chitin synthesis and degradation as targets for pesticide action
TRANSCRIPT

Archives of Insect Biochemistry and Physiology 22:245-261 (1 993)
Chitin Synthesis and Degradation as Targets for Pesticide Action Ephraim Cohen Department of Entornolo#, Faculty of Agriculture, The Hebrew University of Jerusalem, Rehovot, Israel
Various pesticides are being used to destabilize, perturb, or inhibit crucial biochemical and physiological targets related to metabolism, growth, develop- ment, nervous communication, or behavior in pestiferous organisms. Chitin is an eukaryotic extracellular aminosugar biopolymer, massively produced by most fungal systems and by invertebrates, notably arthropods. Being an integral supportive component in fungal cell wall, insect cuticle, and nematode egg shell, chitin has been considered as a selective target for pesticide action. Throughout the elaborate processes of chitin formation and deposition, only the polymeriza- tion events associated with the cell membrane compartment are so far available for chemical interference. Currently, the actinomycetes-derived nucleoside pep- tide fungicides such as the polyoxins and the insecticidal benzoylaryl ureas have reached commercial pesticide status. The polyoxins and other structurally-related antibiotics like nikkomycins are strong competitive inhibitors of the polymerizing enzyme chitin synthase. The exact biochemical lesion inflicted by the benzoylaryl ureas i s still elusive, but a post-polymerization event, such as translocation of chitin chains across the cell membrane, i s suggested.
Hydrolytic degradation of the chitin polymer is essential for hyphal growth, branching, and septum formation in fungal systems as well as for the normal molting of arthropods. Recently, insect chitinase activity was strongly and specifically suppressed by allosamidin, an actimomycetes-derived metabolite. In part, the defense mechanism in plants against invasion of pathogens is associated with induced chitinases. Chitin, chitosan, and their oligomers are able to act as elicitors which induce enhanced levels of chitinases in various plants. Lectins which bind to N-acetyl-D-glucosamine strongly interfere with fungal and insect chitin synthases. Plant lectins with similar properties may be involved in plant-pathogen interaction inter alia by suppressing fungal invasion. 0 1993 Wiley-Liss, Inc.
Key words: allosamidin, benzoylaryl ureas, chitin synthesis inhibitors, chitin synthase, chitinase
Received January 21, 1992; accepted March 18, 1992.
Address reprint requests to Ephraim Cohen, Department of Entomology, Faculty of Agriculture, The Hebrew University of Jerusalem, Rehovot 76 100, Israel.
0 1993 Wiley-Liss, Inc.

246 Cohen
INTRODUCTION
Broad-spectrum neuroactive pesticides have been for decades the major tool for controlling arthropod pests in agriculture, forestry, stored-products, and in public health. Toxicological and environmental problems posed by the massive and widespread use of such nonselective pest control agents are amply documented. Moreover, the increasing awareness of the public to hazards associated with toxic residues in drinking water and edible crops has become a considerable stimulus to devise alternative pest control measures including more selective pesticides. Among the selective targets available in arthropods for pesticide interference, only the hormonal balance and the synthesis of the aminopolysaccharide chitin yielded commercial products.
Chitin is peculiar among other targets by being an abundantly produced extrxellular biopolymer [ 11 deposited as orderly oriented microfibrils notably in exoskeletons of arthropods and in cell walls of most fungi species. Its main function is to contribute strength and rigidity to structural elements of the organism. Being part of suprastructures such as cuticles, peritrophic mem- branes, or cell walls, any interference with chitin deposition or its untimely degradation is detrimental to the organisms involved.
A number of reviews concerning the biochemistry of chitin and its relevance as a target for pesticide activity have been recently published [2-6]. The present study will examine the complexity of cellular events leading to formation of the end product of chitin, e.g., the microfibrillar crystallite and the possible sites for interference throughout the pathway of chitin formation and deposition. Chitin degradation as a target for pest control by either enhancement or inhibition of the hydrolytic enzymes involved will also be considered.
CHITIN: PROPERTIES, FUNCTION, OCCURRENCE
Chitin is a straight chain homopolymer composed of p( 1,4)-linked GlcNAc units (Fig. 1) with a three-dimensional a-helical configuration stabilized by intramolecular hydrogen bonding [7]. The chains coalesce at the cell mem- brane surface via intermolecular hydrogen bonding to form insoluble crystal- line microfibrils. X-ray diffraction analysis of chitin crystallites revealed two major polymorphs. a-Chitin, in which the polysaccharide chains in the microfibril are arranged in an antiparallel pattern [B], is the most abundant crystallite polymorph in nature. A @-chitin polymorph, which was observed in limited cases, is characterized by a parallel arrangement of the chains [9].
0 H,?.
R: N CHJ- CHITIN
n R = NH2- R
CHITOSAN
Fig. 1. Dirner units of chitin and chitosan polymers.

