chemical structure and antigenic aspects of complexes obtained from epimastigotes of trypanosoma...
TRANSCRIPT

4980 Biochemistry 1983, 22, 4980-4987
Doyle, D., Chalovich, J. M., & Barany, V. M. (1981) FEBS Lett. 131, 147.
Edwards, R. H. T., Dawson, M. J., Wilkie, D. R., Gordon, R. E., & Shaw, D. (1982) Lancet, 725.
Gadian, D. G., Radda, G. K., Richards, R. E., & Seeley, P. J. (1979) in Biological Applications of Magnetic Resonance (Shulman, R. G., Ed.) p 463, Academic Press, New York.
Gordon, R. E., Hanley, P. E., Shaw, D., Gadian, D. G., Radda, G. K., Styles, P., Bore, P. J., & Chan, L. (1980) Nature (London) 287, 736.
Gordon, R. E., Hanley, P. E., & Shaw, D. (1982) Prog. Nucl. Magn. Reson. Spectrosc. 15, 1.
Hamilton, J. A., Talkowski, C., Childers, R. F., Williams, E., Allerhand, A., & Cordes, E. H. (1974) J . Biol. Chem. 249, 4872.
Hanley, P. E., & Gordon, R. E. (1981) J . Magn. Reson. 45, 520.
Hoult, D. I. (1978) Prog. Nucl. Magn. Reson. Spectrosc. 12, 41.
Norton, R. S. (1981) Bull. Magn. Reson. 3, 29. Ross, B. D., Radda, G. K., Gadian, D. G., Rocker, G., Esisri,
M., & Falconer-Smith, J. (1982) N . Engl. J . Med. 304, 1338.
Scott, A. I., & Baxter, R. L. (1981) Annu. Reu. Biophys. Bioeng. 10, 15 1.
Sears, B. (1975) J . Membr. Biol. 20, 59. Sharp, R . R., & Richards, E. P. (1977a) Biochim. Biophys.
Acta 497, 14. Sharp, R. R., & Richards, E. P. (1977b) Biochim. Biophys.
Acta 497, 260. Shulman, R. G., Brown, T. R., Ugurbil, K., Ogawa, S., Cohen,
S. M., & den Hollander, J. A. (1979) Science (Washington, D.C.) 205, 160.
Sillerud, L. O., & Shulman, R. G. (1983) Biochemistry 22, 1087.
Ugurbil, K., Shulman, R. G., & Brown, T. R. (1979) in Bi- ological Applications of Magnetic Resonance (Shulman, R. G., Ed.) p 537, Academic Press, New York.
Chemical Structure and Antigenic Aspects of Complexes Obtained from Epimastigotes of Trypanosoma cruzi? Lucia MendonGa-Previato, Philip A. J. Gorin,* Arnaldo F. Braga, Julio Scharfstein, and JOSE 0. Previato
ABSTRACT: Cells of Trypanosoma cruzi, Y strain, were sub- mitted to water extraction by successive freezing and thawing. Fractionation of soluble material on a P-10 column gave an antigenic glycoprotein (fraction I) whose carbohydrate portion (40%) contained galactose, mannose, glucose, and xylose in a molar ratio of 35: 13: 1 : 1. It was electrophoretically homo- geneous ( M , -25 000) and contained short chains of man- nopyranosyl (23%) and galactopyranosyl (10%) nonreducing end units and 2-0-substituted mannopyranosyl units (1 9%). Extraction of the remaining cell fragments with phenol-water gave an antigenic CHC1,-MeOH-H,O-soluble fraction I1 (LPPG), which yielded a galactomannan (fraction 111) (ga- lactose and mannose in a molar ratio of 1:2,1) on degradation with hot aqueous NaOH-NaBH,. It contained end units of
C h a g a s ’ dissease is a complex clinical entity caused by the flagellate protoazoan Trypanosoma cruzi. This protozoan has a complex life cycle with developmental stages in different hosts (Brener, 1973). In the mammalian host, T . cruzi multiplies intracellularly as amastigotes and is subsequently released into the bloodstream as trypomastigotes. These are nondividing forms that can infect new host cells or be ingested by a triatomine bug. In the insect midgut lumen, trypomas- tigotes differentiate to epimastigotes, which are the invertebrate multiplying forms. In the insect rectum, epimastigotes dif- ferentiate to the nondividing metacyclic trypomastigotes, which
From the Departamento de Microbiologia Geral, Instituto de Mi- crobiologia, Universidade Federal do Rio de Janeiro, B r a d (L.M.-P., A.F.B., J.S., and J.O.P.), and Prairie Regional Laboratory, National Research Council, Saskatoon, Saskatchewan, Canada (P.A.J.G.). Re- ceived February 18, 1983; revised manuscript received May 25, 1983. N.R.C. Canada No. 21334. This work was supported by grants from the Conselho Nacional de Desenvolvimento Cientifico e Tecnologico (CNPq), the Conselho de Ensino e Pesquisa da UFRJ (CEPG), and the Financiadora de Estudos e Projetos (FINEP).
0006-2960/83/0422-4980$01.50/0
galactofuranose (25%) and 2-0- (5%), 3-0- (32%), and 2,3- di-0- (1 9%) substituted mannopyranosyl units. Galacto- furanose was directly linked (1-3) to mannopyranose and not via a phosphorodiester bridge. P-D-Ga?f-( 1-3)-Me-a-~- Manp, in contrast with Me-@+Galf, was effective in inhibiting the precipitin reaction between LPPG and antiserum raised against LPPG. Fraction IV, insoluble in CHC1,-MeOH-H,O, contained galactose and mannose in a ratio of 1.4:l. After degradation with hot aqueous NaOH-NaBH, it gave a product (fraction V) containing galactose and mannose in a 1:2 ratio. Methylation analysis showed it to differ from fraction I11 since it contained a high proportion of nonreducing end units (41%) and 2-0-substituted units (16%) of manno- pyranose.
are discharged in the feces and urine onto the vertebrate skin when the insect is feeding. Through skin discontinuities or through mucosal membranes, these forms can reach the blood circulation and thereafter penetrate cells, thus completing its biological cycle.
The clinical features of Chagas’ disease are highly variable and depend on the interaction of several different host and/or parasite factors (Tafuri, 1979). Studies on the cellular membrane of T. cruzi have been performed with the aim of obtaining a better understanding of some of those complex interactions.
