characterization of defensin precursors in mature human ...€¦ · hnp-2, and hnp-3) are highly...
TRANSCRIPT

Characterization of Defensin Precursors in Mature Human Neutrophils
By Sylvia S.L. Harwig, Alane S.K. Park, and Robert I. Lehrer
Human defensins HNP-1 and -3 are broad spectrum antimicro- bial peptides that are synthesized by human neutrophils as 94 amino acid (aa) precursors that require proteolytic re- moval of 64 amino-terminal residues to produce the mature defensins. Recent studies have shown that the early proteo- lytic processing events include two sequential cleavages, each removing 19 amino-terminal aa residues, that yield 75 aa and 56 aa prodefensins, respectively. The subsequent processing steps that convert these 56 aa prodefensins to mature (30 aa) HNP-1 and HNP-3 are not yet known. We identified four new defensin precursors in mature normal neutrophils. The most abundant of these were two 39 aa forms that resulted from the monobasic endoproteolytic
EFENSINS ARE SMALL, cysteine-rich antimicrobial D peptides that are abundant in human, rabbit, guinea pig, and rat neutrophils.'a2 Although attention has centered on their broad spectrum microbicidal and cytotoxic activity, some defensins are selective monocyte chemoattractants? and others cause nonspecific opsonization of bacteria: interference with ACTH-receptor-mediated functions,536 release of histamine from mast cells,' or potent inhibition of protein kinases.' Thus far, four human defensins have been described in human neutrophils. Three of these (HNP-1, HNP-2, and HNP-3) are highly homologous9 and constitute between 30% and 50% of the total protein in azurophil granules of human neutrophils." The fourth, HNP-4, has a distinctly different primary structure" and accounts for only 1% to 2% of the neutrophil's total defensins."
We began this project to see if any additional, quantita- tively minor defensins existed in mature human neutro- phils. Because HNP-4 and all of the currently known nonhuman defensins are more cationic than HNP-1, -2, or -3, we concentrated our search on the neutrophil's small cationic peptides, using acid-urea polyacrylamide gel elec- trophoresis (AU-PAGE) as a screening tool. This strategy led us to identify four "new" peptides, all of which proved to be incompletely processed forms of either HNP-1 or HNP-3. The newly characterized prodefensins provide
From the Department of Medicine, UCLA-Center for the Health Sciences; School of Medicine, University of Southem Califomia; and the Department of Medicine, Wadsworth Veterans Administration Hospital, Los Angeles, CA.
Submitted August 19, 1991; accepted November 6, 1991. Supported by Grant No. AI22839 from the National Institutes of
Health. Protein sequencing was performed at the UCLA Protein Microsequencing Facility, which is supported by a BRS Shared Instrumentation Grant (ISlORR05554) from the National Institutes of Health.
Address reprint requests to Robert I. Lehreq MD, Department of Medicine, UCLA-Center for the Health Sciences, 10833 LeConte Ave, Los Angeles, CA 90024-1876.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section 1734 solely to indicate this fact.
0 1992 by The American Society of Hematology. 0006-4971 1921 7906-0015$3.00/0
cleavage of proHNP-I and proHNP-3. The presence of two proline residues in the vicinity of this newly defined scission site suggested that this cleavage might be "proline-directed." Smaller amounts of the 34 aa and 32 aa prodefensin forms were also found. It remains to be established if these 39,34, and 32 aa prodefensins are obligate intermediates in the prodefensin processing pathway, or arise from side reac- tions. In either event, because these prodefensin intermedi- ates accounted for only 0.25% of the total defensin content, proteolytic conversion of 56 aa prodefensins to mature defensins appears to be a highly efficient process. o 1992 by The American Society of Hematology.
clues that may be relevant to the final processing steps involved in forming mature defensins from their precursors.
MATERIALS AND METHODS
Chemicals. High performance liquid chromatography (HPLC)- grade water and acetonitrile, as well as analytical reagent grade acetic acid, formic acid, and hydrogen peroxide were purchased from Fisher Scientific (Pittsburgh, PA). Trifluoroacetic acid (TFA), heptafluorobutyric acid (HFBA), ethyldimethylaminopropyl carbo- diimide (EDC), phenylisothiocyanate (PITC), and triethylamine were from Pierce Chemical (Rockford, IL). Acrylamide and molecular weight markers were from Bethesda Research Laborato- ries, Inc (Bethesda, MD). Lysine methyl ester, dithiothreitol, iodoacetamide, nitro blue tetrazolium (NBT), and 5-bromo-4- chloro-3-indolyl phosphate (BCIP) were obtained from Sigma (St Louis, MO). Sephacryl S-200 for gel permeation chromatography was from Pharmacia LKB Biotechnology (Piscataway, NJ). Alka- line phosphatase-conjugated goat antimouse or antirabbit IgG were purchased from Biorad (Richmond, CA).
