chemical and organoleptic quality changes of frozen stored
TRANSCRIPT

Louisiana State UniversityLSU Digital Commons
LSU Historical Dissertations and Theses Graduate School
1973
Chemical and Organoleptic Quality Changes ofFrozen Stored and Canned Channel Catfish.Joseph Richard BottaLouisiana State University and Agricultural & Mechanical College
Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses
This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion inLSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please [email protected].
Recommended CitationBotta, Joseph Richard, "Chemical and Organoleptic Quality Changes of Frozen Stored and Canned Channel Catfish." (1973). LSUHistorical Dissertations and Theses. 2520.https://digitalcommons.lsu.edu/gradschool_disstheses/2520

INFORMATION TO USERS
This material was produced from a microfilm copy of the original document. While the most advanced technological means to photograph and reproduce this document have been used, the quality is heavily dependent upon the quality of the original submitted.
The following explanation of techniques is provided to help you understand markings or patterns which may appear on this reproduction.
1.The sign or "target" for pages apparently lacking from the document photographed is "Missing Paga(s)". If it was possible to obtain the missing page(s) or section, they are spliced into the film along with adjacent pages. This may have necessitated cutting thru an image and duplicating adjacent pages to insure you complete continuity.
2. Whan an image on the film is obliterated with a large round black mark, it is an indication that the photographer suspected that the copy may have moved during exposure and thus cause a blurred image. You will find a good image of the page in the adjacent frame.
3. When a map, drawing or chart, etc., was part of the material being photographed the photographer followed a definite method in "sectioning" the material. It is customary to begin photoing at the upper left hand corner of a large sheet and to continue photoing from left to right in equal sections with a small overlap. If necessary, sectioning is continued again - beginning below the first row and continuing on until complete.
4. The majority of users indicate that the textual content is of greatest value, however, a somewhat higher quality reproduction could be made from "photographs" if essential to the understanding of the dissertation. Silver prints of "photographs" may be ordered at additional charge by writing the Order Department, giving the catalog number, title, author and specific pages you wish reproduced.
5. PLEASE NOTE: Some pages may have indistinct print. Filmed as received.
Xerox University Microfilms300 North Zaab RoadAnn Arbor, Michigan 46106

; I ' I
7 4 -1 8 ,3 1 6
BOTTA, Joseph R ich ard , 1946- CHEMICAL AND ORGANOLEPTIC QUALITY CHANGES OF FROZEN STORED AND CANNED CHANNEL CATFISH.
The L o u is ia n a S t a t e U n iv e r s ity and A g r ic u ltu r a l and M echanical C o lle g e , P h . D . , 1973 Food T echnology
University Microfilms, A XEROX Company, Ann Arbor, Michigan
THIS DISSERTATION HAS BEEN MICROFILMED EXACTLY AS RECEIVED.

CHEMICAL AND ORGANOLEPTIC QUALITY CHANGES OF FROZEN STORED AND CANNED CHANNEL CATFISH
A Dissertation
Submitted to the Graduate Faculty of the Louisiana State University and
Agricultural and Mechanical College in partial fulfillment of the requirements for the degree of
Doctor of Philosophy
in
The Department of Food Science
byJoseph Richard Botta
B.Sc.(Agr.), University of British Columbia/ 1969 M.Sc./ University of British Columbia/ 1971
December, 1973

ACKNOWLEDGEMENTS
The author is indebted to and expresses sincere thanks
to Dr. J . E. Rutledge for his advice, encouragement, and
help throughout the course of this study.
The author also wishes to sincerely thank Dr. W. F.
McKnight and Dr. J . E. Love for their help and suggestions.
The author sincerely thanks Dr. J . W. Avault for
providing the fish for the frozen storage study.
The author thanks all the people that served on the
taste panels.
Appreciation is also extended to Dr. R. M. Grodner,
Dr. S. P. Meyers, and Dr. A. F. Novak for serving as members
of his advisory committee.
The writer also gratefully acknowledges Louisiana
State University Sea Grant for providing financial
assistance.
i i

TABLE OF CONTENTS
P age
ACKNOWLEDGEMENTS...................................................................................... i i
TAB IE OF CONTENTS.................................................................................... i i i
LIST OF TABIES............................................................................................ vi
LIST OF FIGURES......................................................................................... v iii
ABSTRACT............................................................................................................ ix
INTRODUCTION................................................................................ 1
LITERATURE REVIEW.................................................................................... 3
Preservation of Catfish....... 3
Storage Studies.............................................................. 3
Thermal Processing...................................................... 5
Off-Flavor............................................................................ 6
Frozen Storage of Fish......................................................... 7
Lipid Hydrolysis............................................................ 7
Protein Extract ab ility ............................................ 13
Nucleotide Changes...................................................... 17
MATERIALS AND METHODS......................................................................... 24
Procurement of Samples.................................. 25
Canning and Frozen Storage Conditions 25
Canning Studies............................................................................ 26

P age
Content and Unsaturation of Lipid Present in Dark and Light Muscle of Catfish.............. 26
Determination of Iodine Number............................... 29
Organoleptic Analyses....................................................... 29
Yields of Canned product............................................... 31
Frozen Storage Studies.................................................................. 32
Determination of Free Fatty Acids....................... 33
Determination of Extractable Protein.............. 34
Determination of Hypoxanthine and Inosine Monophosphate................................................ 34
Organoleptic Analysis.................................................... 36
S tatistical Analysis.................................................................... 37
RESULTS AND DISCUSSION............................................................................ 39
Canning Studies.................................................................................. 39
Content and Unsaturation of Lipid Present In Dark and Light Muscle of Catfish 39
Organoleptic Analysis.................................................... 43
Yields of Canned Product............................................ 49
Frozen Storage Studies............................................................... 51
Free Fatty Acid Concentration............................... 51
Extractable Protein.......................................................... 53
Hypoxanthine C o n c e n t r a t i o n . . 55
Inosine Monophosphate Concentration 57
Organoleptic Analysis............................................. 59
Correlations............................................................................ 61
i v

SUMMARY.. . .
LITERATURE
VITA

LIST OF TABLES
T a b le P age
1 Content and unsaturation of lipidpresent in catfish ......................................................... 40
2 Content and unsaturation of lipidpresent in various fish ............................................ 42
3 Analyses of variance of odor, flavor,texture, and overall quality of canned catfish processed four different ways prior to thermal processing...................... 44
4 Treatment means from analyses of varianceof odor, flavor, texture, and overall quality of canned catfish processed four different ways prior to thermal processing.................................................................. 45
5 Taste panel treatment combination meansfor odor, flavor, texture, and overall quality of canned catfish processed four different ways and evaluated at two time intervals............................................................ 47
6 Free fatty acid content of channelcatfish stored at -10°C.................................... 52
7 Analysis of variance of free fatty acidsof channel catfish stored at -10 C ... 52
8 Extractable protein content of channelcatfish stored at -10 C.................................... 54
9 Analysis of variance of extractableprotein of channel catfish stored at-10 C.................................................................................... 54
10 Hypoxanthine content of channel catfishstored at -10°C ............... 56
v i

T a b le P age
11 Analysis of variance of hypaxanthigeof channel catfish stored at -10 C .... 56
12 Mean absorbance values of apparentinosine monophosphate of channelcatfish stored at -10°C.......................... 58
13 Analysis of variance of absorbance valuesof apparent inosine monophosphate of channel catfish stored at -10 C.. 58
14 Mean taste panel scores for odor,flavor, texture, and overall quality of channel catfish stored at -10 C .... 60
15 Analyses of variance of odor, flavor,texture, and overall auality of channel catfish stored at -10 C.................. 60
v i i

LIST OF FIGURES
F ig u r e P age
1 A diagramatic representation of across-section of a fish showing the areas of light and dark muscle............................. 27
2 Diagramatic presentation of processingprocedures prior to thermal processing 30
3 Yields of canned product processed fourdifferent ways prior to thermal processing......................................................................... 50
v i i i

ABSTRACT
Experiments were conducted to investigate the possi
b ility of canning catfish as a method of preservation .and
to determine if processing methods prior to canning could
eliminate off-flavor. Experiments were also conducted
to determine if certain chemical tests could be used as
an index of texture and flavor changes in frozen stored
catfish.
During the development of a canned catfish product,
characteristics of lipid present in dark and light muscle
were determined. Dark muscle contained 2 5.51% lipid with
an iodine number of 69.99. Light muscle contained 1.26%
lipid with an iodine number of 93.86.
A technically feasible canned catfish product was
developed. I t was found desirable to remove the bone as
well as add salt and lemon juice to the catfish flesh
prior to thermal processing. Removing the dark muscle did
not significantly affect odor, flavor, texture, and overall
quality. Steaming the fish prior to canning decreased
odor, removed off-flavor, decreased flavor, improved tex
ture, and decreased overall quality. Storing the canned

product for 6 months at room temperature tended to improve
texture without affecting odor, flavor, and overall quality.
The canned product of steamed, with dark muscle le f t intact,
gave the greatest yield (0.88 lb of whole catfish required
to f i l l one 307 x 113 can). The product of non-steamed
with dark muscle removed, gave the lowest yield (1.10 lb
of whole catfish required to f i l l one 307 x 113 can).
Odor, flavor, texture, and overall quality did not
significantly change during storage at -10°C for 15 weeks.
Free fatty acids increased, extractable protein decreased,
and hypoxanthine concentration did not significantly change
during storage at -10°C for 15 weeks. Neither free fatty
acids, extractable protein nor hypoxanthine correlated with
texture and flavor. Apparent inosine monophosphate appeared
to hold the most promise as an index of flavor.
x

INTRODUCTION
Recent developments in the catfish industry with
regard to farming practices have resulted in substantial
increases in production. The amount of catfish processed
has increased from 1.9 million pounds in 1969 to approxi
mately 10 million pounds in 1972 (Ammerman, 1973) . However
at the present time several problems s t i l l exist. Two
serious problems are the presence of off-flavor in pond
raised catfish during certain times of the year and the
reluctance of consumer to purchase frozen catfish (Lovell/
1971; Heaton et a l./ 1972).
Consequently, the present research was conducted to
investigate canning as a method of preserving catfish and
to determine if certain chemical tests could be used as
indices of flavor and texture changes of frozen stored
catfish. These tests could possibly be used to evaluate
the quality of frozen catfish available to the consumer.
This study consisted of: (1) determination of lipid
characteristics of catfish, (2) organoleptic analyses of
catfish processed four different ways prior to canning,
1

2
(3) determination of yields of canned products, (4) organ
oleptic analyses of fresh and frozen stored catfish, (5)
determination of extractable protein (accompanied by
determination of free fatty acids) as an index of texture
of frozen stored catfish, (6) determination of inosine
monophosphate and hypoxanthine changes as indices of
flavor of frozen stored fish.

LITERATURE REVIEW
Preservation of Catfish
Storage Studies: Boggess et al. (1971) studied the
storage stab ility of catfish at -18°C for up to 270 days.
They observed significant changes in pH, moisture, and
to tal lipid content. The rate of decrease in appearance,
aroma, color, texture, and flavor scores varied according
to skinning practices, freezing methods, and pond condi
tions (during the production of catfish ). In addition,
shear press firmness values of raw flesh did not appear
to be a reliable index of product texture and 2-thiobarbi-
turic acid (T3A) values were not a satisfactory index of
product stab ility .
Drake and Ammerman (197 3) precooked channel catfish
(fried at 375°F. for 10 min.) and then stored them at 0
or 10°F. At the end of 12 months of storage, a ll 2-thio-
barbituric acid (TBA) values were within acceptable levels.
Also triangle difference tests conducted at the end of the
storage period indicated that precooked (deep-fried) fish
packaged in polyethylene and not treated with polyphosphates
or butylated hydroxyanisole (BHA) could not be differentiated
3

4
from fresh cooked fish. However, Dyer et al. (1964)
reported that baking or possibly steaming is the best
cooking method for detection of changes in quality of
frozen stored fish whereas frying largely obscured the
characteristic frozen storage quality change. Application
of polyphosphates, BHA, or butylated hydraxytoluene (BHT)
did not improve the product, whereas, treatment with
propyl gallate did improve the product. Packaging of
the product in polyethylene bags heat sealed with 2 5
pound vacuum was beneficial.
Several studies concerning ice or refrigeration storage
of catfish have also been conducted. I t has been reported
that the pentasodium salt of diethylenetriaminepentaacetic / /
acid, 2,4,4-trichloro-2-hydroxydiphenyl ether, and methyl
2-chloroacetoacetic acid exhibit preservation action on
chilled channel catfish (Beuchat et a l., 1973). However,
none of these chemicals are presently approved by the FDA
as food additives.
Quality, especially appearance and flavor, change
more during the f ir s t 48 hr than between 48-96 hr on ice
(Heaton et a l., (1972). They also observed that the aroma
and flavor changes associated with freshness f ir s t became
apparent as the loss of natural freshness followed by a

5
strong fishy flavor and the development of "off-flavor".
There was also a great variation in aroma and texture
among fish. Dressed catfish could be held in crushed
ice for as long as 12 days and retain acceptable quality
whereas those held in plastic bags at 33°F were acceptable
for 8 days (Heaton et a l./ 1972). I t was also stated that
processors and consumers desire catfish marketed fresh
rather than frozen.
Determination of hypoxanthine concentration shows its
potential importance as an index of freshness of ice-
stored catfish (Beuchat/ 1973).
Thermal Processing: In itia l experiments by Botta and
Rutledge, (197 3) indicated that whole dressed catfish plus
salt produced an unacceptable product. The addition of
c itric acid improved the product slightly and removing the
dark muscle further improved the canned product. Steamed
f ille ts , with dark muscle removed, plus sa lt and c itr ic
acid yielded a "good" product except i t was too salty and
possessed a slight chemical flavor due to the c itr ic acid.
I t was found that the saltiness could be reduced by d is tr i
buting the salt more evenly throughout the product. The
chemical flavor could be eliminated by using lemon juice
instead of c itric acid.
After the present canning experiments were completed,