Surface glycoconjugates are the membrane components that are currently receiving special attention. Snary & Hudson (1979) isolated a surface glycoprotein of molecular weight (M,) 90 000 that is present in epimastigote, trypomastigote, and amastigote forms and which was able to induce immu- noprotection against experimental infections in mice (Scott & Snary, 1979). However, Nogueira et al. (1981) found that only bloodstream trypomastigotes contain a surface glyco- protein of M , 90 000, whereas epimastigotes and trypomas-
Published 1983 by the American Chemical Society

C H E M I C A L S T R U C T U R E O F T R Y P A N O S O M A C R U Z I C O M P L E X E S V O L . 2 2 , N O . 2 1 , 1 9 8 3 4981
80 g of T. crua (epimastigote form)
freezing and thawing: dilution with cold water ( 2 x 300 mL);
residue
heat a t 100 ' C f o r 15 min; centrifugation (12009 for 20 min)
ext ract ion wi th phenol (45%) o t 7 5 'C for 10 min; cen t r i f ugo t ion (7009 f o r I h l
supernatant
residue supernutant lyophilized phenolic phase aqueous phase (39)
dia lys is and then lyophil ization: 630 mg; extraction with CHCI.-
supernatant residue (precipitation with 1 volume of methanol)
LPPG (60 mg) (Con A column chromatography)
retained material (40 mg) fraction I1 fraction I V
FIGURE 1: Fractionation scheme of methodology used in isolation of materials from cells of Trypanosoma cruzi.
tigotes from acellular cultures contain a glycoprotein with a molecular weight of 75 000.
Employing specific monoclonal antibodies raised against epimastigotes, Snary et al. (1981) isolated a minor glycoprotein ( M , 72 000) present in epimastigotes and metacyclic trypo- mastigotes. This glycoprotein induced immunoprotection in mice challenged with metacyclic trypomastigotes but did not protect when challenged with bloodstream forms (Snary & Ferguson, 1982). Of the complex, 52% consisted of carbohy- drate whose components were glucosamine, glucose, galactose, mannose, ribose, and the previously unobserved fucose. The protein component was rich in serine (Snary et al., 1981).
In other studies, direct chemical analysis was used to as- certain the chemical structure of carbohydrate-containing material obtained from T. cruzi. Aqueous chloral extracts of epimastigotes have been examined by Gonqalves & Yamaha (1969), who isolated an antigenic material containing glucose, galactose, mannose, glucosamine, and xylose. Aqueous alkali extracts of epimastigotes (Gorin et al., 1981) contained non- reducing end units of a-D-mannopyranose, D-galactopyranose, and 0-D-galactofuranose with short chains of 1-2-linked a-D-mannopyranosyl residues. Galactofuranosyl units were linked (1-3) to a-D-mannopyranosyl residues. Alves & Colli (1975) obtained, after phenol extraction, four different com- ponents with distinct electrophoretic mobilities. The three glycoprotein components contained, collectively, galacto- pyranosyl residues (Alves et al., 1979) and a lipopeptido- phosphoglycan (LPPG) consisting of galactose, mannose, and glucose in a molar ratio of 22:35:1, respectively, and also traces of glucosamine (Lederkremer et al., 1976). Galactosyl units were shown to be in the furanosyl form (Lederkremer et al., 1980).
In the present investigation, we report the partial chemical structures and the antigenic properties of the carbohydrate moieties of two molecules, isolated from epimastigote forms of T. cruzi, which are present in all developmental stages of the parasite. Another complex was also isolated and inves- tigated.
Materials and Methods Culture Conditions. Epimastigote forms of T. cruzi (Y
strain) were grown in 500-mL flasks containing agar plus rabbit blood as the solid phase and Brain-Heart Infusion (BHI, Difco) as the liquid phase. The latter (100 mL) was used to inoculate media (1 L) in 3-L Erlenmeyer flasks containing the
following components (in units of grams per liter): BHI, 37; folic acid, 0.02; hemin, 0.01, dissolved in M NaOH (1.0 mL). After 7 days at 25 OC with slow agitation (80 rpm), the cells were harvested by centrifugation, washed 3 times with 0.9% aqueous NaC1, and frozen at -20 OC.
Extraction and Purification of Glycoconjugates. Frozen cells were rapidly thawed, cold water was added, and the resulting suspension was removed by centrifugation. The resulting pellet was dispersed in cold water and the suspension recentrifuged. The combined supernatants were heated to 100 "C and lyophilized, and the residue was fractionated on a column (2 X 120 cm) of Bio-Gel P-10, giving fraction I. This and further procedures are summarized in Figure 1. The pellet remaining after centrifugation was extracted with aqueous phenol at 75 OC (Osborn, 1966) and the aqueous layer lyophilized. The residue was extracted by the method of Lederkremer et al. (1977) with CHCl3-MeOH-H2O (10:10:3 v/v/v) to provide soluble fraction 11. Insoluble material was dissolved in 0.018 M sodium phosphate buffer, pH 7.2 in aqueous 0.9% NaC1, and fractionated on a column (1 X 10 cm) of concanavalin A (Con A)-Sepharose 4B according to the method of Lloyd (1970). Bound material was eluted with aqueous 0.1 M methyl a-D-mannopyranoside in the same buffer to yield fraction IV.
Sodium Dodecyl Sulfate-Polyacrylamide Gel Electro- phoresis (SDS-PAGE). Electrophoresis was performed in a discontinuous buffer system (Laemmli, 1970) with a 7 5 1 5 % gradient in the concentration of the polyacrylamide gel. Gels were stained with Coomassie Brilliant Blue, specific for protein. The carbohydrate-containing bands were detected with the periodate-Schiff (PAS) reagent (Fairbanks et al., 1971).
Gel Column Chromatography. Columns of Bio-Gel P-4 (-400 mesh) and P-10 (200-400 mesh) were equilibrated and eluted with distilled water (for details, see figures).
Detection and Analysis of Sugar Components of Carbohy- drate-Containing Materials. These were hydrolyzed by using 3 M trifluoroacetic acid at 100 OC for 3 h. Following evap- oration of the hydrolysate, the residue was successively reduced with aqueous sodium borohydride and acetylated to yield alditol acetates, which were analyzed by gas-liquid chroma- tography (GLC) by the method of Sawardeker et al. (1965).
Methylation Analysis of Carbohydrate-Containing Mate- rials. These were methylated successively by the methods of Haworth (1915) and Kuhn et al. (1955). Per-0-methylated products were degraded to partially 0-methylated alditol

4982 B I O C H E M I S T R Y M E N D O N C A - P R E V I A T O E T A L .
acetates, which were identified and quantitated (Jansson et al., 1976). In the present investigation, GLC-mass spec- trometry (MS) was carried out with a Finnigan Model 4000 GLC-MS unit interfaced with an Incos 2300 data system. Electron-impact spectra were obtained respectively every 2 s, scanning from m l z 40 to m l z 420. GLC was performed on capillary columns (0.25-mm diameter X 30 m), one coated with OV-17 (Barreto-Bergter et al., 1981) and the other with OV-17 blended with OV-225 (1:3 ratio by weight) according to Gorin et al. (1982). Products were identified by their electron-impact MS spectra and characteristic retention times.
