characterization ofthe light-regulated operonencoding the ... · proteins cpec and cped with...
TRANSCRIPT

Vol. 172, No. 7JOURNAL OF BACTERIOLOGY, JUlY 1990, p. 4072-40810021-9193/90/074072-10$02.00/0Copyright C) 1990, American Society for Microbiology
Characterization of the Light-Regulated Operon Encoding thePhycoerythrin-Associated Linker Proteins from the Cyanobacterium
Fremyella diplosiphontNANCY A. FEDERSPIEL'* AND ARTHUR R. GROSSMAN2
Department ofBacteriology and Biochemistry, University of Idaho, Moscow, Idaho 83843,1 and Department of PlantBiology, Carnegie Institution of Washington, 290 Panama Street, Stanford, California 943052
Received 12 March 1990/Accepted 3 May 1990
Many biological processes in photosynthetic organisms can be regulated by light quantity or light quality or
both. A unique example of the effect of specific wavelengths of light on the composition of the photosyntheticapparatus occurs in cyanobacteria that undergo complementary chromatic adaptation. These organisms alterthe composition of their light-harvesting organelle, the phycobilisome, and exhibit distinct morphologicalfeatures as a function of the wavelength of incident light. FremyeUa diplosiphon, a filamentous cyanobacterium,responds to green light by activating transcription of the cpeBA operon, which encodes the pigmentedlight-harvesting component phycoerythrin. We have isolated and determined the complete nucleotide sequence
of another operon, cpeCD, that encodes the linker proteins associated with phycoerythrin hexamers in thephycobilisome. The cpeCD operon is activated in green light and expressed as two ma,jor transcripts with thesame 5' start site but differing 3' ends. Analysis of the kinetics of transcript accumulation in cultures of F.diplosiphon shifted from red light to green light and vice versa shows that the cpeBA and cpeCD operons are
regulated coordinately. A common 17-base-pair sequence is found upstream of the transcription start sites ofboth operons. A comparison of the predicted amino acid sequences of the phycoerythrin-associated linker
proteins CpeC and CpeD with sequences of other previously characterized rod linker proteins shows 49invariant residues, most of which are in the amino-terminal half of the proteins.
Cyanobacteria are oxygenic photosynthetic procaryoteswhose primary light-harvesting apparatus is the phycobili-some (12, 17, 22). These structures are peripherally associ-ated with the thylakoid membranes and efficiently transfer abroad spectrum of light energy to the photosynthetic reac-tion centers. In most cyanobacteria, the phycobilisome is ahemidiscoidal structure that is composed of two distinctsubstructures: a core complex and six rods radiating fromthe core (21). The pigmented proteins of the phycobilisome(phycobiliproteins) are hydrophilic polypeptides with cova-lently attached tetrapyrrole chromophores. These proteinsconstitute -85% of the mass of the complex, with theremainder comprising nonpigmented "linker" proteins (48).Both the pigmented and nonpigmented constituents of thephycobilisome rods can vary depending on growth condi-tions, such as temperature, light quality and quantity, andthe nutritional status of the growth medium (2, 5, 25, 35, 38,47, 49).The phycobilisome contains three classes of phycobilipro-
teins. Allophycocyanin is the major component of the corecomplex (21). Phycocyanin constitutes the portion of therods adjacent to the core complex, while phycoerythrin orphycoerythrocyanin, when present, is found at the peripheryof the rods. Each type of phycobiliprotein monomer iscomposed of equimolar amounts of two subunits, a and 1,which assemble into trimers and hexamers. Assembly intohigher order structures is mediated by association withparticular linker proteins that determine the assembly path-way of the macromolecular complex and optimize energytransfer from the periphery of the rods to the phycobilisome
* Corresponding author.t Carnegie Institution of Washington publication 1071 and Idaho
Agricultural Experiment Station publication 9051.
core and ultimately to the photosynthetic reaction center(56).Many cyanobacteria alter the composition of their phyco-
bilisomes in response to light quality, or wavelength, aphenomenon known as complementary chromatic adapta-tion (7, 48). The altered phycobilisome composition resultsin optimal absorption of prevalent wavelengths of light and isa consequence of de novo synthesis of the appropriatephycobiliproteins and linker proteins (9). In Fremyelladiplosiphon, phycoerythrin is synthesized in response togreen light but not to red light (6, 18, 51), whereas increasedlevels of phycocyanin are found in red light but not in greenlight (26, 53). These responses appear to be regulated pri-marily at the transcriptional level (13, 33, 36).
In F. diplosiphon, there are three operons encoding phy-cocyanin subunits that are expressed under different condi-tiorns. The cpcl operon encodes the ,B and a subunits of aphycocyanin which is expressed at similar levels in greenand red light (constitutive phycocyanin) (14). The cpc2operon encodes phycocyanin subunits and associated linkerproteins that are expressed in red, but not in green, light(inducible phycocyanin) (14, 32). In phycobilisomes fromcells grown in both red and green light, the constitutivephycocyanin probably occupies the portion of the rodsproximal to the core; the rod substructure is completed inred light-grown cells by the inducible phycocyanin hexamerswith associated linker proteins. The third phycocyanin op-eron, cpc3, encodes a set of phycocyanin subunits and linkerproteins which replaces the red light-inducible phycocyaninand its associated linkers under conditions of sulfur depri-vation (34, 35).
In contrast to the multiple cpc operons, there is only oneset of genes encoding the ,B and a subunits of phycoerythrinin F. diplosiphon. This operon, cpeBA, is expressed at high
4072
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

CYANOBACTERIAL LIGHT-REGULATED PROTEINS 4073
levels in green light and at very low levels in red light (33,36). In green light, the phycoerythrin hexamers occupy thedistal portions of the phycobilisome rods in place of phyco-cyanin hexamers (17). Also, unlike many of the phycocyaninoperons, the genes encoding the linker proteins that associ-ate with phycoerythrin are not linked to cpeBA. These linkerproteins are highly expressed in green light and are requiredfor the proper assembly of phycoerythrin into the rods of thephycobilisome (19, 20). No genes encoding phycoerythrin-associated linker proteins have previously been describedfor any organism, although a partial amino acid sequence ofa linker protein associated with phycoerythrocyanin fromMastigocladus laminosus has been published (16).
Here, we describe the isolation and characterization of thecpeCD operon encoding the phycoerythrin-associated linkerproteins in F. diplosiphon. The cpeC and cpeD genes arecotranscribed at high levels in green light and at barelydetectable levels in red light. We have examined changes insteady state transcript levels from both of the green light-regulated operons (cpeBA and cpeCD) after changing condi-tions of illumination during growth from green to red lightand vice versa. In addition, we have determined the sites oftranscription initiation for both operons. A comparison ofthe amino acid sequences of the two linker proteins CpeCand CpeD to each other and to published sequences of linkerproteins associated with other phycobiliproteins defines re-gions of sequence conservation which may have both struc-tural and functional importance.
MATERIALS AND METHODS
Materials. All chemicals were reagent grade. Restrictionenzymes were obtained from Bethesda Research Laborato-ries, Inc., Boehringer Mannheim Biochemicals, U.S. Bio-chemicals Corp., and Pharmacia, Inc. Klenow fragment ofDNA polymerase I, T4 DNA ligase, Sequenase, calf intes-tinal alkaline phosphatase, and urea were purchased fromU.S. Biochemicals; T4 polynucleotide kinase, reverse tran-scriptase, and Si nuclease were from Pharmacia. Radioac-tive nucleotides ([y-32P]ATP [>5,000 Ci/mmol], [a-32P]dCTP[>3,000 Ci/mmol], and [a-35S]dATP [>600 Ci/mmol]) wereobtained from Amersham Corp.
Phycobilisome preparation and analysis. F. diplosiphon 33(a strain of Calothrix sp., PCC 7601; a subculture of UTEX481) was grown at 30°C in BG-11 medium with 50 mMHEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid) (pH 8.0) in an atmosphere of 5% CO2 in air) in red orgreen light at 15 microeinsteins/cm2 as described by Bruns etal. (8). Phycobilisomes were isolated from red light- andgreen light-grown cells as previously described (2). Sodiumdodecyl sulfate (SDS)-polyacrylamide gel electrophoresiswas performed as described by Laemmli (31) and as modifiedby Anderson et al. (2). The linker polypeptides specificallyexpressed in green light (CpeC and CpeD) were isolatedfrom a preparative polyacrylamide gel and were injected as amixture into rabbits to raise polyclonal antibodies. Proteinswere transferred from polyacrylamide gels to nitrocelluloseby the method of Towbin et al. (51) and were stainedimmunologically with the polyclonal antibodies and proteinA-conjugated horseradish peroxidase (Boehringer Mann-heim) (3).