Chitin Metabolism as Target for Pesticides 247
The occurrence of y-chitin [lo] where the arrangement of the crystallite alternates so that for every three polymer chains two are parallel, is still debatable. Chitin, in particular the a-polymorph, is chemically stable and is insoluble in water and most organic solvents. It is degradable in very strong acid solutions or in concentrated hot alkali.
Chitosan (Fig. 1), which is a P(1-4)-linked D-glucosamine polymer formed by enzymatic deacetylation of chitin (GlcNAc), can be found as a structural component of the cell wall of various fungi, particularly in Zygomycetes [ll] as well as in arthropod cuticles [12]. The importance of chitosan and its oligomers in plant-pathogen interactions will be discussed later.
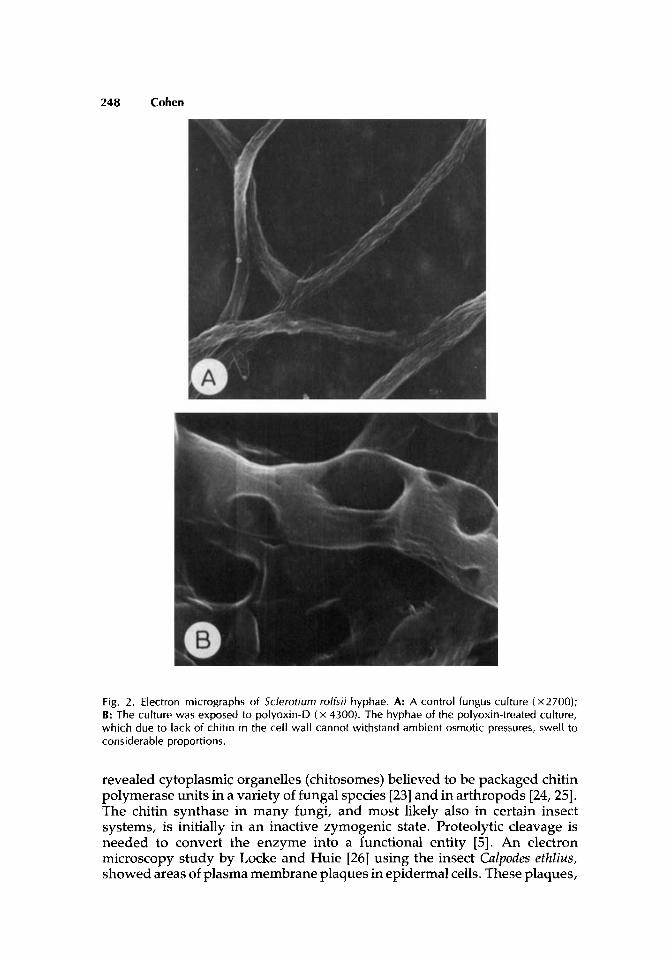
Chitin microfibrils, except in certain centric diatom algae [13], are covalently or noncovalently associated with various proteins (amoebae, arthropods, nematodes) [12] or polysaccharides (fungi) [14]. Such associations largely compose the supramolecular structures of cuticles and peritrophic membranes of arthropods, cell walls, and septa of fungi and yeast; also cyst walls in certain amoebae or egg shells of nematodes. A unique combination between chitin and keratin was recently reported in setae of a brachiopod species [15]. The importance of chitin as an integral part of the above supportive structures can be demonstrated by specific inhibition of its formation. Lack of chitin results in weak insect exoskeletons, difficulties in molting, bleeding, and mortality due to dehydration [16]. Inhibition of chitin synthesis in fungi leads to fragile cell walls which cannot withstand ambient osmotic pressures. As a result swollen and blown-up hyphae are often observed (Fig. 2) [17].
Chitin has become an attractive target for pest control since it occurs in invertebrates, notably arthropods, and in cell walls of most fungi except the Oomycetes where cellulose functions as the supportive component. How- ever, all vertebrates and vascular plants use alternative polymers (cellulose, collagen) as supportive components. The differential distribution of chitin among organisms (Table 1) gives the basis for the selectivity of pest control agents acting by interference with chitin metabolism as their primary mode of action. It should be stressed, nevertheless, that the above selectivity is limited since some nontarget aquatic arthropods [18-201 or beneficial preda- tors [21,22] were reported to be affected by commercial pesticides which act as inhibitors of chitin synthesis.
TARGETS FOR INHIBITION
Chitin Synthesis From Sugar Precursors to Polymer
The pathways of chitin biosynthesis and degradation, which consist of catalytic units, precursors, biotransformations, polymerization, fibrillogene- sis, and hydrolysis, are depicted in Figure 3 . The unequivocal commitment to chitin synthesis occurs at the membrane-bound polymerizing system (chitin synthase) site. Apparently, catalytic units synthesized at the endoplasmic reticulum compartment are packaged in clusters which involve the Golgi apparatus. Subsequently, the assembled multiunits are translocated and integrated into the plasma membrane at the apical compartments of epidermal cells of arthropods or at tips of fungal mycelium. Electron microscopy studies
248 Cohen
Fig. 2. Electron micrographs of Sclerotiurn rolfsii hyphae. A: A control fungus culture (X2700); B: The culture was exposed to polyoxin-D ( x 4300). The hyphae of the polyoxin-treated culture, which due to lack of chitin in the cell wall cannot withstand ambient osmotic pressures, swell to considerable proportions.
revealed cytoplasmic organelles (chitosomes) believed to be packaged chitin polymerase units in a variety of fungal species [23] and in arthropods [24,25]. The chitin synthase in many fungi, and most likely also in certain insect systems, is initially in an inactive zymogenic state. Proteolytic cleavage is needed to convert the enzyme into a functional entity [ 5 ] . An electron microscopy study by Locke and Huie [26] using the insect Calpodes efhZius, showed areas of plasma membrane plaques in epidermal cells. These plaques,

Chitin Metabolism as Target for Pesticides 249
TABLE 1. Phylogenetic Occurrence of Chitina
Fungi (cell wall) Amoebae (cyst wall) Diatom algae Rotifers (egg shell) Nematodes (egg shell) Anthropods (cuticle, peritrophic membrane) Molluscs (radula, shell) Sauids
aEstimated annual production of chitin: 10'o-lO" tons.
which presumably represent membrane-embedded clusters of chitin poly- merase units, underwent hormonally-controlled cyclic turnovers.
To provide a flow of substrate molecules for the functional catalytic units, a cascade of cytoplasmic biotransformations of sugar precursors takes place. Trehalose, glucose, or the polysaccharide glycogen could be the starting points for a chain of bioconversions which include major steps such as phosphorylation, amination, and acetylation (Fig. 3). This metabolic pathway culminates in the formation of UDP-GlcNAc, the ultimate substrate of chitin synthase. Transformation of fructose-6-phosphate to glucosamine-6-phos- phate by glutamine-fructose-6-phosphate aminotransferase can be considered as the first turning point on the pathway to chitin synthesis. Indeed, the
CHITIN SYNTHASE PRECURSORS synthesis biotransfomation: translocation phosphorylation insertion amination activation acetylation
turnover +
UDP-GlcNAc formation
POLYMERIZATION + +
HYDROGEN BOND FORMATION (crystallization, fibrillogenesis)
ORIENTATION OF MICROFIBRILS (cytoskeleton components?)
t SUPRAMOLECULAR ORGANIZATION
(covalent and noncovalent interactions) (with proteins, polysaccharides)
DEGRADATION (chitinase, chitobiase, chitodextrinase)
RECYCLING MINERALIZATION
Fig. 3 . Chitin metabolism.