6
Drake et al. (197 3) reported on studies concerning canning
process calculations and evaluation of canned presteamed
channel catfish. For 304 x 406 cans, the processing time
in a s t i l l retort was 65 min at 250°F. Proximate analyses
indicated that canned channel catfish contained 7 5.13%
moisture, 21.44% crude protein, 3.33% crude fat, 1.18%
ash, and 0.24% crude fiber. Limited tests on fish cakes
and salads prepared from the canned product indicated
that they were very acceptable.
Off-Flavor: Certain types of Actinomycetes have been
shown to impart an earthy odor to water and to fish present
in the water. The organic compound causing the odor was
slightly soluble in water, volatile in steam, soluble in
ether, and partially soluble in alcohol (Thaysen, 1936).
The compound, called geosmin, has been shown to be a two-
ring tertiary alcohol with the hydroxyl group very s te ri-
cally hindered to have a boiling point of 270°C and an
imperical formula of C12H22° (Gerber 3,1(1 Lechevalier, 1965;
Medsker et a l., 1968). Organisms other than Actinomycetes
such as Symploca muscorum (a blue-green alga) may also
product geosmin (Safferman et al„ 1967).
At concentrations as low as 2 ppm, geosmin produced
an earthy-musty odor in trout flesh within 1 hr after the
fish were placed in the solution (Thaysen and Pentelow,

7
1936) . Geosmin has also been shown to produce typical
off-odors in catfish (Lovell/ 1973). Maligalig et al.
(197 3) observed that channel catfish developed the flavor
of their feeds.
In the presence of acid/ odoriferous geosmin is
converted to argosmin which possesses absolutely no odor
(Gerber and Lechevalier/ 1965).
Frozen Storage of Fish
Lipid Hydrolysis: In contrast to oxidative changes
in fish lipids, hydrolysis by itse lf has no obvious nutri
tional significance (Lovem, 1962) . In fish tissue, as
such, any effects of lipid hydrolysis on product quality
are likely to be due to secondary changes, e.g ., possible
increased susceptibility to oxidation, development of
off-flavors. Since the lipids, particularly the phospho
lipids, are intimately associated with protein i t is
possible that lipid hydrolysis may affect the stab ility
of protein toward denaturing influences, e.g ., during
freezing and frozen storage.
Brocklesby (1933) observed a gradual increase in free
fatty acids during frozen storage of Chinook and coho
salmon.
Dyer et al. (1958) reported almost no hydrolysis in

8
rosefish stored at either +10°F or -10°F but in Atlantic
halibut/ while there was no hydrolysis at -10°F, some
free fatty acid formation did occur at 0°F (up to about
1096 in 6 months) . When the halibut were stored at +10°F,
hydrolysis was more rapid (about 2096 free fatty acids in
6 months). With plaice there was an increase in free
fatty acids to about 3296 in 6 months at -10°F. Fresh
Atlantic cod had free fatty acid values of about 1596,
however/ when stored at -10°F the values increased to
about 5096 in 16 months and when stored at +10°F/ hydrolysis
was very rapid, the free fatty acids reaching values of
about 6096 in one month and eventually reaching values of
80 to 9096 in 16 months.
Wood and Haqg (1962) observed lipid hydrolysis and
free fatty acid formation in lingcod and Pacific gray cod
stored at +10°F. Fresh lingcod and Pacific gray cod
contained 3.4 and 5.296 free fatty acid (96 of total lipid),
respectively. After 15 weeks of storage at +10°F, these
values increased to 28 and 4296, respectively. Gray cod
resembled Atlantic cod in that there was a period of rapid
hydrolysis followed by a slower more uniform rate of hydro
lysis. This period of rapid hydrolysis was not observed
with lingcod.
Olley et al. (1962) showed that with Atlantic cod,

9
lemon sole, Atlantic halibut, dogfish, and eleven other
species there was a considerable increase in titra tab le
free fatty acids after 16 weeks at -14°C. The formation
of free fatty acids, expressed as a percentage of the
totc.1 lipid, was very similar in cod, lemon sole, and
halibut but was much less in dogfish. Phospholipase
activity appeared to be negligible in the Elasmobranchs
studied, but in a ll other species, phospholipase was at
least as important as lipase in producing free fatty acids.
In the Gadoids and related species almost a ll the free
fatty acids was from hydrolysis of phospholipids. I t
was also observed that the average lipid content (% of
wet muscle) of halibut varied from 0.7 5% in June to 1.05%
in December, lemon sole varied from 0.78% in July to 1.04%
in January while dogfish varied from 4.27% in July to 14.0%
in February.
The course of free fatty acid formation in rainbow
trout stored at -4°C has been found to be similar to that
in cod (Jonas and Bilinski, 1967).
Free fatty acids of fresh-water whitefish muscle,
stored at -10°C for 16 weeks, have been reported to
increase from 4.6% of the total lipid to 21.4% (Awad et a l.,
1969). Approximately 61% and 39% of the free fatty acid

10
increase was derived from phospholipids and triglycerides,
respectively.
Olley et al. (1969) showed that the extent of the
in itia l rapid, f ir s t order, hydrolysis reaction, appeared
to be limited by the amount of free water available in
the frozen state. Other results of Olley et al. (1969)
showed that with haddock there was a preferential hydrolysis
C16*0‘ C18*0 311(1 C20x5 311(1 that the rates
of hydrolysis of phosphatidylcholine and phosphatidyl-
ethanolamine were similar. Previously, Bligh (1961) had
shown that phosphatidylethanolamine and phosphatidylcholine
hydrolysis were mainly responsible for free fatty acid
increase in frozen stored Atlantic cod. Also, Bligh and
oScott (1966) showed that with cod stored at -12 C the rate
of breakdown of phosphatidylcholine was faster than that
of phosphatidylethanolamine. Bosund and Ganrot (1969),
while studying lipid hydrolysis in frozen Baltic herring,
also observed that phosphatidylcholine was hydrolyzed faster
than cephalin (phosphatidylethanolamine + phosphatidylserine).
They also observed considerably more phospholipid breakdown
and free fatty acid formation in red muscle than in white
muscle. They estimated that only 45% of the free fatty
acids in the red muscle and 7 5% of those in the white muscle
are formed by hydrolysis of phospholipids, the remainder

11
being formed by hydrolysis of triglycerides.
Until recently fish muscle was known to contain only
a lipase able to catalyze the hydrolysis of short chain
triglycerides (Bilinski, 1969)• However, the results
Bilinski and Lau (1969) indicated that rainbow trout muscle
also possesses lipolytic activity capable of hydrolyzing
depot fat which, in fish, is composed of triglycerides
containing predominantly fatty acids with 12-24 carbons.
Thus, hydrolysis of long-chain triglycerides can occur in
herring (Bosund and Ganrot, 1969) and in rainbow trout
(Bilinski and Lau, 1969). Whether other species of fish
possess this ability is not yet known.
The work of Yurkowski and Brockerhoff (1965) indicated
that the phospholipids of frozen stored cod are broken down
by two enzymes, phospholipase and lysophospholipase. Also
their studies on lysolecithinase showed that oleic acid
had a strong inhibitory effect.
Although many researchers have investigated the increase
in free fatty acid formation during frozen storage of various
species of fish, l i t t l e work has been done to determine if
this free fatty acid increase related to organoleptic changes
during frozen storage. Fraser and Dyer (1959) observed that
when Atlantic cod was stored at -10°F and +10°F for 12 months
the percent free fatty acids increased to approximately 50%

12
and 8096, respectively, whereas taste panel scores did not
change. They concluded that free fatty acids probably do
not affect taste.
Botta et al. (1973a and 1973b) observed that for both
Pacific halibut and chinook salmon neither to tal long-chain
free fatty acids nor any of the individual long-chain (C^ ~
C22*6 fa tty significantly (P ^ 0.05) correlated with
flavor changes during storage at -30°C for up to 80 weeks.
In contrast, Peters et al. (1968), while investigating
the effect of stage rigor, method of freezing and method
of thawing on the storage of frozen cod stored at -18°C,
observed that the correlation coefficient of free fatty
acid production with the average taste panel scores (average
of odor, flavor, texture, and overall quality scores) of
the frozen stored samples was -0.974 (p sf 0.01). However,
Olley et al. (1969) stated that if Peters had attempted
the correlation with samples stored at a different tempera
ture the significance might not have been so high. Olley's
results indicated that free fatty acid production does not
go to completion at all temperatures or, if i t does so,
i t is at two distinct rates, an in itia l rapid reaction
followed by a much slower one. Thus, she was of the opinion
that free fatty acid production and actomyosin insolubili
zation cannot both equate to a taste panel for texture at all

13
temperatures of frozen storage.
Protein Extractability: Some workers have observed
that, depending upon the species, protein extractability
(in 5% NaCl) tends to decrease during frozen storage. This
phenomenon has been observed in frozen plaice f il le ts (Dyer
and Morton, 1956); in rosefish (Dyer et a l., 1956) in
Atlantic cod (Dyer and Frasex; 1959; Olley and Lovern,
1960) in Atlantic halibut, lemon sole, and dogfish (Olley
et a l., 1962); in saithe, haddock, whiting, and mackerel
(Olley et a l., 1967); in Pacific cod and pacific halibut
(Tomlinson et a l., 1969); and in fresh water whitefish
(Awad et a l., 1969). No detectable change in the extrac
table protein nitrogen of lingcod, ocean perch, red snapper,
orange spotted rockfish, and rock sole, stored for 8
months at -30°C, was noticed by Tomlinson et al. (1969).
The decrease in protein extractability was also quite
slow for lemon sole and dogfish (Olley et a l., 1962).
Castell and Bishop (1973) showed that protein extrac
tab ility of Atlantic cod decreased slightly during the
spring spawning period (March, April, and May). Muscle
lipid content also decreased during the spring spawning
period. Ironside and Love (1956) observed that protein
extractability values of Atlantic cod decreased as the
size of the cod increased and concluded that i t was due

14
to increasing amount of myocomatta in the larger cod. Love
(1956) observed that protein extractability was effected
by rate of freezing. Extractability was high when fast-
freezing produced small intracellular ice crystals and
decreased steadily with slower freezing to a minimum value
at the point of maximum intracellular crystallization. It
has been shown that protein extractability of Atlantic cod
if also affected by the temperature of frozen storage
(Luijpen, 1957). Temperature decreased along with the
rate of protein extractability.
The relationship between the increase in free fatty
acid formation and the decrease in protein extractability
that occurs during frozen storage appears to vary among
species. Olley et al. (1962) while studying frozen stored
Atlantic cod, lemon sole, Atlantic halibut, and dogfish
observed that the rate of increase of free fatty acids
was twice as high for dogfish as for the other species,
but the rate of decrease in protein extractability was
less for dogfish than i t was for cod. Also, lemon sole,
which was similar to cod in free fatty acid production,
showed much less protein denaturation than cod. However,
Hanson and Olley (1965) hypothesized that neutral lipids
protect protein from feee fatty acid denaturation in situ
and not only at the homogenization stage of the soluble

15
protein determination. This hypothesis was supported by
the work of Olley et al. (1967) who showed that small
quantities of neutral lipid may have a protective effect
on fish muscle proteins.
Using model systems, King et al. (1962) showed that
the addition of a small amount of linoleic or linolenic
acid caused cod actcmyosin to precipitate from a solution
of isolated cod actcmysin. Also, using model systems,
Anderson et al (1965) concluded that the interaction of
protein with fatty acid, resulting in insolubilization of
the protein, is optimal at an ionic strength of 0.5 pit pH
7.2 and that 5 u is the ionic strength that should exist
in the cellular fluid of cod muscle as a result of freez-
omg to -1.5 C.
In his review, Connell (1968) stated that the protein
extractability of rosefish stored at +10°F and of skate
and nursehound at -7°C declined considerably without the
formation of free fatty acid. He refers to unpublished
data and concluded that free fatty acid production is
merely one change which coincided in some species with
the decline in protein extractability.
Dyer and Morton (1956) observed that the texture ra t
ings of plaice f ille ts stored at -12°C showed an increase in
toughness parallel to the decrease in protein extracts-

16
b ility . Also, Dyer et al. (1956) found that with rosefish
stored at -12°C, taste panel results correlated reasonably
well with protein extractability. However, with rosefish
stored at -23°C (Dyer et a l., 1956), and Atlantic cod
stored at -30°C (Love, 1956), marked increase in toughness
occurred before any appreciable decrease in protein extra
ctability had taken place. Luijpen (1957) observed a
consistent close relationship between the increase in
toughness after boiling and the decrease in the ratio of
soluble protein to total nitrogen in cod sampels that had
been stored at -10°C. However, no correlation was observed
with the samples that had been stored at -20°C and at -30°C.
Moorjani et al. (1960) reported that with fille ted morwong
packed in evacuated cans or sealed in cellophane bags and
stored at -18°C, the differences in protein extractability
were associated with texture differences after storage times
of 2 to 6 months. Cowie and Little (1966) studied toughness
and protein solubility of cod f ille ts during storage at -29°C
for 82 months. There was a steady decrease in protein
solubility from 72% to 45% but the development of toughness
during frozen storage was extremely variable. In fact,
cod which had been stored for 82 months were more tender
than some of the control f i l le ts which had been freshly
frozen but not stored. Also Cowie and L ittle (1967) found

