chapter 7 operons: fine control of bacterial transcription student learning outcomes : explain basic...
TRANSCRIPT

Chapter 7 Operons: Fine Control ofBacterial Transcription
Student learning outcomes:• Explain basic features of regulation of operons:
classic catabolic lac and ara; anabolic system trp.
role of repressors, operators, positive controls• Describe briefly role or riboswitches in regulation
7-1
Lac repressor (pink) and CAP-cAMP (blue) binding lac operator/promoter region

Bacterial transcription
• Regulation of gene expression at transcriptional level is very important for prokaryotes:
- conserves energy, permits rapid response
- mRNA in prokaryotes is short-lived
- regulation involves protein-ligand and protein-DNA
interactions primarily
• Important Figs: 1, 2, 3*, 5*, 6, 7, 10, 11, 12, 13, 14, 15*, 16, 18, 19, 20, 21*, 26, 27*, 28, 29, 31, 32, 34
• Review Q: 2, 3, 4, 5, 10, 11, 13, 16, 19, 21 • Analyt Q: 1, 2, 3, 4 7-2
7-3
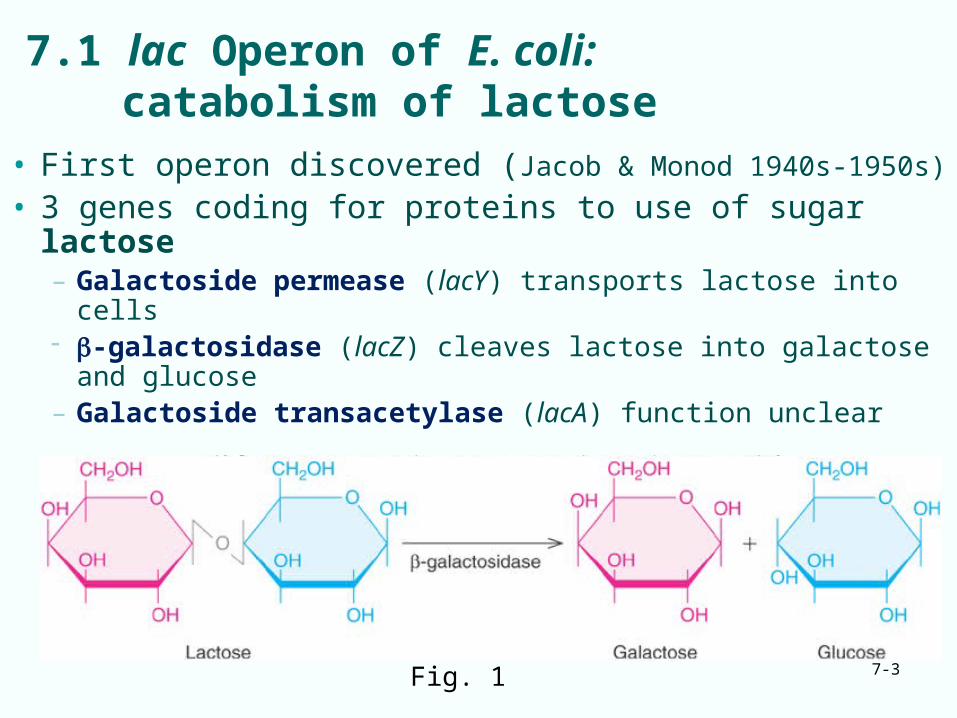
7.1 lac Operon of E. coli:catabolism of lactose
• First operon discovered (Jacob & Monod 1940s-1950s)
• 3 genes coding for proteins to use of sugar lactose– Galactoside permease (lacY) transports lactose into cells -galactosidase (lacZ) cleaves lactose into galactose and
glucose– Galactoside transacetylase (lacA) function unclear
Fig. 1
7-4
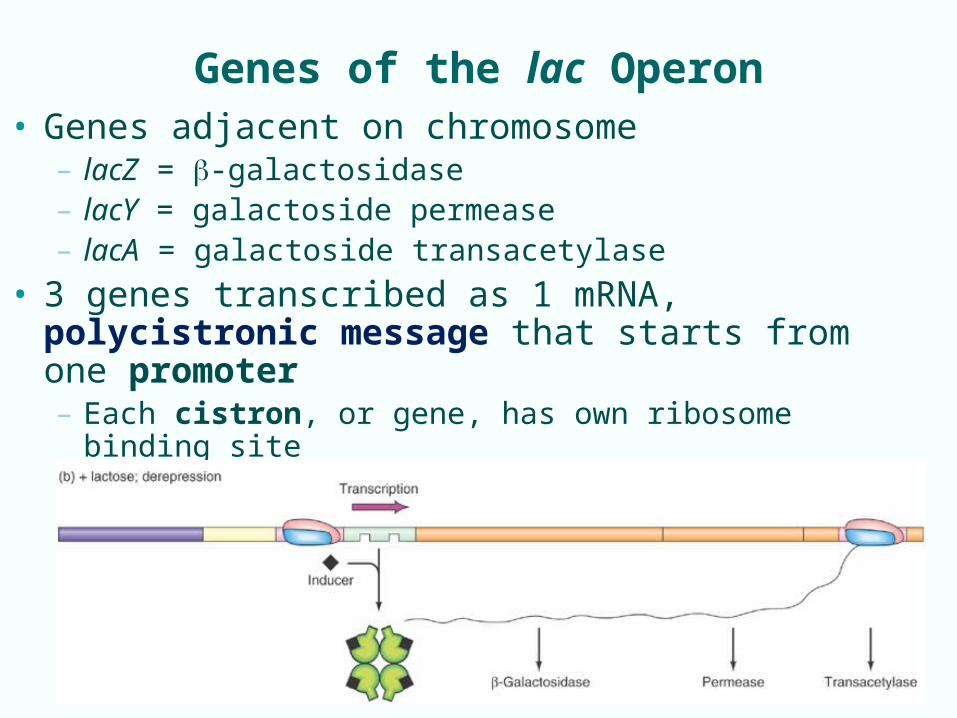
Genes of the lac Operon• Genes adjacent on chromosome
– lacZ = -galactosidase– lacY = galactoside permease– lacA = galactoside transacetylase
• 3 genes transcribed as 1 mRNA, polycistronic message that starts from one promoter– Each cistron, or gene, has own ribosome binding site– Each cistron can be translated by separate ribosomes

7-5
Control of lac Operon
• Glucose preferred substrate for E. coli; only make lac enzymes if need to use lactose as carbon source
• lac operon is tightly controlled, 2 types of control:– Negative control for catabolic systems:– ‘transcription is off unless turn on’; – inducer must remove repressor from operator – Activator, positive factor CAP protein:– responds to low glucose (energy) by stimulating
transcription of lac operon