Procurement of human neutrophil granules. Human neutrophils were obtained from seven normal donors by leukophoresis and were processed within 2 hours of harvest as previously described," except that the cells were disrupted by nitrogen cavitation at 750 psi essentially as described by Klempner et al,I4 rather than by homogenization. The granule-rich, 27,OOOg postnuclear sediments were collected and pooled in two separate batches, derived from four and three donors, respectively, and stored at -70°C until used. The pooled granules were extracted overnight in 5% acetic acid at 4"C, and after the extract was cleared at 27,OOOg for 20 minutes, the pellet was extracted two more times in the same way. The supernatants were pooled and concentrated to approximately 60 mL containing 8 to 10 x 108 cell-equivalentslml in a vacuum centrifuge (Speed Vac; Savant Instruments, Hicksville, NY).
The concen- trated extract of human neutrophil granules was loaded on a 4.8 x 105 cm (V, = 1,900 mL) Sephacryl S-200 column that had been equilibrated and was eluted with 5% acetic acid at a flow rate of 108 mL/h. The column effluent was monitored at 280 nm and 18-mL fractions were collected. Fractions containing peptides for further purification were identified by their electrophoretic mobil- ity on AU-PAGE. Appropriate fractions were pooled and lyophi- lized, and their contents were further purified by reversed-phase HPLC (RP-HPLC) on a 4.6 x 250 mm Vydac C-18 column (Separations Group, Hesperia, CA) using water-acetonitrile gradi- ents that contained 0.1% TFA. Fractions that contained mature defensins HNP-1, -2, and -3 were further purified by a combination of RP-HPLC (Vydac C-18) and Hydrophilic Interaction HPLC (Hydroxyethyl aspartamide column, PolyLC, Columbia, MD).
Purification of prodefensin peptides and defensins.
1532 Blood, Vol79, No 6 (March 15), 1992: pp 1532-1537
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom
DEFENSIN PRECURSORS 1533
Purity of the final product was assessed by tricine sodium dodecyl sulfate-PAGE (Tricine SDS-PAGE), by AU-PAGE, and by analyt- ical RP-HPLC, as previously described."
The purified peptides were hydrolyzed in vacuo in 5.7 N HCI for 36 to 40 hours. Amino acids were quantitated by RP-HPLC on a Novapak C-18 column (Millipore Waters, Milford, MA) after derivatization with phenylisothiocyanate (Picotag technique; Millipore Waters). For quantification of half-cystine residues, hydrolysis was performed on performic acid-oxidized peptides as previously described? Prode- fensins were reduced and carboxyamidomethylated, desalted by RP-HPLC, and subjected to gas-phase Edman degradation on a model 475 A sequencing system (Applied Biosystem, Inc, Foster City, CA).
Because some of the purified fractions were available only in minute amounts, the protein amounts reported in Table 2 were determined by quantitative amino acid analysis. This calculation was based on measuring their total content of phenylthiocarbamy- lated (FTC) amino acids and converting this data to total protein content by applying appropriate weighting factors derived from the sequence data.
A 20-fold molar excess of EDC (40 mg/mL) in 6 mol/L guanidine-HC1 was added to 1 mg of lyophilized HNP-1 in 0.5 mL of 6 mol/L guanidine-HC1, pH 4.75, and the pH was adjusted to between 5.0 and 6.0with 0.1 N NaOH. After this solution was incubated and mixed overnight, 50 pL of 1 mol/L sodium acetate was added and the mixture was dialysed against a 1,000-fold excess of 1% acetic acid for 3 hours, with hourly exchanges. At this point, SDS-PAGE analysis showed that the mixture contained both monomers and covalent dimers of HNP-1, in an approximate ratio of 3:l. Lysine methyl ester (0.29 pmol) was added to the mixture and the EDC coupling reaction was repeated, as above. After this step, we observed higher order multimers of HNP-1 (n = 3,4,5, and 6) on the SDS-PAGE gels as well as additional conversion of the HNP-1 monomers to dimers. This material was combined with Ribi MPL/TDM/CWS adjuvant (RIB1 ImmunoChem, Hamilton, MT) and used to immunize rabbits by conventional procedure^.'^ The resulting antibody, which showed the same specificity as a murine monoclonal antibody (MoAb) to HNP-1,16 recognized HNP-1, -2, and -3 but not HNP-4 on Western blots (data not shown).