250 Cohen
antibiotic bacilysin (tetaine), which is a specific inhibitor of the above enzyme, interfered with chitin biosynthesis in the fungus Candida ulbicans [27]. How- ever, since glucosamine-6-phosphate can still be diverted to and used by other metabolic pathways, inhibition of the amination step by the antibiotic com- pound is not specific with respect to chitin synthesis.
Chitin Synthase Catalytic Activity and Inhibition
The most extensively investigated target in chitin metabolism is the poly- merizing enzyme chitin synthase. Using the fungus Neurosporu crassu Glaser and Brown [28] back in 1957 were the first to demonstrate chitin polymeriza- tion in a cell-free system. Later, the enzyme was obtained from numerous filamentous fungi and yeast species [5,14]. Due to methodological difficulties, cell-free preparations from insects were hard to obtain for a long time. The first reports describing insect chitin synthases from the Coleopteran TriboZium custaneum [29] and the Dipteran Stomoxys calcitruns [30] were published in 1980, Since then, polymerizing enzyme activity has been demonstrated in one crustacean [31] and 5 additional insect species [32-351. In insect and fungal systems the polymerization reaction for in vitro chitin synthesis is usually carried out in simple buffer solutions containing divalent cations like magne- sium and the (radioactive) substrate, UDP-GlcNAc [3,5]. Largely, both en- zyme systems share similar properties such as the stimulatory response to the allosteric effector glucosamine, inhibition by nucleoside peptides and UDP, and lack of requirement for a primer or a lipid intermediate for their catalytic activity [3,5]. Unlike the insect chitin synthase, most of the fungal enzyme preparations, being zymogenic in nature, require proteolytic activation and are apparently insensitive to the fungicide captan [36].
Chitin synthase from the yeast Succharomyces cerevisiue was isolated and purified using the detergent digitonin [37]. At present, three different yeast chitin synthase enzymes (Chs 1,2,3) with different functions and their corre- sponding genes (CHS 1,2,3) were reported [3842]. Chitin synthase genes of the zoopathogen C. albicans [43] and of the filamentous fungus N . crussa [44], were recently cloned and sequenced. At present, no insect chitin synthase gene has been cloned. Based on differences in properties of chitin synthases [29,32,36], the existence of at least separated genes coding for integumental and peritrophic membrane enzymes can be anticipated.
The first documented inhibition of chitin biosynthesis involved nucleoside peptide metabolites (Fig. 4) isolated from actinomycetes. Resembling the structure of the chitin synthase substrate, UDP-GlcNAc, they act as potent competitive inhibitors of the enzyme. These structurally-related antibiotics are the polyoxins (A-M) [45] produced by cultures of Streptomyces cacaoi var. asoensis and nikkomycins ( Z , X, J, I [46] and KZ, K,, Q, 0, [47]) which are metabolites of Streptomyces tendue. Recently, two additional nucleosides re- lated to nikkomycins (Wz, W,) were obtained from the culture broth of a fluorotyrosine-resistant strain of S. tendue [48]. It was suggested that the uridyl moiety, which is shared by the nucleoside antibiotics and the UDP-GlcNAc substrate, attaches to the binding site of the polymerizing enzyme [49,50]. The catalytic site of the enzyme, suggested to be hydrophobic in nature, appar- ently binds to the dipeptidyl (nikkomycins) or the tripeptidyl (polyoxins) part

Chitin Metabolism as Target for Pesticides 251
I H
I H2N-CH
POLYOXIN - b NIKKOMYCIN- Z
Fig. 4. Chemical structures of nucleoside peptide inhibitors.
of the corresponding inhibitors. Most likely, the nucleoside peptides, being of relatively high molecular weight, enter the fungal cells via a dipeptidyl or tripeptidyl transport system [52,53]. Polyoxins have been commercialized and reported to be used in Japan for the control of a limited number of phy- topathogens causing diseases in fruit trees and vegetables [53] . Developing the nikkomycins as pesticides has been discontinued by Bayer company. The Streptomyces antibiotics are potent inhibitors of insect chitin synthase [3], yet they display poor insecticidal activity due to polar properties, relatively high molecular weights, and perhaps lack of appropriate transport systems in insect cells.
Inhibition of arthropod chitin synthases by several toxicants such as plum- bagin [54], buprofezin [55], avermectin [56], and terpenoyl benzimidazoles [57] was reported. Since relatively high levels of the above compounds are required for inhibitory effects, their action was regarded as nonspecific, A large number of pesticides and other toxicants were reported to inhibit the polymerizing enzyme extracted from the fungus Phycomyces [58]. Their actions were attributed to a general disruption of the plasma membrane lipid bilayer, nonspecifically affecting the membrane-bound chitin synthase [59].
Chitin From Polymer to Microfibril
Based on studies with fungal chitin synthase, the membrane-bound en- zyme is most likely exposed to the cytoplasm [60-62]. Apparently, the insect polymerase obeys the same topography. However, studying cultured insect integuments, Mitsui and coworkers [63,64] claimed that the catalytic site faces the outer cell surface. Consequently, such a location invokes a transport system for the high molecular weight UDP-GlcNAc substrate which cannot diffuse through the plasma membrane. No evidence for such sidedness or proof for a transport system was provided. Electron microscopic studies of cell membranes using freeze-fracture techniques showed structures of rosettes and globules, which were suggested to be part of the cellulose biosynthesis system in algae and higher plants [65] . The molecular organization of the chitin synthase units in the cell membrane and the intricate processes which involve the translocation of chitin polymers across this barrier are mysterious. The energy required for the movement or translocation of the chitin polymers may be provided by the formation of 1,4-P-glycoside bonds or by the crystal-