7-6
Negative Control of the lac Operon
• ‘Off until needed to turn on’• Off-regulation by lac repressor:
– Product of lacI gene– Tetramer of 4 identical polypeptides– Binds operator just upstream of promoter
• When repressor binds operator, operon is repressed– Repressor bound to operator prevents RNA polymerase
from initiating at promoter
• lac operon is repressed as long as no lactose is available or needed (i.e. glucose available)
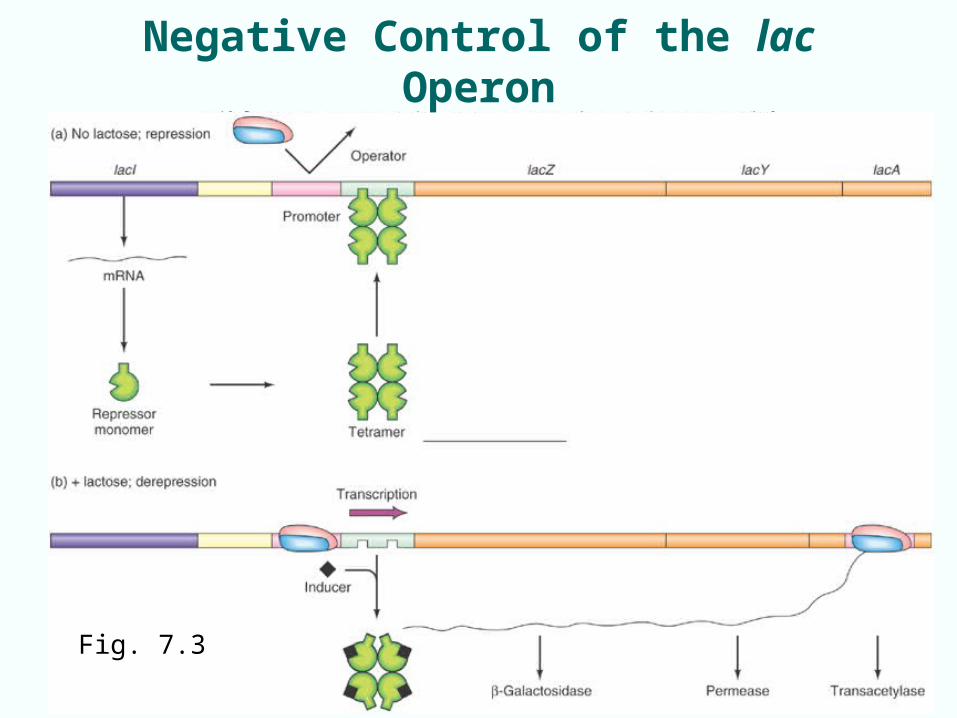
7-7Fig. 7.3
Negative Control of the lac Operon
7-8
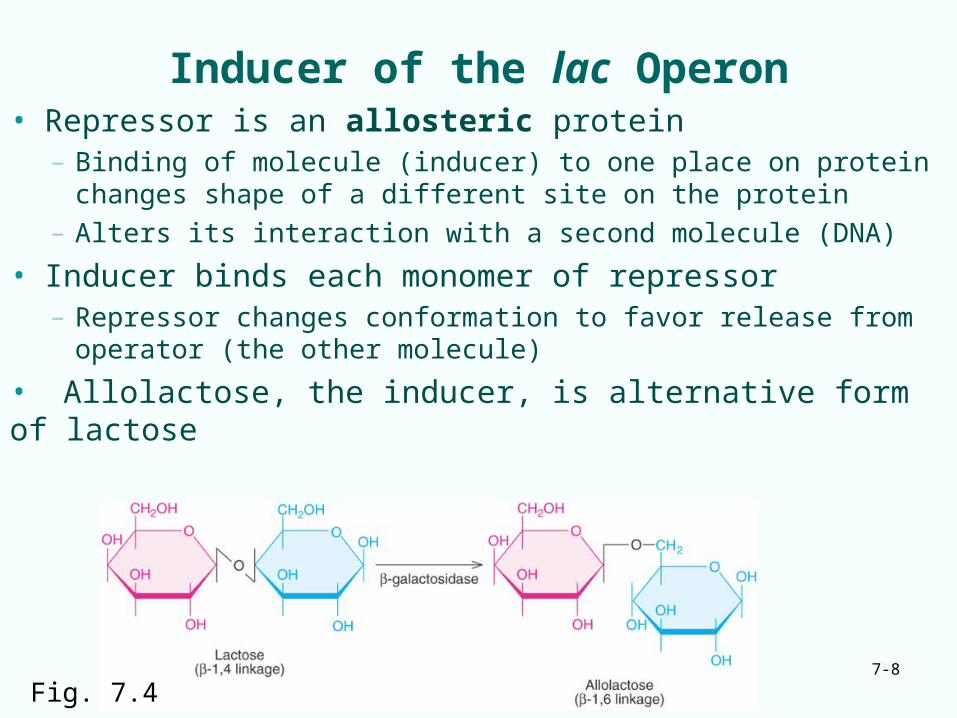
Inducer of the lac Operon• Repressor is an allosteric protein
– Binding of molecule (inducer) to one place on protein changes shape of a different site on the protein
– Alters its interaction with a second molecule (DNA)
• Inducer binds each monomer of repressor– Repressor changes conformation to favor release from
operator (the other molecule)
• Allolactose, the inducer, is alternative form of lactose
Fig. 7.4

7-9
Discovery of the lac Operon1940s -1950s, Jacob & Monod studied metabolism of lactose by E. coli: biochemistry, genetics: cis/trans tests, antibodies to -galactosidase, synthetic non-cleavable inducers
• Three enzyme activities / three genes were induced together by galactosides• Constitutive mutants need no induction; genes are active all the time• Merodiploids, partial diploid bacteria, constructed by conjugation of F’lac into F- lac cells, were critical to analysis of mutants, to decipher cis/trans relationships to define proteins, DNA sites

7-10
Effects of Regulatory Mutations:Wild-type and Mutated Repressors
Fig. 7.5 Lac I- mutations are recessive; repressor can act in trans to affect genes on both pieces of DNA

7-11
Effects of Regulatory Mutations:Wild-type and Mutated Operators (with Defective Binding)
Fig. 7.5 Lac Oc (constitutive) mutation is cis-dominant; affects only gene adjacent to mutant operator
7-12
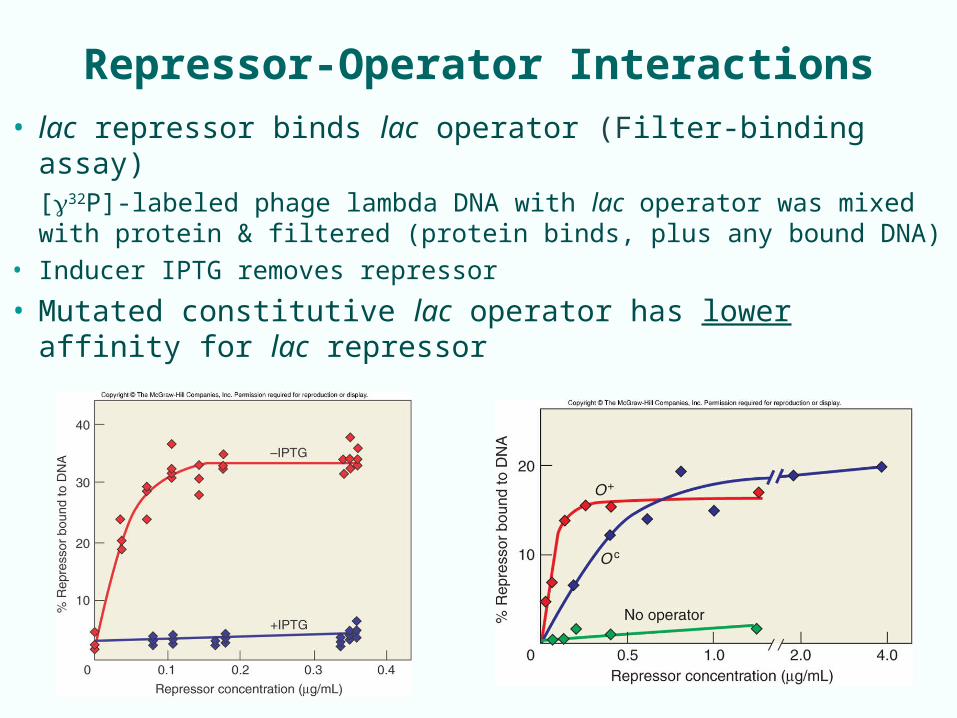
Repressor-Operator Interactions
• lac repressor binds lac operator (Filter-binding assay)[32P]-labeled phage lambda DNA with lac operator was mixed with protein & filtered (protein binds, plus any bound DNA)
• Inducer IPTG removes repressor
• Mutated constitutive lac operator has lower affinity for lac repressor
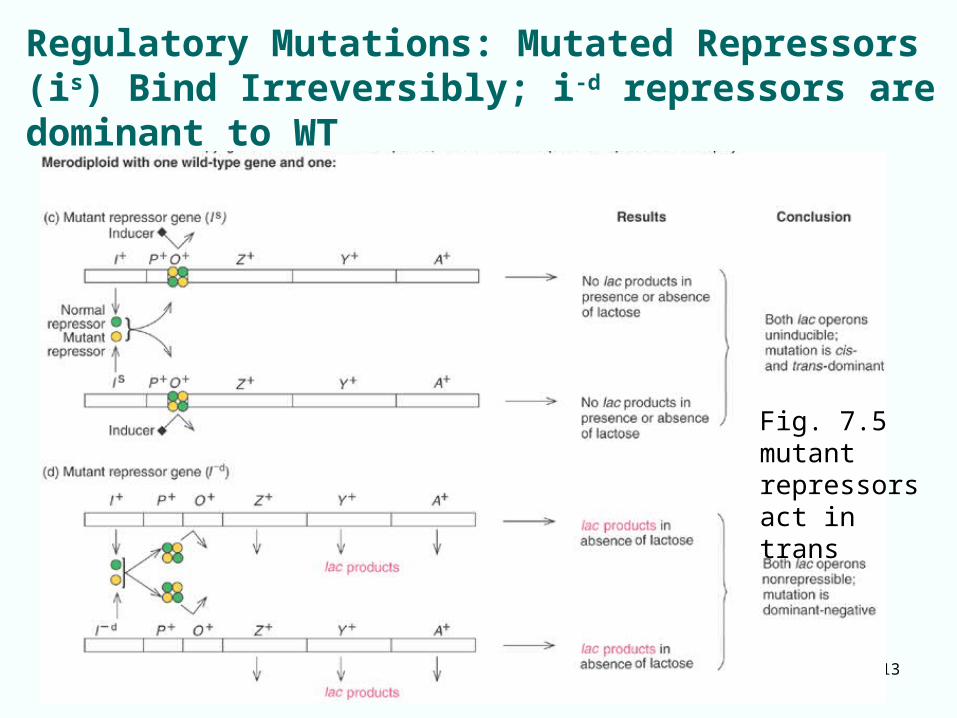
7-13
Regulatory Mutations: Mutated Repressors (is) Bind Irreversibly; i-d repressors are dominant to WT
Fig. 7.5 mutant repressors act in trans

