cloning and characterization of a novel human gene, tm4sf6, encoding a protein belonging to the...
TRANSCRIPT

BRIEF REPORTS
Cloning and Characterization of aNovel Human Gene, TM4SF6,Encoding a Protein Belonging to theTransmembrane 4 Superfamily, andMapped to Xq22Kenji Maeda,*,† Sachiko Matsuhashi,*,1 Katsuji Hori,*Zhenghan Xin,* Tsunehiro Mukai,* Kazuo Tabuchi,†Masayuki Egashira,‡,§ and Norio Niikawa‡
*Department of Biochemistry and †Department of Neurosurgery, SagaMedical School, Nabeshima 5-1-1, Saga 849-8501, Japan; and‡Department of Human Genetics and §Department of Orthopaedics,Nagasaki University School of Medicine, Sakamoto 1-12-4,Nagasaki 852-8523, Japan
Received February 24, 1998; accepted May 22, 1998
The transmembrane 4 (or tetraspanin) superfamily(TM4SF) proteins include nearly 20 members that arestructurally closely related with 20 –30% sequence homol-ogy (7, 11). The polypeptides in this family contain fourhydrophobic, presumably membrane-spanning sequencesand a major presumed extracellular loop between the thirdand the fourth hydrophobic domains. Although the TM4SFprotein was first identified as the CD9 surface antigen ofpre-B lymphocytes (9), members of this family includingCD9 have been reported to be widely expressed in manydifferent tissues (11). It has been reported that expressionof the CD82 (KAI1) and CD9 genes suppressed experimen-tal metastasis of a rat prostate cell line (2) and a mousemelanoma cell line (8), respectively. Several members ofthe family have been shown to stimulate or modulate cellgrowth (1, 6, 10, 11). These biological activities may becaused by the ability of the proteins to associate with oneanother and with other membrane proteins to form com-plexes. Recently, it has been shown that some of theseproteins may associate with integrin and control cell ad-hesion and cell movement (4, 5, 11).
During an investigation of NELL2 gene expression inhuman glioma tissues, a cDNA (N273-9) homologous toA15/TALLA1 (3, 14), a member of the TM4SF, was iso-lated. An EST sequence, HSC15C081 (Accession No.F0634800), partly identical to N273-9, was found in thedatabase. The combined sequence of N273-9 andHSC15C081, contained an ORF encoding a 245-amino-acidsequence homologous to the A15/TALLA-1 protein. A hu-man fetal lung cDNA library (Strategene) was screenedwith a 32P-labeled probe corresponding to the combined
sequence of HSC15C081 and N273-9 (the library gavestrong signal for the sequence by a PCR assay). Threedifferent clones thus isolated were sequenced. All threeclones contained an ORF identical to that obtained fromthe combined sequence of N273-9 and HSC15C081.
The deduced amino acid sequence of the clones (seeGenBank Accession No. AF043906) contained four hydro-phobic, possibly transmembrane domains and a presumedmajor extracellular domain between the presumed thirdand forth transmembrane segments. The well-conservedshort motifs including cysteine residues (CCG, PXSC, andGC) in the major extracellular loop of TM4SF (11) areconserved in the putative major extracellular loop. A pos-sible N-glycosylation site is present in the putative majorextracellular loop. The deduced amino acid sequenceshowed 58% homology to human A15/TALLA-1 protein and26 –36% to other members of the human TM4SF group.From these results, it was concluded that the gene is a newmember of the TM4SF group, and we named the genetransmembrane 4 superfamily member 6 (TM4SF6, alsoknown as T245).
TM4SF6 (T245) gene expression was widely observed inhuman adult tissues (Figs. 1A and 1B). Among the tissuestested, high-level expression was observed in the liver,pancreas, kidney, and ovary, while low-level expressionwas observed in the skeletal muscle, lung, and brain. Theexpression level of fetal lung might be higher than in theadult, because the fetal lung cDNA library used for thescreening of T245 gave a strong signal for the gene. Ex-pression in the spleen, peripheral lymphocytes (Fig. 1B,lanes 1 and 3, respectively), and a T-cell lymphoma CEM
The nucleotide and deduced amino acid sequences from this articlehave been deposited with the GenBank Data Library under Acces-sion No. AF043906.