17
a poor correlation between toughness and protein extracta-
o ob ility with cod stored at -7 C and at -14 C. They concluded
that protein extractability alone cannot accurately describe
toughness and suggested that pH must also be considered.
In contrast, Peters et al. (1968) found that taste panel
scores of cod stored at -18°C for 12 months correlated
(r = +0.92) significantly (P * 0.05) with extractable
protein. Connell (1969), while investigating changes in
the eating quality of frozen stored cod, observed that
protein extractability correlated much better with flavor
(r = -0.730) or firmness (r = -0.664) of cold stored fish
than either color ratio or cell fragility methods.
Gould (1971), in her review, concluded that protein
extractability test was the best test for estimating
texture and that i t should be accompanied by determination
oof free fatty acids at storage temperatures above -13 C.
At temperatures below -14°C, pH may be a more reliable index
of texture than extractable protein.
Nucleotide Changes; Jones (1965) stated that nucleo
tide degradation in most fish muscle follows the well-
defined pattern given below:
f f f e f OIT S 8ATP -=■* ADP -=■♦ AMP -----> IMP -=* Ino------- ■ ■» Hy-2* UAa a a a a, b ■* a, b

18
a = autolytic Hy = hypoxanthineATP = adenosine 5 '-triphosphate IMP = inosine 5 '-mono-ADP = adenosine 5 '-diphosphate phosphateAMP = adenosine 5*-monophosphate Ino = inosineb = bacterial s = slowf = fast UA = uric acid
This pattern has been observed in frozen carp (Saito and
Arai, 1957) in chilled Atlantic cod (Jones and Murray,
1962) and haddock, lemon sole and plaice (Kassemsam et al.,
1963).
In resting muscle, the predominant nucleotide is ATP
which rapidly undergoes autolytic dephosphorylation to
form ADP. ADP also rapidly undergoes autolytic dephos
phorylation to form AMP. AMP is deaminated by the tissue
enzyme AMP deaminase to yield IMP (inosinic acid).
If fish are chilled stared, degradation of ATP to IMP
may be substantially completed within 24 hr, particularly
if the fish have struggled during catching and death, thus
depleting ATP and creatine phosphate stores (Jones, 1965;
Kassemsarn et a l., 1963). Jones and Murray (1962) reported
that trawled cod had l i t t l e ATP and much IMP at death
compared with rested well-fed fish. In addition, Saito
and Arai (1957) showed that degradation of ATP to IMP took
place rapidly during the slow freezing of carp. Nowlan
and Dyer (1969) studied glycolytic and nucleotide changes
in the c ritica l freezing zone (-0.8 to -5.0°C) in pre-rigor

19
cod muscle (placed in the freezer within 40 min post mortem) .
They observed that the amount of change depended on the
thermal arrest time (time from -0.8 to -5.0°C). At a rapid
freezing rate (a thermal arrest time of 13 min), creatine
phosphate decreased 39%, in itia l glycogen decreased 15%
and ATP and ADP combined decreased 11%. With an intermediate
freezing rate (a thermal arrest time of 5.4 hr), creatine
phosphate was almost depleted, in itia l glycogen decreased
50% and ATP and ADP combined decreased 29%. After very
slow freezing (a thermal arrest time of 20 hr), creatine
phosphate was completely dephosphorylated and only traces
of glycogen, ATP, and ADP remained. Also during both slow
and intermediate freezing, deamination of adenosine com
pounds to inosine compounds occurred.
The dephosphorylation of IMP to form inosine is carried
out by a 5 '-nucleotidase (IMP phosphorylase) (Jones and
Murray, 1962). This dephosphorylation reaction varies
considerably with species, but in general, the formation
of inosine from IMP is much slower than the formation of
IMP (Jones, 1965).
The cleavage of inosine by muscle ribosidase (or by
nucleoside phosphorylase) to form hypoxanthine and ribose
(or ribose 1-phosphate) shows a much more variable rate
with different species than the preceding reactions. In

20
chilled-stored Pacific salmon/ inosine has been shown to
accumulate/ indicating a slow breakdown to hypox anth ine
(Creelman and Tomlinson/ 1960). I t has been observed that
with iced haddock and plaice, inosine levels rise to a
maximum in about 8 to 10 days and then rapidly decrease
(Kassemsarn et a l., 1963). Murray et al. (1966) found
l i t t l e inosine accumulation in Atlantic salmon indicating
a high ribosidase activity. L ittly inosine accumulation
and hence a rapid inosine to hypaxanthine conversion has
been observed with redfish (Fraser et a l., 1966), petrale
sole (Spinelli et a l., 1964), and lemon sole (Kassemsarn
et a l., 1963). I t should be noted that the breakdown of
inosine to hypaxanthine differs from the preceding reac
tions in that hypaxanthine is formed by the action of
tissue enzymes and later by bacterial enzymes, whereas,
a ll the preceding ractions were catalyzed by only tissue
enzymes.
Several workers have observed differences in nucleo
tide degradation between dark and light muscle. Saito
et al. (1959) found higher amounts of purine compounds
(especially nucleotides) in dorsal muscle of rainbow
trout as compared with dark lateral muscle. However, as
IMP was rapidly sp lit in the dark muscle but accumulated
in dorsal muscle, inosine and hypaxanthine rose more rapidly

21
in the dark muscle. In both swordfish (Dyer et a l., 1966)
and in mackerel (Fraser et a l., 1968), the pattern of nuc
leotide degradation was similar with both dark and light
muscle but the rate of degradation was faster in the dark
muscle.
Since the present research was concerned with hypo-
xanthine and IMP changes in frozen stored channel catfish
only research concerning purine nucleotide changes in
frozen fi3h of various species or in iced (chilled) cat
fish will be presented here, although, some research papers
concerning nucleotide degradation in various ice-stored
fish were studied for the preliminary review.
Beuchat (1973) observed that measurement of hypoxan-
thine in chilled channel catfish may be a useful, inex
pensive, objective method to assess the degree of freshness
prior to incipient spoilage. I t was also stated that no
chemical test holds as much promise as the hypoxanthine
test as an index of freshness for freshwater fish.
Saito and Arai (1957) found that during the course of
slow freezing of carp there was a rapid breakdown of ATP
and formation of IMP. The IMP formed, decreased slowly
with length of frozen storage and was converted to inosine
and hypoxanthine. In contrast, Spinelli et al. (1964)
observed that with ocean perch f il le ts stored at -20°C

22
there was practically no change in hypoxanthine content
during 4 months of storage. In addition, Tomlinson and
Geiger (1963) found no quantitative difference between
nucleotide (principally ADP) bound in freshly frozen
(pre-rigor) lingcod muscle and those from muscle severely
denautred by frozen storage. Hcwever, Jones (1963) observed
that with Atlantic cod frozen pre-rigor and stored at -14°C,
-20°C and -30°C for 60 weeks, accumulated inosine at -14°c
and slowly accumulated at -20°C. In contrast, hypoxanthine
increased very l i t t l e at -14°C and remained constant at
-20°C and -30°C. Haddock stored at -14°C showed changes
very similar to that of cod.
Connell and Howgate (1968) observed that for fish
(cod) iced for 2, 5 and 8 days and for all conditions of
forzen storage, a linear regression accounting for 35.6%
of the variance was the following:
GA = 4.10 -0.493** Hx + 0.0266** PE
Where OA = overall acceptability and PE = protein extracta
b ility . ** significant at the 1% level.
Dyer et al. (1966) observed no change in nucleotide
levels in swordfish during storage at -26°C for 4 to 5
months. I t was also observed that for swordfish IMP or
hypoxanthine did not appear to be a good chemical indices
for frozen storage quality deterioration. During frozen

23
storage of mackerel at -26°C for 6 weeks and during thaw
ing, l i t t l e change occurred in IMP, and inosine increased
slightly (Fraser et a l., 1968). They also observed good
correlation of taste with both IMP and hypoxanthine content.
I t appears then that the usefulness of IMP and hypoxanthine
as indices of frozen storage quality (flavor) is species
dependent.
I t was considered unwise to rely upon hypoxanthine
alone as an index of quality (flavor) of frozen fish
(Taste Panel Discussion 1969). Spinelli has suggested
that IMP and hypoxanthine should be determined simultane
ously on frozen fish (Taste Panel Discussion 1969).
Gould (1971), in her review, concluded that the very
rapidity of the nucleotide changes limits their use in
iced fish to the f ir s t week of storage. Consequently,
she recommends the use of nucleotide changes only in
fresh-frozen fish. Apart from nucleotide changes and
glycolytic changes there is no existing index for flavor
changes during frozen storage (apart for test for rancidity).
Furthermore, Boggess et al. (1971) observed that TBA values
did not correlate with storage stab ility of frozen stored
channel catfish.

MATERIALS AND METHODS
It has been reported that processors and consumers
desire catfish marketed fresh rather than frozen (Heaton
et a l./ 1972). Consequently, the results of experiments
reported in the literature and those of our in itia l studies
prompted the present research concerning the possibility
of canning as a method of preserving catfish and studies
ascertaining if certain chemical tests could be used as
indices of quality of frozen stored catfish and hence
possibly be used to evaluate the market quality of frozen
stored catfish.
The canning studies consisted of evaluation of the
characteristics of catfish lipids, organoleptic analyses
of the canned products, and determination of the yields
of the canned products.
The frozen storage studies consisted of evaluating
chemical indices of flavor (hypoxanthine and inosine
monophosphate) and texture (extractable protein and free
fatty acids) and their subsequent correlation with organ
oleptic analyses.
24

25
Procurement of Samples
Catfish used in the canning experiments were obtained
from The Catfish Farm, near Gonzales, Louisiana. Immed
iately following collection the catfish were brought to
the Food Science Department, Louisiana State University
where the fish were skinned, dressed, and stored in poly
ethylene bags at 4°C until further processed. All the
experiments were initiated with 48 hr of obtaining the
samples. Catfish samples used in the assessment of frozen
storage quality (organoleptic and chemical) were obtained
from experimental ponds of the Department of Wildlife and
Fisheries, Louisiana State University.
Canning and Frozen Storage Conditions
For all experiments related to canning, 307 x 113
C-enameled cans, obtained from Continental Can Co., New
Orleans, Louisiana were used. All samples were thermally
processed in a s t i l l retort for 50 min at 121°C (250°F),
Fish used in the frozen storage studies were filleted,
wrapped in oxygen-impermeable film (Saran Wrap), and stored
in polyethylene bags at -10°C for 0, 5, 10, and 15 weeks
prior to the various analyses. All experiments were
initiated within 30 hr of obtaining the samples.

26
Canning Studies
In itia l experiments by Botta and Rutledge (1973)
prompted the determination of total content and unsatura
tion of lipid present in both dark and light muscle (Fig. 1).
Removing the dark muscle greatly reduced the content of
o il present in the canning liquor. The level of content
and unsaturation of lipid in fish flesh affects their
storage quality (rancidity).
In itia l studies also prompted a detailed organoleptic
analysis of canned catfish processed four different ways
prior to thermal processing. Samples were analyzed shortly
after canning and after the canned product had been stored
for 6 months at room temperature. The yields of the
canned product processed by each of the four different
methods were also determined.
Determination of Total Content of Unsaturation of
Lipid Present in Dark and Light Muscle of Catfish: Using
a scapel, the dark muscle was removed from 22 f i l le ts (11
catfish) . One twenty gram sample of dark and of light
muscle was obtained from each f i l le t . Each sample was
wrapped in an oxygen-impermeable film and stored at -18°C
until analyzed. All samples were analyzed within a month
of storage.
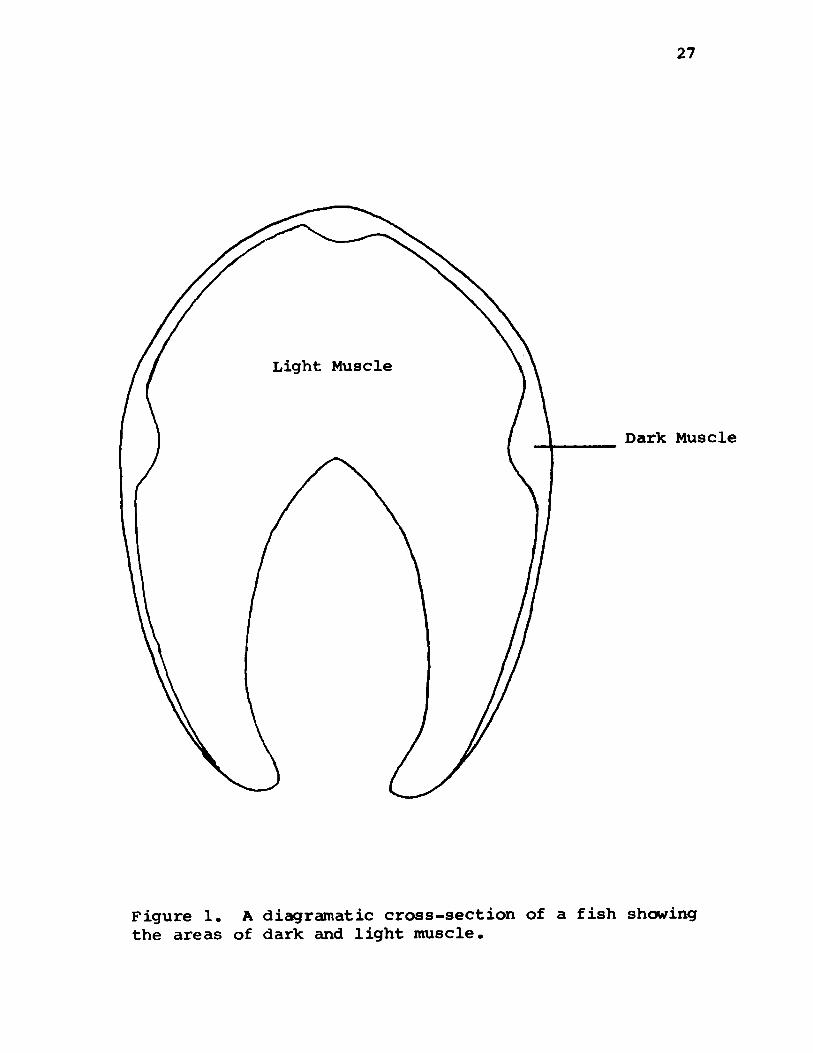
27
Light Muscle
Dark Muscle
Figure 1. A diagramatic cross-section of a fish shewing the areas of dark and light muscle.

