipitation experiments to be as-sociated with the cytoskeletal and cytoskeletal-associ-ated proteins spectrin and ankyrin (Srninvasan et al.,1998). A specific form of ankyrin (ankyrin G) has beenshown to be concentrated at the neuromuscular junc-tion (Kordeli, 1998), and both beta-spectrin andankyrin G colocalize with NaChs at the neuromuscularjunction (Wood and Slater, 1998). Immunoelectronmi-croscopy studies (Flucher and Daniels, 1989) showedthat NaChs line the troughs of the postjunctional foldsat the neuromuscular junction, with ankyrin localizedto the adjacent area of the intracellular side of the cellmembrane. This arrangement is nearly identical tothat of AChRs and rapsyn (also known as the 43kDprotein) at the crests of the folds. Rapsyn is a moleculethat is closely associated with the AChR, is found onthe cytoplasmic aspect of the cell membrane, and isbelieved to stabilize AChR clusters as they form in themembrane by anchoring the receptors to the underly-ing actin filament network. It is possible that ankyrinstabilizes NaCh clusters in a similar manner.
Syntrophin is an intracellular protein that binds toall forms of dystrophin. Three syntrophin genes havebeen isolated. Each subtype has one PDZ domain, twopleckstrin homology domains, and one domain uniqueto syntrophin. PDZ domains bind membrane proteinsthat have a carboxyl terminus ending in (S/T) XV.Muscle NaChs (but none of the brain NaChs) have thissignature sequence. Gee et al. (1998) showed that mus-cle NaChs (both the embryonic and mature skeletalmuscle isoforms) bind to the PDZ domain of all threesyntrophin subtypes. Peptides corresponding to thelast ten amino acids of the two muscle NaCh isoformsbind directly to the PDZ domain of syntrophin. Thus, itis likely that syntrophin tethers muscle NaChs in acomplex with dystrophin (Fig. 4).
Local Regulation of NaCh ExpressionThe cellular arrangement at the neuromuscular
junction allows another type of mechanism to regulateNaCh density and distribution. The synaptic nucleiclustered under the postsynaptic membrane may bespecialized or induced to produce a high level of NaChsin this domain of the muscle, or to express a novelNaCh isoform that has the ability to aggregate at thesynapse. There is evidence, for example, that the nerveterminal secretes two proteins, neuregulin and CGRP,which may act to increase expression of NaChs. Neu-regulins are ligands that activate receptor protein ty-rosine kinases (Mudge, 1993). Several lines of evidencesuggest that neuregulins and their receptors (whichare similar to the epithelial growth factor receptor,termed erbB) might play a role in NaCh clustering.First, ARIA (AChR-Inducing Activity), which has beenshown to be a neuregulin, causes a 3–4-fold increase in
87ENDPLATE SODIUM CHANNELS

functional NaChs when applied to cultured musclecells (Corfas and Fischbach, 1993). Second, neuregu-lins are synthesized by motor neurons in the (Mar-chionni et al., 1993)embryo. Third, neuregulins aredeposited and concentrated in the extracellular matrixat the neuromuscular junction (Jo et al., 1995). Finally,neuregulin accumulation is a late event in synapseformation (Zhu et al., 1995) with a time course verysimilar to that which was described for NaChs (Lupa etal., 1993).
The second protein, CGRP (Calcitonin Gene-RelatedPeptide), has been shown to increase the level of cAMPin muscle cells (Falls et al., 1990), and a change inintracellular cAMP levels has been shown to affectNaCh expression in the brain (Li et al., 1992). Both ofthese molecules are secreted by nerve terminals andmay act as trophic factors to regulate NaCh density inthe postsynaptic membrane.