1 To whom correspondence should be addressed. Telephone: 0952-34-2264. Fax: 0952-34-2067. E-mail: [email protected].
FIG. 1. Northern blot analysis. (A) Northern blot (Clontech) ofpoly(A) RNAs derived from human tissues. Pancreas (lane 1); kidney(2); skeletal muscle (3); liver (4); lung (5); placenta (6); brain (7); andheart (8). M represents DNA size marker. (B) Northern blot (Ori-Gene) of poly(A) RNAs derived from human tissues. Spleen (1); liver(2); peripheral lymphocytes (3); prostate (4); placenta (5); and ovary(6). Approximately 2 mg poly(A) RNA was loaded in each lane in Aand B. (C) 20 mg total RNA derived from human cell lines: HeLa (1),CEM (2), IMR 32 (3), and human tumor tissues excised from pa-tients: neuroblastoma (4), ependymoma 383 (5), meningioma 384 (6),and meningioma 468 (7), was loaded on each lane. Hybridization wascarried out according to the GeneScreen instruction manual with a32P-labeled T245 probe.
GENOMICS 52, 240–242 (1998)ARTICLE NO. GE985415
240
0888-7543/98 $25.00Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.
cell line (Fig. 1C, lane 2) was low, though TALLA1, whichhas the highest homology with T245, was highly expressedin the spleen and in T-cell lymphoma cell lines includingCEM (14). Surprisingly, extremely high expression wasobserved in an ependymoma excised from a patient (Fig.1C, lane 5) although only weak signal was observed in thebrain or in other brain tumors such as neuroblastoma ormeningioma (Figs. 1A and 1C). The expression levels ofglioma tissues or cell lines were also low (data not shown),although a T245 cDNA (N273-9) was initially isolated froma glioma tissue. Two bands of 1.9- and 1.3-kb T245 mRNAwere observed in all tissues and cell lines tested. It isexpected that these bands are derived from the same geneby alternative splicing or poly(A) tail addition, becauseonly one gene was detected in the genome by Southern blotanalysis (data not shown).
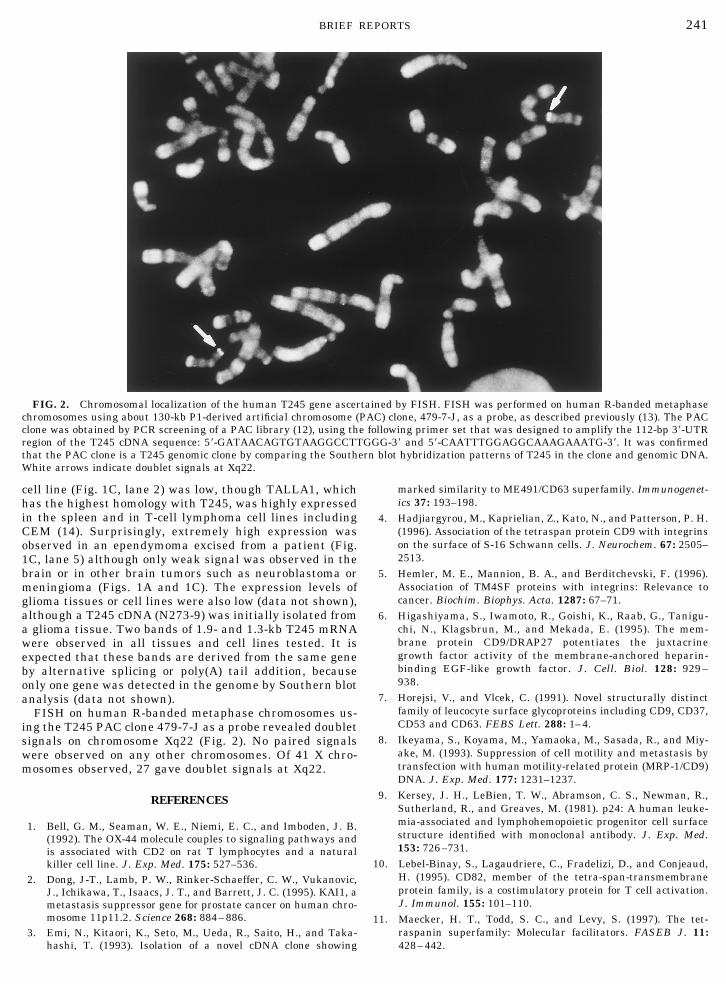
FISH on human R-banded metaphase chromosomes us-ing the T245 PAC clone 479-7-J as a probe revealed doubletsignals on chromosome Xq22 (Fig. 2). No paired signalswere observed on any other chromosomes. Of 41 X chro-mosomes observed, 27 gave doublet signals at Xq22.
REFERENCES
1. Bell, G. M., Seaman, W. E., Niemi, E. C., and Imboden, J. B.(1992). The OX-44 molecule couples to signaling pathways andis associated with CD2 on rat T lymphocytes and a naturalkiller cell line. J. Exp. Med. 175: 527–536.
2. Dong, J-T., Lamb, P. W., Rinker-Schaeffer, C. W., Vukanovic,J., Ichikawa, T., Isaacs, J. T., and Barrett, J. C. (1995). KAI1, ametastasis suppressor gene for prostate cancer on human chro-mosome 11p11.2. Science 268: 884–886.