Carbon-1 3 Nuclear Magnetic Resonance (13C N M R ) Spectroscopy. NMR spectra were obtained from carbohy- drate-containing materials (25 mg of fraction I11 and 10 mg of fraction V) by using a Varian XL-100-15 spectrometer with Fourier transform from D 2 0 solutions (0.85 mL) of a com- pound contained in a coaxial glass cylinder fitting snugly within a 12-mm diameter X 20.3-cm tube maintained at 70 "C. The spectral width was 5000 Hz, the acquisition time 0.8 s, the pulse width 9.5 p s , and the number of transients 150000 (Gorin et al., 1977). Chemical shifts are expressed in 6 relative to the resonance of tetramethylsilane (Me,Si), obtained in a separate experiment.
Amino Acid Analyses. Protein (0.1 mg) was hydrolyzed in 0.5 mL of 6 M HCl at 110 "C for 18 h. The resulting amino acids were analyzed by using a Beckman Model 21 amino acid analyzer.
P-Elimination Reaction of Fraction Z (GP-25). Alkali- catalyzed p-elimination of serine- and threonine-linked car- bohydrates was performed on GP-25 (5 mg) in 0.1 M NaOH in the presence of 0.1 M NaBH, and a trace of NaBT, at 25 "C for 18 h. The solution was neutralized with acetic acid and concentrated to a small volume, and the products were fractionated on a column (0.8 X 50 cm) of Bio-Gel P-10. The resulting fractions were assayed for protein (Lowry et al., 1951), carbohydrate (Dubois et al., 1956), and radioactivity.
Alkaline Borohydride Treatment of Fractions ZI (LPPG) and ZV. This was carried out by using 0.1 M NaBH4 in 0.1 M NaOH for 5 h at 100 "C (Lee & Scocca, 1972). The reaction mixture was neutralized with acetic acid, and the products of degradation were fractionated by column chro- matography on Bio-Gel P-4.
Successive Sodium Periodate Oxidation and Sodium Bo- rohydride Reduction of Fraction ZZ. This fraction (LPPG) was treated with an excess of aqueous sodium periodate for 20 min and the reaction terminated by the addition of ethylene glycol. The product was reduced with sodium borohydride, and after 1 h, acetic acid was added and the solution dialyzed (Gorin & Spencer, 1959). The product contained arabino- furanosyl in place of galactofuranosyl units.
Other Analytical Methods. Phosphate was determined by the method of Ames (1 966) and hexosamine by a modification of the Elson-Morgan reaction (Belcher et al., 1954). Sialic acid was assayed by the thiobarbituric acid method (Warren, 1959).
Antisera. Anti T. cruzi sera were obtained by immunizing rabbits with lo8 epimastigote forms emulsified in complete Freund's adjuvant and injected twice, with an interval of 7 days by the intramuscular route. One week after the second in- jection, the rabbits were boosted 3 times (every other day) by the intravenous route using 1 O7 epimastigotes. The animals were bled 5-7 days after the last intravenous injection.
Ouchterlony immunodiffusion tests on agarose gel plates (Kabat & Mayer, 1961) were performed at 5 OC overnight, whereas quantitative immunoprecipitation was carried out
2.0
1.0 0
3 Q:
0 20 50 t o o Fract ion Number
FIGURE 2: Gel filtration on a 2 X 120 cm column of Bio-Gel P-10 of aqueous extract of epimastigotes from T. cruzi. Fractions were assayed for carbohydrate by using the phenol-sulfuric acid method. Fraction size, 5.0 mL.
Table I: Sugar Analyses (in Percent) of Fractions Obtained after P-10 Column Chromatography of the Aqueous Extract of T. cruzi
fraction
sugar I (PG25) I1 111 IV V
ribose 17.0 21.2 xylose 1.9 mannose 29.3 41.7 48.8 9.8 tr galactose 67.1 14.8 tra 10.6 tr glucose 1.6 43.5 51.0 tr tr inositol 75.8 78.8
Trace.
according to Raschke & Ballou (1971). For precipitin in- hibition studies, serum and inhibitor were incubated at 37 OC for 2 h, after which the antigen was added and the incubation continued at 4 "C for 48 h.
Radiolabeling and Radioimmunoprecipitation Assay of GP-25. GP-25 was radioiodinated by the iodogen method (Fraker & Speck, 1978) to a specific activity of 2 X los cpm/pg. The radiolabeled preparation was specifically im- munoprecipitated by chronic Chagas' disease human serum, and complexes were isolated by absorption on Staphylococcus A particles (New England Enzyme Center, Boston, MA) by described procedures (Kessler, 1975).
Results Aqueous Extraction of Cells. Epimastigote forms of T.
cruzi were obtained by growth on a BHI-based medium. Glycoconjugates were extracted by thawing frozen cells, followed by addition of cold water to form a cell suspension. The procedure was carried out at low temperature to minimize possible proteolytic degradation. After centrifugation, the solution was heated at 100 OC for 15 min, and the precipitate was centrifuged off. The supernatant was lyophilized and the residue fractionated by column chromatography on Bio-Gel P- 10. Elution with water provided five fractions (Figure 2) whose sugar composition is presented in Table I.