Preparation of an expression library. Genomic DNA fromF. diplosiphon 33 was digested with Sau3A in four separatereactions ranging from -10 to 90% completion. DNA frag-ments 1 to 3 kilobases (kb) in size were excised afterelectrophoresis in 0.7% agarose. The DNA was isolated
from the gel by the freeze-squeeze method (50) and precip-itated with ethanol. XZAP (Stratagene [46]) DNA was di-gested to completion with XhoI. Klenow fragment and theappropriate nucleotides were used to fill in the first two basesof the Sau3A and XhoI sites, generating a two-base compat-ible sequence. The DNA was ligated with T4 DNA ligase,plated on Escherichia coli BB4, and screened with theantibodies raised against CpeC and CpeD as describedabove. One immunopositive X clone was obtained, and theinternal Bluescript SK- plasmid containing a 1.6-kb insertwas rescued in vivo from this X clone in E. coli XL-1 Blue(46). Larger clones containing 5' and 3' sequences wereobtained by screening an EMBL3 library and the XZAPlibrary with fragments of the 1.6-kb immunopositive clone.Single-stranded template DNA was prepared from Blue-script, pUC118, and pUC119 plasmids containing fragmentsof the cpeCD genes by using the helper phage M13KO7 (52).DNA sequencing was performed by the dideoxy-chain ter-mination method (41) on single-stranded and double-stranded templates by using Sequenase, according to theprotocols of the manufacturer, with the M13 primer, reverseprimer, and other appropriate primers synthesized on aBiosearch 8600 oligonucleotide synthesizer. DNA sequenceanalyses and protein homology alignments were performedby using the IBI Pustell Sequence Analysis Programs ver-sion 2.02. Predictions of RNA folding were obtained byusing PCFOLD version 3.0 by Michael Zuker (57).RNA isolation and analyses. RNA was isolated from F.
diplosiphon 33 grown in red or green light by lysis inguanidinium chloride and CsCl centrifugation as describedpreviously (13, 24). Electrophoresis of RNA (2 jig) wasperformed on 1% agarose-formaldehyde gels in 20 mMMOPS (morpholinepropanesulfonic acid) buffer (pH 8.0); theRNA was transferred to nitrocellulose and probed withfragments labeled with [a-32P]dCTP by the random oligonu-cleotide priming method (15). Primer extension analysis (54)was used to determine the 5' end of the mRNA encodingCpeC and CpeD. A 24-base oligomer complementary tonucleotides 63 through 86 of the coding region for CpeC wassynthesized, end labeled with [_-32P]ATP, and used as aprimer for reverse transcriptase with 10 Rxg of RNA isolatedfrom cells grown in red or green light. The products wereanalyzed on a denaturing polyacrylamide gel with a sequenc-ing ladder generated by using the same primer as a standard.The position of the transcription start site was confirmed byS1 nuclease protection (44) by using as probe the 890-base-pair (bp) AvaII-PstI fragment labeled at the 5' end atthe AvaII site. The probe was hybridized to 50 ,g ofRNA at30 and 40°C in a total volume of 30 RI; digestion of thehybrids with 38 U of S1 nuclease per 330 ,ul at 37°C gaveoptimal results.
S1 nuclease protection was also used to determine thetranscription start site for the phycoerythrin operon(cpeBA). A 270-bp XbaI-HaeIII fragment starting 20 nucle-otides (nt) within the coding region for CpeB was labeled atthe XbaI site, and -0.25 pmol was annealed to 50 ,g ofRNAat 35°C overnight in a total volume of 50 RI. The hybridswere digested for 60 min at room temperature with 2,000 Uof S1 nuclease per 550 pI.
RESULTS
Cloning and characterization of the phycoerythrin-associ-ated linker genes (cpeCD). Preparations of phycobilisomesfrom cultures of F. diplosiphon 33 grown in red or green lightcan be resolved by SDS-polyacrylamide gel electrophoresis
VOL. 172, 1990
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

4074 FEDERSPIEL AND GROSSMAN
a) b)R G R G
97.4
66.2
42.7
Linker 1310- -..-
Phycobifiprotein 21.5Subunits 3
14.4
FIG. 1. Polyacrylamide gel electrophoresis of phycobilisomesisolated from F. diplosiphon 33 cultures grown in red (lanes R) or
green (lanes G) light. (a) The gel was stained with Coomassie blue;size standards are in kilodaltons, and the linker proteins associatedwith phycoerythrin are marked with arrows. (b) Western blot of a
duplicate gel as described for panel a developed with antibodiesraised to a mixture of the phycoerythrin-associated linker proteins.
(Fig. 1). Differences in both the phycobiliproteins and thelinker proteins are evident in phycobilisome preparationsisolated from different light regimens as previously described(9, 32). The phycoerythrin-associated linker proteins (CpeCand CpeD) are present at high levels in green light (Fig. la)and can be detected by specific antibodies on Western blots(immunoblots) (Fig. lb). The low levels of CpeC and CpeDdetected by the antibodies in red light are consistent with thelevels of phycoerythrin (CpeA and CpeB) which are ob-served under these growth conditions (8). The two smallerbands detected on the Western blot below the predominantlinker protein bands are probably degradation products ofthe linker proteins and cannot be detected on the Coomassie
Resitictlon Map
Clones
blue-stained gel. No cross-reactivity is seen with the phyco-cyanin-associated linker proteins expressed in red light orwith the constitutive 29-kilodalton (kDa) linker protein.The antibodies raised to the phycoerythrin-associated
linker proteins were used to screen an expression library 4fF. diplosiphon 33 DNA constructed in XZAP; one immuno-positive clone containing an insert of -1.6 kb was obtainedand is designated in Fig. 2 as clone 1. This clone containedthe entire coding region for CpeC and the amino-terminalhalf of CpeD. Fragments of this clone were used to screen anEMBL3 library and to rescreen the XZAP library to obtainclones containing flanking regions. A 3.0-kb HindIII-Sau3Afragment (Fig. 2, clone 2) subcloned from an EMBL3 phageadded both the region upstream of cpeC and the remainderof the coding region of CpeD, including -200 bp down-stream. A 3.0-kb ClaI fragment (Fig. 2, clone 3) was ob-tained from another XZAP clone which extended further inthe 3' direction. Restriction mapping, Southern blotting, andDNA sequencing were used to construct the composite mapshown in Fig. 2. The two open reading frames of the cpeCDoperon have predicted amino acid sequences that are highlyhomologous to each other and to the phycocyanin-associ-ated linker proteins (CpcH2 and CpcI2) from F. diplosiphon(32); Table 1 gives the nomenclature used for the variouslinker proteins (30). The nucleotide sequence for this region,including -450 bp upstream of the presumed start codon forCpeC, and the derived amino acid sequences for CpeC andCpeD are presented in Fig. 3.
In F. diplosiphon, the phycoerythrin-associated linkergenes (cpeCD) are not closely linked to the phycoerythringenes (cpeBA); an EMBL3 clone containing cpeBA approx-imately centered within a 15-kb insert did not hybridize on aSouthern blot to a fragment containing the linker genes (datanot shown). The first gene in the cpeCD operon, cpeC, ispredicted to encode a protein of 31.8 kDa with a pI of 9.2. Aconsensus ribosome binding site (AGGAG) (23, 45) precedesthe initial methionine codon by 9 nt. The second gene (cpeD)encodes a protein of 27.9 kDa with a pI of 8.7 and isseparated from cpeC by 40 nt, with a consensus ribosomebinding site (AGGAG) 11 nt upstream of the initial methio-
200 bp
1.2peC cpeD
2
4
4.-* 44- 4-
4-
b c
FIG. 2. Partial restriction map and sequencing strategy for the cpeCD operon encoding the phycoerythrin-associated linker proteins.Templates were derived from the following clones: 1, a 1.6-kb Sau3A fragment isolated from a XZAP clone; 2, a 3.0-kb HindIII-Sau3Afragment isolated from an EMBL3 clone; 3, a 3.0-kb ClaI fragment obtained from a XZAP clone. The initiation site for transcription isindicated by the arrow above the restriction map. The regions encoding the two linker proteins CpeC and CpeD are indicated by open boxeson the restriction map. The fragments indicated in the bottom line were used as probes of Northern blots as shown in Fig. 4. The indicatedrestriction sites are as follows: H, HindIII; P, PstI; S, Sau3A; A, Avall; K, KpnI; C, ClaI.
SequencingStrategy
Transcripts
NorthernProbes
3200 nt
d
J. BACTERIOL.
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

CYANOBACTERIAL LIGHT-REGULATED PROTEINS 4075
TABLE 1. Cloned cyanobacterial phycobilisome linker genes and characteristics of light-regulated expression
Cloned gene Organism Protein product and regulation Reference
cpeCD F. diplosiphon CpeC and CpeD; expressed in green light with phycoeryth- This reportrin (cpeBA)
cpcH2I2D2 F. diplosiphon CpcH2, CpcI2, and CpcD2 (formerly known as LR75, L39, 32and L8 ); expressed in red light with inducible phycocya-nin (cpcB2A2)
cpcH3I3D3 F. diplosiphon CpcH3, CpcI3, and CpcD3; induced under sulfur stress 35with phycocyanin (cpcB3A3)
cpcCD Anabaena strain 7120 CpcC and CpcD; expressed in white light with phycocyanin 5(cpcBA)
cpcCD Synechococcus strain 7002 CpcC and CpcD; expressed in white light with phycocyanin 10(cpcBA)
apcC Synechococcus strain 6301 ApcC; associated with the core (L78); expressed in white 29light with allophycocyanin (apcAB)
apcC F. diplosiphon ApcC; associated with the core (L78); expressed in white 28light with allophycocyanin (apcAIBl)
nine codon. In the -275 bp 3' of cpeD, there are no openreading frames encoding peptides longer than 27 aminoacids. However, there are several small repeated sequencesin this 3' region; for example, TTACGAA is directly re-
peated four times (Fig. 3) and TAATTCGT is repeatedtwice. These sequences may form secondary structures,which could be involved in determining the ratio of thedifferent transcripts in this region of the operon (see below).