After Tricine SDS-gel electrophoresis, the peptides of interest were electrotransferred onto polyvinylidenedifluoride (PVDF) membranes using the buffer system recommended by the apparatus manufacturer (Hoefer Scientific Instruments, San Francisco, CA). The membranes were then reacted with monoclonal or polyclonal antibody against HNP-1. Detection was accomplished by an alkaline phosphatase- conjugated second antibody system, using NBT and BCIP for visualization.
Composition analysis and sequence determination.
Preparation of a polyclonal antibody to HNP-1.
Immunostaining of prodefensin peptides.
RESULTS
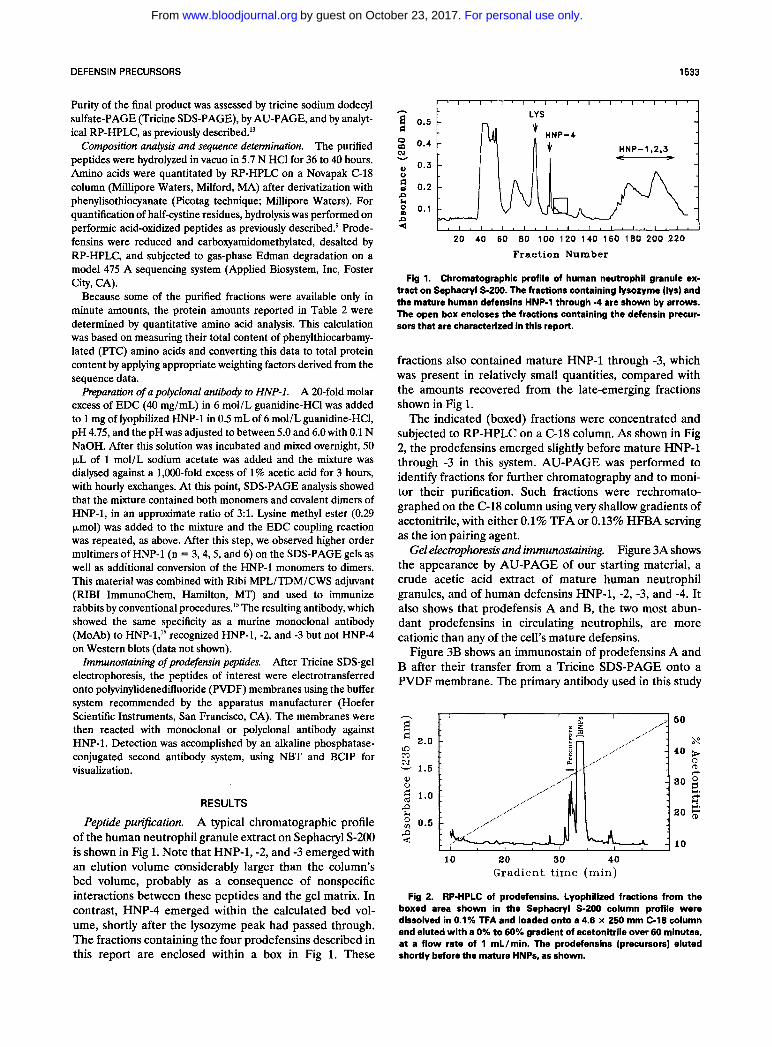
Peptide purification. A typical chromatographic profile of the human neutrophil granule extract on Sephacryl S-200 is shown in Fig 1. Note that HNP-1, -2, and -3 emerged with an elution volume considerably larger than the column's bed volume, probably as a consequence of nonspecific interactions between these peptides and the gel matrix. In contrast, HNP-4 emerged within the calculated bed vol- ume, shortly after the lysozyme peak had passed through. The fractions containing the four prodefensins described in this report are enclosed within a box in Fig 1. These
0.5 1 0.4
3 0.2
p 0.1 P
LY s 1
4 20 40 60 80 100 120 140 160 180 200 220
Fraction Number
Fig 1. Chromatographic profile of human neutrophil granule ex- tract on Sephacryl S-200. The fractions containing lysozyme (lys) and the mature human defensins HNP-1 through -4 are shown by arrows. The open box encloses the fractions containing the defensin precur- sors that are characterized in this report.
fractions also contained mature HNP-1 through -3, which was present in relatively small quantities, compared with the amounts recovered from the late-emerging fractions shown in Fig 1.
The indicated (boxed) fractions were concentrated and subjected to RP-HPLC on a C-18 column. As shown in Fig 2, the prodefensins emerged slightly before mature HNP-1 through -3 in this system. AU-PAGE was performed to identify fractions for further chromatography and to moni- tor their purification. Such fractions were rechromato- graphed on the C-18 column using very shallow gradients of acetonitrile, with either 0.1% TFA or 0.13% HFBA serving as the ion pairing agent.