3. Emi, N., Kitaori, K., Seto, M., Ueda, R., Saito, H., and Taka-hashi, T. (1993). Isolation of a novel cDNA clone showing
marked similarity to ME491/CD63 superfamily. Immunogenet-ics 37: 193–198.
4. Hadjiargyrou, M., Kaprielian, Z., Kato, N., and Patterson, P. H.(1996). Association of the tetraspan protein CD9 with integrinson the surface of S-16 Schwann cells. J. Neurochem. 67: 2505–2513.
5. Hemler, M. E., Mannion, B. A., and Berditchevski, F. (1996).Association of TM4SF proteins with integrins: Relevance tocancer. Biochim. Biophys. Acta. 1287: 67–71.
6. Higashiyama, S., Iwamoto, R., Goishi, K., Raab, G., Tanigu-chi, N., Klagsbrun, M., and Mekada, E. (1995). The mem-brane protein CD9/DRAP27 potentiates the juxtacrinegrowth factor activity of the membrane-anchored heparin-binding EGF-like growth factor. J. Cell. Biol. 128: 929 –938.
7. Horejsi, V., and Vlcek, C. (1991). Novel structurally distinctfamily of leucocyte surface glycoproteins including CD9, CD37,CD53 and CD63. FEBS Lett. 288: 1–4.
8. Ikeyama, S., Koyama, M., Yamaoka, M., Sasada, R., and Miy-ake, M. (1993). Suppression of cell motility and metastasis bytransfection with human motility-related protein (MRP-1/CD9)DNA. J. Exp. Med. 177: 1231–1237.
9. Kersey, J. H., LeBien, T. W., Abramson, C. S., Newman, R.,Sutherland, R., and Greaves, M. (1981). p24: A human leuke-mia-associated and lymphohemopoietic progenitor cell surfacestructure identified with monoclonal antibody. J. Exp. Med.153: 726–731.
10. Lebel-Binay, S., Lagaudriere, C., Fradelizi, D., and Conjeaud,H. (1995). CD82, member of the tetra-span-transmembraneprotein family, is a costimulatory protein for T cell activation.J. Immunol. 155: 101–110.
11. Maecker, H. T., Todd, S. C., and Levy, S. (1997). The tet-raspanin superfamily: Molecular facilitators. FASEB J. 11:428–442.
FIG. 2. Chromosomal localization of the human T245 gene ascertained by FISH. FISH was performed on human R-banded metaphasechromosomes using about 130-kb P1-derived artificial chromosome (PAC) clone, 479-7-J, as a probe, as described previously (13). The PACclone was obtained by PCR screening of a PAC library (12), using the following primer set that was designed to amplify the 112-bp 39-UTRregion of the T245 cDNA sequence: 59-GATAACAGTGTAAGGCCTTGGG-39 and 59-CAATTTGGAGGCAAAGAAATG-39. It was confirmedthat the PAC clone is a T245 genomic clone by comparing the Southern blot hybridization patterns of T245 in the clone and genomic DNA.White arrows indicate doublet signals at Xq22.
241BRIEF REPORTS

12. Matsumoto, N., Soeda, E., Ohashi, H., Fujimoto, M., Kato, R.,Tsujita, T., Tomita, H., Kondo, S., Fukushima, Y., and Niikawa,N. (1997) A 1.2-megabase BAC/PAC contig spanning the 14q13breakpoint of t(2;14) in a mirror-image polydactyly patient.Genomics 45: 11–16.
13. Ohta, T., Tohma, T., Soejima, H., Fukushima, Y., Nagai, T.,Yoshiura, K., Jinno, Y., and Niikawa, N. (1993). The origin ofcytologically unidentifiable chromosome abnormalities: Sixcases ascertained by targeted chromosome-band painting.Hum. Genet. 92: 1–5.
14. Takagi, S., Fujikawa, K., Imai, T., Fukuhara, N., Fukudome,K., Minegishi, M., Tsuchiya, S., Konno, T., Hinuma, Y., andYoshie, O. (1995). Identification of a highly specific surfacemarker of T-cell acute lymphoblastic leukemia and neuroblas-toma as a new member of the transmembrane 4 superfamily.Int. J. Cancer 61: 706–715.