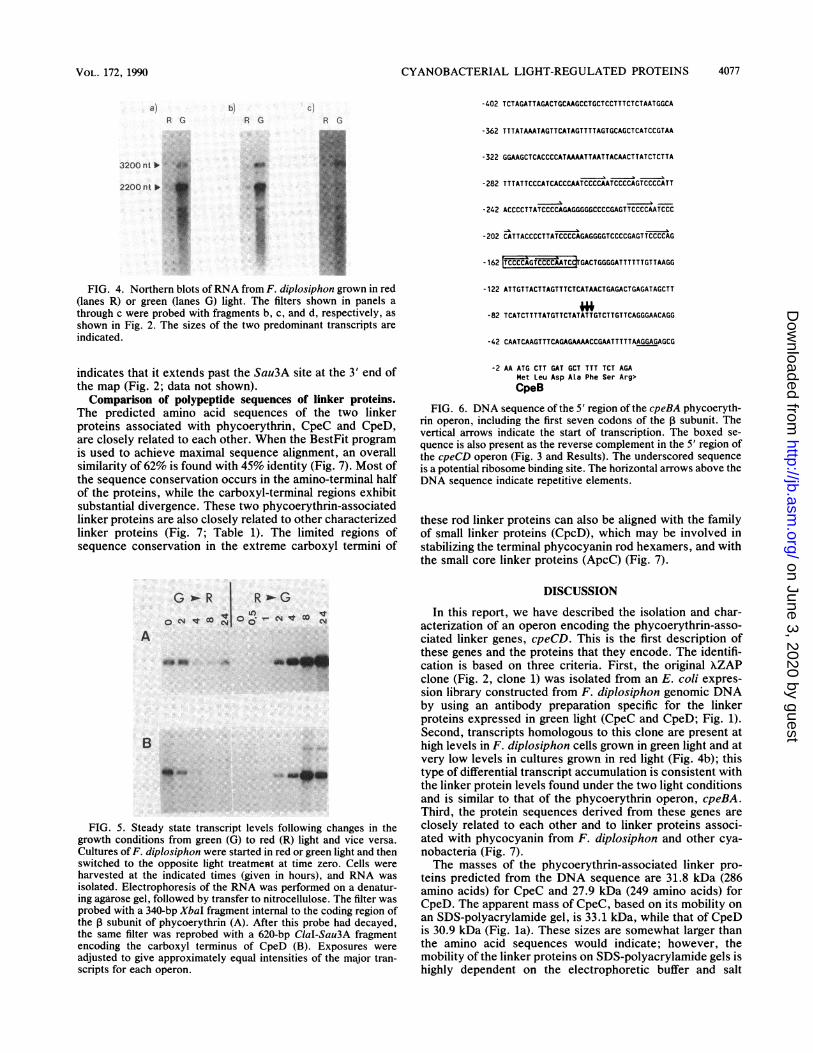
Transcription of the cpeCD operon. Restriction fragmentswithin and flanking the cpeCD genes were used to probeNorthern blots (RNA blots) ofRNA isolated from F. diplosi-phon grown in red or green light. In green light, the cpeCDoperon expresses a major 2,200-nt transcript and a lessabundant 3,200-nt transcript, while in red light, very lowlevels of both transcripts can be detected (Fig. 4). The2,200-nt transcript is large enough to encode the two linkerproteins, while the 3,200-nt transcript extends an additional-1 kb in the 3' direction (see below). A probe 5' to thepredicted start of transcription (fragment a in Fig. 2) does nothybridize to any transcripts (data not shown), while probesfrom within the coding regions of cpeC and cpeD (fragmentsb and c in Fig. 2) hybridize to both the 2,200- and 3,200-nttranscripts (Fig. 4a and b). A probe from the region down-stream of cpeD (fragment d in Fig. 2) hybridizes to the3,200-nt transcript and to several smaller transcripts whichmay be processing intermediates or degradation products orboth (Fig. 4c). In addition, on long exposures, a largertranscript of -3,800 nt can be detected with both codingregion and 3' probes. A smaller band (-1,500 nt) may be anartifact on the basis of its position immediately below thesmall rRNA band.
Regulated expression of cpeBA and cpeCD by green light.Transcription of the cpeBA and cpeCD operons is coordi-nately regulated. When cultures of F. diplosiphon are grownin green light and then transferred to red light, the levels oftranscripts encoding phycoerythrin and the phycoerythrin-associated linkers decline to low levels within 4 h (Fig. 5);however, the phycoerythrin transcripts are maintained athigh levels somewhat longer than are the transcripts encod-ing the linker proteins. Conversely, when red light-growncultures are transferred to green light, the initially low levelsof cpeBA and cpeCD transcripts increase rapidly, reaching a
maximum between 4 and 8 h. Both cpeCD transcriptsaccumulate at similar rates, although the steady state level ofthe 3,200-nt cpeCD transcript is -20% of the level of the2,200-nt transcript in green light (as determined by densito-metry); the level of the 3,200-nt transcript is too low in redlight to quantitate (Fig. 4 and Sb).
By using probes specific for the cpeCD operon, primerextension and Si nuclease protection analyses ofRNA fromF. diplosiphon predicted a single transcription start site 187bases upstream of the presumed AUG initiation codon ofcpeC (data not shown); the start site is indicated in Fig. 3.The primer extension analysis established this same tran-scription start site in both green and red light. Since only onetranscription start site was observed, both the 2,200- and3,200-nt transcripts must have the same 5' end and differ intheir 3' ends. The locations of the two transcripts relative tothe cpeCD coding regions are shown in Fig. 2. The longleader sequence of the transcripts for the linker genescontains several small (14 to 21 amino acids) overlappingopen reading frames that have potential ribosome bindingsites and that start with methionine. At this time, theregulatory significance of this observation is unknown.
Although the DNA sequence of the coding region of thephycoerythrin operon (cpeBA) has been previously reported(33), only a small region of DNA sequence 5' of the codingregion has been published (25). The sequence of a 423-bpXbaI fragment containing the 5' region of cpeBA includingthe first seven codons is shown in Fig. 6. The start oftranscription of this operon was determined by S1 nucleaseprotection to be 62 to 64 bases 5' of the translation initiationcodon, as indicated in Fig. 6 (data not shown).Having defined the transcription start sites for the cpeBA
and cpeCD operons, we examined the upstream regions forDNA sequence homologies that might be involved in theircoordinate regulation. A 17-bp element is found 83 bp 5' ofthe transcription start site for the cpeBA operon (5'-TCCCCAGTCCCCAATCC) and is also found as the reversecomplement 195 bp upstream of the transcription start sitefor the cpeCD operon (5'-GGATTGGGGACTGGGGA)(Fig. 3 and 6). This element is located in both promoterswithin stretches ofDNA which contain multiple small repet-itive sequences. We are currently investigating the signifi-cance of this sequence motif in the light-regulated expressionof these operons.As described above, transcription of the cpeCD operon
results in the accumulation of two major transcripts with thesame 5' end but with different lengths of 3' sequences. It isnot known whether transcriptional termination or RNAprocessing is responsible for this differential transcript accu-mulation. However, examination of the sequence 3' of thetranslation stop codon for CpeD revealed a potential RNAsecondary structure with a AGO of -101.4 kcal/mol (1 cal =4.184 J). The precise endpoint of the 2,200-nt transcript hasnot yet been mapped, but S1 nuclease protection analysis
VOL. 172, 1990
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