Figure 3A shows the appearance by AU-PAGE of our starting material, a crude acetic acid extract of mature human neutrophil granules, and of human defensins HNP-1, -2, -3, and -4. It also shows that prodefensis A and B, the two most abun- dant prodefensins in circulating neutrophils, are more cationic than any of the cell's mature defensins.
Figure 3B shows an immunostain of prodefensins A and B after their transfer from a Tricine SDS-PAGE onto a PVDF membrane. The primary antibody used in this study
Gel electrophoresis and immunostaining.
..... $9 8 - 2.0
- 1.5
M c3 40 nl %
c Q,
$ 1.0 t $ 0.5 e d
30 0 2. z.
20 &- r+
l o 10 20 30 40
Gradient time (min)
Fig 2. RP-HPLC of prodefensins. Lyophilized fractions from the boxed area shown in the Sephacryl S-200 column profile were dissolved in 0.1% TFA and loaded onto a 4.6 x 250 mm C-18 column and eluted with a 0% to 60% gradient of acetonitrile over 60 minutes, at a flow rate of 1 mL/min. The prodefensins (precursors) eluted shortly before the mature HNPs, as shown.
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom

1534 HARWIG, PARK, AND LEHRER
Fig 3. PAGE of mature human defen- sins and prodefensins. (A) AU-PAGE (12.5%): lane 1, human PMN granule extract equivalent to 2 x 10' cells; lane 2.2 pg each of HNP-1, HNP-2, and HNP-3; lane 3, 2 pg of HNP-4; lane 4, 2 pg of purified prodefensin A; lane 5, 2 pg of purified prodefensin B. The gel was stained with Coomassie Brilliant Blue. (6) lmmunostaining of mature defensin peptides and prodefensin peptides after transfer onto PVDF membrane. Lane 1, prodefensin B; lane 2, prodefensin A; lane 3, 2 pg HNP 1; lane 4, prestained molecular weight markers (faintly visi- ble). The reagents included a polyclonal primary antibody against polymeric HNP1 and an alkaline phosphatase- conjugated second antibody with the BCIP-NBT substrate system. (C) Tricine SDS-PAGE (16.5%): lane 1, molecular weight markers; lane 2, 2 pg HNP-1; lane 3,2 pg HNP-3; lane 4,2 pg prodefen- sin A; lane 5,2 pg of prodefensin 6. Each sample was boiled for 5 minutes in sam- ple buffer containing 40 mmol/L dithio- threitol. The gel was stained with Coomassie Brilliant Blue.
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom
DEFENSIN PRECURSORS 1535
detected HNP-1, -2, and -3 with equal efficacy, but did not recognize HNP-4.
Figure 3C shows a Tricine SDS-PAGE gel, run under reducing conditions, and indicates that prodefensins A and B migrated with an apparent molecular weight of approxi- mately 4.3 Kd. The migration of HNP-1 and HNP-3 was consistent with their known molecular weights (Mr), as previously determined from sequence analysis (Mr HNP- 1 = 3,442; Mr HNP-3 = 3,486).
Taken together, these studies indicate that prodefensins A and B are slightly larger and more cationic than any of the mature human neutrophil defensins, and are immuno- logically related to HNP-1 through -3, but not to HNP-4.
Amino acid analysis (Table 1) showed that the compositions of prode- fensin peptides A and B were similar to each other and that both resembled mature defensin peptides HNP-1 or HNP-3. However, both prodefensins A and B also contained certain residues (most notably lysine and methionine) that were not present in the aforementioned mature defensins. Prode- fensins C and D were also subjected to amino acid analysis (Table 1). Both contained a methionine residue and prode- fensin C also contained a lysine in addition to those residues also present in mature HNP-1 and HNP-3.
Prodefensin A was composed of 39 amino acids (aa), and included the entire
Amino acid composition of the peptides.
Primaiy sequences of prodefensins.