Chemical Structure of Fraction Z. On SDS-PAGE, fraction I (GP-25) gave a single band of apparent M, -25 000, de- tected by Coomassie Brilliant Blue and giving an intense color with PAS (Figure 3). Fraction I contained 60% protein, and the remaining carbohydrate component yielded, on hydrolysis, galactose, mannose, glucose, and xylose in a molar ratio of 35:13:1:1, The glycosidic linkage positions contained in fraction I were determined by methylation analysis. The per-0-methylated derivative of fraction I was degraded to partially 0-methylated alditol acetates, which were identified
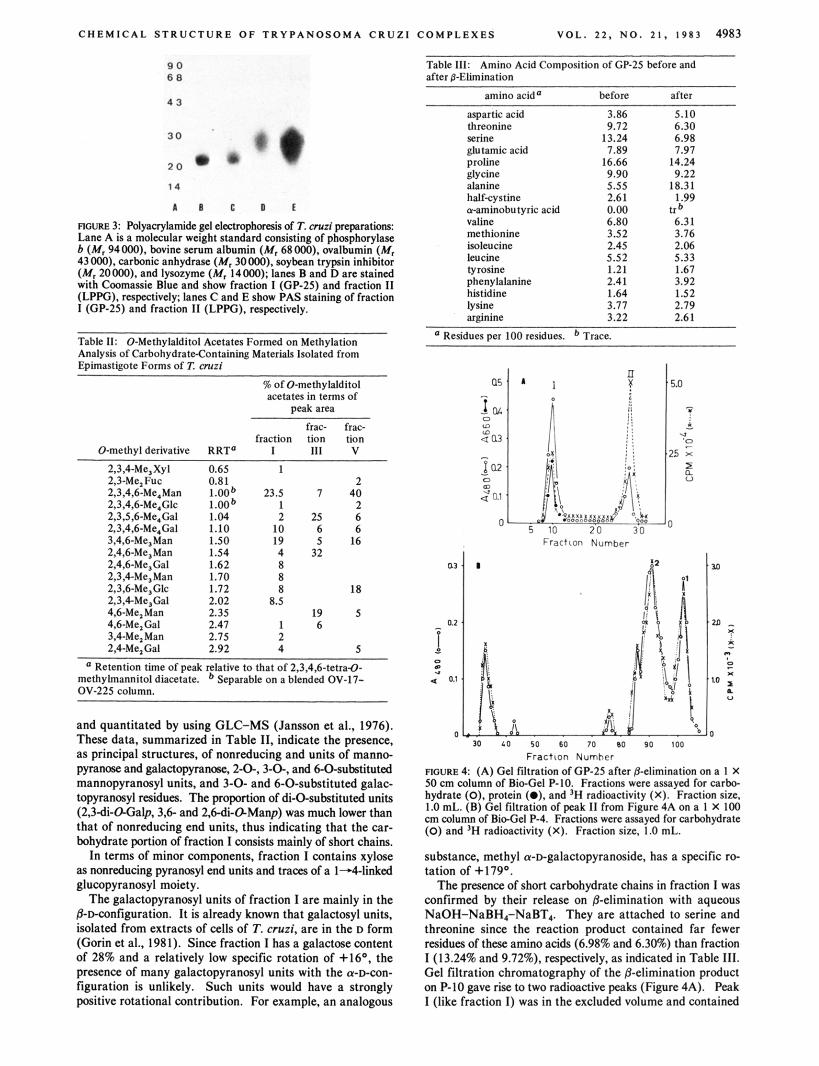
C H E M I C A L STRUCTURE O F TRYPANOSOMA CRUZl COMPLEXES V O L . 2 2 , N O . 2 1 . 1 9 8 3 4983
9 0 6 8
4 3
3 0
2 0 I.* 14
A I I c 0 t
ROURE 3 Polyacrylamide gel clcctmphorrris of T. cmzi p q ~ r a t i o m Lane A is a molecular weight standard consisting of phosphorylase b (M, 94000). bovine serum albumin (M, 68000). ovalbumin (M, 43000). carbonic anhydrase (M, 30000). soybean trypsin inhibitor (M, 20000). and lysozyme (M, 14000); lanes Band Dare stained with Cmmassie Blue and show fraction I (GP-25) and fraction I t (LPPG), respectively; lanes C and E show PAS staining of fraction I (GP-ZS) and fraction I I (LPPG), respectively.
Table 11: 0-Methylalditol Acetates Formed on Methylation Analysis of CarbohydrateContaining Materials Isolated from Epimastigote Forms of T. m r i
%of 0-methylalditol acetates in terms of
Deak area frac- frac-
fraction tion tion 0-methyl derivative RRTD I 111 v
2,3,4Me,Xyl 0.65 1 2,3-Me2Fuc 0.81 2 2,3,4,6-Me,Man L O O b 23.5 7 40 2.3.4.6-Me.Glc l . O O b I 2 ~ ~~ . . . . . 2,3,5,6-Me.Gal 1.04 2 25 6 2,3,4,6-Me.Gal 1.10 10 6 6 3,4,6-Me,Man 1.50 19 5 16 2,4,6-Me3Man 1.54 4 32 2.4.6-Me;Gal 1.62 8 2,3,4-Me,Man 1.70 a 2,3,6-Me,Glc 1.72 8 l a 2.3.4-Me.Gal 2.02 R.5 4;6:Me,hi;n 2.35 19 5 4,6-Me,Gal 2.47 1 6 3,4-Me2Man 2.75 2 2,4-Me,Gal 2.92 4 5
a Retention time of peak relative to that of 2,3.4,6-tetra-0- Separable on a blended OV-17- methylmannitol diacetate.
OV-225 column.
and quantitated by using GLC-MS (Jansson et al.. 1976). These data, summarized in Table 11, indicate the presence, as principal structures, of nonreducing and units of mannc- pyranose and galactopyranose, 2 -0 , 3-0, and 60substituted mannopyranosyl units, and 3-0- and 6-0-substituted galac- topyranosyl residues. The proportion of di-@substituted units (2,3di-@Galp. 3.6- and 2,ai-OManp) was much lower than that of nonreducing end units, thus indicating that the car- bohydrate portion of fraction I consists mainly of short chains.
In terms of minor components, fraction I contains xylose as nonreducing pyranosyl end units and traces of a 1 4 l i n k e d glucopyranosyl moiety.
The galactopyranosyl units of fraction I are mainly in the 8-o-configuration. It is already known that galactosyl units, isolated from extracts of cells of T. cruri, are in the D form (Gorin et al., 1981). Since fraction I has a galactose content of 28% and a relatively low specific rotation of + 1 6 O , the presence of many galactopyranosyl units with the a-o-con- figuration is unlikely. Such units would have a strongly positive rotational contribution. For example, an analogous
......
aspartic acid 3.86 5.10 threonine 9.72 6.30 serine 13.24 6.98 glutamic acid 7.89 7.97 proline 16.66 14.24 glycine 9.90 9.22 alanine 5.55 18.31 halfcystine 2.61 1.99
valine 6.80 6.31 methionine 3.52 3.16 isoleucine 2.45 2.06 leucine 5.52 5.33 tyrosine 1.21 1.67 phenylalanine 2.41 3.92 histidine 1.64 1.52 lysine 3.77 2.79 arginine 3.22 2.61
a-aminobutyric acid 0.00 trb
a Residues Der 100 residues. Trace.
a3. I
FractLon Number
M
L o - x 6
' 0 L
" x . "
Fraction Numher mouRe 4 (A) Gel filtration of GP-2S after &elimination on a I X SO cm column of Bic-Gel P-IO. Fractions were assayed for c a r b hydrate (0). protein (0). and 'H radioactivity (X). Fraction size, 1.0 mL. (E) Gel filtration of peak 11 from Figure 4A on a 1 X 100 em column of Bidiel P-4. Fractions were assayed for carbohydrate (0) and 'H radioactivity ( X ) . Fraction size, 1.0 mL.
substance, methyl a-D-galactopyranoside, has a specific ro- tation of +179".