4076 FEDERSPIEL AND GROSSMAN J. BACTERIOL.
- 453 GATcrTGCTCAAGCAGTAGATTrTArTTAArTCAAAGCAGGGGCGACGGAGT GAAGTGGGATTGGGGA GGl GAGGAGGATGAGGGGGATGAGGAGGATGAGGGGGATGAGGGGAAAT AA
-332 CCATTACCCATTCCCCATTCCCCATTACCCAATTTCCCATGCCCATTACAAATAGTTTGTGCMAAATGAGTGCAAAATCTCTCATTTCATCAAGATTTACAAATCTTGATGTACATCrTTT
- 21 1 CCGCGCTCATAAGAAAATGArTCATACAAAGCGTATAAMACGCCTAAAACCTTrGAGcArTCTAGGAAATCACAGCTTrTrCATGCTTrATGGAAGCTGGTAATTTGGGTTrGCMAACAAACAAT CC
-90 AAAGAATTGATGAGCCTGGATGAGCAATCTGAAATTTATTTACAAATCGAAACAAATTTCTTAAACTTTCGTTAACAGGAGAAACAATTA ATG CCA TTT GGA CCA GCT TCAM P F G P A S>CpeC
22 CGC TTG GGA GTC AGC CTA TTT GAT GAA ACT CCT CCC GTT GAG TGG GTA CCA GGT CGC TCA CAA GAA GAA GCA GAA ACA ATC ATT CGG GCAR L G V S L F D E T P P V E W V P G R S 0 E E A E T I I R A>
112 ATC TAT CGG CAA GTA TTA GGT AAT GCC TAT GTG ATG GAA AGT GAG CGG CTT GCT GTG CCT GAA TCC CAG TTT AAG CGG GGT GAG TTG AGCI Y R Q V L G N A Y V M E S E R L A V P E S 0 F K R G E L S>
202 GTC CGC GAG TTT GTC AGA GCA GTG GCT AAA TCT GAA CTA TAT CGT TCT CGC TTT TTC ACC AGT TGT GCG CGC TAC CGA GCC ATT GAM CTCV R E F V R A V A K S E L Y R S R F F T S C A R Y R A I E L>
292 AAC TTC CGC CAT CTA TTG GGT CGT CCA CCA CTA GAT TTA G GMA ATG CGC TCC CAC AGC ACA ATC CTT GAT ACT CAA GGG TTT GAA GCTN F R H L L G R P P L D L E E M R S H S T I L D T 0 G F E A>
382 GAG ATT GAT TCT TAT ATC GAY GGT GAT GAG TAT CAG TCT ACT TTT GGC GAG AAC ATT GTA CCT TAC ATC CGA GGC TAT AAA ACC GM GCGE I D S Y I D G D E Y 0 S T F G E N I V P Y I R G Y K T E A>
472 CTT CAG AGC ATG GYG CAA TTT ACT CAT ACC TTC CM CTG GTA CGA GGT GCT TCT AGC AGC AGC CTG MG GGT GAC TTA TCT GGC AAG GCTL Q S M V 0 F T H T F 0 L V R G A S S S S L K G D L S G K A>
562 CCT AAG CTG AAT GCA TTA GTG ATT CAA AGC ACA CCA ACA GCA GTA ATT TCA CCT GCT AGC GCT GGA GCA ACC TTC TCG ACA CCA CCT ACTP K L N A L V I 0 S T P T A V I S P A S A G A T F S T P P T>
652 GGT GCC CGT ACC CGT CTT GGA GYC GAY GCT AGT GCT GGT GGC AAA GTT TAC CGC ATT GMA GrT ACA GGT TAT CGT GCC AAA ACC TTC AATG A R T R L G V D A S A G G K V Y R I E V T G Y R A K T F N>
742 AAT ATY TCC AAG TYT CGC CGT TCC AAT CAA GTC TTT CTG GTG CCC TAC GAA AAG CTC TCT CAA GAG TAT CAA CGG ATY CAC CAG CM GGCN I S K F R R S N 0 V F L V P Y E K L S 0 E Y 0 R I H 0 0 G>
832 GGC GTG ATC GCA AGT ATC ACT CCT GTA TAAATTAGGTGCAMCTTAAMAATTGAGGAGCAGAAATTTTA ATG GCA TCC CAG ACA ATT CTT GAA CYT TGGG V I A S I T P V> M A S 0 T I L E L W>
Cpe D932 CCC TCT AGT AGC TTA GAA GAA GTT CAA ACT ATT ATC CGT GCA GYT TAC AAA CAG GTT TTA GGC MC CCT CAT GYT ATG GAG AGY GAG CGG
P S S S L E E 0 T I I R A V Y K 0 V L G N P H V M E S E R>
1022 TTG GTG ACA GCA GAA TCA CAA rTA TGC GAT CGC TCC ATC ACC GTG CGG GM TYT GYC CGC AGC GrT GCC AAG TCT GAY TTT TAT CGC MCL V T A E S Q L C D R S I T V R E F V R S V A K S D F Y R N>
1112 CGC TAC TTC CAA TCC TGC GCT CCC TAC CGA TTT GTA GAA CTT AAC TTC TTA CAT TTG CTT GGT CGC GCA CCC CAG GAY CAA AGA GAA GTTR Y F S C A P Y R F V E L N F L H L L G R A P D 0 R E V>
1202 TCC GAA CAC AYC GTT CGT ACT GTA GCT GAA GGC YAC GAY GCT GM ATT GAC TCC TAT ATC GAY AGT AGT GAA TAT GMA GCA GCC TYT GGTS E H I V R T V A E G Y D A E I D S Y I D S S E Y E A A F G>
1292 GM AAC GTA GTG CCT TAC TAT CGT GGT AGA AGT AGC GMA GCC MC TCC MG CM GTA GGC TTC MC CGC ATA TTT GCC CTT GAT CGC GGCE N V V P Y Y R G R S S E A N S K 0 V G F N R I F A L D R G>
1382 CCT GCC CAA ATY GAY AGY GCA GrT AAA TCG GCT CM TTG GTC TAT GCT GrT GCT ACT MC AGC GCC AAC GCG ATC MA GCC TCT TCA TCCP A 0 1 D S A V K S A 0 L V Y A V A T N S A N A I K A S S S>
1472 ACA GYC ATT GGC TCT GGA ACT GAA AAA CGA TTC AAA ATC TYG GTG CAA GGT TCC AAA TTC GAC AGT CCC CGA CGC ATC AGT ACC ACT GAGT V I G S G T E K R F K I L V 0 G S K F D S P R R I S T T E>
1562 TAC ATT GrT CCA GCT AGT AAG ATG ACT CCC CM ATT CAG CGG ATT MT CGT ACT TCT GGC AAA ATC GTC AGC ATY ACT GAA ATT GTC TAACCY I V P A S K M T P 0 I 0 R I N R T S G K I V S I T E I V>
1654 TTTAACAGGGTGGGCATTrAATAcTTrGTCTGTTrCTAAATGrTTGAACCGATTAGTATTrAAcAAcrTCGTAATTcGTAGrTTAcTAATTrcGTAArTrAcGrTTACGGATAGGGATTTAGcT CCCACCT
1775 GTAAcGGcArTGArTrTrTATAGAAGrTCGGAGACTCAAAcCCCTCAGCTTrAArTTACGAATTrAcGAArTrAcGAATTAcGAATTAGTAATTTAGGTTrGAGACATTrGCCCGCCCCAGATCATTrCACrTT
1896 TTTTAATTTGTAATTTTCMAAA
FIG. 3. DNA sequence of the cpeCD operon encoding the phycoerythrin-associated linker proteins, beginning with the second Sau3A siteshown in the restriction map in Fig. 2. The boxed 17-bp sequence is found as the reverse complement in the promoter of the cpeBA(phycoerythrin) operon (Fig. 6). The underscored sequences are potential ribosome binding sites. The horizontal arrows above the DNAsequence indicate repetitive sequences. The arrowhead indicates the nucleotide at which transcription starts. The predicted amino acidsequence for the two linker proteins is shown beneath the DNA sequence. The GenBank accession number is M33832.
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from
CYANOBACTERIAL LIGHT-REGULATED PROTEINS 4077
a)R G
3200 nt
.~2200 nt lo .::;
bJR G
c,
R G
so
_I
FIG. 4. Northern blots ofRNA from F. diplosiphon grown in red(lanes R) or green (lanes G) light. The filters shown in panels athrough c were probed with fragments b, c, and d, respectively, asshown in Fig. 2. The sizes of the two predominant transcripts areindicated.
indicates that it extends past the Sau3A site at the 3' end ofthe map (Fig. 2; data not shown).Comparison of polypeptide sequences of linker proteins.
The predicted amino acid sequences of the two linkerproteins associated with phycoerythrin, CpeC and CpeD,are closely related to each other. When the BestFit programis used to achieve maximal sequence alignment, an overallsimilarity of 62% is found with 45% identity (Fig. 7). Most ofthe sequence conservation occurs in the amino-terminal halfof the proteins, while the carboxyl-terminal regions exhibitsubstantial divergence. These two phycoerythrin-associatedlinker proteins are also closely related to other characterizedlinker proteins (Fig. 7; Table 1). The limited regions ofsequence conservation in the extreme carboxyl termini of
G R|-GR
Oc NtcoN
A
0.4
FIG. 5. Steady state transcript levels following changes in thegrowth conditions from green (G) to red (R) light and vice versa.
Cultures of F. diplosiphon were started in red or green light and thenswitched to the opposite light treatment at time zero. Cells wereharvested at the indicated times (given in hours), and RNA was
isolated. Electrophoresis of the RNA was performed on a denatur-ing agarose gel, followed by transfer to nitrocellulose. The filter wasprobed with a 340-bp XbaI fragment internal to the coding region ofthe subunit of phycoerythrin (A). After this probe had decayed,the same filter was reprobed with a 620-bp ClaI-Sau3A fragmentencoding the carboxyl terminus of CpeD (B). Exposures were
adjusted to give approximately equal intensities of the major tran-scripts for each operon.