Table 1. Amino Acid Compositions of Mature Defensins and Prodefensins
Mature Defensins Prodefensins
HNP-1 HNP-3 A B C D
Amino acids
Ala Arg Asx Cyst Glx GlY His Ile Leu
Met Phe Pro Ser Thr Trp Tyr
LY s
30 3.9 (4) 4.1 (4) 0.0 (0) 6.0 (6) 1.9 (2) 3.0 (3) 0.0 (0) 2.9 (3) 1.3 (1) 0.0 (0) 0.0 (0) 1.0 (1) 1.0 (1) 0.0 (0) 1.1 (1) ND (1) 3.0 (3)
30 3.0 (3) 4.1 (4) 0.9 (1) 5.5 (6) 1.9 (2) 3.0 (3) 0.0 (0) 2.9 (3) 1.2 (1) 0.0 (0) 0.0 (0) 1.0 (1) 1.0 (1) 0.0 (0) 1.1 (1) ND (1) 3.0 (3)
39 39 2.9 (3) 5.2 (5) 5.0 (5) 2.3 (2) 4.7 (6) 4.8 (6) 2.2 (2) 2.2 (2) 3.9 (4) 4.2 (4) 0.8 (1) 1.0 (1) 2.8 (3) 3.1 (3) 1.3 (1) 1.3 (1) 2.0 (2) 2.1 (2) 1.0 (1) 0.7 (1) 0.9 (1) 1.1 (1) 1.9 (2) 2.3 (2) 1.2 (1) 1.2 (1) 1.0 (1) 1.2 (1) ND (1) ND (1) 2.6 (3) 2.9 (3)
4.0 (3 or 4)'
1.6 (1 or 2)*
34 3.1 (3) 4.8 (5) 2.1 (2) 5.0 (6) 2.2 (2) 3.4 (3) 0.0 (0) 2.8 (3) 1.1 (1) 1.0 (1) 0.6 (1) 0.9 (1) 1.3 (1) 0.4 (0) 1.2 (1) ND (1) 3.2 (3)
32 3.1 (3) 3.8 (4) 2.2 (2) 4.6 (6) 2.1 (2) 3.0 (3) 0.0 (0) 2.8 (3) 1.1 (1) 0.0 (0)
0.9 (1) 1.2 (1) 0.4 (0) 1.1 (1) ND (1) 2.7 (3)
0.7 (1)
Values determined from analysis of 40-hour hydrolysates. Numbers within parentheses indicate residues as determined by sequence analysis.
Abbreviations: Asx, aspartic acid or asparagine; Glx, glutamic acid or glutamine; ND, not determined.
"This peptide contained a mixture of asp and ala in a ratio of 57:43 (D:A) on the 10th cycle of its sequence analysis.
tCysteine was determined as cysteic acid
I O I
MRTLA I LAA I LLVALQAQA Signal SEqUeilCe
20 30 Y O SO 60 I I I I i EPLQARADEVAAAPEQ IAA D I PEVVVSLAWDESLAP KHPGS RK NM proplece
! + 0 00 70 EO 90
A I I I DCYCR I PAC IAGERRYGTC IYQGRLWAFCC mature - ...t+ HNRI
HNW
Fig 4. Proteolytic cleavage sites of HNP-1 through -3 precursor peptides. The amino acid sequences of the HNP-1 and HNP-3 prepro- defensins are shown in standard single letter code. The cleavage sites that generate the 75 aa and 56 aa precursors" are shown by closed arrows, those which generate the 39 aa, 34 aa, and 32 aa proHNPs described in this report are indicated by open arrows. The sites whose cleavage would generate mature HNP-1 and -3 and HNP-2 and the intramolecular cysteine disulfide pairs of the mature defensin mole- cule are also shown.
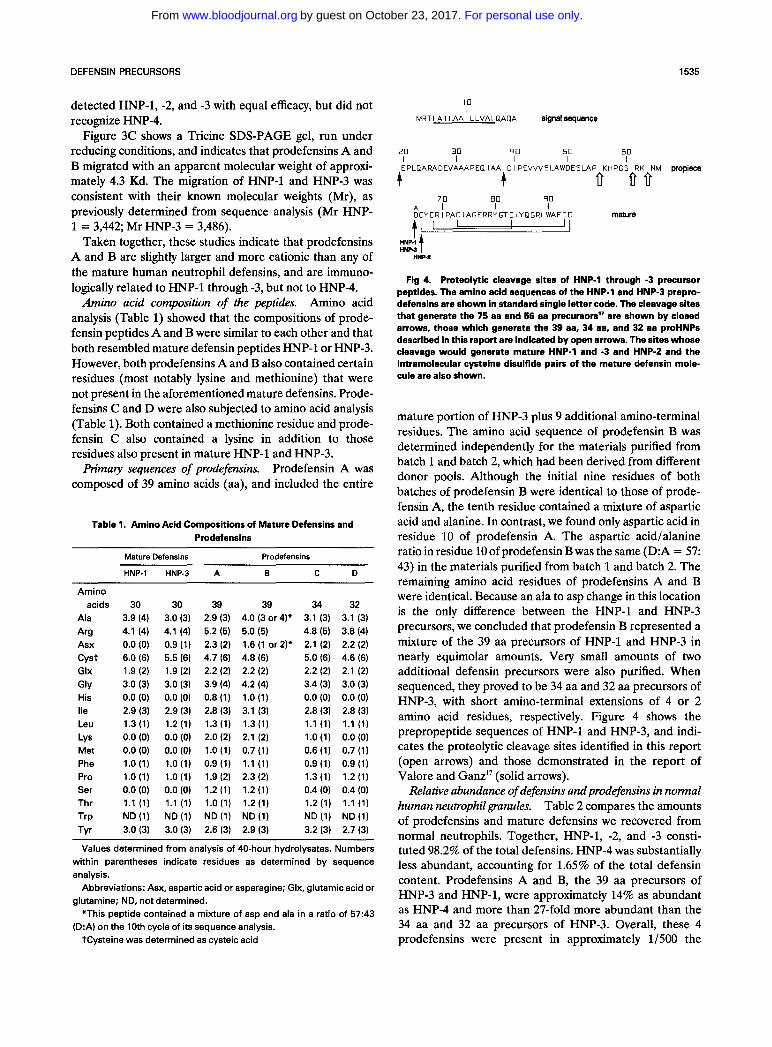
mature portion of HNP-3 plus 9 additional amino-terminal residues. The amino acid sequence of prodefensin B was determined independently for the materials purified from batch 1 and batch 2, which had been derived from different donor pools. Although the initial nine residues of both batches of prodefensin B were identical to those of prode- fensin A, the tenth residue contained a mixture of aspartic acid and alanine. In contrast, we found only aspartic acid in residue 10 of prodefensin A. The aspartic acid/alanine ratio in residue 10 of prodefensin B was the same (D:A = 57: 43) in the materials purified