chapter 3 enzymes. almost all processes in the living cell are catalyzed by the specific...
TRANSCRIPT

Chapter 3 Enzymes

• Almost all processes in the living cell are catalyzed by the specific biocatalyst. Enzymes are catalysts that change the rate of a reaction without being changed themselves. Enzymes are highly specific and their activity can be regulated..

•Biocatalyst: enzymes and ribozyme.•One of the most important functions of proteins is their role as catalysts. Until recently, all enzymes were considered to be proteins. Several examples of catalytic RNA molecules have now been vertified. Living processes consist almost entirely of biochemical reactions. Without catalysts these reactions would not occur fast enough to sustain life.

• Enzymes bind to one or more ligands, called substratee, and convert them into one or more chemically modified products.

1 Composition of enzymes
• Simple enzyme and conjugated enzyme. • Conjugated enzyme:• apoenzyme + cofactor holoenzyme. • Cofactor : prosthetic group+ coenzyme • prosthetic group: tightly bond with apoenzyme.
FAD, metal, etc. • coenzyme loosely bond with apoenzyme. NAD,
NADP, etc.

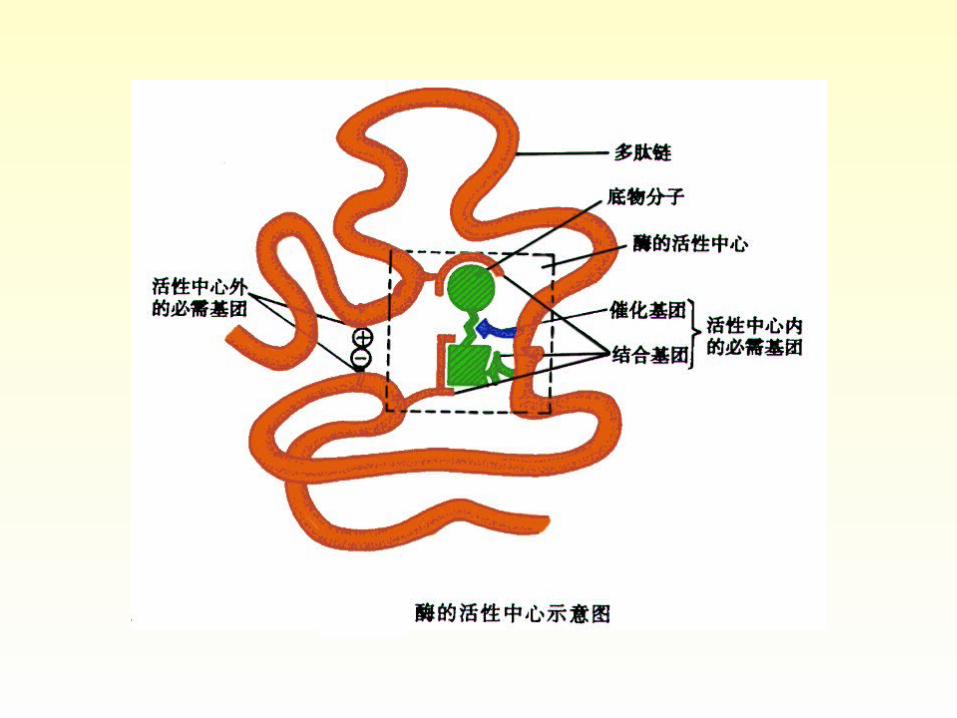
• Active site: Each type of enzyme molecule contains a unique, intricately shaped binding surface called an active site.
• Catalytic residues are highly conserved. Certain amino acids, notably cysteine and hydroxylic, acidic, or basic amino acids, perform key roles in catalysis.
• Essential group in active site: binding group +catalytic group. Cofactors always be a part of the active site.

Active site
• The active site is the region of the enzyme that binds the substrate, to form an enzyme-substrate complex, and transforms it into product. The active site is a three-dimensional entity, often a cleft or crevice on the surface of the protein, in which the substrate is bound by multiple weak interactions. Two models have been proposed to explain how an enzyme binds its substrate: the lock-and –key model and the induced-fit model.

2 Characteristics and mechanisms
of enzymatic reactions • Characteristics• Enzymes have several remarkable properties. First,
the rates of enzymatically catalyzed reactions are often phenomenally high. (Rate increases by factors of 106or greater are common.) . Second, in marked contrast to inorganic catalysts, the enzymes are highly specific to the reactions they catalyze. Side products are rarely formed. Finally, because of their complex structures, enzymes can be regulated. This is an especially important consideration in living organisms, which must conserve energy and raw materials.

• Specificity: Absolute specificity, relative specificity, and stereospecificity.
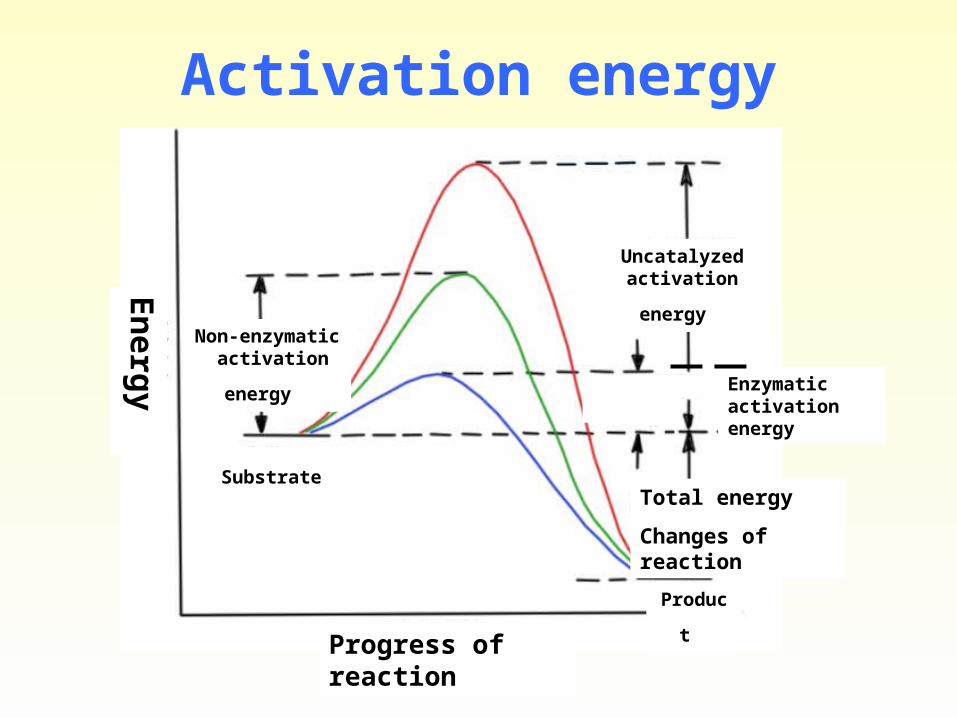
• Activation energy: To proceed at a viable rate, most chemical reactions require an initial input of energy. In the laboratory this energy is usually supplied as heat. At temperatures above absolute zero (-273.1ºC), all molecules possess vibrational energy, which increases as molecules are heated. Consider the following reaction:
A+B C
As the temperature rises, vibrating molecules (A and B) are more likely to collide, A chemical reaction occurs when the colliding molecules possess a minimum amount of energy called the activation energy.
Uncatalyzed
activation energy
Enzymatic activation energy
En
ergy
Progress of reaction
Total energy
Changes of reaction
Non-enzymatic
activation energy
Substrate
Product
Activation energy