28
Total lipid was extracted from 20 g of frozen flesh
by a procedure similar to that of Bligh and Dyer (1959).
Twenty grams of flesh, 40 ml methanol, and 20 ml chloroform
were homogenized for 2 min in a VirTis "45" hanogenizer,
to give a methanol:chloroform:water ratio of 2:1:0.8.
Twenty m illileters of chloroform were then added and
homogenization continued for 30 sec after which 20 ml of
water were added and the mixture homogenized for an addi
tional 30 sec. This gave a homogenate with a methanol:
chloroform:water ratio of 2:2:1.8. The homogenate was
then vacuum filtered through Whatman No. 1 f i l te r paper
on a No. 2 Buchner funnel into a 250 ml side-arm flask.
The tissue residue, f i l te r paper and 40 ml of chloroform
were homogenized for 1 min and vacuum filtered as before.
The hcmogenizer cup and the extracted tissue residue were
rinsed with 20 ml of a chloroform:methanol solution, 1:1
(v/v). The combined filtra tes were then transferred to a
250 ml separatory funnel and the side-arm flask was rinsed
with 20 ml of a 1:1 (v/v) chloroform:methanol solution.
After the chloroform and methanolic layers had
completely separated, the chloroform layer was removed.
The extracted lipid present in the chloroform layer was
dried to constant weight. The major portion of the

2 9
chloroform was evaporated by heating the solution on a
steam bath. The residual chloroform was evaporated by
heating the solution at 105°C until the sample obtained
constant weight (approximately 45 min at 105°C).
Determination of Iodine Number: The iodine number
(an index of unsaturation) of oil extracted from the dark
and light muscle of 8 fish was determined by reacting
0.25 to 0.50 g of extracted oil with Hanus iodine reagent
as described in A.O.A.C. (1970). The reacted mixture was
titra ted with 0.1N sodium thiosulphate solution using a
starch indicator.
Organoleptic Analysis of Canned Catfish Processed
Four Different Ways Prior to Thermal Processing: The
methods of processing are outlined in Fig. 2.
One lot of uneviscerated skinned catfish were divided
into two major groups. Catfish in one major group were
eviscerated, steamed for 20 min in free flowing steam,
the flesh separated from the bone, and then divided into
two sub-groups. The dark muscle was le ft intact in one
sub-group and removed in the other sub-group. Catfish in
the other major group were eviscerated, fille ted , and
divided into two sub groups. The dark muscle was le ft
intact in one sub-group and removed from the catfish in
the other sub-group. Thus, the four treatment combinations

30
UNEVISCERATED SKINNED CATFISH
EVISCERATED
FI LUSTEDI
NOT STEAMED
DARK MUSCLE DARK MUSCLE DARK MUSCLE DARK MUSCLEIE FT INTACT REMOVED LEFT INTACT REMOVED
Figure 2. Diagramatic presentation of processing procedures prior to thermal processing.
EVISCERATEDI
STEAMED (20 min)
FLESH SEPARATED FROM THE BONE

31
were: (1) steamed with dark muscle not removed# (2)
steamed with dark muscle removed, (3) non-steamed with
dark muscle not removed, and (4) non-steamed with dark
muscle removed.
It should be emphasized that not all of the dark
muscle was removed. Only the dark muscle in the dorsal
and lateral lines and within approximately a half inch
on each side of the lateral lines was removed. The
remainder was too d ifficu lt to remove.
In addition, 3.0 ml of lemon juice and one-half
teaspoon of sa lt were evenly distributed among the f il le ts
in the can prior to thermal processing.
The canned product of each treatment combination was
organoleptically evaluated, at the end of both storage
periods, by a 8 member experienced analytical taste panel
on 3 different days gave a total of 24 observations per
treatment combination per storage time. Odor, flavor, and
overall quality were evaluated using a six-point hedonic
scale (a score of 6 being excellent and a score of 1 being
very poor). Texture was evaluated using a five-point
hedonic scale (a score of 5 being very firm and a score
of 1 being very mushy)•
Determination of Yields of Canned product: Yields of
canned product were calculated for each treatment combination

32
shewn in Fig. 2. This entailed determining yields from
skinned eviscerated catfish for each treatment combina
tion and determining the weight of flesh required to f i l l
a 307 x 113 can. Yields of five fish for each treatment
combination were determined. Privious experiments had
been conducted on the yield of skinned eviscerated catfish
from unskinned and uneviscerated catfish. Using this
information, yields of each treatment combination from
unskinned and uneviscerated catfish were determined.
Frozen Storage Studies
Gould (1971), in her comprehensive review on testing
the freshness of frozen fish, concluded that the protein
extractability test accompanied by determination of free
fatty acids, if the fish are stored above -13°C, appears
to be the best test for estimating texture. She also stated
that nucleotide changes, i .e . , hypoxanthine increase and
particularly IMP disappearance, appears to be the best test
for estimating flavor. Consequently, i t was decided to
determine if extractable protein coupled with free fatty
acid determination can estimate texture in frozen stored
catfish and if changes in hypaxanthine or IMP concentration
could be correlated with flavor changes in frozen stored
fish.

33
Determination of Free Fatty Acids: Lipid was extracted
by the procedure that was used for the determination of
total content and unsaturation of lipid present in dark
and light muscle except that 40 g of sample and the corres
ponding volume of reagents were used. Anhydrous sodium
sulphate was added to remove water from the chloroform
layer. Total lipid was determined on an aliquot of the
water-free chloroform layer by the method that was described
previously for total content and unsaturation of lipid pre
sent in dark and light muscle of catfish. The water free
lipid fraction was then gravity filtered through Whatman
No. 1 f i l te r paper into a 2 50 ml round bottom flask. The
sodium sulphate was rinsed with 2 5 ml of reagent grade
chloroform. The lipid chloroform solution was then flash
evaporated to almost dryness, the water bath temperature
was 45°C.
The concentrated lipid was then dissolved in 50 ml of
petroleum ether (b.p. 65°C) and the solution transferred
to a column containing 15 g of activated s ilic ic acid to
remove the phospholipids. S ilicic acid (mallinckrodt 100
mesh) was activated by heating overnight at 120°C. The
column was washed with 150 ml of diethyl ether (Olley and
Lovern, 1960). The eluate was taken to almost dryness in
a rotary vacuum evaporator with a water bath temperature of
35°C. The concentrated lipid was then dissolved in 2 5 ml

34
of ethanol and the solution titra ted with 0.1N ethanolic
KOH using 1% phenolphthalein as an indicator (Olley and
Lovern, 1960). The free fatty acids were expressed as a
weight percentage of total lipid .
Determination of Extractable Protein: Protein extrac
table from frozen flesh in 5% sodium chloride solution was
determined by a method similar to that of Dyer et al.
(1950). 1/enty-five grams of frozen catfish flesh was blended
with 490 ml of 5% NaCl solution (adjusted to pH 7.0 with
NaHCO^) for 4 min in a Waring blender. Extraction was
carried out at 4°C and in order to prevent foaming, a
500 ml screw cap blender jar was filled completely with
5% NaCl solution and the cap tightly screwed on. In this
manner foaming was completely eliminated. The homogenate
was centrifuged for 30 min at 1,500 G. Extractable protein
was determined on an aliquot of the supernatant by the
direct biuret method of Snow (1950). The results were
expressed as a percentage of crude protein present in
catfish. Crude protein was determined on 6 catfish by
multiplying the percent to tal N (macro-kjeldahl method)
by 6.25.
Determination of Hypoxanthine and Inosine Monophosphate
Concentration: Hypoxanthine and inosine monophosphate (IMP)

35
were extracted by the method of Spinelli et al. (1964).
In this method/ 40 g of frozen catfish/ with dark muscle
along the lateral line removed/ were blended with 80 g of
0.7N chilled perchloric acid for 1 min in a VirTis "45"
homogenizer. The mixture was gravity filtered through
Whatman No. 1 f i l te r paper and 20 g of the f iltra te was
neutralized to a pH near 6.5 with ION KOH. Final adjust
ment to a pH of 6.5 was made with 1.8 N KOH. After cooling
ofor 15 min at 0 C, the solution was centrifuged at 7 55 G
for 15 min to remove the KC10. from solution and the4
supernatant liquor was brought to 2 5 ml with water. Five
m illilite rs of the neutralized f iltra te was passed over
Dcwex 1-X8 column in the formate form to remove nucleotide
and phosphorylated sugars.
The column prepared was similar to that of Jones (1960)•
Dowex 1-X8 (chloride), 200 to 400 mesh was suspended in
water and poured into a 0.8 cm diameter column until a
sedimented bed 4 cm deep was obtained. The bed was washed
with 10 times its volume with a solution containing 6N
formic acid and 1M sodium formate and washed an additional
10 times its volume with 8896 formic acid. I t was then
washed with water and weak alkali until the eluant was
neutral.

36
Five m illilite rs of neutralized filtra te was passed
through the column and washed with water until the absor
bance of the eluant had decreased to 0.01 at 260 run. The
column eluant was analyzed for hypaxanthine using a xanthine
oxidase method similar to that of Fujii (1968). In this
method 0.67 units of xanthine oxidase in 0.5 ml of water
was added to a solution containing 2.0 ml of water, 2.0
ml of 0.15M P04 buffer (pH 7.6), and 0.5 ml of eluant.
The solution was incubated at 35°C for 45 min and the
absorbance measured at 290 nm. Hypaxanthine was calculated
as mol per g of tissue.
The column was then eluted with three 5-ml portions
of 4n HC1 followed by two 5-ml portions of d istilled
water. Ten m illilite rs of this eluant was diluted to
50 ml in d istilled water and the absorbance measured at
2 56 nm. Both IMP and adenosine nucleotides are measured,
however, with iced fish the major nucleotide after a few
days has been shown to be IMP for several species of fish
(Jones and Murray, 1962; Kassemsarn et a l., 1963). No
interference at 256 nm would result from phosphorylated
sugars as they show no absorption in the ultraviolet. The
results were reported as absorbance values of apparent IMP.
Organoleptic Analysis of Fresh and Frozen Stored

37
Catfish: Fresh catfish and catfish that had been stored
at -10°C for 5, 10, and 15 weeks were organoleptically
evaluated by a nine member experienced analytical taste
panel using a six-point hedonic scale (a score of 6 being
excellent and a score of 1 being very poor). Each sample
was evaluated according to odor, flavor, texture, and over
all quality. The samples were prepared by wrapping them
in aluminum foil then baked for 15 min at 425°F. and
immediately served. I t was found necessary to spray the
aluminum foil with "PAM" (a pure vegetable spray-on
coating for cookware which prevents food from sticking,
distributed by Boyle-Midway, Inc., New York, N.Y. 10017)
to prevent the samples from sticking to the fo il.
S tatistical Analyses
Lipid content and unsaturation of catfish flesh was
s ta tis tica lly analyzed using a paired t- te s t as described
by Li, 1964.
Odor, flavor, texture, and overall quality of canned
catfish processed four different ways prior to thermal
processing were sta tis tica lly analyzed using a randomized
complete block design, blocking on judges, with a factorial
arrangement of treatments.

38
Changes in concentration of free fatty acids, extrac
table protein, hypaxanthine, and apparent IMP during frozen
storage were sta tis tica lly analyzed using a completely
randomized design. Odor, flavor, texture, and overall
quality of fresh and frozen catfish were sta tistica lly
analyzed using a randomized complete block design with a
factorial arrangement of treatments.