from batch 1 and batch 2. The remaining amino acid residues of prodefensins A and B were identical. Because an ala to asp change in this location is the only difference between the HNP-1 and HNP-3 precursors, we concluded that prodefensin B represented a mixture of the 39 aa precursors of HNP-1 and HNP-3 in nearly equimolar amounts. Very small amounts of two additional defensin precursors were also purified. When sequenced, they proved to be 34 aa and 32 aa precursors of HNP-3, with short amino-terminal extensions of 4 or 2 amino acid residues, respectively. Figure 4 shows the prepropeptide sequences of HNP-1 and HNP-3, and indi- cates the proteolytic cleavage sites identified in this report (open arrows) and those demonstrated in the report of Valore and Ganz17 (solid arrows).
Relative abundance of defensins andprodefensins in normal human neutrophilgranules. Table 2 compares the amounts of prodefensins and mature defensins we recovered from normal neutrophils. Together, HNP-1, -2, and -3 consti- tuted 98.2% of the total defensins. HNP-4 was substantially less abundant, accounting for 1.65% of the total defensin content. Prodefensins A and B, the 39 aa precursors of HNP-3 and HNP-1, were approximately 14% as abundant as HNP-4 and more than 27-fold more abundant than the 34 aa and 32 aa precursors of HNP-3. Overall, these 4 prodefensins were present in approximately 1/500 the
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom

1536 HARWIG, PARK, AND LEHRER
Table 2. Relative Recovery of Prodefensin Peptides From Normal Human Neutrophils
No. of Mature Defensins Prodefensins Leukophoresis
Batch Units Total Cells HNP-1 Through -3 HNP-4 A B C D
1 4 5.9 x 10'0 72.5' 1.1 0.043 0.061 NA NA 2 3 4.7 f 10'0 52.8 1 .o 0.031 0.157 0.0023 0.0045 Combined 7 10.6 x 10" 125.3 2.1 0.074 0.157 0.0023 0.0045
% of total 98.2 1.65 0.058 0.171 0.002 0.004
Abbreviation: NA, not analyzed. 'Amount purified (mg), determined by amino acid analysis.
abundance of the mature defensins HNP-1 through -4, suggesting that the final steps in proteolytic processing of defensins by neutrophils were highly efficient.
DISCUSSION
The human neutrophil's principal defensins (HNP-1, -2, and -3) are synthesized as 94 aa prepropeptides.'* Conse- quently, to form the mature defensins, which are either 29 (HNP-2) or 30 (HNP-1 and -3) aa long, these precursors must undergo proteolytic processing. Valore and Ganz recently showed that the early processing steps entail sequential formation of two intermediate prodefensin forms, 75 and 56 aa long, re~pectively.'~ This report describes an additional 39 aa prodefensin precursor for both HNP-1 and HNP-3, as well as the recovery of much smaller amounts of 32 and 34 aa precursors of HNP-3. Together, these prode- fensins constituted approximately 0.23% of the total defen- sins recovered from circulating normal neutrophils.