• Not all collisions result in chemical reactions because only a fraction of the molecules have sufficient energy.
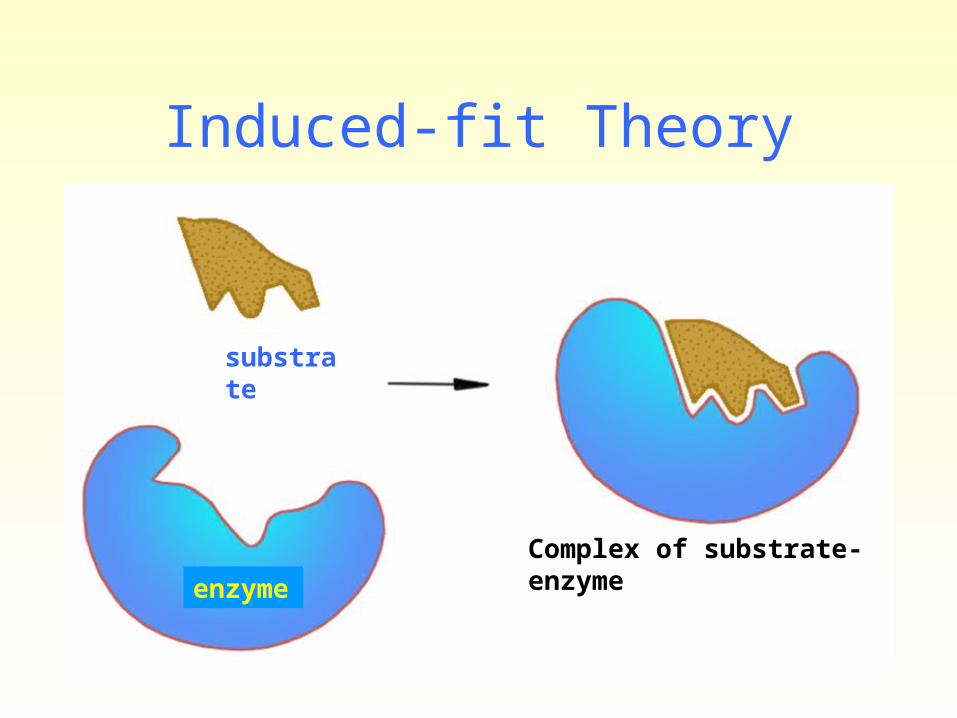
• Induced-fit hypothesis and transition state.
Substrates induce conformational changes in enzymes. During any chemical reaction reactants with sufficient energy will attain transition state (a strained intermediate form) when the substrate binds to the enzyme (inducing).
Induced-fit Theory
substrate
enzyme
Complex of substrate-enzyme

• Mechanisms
Proximity effect and orientation arrange: For a biochemical reaction to occur, the substrate must come into close proximity to catalytic functional groups (side chain groups involved in a catalytic mechanism ) within the active site. In addition, the substrate must be precisely, spatially oriented to the catalytic groups. Once the substrate is correctly positioned, a change in the enzyme’s conformation may result in a strained enzyme-substrate complex. This strain helps to bring the enzyme-substrate complex into the transition state.

• Multielement catalysis (Acid-Base catalysis ) : Chemical groups can often be made more reactive by adding or removing a proton. Enzyme active sites contain side chain groups that act as proton donors or acceptors. These groups are referred to as general acids or general bases.
• Surface effect: The strength of electrostatic interactions is related to the capacity of surrounding solvent molecules to reduce the attractive forces between chemical groups. Water is largely excluded from the active site as the substrate binds.