7-14
Mechanism of lac Repression still unclear
• Initial binding in vitro: repressor does not block access by RNAP to lac promoter
• RNAP and repressor bind together to lac promoter• Polymerase-promoter complex is in equilibrium with
free polymerase and promoter
• Two hypotheses remain:– RNAP can bind lac promoter in presence of repressor
• Repressor inhibits transition from abortive transcription to processive transcription
– Repressor, by binding to operator, blocks access by RNAP to adjacent promoter
7-15
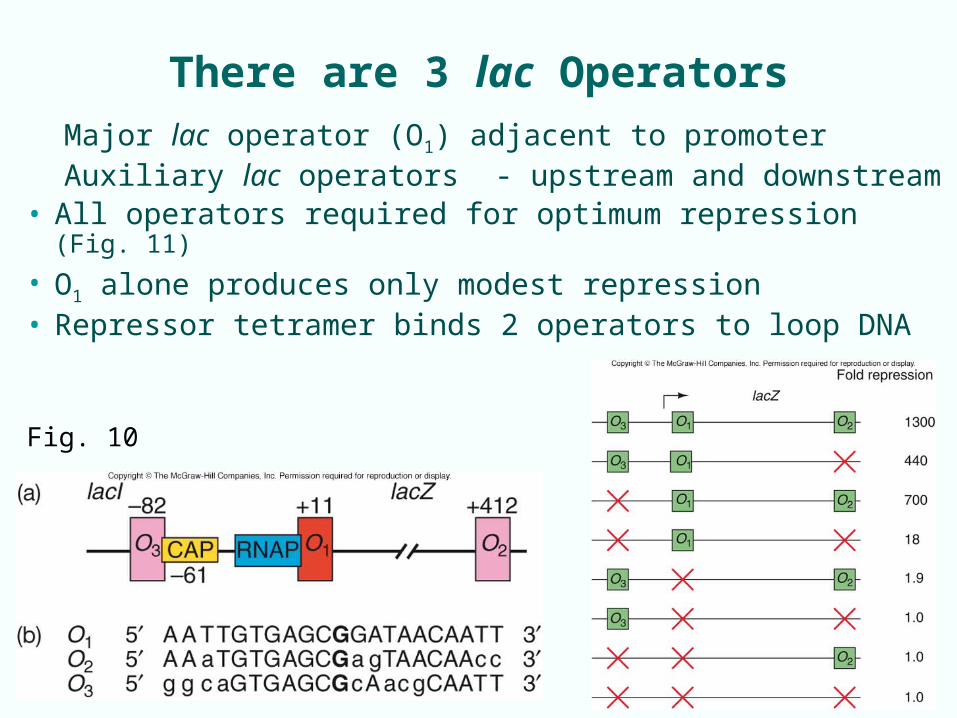
There are 3 lac Operators
Major lac operator (O1) adjacent to promoterAuxiliary lac operators - upstream and downstream
• All operators required for optimum repression (Fig. 11)
• O1 alone produces only modest repression• Repressor tetramer binds 2 operators to loop DNA
Fig. 10
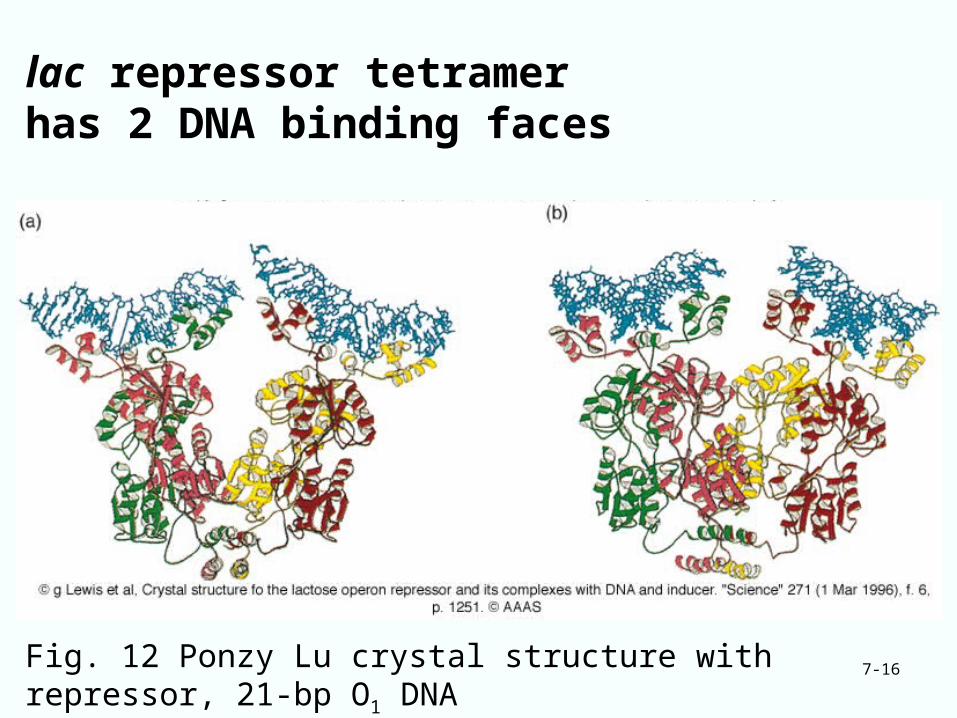
7-16
lac repressor tetramer has 2 DNA binding faces
Fig. 12 Ponzy Lu crystal structure with repressor, 21-bp O1 DNA
7-17
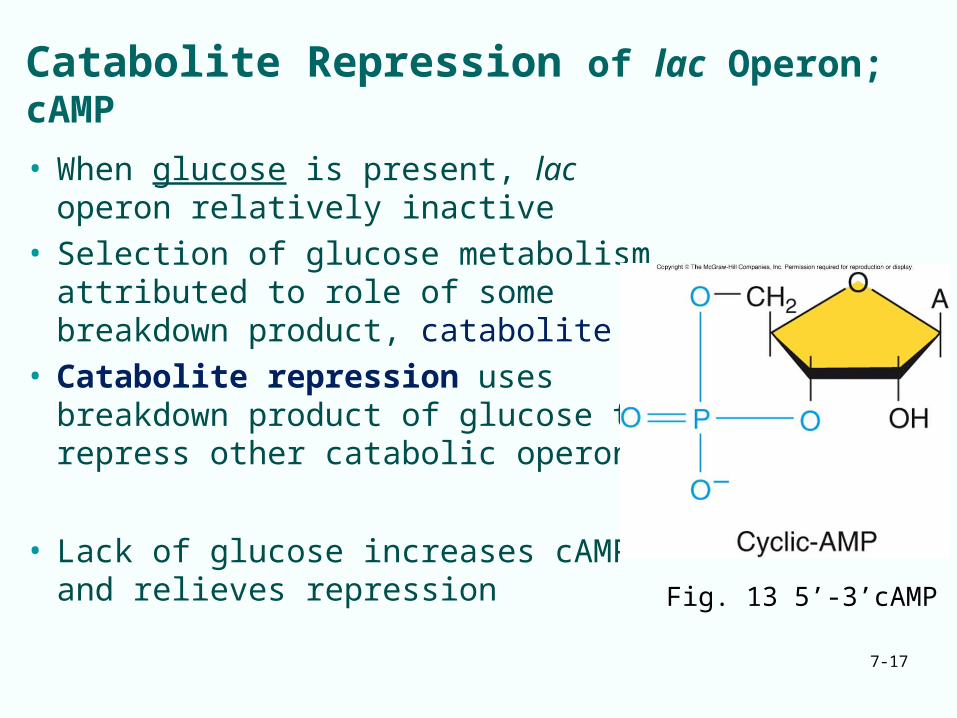
Catabolite Repression of lac Operon; cAMP
• When glucose is present, lac operon relatively inactive
• Selection of glucose metabolism attributed to role of some breakdown product, catabolite
• Catabolite repression uses breakdown product of glucose to repress other catabolic operons
• Lack of glucose increases cAMP and relieves repression
Fig. 13 5’-3’cAMP