The p r a n c e of short carbohydrate chains in fraction I was confirmed by their release on @-elimination with aqueous NaOH-NaBHrNaBT,. They are attached to serine and threonine since the reaction product contained far fewer residues of these amino acids (6.98% and 6.30%) than fraction I (13.24% and 9.72%). respectively, as indicated in Table 111. Gel filtration chromatography of the @-elimination product on P-IO gave rise to two radioactive peaks (Figure 4A). Peak I (like fraction I) was in the excluded volume and contained

4984 B I O C H E M I S T R Y
l a c 0
M E N D O N C A - P R E V I A T O E T A L .
N
11
muRe 5: lmmunoprccipitation profiles of 'z'I-GP-25. (Lane A) "'I-GP-25 alone; (lanes B and C) "'I-GP-25 mixed with normal human serum and serum for patients with chronic Chagas' disease, respectively; (lane D) molccular weight markers.
carbohydrate and protein. However, this does not necessarily signify that it represents undegraded material. Peak 11, which had a lower molecular weight, consisted of carbohydrate only, and when it was subjected to further column chromatography on P-4, two main fractions were obtained with elution char- acteristics corresponding to di- and tetrasaccharides (Figure 48).
As can be seen from Table 111, the predominant amino acid of fraction I is proline followed by successively smaller pro- portions of serine, glycine. threonine, glutamic acid, and valine.
Antigenic Properties of Fraction I . Fraction I was found to be specifically precipitated by chronic Chagas' disease serum (Scharfstein et al., 1982a,b). Analysis of the corresponding immune complexes on SDS-PAGE confmed the glycoprotein of M, -25000 as the isolated antigen (Figure 5).
Isolation and Chemical Characterization of Fraction II (UPG). Hot phenolwater extraction of insoluble cell debris, obtained after the aqueous extraction procedure described above, provided material which was retained in the aqueous phase. Extraction of this material with CHC13-MeOH-Hz0 provided soluble fraction I1 (LPPG), which contained carbo- hydrate (62%). protein (10%). hexosamine (1.5%). phosphate (7.8%). and lipid, but no sialic acid. The carbohydrate portion contained galactose and mannose in a molar ratio of 1:2 and traces of glucose and inositol, as determined by examination of the product obtained on successive hydrolysis, sodium bo- rohydride reduction. and acetylation. Fraction 11 gave on SDS-PAGE (Figure 3) a single band that was positive for protein (Coomassie Blue), carbohydrate (PAS), and lipid (Sudan Black; result not shown). In terms of t h s c analytical data, fraction I1 corresponds to the LPPG of Lcderkremer et al. (1976, 1977, 1978) despite the difference in extraction procedure.
Degradation of fraction 11 with hot aqueous NaOH-NaBH, gave rise to a carbohydrate, which was isolated as a single peak by column chromatography on P-4. Its elution characteristics were those of a moiety having approximately IO sugar units, when a I-blinked series of a-Dglucopyranosyl oligo- saccharides were used as standards. It was designated fraction 111 and was found to contain galactose and mannm in a ratio of 1:2.1. hexosamine (2%). phosphate (5%). and traces of glucose and inositol. Its "C NMR spectrum (Figure 6) contained a complex C-l region indicating the presence of many carbohydrate structures. This region extended from 6 96.8 to 106.8. One of the predominant signals at 6 106.5 is at unusually low field and corresponds to @-D-galactofuranosyl units (Gorin et al., 1981). Its sharpness, having a line width of 2 Hz, showed that the units are glycosidically linked to an adjacent sugar unit, and although phosphate is present, it is not, as previously suggested (Lederkremer et al., 1980). prsent in a phosphorodiester bridge between the units. If this was so, the signal would appear as a doublet due to "CXI-~'P
FIGUFS 6 Carbon-13 NMR spectrum of fraction 111 isolated from cpimastigotes of T. cruzi.
0 -
0, \o A
mme 7: Stained immunodillusion plate demonstrating reaction between rabbit hypcrimmune serum in the center well and the fol- lowing: A/C/E, fraction 11 (LPFG) intact; B/D/F. fraction I1 (LPPG) after sodium periodate oxidation.
coupling of 3-6 Hz (Lapper et al., 1972, 1973). Examples of line splitting in polysaccharide spectra have becn summa- rized (Gorin, 1981).
Methylation analysis of fraction 111, using the methodology described above. gave rise to a complex mixture of partly @methylated alditol acetates (Table 11). Their structures and proportions indicated that mast of the nonreducing end units were galactofuranose (25%) with smaller proportions of mannopyranose (7%) and galactopyranose (6%). Since the mannopyranosyl units were 2-0- (5%). 3-0- (32%). and 2.3-di-0- (19%) substituted, the nonreducing end units of @-c-galactofuranose are mainly linked 1-3 to adjacent D mannopyranosyl residues. These have the a-configuration since the specific rotation of fraction 111 is +31°. Since this value is much higher than that (-84") of a @-D-galactofuranan (Haworth et al., 1937). the rotational contribution of the o-mannopyranosyl units must be strongly positive and thus a. The presence of the @-o-Gal/-( I -3) -a -~Manp structure is confirmed by the presence in the "C NMR spectrum of the signal at 6 106.5. typical for this structure (Gorin et al.. 1981).
Another interesting feature of the "C NMR spectrum are two C-l signals a t high field (6 97.6 and 96.8). Such reso- nances have only been encountered rarely (Gorin, 1981). one example being from 2-acetamido-2deoxy-a-o-gluwp~nosyl units, which would also give a C-2 signal at 6 55.8 and a CH,-CO-NH signal a t 6 23.4. However, fraction 111 only contains 2% hexosamine, and such units would have to give rise to signals that are disproportionately large.
Antigenic Properties of Fraction I I . Fraction I1 (LPPG) gave a precipitation line in immunodiffusion p la t s with rabbit immune serum against epimastigote forms (Figure 7). After rapid oxidation of fraction 11 with aqueous sodium periodate followed by sodium borohydride reduction, a process that converts nonreducing end units of @-D-galactofuranose to a-L-arabinofuranw, without affecting other units appreciably, the modified complex became nonimmunoprecipitable by the same antiserum. The precipitin curve of LPPG antiserum against intact LPPG shows an equivalence point a t 20 pg of antigen with 100 pL of antiserum in a total volume of 500 pL. The precipitate contained about 40 pg of protein. For hapten

C H E M I C A L S T R U C T U R E O F T R Y P A N O S O M A C R U Z I C O M P L E X E S V O L . 2 2 , N O . 2 1 , 1 9 8 3 4985
100
75 8
f 50 9
- 25
C 0
r c
0 0.5 1.c
Hap ten concent ra t ton , pM
FIGURE 8: Hapten inhibition of the precipitin reaction between T. cruzi LPPG and antiserum raised against intact LPPG: (X) oligo- saccharide obtained after alkaline degradation of LPPG (fraction 111); (0) @-D-Galf-( 1+3)-Me-a-~-Manp; (0) methyl 8-D-galacto- furanoside; (A) methyl a-L-arabinofuranoside.