Because the sequences of the preprodefensins are known from their respective cDNAs,'* it is apparent that the 39 aa precursors of both HNP-1 and HNP-3 resulted from en- doproteolytic cleavage between pross and lyss6 in their corresponding precursor peptides (numbered, in this case, from the amino-terminal methionine residue of the prepro- defensin). Although such an event could result from the action of a trypsin-like enzyme, the arginine-rich and potentially trypsin-susceptible nature of defensins suggests that a more specific processing enzyme is likely to be involved in generating the 39 aa forms. Given the existence of vicinal proline residues, both amino- and carboxy- terminal to the indicated scission site of proHNP-l and proHNP-3, a proline-directed monobasic cleavage model, deserves consideration. Although this type of monobasic proteolytic processing has been noted for a wide variety of peptides, including PGLa and xenopsin from frog skin and yeast killer toxin,19 it more often occurs at arginine than lysine residues, as indicated in the formation of the 39 aa prodefensins A and B.
The next most abundant small defensin precursor that we found was the 34 aa form of HNP-3, which would have arisen from hydrolysis of a bond between serm and the dibasic, arg61-1ys62 doublet in the precursor (again num- bered from the amino-terminal methionine of preprodefen- sin). Although dibasic residues are common sites of proteo- lytic processing in prohormones, bond scission usually occurs between or carboy-terminal to the basic residues rather than amino-terminal to them?' However, at least two
processing enzymes that cleave amino-terminal to a dibasic arg-lys pair, as required to generate the 34 aa prodefensin form of HNP-3, have been reported in other systems. One of these cleaves a bond between glulz and lys,,-arg,, in somatostatin*' and the other cleaves the bond between leu, and ark-lys, of neoendorphin as well as the bond between met, and ark-arg, of BAMP-12.22
Although the final trimming of the 34 aa precursors to the 30 aa mature defensins could be mediated by sequential aminopeptidase cleavages, our detection of small amounts of the 32 aa precursor of HNP-3 without discerning the 33 aa and 31 aa forms raises the possibility that this final trimming may involve two sequential cleavages mediated by a dipeptidyl aminopeptidase, analogous to the process recently described for two insect antimicrobial peptides, cecropins A and B.23
Recently, Bateman et a1 isolated two prodefensin pep- tides from human HL-60 promyelocytic leukemia cells." One was a 56 aa long precursor of HNP-1, which they named "HP 1-56." Although they found this 56 aa form to be prominent in HL-60 cells and immature leukocytes, they reported difficulty in detecting it in mature neutrophils. We have also recovered substantial amounts of this 56 aa form from the immature leukocytes of patients with chronic myelogenous leukemia, but not from mature neutrophils (data not shown), consistent with the formation of this prodefensin in immature myeloid cells that are not repre- sented in normal circulating blood. Bateman et a1" did not identify the additional putative HNP precursor. However, speculating that it might result from cleavage at a dibasic sequence, they inferred that it might be the 32 aa prodefen- sin form. Although we identified the hypothecated 32 defensin precursor in the present study, it was present in minute concentrations, compared with the 39 aa precursor forms of HNP-1 and HNP-3. Consequently, we suggest that the unidentified small defensin precursor observed by Bateman et a1" was more likely to have been the same 39 aa prodefensin reported here.
It remains to be determined if the minor prodefensins identified in this report are side products of defensin biosynthesis or obligatory intermediates in this process. If it is assumed that these minor prodefensins are produced by the mainstream defensin processing mechanism, a model for the final steps in the proteolytic processing of defensins by human neutrophils can be formulated (Fig 4). Formal proof that the prodefensins characterized in this report accurately mark the mainstream pathway of defensin pro-
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom

DEFENSIN PRECURSORS 1537
cessing must await further study with defensins whose primary structures have been selectively altered in the putative processing sites.
Although this model deals specifically with the produc- tion of mature (30 aa) HNP-1 and HNP-3, it should be noted that either of these defensins could be converted to HNP-2 simply by removing one additional amino-terminal
residue. Thus, the abundant messenger RNA for HNP-1'8,25 and HNP-318326 that has been found in immature myeloid cells could also suffice for the production of HNP-2.
ACKNOWLEDGMENT
We thank Dr Tomas Ganz for helpful discussions and sugges- tions.