7-18
Catabolite Activator Protein - CAP
• cAMP added to E. coli cultures overcomes catabolite repression of lac operon:– activates lac genes even in presence of glucose
• Positive controller of lac operon has 2 parts:– cAMP– Protein factor known as:
• Catabolite activator protein or CAP• Cyclic-AMP receptor protein or CRP• Gene encoding this protein is crp
7-19
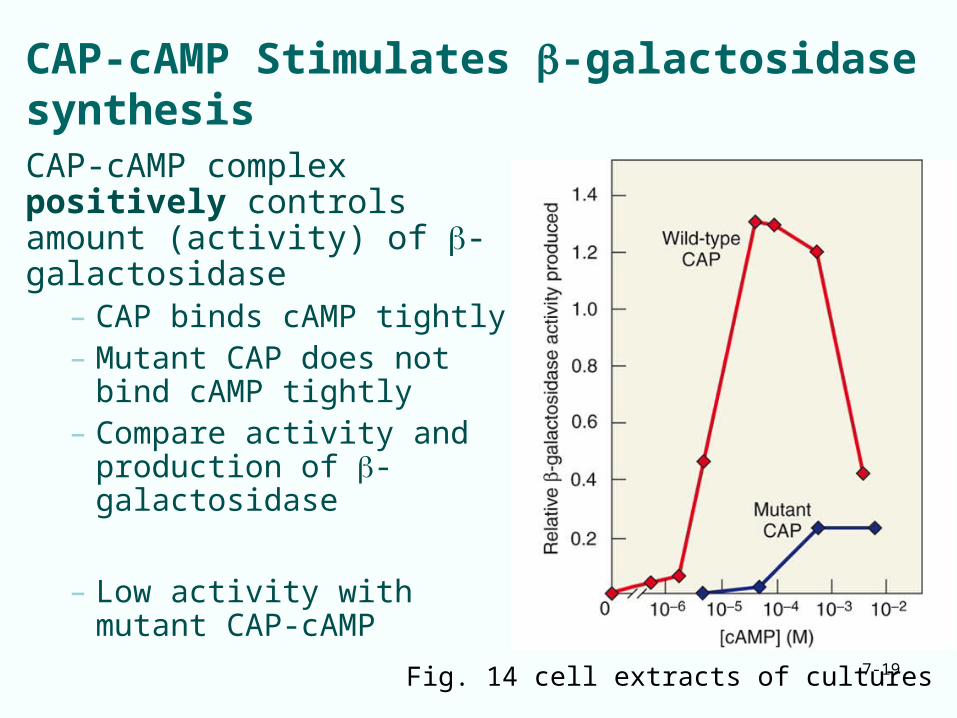
CAP-cAMP Stimulates -galactosidase synthesisCAP-cAMP complex positively controls amount (activity) of -galactosidase
– CAP binds cAMP tightly– Mutant CAP does not bind
cAMP tightly– Compare activity and
production of -galactosidase
– Low activity with mutant CAP-cAMP
Fig. 14 cell extracts of cultures

7-20
Mechanism of CAP Action• CAP-cAMP complex binds lac promoter tightly
– Mutants whose lac gene not stimulated by cAMP had mutations just upstream of lac promoter
– Ex. L1 deletion mutant
• Binding of CAP and cAMP to activator site helps RNAP form open promoter complex (RPo)
• CAP-cAMP activated operons have weak promoters– The -35 boxes are unlike consensus sequence– If promoters were strong, could be activated even when
glucose is present
Fig. 7.16
7-21
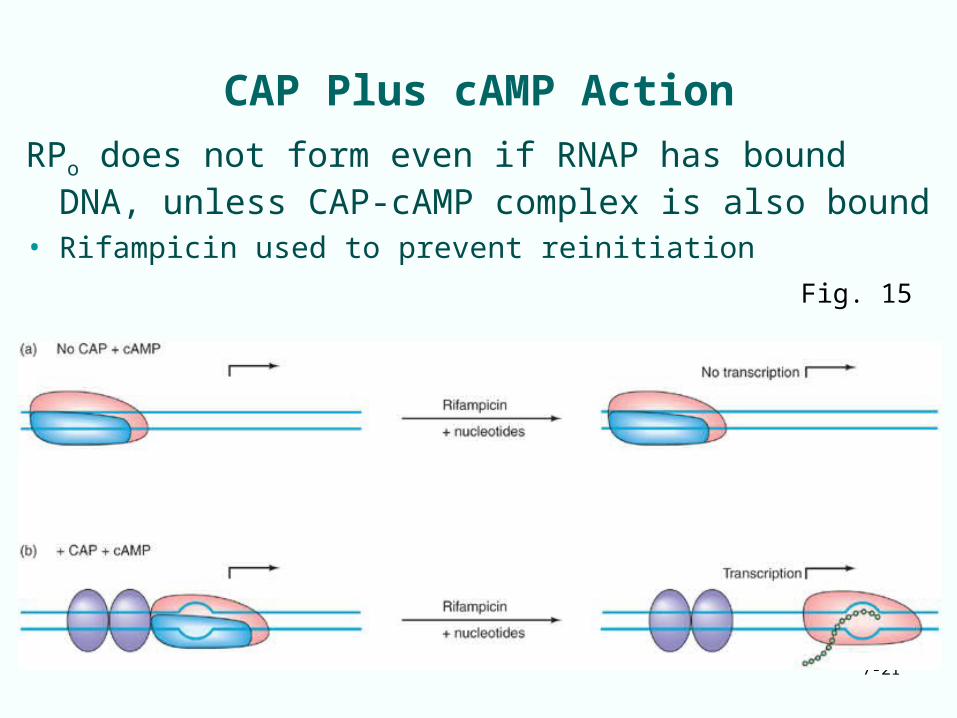
CAP Plus cAMP Action
RPo does not form even if RNAP has bound DNA, unless CAP-cAMP complex is also bound
• Rifampicin used to prevent reinitiationFig. 15