3 Enzyme kinetics• The rate or velocity of a biochemical reaction is
defined as the change in the concentration of a reactant or product per unit time.
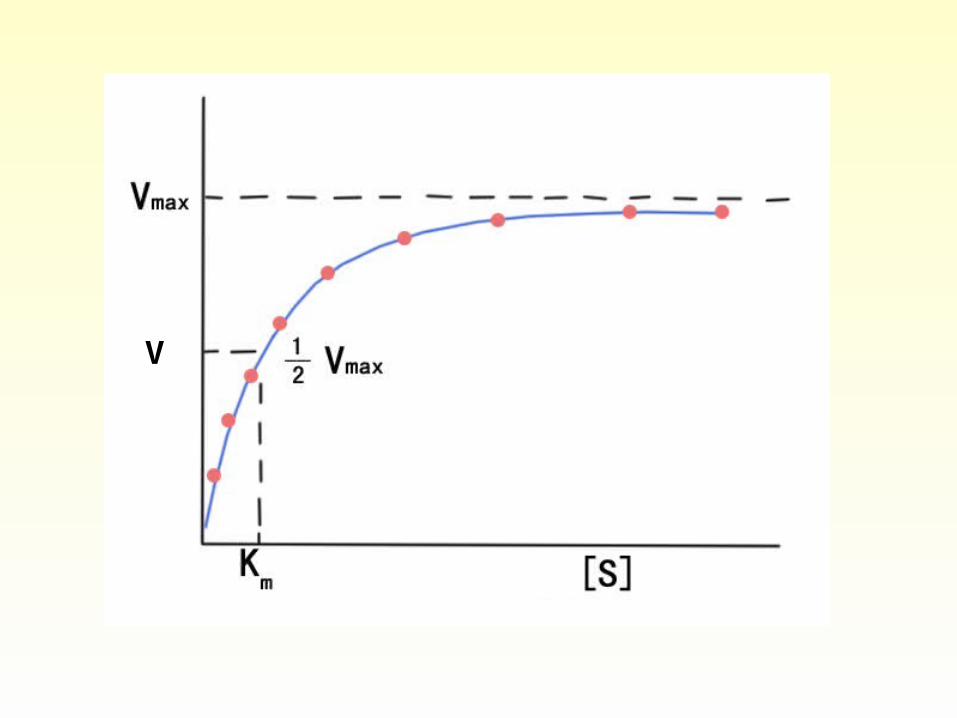
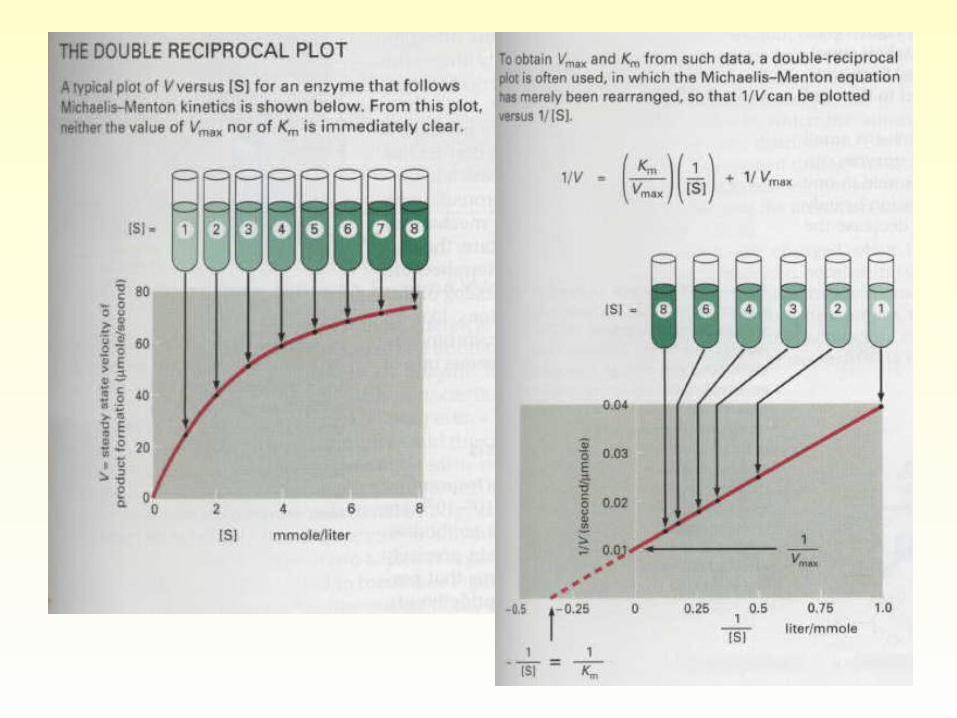
• Plotting initial velocity v versus substrate concentration [S].The rate of the reaction is directly proportional (first order reaction) to substrate concentration only when [S] is low. When [S] becomes sufficiently high that the enzyme is saturated, the rate of the reaction is zero-order with respect to substrate.
V

Michaelis-Menten Equation
K1= rate constant for ES formation
K2= rate constant for ES dissociation
K3= rate constant for product formation
and release from the active site
(1)
S + E ES E + Pk1
k2
k3
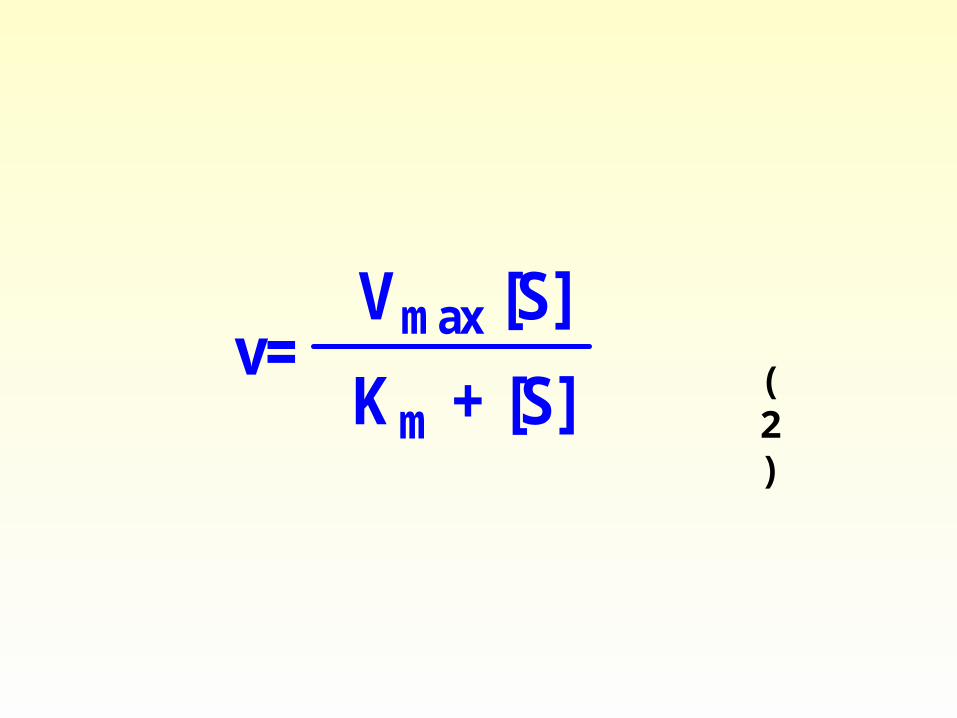
(2)
v=Vmax [S]
Km + [S]