FIGURE 9: Carbon-13 NMR spectrum of fraction V isolated from epimastigotes of T. cruzi.
inhibition studies, we used oligosaccharide obtained after alkaline degradation of LPPG, @-~-Ga?f-( 1-+3)-Me-a-~- Manp, methyl @-D-galactofuranoside, and methyl a-L- arabinofuranoside. The results are shown in Figure 8.
Chemical Characteristics of Fractions ZV and V. The material that was insoluble in CHC13-MeOH-H20 in the above extraction process was further processed. Its yield was lower than that of the soluble fraction 11. It was fractionated by selective absorption on a column of Con A-Sepharose, and fraction IV was obtained by elution with aqueous 0.1 M methyl a-D-mannopyranoside. It contained units of galactose and mannose, in a molar ratio of 1.4:1, hexosamine (6%), and traces of glucose and fucose.
Fraction IV was degraded to fraction V with hot aqueous NaOH-NaBH4. It contained galactose and mannose in a ratio of 1:2 and traces of glucose and fucose. The hexosamine content was not determined. One interesting feature, which is not yet explained, is the reduction of the galactose content on alkaline degradation.
The I3C NMR spectrum of fraction V (Figure 9), although having similarities to that of fraction 111, has one important difference in that a signal is present at 6 103.5. This corre- sponds to nonreducing end units of a-D-mannopyranose linked 1-2 to a-D-mannopyranosyl units (Gorin, 1973), conforming with the methylation data (Table 11). This indicated nonre- ducing end units (41%) and 2-0-substituted units (16%) of mannopyranose. Also characterized were nonreducing end units of galactopyranose (6%) and P-D-galactofuranose (6%; see typical 13C N M R signal at 6 106.6) and a trace of 4-0- substituted fucopyranosyl (or 5-0-substituted furanosyl) residues (2%). Fucose has been recently identified as a com- ponent of a minor epimastigote glycoprotein (Snary et al., 1981). A trace of a short-chain 1-4-linked glucopyranosyl component is also apparently present in fraction V.
Discussion The knowledge of the structure and function of cell-surface
glycoconjugates of T cruzi may be of importance for the un- derstanding of the relationship between parasites and host cells. These glycoconjugates have been prepared, in most cases, from
detergent extracts of radioiodinated parasites by a process of immunoprecipitation (Snary & Hudson, 1979; Snary, 1980; Araujo & Remington, 1981; Nogueira et al., 1981). However, such preparations have not yet been subjected to analysis that would give an insight into their detailed chemical structures.
In the present study, we have investigated the chemical structures of the carbohydrate components of three fractions, designated I, 11, and IV, isolated from epimastigote forms of T. cruzi.
The fractionation procedure, summarized in Figure 1, was as follows. Cells were disrupted and extracted by successive freezing and thawing, and liberated soluble material was fractionated on a column of P-10. Five fractions were ob- tained, but only the first one that was eluted, fraction I, was investigated further. The cell debris, isolated after extraction, was then extracted with hot phenol-water. The aqueous phase was lyophilized and the residue extracted with CHC1,- MeOH-H20. Soluble material (fraction 11) appears to cor- respond to the LPPG of Lederkremer et al. (1976, 1977, 1978), who extracted cells directly with phenol-water without a prior aqueous treatment. The preparations resemble each other closely in terms of chemical composition. Our other prepa- ration, termed fraction IV, was obtained by fractionation of the CHC13-MeOH-H20-insoluble material on a column of Con A-Sepharose and consisted of the bound material.
Fraction I (GP-25) is a glycoprotein, which on SDS-PAGE electrophoresis gave rise to a single protein- and carbohy- drate-positive band with an apparent molecular weight of -25 000. The glycoprotein was radioiodinated and found to be specifically precipitated by serum obtained from chronic Chagas’ disease patients. The antigenic role of the glycoprotein was confirmed by its recovery from the insoluble immune complex and examination by gel electrophoresis.
@-Elimination of fraction I with cold aqueous NaOH-Na- BH4-NaBT4 indicated that the glycoprotein contained, at least to some extent, short carbohydrate chains. The reaction product was chromatographed on a column of P- 10, and two labeled fractions were obtained. One was excluded from the column and could consist of an undegraded component of fraction I or a true @-elimination product. The other fraction was refractionated on a column of P-4 and gave rise to two fractions whose elution characteristics corresponded to those of a di- and tetrasacharide. The oligosaccharides appear to be linked to serine and threonine, since these amino acids were decomposed during the @-elimination reaction.
Methylation analysis of fraction I supported the suggested existence of short sugar chains. This resulted in the formation of many 0-methylalditol acetate fragments (Table 11) which show the presence of predominant nonreducing end units of mannopyranose and galactopyranose and 2-0-substituted mannopyranosyl residues. The mannosyl units could exist as a-~-Manp-[(l-*2)-a-~-Manp],- with n = 0-2, as in the po- lysaccharide isolated after vigorous alkaline extraction (Gorin et al., 1981). The detection of galactopyranosyl units agrees with the observation of Alves et al. (1979), who found them in their mixed glycoprotein cell-surface components. The proportion of tetra-O-methylalditol acetates arising from nonreducing end units of fraction I is much greater than di- 0-methyl derivatives which would be products from branch points (present in equal amounts if it were a polysaccharide). Thus, the carbohydrates must be present as short chains linked to protein. The antigenic nature of GP-25 has been recently studied by Scharfstein et al. (1982a), who used affinity-purified human antibodies to demonstrate the presence of GP-25’s antigenic sites at the surface of T . cruzi and in all stages of

4986 B I O C H E M I S T R Y M E N D O N C A - P R E V I A T O E T A L .
of galactofuranose (6%) and galactopyranose (6%) but con- sisted principally of nonreducing end units of mannopyranose (41%) some of which are likely linked 1-2 to internal man- nopyranosyl units (16%). Such structures are consistent with the presence of C-1 signals in the 13C NMR spectrum at 6 103.5 and 102.3, respectively, values that have been previously reported for yeast mannan, which has similar structures (Gorin, 1973) with the a-configuration. This structure is consistent with the specific rotation of galactomannan, which is +29O.
The present results may have interesting implications for the study of Chagas’ disease. It is now possible to investigate the interaction of the host immune system with two cell-surface antigens which are present in all stages of development of T. cruzi. Some details of the chemical structures of the carbo- hydrate portions of these molecules have been determined. Remaining to be elucidated are the cellular function of the antigens and their roles in the infectious process.
