characterization of an abaecin-like antimicrobial peptide identified from a pteromalus puparum cdna...
TRANSCRIPT

Journal of Invertebrate Pathology 105 (2010) 24–29
Contents lists available at ScienceDirect
Journal of Invertebrate Pathology
journal homepage: www.elsevier .com/ locate / j ip
Characterization of an abaecin-like antimicrobial peptide identifiedfrom a Pteromalus puparum cDNA clone
Xiaojing Shen a, Gongyin Ye a,*, Xiongying Cheng b, Chunyan Yu c, Illimar Altosaar d, Cui Hu a
a State Key Laboratory of Rice Biology, Ministry of Agriculture Key Laboratory of Molecular Biology of Crop Pathogens and Insects, Institute of Insect Sciences,College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310029, Chinab Vitegen LLC, Cary NC 27519, USAc The First Affiliated Hospital of College of Medicine, Zhejiang University, Hangzhou 310003, Chinad Centre for Research on Environmental Microbiology, Department of Biochemistry, Microbiology and Immunology, Faculty of Medicine, University of Ottawa, 451 Smyth Road,Ottawa, Canada K1H 8M5
a r t i c l e i n f o
Article history:Received 22 December 2009Accepted 7 May 2010Available online 11 May 2010
Keywords:Antimicrobial peptideAbaecinInsect defensePteromalus puparumPieris rapaeEndoparasitoid wasp
0022-2011/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.jip.2010.05.006
* Corresponding author. Address: Institute of Insecture and Biotechnology, Zhejiang University, Hangzhou8604 9815.
E-mail address: [email protected] (G. Ye).
a b s t r a c t
Abaecin is a major antimicrobial peptide, initially identified from the honeybee. In our effort to discovernew antimicrobial peptides from the endoparasitoid wasp Pteromalus puparum, we identified an antibac-terial cDNA clone that codes a fragment with high amino acid sequence similarity to abaecin. The proline-rich peptide (YVPPVQKPHPNGPKFPTFP, named PP30) was chemically synthesized and characterized inthis study. Antimicrobial assays indicated that the cationic peptide was active against both Gram-nega-tive and positive bacteria, but not active against fungi tested. No hemolytic activity was observed againsthuman erythrocytes after 1 h incubation at concentration of 125 lM or below. The antibacterial activityof PP30 against Escherichia coli was attenuated in the presence of increasing concentrations of NaCl.Transmission electron microscopic (TEM) examination of PP30-treated E. coli cells showed morphologicalchanges in the cells and extensive damage to the cell membranes. The circular dichroism (CD) spectros-copy studies indicated that PP30 formed random coil structures in phosphate buffer (pH 7.4), 50% TFE and25 mM SDS solution. Expression analysis of the gene coding for the peptide indicated that its expressionwas upregulated upon bacterial infection, indicating that the gene may play a role in preventing potentialinfection by microorganisms during parasitization in Pieris rapae.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction
Antimicrobial peptides/proteins (AMPs) (such as cecropins,defensins, proline/glycine-rich antibacterial peptides, and anti-fungal proteins) have been demonstrated to play a very importantrole in insect humoral defense reactions. They are produced andsecreted into the hemolymph in response to microbial infectionor septic body injury (Angus et al., 2001). These peptides are pro-duced in quantity mainly by the fat body, various epithelia, andcertain hemocytes (Bartholomay et al., 2004; Irving et al., 2005),then rapidly released into hemolymph where they act synergisti-cally against bacteria and fungi and even certain viruses (Tzouet al., 2002). Since Boman and co-workers (Steiner et al., 1981)isolated two families of antibacterial peptides, large number of
ll rights reserved.
t Sciences, College of Agricul-310029, China. Fax: +86 571
AMPs have been isolated and characterized. Among about890 AMPs identified from eukaryotic origin so far, more than 180were identified from insects (Cheng et al., 2006; Cytrynska et al.,2007).
Although numerous AMPs have been reported in hymenopteraninsects such as honeybees, wasps, bumble bees and ants (Cerovskyet al., 2008; Xu et al., 2009), there has been few research focused onAMPs of endoparasitoid wasps. Pteromalus puparum (Hymenop-tera: Pteromalidae) is a predominant pupal endoparasitoid waspof Pieris rapae (cabbageworm). Over the last several years, we havebeen investigating the interactions between P. puparum and P. ra-pae, and demonstrated that the venom of P. puparum contains arich array of peptides, including those with biological activity(Cai et al., 2004; Zhang et al., 2005; Wu et al., 2008). In this report,we describe further characterization of an abaecin-like AMPidentified previously from a P. puparum cDNA clone (Cheng et al.,2009). This peptide was found to be active against Gram-negativeand Gram-positive bacteria with a very weak hemolyticactivity, and its coding gene expression upregulated by bacterialinfection.