ES formation = K1 ( [E] - [ES] ) [S] (3)
ES dissociation = K2 [ES ]+ K3 [ES] (4)
S + E ES E + Pk1
k2
k3

K1 ( [E] - [ES] ) [S] = K2 [ES ]+ K3 [ES]
( [E] - [ES] ) [S] K2+ K3
=
[ES] K1

Michaelis and Menton introduced a new constant,
Km ( now referred as the Michaelis constant):
K2+ K3
Km=
K1
( [E] - [ES] ) [S]
Km =
[ ES]

Km [ES] = [E] [S] – [ES] [S]
Km [ES] + [ES] [S] = [E] [S]
[ES] ( Km + [S] ) = [E] [S]
[E] [S]
[ES] = (5)
Km+[S]

Since V= K3 [ES], from ( 5 )
[E] [S]
V= K3 (6)
Km+[S]
When the [S] is much higher than the enzymes, all enzymes form [ES], that is, [E]= [ES], and maximum velocity ( Vmax ) can attain.
Vmax = K3 [ES] = K3 [E] (7)
Vmax
K3 =
[E]

Vmax [E] [S] Vmax [S]
V= = (2)
[E] Km+[S] Km+[S]

Significances of Km and Vmax
1) When [S] = Km,
Vmax [S] Vmax
V = =
[S] + [S] 2
2) When [S] is very much greater than Km,
Vmax [S] Vmax [S]
V= = = Vmax
Km+[S] [S]

3) It may reflect the affinity of the enzyme for its substrate. If K3 is much smaller than K2, that is K3 « K2, Km is the
dissociation constant for the [ES].
K2
Km=
K1
4) From Vmax = K3 [ES] = K3 [E], enzymes are saturated.
Vmax
K3=
[E]
The turnover number (Kcat ) = K3. This quantity is the numb
er of moles of substrate converted to product each second per mole of enzyme.

Vmaxv1
=Km . 1
[S] +
Vmax
1
Lineweaver-Burk Double-reciprocal pl
ot
y = mx + b
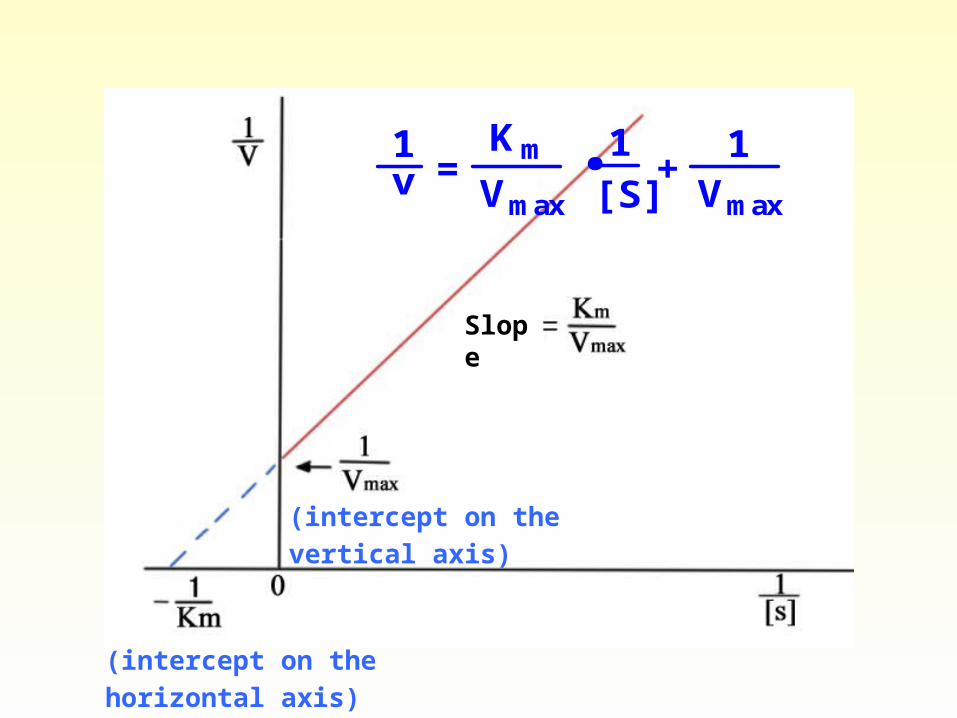
Slope
Vmaxv1
=Km . 1
[S] +
Vmax
1
(intercept on the vertical axis)
(intercept on the horizontal axis)