-402 TCTAGATTAGACTGCAAGCCTGCTCCTTTCTCTAATGGCA
-362 TTTATAAATAGTTCATAGTTTTAGTGCAGCTCATCCGTAA
-322 GGAAGCTCACCCCATAAAATTAATTACAACTTATCTCTTA
-282 TTTATTCCCATCACCCAATCCCCAATCCCCAGTCCCCATT
-242 ACCCCTTATCCCCAGAGGGGGCCCCGAGTTCCCCAATCCC
-202 CATTACCCCTTATCCCCAGAGGGGTCCCCGAGTTCCCCAG
-162 ITCCCCAGTCCCCAATCdrGACTGGGGATTTTTTGTTAAGG
-122 ATTGTTACTTAGTTTCTCATAACTGAGACTGAGATAGCTT
-82 TCATCTTTTATGTTCTATATTGTCTTGTTCAGGGAACAGG
-42 CAATCAAGTTTCAGAGAAAACCGAATTTTTAAGGAGAGCG
-2 AA ATG CTT GAT GCT TTT TCT AGAMet Leu Asp Ala Phe Ser Arg>CpeB
FIG. 6. DNA sequence of the 5' region of the cpeBA phycoeryth-rin operon, including the first seven codons of the 1 subunit. Thevertical arrows indicate the start of transcription. The boxed se-quence is also present as the reverse complement in the 5' region ofthe cpeCD operon (Fig. 3 and Results). The underscored sequenceis a potential ribosome binding site. The horizontal arrows above theDNA sequence indicate repetitive elements.
these rod linker proteins can also be aligned with the familyof small linker proteins (CpcD), which may be involved instabilizing the terminal phycocyanin rod hexamers, and withthe small core linker proteins (ApcC) (Fig. 7).
DISCUSSION
In this report, we have described the isolation and char-acterization of an operon encoding the phycoerythrin-asso-ciated linker genes, cpeCD. This is the first description ofthese genes and the proteins that they encode. The identifi-cation is based on three criteria. First, the original XZAPclone (Fig. 2, clone 1) was isolated from an E. coli expres-sion library constructed from F. diplosiphon genomic DNAby using an antibody preparation specific for the linkerproteins expressed in green light (CpeC and CpeD; Fig. 1).Second, transcripts homologous to this clone are present athigh levels in F. diplosiphon cells grown in green light and atvery low levels in cultures grown in red light (Fig. 4b); thistype of differential transcript accumulation is consistent withthe linker protein levels found under the two light conditionsand is similar to that of the phycoerythrin operon, cpeBA.Third, the protein sequences derived from these genes areclosely related to each other and to linker proteins associ-ated with phycocyanin from F. diplosiphon and other cya-nobacteria (Fig. 7).The masses of the phycoerythrin-associated linker pro-
teins predicted from the DNA sequence are 31.8 kDa (286amino acids) for CpeC and 27.9 kDa (249 amino acids) forCpeD. The apparent mass of CpeC, based on its mobility onan SDS-polyacrylamide gel, is 33.1 kDa, while that of CpeDis 30.9 kDa (Fig. la). These sizes are somewhat larger thanthe amino acid sequences would indicate; however, themobility of the linker proteins on SDS-polyacrylamide gels ishighly dependent on the electrophoretic buffer and salt
VOL. 172, 1990
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

4078 FEDERSPIEL AND GROSSMAN
1 20 40 60 80
CpeC (Fd) MPFG SR SLFDETPPVEWVPGRSQEEAETIIRA YRQVLG YVMER VPESQFKRGFLSVREFVRAVAKSELCpeD (Fd) M--- Sq-T-------Til-ElwPssSlEEvqT IIRAv G hVMEER taESQlcdrsitVREFVRsVAKSdfCpcI2 (Fd) pits SRLGtay-qTnPiElrPnwtaEdAkivIqAvRQVLG YiM ER slESlltnGLSVRdFVRAVAKSELCpcI3 (Fd) mpitt SRLGtSaFsnaaPiElrsntnkaEiaqvIaA RQVLG YVi ER glESlItnGritVqEFVRqlAKSnLCpcH2 (Fd) mtsst rqLGfepFasTaPtE-1rasS--dvpavIhAaQ hVM ER saErllqqGniSVRdFVR11AqSELCpcH3 (Fd) maPlt VSRrpFadsdkVElrfvktaEEvrsvIws RQVLG hlf ER saESllqqa iSVRdFVRAiAqSELCpcC (An) maitt tSRepFsdaPkVElrPkaSrEEvEsvIRA RhVLG Yil ER saESllrdGr#LtVREFVRsVAKSELCpcC (Sy) mpvtv SRLGaaFDq-sPVElranySrddAqTvIRA v YVM ER aaESlFtnGfSVRdFVRAVAqSEL
100 120 140 160
CpeC (Fd) Y SCARYRAIELNFRHLL4GRPPL EEMRSHSTILDT FEAEIDSYI 3EYQSTFGENIVPYIRGYKTEA-LQSCpeD (Fd) Y y SCApYRfvELNFLHLLGRaPq rEvseHivrtvaelydAEIDSYIDslsEYeaaFGENvVPYyRGrssEA-nskCpcI2 (Fd) Y Fl phfqtRvIELNFkHLLGRaPy sEvieHldryqnQ FdAdIDSYIDsiaEYdtyFGdsIVPYyRdlvTtgvgQrCpcI3 (Fd) Y F nnfhsRvtIELNFkHLLGRaPy esEiiyHldlyqT GyEAdIDSYIDsaEYQtnFadNIVPYyRGfnnql-gQkCpcH2 (Fd) Y F StpqrRfI|ELNykHLLGRaPy esEisyHvnlytek 3yEAEInSYIDsa iYQesFGErIVPhyR feTqp-gQkCpcH3 (Fd) Y F SnsqvRfIELNykHLLjGRaPy1sEiayHvdIyts1jyEAEInSYID EYQqnFGdsIVPYyRl qTtv-gQkCpcC (An) Y F nsfqtRlIIELNykHLLGRaPy sEvvyHldlyqnk3ydAEIDSYIDs EYQSnFGdNvVPYyR feTqv-gQkCpcC (Sy) Y Fl nnfqtRvIELNFkHLLCRaP aEvieHldryqne3FEAdInSYID EYtenGdNIVPYIR vvqt-ghr
CpeC (Fd)CpeD (Fd)CpcI2 (Fd)CpcI3 (Fd)CpcH2 (Fd)CpcH3 (Fd)CpcC (An)CpcC (Sy)PEC-L (Ma)CpcD (Ma)CpcD2 (Fd)CpcD3 (Fd)CpcD (An)CpcD (Sy)
CpeC (Fd)CpeD (Fd)CpcI2 (Fd)CpcI3 (Fd)CpcH2 (Fd)CpcH3 (Fd)CpcC (An)CpcC (Sy)PEC-L (Ma)CpcD (Ma)CpcD2 (Fd)CpcD3 (Fd)CpcD (An)CpcD (Sy)ApcC (Ma)ApcC (Sy)ApcC (Fd)
J. BACTERIOL.
180 200 220 240* * **
MVQFTHTFQLVRGASSSSLK(GDLSGiKA-PKLNA-LVIQSTPTAVISPAS--AGAT---FSTPP--TGARTR-LG-VDAS--qVgFnriFaLdRGpa----qidsavKs-aqLvy-aVatnsanAikassS-----------------------t-VigS--tVgFTrmFrLyRGyanSd-rsqLaGss-srLas-dlatnsaTAiIaPsg--gtqgwsylpskq--gtApsRtfGrssqg--tVgFTriFQLyRGyatSd-rsqipGas-arLan-elarnsastVIaPAg--snng---Fay-r--asvkgk-tp-stAf--tVgFnrmFQiyRGyanSd-rsq--GKn--K-sA-wltQdlaln-lasni--qtpn---Fg-----kGl-T---G-VvAg--tagFprfFQLyRGyantd-raqnksK--gq-lt-wdlaknlvspIyPAd--A-gs---1 tgvs--TGnRgg-nt-yri r--tagFnriFrLyRGyanSd-raqveGtksrigAgnLasnkasTiVg-Psg--tndswg-Frasad-vapkkn-LG-navg--tVgFTrmFsL.qRGyanSd-raqiaGnA-srLaq-elarnTtsAVvgPsgvneGwa---Frsaaddyhpgqs-LG-gstglstVgFnriFeLyRGranSd-naqfgGKs-arLrs-kismnlantivpPtS--piAa----ST----ssART--L--V-----
mfgqTt-LG-iDsvssmlgsvlTRrs----sSg-
mvyqsmfgqTt-LG-agsvss
mlsqfAgte
260
AGGKVYRIEVTGYRAKTFNNISKF-- VRRQV-FL-VAL_ __
_V
280
KI--LSQEr_
K HQI300
VIAS ITPV
stprlYRIEVTGislpry---pKv-- RRS ke-Fi- VPY q--LSst QqInkl GkvASITfaqqGsqafgsgrlyrvevaaisqpaiqvRRi Nkrsihr rti lfptSst Qwq---_--- IASvTPldrGqlYRvrVi--qAdr-grttqi- SiQe-yL-jVsY q--LSpt QR1nQrGsfrvvnIsPattqaaspnsprirqs-----IS--- R--vV---- VP q--LSnll qlnrQGrkviSIalseGdrVYRlEVTGiRspgypsv----- SRRstV-Fi- VPY r--LSdk qvHkQGGkIvSvTsaAddqVvRvEVaalstprypri -----vG--mf-I-Vea i-AgTlNtnvav--sasrVfRfEVvGmRqneeNdknKynisdnrVfvyEVeClRqneqtdnnryqirsfqVevsglhqnevtnqNN-ypi--sasrVfRyEVvGlRqssetdknKyniAasrVftyEVqGlqteetdNqeya f-
Grl fkItacvpsqtrirtqrel--mrmfRItaclpspskirtqrel--GrlfkItasvpsqtrirtqrel--
RRSRRSRRSRnSSs5
RnSRRSqntantqnt
srV-Ff -
rQV-yt-gsV-yitstieiq-gsV-FitgsV-FitgsV-Firyft-kl-fft-kl-yft-kl-
VPVP
t VE-Vit iptVFnVP
_-VP_-VP
'5
if
isis
piPNPNPfPNPN
sr--LSQkl2edr--LSatY Qe'nr--mSeE R:sr--mneE R'srf--neEl QR
ar--mnQE REnw-- frE RQdaw--frE R'Enw--frE R.'