X. Shen et al. / Journal of Invertebrate Pathology 105 (2010) 24–29 25
2. Materials and methods
2.1. cDNA clone isolation, sequencing, and peptide synthesis
The cDNA clone isolation and sequencing of an abaecin-likeAMP (PP30) have been described previously with a vital dye-stain-ing method (Cheng et al., 2009). It was screened from a cDNAexpression library, which was developed with the mixture wholebody of P. puparum pupae, pharate adults and adults (1:1:1.5).The synthetic peptide was acquired as described previously (Shenet al., 2010). Briefly, 20 mg of peptides were commercially synthe-sized by solid-phase methods at the Shanghai Bootech BioScience(Shanghai, China) and Technology Company, using N-9 FMOCchemistry with Fmoc-Ile-Wang resin (STAM, USA) on a 431A Pep-tide Synthesizer (Applied Biosystems). The peptides were purifiedby HPLC to 90% purity for use in this study.
2.2. Assay of antibacterial activity, minimal inhibitory concentrations(MIC) determination and salt-dependence test
To assay the antibacterial activity of the clone containing thecDNA of PP30, the cells from the isolated Escherichia coli colonywere used to compare their growth in non-induction mediumand inductive medium as previously described (Cheng et al., 2009).
MIC determination was performed as described previously(Shen et al., 2010). Briefly, bacterial strains Staphylococcus aureus(CVCC1882), E. coli (CVCC1570), Sarcina lutea (CVCC1600), Bacilluspumilus (CVCC709) and B. subtilis (CVCC717) were purchased fromChina Institute of Veterinary Drug Control. The MIC of the syntheticpeptide PP30 were determined based on a broth microdilution as-say modified from the method reported elsewhere (Otvos, 2002;Shen et al., 2010).
Salt-dependence test was carried out as previously described(Shen et al., 2010). The diluted E. coli culture was aliquoted into96-well plates and incubated with the peptide at 50% MIC in M9medium containing different concentrations of NaCl. The bacterialculture was incubated for 12 h at 37 �C and the absorbance wasmeasured at 600 nm. All assays were performed in triplicate.
2.3. Hemolytic assay
Hemolytic assay was performed using human erythrocytes aspreviously described with slight modification (Ryge and Hansen,2005; Shen et al., 2010). Briefly, the erythrocytes were washedthree times (3000 rpm) with phosphate buffered saline (0.15 M,pH 7.2) and diluted to a final concentration of 0.5% in the same buf-fer. To each well of a polypropylene microtiter plate 75 ll of the di-luted erythrocytes and 75 ll of peptide solution at 250, 125, 62.5,31.2, 15.6, 7.8, 3.9, and 1.95 lM were added. After incubated for 1 hat 37 �C, samples were centrifuged at 4000 rpm for 10 min and theabsorbance of supernatant was determined at 414 nm with an ELI-SA plate reader. Negative control (zero hemolysis (blank)) and po-sitive control (100% hemolysis) were achieved by suspending theerythrocytes in PBS and 0.1% PBS with Triton X-100, respectively.The hemolysis percentage was calculated as follows: [(Apep-
tide � APBS)/(ATriton � APBS)] � 100. All hemolysis determinationswere performed in duplicate and are the average of three indepen-dent determinations using the same stock solution.
2.4. Transmission electron microscopy (TEM)
To investigate the effect of the newly identified peptide on cellmembrane, we examined the membrane integrity of E. coli cellsafter treatment with the peptide at MIC (Shen et al., 2010). The sec-
tions were observed in TEM of Model JEM-1230. The negative con-trol was run in the absence of the peptide.
2.5. Circular dichroism (CD)
To analyze the circular dichroism (CD) of the synthetic peptidePP30, it was performed as described previously (Shen et al., 2010).In a brief, the CD spectra were recorded at room temperature on aJasco J-815 spectropolarimeter (Tokyo, Japan) using a quartz cell of1 mm path length. Spectra were obtained from 260 to 190 nm byaveraging two scans at a scan speed of 10 nm/min CD spectra werereported in terms of ellipticity units per mole and the a-helicitywas calculated from the mean residue ellipticity at 222 nm.
2.6. RT-PCR
RT-PCR was used to detect transcripts of the gene encodingPP30 after feeding the insects with different concentrations of E.coli, P. puparum female adults were reared with honey water(20%) containing different concentrations of E. coli cells (107, 106,105, 104 colony forming units (CFU)/ml) for 24 and 48 h. TotalRNA was then extracted using a Total RNA Extraction Kit (Prome-ga). First strand cDNA was synthesized from 2 lg of the totalRNA using RevertAid™ First Strand cDNA Synthesis Kit #K1621(Fermentas) and used as template for RT-PCR using the followingprimers 50-GCTCTAGAATGAAGTTCGTCCTCAGT-30 (forward primer)and 50-ACGCTAGCTGACTGAAATTGGTGTTA-30 (reverse primer).Elongation factor 18S rRNA was amplified as an internal control(Wang et al., 2008). The PCR reaction was conducted with the ini-tial pre-denaturation step at 94 �C for 5 min followed by 30 cyclesof denaturing at 94 �C for 30 s, annealing at 52 �C for 60 s andextension at 72 �C for 30 s, and the final extension step at 72 �Cfor 5 min. 10 ll of the amplification products was analyzed andvisualized by electrophoresis in a 1% agarose gel containing ethi-dium bromide.