Multiple factors affect the rates of enzyme-catalyzed reactions.
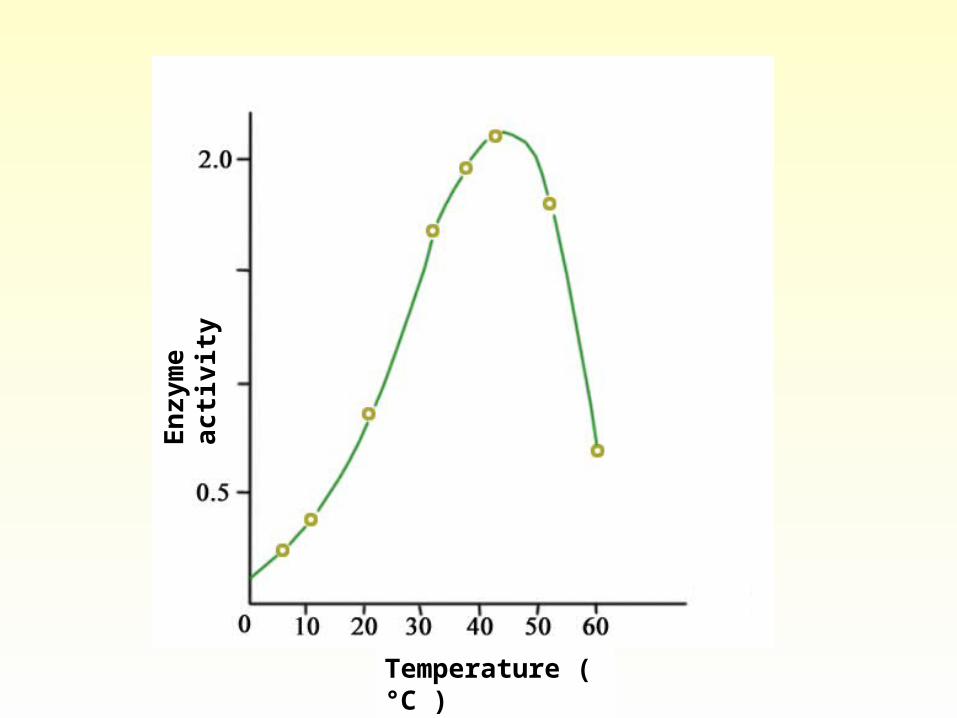
• Temperature While raising temperature increases the rate of an enzy
me-catalyzed reaction, this holds only over a strictly limited range of temperatures. The reaction rate initially increases as temperature rises owing to increased kinetic energy of the reacting molecules. Eventually, however, the kinetic energy of the enzyme exceeds the energy barrier for breaking the weak bonds that maintain its secondary-tertiary structure. At this temperature, denaturation, with an accompanying precipitate loss of catalytic activity, predominates.

• Enzymes from humans, who maintain a body temperature of 37 ºC, generally exhibit stability at temperature up to 45-55 ºC. Enzymes from microorganisms that inhabit natural hot springs or hyperthermal vents on the ocean floor may be stable at or above 100 ºC.
• Optimum temperature: Temperature at which it operates at maximal efficiency.
En
zym
e ac
tivi
ty
Temperature ( °C )

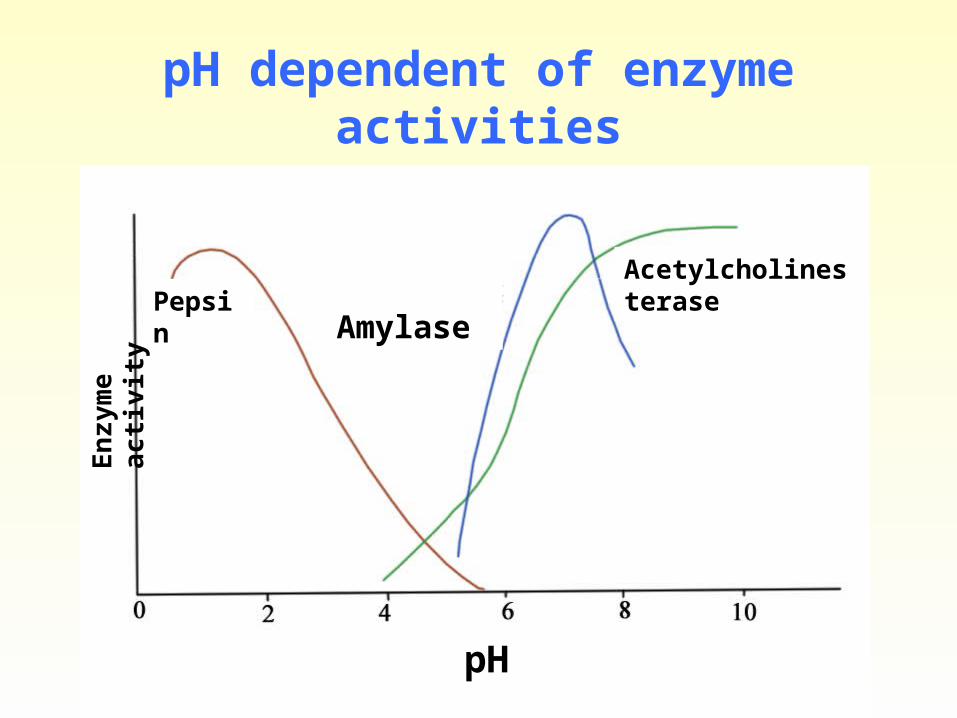
• pH
When enzyme activity is measured at several pH values, optimal activity typically is observed between pH values of 5 and 9. However, a few enzymes are active at pH values well outside this range.
pH optimum: The pH value at which an enzyme’s activity is maximal is called the pH optimum.

• Initial rate is proportionate to enzyme concentration
The initial rate of a reaction is the rate measured before sufficient product has been formed to permit the reverse reaction to occur. The initial rate of an enzyme-catalyzed reaction is always proportionate to the concentration of enzyme. Note, however, that this is statement holds only for initial rates.
• Substrate concentration
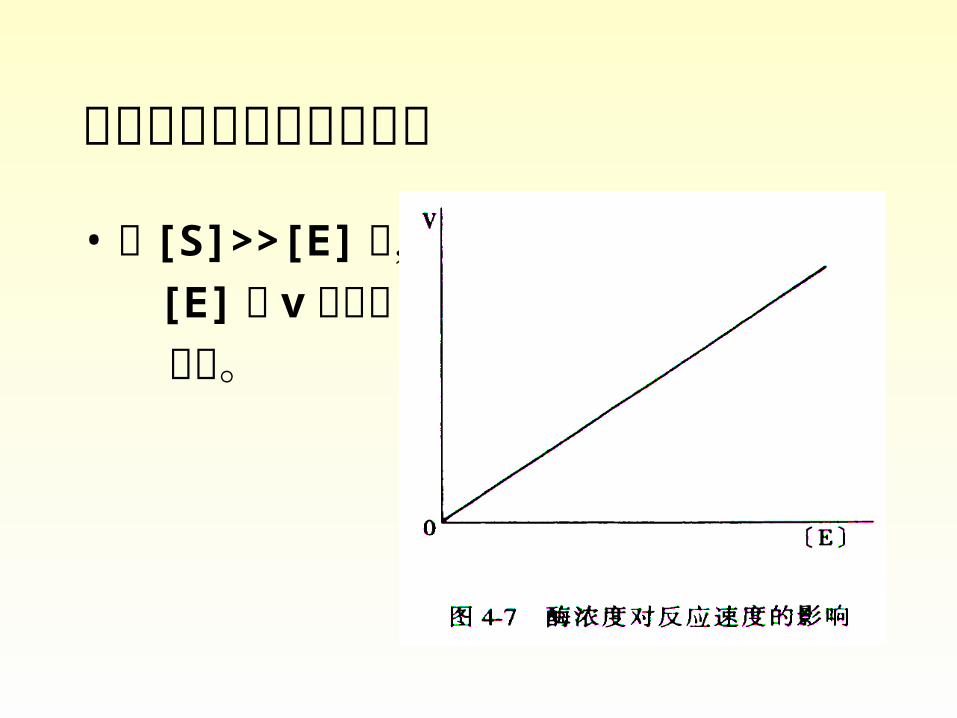
酶浓度对反应速度的影响
• 当 [S]>>[E] 时, [E] 与 v 呈正比 关系。
pH dependent of enzyme activities
Pepsin AmylaseAcetylcholinesterase
En
zym
e ac
tivi
ty
pH