During muscle maturation, a change occurs in thetype of NaCh expressed, from an embryonic to a ma-ture form. These two NaCh isoforms are encoded bydifferent genes and are differentially sensitive to block-ade by the neurotoxin tetrodotoxin (TTX). RNase pro-tection assays have shown that the developmentalswitch in expression of these two proteins occurs overthe same time period as the onset of NaCh aggregationmeasured electrophysiologically at the synapse (Lupaet al., 1993). Thus, it was possible that only the mature,TTX-sensitive form of the NaCh could become concen-trated at the endplate. This was tested by comparingthe TTX-sensitivity of Na currents recorded at end-plate and extrajunctional regions on developing musclecells. The evidence showed that, as the endplate NaChdensity developed, similar proportions of the embry-onic, TTX-resistant and adult, TTX-sensitive NaChscould be detected in both synaptic and extrajunctionalareas of the muscle membrane. Thus, both types of
NaCh had the ability to become aggregated at theneuromuscular synapse. This conclusion was also sup-ported by previous studies showing that the NaCh con-centration persists at neuromuscular junctions aftermany weeks of denervation, and both TTX-sensitiveand TTX-insensitive NaChs were concentrated at thedenervated endplate (Caldwell and Milton, 1988; Lupaet al., 1995).
SUMMARYAlthough both NaChs and AChRs are concentrated
at the neuromuscular junction, they differ in manydetails of localization and regulation. NaChs are in thedepths of the postsynaptic folds while AChRs are at thecrests. NaChs are reduced in density near the tendon ofsoleus muscle where AChRs are increased in density.AChRs are aggregated within minutes or hours of neu-rite contact (about a week before birth in rodents), butNaChs do not aggregate until 2 weeks later. Denerva-tion causes a large increase in the density of extrajunc-tional AChRs but little change in the density of NaChs.NaChs and AChRs bind to different cytoskeletal-asso-ciated proteins (ankyrin and rapsyn, respectively).Thus, it is reasonable to expect that there will be ad-ditional differences in the details of regulation ofAChRs and NaChs.
The following sequence of events is consistent withthe experimental results. NaCh aggregation is initi-ated by an isoform of agrin that is upregulated about 1week after birth and that induces clustering of NaChspresent in the membrane. NaCh synthesis may be reg-ulated by an isoform of neuregulin, which increasestranscription in nuclei near the synapse, producing alocal increase in NaCh density. Ankyrin and syntro-phin are recruited to the synapse and enable NaChs toaccumulate at a high density by tethering or immobi-lizing them.
Fig. 4. Linkage of NaChs to dystrophin by binding to syntrophin.The PDZ domain of syntrophin (SYN) binds the carboxyl-terminalSXV sequence of muscle NaChs. Syntrophin is bound to dystrophin(DYS) which in turn binds beta-dystroglycan (-DG). The sarcoglycancomplex (SG) and alpha-dystroglycan (-DG) are also associated with
dystrophin. Reproduced from Gee SH, Madhavan R, Levinson SR,Caldwell JH, Sealock R, Froehner SC. 1998. Interaction of muscle andbrain sodium channels with multiple members of the syntrophinfamily of dystrophin-associated proteins. J Neurosci 18:128–137 withpermission of the publisher.
88 J.H. CALDWELL

REFERENCESAngelides KJ. 1986. Fluorescently labelled Na1 channels are local-
ized and immobilized to synapses of innervated muscle fibres. Na-ture 321:63–66.
Beam KG, Caldwell JH, Campbell DT. 1985. Na channels in skeletalmuscle concentrated near the neuromuscular junction. Nature 313:588–590.
Betz WJ, Caldwell JH, Kinnamon SC. 1984. Increased sodium con-ductance in the synaptic region of rat skeletal muscle fibres.J Physiol (Lond ) 352:189–202.
Bloch RJ, Steinbach JH, Merlie JP, Heinemann S. 1986. Collagenasedigestion alters the organization and turnover of junctional acetyl-choline receptors. Neurosci Lett 66:113–119.
Boudier JL, Le Treut T, Jover E. 1992. Autoradiographic localizationof voltage-dependent sodium channels on the mouse neuromuscularjunction using 125I-alpha scorpion toxin. II. Sodium distribution onpostsynaptic membranes. J Neurosci 12:454–466.