qrnIkitrnrtr1rlqknqklakn
rvASIsPagqkIvklTPaskIvklePltraagIvnI rPagenptedasen
kIvnIqPlnlqinenklvkIeqlvsaeakIvSI kPytgatasdeekIvkvelatgkqg intglakI ik-elatgrpntntgl 1kIvk-elatgkqq intgla
286 aa249 aa288 aa285 aa269 aa271 aa287 aa290 aa115 aa80 aa85 aa70 aa80 aa79 aa67 aa67 aa67 aa
FIG. 7. Comparison of the amino acid sequences of linker proteins from F. diplosiphon and other cyanobacteria. Single-letter amino acidsymbols are used. Residues of linker proteins identical to those in CpeC are indicated by capital letters, nonidentical residues are indicatedby lowercase letters, and gaps are indicated by hyphens. Boxed regions indicate areas having a high degree of homology; asterisks indicateresidues which are identical among all the proteins. An arbitrary numbering system is used starting with the first amino acid of CpeC. PEC-L(Ma), CpcD (Ma), and ApcC (Ma) are the reported amino acid sequences of the carboxyl-terminal portion of the 34.5-kDa linker associatedwith phycoerythrocyanin, the 8.9-kDa rod/core linker, and the 8.9-kDa core linker, respectively, from M. laminosus (15). References and briefdescriptions of the other proteins are provided in Table 1. Abbreviations: Fd, F. diplosiphon; An, Anabaena strain 7120; Sy, Synechococcusstrains; Ma, M. laminosus.
concentrations. For example, the apparent masses of thephycocyanin-associated linker proteins CpcH2 and CpcI2range from 33.3 and 32.0 kDa, respectively (Fig. la), to 39.0and 37.5 kDa on a different gel system (32); the predictedmasses of these two proteins based on the DNA sequenceare 32.5 and 30.5 kDa (32). This behavior may be due to thebasic nature of these proteins, resulting in anomalous SDSbinding. In addition, phycocyanin-associated linker proteinsfrom Anacystis nidulans have been reported to containcovalently linked, glucose-containing polysaccharides (39,40). By analogy, the linker proteins from F. diplosiphon,which are associated with both inducible phycocyanin andphycoerythrin, may also be glycosylated, consistent withtheir larger apparent molecular masses on SDS-polyacryl-amide gels than were predicted from their amino acid com-positions.The structural roles of the individual linker proteins CpeC
and CpeD in the assembly of phycobilisomes in F. diplosi-phon are still undefined. However, reconstitution studieswith isolated phycoerythrin subunits and the two associatedlinker proteins from Synechocystis strain 6701 (19, 20) have
shown that the larger linker protein (31.5 kDa) associateswith phycoerythrin hexamers internal to the phycobilisomerods, while the smaller linker protein (30.5 kDa) is foundonly in the phycoerythrin hexamers most distal to the core.If the relative migration of the phycoerythrin-associatedlinker proteins of F. diplosiphon on SDS-polyacrylamidegels correlates with their predicted sizes based on amino acidsequence (that is, if the mobilities of CpeC and CpeD are notreversed on gels as a result of more extensive glycosylationof the smaller protein), then the larger linker protein (CpeC)would correspond to the internal rod linker, while thesmaller linker protein (CpeD) would be the linker associatedwith phycoerythrin hexamers at the ends of the rods in F.diplosiphon.When the predicted amino acid sequences of CpeC and
CpeD are compared with each other and with the sequencesof previously characterized rod linker proteins, 49 aminoacid residues (-17 to 20%) are invariant among all eight rodlinker proteins, with 115 invariant amino acids betweenCpeC and CpeD (45%). While there is very little similarity atthe amino-terminal end of the proteins, except for a cluster
tsG
rGG
1 GG
1GmG
!I.ltititi
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

CYANOBACTERIAL LIGHT-REGULATED PROTEINS 4079
of five amino acids (residues 6 through 10 in CpeC), a largeblock of conserved domains starts near residue 35 for mostof the linker proteins and continues approximately throughresidue 175 (Fig. 7). Analysis of the amino acid distributionwithin the conserved regions does not provide any obviousinsights into possible relationships between structure andfunction. For CpeC and CpeD, these regions exhibit aslightly lower percentage of polar amino acids and a slightlyhigher ratio of acidic amino acids than is represented in eachprotein as a whole, while the content of basic and nonpolaramino acids in these regions closely reflects their overallcomposition in each protein. The carboxyl-terminal regionsof the rod linker proteins are much more divergent, withmany more deletions, insertions, and substitutions than arefound in the amino-terminal domains. Interestingly, it isthese less conserved carboxyl-terminal regions that havelimited homology to the family of small linker proteins suchas CpcD2 and CpcD3 (Fig. 7), which have been proposed tofunction in the formation and assembly of the terminalphycocyanin rod hexamers (10). This similarity had beenpreviously described between the L89 linker protein fromM. laminosus and the phycoerythrocyanin-associated linkerL34 5 (CpcD and PEC-L in Fig. 7, respectively); additionally,another 8.9-kDa protein (ApcC in Fig. 7) was describedwhich is believed to be associated with the phycobilisomecore (16). The genes encoding ApcC have recently beencloned from Synechococcus strain 6301 and F. diplosiphon(28, 29). The degree of sequence conservation among linkerproteins which have different modes of regulation, whichassociate with different phycobiliproteins, and which derivefrom different organisms, implicates the conserved regionsin important functional or structural interactions or both.
Schirmer et al. (42, 43) have determined the crystalstructure of phycocyanin hexamers from Agmenellum qua-druplicatum and phycocyanin trimers from M. laminosus inthe absence of linker proteins. The phycocyanin complexesare arranged in a toroidal structure with an internal cavitywith a maximum diameter of -4 nm (43). The crystalstructure of the phycocyanin hexamers predicts that somenegatively charged residues protrude into the internal chan-nel; these residues may be involved in specific interactionswith the basic linker proteins. Evidence regarding the site ofbinding of the linker proteins to the hexamers was providedby studies of the susceptibility of Anabaena variabilis phy-cocyanin-linker protein aggregates to proteolysis (55). Thoseauthors showed that the 32.5-kDa linker protein was di-gested to a 28-kDa polypeptide when present in phycocyanintrimers or hexamers but was completely protected fromdegradation in higher order aggregates. This implies that a28-kDa domain of the linker protein may be bound within theinternal cavity of the phycocyanin hexamers, while the4.5-kDa domain is involved in the stacking interactionsbetween hexamers. Another interesting point derived fromthe phycocyanin structure is that there is remarkable simi-larity in three-dimensional structure between the a and ,Bsubunits, even though only 46 residues are conserved (28and 27%, respectively) (42). This characteristic of conserva-tion of structure in spite of sequence divergence may also bea feature of the linker proteins associated with both phyco-cyanin and phycoerythrin because of the constraints im-posed by the functional assembly of the phycobilisome.We have shown that the cpeCD operon is expressed on
two major transcripts which have the same 5' ends but whichdiffer in the 3' ends. The smaller, 2,200-nt transcript is ofsufficient length to encode both CpeC and CpeD, while thelarger, 3,200-nt transcript extends another -1 kb in the 3'
direction. These transcripts are abundant in F. diplosiphoncells grown in green light but are barely detectable incultures grown in red light, which correlates with the ob-served levels of the phycoerythrin-associated linker proteinsin these cultures. The precise 3' end of the 2,200-nt tran-script has not yet been determined; however, it probably lies3' of the very stable hairpin which is predicted to form justdownstream of the coding region of CpeD. This potentialstructure is followed by a string of uridine residues, consti-tuting a classic procaryotic simple terminator as defined in E.coli (37). Similarly, a smaller hairpin is predicted to occur inthe cpc2 operon of F. diplosiphon, between the genesencoding the ,B and a subunits of the inducible phycocyanin(cpcB2A2) and the genes encoding the associated linkerproteins (cpcH2I2D2) (14). If the predicted secondary struc-ture in the cpeCD operon acts as a simple terminator, theratio of the 2,200- and 3,200-nt transcripts could be deter-mined primarily by the percentage of transcriptionalreadthrough past the hairpin. In addition, this structurecould act as a barrier to degradation from the 3' end of theoperon-length transcript. Such segmental stability of tran-scripts has been observed for the polycistronic pufoperon inRhodobacter capsulatus (4, 11). At this point, we have noevidence regarding these alternative mechanisms (which arenot mutually exclusive) of regulating the levels of the twomajor cpeCD transcripts. We are currently continuing thesequence analysis of the region 3' of cpeD to define anyadditional genes which might be encoded in this operon. Noother proteins have yet been identified in F. diplosiphonwhich are known to be expressed preferentially in greenlight, but possibilities include other phycobilisome compo-nents or factors required for assembly, chromophorylationenzymes involved in phycobiliprotein modification, determi-nants of morphological changes mediated by green light, orregulatory factors involved in the signal transduction path-way.