(4) Enzyme inhibition The activity of enzymes can be inhibited. Many substances c
an reduce or eliminate the catalytic activity of specific enzymes. Inhibition may be irreversible or reversible.
Irreversible inhibitors usually bond covalently to the enzyme, often to a side chain group in the active site. For example, enzymes containing free sulfhydryl groups can react with alkylating agents such as iodoacetate and heavy metals. This process is not readily reversed either by removing the remainder of the free inhibitor or by increasing substrate concentration.
Specific inhibitor: specifically bind to essential amino acid on active site. Some organic phosphor compounds could specifically bind to –OH of serine.

Non specific inhibitor: not only binds to essential group, but also to outsides of essential group. Hg2+, Ag2+ and As3+ .
In reversible inhibition:
the inhibitor can dissociate from the enzyme because it binds through noncovalent bonds. The most common forms of reversible inhibition are competitive and noncompetitive.

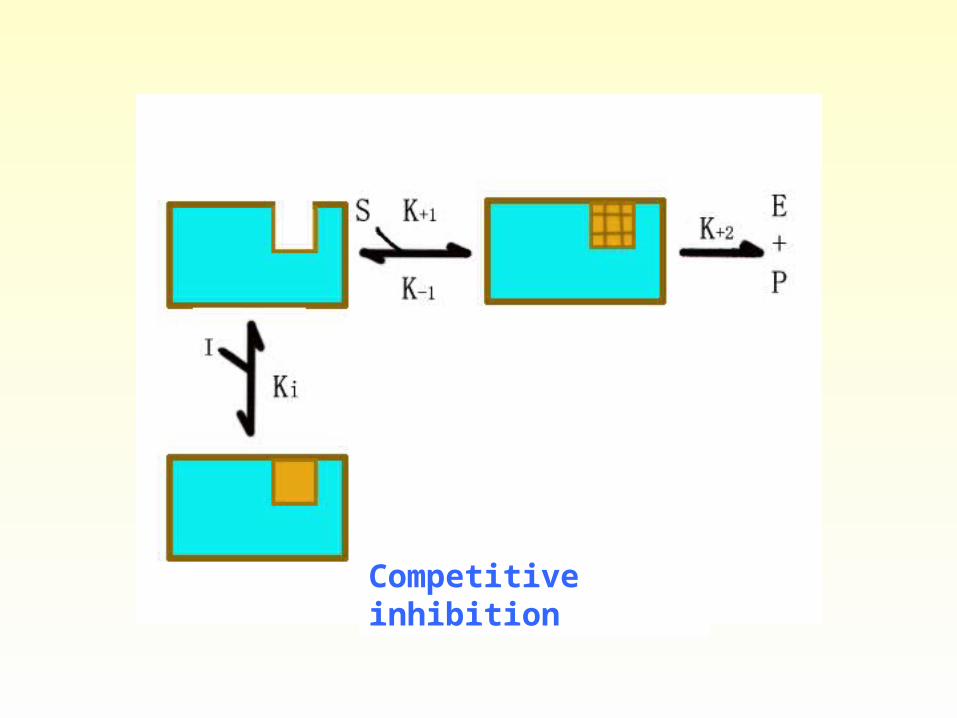
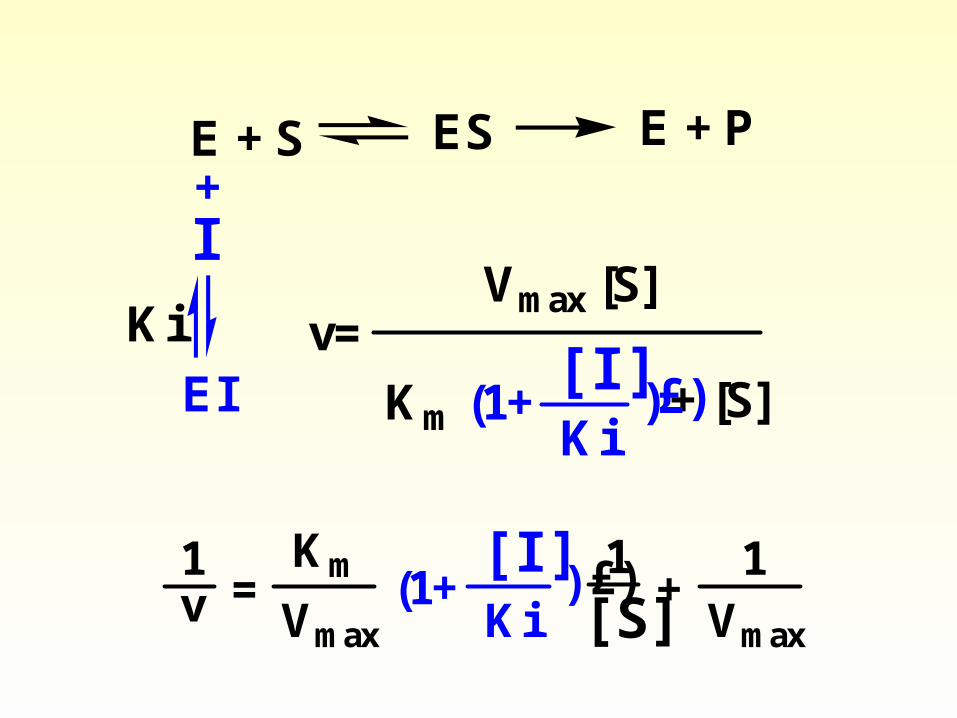
1) Competitive inhibition
• Competitive inhibitors typically resemble the substrate
• Classic competitive inhibition occurs at the substrate-binding (catalytic) site. The chemical structure of a substrate analog inhibitor (I) generally resembles that of the substrate (S). It therefore combines reversibly with the enzyme, forming an enzyme-inhibitor (EnzI) complex rather than an EnzS complex.
Competitive inhibition
Vmaxv1
=Km 1
[S] +
Vmax
1 (1+
Ki
[I] £))
v=Vmax [S]
Km (1+ + [S]Ki[I] £))
E + S E + P+I
EI
ES
Ki