7-22
CAP-cAMP Recruits RNAP• Two steps:
– Formation of closed promoter complex RPo– Conversion of RPo into open promoter complex
• CAP-cAMP bends target DNA about 100° when binds– DNA red; CAP blue; cAMP thin pink; green
• CAP contacts subunit of RNAP (Fig. 17a)
okcKRPRPPR
B2
7-23
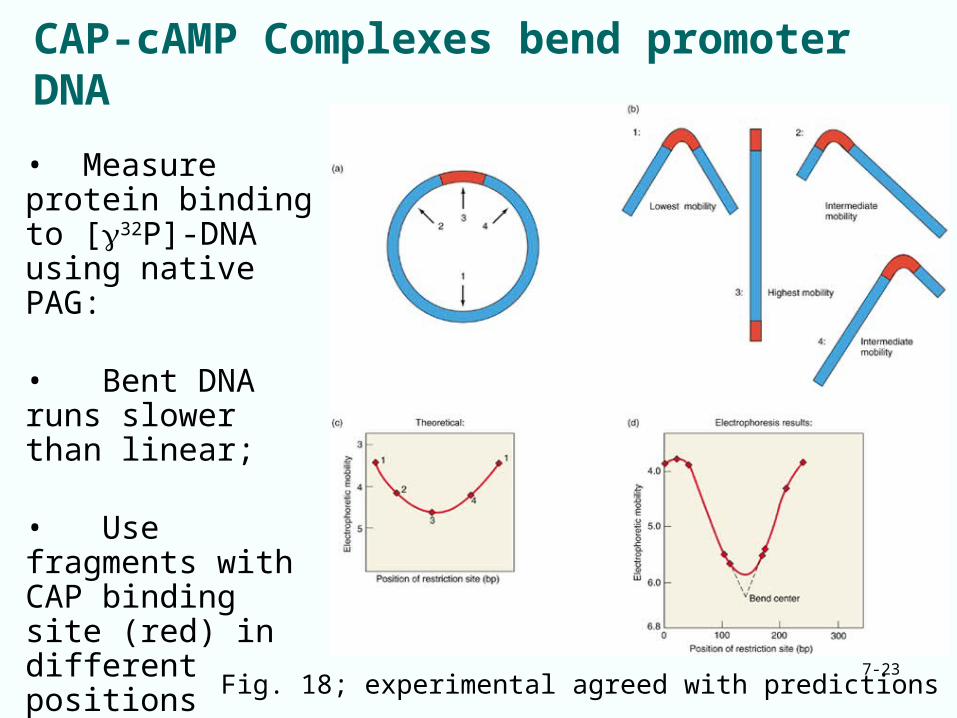
CAP-cAMP Complexes bend promoter DNA
Fig. 18; experimental agreed with predictions
• Measure protein binding to [32P]-DNA using native PAG:
• Bent DNA runs slower than linear;
• Use fragments with CAP binding site (red) in different positions relative to restriction enzyme sites• See which fragment runs slowest
7-24
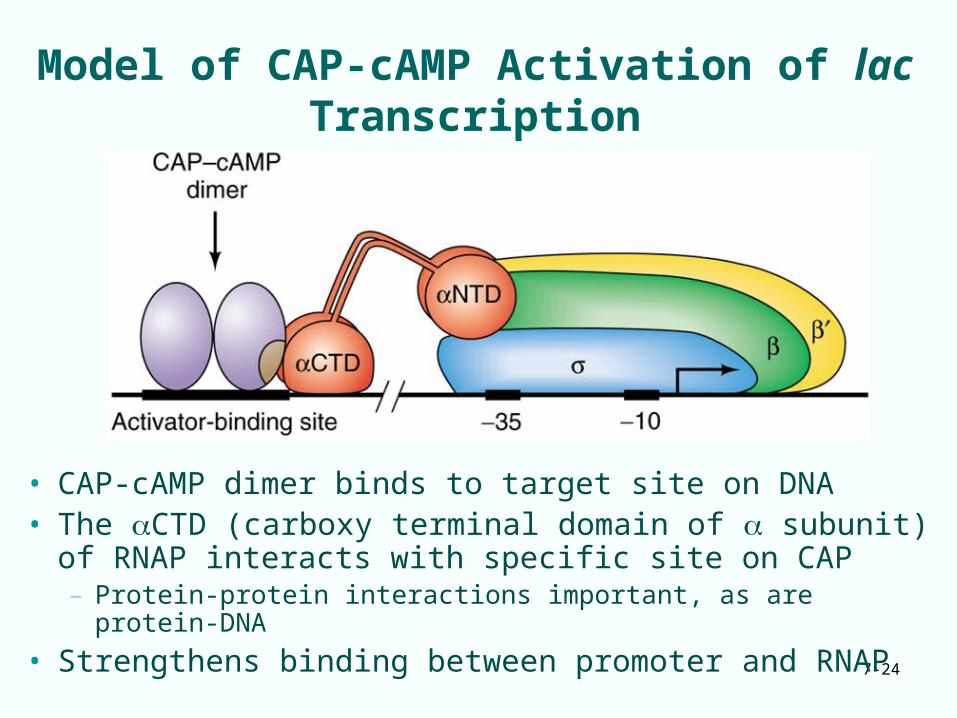
Model of CAP-cAMP Activation of lac Transcription
• CAP-cAMP dimer binds to target site on DNA• The CTD (carboxy terminal domain of subunit) of
RNAP interacts with specific site on CAP – Protein-protein interactions important, as are protein-DNA
• Strengthens binding between promoter and RNAP
7-25
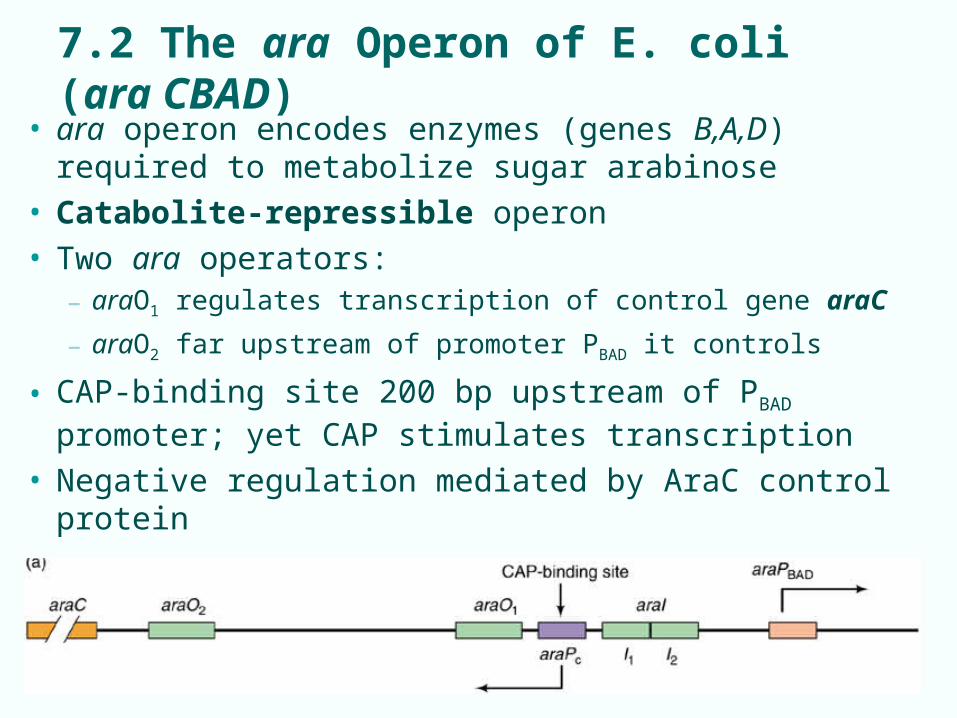
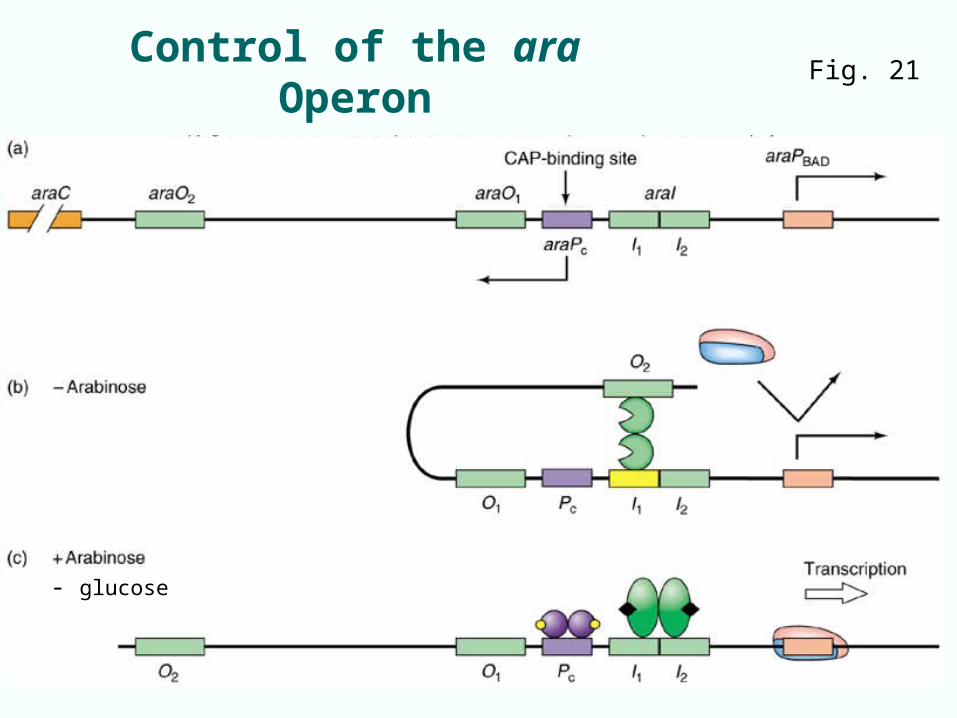
7.2 The ara Operon of E. coli (ara CBAD)