3. Results and discussion
3.1. Isolation of antibacterial clone containing an abaecin-like peptidedomain
We screened a cDNA expression library from P. puparum andisolated clones that contain DNA sequences with E. coli inhibitoryeffect (Cheng et al., 2009). One of the clones PP30 (GenBank acces-sion No. EF444537.1) contains a cDNA with an open reading frameof 354 bp (Fig. 1A). While no significant nucleotide sequencehomology was found after blast search against the public database(http://blast.ncbi.nlm.nih.gov/Blast.cgi), its translated amino acidsequence has an N-terminal segment that is 52% identical to thefirst 19 amino acids of abaecin (GenBank accession No.AAL35348) (Fig. 1B), a major antibacterial peptide isolated fromhoneybee (Casteels et al., 1990). This segment is also highly similarto abaecins from other insects such as Myrmica ruginodis and Man-ica rubida (Viljakainen et al., 2009). In comparison with abaecinprecursor from honeybee, which has a 19 amino acid signal pep-tide, the deduced wasp protein is likely to have a signal peptideof 22 amino acids. When E. coli cells containing this cDNA clonewere grown under induced condition (with addition of 1 mM IPTG(isopropyl-b-D-thiogalactoside) in culture medium), their growthrate was reduced by 50% in 10 h growth period compared to unin-duced cells (Table 1). We therefore reasoned that the abaecin-likesegment (PP30, YVPPVQKPHPNGPKFPTFP) might be attributed tothe cell inhibition effect. The segment spans amino acid 21–40 inthe deduced protein and is a proline-rich and cationic peptide. Pro-line-rich and positive charges are typical characteristics of many

Fig. 1. (A) The nucleotide sequence of cDNA clone coding for PP30 from P. puparum and deduced amino acid sequence. The sequence of PP30 is underlined. The asterisk (�)indicates stop condon; (B) amino acid sequence alignment between abaecin (Casteels et al., 1990) and PP30 conserved amino acid residues are highlighted.
Table 1Growth of E. coli cells harboring cDNA coding for PP30 in non-inductive and inductivemedium.
Colony ID Growth medium Growth reduction (%)
Non-inductive Inductive
Control 1.079a ± 0.140 1.013 ± 0.089 6.11PP30 0.693 ± 0.025 0.287 ± 0.019 50.0
a A600 measured 10 h after culture. Ten microliter aliquots of E. coli cells wereinoculated in 5 ml of LB medium (0.5% yeast extract, 1% peptone, 1% NaCl, 100 lg/ml ampicillin) with and without inducer (1 mM IPTG). The A600 was measured after10 h incubation as an index of cell growth. The data are represented asmean ± standard deviation.
26 X. Shen et al. / Journal of Invertebrate Pathology 105 (2010) 24–29
AMPs. For examples, several AMPs isolated from sheep and goatleukocytes are proline-rich (Shamova et al., 1999). Using an agardiffusion assay, we demonstrated that a peptide synthesized basedon the 19 amino acid sequence did display clear zone when 5 ll of
1 mM peptide solution was applied. The peptide is therefore fur-ther characterized in the study to explore its potential use.
3.2. MICs
The HPLC-purified synthetic peptide was used to determineits MICs against several Gram-negative and Gram-positive bacte-ria. Table 2 summarizes the results of MIC determinationsagainst both Gram-negative and Gram-positive bacteria withMIC varied between 13.3 and 133.3 lM. The abaecin-like peptidehas similar MIC against E. coli in comparison with abaecin(Casteels et al., 1990). It is active against both Gram-positiveand negative bacteria, as demonstrated with abaecin (Kuhn-Nentwig, 2003).