inhibitor
No inhibitor

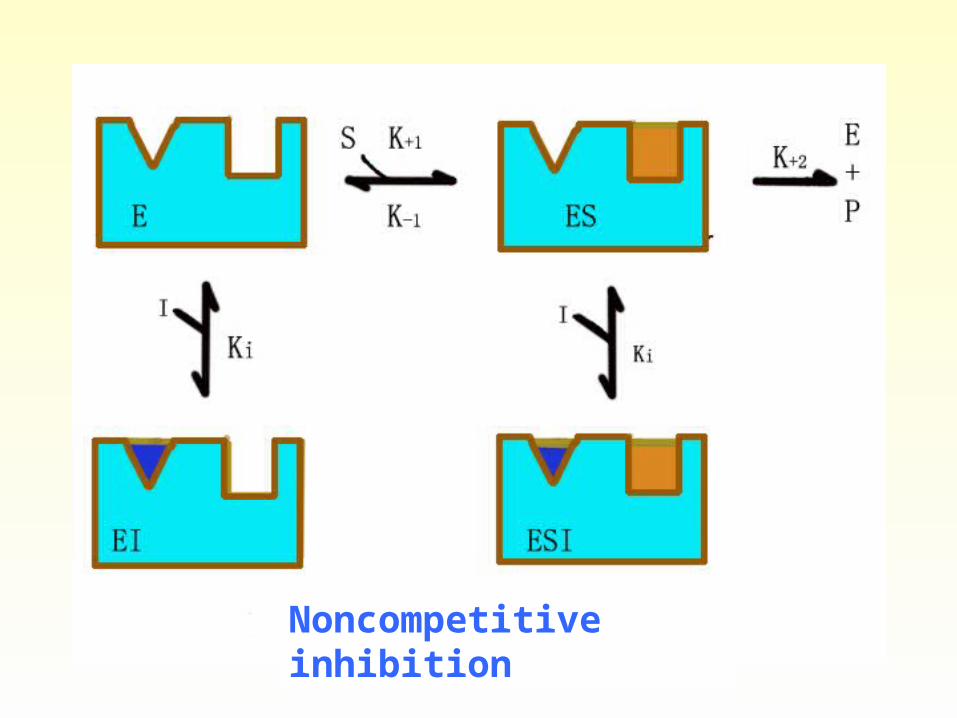
Noncompetitive inhibition • In noncompetitive inhibition, no
competition occurs between S and I. The inhibitor usually bears little or no structural resemblance to S and may be assumed to bind to the enzyme at a site other than the active site. Both EI and EIS complexes form. Inhibitor binding alters the enzyme’s three-dimensional configuration and blocks the reaction.
Noncompetitive inhibition

E + S ES E + P+I
ESI
+I
EI + S
E + S ES E + P+I
ESI
+I
EI + S
Ki Ki

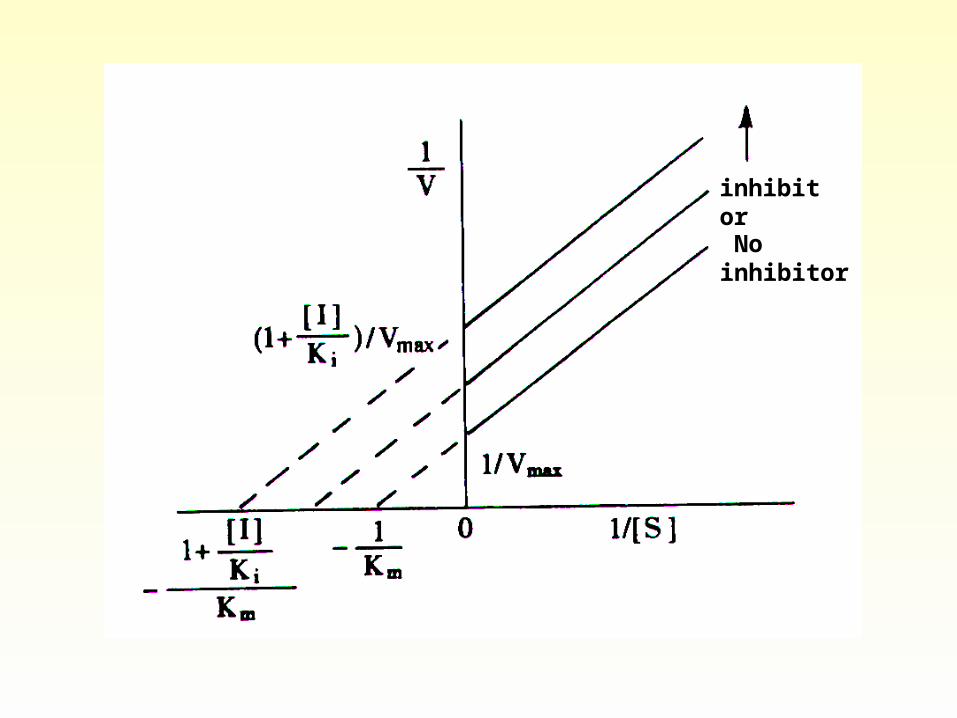
Plots of 1/V versus 1/[S] in the presence of several concentrations of the inhibitor intersect at the same point on the horizontal axis, -1/Km. In
noncompetitive inhibition the dissociation constants for ES and EIS are assumed to stay the same.