252 Cohen
DIFLUBENZURON (Dimilin)
Fig. 5 . Chemical structure of a representative benzoylaryl urea compound.
lization process. The implication of different membrane proteins in transloca- tion of chitin polymers has been suggested [59]. The involvement of auxiliary membrane protein or proteins having lectin-like properties in the transloca- tion of chitin polymers is at present speculative. It is nevertheless tempting to speculate that the mode of action of the insecticidal benzoylaryl ureas (Fig. 5) is related to inhibition of such proteins. This group of commercial insecti- cides, which are highly potent inhibitors of chitin synthesis in insects, but not in fungi, does not inhibit the chitin polymerase. The exact biochemical lesion inflicted by these inhibitors is not known. Among the suggestions forwarded were 1) enhanced activities of phenoloxidase and chitinase [66]; 2) inhibition of proteases which convert the inactive zymogenic chitin synthase into an active enzyme [67]; 3) inhibition of DNA synthesis [68]; 4) inhibition of influx of nucleosides [69]; 5) interference with the insect hormonal balance [70]; and 6) inhibition of the speculative GlcNAc transporter [64]. Some of the suggested mechanisms of action such as 1, 3, and 4 have been regarded as secondary effects [3]. Hypotheses 2 and 5 were refuted since an on-going chitin synthesis is rapidly blocked [71,72] and the insect endocrine system is not affected by benzoylaryl ureas [72-741.
The last step in chitin biotransformation involves the formation of micro- fibrils at the cell surface, following the coalescence of adjacent nascent chitin chains via extensive hydrogen bonding. Disrupting these hydrogen bonds by dyes such as Calcofluor white, Congo red, and Primulin inhibited chitin synthesis in a number of fungal systems [75-791, insects [59,80], and a diatom alga [HI.
Albeit chitin polymerization and crystallization are coupled, they are con- secutive processes, evidence of which emerged from experiments using dye compounds such as Calcofluor white and Congo red [38,79]. It has been reported that chitin synthase activity was stimulated in the yeast S. cerevisiue [38] and the flour beetle T. castaneum [82] by Calcofluor white and chito- oligosaccharide-binding lectins, respectively. In the case of GlcNAc-binding lectins, it was suggested [82] that disruption of polymerization and detach- ment of polymers from the catalytic sites were followed by renewed biosyn- thesis of chitin polymers. Presumably, the observed stimulatory effect was due to a more efficient and rapid incorporation of the radioactive substrate into newly-synthesized shorter chitin polymers.
The characteristic helicoidal pattern of the arthropod cuticle suprastructure may indicate some sort of association between the cytoskeletal components of the epidermal cells and the membrane chitin synthase units. This probable connection also invokes an appreciable fluidity of the plasma membrane to

Chitin Metabolism as Target for Pesticides 253
facilitate mobility of the embedded chitin synthase clusters. Such mobility may account for changes in orientation of the chitin microfibrils and, thus, for the helicoidal appearance. It is worthy to note that the fungicide benomyl as well as colchicine and vinblastine, known to disrupt cytoskeletal microtubular proteins, inhibited chitin synthesis in yeast [77] and in imaginal disks of an insect species [83].
In summary, throughout the pathway of chitin synthesis from simple precur- sors to microfibrils, a number of target sites for pest control can be exploited. Practically, the catalytic event is susceptible to strong inhibition by structural analogs of the substrate. There is no experimental proof, but it is plausible that post-catalytic events associated with translocation of chitin polymers across the cell membrane are perturbed by insecticidal benzoylaryl ureas.
Chitin Degradation
Chitin degradation involves a joint action of an endo enzyme, chitinase, which yields oligomers, and an exo enzyme, P-N-acetyl-glucosaminidase, which hydrolyzes dimers or terminal (GlcNAc), polymers [84]. A third enzyme (chitodextrinase), which hydrolyzes small chitooligosaccharides, has been recently reported in a chitinolytic marine bacterium [85]. Since chitin microfibrils are tightly associated with various proteins (arthropods) or poly- saccharides such as glucan (fungi), proteolytic or glucanase activities often accompany and facilitate chitin hydrolysis.
Degradation of chitin is physiologically necessary to maintain normal hyphal growth and branching in filamentous fungi, and septum formation in yeast [86]. It has been shown that the dual action of chitin synthesis and degradation exists at the hyphal tips. Hydrolysis of chitin at specific sites on the hypha is a prerequisite for branching. Chitin degradation also facilitates spore release and germination [86]. The cyclic phenomenon of molting in arthropods is highly complex and involves a concerted and coordinated flow of hormonally-controlled biochemical events [84]. One such major event is the degradation of the endocuticular chitin, followed by the subsequent recycling of the aminosugar monomers into newly-formed polymers. The normal chitin degradation, be it in insects or fungi, could serve as a prime target site for interfering with essential biochemical and physiological pro- cesses. Indeed, in addition to organisms which form chitin (Table 2), chitin degradation systems have been evolved in nonchitinous organisms such as in prokaryotic microflora, certain vertebrates, and vascular plants [12,86]. Chitin is a nutritional source for chitinolytic microorganisms [12], crustacean-
TABLE 2. Phylogenetic Occurrence of Chitin Metabolic Svstems
Chitin synthesis Chitin degradation
Fungi Fungi Protozoa Protozoa Nematodes Nematodes Invertebrates Invertebrates
Bacteria Vertebrates Vascular ulants