• ara operon encodes enzymes (genes B,A,D) required to metabolize sugar arabinose
• Catabolite-repressible operon• Two ara operators:
– araO1 regulates transcription of control gene araC
– araO2 far upstream of promoter PBAD it controls
• CAP-binding site 200 bp upstream of PBAD promoter; yet CAP stimulates transcription
• Negative regulation mediated by AraC control protein
7-26
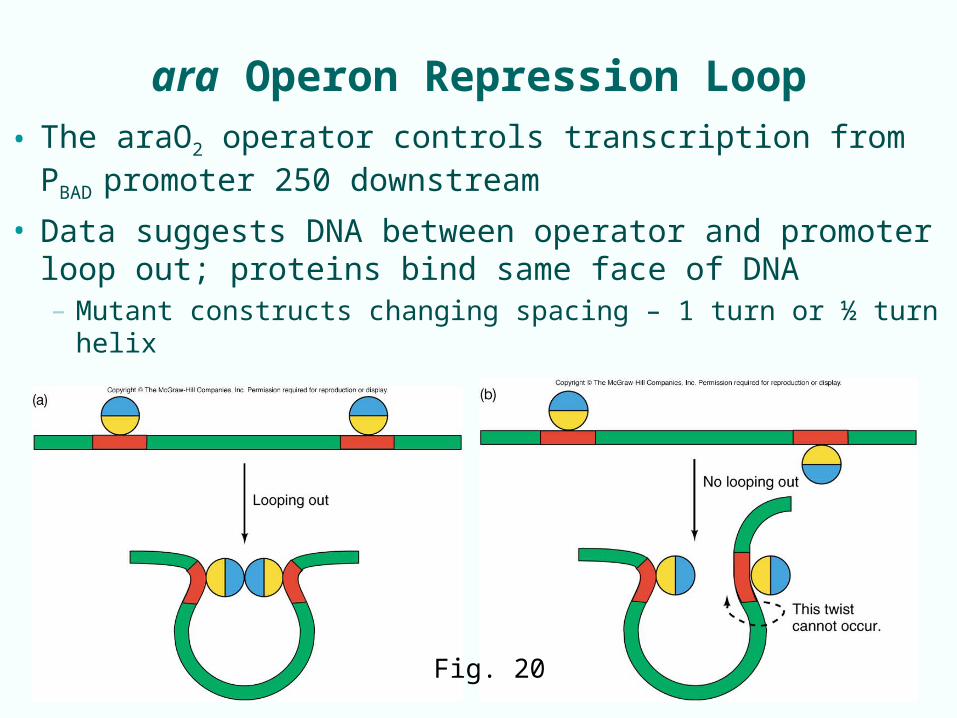
ara Operon Repression Loop
• The araO2 operator controls transcription from PBAD
promoter 250 downstream• Data suggests DNA between operator and promoter
loop out; proteins bind same face of DNA– Mutant constructs changing spacing – 1 turn or ½ turn helix
Fig. 20
7-27
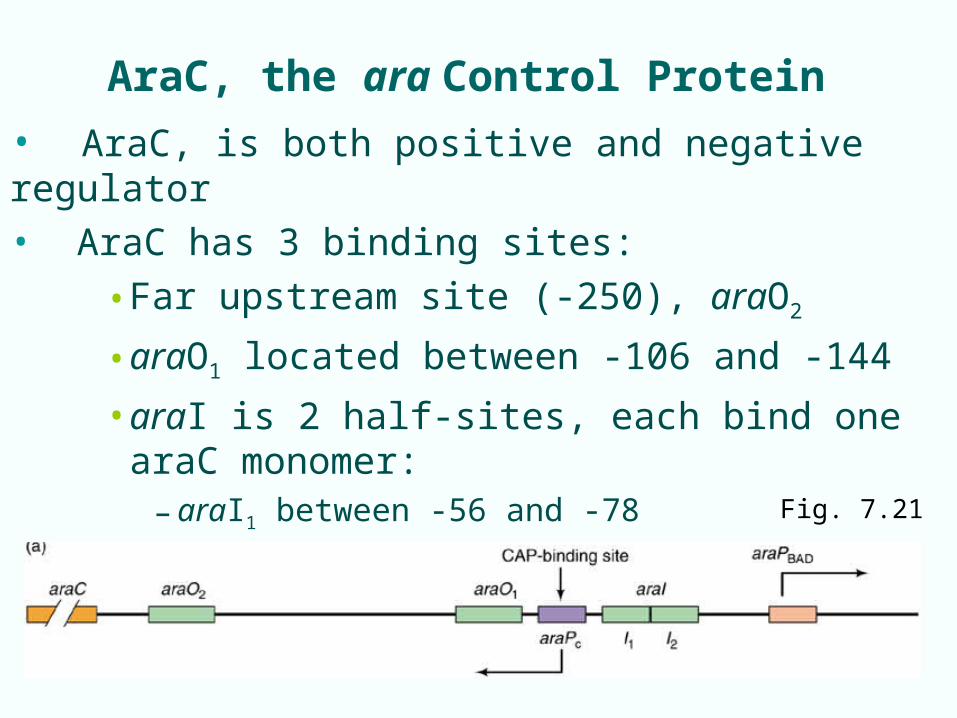
AraC, the ara Control Protein
• AraC, is both positive and negative regulator
• AraC has 3 binding sites:
• Far upstream site (-250), araO2
• araO1 located between -106 and -144
• araI is 2 half-sites, each bind one araC monomer:– araI1 between -56 and -78
– araI2 -35 to -51 Fig. 7.21
7-28
Control of the ara Operon Fig. 21
- glucose