It was also observed that the peptide acted as a bactericidalagent against these bacteria except S. aureus, since the co-cultureof the peptide with the bacteria at concentration above MICs re-sulted in loss of viability of the bacteria. However, antifungal assay
0
0.05
0.1
0.15
0.2
0.25
0.3
0.35
0 20 40 60 80 100 150NaCl concentration(µM)
Cel
l gro
wth
(OD
600)
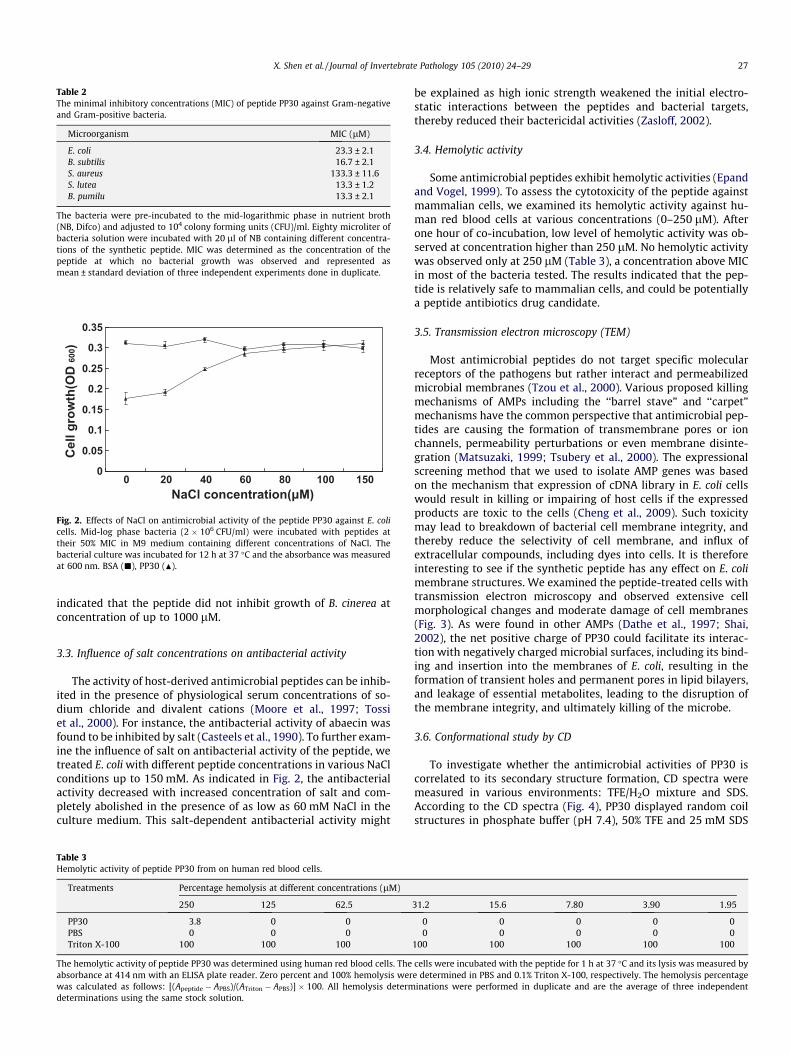
Fig. 2. Effects of NaCl on antimicrobial activity of the peptide PP30 against E. colicells. Mid-log phase bacteria (2 � 106 CFU/ml) were incubated with peptides attheir 50% MIC in M9 medium containing different concentrations of NaCl. Thebacterial culture was incubated for 12 h at 37 �C and the absorbance was measuredat 600 nm. BSA (j), PP30 (N).
Table 2The minimal inhibitory concentrations (MIC) of peptide PP30 against Gram-negativeand Gram-positive bacteria.
Microorganism MIC (lM)
E. coli 23.3 ± 2.1B. subtilis 16.7 ± 2.1S. aureus 133.3 ± 11.6S. lutea 13.3 ± 1.2B. pumilu 13.3 ± 2.1
The bacteria were pre-incubated to the mid-logarithmic phase in nutrient broth(NB, Difco) and adjusted to 104 colony forming units (CFU)/ml. Eighty microliter ofbacteria solution were incubated with 20 ll of NB containing different concentra-tions of the synthetic peptide. MIC was determined as the concentration of thepeptide at which no bacterial growth was observed and represented asmean ± standard deviation of three independent experiments done in duplicate.
X. Shen et al. / Journal of Invertebrate Pathology 105 (2010) 24–29 27
indicated that the peptide did not inhibit growth of B. cinerea atconcentration of up to 1000 lM.
3.3. Influence of salt concentrations on antibacterial activity
The activity of host-derived antimicrobial peptides can be inhib-ited in the presence of physiological serum concentrations of so-dium chloride and divalent cations (Moore et al., 1997; Tossiet al., 2000). For instance, the antibacterial activity of abaecin wasfound to be inhibited by salt (Casteels et al., 1990). To further exam-ine the influence of salt on antibacterial activity of the peptide, wetreated E. coli with different peptide concentrations in various NaClconditions up to 150 mM. As indicated in Fig. 2, the antibacterialactivity decreased with increased concentration of salt and com-pletely abolished in the presence of as low as 60 mM NaCl in theculture medium. This salt-dependent antibacterial activity might
Table 3Hemolytic activity of peptide PP30 from on human red blood cells.
Treatments Percentage hemolysis at different concentrations (lM)
250 125 62.5
PP30 3.8 0 0PBS 0 0 0Triton X-100 100 100 100
The hemolytic activity of peptide PP30 was determined using human red blood cells. Theabsorbance at 414 nm with an ELISA plate reader. Zero percent and 100% hemolysis werwas calculated as follows: [(Apeptide � APBS)/(ATriton � APBS)] � 100. All hemolysis determdeterminations using the same stock solution.
be explained as high ionic strength weakened the initial electro-static interactions between the peptides and bacterial targets,thereby reduced their bactericidal activities (Zasloff, 2002).