Caldwell JH, Campbell DT, Beam KG. 1986. Na channel distributionin vertebrate skeletal muscle. J Gen Physiol 87:907–932.
Caldwell JH, Milton RL. 1988. Sodium channel distribution in normaland denervated rodent and snake skeletal muscle. J Physiol(Lond ) 401:145–161.
Chapron J, Koenig J. 1989. In vitro synaptic maturation. NeurosciLett 106:19–22.
Cohen I, Rimer M, Lomo T, McMahan UJ. 1997. Agrin-inducedpostsynaptic-like apparatus in skeletal muscle fibers in vivo [pub-lished erratum appears in Mol Cell Neurosci 1997;10(3-4):208]. MolCell Neurosci 9:237–253.
Corfas G, Fischbach GD. 1993. The number of Na1 channels incultured chick muscle is increased by ARIA, an acetylcholine recep-tor-inducing activity. J Neurosci 13:2118–2125.
Falls DL, Harris DA, Johnson FA, Morgan MM, Corfas G, FischbachGD. 1990. Mr 42,000 ARIA: a protein that may regulate the accu-mulation of acetylcholine receptors at developing chick neuromus-cular junctions. [Review] [50 refs]. Cold Spring Harbor Symp QuantBiol 55:397–406.
Flucher BE, Daniels MP. 1989. Distribution of Na1 channels andankyrin in neuromuscular junctions is complementary to that ofacetylcholine receptors and the 43 kd protein. Neuron 3:163–175.
Gee SH, Madhavan R, Levinson SR, Caldwell JH, Sealock R, Froeh-ner SC. 1998. Interaction of muscle and brain sodium channels withmultiple members of the syntrophin family of dystrophin-associatedproteins. J Neurosci 18:128–137.
Haimovich B, Schotland DL, Fieles WE, Barchi RL. 1987. Localizationof sodium channel subtypes in adult rat skeletal muscle usingchannel-specific monoclonal antibodies. J Neurosci 7:2957–2966.
Hong SJ, Chang CC. 1989. Use of geographutoxin II (mu-conotoxin)for the study of neuromuscular transmission in mouse. Br J Phar-macol 97:934–940.
Jo SA, Zhu X, Marchionni MA, Burden SJ. 1995. Neuregulins areconcentrated at nerve-muscle synapses and activate ACh-receptorgene expression. Nature 373:158–161.
Kordeli E, Ludosky MA, Deprette C, Frappier T, Cartaud J. 1998.AnkyrinG is associated with the postsynaptic membrane and thesarcoplasmic reticulum in the skeletal muscle fiber. J Cell Sci111:2197–2207.
Li M, West JW, Lai Y, Scheuer T, Catterall WA. 1992. Functionalmodulation of brain sodium channels by cAMP-dependent phos-phorylation. Neuron 8:1151–1159.
Lupa MT, Caldwell JH. 1991. Effect of agrin on the distribution ofacetylcholine receptors and sodium channels on adult skeletal mus-cle fibers in culture. J Cell Biol 115:765–778.
Lupa MT, Caldwell JH. 1994. Sodium channels aggregate at formersynaptic sites in innervated and denervated regenerating muscles.J Cell Biol 124:139–147.
Lupa MT, Krzemien DM, Schaller KL, Caldwell JH. 1993. Aggrega-tion of sodium channels during development and maturation of theneuromuscular junction. J Neurosci 13:1326–1336.
Lupa MT, Krzemien DM, Schaller KL, Caldwell JH. 1995. Expressionand distribution of sodium channels in short- and long-term dener-vated rodent skeletal muscles. J Physiol (Lond ) 483:109–118.
Marchionni MA, Goodearl AD, Chen MS, Bermingham MD, Kirk C,Hendricks M, Danehy F, Misumi D, Sudhalter J, Kobayashi K. 1993.Glial growth factors are alternatively spliced erbB2 ligands expressedin the nervous system [see comments]. Nature 362:312–318.
Martin AR. 1994. Amplification of neuromuscular transmission bypostjunctional folds. Proc R Soc Lond [Biol ] 258:321–326.