References Alves, M. J. M., & Colli, W. (1975) FEBS Lett. 22, 188-190. Alves, M. J. M., Franco da Silveira, J., De Paiva, C. H. R.,
Tanaka, C. T., & Colli, W. (1979) FEBS Lett. 99, 81-85. Ames, B. N. (1966) Methods Enzymol. 8, 115-118. Araujo, F. G., & Remington, J. S . (1981) J . Zmmunol. 127,
Barreto-Bergter, E., Hogge, L., & Gorin, P. A. J. (1981)
Belcher, R. A,, Nutten, A. J., & Sambrook, C. M. (1954)
Brener, Z . (1973) Annu. Rev. Microbiol. 27, 347-382. Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., &
Fairbanks, G., Steck, T. L., & Wallach, D. F. H. (1971)
Fraker, P. J., & Speck, J. C. (1978) Biochem. Biophys. Res.
Gonsalves, J. M., & Yamaha, T. (1969) Am. J . Trop. Med.
Gorin, P. A. J. (1973) Can. J . Chem. 51, 2375-2383. Gorin, P. A. J. (1981) Adv. Carbohydr. Chem. Biochem. 38,
Gorin, P. A. J., & Spencer, J. F. T. (1959) Can. J . Chem. 37,
Gorin, P. A. J., Haskins, R. H., Travassos, L. R., & Mendonsa-Previato, L. (1 977) Carbohydr. Res. 55, 2 1-33.
Gorin, P. A. J., Barreto-Berger, E., & Cruz, F. S . (1981) Carbohydr. Res. 88, 177-1 88.
Gorin, P. A. J., Giblin, E. M., Slater, G. P., & Hogge, L. (1982) Carbohydr. Res. 106, 235-241.
Haworth, W. N. (1915) J . Chem. SOC., 8-16. Haworth, W. N., Raistrick, H., & Stacey, M. (1937) Biochem.
Jansson, P.-E., Kenne, L., Liedgren, H., Lindberg, B., & Lonngren, J. (1976) Chem. Commun. Univ. Stockholm.
Kabet, E. A., & Mayer, M. (1961) Experimental Immuno- chemistry, 2nd ed., pp 675-686, Charles C Thomas, Springfield, IL.
855-859.
Carbohydr. Res. 97, 147-150.
Analyst (London) 79, 201-208.
Smith, F. (1956) Anal. Chem. 28, 350-356.
Biochemistry 10, 2606-26 16.
Commun. 80, 849-857.
Hyg. 72, 39-44.
13-104.
499-502.
J . 31, 640-644.
Kessler, S. W. (1975) J . Zmmunol. 115, 1617-1624. Kuhn, R., Trischmann, H., & Low, I. (1955) Angew. Chem.
Laemmli, U. K. (1970) Nature (London) 227, 680-685. Lapper, R. D., Mantsch, H. H., & Smith, I. C. P. (1972) J .
Lapper, R. D., Mantsch, H. H., & Smith, I. C. P. (1973) J .
67, 32.
Am. Chem. SOC. 94, 6243-6244.
Am. Chem. SOC. 95, 2878-2880.
parasite development (Scharfstein et al., 1982b). A glycoprotein having a molecular weight of 90 000 which
has been isolated from all forms of T. cruzi by Snary (1980) was found to protect mice against a challenge infection with T. cruzi (Scott & Snary, 1979). On the other hand, Nogueira et al. (1981) identified a protein with a molecular weight of 90 000 presumably implicated in the antiphagocytic activity of blood-form trypomastigotes. Glycoproteins with molecular weights of 72 000 and 75 000 were respectively detected by Snary et al. (1 98 1) and Nogueira et al. (1 98 1) exclusively in epimastigote and metacyclic forms. On the basis of presently available data, we cannot yet establish the structural rela- tionship, if any, of these glycoproteins with GP-25.
Fraction 11, which corresponds to LPPG, was also found to be antigenic. This was observed in an immunodiffusion gel using rabbit immune serum against epimastigote forms. It was found that the nonreducing end units of @-D-galacto- furanose were involved in its antigenicity since on conversion of these units to a-L-arabinofuranose the antigenic activity disappeared. Also, when fraction I11 (see below) and /3-D- Gal$( 1-3)-Me-a-~-Manp were used as competitive inhib- itors, precipitation inhibitions of 85% and 75%, respectively, were observed. Methyl 8-D-galactofuranoside was relatively ineffective. This is the first time in the T. cruzi system that specific sugar units have been demonstrated as being respon- sible for antigenic activity.
LPPG was only detectable in epimastigote forms, as indi- cated by using surface labeling techniques (Zingales et al., 1982). It was speculated that LPPG is involved in specific phenomena of epimastigote cells, for example, their lability to the alternative pathway of complement (Nogueira et al., 1975) and their capacity to divide in the midgut of the insect vector. However, using the indirect immunofluorescence technique, Previato et al. (1982) showed the presence of this substance in all stages of the life cycle of T. cruzi.
In the present investigation, the carbohydrate portion of fraction I1 was prepared by reductive cleavage with hot aqueous NaOH-NaBH, and purified by column chromatog- raphy on P-4. It was named fraction 111, had a degree of polymerization (dp) in the range of 10, and had nonreducing end units of @-D-galactopyranose (25%) linked mainly 1-3 directly to a-D-mannopyranosyl units and not via a phospho- rodiester bridge as previously suggested (Lederkremer et al., 1980). High proportions of 3-0- (32%) and 2,3-di-O-sub- stituted (19%) a-D-mannopyranosyl units were also present. In general, the methylation and I3C NMR spectral data are in agreement. The C-1 signals of 8-D-galactofuranosyl units have already been mentioned under Results. In terms of the other main C-1 signals (Figure 6), which arose from a - ~ - mannopyranosyl units, the following assignments can be made. Those at 6 102.2 and 100.1 belong to C-1’s of 3-0-substituted and 2,3-di-Osubstituted Pmannopyranosyl units, respectively, since these major structures are indicated by the methylation data and the fact that the signal at high field of 6 100.1 has undergone a strong upfield ,&shift due to 2-0-substitution. This has been observed in the case of the C-1’s of 2,6-di-O- substituted a-D-mannopyranosyl residues of baker’s yeast mannan, which give rise to a signal at 6 100.0 (Gorin, 1973).
Fraction IV was degraded to fraction V by using hot aqueous NaOH-NaBH4. This carbohydrate had a chemical structure different from those of other fractions. It contained galactose and mannose in a molar ratio of 1 :2 and traces of glucose and fucose and was structurally different from fraction 111. According to the methylation analysis (Table 11), it contained only a small proportion of nonreducing end units

Biochemistry 1983, 22, 4987-4993 4987
Lederkremer, R. M., Alves, M. J. M., Fonseca, G. C., & Colli, W. (1976) Biochim. Biophys. Acta 441, 85-96.