254 Cohen
N CH3 OH
ALLOSAMIDIN
Fig. 6. Chemical structure of a chitinase inhibitor.
eating fish [87], and in insectivorous plants [88]. Mycopathogens [89,90] and entomopathogens [91] use their respective chitinolytic systems to penetrate arthropod cuticules and fungal cell walls.
Plants contain inducible systems which react to invasion of pathogens [92] inter alia by de novo synthesis of pathogenesis-related (PR) proteins. Among these proteins are glucanases, chitinases, and chitosanases, believed to act as a defense mechanism against phytopathogenic fungi [93-961. There is no direct evidence for the protective role of chitinase. However, recent studies in which the expression of the chitinase genes in the plant was amplified, should confirm the above role. It is noteworthy that externally applied chitosan oIigomers functioned as elicitors of chitinase formation in pea [97]. It was also suggested that chitosan acts as a transducing signal in the interaction between a gall-producing mite and a plant [98]. Moreover, exter- nally applied polycationic chitosan or chitosan oligomers, which have high affinity for DNA, were detected within plant cell nuclei [99]. This observation is a clear indication that the above polysaccharides act at the genomic level to induce PR proteins.
Only recently allosamidin (Fig. 6) and desmethylallosamidin, two metabo- lites isolated from a Streptomyces sp., were reported to strongly and specifically inhibit insect chitinase and disrupt insect ecdysis [loo, 1011, These competitive inhibitors of chitinase are dimers of the aminosugar P-N-acetylallosamin attached to an aminocyclitol moiety. It was observed that while allosamidin is not effective against bacterial and plant chitinases, it dispIays appreciable inhibitory effects against chitinases of a nematode species [lo21 and the fungus N. CYUSSU [103]. Currently, the prospects for developing the allosamidins or similar effective products as commercial pesticides are unclear. However, the possibility of using chitin degradation as a new target for pest control is attractive.
CONCLUSIONS
In principle, the elaborate processes which involve the biosynthesis of chitin and its degradation should offer a number of interesting selective targets for the control of insect pests as well as phyto and zoopathogens. So far in practice, only the chitin polymerization step was proven to be a useful target and has been exploited. The polyoxins, which have been developed in Japan, reached the status of commercial fungicides, and are being applied on a limited scale. Since the introduction of diflubenzuron, a number of potent

Chitin Metabolism as Target for Pesticides 255
analogs of the insecticidal benzoylaryl urea group have been developed and registered for commercial application. The use of these insect growth regula- tors is now widespread for the control of a variety of pests. Being selective to an appreciable degree, they are excellent candidates to be incorporated in integrated pest management (IPM) programs [104,105].
LITERATURE CITED
1. Gooday GW: The ecology of chitin degradation. In: Advances in Microbial Ecology. Marshall KC, ed. Plenum, New York, vol11, pp 387430 (1990).
2. Gooday GW: Inhibition of chitin metabolism. In: Biochemistry of Cell Walls and Mem- branes in Fungi. Kuhn FJ, Trinci APJ, Goosey MW, Copping LG, eds. Springer-Verlag, Berlin, pp 61-79 (1990).
3. Cohen E: Chitin biochemistry: Synthesis and inhibition. Annu Rev Entomol32, 71 (1987).
4. Cohen E: Extracellular biopolymers as targets for pest control. In: Pesticides and Alterna- tives. Casida JE, ed. Elsevier, Amsterdam, pp. 23-32 (1990).
5. Cabib E: The synthesis and degradation of chitin. In: Advances in Enzymology and Related Areas of Molecular Biology. Meister A, ed. John Wiley, New York, pp 59-101 (1987).
6. Mayer RT, Cunningham G, Gupton J: Insecticides based on differences in metabolic pathways. In: Safer Insecticides, Development and Use. Hodgson E, Kuhr RJ, eds. Marcel Dekker, New York, pp 209-251 (1990).
7. Carlstrom D: The crystal structure of chitin (poly-N-acetyl-glucosamine). J Biophys Bio- them Cytol3, 669 (1957).
8. Minke R, Blackwell J: The structure of a-chitin. J Mol Biol 120, 167 (1978).
9. Gardner KH, Blackwell 1: Refinement of the structure of P-chitin. Biopolymers 14, 547 (1975).
10. Rudall KM, Kenchington W: The chitin system. Biol Rev48, 597 (1973).
11. Bartnicki-Garcia S: Cell wall chemistry, morphogenesis and taxonomy of fungi. Annu Rev Microbiol22, 87 (1968).
12. Muzzarelli RAA: Chitin. Pergamon Press, Oxford, 309 pp (1977).
13. Herth W, Zugenmaier P: Ultrastructure of the chitin fibrils of the centric diatom Cyclofella crypfiea. J Ultrast Res61, 230 (1977).
14. Gooday GW, Trinci APJ: Wall structure and biosynthesis in fungi. In: Eukaryotic Microbial Cell. Gooday GW, Lloyd D, Trinci APJ, eds. 30th Symposium of Society for General Microbiology. Cambridge University Press, Cambridge, England, pp 207-251 (1980).
15. Tanaka K, Katsura N, Saku T, Kasuga S: Composite texture of chitin and keratin in an animal organ, Lingda seta. Polymer J 20, 119 (1988).
16. Verloop A, Ferrell CD: Benzoylphenyl ureas-A new group of larvicides interfering with chitin deposition. In: Pesticide Chemistry in the 20th Century. Plimmer JR, ed. ACS Symposium, Series 37. American Chemical Society, Washington, DC, pp 237-270 (1977).