AraC loops DNA by binding two operators
7-29
Add AraC to labeled minicircles with wt or mutant AraC binding sites -> looping makes more supercoiled and moves faster; At to, add excess unlabeled DNA of strong araI site;Electrophorese to see if still looped; mutants bind weaker
Fig. 22

7-30
Positive Control of the ara Operon
• Positive control is also mediated by CAP and cAMP• CAP-cAMP complex attaches to binding site Pc
upstream of araBAD promoter• DNA looping would allow CAP to contact subunit
of RNAP, stimulate its binding to the promoter
- glucose
Fig. 7.21

7-31
ara Operon Summary
• ara operon controlled by AraC protein– Represses by looping out DNA between araO2 and araI1
that are 210 bp apart
• Arabinose derepresses operon by binding AraC; AraC loosens attachment to araO2 and binds araI2
– Breaking loop allows transcription of operon from PBAD
• CAP-cAMP stimulate transcription(bind upstream araI)• AraC controls own synthesis by binding araO1 to
prevent leftward transcription of araC gene from Pc
Fig. 7.26

7-32
7.3 The trp Operon – anabolic - tryptophan
• E. coli trp operon contains 5 genes (E,D,C,B,A) for enzymes to make amino acid tryptophan (anabolic)
• Anabolic enzyme synthesis typically on, and turned off by high level of product of pathway (repressed)
• Operon subject to negative control of transcription by repressor when high tryptophan levels– The trp operator lies wholly within trp promoter– Tryptophan helps trp repressor bind operator
• trp operon also exhibits attenuation of transcription
7-33
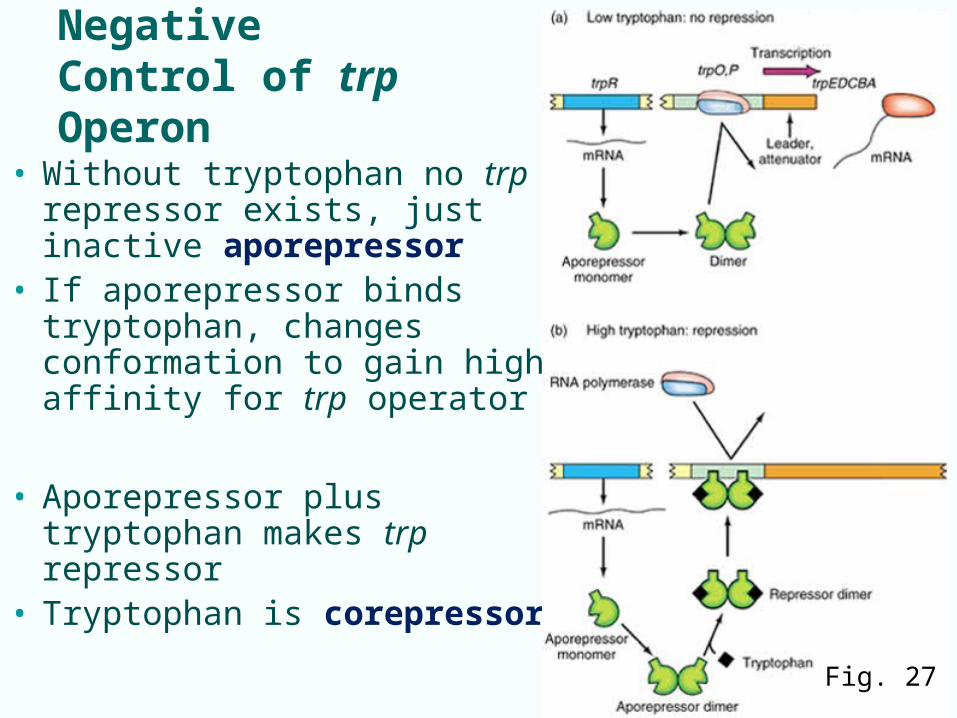
Negative Control of trp Operon
• Without tryptophan no trp repressor exists, just inactive aporepressor
• If aporepressor binds tryptophan, changes conformation to gain high affinity for trp operator
• Aporepressor plus tryptophan makes trp repressor
• Tryptophan is corepressor
Fig. 27

7-34
Attenuation of transcription in trp Operonextra level of control than just repressor-operator
Fig. 28: Attenuation is premature termination in 5’ UTR of trp E
7-35
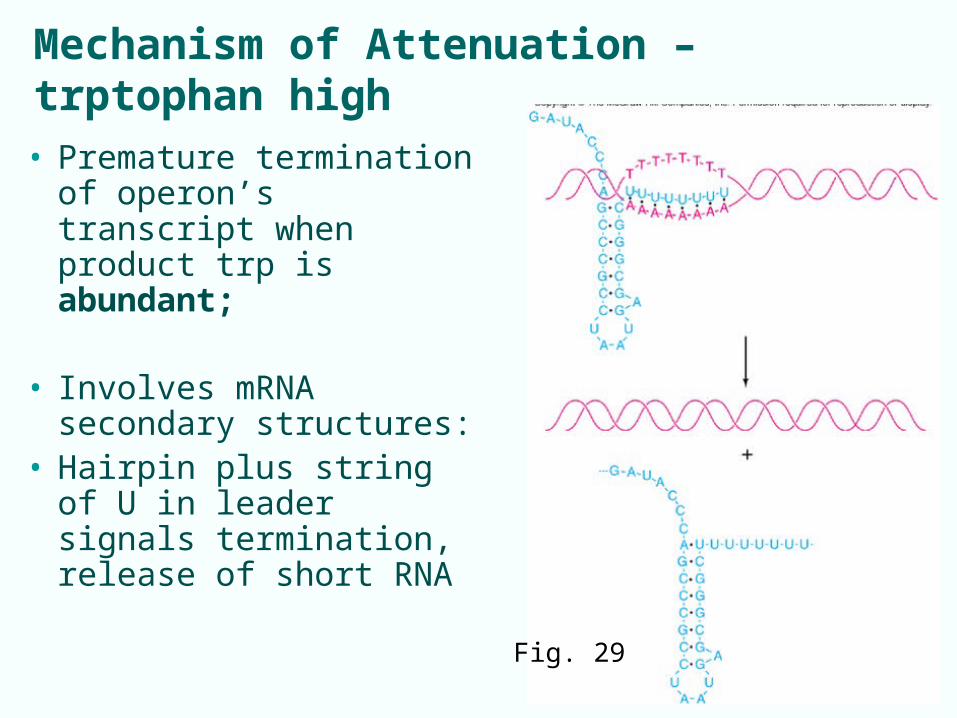
Mechanism of Attenuation – trptophan high
• Premature termination of operon’s transcript when product trp is abundant;
• Involves mRNA secondary structures:
• Hairpin plus string of U in leader signals termination, release of short RNA
Fig. 29

7-36
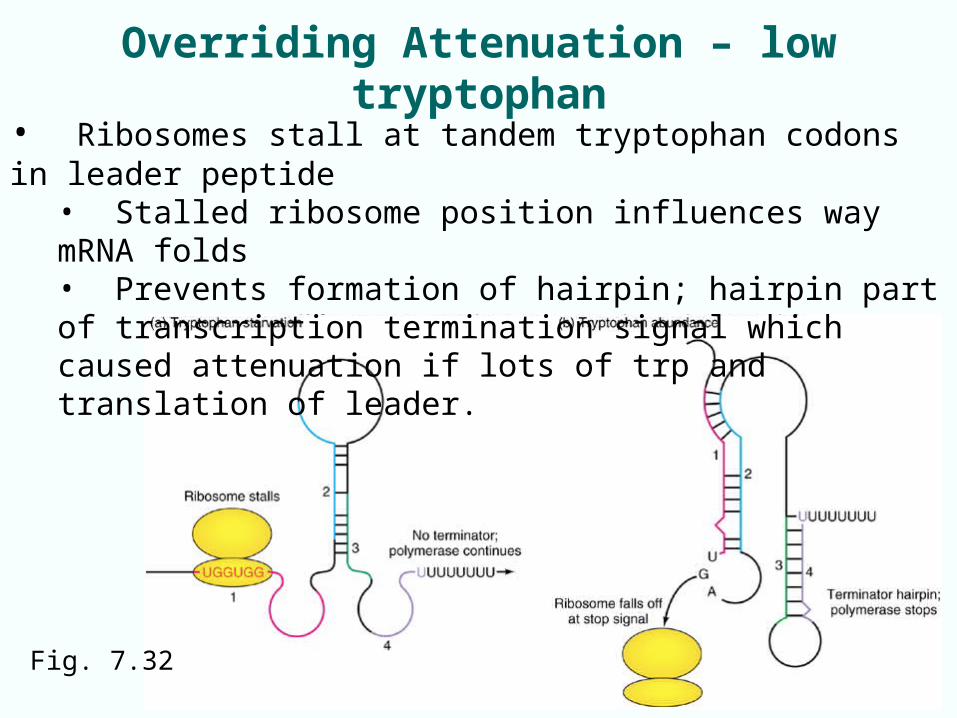
Defeating Attenuation – tryptophan low
• Prokaryotes start making protein before mRNA completely finished – only one cell compartment.
• If amino acid supply is low, ribosomes stall at tandem tryptophan codons in trp leader peptide– stalled protein synthesis -> – ribosome position influences mRNA folding– Prevents formation of hairpin (part of transcription
termination signal which causes attenuation)
Fig. 7.31 Trp leader has tandem trp codons
7-37
Overriding Attenuation – low tryptophan
• Ribosomes stall at tandem tryptophan codons in leader peptide• Stalled ribosome position influences way mRNA folds• Prevents formation of hairpin; hairpin part of transcription termination signal which caused attenuation if lots of trp and translation of leader.
Fig. 7.32