Unlike the red light-inducible phycocyanin subunits andassociated linker proteins (cpc2) (14, 32), the linker proteinsassociated with phycoerythrin are not closely linked in theF. diplosiphon genome to the phycoerythrin operon(cpeBA). This makes their coordinate light-regulated expres-sion more complex. While the ratio of the phycocyanin-associated linker proteins to phycocyanin subunits is appar-ently maintained by lower levels of full-length transcriptsrelative to transcripts encoding only the phycocyanin sub-units (as the result of transcriptional termination,readthrough, and/or processing, as discussed above), regu-lation of the ratio of phycoerythrin-associated linker proteinsto phycoerythrin subunits must be achieved by independenttranscriptional events. We have determined the transcrip-tional start sites for both green light-activated operons;transcription initiates 62 to 64 bp upstream of the first AUGin the phycoerythrin operon cpeBA, while the start of thecpeCD transcripts encoding the linker proteins is 187 bp 5' ofthe putative translation start site for CpeC. The short leadersequence of the cpeBA transcript contains no open readingframes prior to the start of the CpeB coding region; incontrast, the long leader sequence of the cpeCD transcriptscontains several small (14 to 21 amino acids) overlappingopen reading frames with potential ribosome binding sitesupstream of the AUG initiating translation of CpeC. Wehave no evidence at this time suggesting whether or not theshort peptides are synthesized and involved in translationalcontrol of this operon.The steady state level of the cpeBA transcript appears to
be approximately 5- to 10-fold higher than that of the
VOL. 172, 1990
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

4080 FEDERSPIEL AND GROSSMAN
2,200-nt cpeCD transcript, on the basis of Northern hybrid-izations with probes of similar specific activity; however,since these transcripts do not share any common sequences(unlike the transcripts of the inducible phycocyanin operonand its linker proteins), it is difficult to make very accurateestimates of relative transcript abundance. The kinetics oftranscript accumulation in response to changes in lightquality are very similar between the two operons (Fig. 5aand b), implying that similar features in the 5' regulatoryregions of these genes allow for their coordinate regulation.However, there is no obvious homology in the regionsimmediately upstream of the predicted transcription startsites for these two operons. In contrast, there is a highdegree of sequence conservation in the regions immediately5' of the transcription start sites for the cpeBA operon in F.diplosiphon and in Synechocystis strain 6701 (1). Thus, incomparing these green light-regulated operons, it is stillunclear what constitutes a basic promoter recognition se-quence.The sequences upstream of the transcription start sites for
the cpeBA and cpeCD operons in F. diplosiphon contain ahigh number of repetitive elements. For example, in the 5'region of cpeCD, the sequence GGATGAGG is found fivetimes in a much larger region of high strand asymmetrycontaining predominantly guanosine residues (49 of 80 =61% G versus 3 of 80 = 4% C, from -339 to -418 bp relativeto the translation start site) (Fig. 3). This is followed by aregion containing a high ratio of thymidine and cytosineresidues, including elements such as the palindrome CCATTACC, which is repeated directly and in slightly variantforms several times. Numerous other elements are found asdirect or inverted repeats and as regions of dyad symmetry.Similarly, numerous repetitive elements can be found in the5' region of cpeBA, such as the sequence TCCCCA repeated10 times (Fig. 6). Both promoter regions possess significantdyad symmetry over several hundred base pairs, and thepossibility exists that cruciform DNA structures could play arole in the regulation of these operons (27). Embedded inthese repetitive structures (which in general are not the samebetween the two operons) is one 17-bp element which iscommon to both operons, TCCCCAGTCCCCAATCC. Thismotif is found 83 bp 5' of the transcription start site for thecpeBA operon and is found as the reverse complement 195bp upstream of the transcription start site for cpeCD. Thiselement is contained within the cpeBA promoter region in a190-bp fragment which binds to a factor in extracts from F.diplosiphon cultures grown in green light, but not in extractsfrom cultures grown in red light, as determined by DNArmobility shift gel assays (N. A. Federspiel, unpublishedresults). If this element is important in green light activationof transcription, the lower steady state levels of transcriptsfrom cpeCD may be due to the distance of this element fromthe transcription start site, its orientation, or the sequencesflanking the element. However, this element is not found inthe -450 bp which have been reported 5' of the cpeBAoperon in Synechocystis strain 6701 (1). Further investiga-tions of the specific DNA sequences and protein factorsinvolved in the transcriptional activation of these operons bygreen light will lead to a fuller understanding of the signaltransduction pathway between perception of light qualityand changes in gene expression in cyanobacteria.
ACKNOWLEDGMENTS
We are grateful to Lamont Anderson for valuable discussionsduring the course of this work, to LuAnn Scott for excellent
technical assistance, and to Lamont Anderson, Robert Fisher,Cindy Orser, and Scott Minnich for comments on the manuscript.
This work was supported by grant no. DCB8615606 from theNational Science Foundation (Carnegie Institute of Washington)and by BRSG grant no. 2507RR07170-13 from the National Insti-tutes of Health (University of Idaho).
LITERATURE CITED1. Anderson, L. K., and A. R. Grossman. 1990. Structure and
light-regulated expression of phycoerythrin genes in wild-typeand phycobilisome assembly mutants of Synechocystis sp.strain PCC 6701. J. Bacteriol. 172:1297-1305.
2. Anderson, L. K., M. C. Rayner, R. M. Sweet, and F. A.Eiserling. 1983. Regulation of Nostoc sp. phycobilisome struc-ture by light and temperature. J. Bacteriol. 155:1407-1416.
3. Ausubel, F. M., R. Brent, R. E. Kingson, D. D. Moore, J. G.Seidman, J. A. Smith, and K. Struhl (ed.). 1989. Currentprotocols in molecular biology, p. 10.8.1-10.8.6. John Wiley &Sons, Inc., New York.
4. Belasco, J. G., J. T. Beatty, C. W. Adams, A. vonGabain, andS. N. Cohen. 1985. Differential expression of photosynthesisgenes in R. capsulata results from segmental differences instability within the polycistronic rxcA transcript. Cell 40:171-181.
5. Belknap, W. R., and R. Haselkom. 1987. Cloning and lightregulation of expression of the phycocyanin operon of thecyanobacterium Anabaena. EMBO J. 6:871-884.
6. Bennett, A., and L. Bogorad. 1973. Complementary chromaticadaptation in a filamentous blue-green alga. J. Cell Biol. 58:419-435.
7. Bogorad, L. 1975. Phycobiliproteins and complementary chro-matic adaptation. Annu. Rev. Plant Physiol. 26:369-401.
8. Bruns, B. U., W. R. Briggs, and A. R. Grossman. 1989.Molecular characterization of phycobilisome regulatory mu-tants of Fremyella diplosiphon. J. Bacteriol. 171:901-908.
9. Bryant, D. A. 1981. The photoregulated expression of multiplephycocyanin species: general mechanism for the control ofphycocyanin synthesis in chromatically adapting cyanobacteria.Eur. J. Biochem. 119:425-429.
10. Bryant, D. A. 1988. Genetic analysis of phycobilisome biosyn-thesis, assembly, structure, and function in the cyanobacteriumSynechococcus sp. PCC 7002, p. 62-90. In S. E. Stevens, Jr.,and D. A. Bryant (ed.), Light-energy transduction in photosyn-thesis: higher plant and bacterial models. American Society ofPlant Physiologists, Rockvilie, Md.
11. Chen, C.-Y., J. T. Beatty, S. N. Cohen, and J. G. Belasco. 1988.An intercistronic stem-loop structure functions as an mRNAdecay terminator necessary but insufficient for puf mRNAstability. Cell 52:609-619.
12. Cohen-Bazire, G., and D. A. Bryant. 1982. Phycobilisomes:composition and structure, p. 143-189. In N. G. Carr and B.Whitton (ed.). The biology of cyanobacteria. University ofCalifornia Press, Berkeley, Calif.
13. Conley, P. B., P. G. Lemaux, and A. R. Grossman. 1985.Cyanobacterial light-harvesting complex subunits encoded intwo red light-induced transcripts. Science 230:550-553.
14. Conley, P. B., P. G. Lemaux, and A. R. Grossman. 1988.Molecular characterization and evolution of sequences encod-ing light-harvesting components in the chromatically adaptingcyanobacterium Fremyella diplosiphon. J. Mol. Biol. 199:447-465.