3.4. Hemolytic activity
Some antimicrobial peptides exhibit hemolytic activities (Epandand Vogel, 1999). To assess the cytotoxicity of the peptide againstmammalian cells, we examined its hemolytic activity against hu-man red blood cells at various concentrations (0–250 lM). Afterone hour of co-incubation, low level of hemolytic activity was ob-served at concentration higher than 250 lM. No hemolytic activitywas observed only at 250 lM (Table 3), a concentration above MICin most of the bacteria tested. The results indicated that the pep-tide is relatively safe to mammalian cells, and could be potentiallya peptide antibiotics drug candidate.
3.5. Transmission electron microscopy (TEM)
Most antimicrobial peptides do not target specific molecularreceptors of the pathogens but rather interact and permeabilizedmicrobial membranes (Tzou et al., 2000). Various proposed killingmechanisms of AMPs including the ‘‘barrel stave” and ‘‘carpet”mechanisms have the common perspective that antimicrobial pep-tides are causing the formation of transmembrane pores or ionchannels, permeability perturbations or even membrane disinte-gration (Matsuzaki, 1999; Tsubery et al., 2000). The expressionalscreening method that we used to isolate AMP genes was basedon the mechanism that expression of cDNA library in E. coli cellswould result in killing or impairing of host cells if the expressedproducts are toxic to the cells (Cheng et al., 2009). Such toxicitymay lead to breakdown of bacterial cell membrane integrity, andthereby reduce the selectivity of cell membrane, and influx ofextracellular compounds, including dyes into cells. It is thereforeinteresting to see if the synthetic peptide has any effect on E. colimembrane structures. We examined the peptide-treated cells withtransmission electron microscopy and observed extensive cellmorphological changes and moderate damage of cell membranes(Fig. 3). As were found in other AMPs (Dathe et al., 1997; Shai,2002), the net positive charge of PP30 could facilitate its interac-tion with negatively charged microbial surfaces, including its bind-ing and insertion into the membranes of E. coli, resulting in theformation of transient holes and permanent pores in lipid bilayers,and leakage of essential metabolites, leading to the disruption ofthe membrane integrity, and ultimately killing of the microbe.
3.6. Conformational study by CD
To investigate whether the antimicrobial activities of PP30 iscorrelated to its secondary structure formation, CD spectra weremeasured in various environments: TFE/H2O mixture and SDS.According to the CD spectra (Fig. 4), PP30 displayed random coilstructures in phosphate buffer (pH 7.4), 50% TFE and 25 mM SDS
31.2 15.6 7.80 3.90 1.95
0 0 0 0 00 0 0 0 0
100 100 100 100 100
cells were incubated with the peptide for 1 h at 37 �C and its lysis was measured bye determined in PBS and 0.1% Triton X-100, respectively. The hemolysis percentageinations were performed in duplicate and are the average of three independent
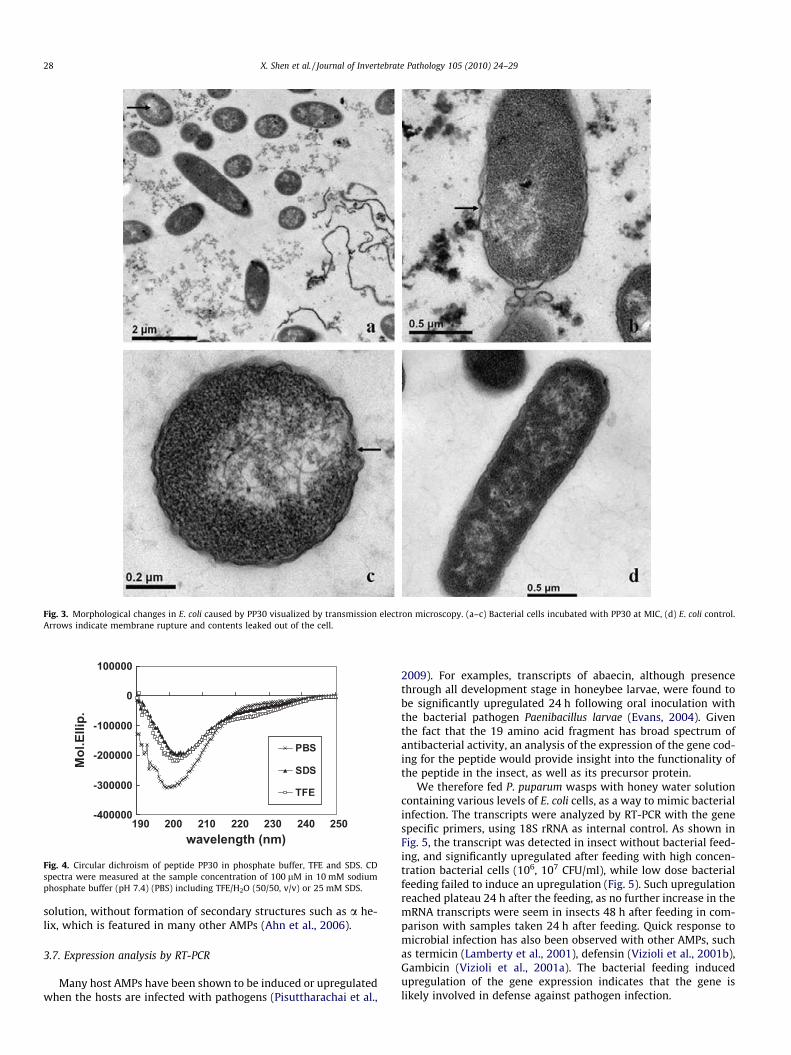
Fig. 3. Morphological changes in E. coli caused by PP30 visualized by transmission electron microscopy. (a–c) Bacterial cells incubated with PP30 at MIC, (d) E. coli control.Arrows indicate membrane rupture and contents leaked out of the cell.