7-38
7.4 Riboswitches• Small molecules can act directly on 5’-UTRs of
mRNAs to control gene expression• Riboswitches –regions of 5’-UTRs that can alter
structure to control gene expression after bind ligand• Region that binds ligand is called aptamer• Expression platform is other module in riboswitch:
– Terminator – Ribosome-binding site– Another RNA element that affects gene expression
• Operates by depressing gene expression– Some work at transcriptional level– Others can function at translational level
7-39
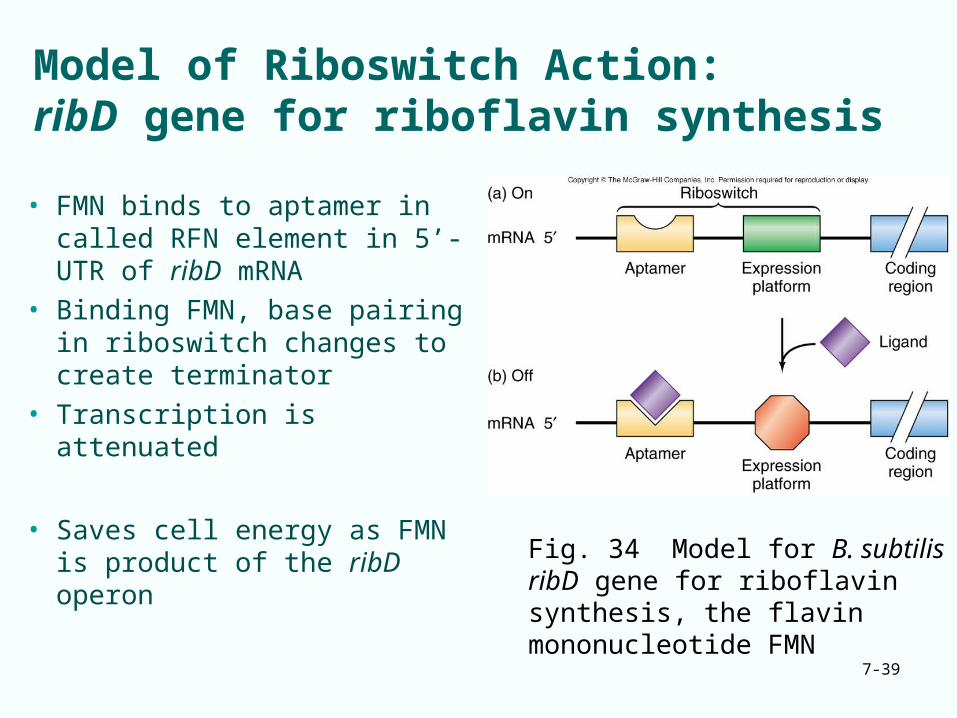
Model of Riboswitch Action:ribD gene for riboflavin synthesis
• FMN binds to aptamer in called RFN element in 5’-UTR of ribD mRNA
• Binding FMN, base pairing in riboswitch changes to create terminator
• Transcription is attenuated
• Saves cell energy as FMN is product of the ribD operon
Fig. 34 Model for B. subtilis ribD gene for riboflavin synthesis, the flavin mononucleotide FMN
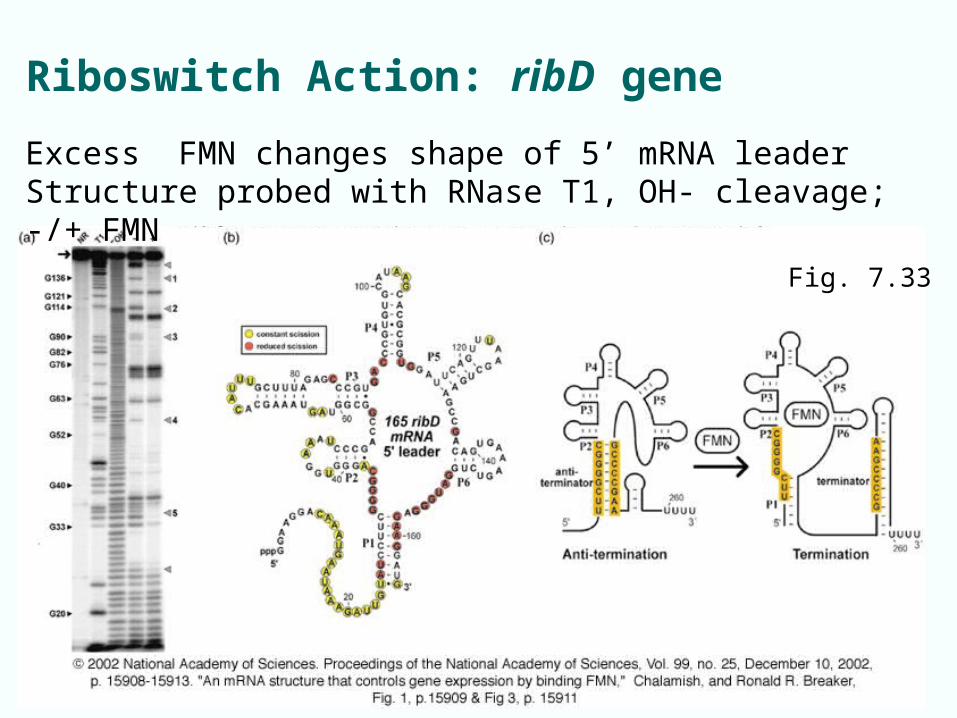
Riboswitch Action: ribD gene
7-40
Fig. 7.33
Excess FMN changes shape of 5’ mRNA leaderStructure probed with RNase T1, OH- cleavage; -/+ FMN

Review problems• 5. Describe and give results of experiment that shows lac
operator is site of repressor binding.• 21. Why does translation of the trp leader region not simply
continue into the trp structural genes (trpE, etc.) in E. coli ?
• AQ 3. Consider E. coli cells each having one of the following mutations: Indicate effect each mutation on function of lac operon (assuming no glucose is present):
a. mutant lac operator (Oc locus) that doesn’t bind repressor.
b. mutant lac repressor (I-) that doesn’t bind operator
c. Mutant lac repressor (Is) that doesn’t bind inducer
d. Mutant lac promoter that doesn’t bind CAP-CAMP7-41

Review problems
4. Why are negative and positive control of the lac operon important to the neergy efficiency of E. coli cells?
14. Diagram how arabinose relieves repression in the araBAD operon. Show whereAraC is located (a) in the absence of arabinose; (b) in the presence of arabinose and lack glucose.
See Fig. 31. Because the his operon for synthesis of histidine operates much like trp operon. What might you predict about the sequence of amino acids encoded by the his leader?
7-42