Lederkremer, R. M., Tanaka, C. T., Alves, M. J. M., & Colli, W. (1977) Eur. J . Biochem. 74, 263-267.
Lederkremer, R. M., Casal, 0. L., Tanaka, C. T., & Colli, W. (1978) Biochem. Biophys. Res. Commun. 85,
Lederkremer, R. M., Casal, 0. L., Alves, M. J. M., & Colli,
Lee, Y.-C., & Scocca, J. R. (1972) J . Biol. Chem. 247,
Lloyd, K. 0. (1970) Arch. Biochem. Biophys. 137,460-468. Lowry, 0. H., Rosebrough, N. J., Farr, A. L., & Randall, R.
Nogueira, N., Bianco, C., & Cohn, Z. (1975) J . Exp. Med.
Nogueira, N., Chaplan, S., Tydings, J. D., Unkeless, J., &
Osborn, M. J. (1966) Methods Enzymol. 8, 161-164. Previato, J . O., Andrade, A. F. B., Gorin, P. A. J., &
Mendonga-Previato, L. (1982) Reuniiio Anual de Pesquisa Basica em Doenca de Chagas, IX, p 71, Imprensa Universitbria, Brasil.
Raschke, W. C., & Ballou, C. E. (1971) Biochemistry 10,
1268-1274.
W. (1980) FEBS Lett. 116, 25-29.
5753-5758.
J. (1951) J . Biol. Chem. 193, 265-275.
142, 224-229.
Cohn, Z. (1981) J . Exp. Med. 153, 629-639.
4 130-4 1 35.
Sawardeker, J. S., Sloneker, J. H., & Jeanes, A. (1965) Anal. Chem. 37, 1602-1604.
Scharfstein, J., Previato, J . O., Rodrigues, M. M., Barcinski, M. A., & Mendonga-Previato, L. (1982a) Abstracts of the International Congress of Parasitology, V, Toronto, Canada, p 37.
Scharfstein, J., Rodrigues, M. M., Alves, C. A., De Souza, W., & Mendonqa-Previato, L. (198213) Reuniiio Anual de Pesquisa Basica em Doenca de Chagas, IX, p 77, Imprensa Universitbria, Brasil.
Scott, M. T., & Snary, D. (1979) Nature (London) 282, 73-74.
Snary, D. (1980) Exp. Parasitol. 49, 68-77. Snary, D., & Hudson, L. (1979) FEBS Lett. 100, 16-170. Snary, D., & Ferguson, M. A. J. (1982) Abstracts of the
International Congress of Parasitology, V, Toronto, Canada, p 665.
Snary, D., Ferguson, M. A. J., Scott, M. T., & Allen, A. K. (1981) Mol. Biochem. Parasitol. 3, 343-356.
Tafuri, W. L. (1979) in Biology of the Kinetoplastida (Lumsden, W. H. R., & Evans, D. A., Eds.) Vol. 2, pp 547-618, Academic Press, New York.
Warren, L. (1959) J . Biol. Chem. 234, 1971-1975. Zingales, B., Martin, N. F., Lederkremer, R. M., & Colli, W.
(1982) FEBS Lett. 142, 238-242.
Functions of the 5’-Phosphoryl Group of Pyridoxal 5’-Phosphate in Phosphorylase: A Study Using Pyridoxal-Reconstituted Enzyme as a Model System+ Yen Chung Chang, Timothy McCalmont, and Donald J. Graves*
ABSTRACT: Pyridoxal-reconstituted phosphorylase was used as a model system to study the possible functions of the 5’- phosphoryl group of pyridoxal 5’-phosphate (PLP) in rabbit muscle glycogen phosphorylase. Kinetic study was conducted by using competitive inhibitors of phosphite, an activator, and a-D-glucopyranose 1 -phosphate (glucose- 1-P) to study the relationship between the PLP phosphate and the binding of glucose- 1-P to phosphorylase. Fluorine- 19 nuclear magnetic resonance ( 19F NMR) spectroscopy of fluorophosphate bound to pyridoxal phosphorylase showed that its ionization state did not change during enzymatic catalysis. Evaluation of the apparent kinetic parameters for the activation of pyridoxal phosphorylase with different analogues having varied pK,, values demonstrated a dependency of KM on pKa2 Molybdate, capable of binding as chelates in a trigonal-bipyramidal con- figuration, was tested for its inhibitory property with pyridoxal
\nous studies of a-glucan phosphorylases reconstituted with vitamin B6 analogues suggest that the 5’-phosphoryl group of PLP’ is likely to be involved in the catalytic process [see reviews by Helmreich & Klein (1980) and Graves & Wang (1972)l. Although phosphorylase reconstituted with pyridoxal
From the Department of Biochemistry and Biophysics, Iowa State University, Ames, Iowa 5001 1. Receiued February 10, 1983; reuised manuscript received June 9,1983. Journal Paper 5-10917 of the Iowa Agriculture and Home Economics Experiment Station, Ames, Iowa, Project 2120.
0006-296018 3 10422-4987$0 1.50/0
phosphorylase. On the basis of the results in this study, several conclusions may be drawn: (1) The bound phosphite in py- ridoxal phosphorylase and, possibly, the 5’-phosphoryl group of PLP in native phosphorylase do not effect the glucose- 1 -P binding. (2) One likely function of the 5’-phosphoryl group of PLP in native phosphorylase is acting as an anchoring point to hold the PLP molecule and/or various amino acid side chains in a proper orientation for effective catalysis. (3) The force between the PLP phosphate and its binding site in phosphorylase is mainly electrostatic; a change of ionization state during catalysis is unlikely. (4) Properties of the central atoms of different anions are important for their effects as either activators or inhibitors of pyridoxal phosphorylase. (5) Our results with molybdate are consistent but do not prove that a trigonal-bipyramidal structure of PLP is involved in the catalytic mechanism of phosphorylase.
is inactive (Illingworth et al., 1958), it was found some phosphate analogues could activate the enzyme (Parrish et al., 1977). These findings were explained by binding of phosphate and analogues at the site where the 5’-phosphoryl group of the coenzyme in native phosphorylase resides. Inorganic pyro- phosphate was found a potent inhibitor to pyridoxal-re-
I Abbreviations: glucose- 1 -P, cu-D-glUCOpyranOSe 1 -phosphate; PLP, pyridoxal 5’-phosphate; NMR, nuclear magnetic resonance; ppm, parts per million; EDTA, ethylenediaminetetraacetic acid.
0 1983 American Chemical Society