-400000
-300000
-200000
-100000
0
100000
190 200 210 220 230 240 250wavelength (nm)
Mol
.Elli
p.
PBS
SDS
TFE
Fig. 4. Circular dichroism of peptide PP30 in phosphate buffer, TFE and SDS. CDspectra were measured at the sample concentration of 100 lM in 10 mM sodiumphosphate buffer (pH 7.4) (PBS) including TFE/H2O (50/50, v/v) or 25 mM SDS.
28 X. Shen et al. / Journal of Invertebrate Pathology 105 (2010) 24–29
solution, without formation of secondary structures such as a he-lix, which is featured in many other AMPs (Ahn et al., 2006).
3.7. Expression analysis by RT-PCR
Many host AMPs have been shown to be induced or upregulatedwhen the hosts are infected with pathogens (Pisuttharachai et al.,
2009). For examples, transcripts of abaecin, although presencethrough all development stage in honeybee larvae, were found tobe significantly upregulated 24 h following oral inoculation withthe bacterial pathogen Paenibacillus larvae (Evans, 2004). Giventhe fact that the 19 amino acid fragment has broad spectrum ofantibacterial activity, an analysis of the expression of the gene cod-ing for the peptide would provide insight into the functionality ofthe peptide in the insect, as well as its precursor protein.
We therefore fed P. puparum wasps with honey water solutioncontaining various levels of E. coli cells, as a way to mimic bacterialinfection. The transcripts were analyzed by RT-PCR with the genespecific primers, using 18S rRNA as internal control. As shown inFig. 5, the transcript was detected in insect without bacterial feed-ing, and significantly upregulated after feeding with high concen-tration bacterial cells (106, 107 CFU/ml), while low dose bacterialfeeding failed to induce an upregulation (Fig. 5). Such upregulationreached plateau 24 h after the feeding, as no further increase in themRNA transcripts were seem in insects 48 h after feeding in com-parison with samples taken 24 h after feeding. Quick response tomicrobial infection has also been observed with other AMPs, suchas termicin (Lamberty et al., 2001), defensin (Vizioli et al., 2001b),Gambicin (Vizioli et al., 2001a). The bacterial feeding inducedupregulation of the gene expression indicates that the gene islikely involved in defense against pathogen infection.

Fig. 5. Expression profiles of PP30 examined by RT-PCR. 18S rRNA was used as aninternal control. Lane 1 and 2: samples taken 24 and 48 h from non-E. coli treatment(control); Lane 3 and 4: samples taken 24 and 48 h after reared with honey watercontaining 104 CFU/ml E. coli; Lane 5 and 6: samples taken 24 and 48 h after rearedwith honey water containing 105 CFU/ml E.coli; Lane 7 and 8: samples taken 24 and48 h after reared with honey water containing 106 CFU/ml E.coli; Lane 9 and 10:samples taken 24 and 48 h after reared with honey water containing 107 CFU/mlE.coli.
X. Shen et al. / Journal of Invertebrate Pathology 105 (2010) 24–29 29
In conclusion, in an effort to identify antimicrobial peptides inthe endoparasitoid wasp P. puparum, we screened an expressioncDNA library from the wasp and isolated a number of cDNA clonesthat inhibited growth of the host E. coli cells. One clone that con-tains an abaecin-like peptide fragment was further studied usingthe synthetic peptide. We demonstrated that the peptide is activeagainst both Gram-negative and positive bacteria with low cyto-toxicity to human erythrocytes. It did not form secondary structurein aqueous solutions. The potency of the peptide reduced as saltsincreased in the culture medium and could cause intensivedamage, including rupture of cell membranes of bacterial cellstreated with the peptide. The expression of gene coding for thepeptide is constitutive and upregulated upon feeding of the insectwith bacteria, indicating its involvement in pathogen defensemechanism. In addition, some topics such as the site and temporalpattern of the peptide synthesis during the life cycle of the waspand the sex dependence of the peptide synthesis remain to be de-fined in future.