RESULTS AND DISCUSSION
Canning studies
Determination of Total Content and Unsaturation of
Lipid present in Dark and in Light Muscle of Catfish: The
results of earlier work (Botta and Rutledge, 1973) prompted
the determination of to tal content and unsaturation of
lipid present in the dark and also in the light muscle.
This was based on the observation that removing the dark
muscle greatly reduced the content of o il present in the
canning liquor.
The mean total lipid content of light muscle was only
1.26% whereas that of dark muscle was 25.52% (Table 1).
Thus, dark muscle contained approximately 20 times as much
lipid as light muscle. The high standard deviation of
lipid extracted from dark muscle was attributed to several
factors. Since fat is deposited in the dark muscle, its
lipid content is therefore a function of the diet of the
fish (Bilinski, 1969). Some of the fish were obtained
during the summer when they possessed off-flavor, and thus,
were on a reduced feeding program. Other fish were obtained
in the fa ll when off-flavor was no longer present and the
fish were on a regular feeding program. Secondly, during
39

4 0
Table 1. Content and Unsaturation of Lipid Present in Catfish.
SampleTotal lipid^
(g/100 g muscle)5
Iodine Number
2. 3Light muscle 1.26 (0.56) 93.86
Dark muscle 25.51 (7.36) 69.99
Total Flesh 4.94 (.147)4
■'"Total lipid extracted from muscle by Chloroform-methanol method (Bligh and Dyer, 1959)2Mean with standard deviation in brackets
3For light and dark muscle n = 22 (11 fish)
4For total flesh n = 11
^For Iodine No. n = 8 (8 fish)* *
Significant at the .01 level

4 1
the sampling i t was extremely d ifficu lt to separate all
of the light muscle from the dark muscle; this may have
contributed to the variability . In addition, part of the
variability may be genetic as Worthington et al. (1971)
also observed a large variability in catfish lipid (fatty
acid) content and composition.
Table 1 also shows the iodine number of lipid extracted
from dark and light muscle of catfish. The mean iodine
number of lipid extracted from light muscle was 93.86
whereas that of lipid extracted from dark muscle was
69.99. A paired t- te s t indicated that, the iodine number
of o il extracted from dark muscle was highly significantly
(P ^ 0.01) lower than the iodine number of oil from light
muscle. Thus although dark muscle contained approximately
20 times as much lipid as light muscle the lipin in dark
muscle was significantly (P ^ 0.01) more saturated than
lipid in light muscle.
The content and unsaturation of lipid present in
various other fish is shown in Table 2. I t indicates
that catfish is a medium fatty fish as i t contains more
lipid than certain fish such as lingcod and rainbow trout,
but less than mackerel and menhaden.
The iodine number of lipid from catfish is far lower
than that of lipid from any of the other fish presented

42
T a b le 2
Content and Unsaturation of Lipid Present in Various Fish
Species
Total lipid (% of wet muscle) Iodine Number
DarkMuscle
LightMuscle
TotalFlesh
DarkMuscle
LightMuscle
TotalFlesh
Catfish ^ 25.52 1.26 4.94 69.99 93.86Coalf islj 2.5 0.5 ---------------------- ---------------------- ---------------------- ----------------------------
Herring ^ 28.2 13.0 ------------------ ---------------------- ---------------------- ----------------------------
Mackerel ^ 29.7 13.1 ---------------------- ---------------------- - — - ----------------------------
Atlanti^ Salmon 15.1 2.0 ---------------------- ---------------------- ---------------------- ----------------------------
Lingcod 2 — - — — 0.85 ---------------------- — ---------------- — —-
Pacific Gray Cod ---------------------- - -— 0.58 ---------------------- ---------------------- ---------------- —
Greater^Silver Smelt 4 • m w m m * 3.5 _____ 314.9
Atlantic Herring ---------------------- ---------------------- ---------------------- ---------------------- ---------------------- 128Pacific Herring 4 ---------------------- ---------------------- ---------------------- ---------------------- ---------------------- 130Pacific Pilchgrd ---------------------- ---------------------- ---------------------- ---------------------- ---------------------- 192Cod liver oi^ ---------------------- ---------------------- ---------------------- ---------------------- ---------------------- 155Atlant ic^Cod ---------------------- ---------------------- 0.7 ---------------------- ---------------------- 184Mackerel ---------------------- ---------------------- 12.9 ---------------------- ---------------------- 140Menhaden (entire
fish) 15.5 150Striped Mullet 4
(entire figh) 2.8 160Lake Herring 4 ---------------------- ---------------------- 2.5 ---------------------- ---------------------- 163Rainbow Trout 4 ---------------------- r.. — — — 2.5 ---------------------- 184Lake Whitefish — — — — — — — — 2 . 2 — — — — — — — — 158
Bilinski, E. (1969). in Fish in Research. Academic Press, Inc., New York, pp. 135-152.
Wood, J . D. and S. A. Hagg. (1962). J . Fish Res. Bd.( . Canada, 19(1): 169-171.
Mackie, P. R. and R. Hardy. (1969). J . Fd. Technol.(4) 4:241-254.
Ackman, R. G. (1966). J . Am. Oil Chem. Sco. 43(6): 385-389.

43
in Table 2. For example/ the iodine number of lipid from
light muscle of catfish is 93.86 whereas the iodine number
of greater silver smelt is 114. In addition/ while lipid
present in dark and light muscle of catfish have an iodine
number of 69.99 and 93.86, respectively, the iodine number
of oleic acid, an 18 carbon fatty acid with one double
bond, is 90.07 (Triebold and Aurand, 1963)•
The low iodine number of lipid present in dark muscle
may be important from a storage point of view, (rancidity).
Organoleptic Analysis of Canned Catfish Processed
Four Different Ways Prior to Thermal processing: The
results of earlier work (Botta and Rutledge, 1973) also
prompted a detailed organoleptic analysis of canned cat
fish processed four different ways prior to thermal
processing and to determine yields of the canned product
processed by each of the four different methods.
S tatistical analysis of the taste panel data revealed
that removing dark muscle did not significantly (P ^ 0.05)
reduce odor, flavor, texture, and overall quality (Table 3
and 4). Steaming the f ille ts prior to thermal processing
significantly (P s? 0.01) reduced odor and flavor, signifi
cantly (P < 0.01) improved texture, and significantly
(p ^ 0.05) reduced overall quality (Table 3 and 4). It
is thought that steaming reduced odor and flavor by

44
T a b le 3
Analyses of Variance of Odor, Flavor, Texture and overall Quality of Canned Catfish Processed Four
Different Ways prior to Thermal Processing
Source d . f .Odor. m.s.
Flavoj m • s •
Textuje m.s.
Overall Qualify m.s •
Time (T) 1 0.521 1.021 1.3332 0.005Rep (time) (R) 4 2.021 0.583 0.208 0.458Judge (J) 7 1-976** 3.902** 2.905** 2.571*Steam (S) 1 65.333 46.021 56.333 27.755Muscle (M) 1 1.688 1.021 3.000 0.130S X M 1 6.021 0.021 0.750 0.630J X S 7 2.452 6.164 1.833 5.100J X M 7 0.902 0.640 0.429 0.785J X S X M 7 2.283 0.426 0.250 0.523J X T 7 5.140 3.949 1.738 3.446T X S 1 1.021 0.520 0.333 0.047T X M 1 0 . 0 0 0 2.521 0.750 0.005T X S X M 1 0.333 0.188 0.333 0.005J X T X S 7 2.830 1.521 1.524 2.202J X T X M 7 0.381 0.283 0.250 0.231J X T X S X M 7 0.476 0.402 0.714 0.517J X R (T) 28 1.511 0.881 0.554 0.541R X S (T) 4 0.411 1.865 1.583 1.760R X M(T) 4 0.734 1.427 1.188 0.865R X S X M(T) 4 2.161 0.166 0.167 0.208J X R X S (T) 28 1.108 0.686 0.786 0.403J X R X M(T) 28 0.514 0.510 0.402 0.555J X R X S X M(T) 28 0.846 0.655 0.643 0.625Total 191 1.658 1.387 1.098 1.094
Time was tested by R(T); steam by R X S(T); muscle by R X M(T); S XMb y R XS X M(T); T X S by R X S(T); T X M by R X M(T); and T X S X Mb y R X S X M(T).
*Significant at the 5% level
★★Significant at the 1% level
2Approaching significance (P ^ 0.065)

4 5
Table 4
Treatment means From Analyses of Variance of Odor, Flavor# Texture, and Overall Quality of Canned Catfish Processed
Four Different Ways Prior to Thermal Processing
Odor FIavor Texture OverallQuality
Dark muscle not removed 3.93 3.65 3.33 3.71ns • ns. ns. ns .
Daxk muscle removed 3.73 3.50 3.58 3.67
Steamed 3.25**
3.08 4.00 ** **
3.31*
Non-steamed 4.16 4.06 2.92 4.07
Freshly canned 3.78 3.64 3.38 3.70ns • ns. ns. ns.
Stored for 6 months 3.88 3.50 3.54 3.70
For each treatment n = 96
ns. = no significant differences at the 5% level
* significant at the 5% level
** Significant at the 1% level

4 6
removing volatile compounds and by forming azeotropes
possibly removing some non-volatiles that are important
in promoting odor and flavor. I t is also believed that
steaming improved texture by removing a large portion of
the water normally present in raw fish flesh.
Storing the canned product at room temperature for
6 months did not significantly affect odor, flavor, tex
ture, and overall quality (Tables 3 and 4). However,
although storage time did not significantly (P ^ 0.05)
improve texture, i t approached significance (P 0.065).
Thus, i t is tempting to speculate that a longer storage
time would significantly improve texture without signifi
cantly reducing odor, flavor, or overall quality.
Development of oxidative rancidity during storage of
the canned product does not appear to be an important
factor as storage time did not significantly reduce flavor
(Table 3 and 4). This agrees with Drake and Ammerman (1973)
who observed that when precooked catfish were frozen and
stored at 0°F the TEA values were within acceptable levels
at the end of the 12 months of storage.
Although steaming significantly reduced odor, flavor
and overall quality, i t should be noted that the product
was s t i l l quite acceptable (Tables 4 and 5). In addition,
there are some advantages to steaming catfish prior to

T a b le 5
Taste Panel Treatment Combination Means for Odor, Flavor, Texture, and Overall Quality of Canned Catfish Processed Four Different Ways and Evaluated at
Two Different Time Intervals 1
0 Storage Time 6 Month Storage Time
Steamed Non-Stearned Steamed Non-SteamedDark Dark Dark Dark Dark Dark Dark Dark
MuscleOn
MuscleOff
MuscleOn
MuscleOff
MuscleOn
MuscleOff
MuscleOn
Muscl<Off
Odor 3.50 3.04 4.25 4.33 3.54 2.91 4.42 4.67
Flavor 3.33 2.87 4.33 4.04 3.00 3.13 3.92 3.95
Texture 3.79 3.96 2.58 3.17 4.08 4.17 2.88 3.04
OverallQuality 3.42 3.25 4.04 4.08 3.38 3.21 4.04 3.13
Each mean is the average of 24 observations

4 8
thermal processing. Steaming facilita tes easy removal
of the bone and steaming may have important applications
regarding catfish processing off-flavor. Experiments with
off-flavor catfish indicated that the treatment combina
tion of steamed, with dark muscle removed, essentially
eliminated the earthy-musty odor and flavor.
Geosmin, C12H22°/ a d Jnetlly1 substituted, saturated,
two-ring teriary alcohol is suspected of possibly causing
off-odors and off-flavors in catfish (Lovell, 1971; 1973;
Medsker et a l., 1968). I t is possible that the compound
forms an azeotrope and thus can be removed by steaming.
Also, Gerber and Lechevalier (1965) reported that geosmin
is unstable to acid. In the presence of acid, odoriferous
geosmin is readily transformed to argosmin which possesses
absolutely no odor (Gerber and Lechevalier, 1965). Thus,
the lemon juice added to the product prior to thermal
processing, may have partially transformed geosmin into
argosmin which possesses no odor.
It appears that for catfish that do not possess
off-flavor the best treatment combination from a flavor
standpoint would be non-steamed with dark muscle on.
From a processing standpoint i t would be steamed with
dark muscle on. For catfish with off-flavor the best
treatment combination would be steamed with dark muscle on.

4 9
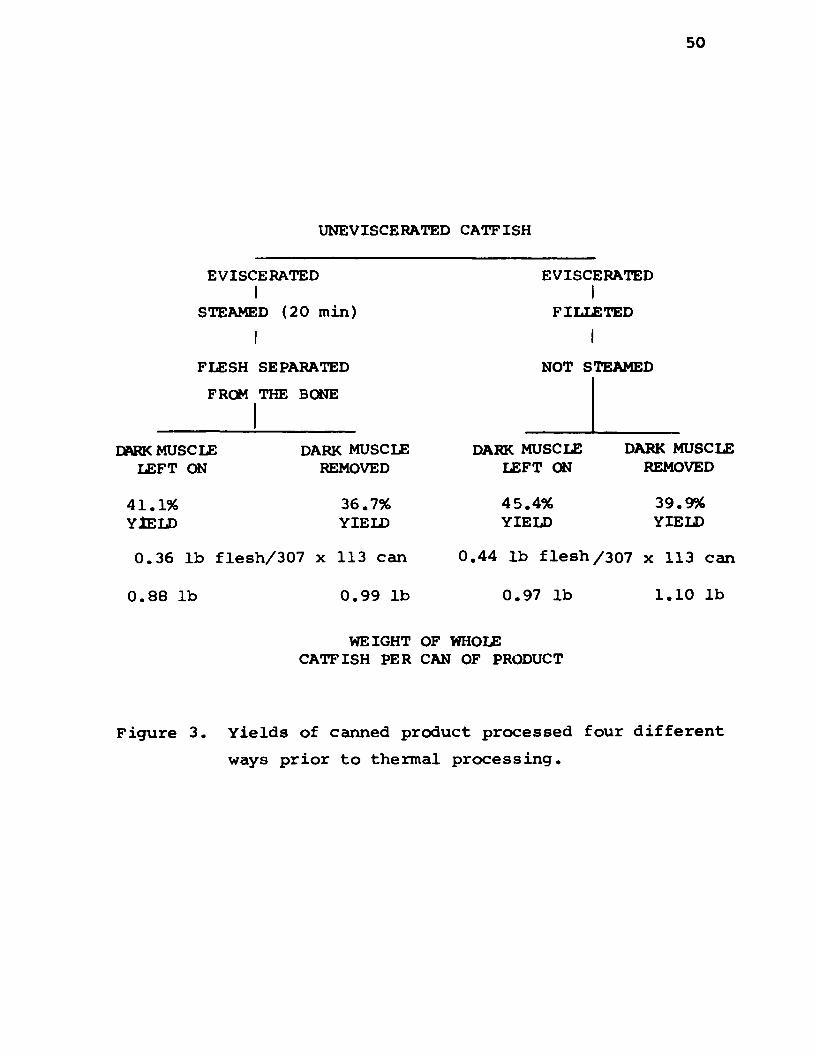
Yields of Canned Product Processed Four Different Wavs
Prior to Thermal Processing: Figure 3 shows the yields
of canned product for each treatment combination. The
percent yield of canned product varied from 45.4% to
36.7%. In order of decreasing yield the treatment
combinations were: unsteamed with dark muscle le ft on,
steamed with dark muscle le ft on, unsteamed with dark
muscle removed, and steamed with dark muscle removed.
However, one 307 x 113 size can of steamed product required
0.36 lb of flesh as compared to 0.44 lb of flesh for the
unsteamed product. This is because the steamed product
was less dense due to water loss during steaming. Figure 3
also shows the weight of whole catfish required per cen of
product for each treatment combination. The amount of
whole catfish required to f i l l a can of product varied
from 1.10 lb for non-steamed with dark muscle removed to
0.88 lb for steamed with dark muscle le ft on.
Greenfield (1970) stated that if a catfish producer
sold catfish for 41C/lb then the cost to the consumer
would be $1.43/lb for retailed frozen catfish. Presently
a local supermarket sells 307 x 113 cans of tuna for 39C
to 65C each, depending on the quality, and the same size
cans of salmon sell for 69C to 89C each, depending on the
species. Bearing in mind that 0.88 to 1.10 lb of whole
50
UNEVISCERATED CATFISH
EVISCERATED
STEAMED (20 min)
EVISCERATED
FI LIE TED
FLESH SEPARATED
FROM THE BONE
NOT STEAMED
DARK MUSCLE LEFT ON
41.1%YIELD
DARK MUSCLE REMOVED
36.7%YIELD
0.36 lb flesh/307 x 113 can
0.88 lb 0.99 lb
DARK MUSCLE LEFT ON
45.4%YIELD
DARK MUSCLE REMOVED
39.9%YIEIE
0.44 lb flesh/307 x 113 can
0.97 lb 1.10 lb
WEIGHT OF WHOLE CATFISH PER CAN OF PRODUCT
Figure 3. Yields of canned product processed four different
ways prior to thermal processing.