McMahan UJ, Slater CR. 1984. The influence of basal lamina on theaccumulation of acetylcholine receptors at synaptic sites in regen-erating muscle. J Cell Biol 98:1453–1473.
McMahan UJ, Horton SE, Werle MJ, Honig LS, Kroger S, Ruegg MA,Escher G. 1992. Agrin isoforms and their role in synaptogenesis.[Review] [22 refs]. Curr Opin Cell Biol 4:869–874.
Meier T, Hauser DM, Chiquet M, Landmann L, Ruegg MA, BrennerHR. 1997. Neural agrin induces ectopic postsynaptic specializationsin innervated muscle fibers. J Neurosci 17:6534–6544.
Milton RL, Lupa MT, Caldwell JH. 1992. Fast and slow twitch skel-etal muscle fibres differ in their distribution of Na channels nearthe endplate. Neurosci Lett 135:41–44.
Mudge A. 1993. New ligands for neu? Curr Biol 3:361–364.Nastuk W, Alexander J. 1973. Non-homogeneous electrical activity in
single muscle fibers. Fed Proc 32:333Rimer M, Cohen I, Lomo T, Burden SJ, McMahan UJ. 1998. Neuregu-
lins and erbB receptors at neuromuscular junctions and at agrin-induced postsynaptic-like apparatus in skeletal muscle. Mol CellNeurosci 12:1–15.
Roberts WM. 1987. Sodium channels near end-plates and nuclei ofsnake skeletal muscle. J Physiol (Lond ) 388:213–232.
Ruegg MA, Bixby JL. 1998. Agrin orchestrates synaptic differentia-tion at the vertebrate neuromuscular junction. [Review] [74 refs].Trends Neurosci 21:22–27.
Ruff RL. 1992. Na current density at and away from end plates on ratfast- and slow-twitch skeletal muscle fibers. Am J Physiol 262:C229–C234
Ruff RL, Lennon VA. 1998. End-plate voltage-gated sodium channelsare lost in clinical and experimental myasthenia gravis. Ann Neurol43:370–379.
Ruff RL, Whittlesey D. 1992. Na1 current densities and voltagedependence in human intercostal muscle fibres. J Physiol (Lond )458:85–97.
Sharp AA, Caldwell JH. 1996. Aggregation of sodium channels in-duced by a postnatally upregulated isoform of agrin. J Neurosci16:6775–6783.
Slater CR, Lyons PR, Walls TJ, Fawcett PR, Young C. 1992. Structureand function of neuromuscular junctions in the vastus lateralis ofman. A motor point biopsy study of two groups of patients. Brain115:451–478.
Srinivasan J, Schachner M, Catterall WA. 1998. Interaction of voltage-gated sodium channels with the extracellular matrix molecules tena-scin-C and tenascin-R. Proc Natl Acad Sci USA 95:15753–15757.
Srinivasan Y, Elmer L, Davis J, Bennett V, Angelides K. 1988.Ankyrin and spectrin associate with voltage-dependent sodiumchannels in brain. Nature 333:177–180.
Thesleff S, Vyskocil F, Ward MR. 1974. The action potential in end-plate and extrajunctional regions of rat skeletal muscle. ActaPhysiol Scand 91:196–202.
Vautrin J, Mambrini J. 1989. Synaptic current between neuromuscu-lar junction folds. J Theoret Biol 140:479–498.
Wood SJ, Slater CR. 1995. Action potential generation in rat slow-and fast-twitch muscles. J Physiol (Lond ) 486:401–410.
Wood SJ, Slater CR. 1998. b-spectrin is colocalized with both voltage-gated sodium channels and ankyrinG at the adult rat neuromuscu-lar junction. J Cell Biol 140:675–684.
Zhu X, Lai C, Thomas S, Burden SJ. 1995. Neuregulin receptors,erbB3 and erbB4, are localized at neuromuscular synapses. EMBOJ 14:5842–5848.
89ENDPLATE SODIUM CHANNELS