256 Cohen
17. Endo A, Kakiki K, Misato T: Mechanism of action of the antifungal agent polyoxin D. J Bacteriol104, 189 (1970).
18. Apperson CS, Schaeffer CH, Colwell AE, Werner GH, Anderson NL, Dupras EF, Long- anecker DR: Effects of diflubenzuron on Chaoborus astictopus and nontarget organisms and persistence of diflubenzuron on lentic habitats. J Econ Entomol71, 521 (1978).
19. Farlow JE, Breaud TP, Steelman CD, Schilling PE: Effects of the insect growth regulator diflubenzuron on non-target aquatic populations in a Louisiana intermediate marsh. Environ Entomol7, 199 (1978).
20. Muzzarelli R: Chitin synthesis inhibitors: Effects on insects and on nontarget organisms. Crit Rev Environ Control 16, 141 (1986).
21. Broadbent AB, Pree DJ: Effects of diflubenzuron and BAY SIR 8514 on beneficial insects associated with peach. Environ Entomol 23, 133 (1984).
22. Peleg BA: Effect of 3 insect growth regulators on larval development, fecundity and egg viability of the coccinelid Chilororus bipusfulatus (Col.: Coccinellidae). Entomophaga 28,117 (1983).
23. Bartnicki-Garcia 5, Bracker CE, Reyes E, Ruiz-Herrera J: Isolation of chitosomes from taxonomically diverse fungi and synthesis of chitin microfibrils in vitro. Exp Mycol2, 173. (1978).
24. Mothes U, Seitz KA: Action of the microbial metabolite and chitin synthesis inhibitor nikkomycin on the mite Tetranychus urticae; and electron microscopy study. Pestic Sci 13, 426 (1982).
25. Cohen E: In vitro chitin synthesis in an insect: Formation and structure of microfibrils. Eur J Cell Bioll6, 284 (1982).
26. Locke M, Huie P: Apolysis and the turnover of plasma membrane plaques during cuticle formation in an insect. Tissue Cell 2 1 , 277 (1979).
27. Milewsky S, Chmara H, Borowsky E: Antibiotic tetaine-a selective inhibitor of chitin and mannoprotein biosynthesis in Candida albicans. Arch Microbiol145, 234 (1985).
28. Glaser L, Brown DH: The synthesis of chitin in cell free extracts of Neztrospora crassa. J Biol Chem228, 729 (1957).
29. Cohen E, Casida JE: Properties of Tribolium gut chitin synthetase. Pestic Biochem Physiol 13, 121 (1980).
30. Mayer RT, Chen AC, DeLoach JR: Characterization of a chitin synthase from the stable fly, Stomoxys calcitrans (L.). Insect Biochem 10, 549 (1980).
31. Horst MN: The biosynthesis of crustacean chitin by a microsomal enzyme from larval brine shrimp. J Biol Chem 256, 1412 (1981).
32. Cohen B, Casida JE: Properties and inhibition of insect integumental chitin synthetase. Pestic Biochem Physiol 2 7, 301 (1982).
33. Mitsui T, Nobusawa C, Tokuda K, Tada M Binding of UDP-N-acetylglucosamine to brush-border membrane vesicles of midgut epithelial cells of the cabbage armyworm, Mamestra brassicae. J Pestic Sci 11, 63 (1986).

Chitin Metabolism as Target for Pesticides 257
34. Turnbull IF, Howells AJ: Integumental chitin synthase activity in cell-free extracts of the Australian sheep blowfly, Lucilia cuprina, and two other species of Diptera. Aust J Biol Sci 36, 251 (1983).
35. Ward GB, Mayer RT, Feldlaufer MF, Svoboda JA: Gut chitin synthase and sterols from larvae of Diaprepes abbrmiatus (COkOptera: Curculionidae). Arch Insect Biochem Physiol 18, 105 (1991).
36. Cohen E, Casida JE: Inhibition of Tribolium gut chitin synthetase. Pestic Biochem Physiol 13, 129 (1980).
37. Kang MS, Elango N, Mattia E, Au-Young J, Robbins PW, Cabib E: Isolation of chitin synthetase from Saccharomyces cerevisiae. Purification of an enzyme by entrapment in the reaction product. J Biol Chem 259, 14966 (1984).
38. Bulawa CE, Slater M, Cabib E, Au-Young J, Sburlati A, Adair WL Jr., Robbins PW: The S. cerevisiae structural gene for chitin synthase is not required for chitin synthesis in vivo. Cell 46, 213 (1986).
39. Shaw JA, Mol PC, Bowers B, Silverman SJ, Valdivieso MH, Duran A, Cabib E: The function of chitin synthases 2 and 3 in the Sacchuromyces cerevisiae cell cycle. J Cell Biol124,lll (1991).
40. Orlean P: Two chitin synthases in Saccharomyces cerevisiae. J Biol Chem 262, 5732 (1987).
41. Valdivieso MH, Mol PC, Shaw JA, Cabib E, Duran A: CAL1, a gene required for activity of chitin synthase 3 in Saccharomyces cerevisiae. J Cell Biol 114, 101 (1991).
42. Bulawa CE, Osmond BC: Chitin synthase I and chitin synthase are not required for chitin synthesis in vivo in Saccharomyces cerevisiae. Proc Natl Acad Sci USA 87, 7424 (1990).
43. Au-Young J, Robbins PW: Isolation of a chitin synthase gene (CHS 1) from Candida albicans by expression in Saccharomyces cerevisiae. Mol Microbiol4, 197 (1990).
44. Yarden 0, Yanofsky C: Chitin synthase 1 plays a major role in cell wall biogenesis in Neurospora crassu. Genes Dev 5, 2420 (1991).
45. Isono K, Asahi K, Suzuki S: Studies of polyoxins, antifungal antibiotics. XII. The structures of polyoxins. J Am Chem SOC 91, 7490 (1969).
46. Dahn U, Hagenmaier H, Hohne H, Konig WA, Wolf GA, Zahner H: Stoffwechsel Produkte von Mikroorganisman. 154. Mitteilung. Nikkomycin, ein neuer Hemmstoff der Chitinsynthese bei Pilzen. Arch Microbiol 207,143 (1976).
47. Bormann C, Huhn W, Zahner H, Rathmann R, Hahn H, Konig WA: Metabolic products of microorganisms. 228. New nikkomycins produced by mutants of Streptomyces tendae. J Antibiot 38, 9 (1985).
48. Decker H, Walz F, Bormann C, Zahner H, Fiedler H-P, Heitsch H, Konig WA: Metabolic products of microorganisms. 255 Nikkomycins W, and W,, new chitin synthetase inhibitors from Streptomyces tendae. J Antibiot 43, 43 (1990).
49. Hori M, Kakiki K, Misato T: Interaction between polyoxin and active center of chitin synthetase. Agric Biol Chem 38, 699 (1974).
50. Misato T, Kakih K, Hori M: Chitin as a target for pesticide action: Progress and prospect. In: Advances in Pesticide Science. Geissbiihler H, Brooks GT, Kearney PC, eds. Pergamon Press, Oxford, pp 45W64 (1979).