Acknowledgments
This work was supported by program for 973 Program(2006CB102005), NSFC (Grant No. 30571251), NCET (NCET-05-0513), and the Innovation Research Team Program of the Ministryof Education of China (Grant No. IRT0535).
References
Ahn, H.S., Cho, W., Kang, S.H., Ko, S.S., Park, M.S., Cho, H., Lee, K.H., 2006. Design andsynthesis of novel antimicrobial peptides on the basis of a helical domain oftenecin 1, an insect defensin protein, and structure–activity relationship study.Peptides 27, 640–648.
Angus, D.C., Linde-Zwirble, W.T., Lidicker, J., 2001. Epidemiology of severe sepsis inthe United States: analysis of incidence, outcome, and associated costs of care.Crit. Care Med. 29, 1303–1310.
Bartholomay, L.C., Cho, W.L., Rocheleau, T.A., Boyle, J.P., Beck, E.T., Fuchs, J.F., Liss, P.,Rusch, M., Butler, K.M., Wu, R.C.C., Lin, S.P., Kuo, H.Y., Tsao, I.Y., Huang, C.Y., Liu,T.T., Hsiao, K.J., Tsai, S.F., Yang, U.C., Nappi, A.J., Perna, N.T., Chen, C.C., Christen-sen, B.M., 2004. Description of the transcriptomes of immune-activatedhemocytes from the mosquito vectors Aedes aegypti and Armigeressubalbatus. Infect. Immun. 72, 4114–4126.
Cai, J., Ye, G.Y., Hu, C., 2004. Parasitism of Pieris rapae (Lepidoptera: Pieridae) by apupal endoparasitoid, Pteromalus puparum (Hymenoptera: Pteromalidae):effects of parasitization and venom on host hemocytes. J. Insect Physiol. 50,315–322.
Casteels, P., Ampe, C., Riviere, L., Van Damme, J., Elicone, C., Fleming, M., Jacobs, F.,Tempst, P., 1990. Isolation and characterization of abaecin, a majorantimicrobial response peptide in the honeybee (Apis mellifera). Eur. J.Biochem. 187, 381–386.
Cerovsky, V., Slaninová, J., Fucír, V., Hulacová, H., Borovicková, L., Jezek, R.,Bednárová, L., 2008. New potent antimicrobial peptides from the venom ofPolistinae wasps and their analogs. Peptides 99, 992–1003.
Cheng, T., Zhao, P., Liu, C., Xu, P., Gao, Z., Xia, Q., Xiang, Z., 2006. Structures,regulatory regions, and inductive expression patterns of antimicrobial peptidegenes in the silkworm Bombyx mori. Genomics 87, 356–365.
Cheng, X.Y., Liu, G.Z., Ye, G.Y., Wang, H.J., Shen, X.J., Wu, K.C., Xie, J.H., Altosaar, I.,2009. Screening and cloning of antimicrobial DNA sequences using a vitalstaining method. Gene 430, 132–139.
Cytrynska, M., Mak, P., Zdybicka-Barabas, A., Suder, P., Jakubowicz, T., 2007.Purification and characterization of eight peptides from Galleria mellonellaimmune hemolymph. Peptides 28, 533–546.
Dathe, M., Wieprecht, T., Nikolenko, H., Handel, L., Maloy, W.L., MacDonald, D.L.,Beyermann, M., Bienert, M., 1997. Hydrophobicity, hydrophobic moment andangle subtended by charged residues modulate antibacterial and haemolyticactivity of amphipathic helical peptides. FEBS Lett. 403, 208–212.
Epand, R.M., Vogel, H.J., 1999. Diversity of antimicrobial peptides and theirmechanisms of action. Biochim. Biophys. Acta 462, 11–28.
Evans, J.D., 2004. Transcriptional immune responses by honey bee larvae duringinvasion by the bacterial pathogen, Paenibacillus larvae. J. Invertebr. Pathol. 85,105–111.
Irving, P., Ubeda, J.M., Doucet, D., Troxler, L., Lagueux, M., Zachary, D., Hoffmann, J.A.,Hetru, C., Meister, M., 2005. New insights into Drosophila larval haemocytefunctions through genome-wide analysis. Cell Microbiol. 7, 335–350.
Kuhn-Nentwig, L., 2003. Antimicrobial and cytolytic peptides of venomousarthropods. Cell Mol. Life Sci. 60 (12), 2651–2668.
Lamberty, M., Zachary, D., Lanot, R., Bordereau, C., Robert, A., Hoffmann, J.A., Bulet,P., 2001. Insect immunity. Constitutive expression of a cysteine-rich antifungaland a linear antibacterial peptide in a termite insect. J. Biol. Chem. 276, 4085–4092.