51
catfish are required for one 307 x 113 can, i t then appears
that canned catfish probably could not compete economi
cally with canned tuna or canned salmon. However, a
detailed economic study of the costs involved would have
to be made before any definite conclusions can be drawn.
In addition, even if the price of canned catfish were
higher than that of tuna or salmon, i t does not necessarily
mean that a limited volume of the product could not be sold,
particularily in the river basins of the Gulf Coast and the
southern half of the Mississippi River drainage system.
In these regions catfish are regarded as high-value premium
food (Greenfield, 1970). Also, through improved culturing
methods the price of whole catfish may in the future be
relatively lower in terms of constant dollars than they
are at the present time. I t should be emphasized that
consumer acceptability and economic feasability requires
further study.
Frozen Storage Studies
Free Fatty Acids: Free fatty acids (FFA) signifi
cantly (P sr 0.01) increased in concentration from 0.36%
for fresh catfish to 5.21% for catfish stored at -10°C
for 15 weeks (Tables 6 and 7). The coefficient of
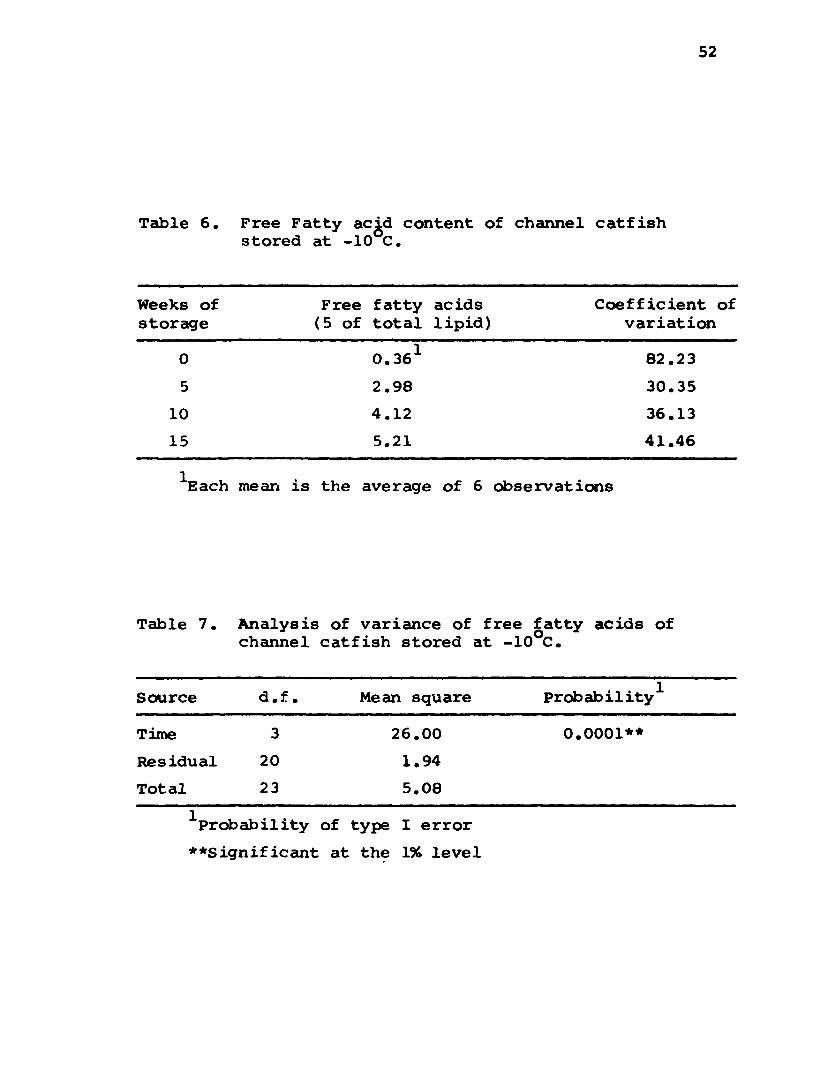
52
Table 6. Free Fatty ac .d content of channel catfish stored at -10 C.
Weeks of storage
Free fatty (5 of total
acids 1ipid)
Coefficient of variation
0 0.361 82.23
5 2.98 30.35
10 4.12 36.13
15 5.21 41.46
Pach mean is the average of 6 observations
Table 7. Analysischannel
of variance of catfish stored
free fatty acids of at -10°C.
Source d .f . Mean square Probability1
Time 3 26.00 0.0001**
Residual 20 1.94
Total 23 5.08
Probability of type I error
**Significant at the 196 level

53
variation indicated that the variability among samples at
a time period (particularly fresh catfish) was quite high.
I t is interesting to note that like Atlantic cod the FPA
of frozen stored channel catfish increased most rapidly
during the in itia l time period (Wood and Haqq, 1962).
The mean value of 0.36% FFA for fresh catfish is much
lower than that observed with other species. This may be
important from a storage point of view as i t is possible
that free fatty acids are more susceptible to rancidity
than neutral lipids or phospholipids, (Lovem, 1962).
Pacific gray cod and lingcod contained 3.4 and 5.2% FFA,
respectively, (Wood and Haqq, 1962)• Fresh Atlantic cod
contained 15% FFA (Dyer et a l., 1958) whereas fresh white-
fish contained 4.6% FFA (Awad et a l., 1969).
Extractable Protein: The mean crude protein content
of 6 catfish was 22.03%. This value agrees with that of
Drake et al. (1973) who observed that the crude protein
content of channel catfish was 21.44%. Fresh channel
catfish contained 91.62% extractable protein (% of crude
protein) (Table 8) which is similar to that of Awad et al.
(1969). The la tte r workers reported that about 93% of the
to tal protein in freshly frozen freshwater whitefish was
extracted with a 5% NaCl solution.
The extractable protein content of channel catfish
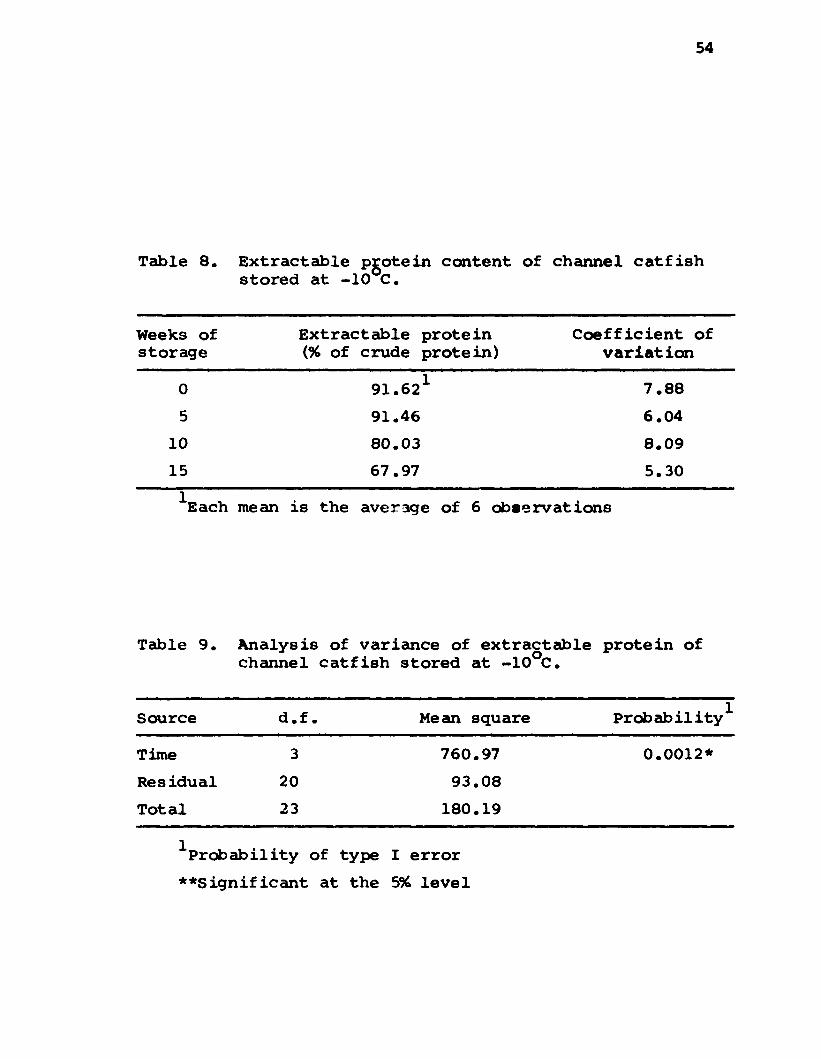
54
Table 8. Extractable protein content of channel catfish stored at -10 C.
Weeks of storage
Extractable protein Coefficient of (% of crude protein) variation
0 91.621 7.88
5 91.46 6.04
10 80.03 8.09
15 67.97 5.30
*Each mean is the average of 6 observations
Table 9. Analysischannel
of variance of extractable catfish stored at -10°C.
protein of
Source d .f . Mean square Probability*
Time 3 760.97 0.0012*
Residual 20 93.08
Total 23 180.19
^■probability of type I error
**Significant at the 5% level

55
significantly (P ^ 0.05) decreased from 91.62% at 0 weeks
of frozen storage to 67.97% after 15 weeks of frozen
storage (Tables 8 and 9). The coefficients of variation
indicated that the variability of extractable protein among
fish (samples) was quite low and relatively constant (Table
8). These attributes would make the extractable protein
test a good index of texture if i t correlated with texture.
It is interesting to note that extractable protein did not
s ta rt to decrease until after free fatty acids increased to
their greatest concentration (Tables 6 and 8). This would
suggest that with channel catfish there may be a relation
ship between free fatty acid increase and extractable
protein decrease. This relationship has been suggested
by several researchers for various species of fish (Olley
et a l., 1962; King et a l., 1962; and Anderson et a l., 1965).
Hypoxanthine: The mean hypaxanthine content of 6 fish
varied from 104.50 ug/g flesh to 61.16 ug/g flesh (Table 10).
S tatistical analysis revealed that hypaxanthine concentra
tion did not significantly change during frozen storage
at -10°C for 15 weeks (Tables 10 and 11). The coefficients
of variation indicated that the variation among samples at
the various storage times was quite high (Table 10). This
would make the hypoxanthine test a poor index of flavor.
One explanation of the high variability is that during the
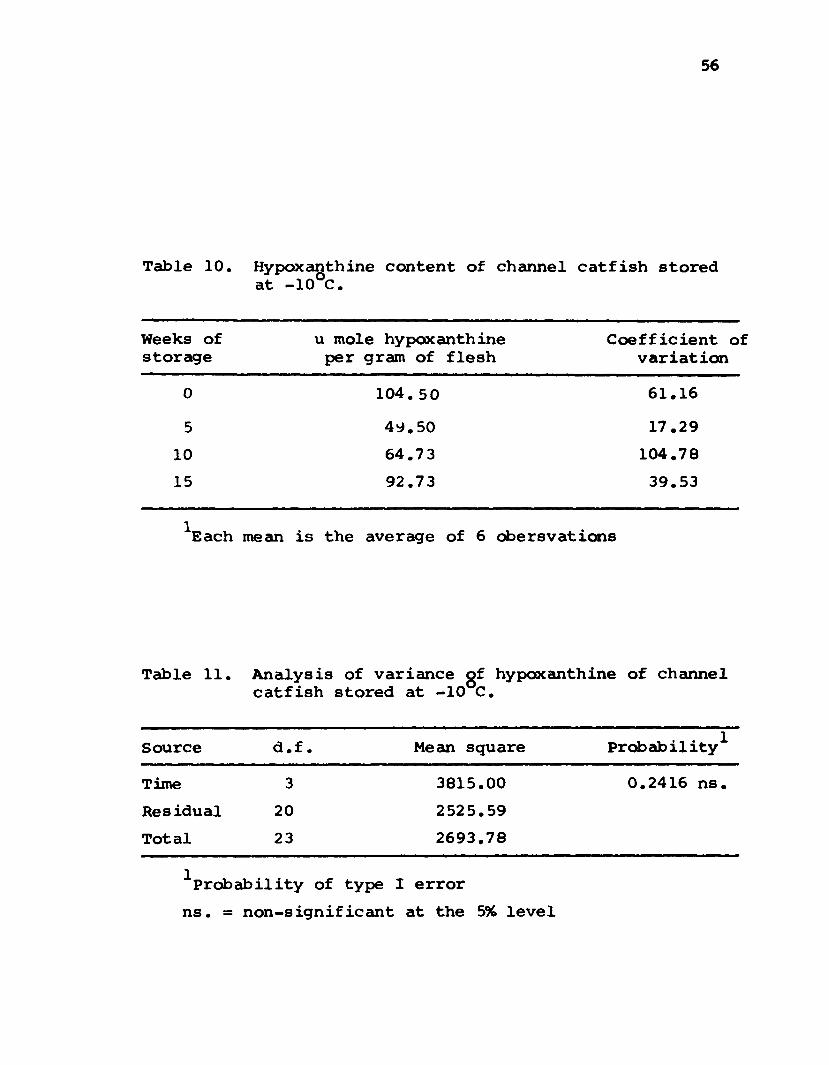
56
Table 10. Hypoxanthine content of channel catfish stored at -10 C.
Weeks of storage
u mole hypoxanthine per gram of flesh
Coefficient of variation
0 104.50 61.16
5 4*.50 17.29
10 64.7 3 104.78
15 92.73 39.53
"Each mean is the average of 6 obersvations
Table 11. Analysiscatfish
of variance of hypaxanthine of channel stored at -10 C.
Source d .f . Mean square Probability1
Time 3 3815.00 0.2416 ns.
Residual 20 2525.59
Total 23 2693.78
^"Probability of type I error
n s . = n o n - s i g n i f i c a n t a t t h e 5% l e v e l