15. Feinberg, A. P., and B. Vogeldstein. 1983. A technique forradiolabeling DNA restriction endonuclease fragments to highspecific activity. Anal. Biochem. 132:6-13.
16. Fuglistafler, P., F. Suter, and H. Zuber. 1985. Linker polypep-tides of the phycobilisome from the cyanobacterium Mastigo-cladus laminosus: amino-acid sequences and relationships.Biol. Chem. Hoppe-Seyler 366:993-1001.
17. Gantt, E. 1981. Phycobilisomes. Annu. Rev. Plant Physiol.32:327-347.
18. Gendel, S., I. Ohad, and L. Bogorad. 1979. Control of phyco-erythrin synthesis during chromatic adaptation. Plant Physiol.64:786-790.
J. BACTERIOL.
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

CYANOBACTERIAL LIGHT-REGULATED PROTEINS 4081
19. Gingrich, J. C., L. K. Blaha, and A. N. Glazer. 1982. Rodsubstructure in cyanobacterial phycobilisomes: analysis of Syn-echocystis 6701 mutants low in phycoerythrin. J. Cell Biol.92:261-268.
20. Gingrich, J. C., R. C. Wifliams, and A. N. Glazer. 1982. Rodsubstructure in cyanobacterial phycobilisomes: phycoerythrinassembly in Synechocystis 6701 phycobilisomes. J. Cell Biol.95:170-178.
21. Glazer, A. N. 1982. Phycobilisomes: structure and dynamics.Annu. Rev. Microbiol. 36:173-198.
22. Glazer, A. N. 1985. Light harvesting by phycobilisomes. Annu.Rev. Biophys. Biophys. Chem. 14:47-77.
23. Gold, L. 1988. Posttranscriptional regulatory mechanisms inEscherichia coli. Annu. Rev. Biochem. 57:199-233.
24. Green, L. S., D. E. Laudenbach, and A. R. Grossman. 1989. Aregion of a cyanobacterial genome required for sulfate trans-port. Proc. Natl. Acad. Sci. USA 86:1949-1953.
25. Grossman, A. R., P. G. Lemaux, P. B. Conley, B. U. Bruns, andL. K. Anderson. 1988. Characterization of phycobiliprotein andlinker polypeptide genes in Fremyella diplosiphon and theirregulated expression during complementary chromatic adapta-tion. Photosynth. Res. 17:23-56.
26. Haury, J. F., and L. Bogorad. 1977. Action spectra for phyco-biliprotein synthesis in a chromatically adapting cyanophyte,Fremyella diplosiphon. Plant Physiol. 60:835-839.
27. Horwitz, M. S. Z., and L. A. Loeb. 1988. An E. coli promoterthat regulates transcription by DNA superhelix-induced cruci-form extrusion. Science 241:703-705.
28. Houmard, J., V. Capuano, T. Coursin, and N. Tandeau deMarsac. 1988. Genes encoding core components of the phyco-bilisome on the cyanobacterium Calothrix sp. strain PCC 7601:occurrence of a multigene family. J. Bacteriol. 170:5512-5521.
29. Houmard, J., D. Mazel, C. Moguet, D. A. Bryant, and N.Tandeau de Marsac. 1986. Organization and nucleotide se-quence of genes encoding core components of the phycobili-somes from Synechococcus 6301. Mol. Gen. Genet. 205:404-410.
30. Houmard, J., and N. Tandeau de Marsac. 1988. Cyanobacterialgenetic tools: current status. Methods Enzymol. 167:808-847.
31. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
32. Lomax, T. L., P. B. Conley, J. Schilling, and A. R. Grossman.1987. Isolation and characterization of light-regulated phycobil-isome linker polypeptide genes and their transcription as apolycistronic mRNA. J. Bacteriol. 169:2675-2684.
33. Mazel, D., G. Gugliehli, J. Houmard, W. Sidler, D. A. Bryant,and N. Tandeau de Marsac. 1986. Green light induces transcrip-tion of the phycoerythrin operon in the cyanobacterium Calo-thrix 7601. Nucleic Acids Res. 14:8279-8290.
34. Mazel, D., J. Houmard, and N. Tandeau de Marsac. 1988. Amultigene family in Calothrix sp. PCC 7601 encodes phycocya-nin, the major component of the cyanobacterial light-harvestingantenna. Mol. Gen. Genet. 211:296-304.
35. Mazel, D., and P. Marliere. 1989. Adaptive eradication ofmethionine and cysteine from cyanobacterial light-harvestingproteins. Nature (London) 341:245-248.
36. Oelmhiller, R., P. B. Conley, N. Federspiel, W. R. Briggs, andA. R. Grossman. 1988. Changes in accumulation and synthesisof transcripts encoding phycobilisome components during accli-mation of Fremyella diplosiphon to different light qualities.Plant Physiol. 88:1077-1083.
37. Platt, T. 1986. Transcription termination and the regulation ofgene expression. Annu. Rev. Biochem. 55:339-372.
38. Riethman, H., G. Bulierjahn, K. J. Reddy, and L. A. Sherman.1988. Regulation of cyanobacterial pigment-protein compositionand organization by environmental factors. Photosynth. Res.18:133-161.
39. Riethman, H. C., T. P. Mawhhney, and L. A. Sherman. 1987.Phycobilisome-associated glycoproteins in the cyanobacteriumAnacystis nidulans R2. FEBS Lett. 215:209-214.
40. Riethman, H. C., T. P. Mawhinney, and L. A. Sherman. 1988.Characterization of phycobilisome glycoproteins in the cyano-bacterium Anacystis nidulans R2. J. Bacteriol. 170:2433-2440.
41. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
42. Schirmer, T., W. Bode, and R. Huber. 1987. Refined three-dimensional structures oftwo cyanobacterial C-phycocyanins at2.1 and 2.5 A resolution. J. Mol. Biol. 196:677-695.
43. Schirmer, T., R. Huber, M. Schneider, and W. Bode. 1986.Crystal structure analysis and refinement at 2.5 A of hexamericC-phycocyanin from the cyanobacterium Agmenellum quadru-plicatum. J. Mol. Biol. 188:651-676.
44. Sharp, P. A., A. J. Berk, and S. M. Berget. 1980. Transcriptionmaps of adenovirus. Methods Enzymol. 65:750-768.
45. Shine, J., and L. Dalgarno. 1974. The 3'-terminal sequence ofEscherichia coli 16S ribosomal RNA: complementarity to non-sense triplets and ribosome binding sites. Proc. Natl. Acad. Sci.USA 71:1342-1346.
46. Short, J. M., J. M. Fernandez, J. A. Sorge, and W. D. Huse.1988. X ZAP: a bacteriophage A expression vector with in vivoexcision properties. Nucleic Acids Res. 16:7583-7600.
47. Tandeau de Marsac, N. 1977. Occurrence and nature of chro-matic adaptation in cyanobacteria. J. Bacteriol. 130:82-91.
48. Tandeau de Marsac, N., and G. Cohen-Bazire. 1977. Molecularcomposition of cyanobacterial phycobilisomes. Proc. Natl.Acad. Sci. USA 74:1635-1639.
49. Tandeau de Marsac, N., D. Mazel, T. Damerval, G. Guglielmi,V. Capuano, and J. Houmard. 1988. Photoregulation of geneexpression in the filamentous cyanobacterium Calothrix sp.PCC 7601: light-harvesting complexes and cell differentiation.Photosynth. Res. 18:99-132.
50. Thuring, R. W., J. B. Sanders, and P. A. Borst. 1975. Freeze-squeeze method for recovering long DNA from agarose gels.Anal. Biochem. 66:213-220.
51. Towbin, H., T. Staehelin, and J. Gordon. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulosesheets: procedure and some applications. Proc. Natl. Acad. Sci.USA 76:4350-4354.
52. Vieira, J., and J. Messing. 1987. Production of single-strandedplasmid DNA. Methods Enzymol. 153:3-11.
53. Vogelmann, T. C., and J. Scheibe. 1978. Action spectra forchromatic adaptation in the blue-green alga Fremyella diplosi-phon. Planta 143:233-239.
54. Williams, J. G., and P. J. Mason. 1985. Hybridisation in theanalysis of RNA, p. 139-160. In B. D. Hames and S. J. Higgins(ed.), Nucleic acid hybridisation: a practical approach. IRLPress, Washington, D.C.
55. Yu, M.-H., and A. N. Glazer. 1982. Cyanobacterial phycobili-somes: role of the linker polypeptides in the assembly ofphycocyanin. J. Biol. Chem. 257:3429-3433.
56. Zuber, H. 1986. Structure of light-harvesting antenna complexesof photosynthetic bacteria, cyanobacteria and red algae. TrendsBiochem. Sci. 11:414-419.
57. Zuker, M., and P. Stiegler. 1981. Optimal computer folding oflarge RNA sequences using thermodynamics and auxiliaryinformation. Nucleic Acids Res. 9:133-148.
VOL. 172, 1990
on June 3, 2020 by guesthttp://jb.asm
.org/D
ownloaded from