258 Cohen
51. Payne JW, Shallow DA: Studies on drug targeting in the pathogenic fungus Cuiidrda albicans: Peptide transport mutants resistant to polyoxins, nikkomycins and bacilysin. E M S Microbiol Lett 28, 55 (1978).
52. Gonneau M, Yadan JC, Sarthou P, Le Goffic F: Nikkomycin Z as inhibitor of Candidu ulbicuns growth. In: Chitin in Nature and Technology. Muzzarelli R, Jeuniaux C, Gooday GW, eds. Plenum, New York, pp 203-205 (1986).
53. MisatoT Present status and future prospects of agricultural antibiotics. J Pestic Sci 7,301 (1982).
54. Kubo I, Uchida M, Klocke JA: An insect ecdysis inhibitor from the African medicinal plant, Plumbago cupensis (Plumbaginaceae); a naturally occurring chitin synthetase inhibitor. Agric Biol Chem 47, 911 (1983).
55. Uchida M, Asai T, Sugimoto T: Inhibition of cuticle deposition and chitin biosynthesis by a new insect growth regulator, buprofezin, in Nilupuruuta lugens Stlil. Agric Biol Chem 49, 1233 (1985).
56. Calcot PH, Fatig RO (111): Inhibition of chitin metabolism by avermectin in susceptible organisms. J Antibiot 37, 253 (1984).
57. Cohen E, Kuwano E, Eto M: The use of Triboliunz chitin synthetase assay in studying effects of benzimidazoles with a terpene moiety and related compounds. Agric Biol Chem 48, 1617 (1984).
58. Leighton T, Marks E, Leighton F: Pesticides: Insecticides and fungicides are chitin synthesis inhibitors. Science 213, 905 (1981).
59. Cohen E: Chitin biochemistry. In: Physiology of the Insect Epidermis. Retnakaran A, Binnington KC, eds. Inkata Press, North Layton, pp 96-114 (1991).
60. Sentandreu R, Martinez-Ramon A, Ruiz-Herrera J: Localization of chitin synthase in Mucor rouxii by an autoradiographic method. J Gen MicrobiolZ30, 1193 (1984).
61. Duran A, Bowers B, Cabib E: Chitin synthetase zymogen is attached to the yeast plasma membrane. Proc Natl Acad Sci USA 72, 3952 (1975).
62. Braun PC, Calderone RA: Chitin synthesis in Cundida albicans: Comparison of yeast and hyphal forms. J Bacteriol 133, 1472 (1978).
63. Mitsui M, Nobusawa C, Fukami J-I: Mode of inhibition of chitin synthesis by diflubenzuron in the cabbage armyworm Mumestru brassicue L. J Pestic Sci 9, 19 (1984).
64. Mitsui M, Tada M, Nobusawa C, Yamaguchi I: Inhibition of UDP-N-acetylglucosamine transport by diflubenzuron across biomembranes of the midgut epithelial cells in the cabbage armyworm Mamestru brassicae L. J Pestic Sci 10, 55 (1985).
65. Delmer DP: Cellulose biosynthesis. Annu Rev Plant Physiol38, 259 (1987).
66. Ishaaya I, Casida JE: Dietary TH-6040 alters composition and enzyme activity of housefly larval cuticle. Pestic Biochem Physiol4, 484 (1974).
67. Marks EP, Leighton T, Leighton F: Mode of action of chitin synthesis inhibitors. In: Insecticide Mode of Action. Coats J, ed. Academic Press, New York, pp 281-313 (1982).

Chitin Metabolism as Target for Pesticides 259
68. DeLoach JR, Meola SM, Mayer RT, Thompson JM: Inhibition of DNA synthesis by diflubenzuron in pupae of the stable fly Stomoxys culcitruns (L.). Pestic Biochem Physiol 15, 172 (1981).
69. Mayer RT, Netter KJ, Leising HB, Schachtschabel DO: Inhibition of the uptake of nucleosides in cultured Harding-Passey melanoma cells by diflubenzuron. Toxicology 30, 1 (1984).
70. Yu SJ, Terriere LC: Activities of hormone metabolizing enzymes in house flies treated with some substituted urea growth regulators. Life Sci 17, 619 (1975).
71. Deul DJ, DeJong BJ, Kortenbach JAM: Inhibition of chitin synthesis by two 1-(2,6-disubsti- tuted benzoyl)-3-phenylurea insecticides. 11. Pestic Biochem Physiol8, 98 (1978).
72. Hajjar NP, Casida JE: Structure-activity relationships of benzoylphenyl ureas as toxicants and chitin synthesis inhibitorsin Oncopeltlrsfusciatus. PesticBiochemPhysiol II, 699 (1979).
73. Cohen E, Casida JE: Insect chitin synthetase as a biochemical probe for insecticidal compounds. In: Pesticide Chemistry. Human Welfare and the Environment. Miyamoto J, Kearney PC, eds. Pergamon Press, Oxford, pp 25-32 (1983).
74. O'Neill MP, Holman GM, Wright JE: P-Ecdysone levels in pharate pupae of the stable fly Sfomoxys catcifrarzs and interaction with the chitin inhibitor diflubenzuron. J Insect Physiol 23, 1243 (1977).
75. Elorza MV, Rico H, Sentandreu R: Calcofluor white alters the assembly of chitin fibrils in Sacchuromyces cerevisiue and Cundidu ulbicuns cells. J Gen Microbiol 129, 1577 (1983).
76. Roncero C, Duran A: Effect of Calcofluor white and Congo red on fungal cell wall morphogenesis. In vivo activation of chitin polymerization. J Bacteriol 263, 1180 (1985).
77. Roncero C, Valdivieso MH, Ribas JC, Duran A: Isolation and characterization of Saccharorny- ces cerevisiue mutants resistant to calcofluor white. J Bacteriol 263, 1180 (1988).
78. Vermeulen CA, Wessels JGH: Chitin biosynthesis by fungal membrane preparation. Evidence for a transient non-crystalline state of chitin. Eur J Biochem 258, 411 (1986).
79. Selitrennikoff CP: Chitin synthetase activity of Neurosyora crussur Effects of primulin and congo red. Exp Mycol9, 179 (1985).
80. Zimmermann D, Peters W: Fine structure and permeability of peritrophic membranes of Calliphoru e ~ ~ ~ ~ r o c e p ~ u ~ u (Meigen) (Insecta: Diptera) after inhibition of chitin and protein synthesis. Comp Biochem Physiol86B, 353 (1987).
81. Herth W: Calcofluor white and congo red inhibit chitin microfibril assembly of Pooterio- ochromonas: Evidence for a gap between polymerization and microfibril formation. J Cell Biol87, 442 (1980).
82. Cohen E, Casida JE: Insect and fungal chitin synthetase activity: Specificity of lectins as enhancers and nucleoside peptides as inhibitors. Pestic Biochem Physiol37, 249 (1990).
83. Oberlander €4, Lynn DE, Leach CE: Inhibition of cuticle production in imaginal discs of Plodia interpunctellu (cultured in vitro): Effects of colcemid and vinblastine. J Insect Physiol 29, 49 (1983).
84. Kramer KJ, Koga D: Insect chitin. Physical state, synthesis, degradation and metabolic regulation. Insect Biochem 16, 851 (1986).