REFERENCES 1. Ganz T, Selsted ME, Lehrer RI: Defensins. Eur J Haematol
44:1,1990 2. Lehrer RI, Gam T: Antimicrobial polypeptides of human
neutrophils. Blood 76:2169,1990 3. Territo MC, Ganz T, Se1ste.d ME, Lehrer RI: Monocyte-
chemotactic activity of defensins from human neutrophils. J Clin Invest 84:2017,1989
4. Fleischmann J, Selsted ME, h h r e r RI: opsonic activity of MCP-1 and MCP-2, cationic peptides from rabbit alveolar macro- phages. Diagn Microbiol Infect Dis 3:233,1985
5. Zhu Q, B~~~~~~ A, singh A, solomon S: ~ ~ ~ l ~ ~ i ~ ~ and biological activity of corticostatic peptides (an t i -Am). Endo- crinol Res 15:129,1989
6. Tominaga T, Fukata J, Nakai Y, Funakoshi S, Fujii N, Imura
14. Klempner MS, Mikkelsen RB, Corfman DH, Andre- Schwartz A Neutrophil plasma membranes. I. High-yield purifica- tion of human neutrophil plasma membrane vesicles by nitrogen cavitation and differential centrifugation. J Cell Biol86:21,1980
15. Harlow E, Lane D: Antibodies, A Laboratory Manual. Cold Spring Harbor, m, Cold Spring Harbor Laboratory, 1988, P 92
16. Panyutich AN, Voitenok N, Lehrer RI, Ganz T An enzyme immunoassay for human defensins. J h"mnol Methods 141:149, 1991
17. Valore EV, Ganz T: Posttranslational processing of defen- sins in immature human myeloid cells. Blood 79:1538,1992
18. D a h Lehrer RI, Ganz T, Kronenberg M: Isolation and characterization of human defensin cDNA clones. Proc Natl
USA 85:73177 1988 H: Effects of c0rticostatin-I on rat adrenal cells in vitro. J
19. Tw: The processing Of peptide precursors' FEBS Endocrinol125:287,1990 Lett 2001,1986
7. Yamashita T, Saito K Purification, primary structure and biological activity of guinea pig neutrophil peptides. Infect Immun 57:2405,1989
Jr, Ganz T, Selsted ME, Lehrer RI, Kuo JF Inhibition of protein kinase C by defensins, antibiotic peptides from human neutrophils. Biochem Pharmacol37951,1988
Primary structures of three human neutrophil defensins. J Clin Invest 76:1436,1985
10. Rice WG, Ganz T, Kinkade JM Jr, Selsted ME, Lehrer RI, Parmley R T Defensin-rich granules of human neutrophils. Blood 70757,1987 2645862,1989
11. Singh A, Bateman A, Zhu Q, Shimasaki S, Esch F, Solomon s: Sh-~cture of a novel granulocyte Peptide with anti-ACTH activity. Biochem Biophys Res Commun 155524,1988
12. Gabay JE, Scott RW, Campanelli D, Grfith J, Wilde C, Marra MN, Seeger M, Nathan C F Antibiotic proteins of human polymorphonuclear leukocytes. Proc Natl Acad Sci USA 865610, 1989 some 8q21.1-22. Blood 71:1713,1988
13. Ganz T, Selsted ME, Szklarek D, Harwig SSL, Daher K, Bainton DF, Lehrer RI: Defensins: Natural peptide antibiotics of human neutrophils. J Clin Invest 76:1427,1985
20. Gluschankof P, Cohen P: Proteolytic enzymes in the post- translational processing of polypeptide hormone precursors. Bio- chem Res 12951,1987
21. Gluschankof P, Morel A, Benoit R, Cohen P: The somato- " Rice WG3 RL7 Khkade JM statin-28 convertase of rat brain cortex generates both somatostat-
in-14 and somatostatin-28 (l-12). Biochem Biophys Res Commun 128:1051, 1987
22. Camargo ACM, Ribeiro MJVF, Schwartz W N Conversion 9. Selsted ME, Harwig SSL, Ganz T, Schilling m, Lehrer RI: and inactivation of opioid peptides by rabbit brain endo-
oligopeptidase A. Biochem Biophys Res Commun 130:932, 1985 23. Boman HG, Boman IA, Andreu D, Li Z-q, Merrifield RB,
Schlenstedt G, Zimmeman R Chemical synthesis and enzymic processing of precursor forms of cecropins A and B. J ~ i ~ l Chem
24. Bateman A, Singh A, Shustik C, Mars WM, Solomon S: The isolation and identification of multiple forms of the neutrophil granule peptides from human leukemic cells. J Biol Chem 286: 7524,1991
25. Mars WM, vanTuinen P, Drabkin HA, White JW, Saunders G F A myeloid-related sequence that localizes to human chromo-
26. Wiedemann LM, Francis GE, Lamb RF, Burns JH, Winnie JN, MacKenzie ED, Birnie GD: Differentiation stage-specific expression of a gene during granulopoiesis. Leukemia 3:227,1989
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom

1992 79: 1532-1537
SS Harwig, AS Park and RI Lehrer Characterization of defensin precursors in mature human neutrophils
http://www.bloodjournal.org/content/79/6/1532.full.htmlUpdated information and services can be found at:
Articles on similar topics can be found in the following Blood collections
http://www.bloodjournal.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://www.bloodjournal.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://www.bloodjournal.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Society of Hematology, 2021 L St, NW, Suite 900, Washington DC 20036.Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by the American
For personal use only.on October 23, 2017. by guest www.bloodjournal.orgFrom