Matsuzaki, K., 1999. Why and how are peptide–lipid interactions utilized for self-defense? Magainins and tachyplesins as archetypes. Biochim. Biophys. Acta1462, 1–10.
Moore, M.A., Hakki, Z.W., Gregory, R.L., Gfell, L.E., Kim-Park, W.K., Kowolik, M.J.,1997. Influence of heat inactivation of human serum on the opsonization ofStreptococcus mutans. Ann. N. Y. Acad. Sci. 832, 383–393.
Otvos Jr., L., 2002. The short proline-rich antibacterial peptide family. Cell. Mol. LifeSci. 59, 1138–1150.
Pisuttharachai, D., Yasuike, M., Aono, H., Yano, Y., Murakami, K., Kondo, H., Aoki, T.,Hirono, I., 2009. Characterization of tow isoforms of Japanese spiny lobsterPanulirus japonicus defensin cDNA. Dev. Comp. Immunol. 33 (4), 434–438.
Ryge, T.S., Hansen, P.R., 2005. Novel lysine-peptoid hybrids with antibacterialproperties. J. Peptide Sci. 11, 727–734.
Shai, Y., 2002. Mode of action of membrane active antimicrobial peptides.Biopolymers 66, 236–248.
Shamova, O., Brogden, K.A., Zhao, C., Nguyen, T., Kokryakov, V.N., Lehrer, R.I., 1999.Purification and properties of proline-rich antimicrobial peptides from sheepand goat leukocytes. Infect. Immun. 67, 4106–4111.
Shen, X.J., Ye, G.Y., Cheng, X.Y., Yu, C.Y., Yao, H.W., Hu, C., 2010. Novel antimicrobialpeptides identified from an endoparasitic wasp cDNA library. J. Pept. Sci. 16,58–64.
Steiner, H., Hultmark, D., Engstrom, A., Boman, H.G., 1981. Sequence and specificityof two antibacterial proteins involved in insect immunity. Nature 292, 246–248.
Tossi, A., Sandri, L., Giangaspero, A., 2000. Amphipathic, a-helical antimicrobialpeptides. Biopolymers 55, 4–30.
Tsubery, H., Ofek, I., Cohen, S., Fridkin, M., 2000. Structure–function studies ofpolymyxin B nonapeptide: implications to sensitization of Gram-negativebacteria. J. Med. Chem. 43, 23085–23092.
Tzou, P., De Gregorio, E., Lemaitre, B., 2002. How Drosophila combats microbialinfection: a model to study innate immunity and host–pathogen interactions.Curr. Opin. Microbiol. 5, 102–110.
Tzou, P., Ohresser, S., Ferrandon, D., Capovilla, M., Reichhart, J.M., Lemaitre, B.,Hoffmann, J.A., Imler, J.L., 2000. Tissue-specific inducible expression ofantimicrobial peptide genes in Drosophila surface epithelia. Immunity 13,737–748.
Viljakainen, L., Evans, J.D., Hasselmann, M., Rueppell, O., Tingek, S., Pamilo, P., 2009.Rapid evolution of immune proteins in social insects. Mol. Biol. Evol. 26 (8),1791–1801.
Vizioli, J., Bulet, P., Hoffmann, J.A., Kafatos, F.C., Müller, H.M., Dimopoulos, G., 2001a.Gambicin: a novel immune responsive antimicrobial peptide from the malariavector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 98, 12630–12635.
Vizioli, J., Richman, A.M., Uttenweiler-Joseph, S., Blass, C., Bulet, P., 2001b. Thedefensin peptide of the malaria vector mosquito Anopheles gambiae:antimicrobial activities and expression in adult mosquitoes. Insect. Biochem.Mol. Biol. 31, 241–248.
Wang, H., Dong, S.Z., Li, K., Hu, C., Ye, G.Y., 2008. A heat shock cognate 70 gene in theendoparasitoid, Pteromalus puparum, and its expression in relation to thethermal stress. BMB Reports 41 (5), 388–393.
Wu, M.L., Ye, G.Y., Zhu, J.Y., Chen, X.X., Hu, C., 2008. Isolation and characterization ofan immunosuppressive protein from venom of the pupa-specific endoparasitoidPteromalus puparum. J. Invertebr. Pathol. 99, 186–191.
Xu, P., Shi, M., Chen, X.X., 2009. Antimicrobial peptide evolution in the Asiatic honeybee Apis cerana. PLoS One 4 (1), e4239.
Zasloff, M., 2002. Antimicrobial peptides of multicellular organisms. Nature 415,389–395.
Zhang, Z., Ye, G.Y., Cai, J., Hu, C., 2005. Comparative venom toxicity betweenPteromalus puparum and Nasonia vitripennis (Hymenoptera: Pteromalidae)toward the hemocytes of their natural hosts, non-target insects and culturedinsect cells. Toxicon 46, 337–349.