260 Cohen
85. Bassler BL, Yu C, Lee YC, Roseman S: Chitin utilization by marine bacteria. Degradation and catabolism of chitin oligosaccharides by Vibrio fumissii. J Biol Chem 266, 24276 (1991).
86. Gooday GW: Physiology of microbial degradation of chitin and chitosan. Biodegradation 1, 177 (1990).
87. Gooday GW: Chitinase activities in animals, fungi and bacteria. In: Chitin in Nature and Technology. Muzzarelli R, Jeunieux C, Gooday GW, eds. Plenum, New York, pp 241-261 (1986).
88. Amagase S, Mori M, Nakayama S: Digestive enzymes in insectivorous plants. IV. Enzy- matic digestion of insects by Nepenthes secretion and Drosera peltata extract: Proteolytic and chitinolytic activities. J Biochem 72, 765 (1972).
89. Chet I, Cohen E, Elster I: The role of chitinase and chitin synthetase inhibitors in controlling plant pathogenic fungi. In: Chitin in Nature and Technology. Muzzarelli R, Jeuniaux C, Gooday GW, eds. Plenum, New York, pp 237-240 (1986).
90, Manoncha MS: Cellular and molecular aspects of fungal host-mycoparasite interactions. Z Pflanzenkr Pflanzenschutz 94, 432 (1987).
91. Coudron TA, Kroha MJ, Ignoffo CM: Levels of chitinolytic activity during development of three entomopathogenic fungi. Comp Biochem Physiol798,339 (1984).
92. Linthorst HJM: Pathogenesis-related proteins in plants. Crit Rev Plant Sciences 10, 123 (1991).
93. Boller T: Chitinase: A defense of higher plants against pathogens. In: Chitin in Nature and Technology. Muzzarelli R, Jeuniaux C, Gooday GW, eds. Plenum, New York, pp 223-230 (1986).
94. Mauch F, Hadwiger LA, Roller T: Antifungal hydrolases in pea tissue. I. Purification and characterization of two chitinases and two p -1,3-glucanases differentially regulated during development and in response to fungal infection. Plant Physiol87, 325 (1988).
95. Rousseau-Limouzin M, Fritig B: Induction of chitinases, 1,3-P-glucanases and other patho- genesis-related proteins in sugar beet leaves upon infection with Cercospora beticola. Plant Physiol Biochem 29, 105 (1991).
96. Grenier J, Asselin A: Some pathogenesis-related proteins are chitosanases with lytic activity against fungal spores. Mol Plant Microbe Interac 3,401 (1990).
97. Kendra DF, Christian D, Hadwiger LA: Chitosan oligomers from Fusarium solanilpea interactions, chitinase/P-glucanase digestion of sporeling and from fungal wall chitin activity inhibit fungal growth and enhance disease resistance. Physiol Molec Plant Pathol 35, 215 (1989).
98. Bronner R, Westphal E, Dreger F: Chitosan, a component of the compatible interaction between Solanum dulcamara L. and the gall mite Eriophyes cladophthrius Nal. Physiol Molec Plant Pathol34, 117 (1989).
99. Hadwiger LA, Kendra DF, Fristensky BW, Wagoner W: Chitosan both activates genes in plants and inhibits RNA synthesis in fungi. In: Chitin in Nature and Technology. Muzza- relli R, Jeuniaux C , Gooday GW, eds. Plenum, New York, pp 209-214 (1986).
100. Koga A, Isogai A, Sakuda S, Matsumoto S, Suzuki A, Kimura S, Ide A: Specific inhibition of Bombyx mori chitinase by allosamidin. Agric Biol Chem 52, 471 (1987).

Chitin Metabolism as Target for Pesticides 261
101. Sakuda S, Isogai A, Makita T, Matsumoto S, Koseki K, Kodama H, Suzuki A: Structures of allosamidins, novel insect chitinase inhibitors, produced by actinomycetes. Agric Biol Chem 51, 3251 (1987).
102. Gooday GW, Brydon LJ, Chappell LH: Chitinase in female Onchocercu gibsoni and its inhibition by allosamidin. Mol Biochem Parasitol29, 223 (1988).
103. McNab R, Glover LA: Inhibition of Neurosporu Crussa cytosolic chitinase by allosamidin. FEMS Microbiol Lett 82, 79 (1991).
104. Retnakaran A, Wright JE: Control of insect pests with benzoylphenyl ureas. In: Chitin and Benzoylphenyl Ureas. Retnakaran A, Wright JE, eds. Junk, Dordrecht, pp 205282 (1987).
105. Granett J: Potential of benzoylphenyl ureas in integrated pest management. In: Chitin and Benzoylphenyl Ureas. Retnakaran A, Wright JE, eds. Junk, Dordrecht, pp 283-302 (1987).