Assignment of the SteroidReceptor Coactivator-1 (SRC-1)Gene to Human ChromosomeBand 2p23Melina Carapeti,* Ricardo C. T. Aguiar,†Andrew Chase,* John M. Goldman,*and Nicholas C. P. Cross*,1
*Department of Haematology, Imperial College School of Medicine,Hammersmith Hospital, Du Cane Road, London W12 ONN, UnitedKingdom; and †Division of Hematologic Malignancies, Dana FarberCancer Institute, 44 Binney Street, Boston, Massachusetts 02115
Received March 16, 1998; accepted June 9, 1998
Nuclear receptor coactivators (NRCoAs) mediate the li-gand-dependent transcriptional activation of specific genesby nuclear receptors, for example, the receptors for steroidand thyroid hormones, retinoids, and vitamin D. Currently,three NRCoAs have been cloned: TIF2 (also known as GRIP1or NCoA-2) (11), SRC-1 (also known as F-SRC-1 or NCoA-1)(8), and AIB1 (also known as RAC3, ACTR, or p/CIP) (2).These proteins are functionally and structurally homologousand are thought to stimulate gene expression by facilitatingthe assembly of basal transcription factors into a stablepreinitiation complex. Recently NRCoAs have been shown topossess histone acetyl transferase (HAT) activity and also tointeract directly with CBP, a general transcriptional co-inte-grator (10). It is likely, therefore, that NRCoAs mediate tran-scriptional activation by a mechanism that involves chroma-tin remodeling.
Several lines of evidence have implicated abnormalities ofNRCoAs in malignancy. The AIB1 gene at 20q12 is fre-quently amplified in breast and ovarian cancers (2), and we
have recently cloned a novel fusion between the TIF2 and theMOZ genes at 8q13 and 8p11, respectively, in acute myeloidleukemia (AML) (1, 3). In addition, the ARA70 gene wasshown to be fused to RET in a case of human thyroid papil-lary carcinoma (9, 12), and TIF1 is fused to B-RAF in themouse hepatoma-derived oncogene T18 (5). Although ARA70
and TIF1 do not share sequence homology with the NRCoAfamily, they are also involved in the ligand-dependent acti-vation function of nuclear receptors.
Here we have mapped the chromosomal localization ofSRC-1 (GenBank Accession No. U40396) to determinewhether this gene is also located at a region that has beenimplicated in malignancy. An SRC-1 probe, designed to aregion of low homology between the NRCoA family mem-bers, was amplified from normal peripheral blood leukocytecDNA with primers SRC1-A (59-CTGAGCTAGCTGAACTA-GAC-39) and SRC1-B (59-GGTGCTGTTGACTGTACAGT-39).The probe was sequenced to confirm its identity and used toscreen the gridded human library RPCI1. Four positive PACclones (Nos. 100-G8, 115-I1, 125-H2, and 200-P18) were iso-lated and used as fluorescence in situ hybridization (FISH)probes on metaphases from phytohemagglutinin-stimulated pe-ripheral blood lymphocytes from a normal individual. PACswere labeled with digoxigenin by nick-translation and detectedwith sheep anti-digoxigenin (Boehringer-Mannheim, Germa-ny), rabbit anti-sheep fluorescein isothiocyanate (FITC; Vector,UK), and finally swine anti-rabbit FITC (Dako, UK). All fourPACs specifically hybridized to chromosome band 2p23 andto no other region (Fig. 1A). A gridded CEPH mega-YAClibrary was also screened with the SRC-1 probe. Severalpositive clones were identified, including Nos. 744h7, 765d3,916f8, and 953d6. Comparison with the existing map of chro-mosome 2 (4) indicates that SRC-1 maps to the short arm inthe vicinity of markers D2S144 and D2S171 (Fig. 1B). Thesemarkers have been mapped to 48 cM and 149 cR of thegenetic and radiation hybrid maps, respectively, and arecontained in the contig WC2.2 (http://www.genome.wi.mit.edu:80/cgi-bin/contig/lookup_contig).
Translocations involving chromosome 2p23 or neighboringbands have been reported in a variety of hematological ma-lignancies (6). The ALK gene at 2p23 is disrupted by t(2;5)(p23;q35) in anaplastic large-cell lymphoma (7), but themolecular etiology of other translocations, for example, thet(2;3)(p23;q26) in AML, is unknown. The mapping of SRC-1 to2p23 provides a further candidate gene for disruption bythese translocations.
ACKNOWLEDGMENTS
We thank the Medical Research Council Human Genome MappingProject (MRC HGMP) Resource Centre (Hinxton, UK) for providingthe PAC clones and CEPH megaYAC clones.
REFERENCES
1. Aguiar, R. C. T., Chase, A., Coulthard, S., Macdonald, D. H. C.,Carapeti, M., Reiter, A., Sohal, J., Lennard, A., Goldman, J. M.,and Cross, N. C. P. (1997). Abnormalities of chromosome band
1 To whom correspondence should be addressed. Telephone: 1 44181 383 3302. Fax: 1 44 181 740 9679. E-mail: [email protected].
242 BRIEF REPORTS
GENOMICS 52, 242–244 (1998)ARTICLE NO. GE9854460888-7543/98 $25.00Copyright © 1998 by Academic PressAll rights of reproduction in any form reserved.