57
spectrophotometric measurements of the hypoxanthine solu
tions the readings varied between 85 and 99% transmittance.
For greatest accuracy solutions should absorb in the range
of 15 to 65% transmittance (Ewing# 1969). At low concen
trations the error inherent in reading the galvanometer
or other indicating device becomes large compared to the
quantity which is being measured (Ewing, 1969). The varia
b ility was probably not due to the presence of dark muscle
as most of i t had been removed.
Apparent Inosine Monophosphate: The mean absorbance
values of apparent IMP from 6 fish varied from 0.5916 to
0.9408. I t should be emphasized that adenosine nucleotide
(ATP, ADP, and AMP) as well as IMP absorb at 256 nm (Spielli
et a l., 1964). The coefficients of variation indicated
that the variability of the absorbance values at any given
storage time was quite low. Consequently, absorbance values
of apparent IMP would make a good index of flavor if they
are correlated with flavor. S tatistical analysis revealed
that the absorbance values significantly (P £ 0.01) changed
during frozen storage at -10°C. Visual analysis of the
data presented in Table 12 indicated that the high absor
bance values at 10 weeks of storage was responsible for
the highly significant change. I t should be noted that
the mean absorbance value ^t 15 weeks of storage was

58
Table 12. Mean absorbance values of apparent inosinemonophosphate (IMP) content of channel catfish stored at -10 C.
Weeks of storage Absorbance at 2 56 nm
Coefficient of variation
0 0.59161 13.30
5 0.5998 8.26
10 0.9408 3.65
15 0.5952 5.30
*Each mean is the average of 6 observations
Table 13. Analysisapparentcatfish
of variance of absorbance inosine monophosphate of
stored at -10 C.
values of channel
Source d .f . Mean square Probability
Time 3 0.179000 0.0001**
Residual 20 0.000985
Total 23 0.024322
^Probability of type I error
* * S i g n i f i c a n t a t t h e 1% l e v e l

59
essentially equal to the mean absorbance values at 0 and
5 weeks of frozen storage. Therefore/ if one considers
the high values at 10 weeks of storage atypical then IMP
did not essentially change.
Organoleptic Analysis: The mean taste panel scores
for odor, flavor, texture, and overall quality, for the
most part varied between scores of 4 and 5 (Table 14).
A score of 6 was excellent and a score of 1 was very poor.
In general, odor, flavor, texture, and overall quality
were rated between very good and good, (Table 15)• In
contrast, Awad et al. (1969) observed that with freshwater
whitefish stored at -10°C, overall quality decreased from
a mean score of 3.5 (a score of 6 being excellent and a
score of 1 being poor) at 0 weeks of frozen storage to a
mean score of 1.5 after 16 weeks. I t should be emphasized
that the whitefish in Awad's study was 5 days post mortem
before the onset of frozen storage, the catfish used in
the present study were less than 30 hr post mortem before
the onset of frozen storage. This is very important as
the longer fish are held post mortem the probability
increases that the fish passed through the induction
period of oxidavite rancidity. The sooner fish are frozen
the longer will be the induction period of oxidative ran
cidity. In addition, the catfish used in the present

6 0
Table 14. Mean taste panel scores for odor, flavor, texture, and overall quality of channel catfish stored at -10 C.
Weeks of storage
Odor Flavor Texture Overall quality
0
i—im00• 4.441 4.701 4.48
5 5.15 4.15 4.56 4.33
10 4.59 3.85 4.51 3.92
15 5.00 4.11 4.77 4 .29
^Each mean is the average of 27 observations
Table 15. Analysis of variance of and overall quality of at -10 C.
odor, flavor, texture, channel catfish stored
Source d.f OdorM.S.
FlavorM.S.
Texture Overall quality M.S. M.S.
Time 3 1.51 ns. 1.59 ns 0.40 ns. 1.51 ns.
Judge 8 1.79 3.15 5.08 4.34
T x J 24 0.65 0.96 1.04 0.70
Residual 72 0.55 0.86 0.58 0.56
Total 107 0.69 1.07 1.01 0.90
n s . = n o n - s i g n i f i c a n t a t t h e 5% l e v e l .

61
study were very well packaged. The fish were tightly
wrapped several times with an oxygen impermeable film
which excluded oxygen which is necessary for the develop
ment of oxidative rancidity. Also, the oxygen impermeable
film appeared to prevent dehydration which if occurs to
a considerable extent can significantly lower the score
which the fish sample would receive. The organoleptic
results of the present study indicate that if catfish are
packaged properly and frozen soon after harvest their
organoleptic attributes do not significantly degrade
during storage at -10°C for 15 weeks.
Visual Correlations: Due to the small size of the
catfish available for the frozen storage study, both
chemical and organoleptic tests could not be conducted on
the same fish. Consequently, this prevented s ta tis tica l
correlations.
Free fatty acids significantly (P ^ 0.01) increased
in concentration during frozen storage, whereas odor,
flavor, texture, and overall quality did not significantly
change. In addition, the variability among samples at a
given time period was quite high. Consequently, free fatty
acids do not appear to be an useful index for any of the
organoleptic attributes tested.

62
Although the variability among samples at any given
storage time was low for extractable protein i t signifi
cantly (P ^ 0.05) decreased during frozen storage. Thus,
extractable protein does not appear to be an useful index
of texture or any of the other organoleptic attributes
tested.
Hypaxanthine like odor, flavor, texture, and overall
quality did not significantly change during frozen storage.
However, the large variability (Table 10) rendered the
test inappropriate as an index of flavor.
Apparent IMP appears to be the most promising as an
index of flavor even though i t significantly changed
during frozen storage (Table 13). Since the values at 5,
10, and 15 weeks of frozen storage were almost identical
(as were the flavor data) further study may prove i t a
useful index.

SUMMARY
The characteristics of lipids present in the dark and
light muscle of channel catfish were determined. Light
muscle contained 1.26% lipid which had an iodine number
of 93.86. In contrast, dark muscle contained 2 5.15%
lipid with an iodine number of 6.99.
The use of canning as a means of preserving catfish
and combating the off-flavor problem was investigated. A
technically feasible canned product was developed. It
was found desirable to remove the bone as well as add salt
and lemon juice to the fish immediately prior to thermal
processing. Removing the dark muscle did not significantly
affect odor, flavor, texture, and overall quality of the
canned product. Steaming of fish prior to canning decreased
odor, removed off-flavor, decreased flavor, improved texture,
and decreased overall quality. Storage of the canned pro
duct for 6 months tended to improve texture but did not
affect odor, flavor, or overall quality. The canned product
of steamed with dark muscle le f t on gave the greatest yield
(0.88 lb of whole catfish required to f i l l one 307 x 113
can). The product of non-steamed with dark muscle removed
63

64
gave the lowest yield (1.10 lb of whole catfish required
to f i l l one 307 x 113 can).
Experiments to determine if certain chemical tests
could correlate with texture and flavor of frozen stored
channel catfish were also conducted. Odor# flavor# tex
ture# and overall quality did not significantly change
oduring storage at -10 C for 15 weeks.
Free fatty acids increased# extractable protein
decreased# and hypoxanthine concentration did not signi
ficantly change during storage at -10°C for 15 weeks.
Neither free fatty acids# extractable protein# nor hypox
anthine correlated with texture and flavor. Apparent
inosine monophosphate appeared to hold the most promise
as an index of flavor.
Consumer acceptability# economic feasability# canning
of catfish packed in oil# and additional work with off-
flavor catfish requires further study.

LITERATURE CITED
Ackman, R. G. 1966. Empirical relationships between iodine value and polyunsaturated fatty acid content in marine oils and lipids. J . Am. Oil Chem. Soc. 43:385-389.
Ammerman, G. R. 1973. Introductry remarks, fish and seafood program, Association of Southern Agricultural Workers Inc., Atlanta, Ga., Feb. 4-7.
Anderson, M. L., M. A. Steinberg, and F. J . King. 1965.Seme physical effects of freezing fish muscle and their relation to protein-fatty acid interaction, in Kreuzer, R. (ed.) The Technology of Fish Utilization, Fishing News (Books) Ltd., London, 1965, pp. 105-110.
A.O.A.C. 1970. Official Methods of Analysis of the Association of Official Analytical Chemists, George Banta Co., Inc., Menasha, Wisconsin.
Awad, A., W. D. Pcwrie, and o . Fennema. 1969. Deterioration of fresh-water whitefish muscle during frozen storage at -10 C. J . Food Sci. 34:1-9.
Beuchat, L. R. 1973. Hypoxanthine measurement in assessing freshness of chilled catfish (Ictalurus punctatus) . J . Agr. Food Chem. 21:453-455.
Beuchat, L. R., E. K. Heaton, and T. S. Boggess J r . 1973. Preservation of channel catfish with some selected chemicals. J . Food Sci. 38:531-535.
Bilinski, E. 1969. Lipid catabolism in fish muscle, inNeuhaus, O. W. and J. E. Halver (eds.), Fish in Research, Academic Press, pp. 135-155.
Bilinski, E. and Y. C. Lay. 1969. Lipolytic activitytoward long-chain triglycerides in the lateral line muscle of rainbow trout (Salmo qairdneri) . J . Fish.Res. Bd. Can. 26:1857-1866,
65

6 6
Bligh, E. G. 1961. Lipid hydrolysis in frozen cod muscle.J . Fish. Res. Bd. Can. 18:143-145.
Bligh/ E. G. and W. J. Dyer. 1959. A rapid method of total lipid extraction and purification. Can. J . Biochem. Physiol. 37:911-917.
Bligh/ E. G. and M. A. Scott. 1966. Lipids cod muscleand the effect of frozen storage. J . Fish. Res. Bd.Can. 23:1025-1036.
Boggess/ T. S. J r ., E. K. Heaton/ and A. L. Shewfelt.1971. Storage stab ility of commercially prepared and frozen pond-raised channel catfish (Ictalurus punctatus# rafinisgue). J . Food Sci. 36:969-973.
Bosund/ I. and B. Ganrot. 1969. Lipid hydrolysis in frozen baltic herring. J . Food Sci. 34:13-18.
Botta/ J . R./ J . F. Richards, and N. Tomlinson. 1973a.Thiobarbituric acid value, total long-chain free fatty acids, and flavor of Pacific halibut (Hippoqlossus stenolepis) and Chinook salmon (Oncorhynus tshawvtscha) frozen at sea. J . Fish. Res. Bd. Can. 30:63-69.
Botta, J . R., J . F. Richards, and N. Tomlinson. 1973b.Flesh concentration of various long-chain free fatty acids of Pacific halibut (Hippoglossus stenolepis) and Chinook salmon (Oncorhynus tshawytscha) frozen at sea. J . Fish Res. Bd. Can. 30:79-82.
Botta, J . R. and J . E. Rutledge. 1973. Chemical and physical factors affecting the organoleptic quality of canned catfish (Ictalurus punctatus, rafinisque). Presented at 70th annual convention, Association of Southern Agricultural Workers, Inc., Atlanta, Ga., Feb. 4-7.
Brocklesby, H. N. 1933. The hydrolysis of body oil of the salmon. Contrib. Can. Biol. Fish. 7:507-519.
Castell, C. H. and D. M. Bishop. 1973. Effect of season on salt-extractable protein in muscle from trawler- caught cod and on its stab ility during frozen storage.J. Fish. Res. Bd. Can. 30:157-160.

67
Connell/ J . J. 1968. The effect of freezing and frozen storage on the proteins of fish muscle/ in Hawthorn/J . and E. j . Rolfe (eds.), Low Temperature Biology of Foodstuffs, Pergamon Press, Toronto, 1968. pp 333-358.
Connell, J . J . 1969. Changes in the eating quality offrozen stored cod and associated chemical and physical changes, in Kreuzer, R. (ed.). Freezing and Irradiation of Fish, Fishing News (Books) Ltd., London, 1969. pp 323-329.
Connell, J . J . and P. Hcwgate. 1968. Sensory and objective measurement of the quality of frozen stored cod of different in itia l freshnesses. J . Sci. Fd. Agric. 19:342-354.
Cowie, W. P. and W. T. L ittle . 1966. The relationshipbetween the toughness of cod stored at -29 C and its muscle protein solubility and pH. J . Fd. Technol. 1:335-343 .
Cowie, W. P. and W. T. L ittle . 1967. The relation between the toughness of cod stored at -7 C and -14 C, its muscle protein solubility, and muscle pH. J . Fd. Technol. 2:217-222.
Creelman, V. M. and N. Tomlinson. 1960. Inosine in themuscle of Pacific salmon stored in ice. J . Fish. Res. Bd. Can. 17:449-451.
Drake, S. R. and G. R. Ammerman. 1973. Shelf-life andacceptability of precooked channel catfish. Presented at 70th annual convention, Association of Southern Agricultural Workers, Inc., Atlanta, Ga., Feb. 4-7,1973.
Drake, S. R., J. B. Magee, and G. R. Ammerman. 1973. process calculations and evaluations of canned channel catfish. Presented at 70th annual convention, Association of Southern Agricultural Workers, Inc., Atlanta, Ga.,Feb. 4-7, 197 3.
Dyer, W. J . and D. I. Fraser. 1959. Proteins in fishmuscle 13. Lipid hydrolysis. J . Fish. Res. Bd. Can. 16:43-53.

6 6
Dyer, W. J . and M. L. Morton. 1956. Storage of frozen plaice f i l le ts . J . Fish. Res. Bd. Can. 13:129-134.
Dyer, W. J ., D. I. Fraser, and B. G. Bligh. 1958. Fathydrolysis in frozen fish I . Free fatty acid formation. Progress Reports Atlantic Coast Stations Fish.Res. Bd. Can. 71:17-19.
Dyer, W. J ., B. I. Fraser, and D. P. Lohnes. 1966. Nucleotide degradation and quality in ordinary and red muscle of iced and frozen swordfish (Xiphias qladius) . J.Fish. Res. Bd. Can. 23:1821-1833.
Dyer, W. J ., H. V. French, and J . M. Snow. 1950. Proteins in fish muscle I. Extraction of protein fractions in fresh fish. J . Fish. Res. Bd. Can. 7:585-593.
Dyer, W. J ., M. L. Morton, D. I. Fraser, and E. G. Bligh.1956. Storage of frozen rosefish f i l le ts . J . Res.Bd. Can. 13:569-579.
Dyer, W. J ., D. I. Fraser, R. G. Macintosh, and M. Myer.1964. Cooking method and palatability of frozen f i l le ts of various qualities. J . Fish. Res. Bd. Can. 21:577-589.
Ewing, G. W. 1969. Instrumental Methods of Chemical Analysis. McGraw-Hill, New York, pp 69-72.
Fraser, D. I. and W. J. Dyer. 1959. Fat hydrolysis infrozen fish 2. Relation to protein stab ility . Progress Reports Atlantic Coast Stations Fish. Res. Bd. Can. 72:643-652.
Fraser, D. I ., D. P. Pitts, and W. J . Dyer. 1968a. Nucleotide degradation and organoleptic quality in fresh and thawed mackerel held at the above ice temperature.J . Fish. ReS. Bd. Can. 25:239-253.
Fraser, D. I ., S. G. Simpson, and W. J . Dyer. 1968b. Very rapid accumulation of hypoxanthine in the muscle of redfish stored in ice. J . Fish. Res. Bd. Can. 25: 817-821.
Fujii, Y., E. Nogouchi, and K. S. Su. 1968. Relationbetween the quality of canned fish and its content of ATP breakdowns-I. ATP breakdowns in canned pink salmon and mackerel in relation to their inspection grades.Bull. Jap. Soc. ,Sci. Fish. 34:1031-1035.

6 9
Gerber, N. N. and H. A. Lechevalier. 1965. Geosmln, anearthy-smelling substance, isolated from Actinomycetes. Appl. Microbiol. 13:935-938.
Greenfield, J . E. 1970. Economic and business dimensions of the catfish farming industry. USDI Report, Bureau of Cornrn. Fish., Ann Arbor, Michigan.
Gould, E. 1971. in Gould, E. and J . A. Peters. On testing the freshness of frozen fish, Fishing News (Books)Ltd., London, 1971. 80 p .
Hanson, S. W. F. and J. Olley. 1965. Observations on the relationship between lipids and protein deterioration. in Kreuzer, R. (ed.). The Technology of Fish U tilization, Fishing News (Books) Ltd., London, 1965. pp 105- 110.
Heaton, E. K., J . Page, J . W. Andrews, and T. S. Boggess Jr . 1972. Changes in quality of channel catfish held on ice before and after processing. J . Food Sci. 37: 841-844.
Ironside, J. I. M. and R. M. Love. 1956. Measurement ofdenaturation of cold-stored fish. Nature. 178:988-989.
Jonas, R. E. E. and E. Bilinski. 1967. Glycerylphospho- choline and related compounds in rainbow trout muscle and its relationship to protein denaturation. J . Fish. Res. Bd. Can. 24:263-280.
Jones, N. R. 1960. The separation and determination of free purines, pyrimidines, and nucleoside in cod muscle. Analyst. 85:111-115.
Jones, N. R. 1965. Hypaxanthine and other purine-containing fractions in fish muscle as indices of freshness* in Kreuzer, R. (ed.)*>,The Technology of Fish Utilization, Fishing News (Books) Ltd., London, pp 179-184.
Jones, N. R. and J . Murray. 1962. Degradation of adenine and hypaxanthine nucleotide in the muscle of chilled- stored trawled cod (Gadus callarias) . J . Sci. Food Agric. 13:475-480.

7 0
Kassemsam, B., B. Sanz Perez, J . Murray, and N. R. Jones. 1963. Nucleotide degradation in the muscle of iced haddock (Gadus aeglefinus), lemon sole (Pleuronectes microcephalus), and plaice (Pleuronectes platessa) •J. Food Sci. 28:28-37.
King, F. J . . , M. L. Anderson, and M. A. Steinberg. 1962.Reaction of cod actcmysosin with linoleic and linolenic acids. J . Food Sci. 27:363-366.
Li, J . C. R. 1964. S tatistical Inference 1. EdwardsBrothers, Inc., Ann Arbor, Michigan.
Love, R. M. 1956. Infleunce of freezing rate on thedenaturation of cold-stored fish. Nature. 178:988-989.
Lovell, R. T. 1971. Off-flavor. The Catfish Farmer.Nov. 1971. pp 7-9.
Lovell, R. T. 1973. Absorption of an earthy-musty flavorby channel catfish from cultures of geosmin-producing algae in controlled environments. Presented at 70th annual convention, Association of Southern Agricultural Workers, Inc., Atlanta, Ga., Feb. 4-7, 197 3.
Lovem, J . A. 1962. The lipids of fish and changesoccurring in them during processing and storage, in Heen, E. and R. Kreuzer (eds.). Fish in Nutrition, Fishing News (Books) Ltd., London, pp 86-110 .
Luijpen, A. F. M. G. 1957. Denaturation of fish proteins. Nature. 180:1422-1423.
Mackie, P. R. and R. Hardy. 1969. A Lipid analysis of the greater silver smelt (Argentina silus, ascanuis) and an evaluation of its potential food and fish meal production. J . Food Technol. 4:241-254.
Maligalig, L. L., J . F. Caul, and O. W. Teimeier. 1973. Aroma and flavor of farm-raised channel catfish: effects of pond condition, storage, and d iet. Food Prod. Develop. 7(4):86-92.
Medsker, L. L., D. Jenkins, and J . F. Thomas. 1968.Odorous compounds in natural waters: an earthy-smellig compound associated with blue-gree algae and Actino- mycetes. Environ. Sci. Tech. 2:461-464.

71
Moorjani, M. W., W. A. Montgomery, and G. G. Coote. 1960. Correlation of taste panel gradings with salt-extrac- table protein of frozen fish f i l le ts . Food Res. 25:263-269.
Murray, J ., N. R. Jones, and J. R. Burt. 1966. Hypaxanthine in the muscle of chilled-stored Atlanta . Salmon (Salmo salgar) . J . Fish Res. Bd. Can. 2? .975-1797.
Ncwlan, S. S. and w. J . Dyer. 1969. Glycc. auxc and nucleotide Qchanges in the critica l freezing zone, -0.8 to -5.0 C, in prefrozen cod muscle frozen at various rates. J . Fish. Res. Bd. Can. 26:2621-2632.
Olley, J . and J . A. Lovem. I960. Phosolipid hydrolysis in cod stored at various temperatures. J . Sci. Food Agric. 11:644-652.
Olley, J ., J . Farmer, and E. Stephen. 1969. The rate ofphospholipid breakdown in frozen fish. J . Food Technol. 4:27-37.
Olley, J ., R. Pirie, and H. Watson. 1962. Lipase and phospholipase activity in fish skeletal muscle and its relationship to protein denaturation. J . Sci.Food Agric. 13:501-516.
Olley, J ., E. Stephen, J . Farmer, and I. Robertson. 1967.A critica l look at two objective tests for cold storage deterioration. J . Fd. Technol. 2:207-216*
Peters, J . A., W. A. MacCallum, W. J . Dyer, D. R. Idler,J . W. Slavin, J . P* Lane, D. I . Fraser, and E. J . Laishley. 1968. Effect of stage of rigor and of freezing-thawing processes on storage quality of refrozen cod. J . Fish. Res. Bd. Can. 25:299-320.
Safferman, R. S., A. A. Rosen, C. I. Mashni, and M. E.Morrie. 1967. Earthy-smelling substance from a blue- green alga. Environ. Sci. Tech. 1:429-431.
Saito, T. and K. Arai. 1957. Studies on the organicphosphates in muscles of aquatic animals-V. Changes muscular nucleotides of carp during freezing and storage. Bull. Jap. Soc. Sci. Fish. 23:265-268 .

72
Saito, T., K. Arai, and T. Yajima. 1959. Changes inpurine nucleotides of red lateral muscle of rainbow trout. Nature. 184:1415.
Spinelli, J ., M. Eklund, and D. Miyauchi. 1964. Measurement of hypoocanthine in fish as a method of assessing freshness. J . Food Sci. 29:710-714.
Snow, J . M. 1950. Proteins in fish muscle. I I . Colorimetric estimation of fish muscle protein. J . Fish.Res. Bd. Can. 7:594-598.
Thaysen A. C. 1936. The origin of an earthy or muddytaint in fish. I. The nature and isolation of the ta in t. Ann. Appl. Biol. 23:99-104.
Thaysen/ A. C. and F. T. K. Pentelow. 1936. The originof an earthy or muddy tain t in fish. I I . The effect on fish of the tain t produced by an odoriferous species of Actinomyces. Ann. Appl. Biol. 23:105-109.
Tomlinson/ N. and S. E. Geiger. 1963. The bound nucleotides of freshly frozen and severly denaturated lingcod muscle. J . Fish. Res. Bd. Can. 20:187-194.
Tomlinson/ N., S. E. Geiger, and S. W. Roach. 1969.Quality of sea frozen fish from the North-eastern Pacific Ocean, iji Kreuzer, R. (ed.), Freezing and Irradiation of Fish. Fishing News (Books) Ltd., London, pp 68-75.
Triebold, H. O. and L. W. Ayr and. 1963. Food Composition and Analysis, D. Van Nostrand Co., Inc., Princeton,N. J . pp 172.
Yurkowski, M. and H. Brockerhoff. 1965. Lysolechithinase of cod muscle. J . Fish. Res. Bd. Can. 30:63-69.
Wood, J . O. and S. A. Haqq. 1962. Fat hydrolysis infrozen f il le ts of lingcod and Pacific cod. J . Fish. Res. Bd. Can. 19:169-171.
Worthington, R. E., T. S. Boggess, J r ., and E. K. Heaton.1972. Fatty acids of channel catfish (Ictalurus punctatus) . J . Fish Res. Bd. Can. 29:113-115.

VITA
Joseph Richard Botta was bom on March 18# 1946/ at
O'Brian Bay, British Columbia/ Canada. He received his
primary and elementary education via British Columbia
Correspandance School. He graduated from Kitsilano Senior
Secondary School in June, 1965. In September/ 1965, he
enrolled in the Faculty of Agriculture/ University of
British Columbia which granted him the degree of Bachelor
of Science in Agriculture (majoring in Food Science) in
May, 1969/ and the degree of Master of Science in Food
Science in 1971. In that same year he undertook graduate
work, majoring in Food Science and minoring in Heme
Economics (Nutrition) and Horticulture at Louisiana State
University. He is presently a candidate at Louisiana State
University for the degree of Doctor of Philosophy.
73

EXAMINATION AND THESIS REPORT
Candidate:
Major Field:
Title of Thesis:
Josep h R ichard B o tta
Food S c ie n c e
"Chem ical and O r g a n o lep tic Q u a lity Changes o f Frozen S tored and Canned Channel C a tf ish "
Approved:
Major Professor and Chairrrtan
Dean of the Graduate School
EXAMINING COMMITTEE:
✓f*!- - 3 YLnraA.
Date of Examination:
November 20 , 1973