inhibitor
No inhibitor

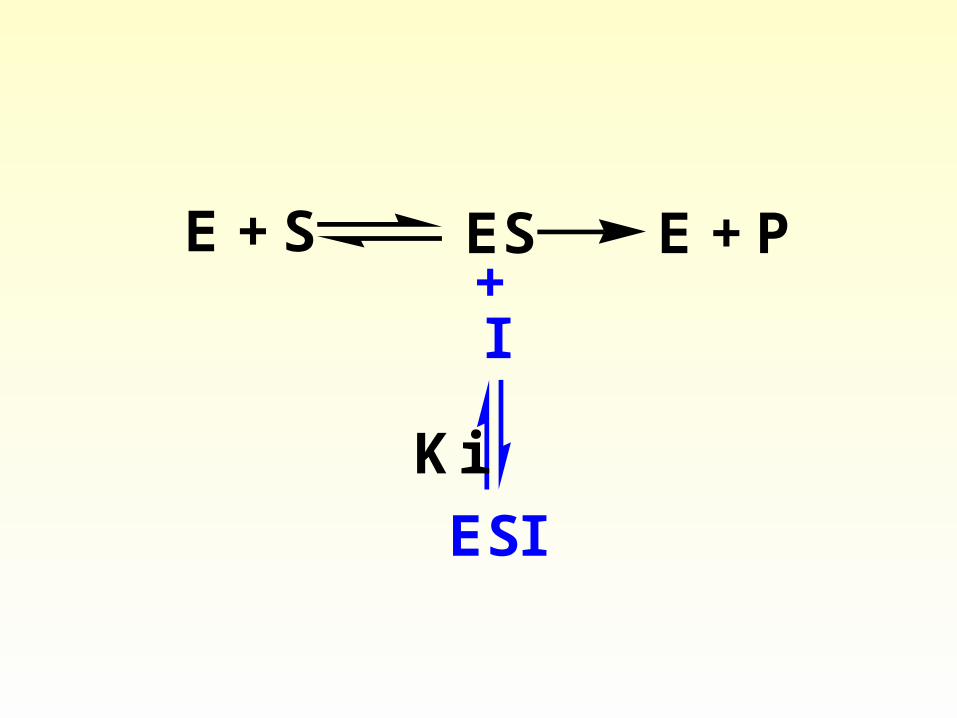
3) Uncompetitive inhibition
• The inhibitor bind to ES and results in decrease of both ES and P (also free E).
• E + S ES E+S
+
I
Ki
ESI

Uncompetitive inhibition
Ki
E + S ES E + P
ESI
+I
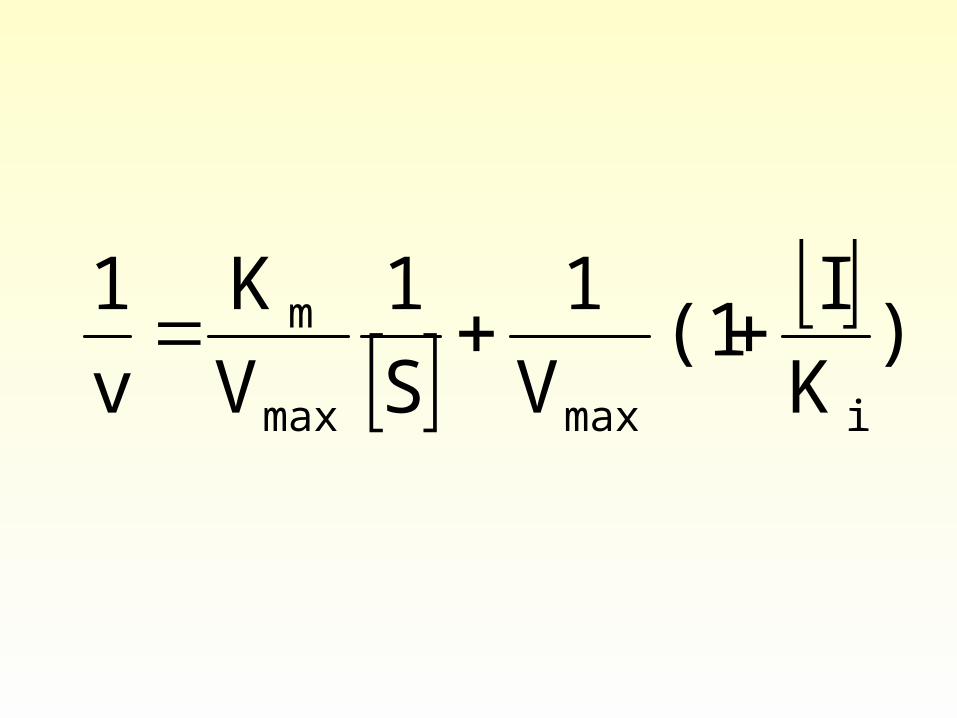
)K
I(1
V
1
S
1
V
K
v
1
imaxmax
m
No inhibitor
inhibitor

4) Effect of activator on the enzyme activities
• Activator: substances enable non-active enzyme to become active one. Metals such as Mg2+, K+, Mn2+, etc.
• Essential activator and non-essential activator.

5) Enzyme activity assay and unit of
enzyme activity • Enzyme activity is measured in international units (I.
U.) One I.U. is defined as the amount of enzyme that produces 1μmol of product per minute. An enzyme specific activity, a quantity that is used to monitor enzyme purification, is defined as the number of international units per milligram of protein.
• A new unit for measuring enzyme activity called the katal, has recently been introduced. One katal (kat) indicates the amount of enzyme for the transformation of 1 mole of substrate per second.
• 1 IU =16.67×10-9 kat

4 Regulation of enzyme • The thousands of enzyme-catalyzed chemical
reactions in living cells are organized into a series of biochemical or metabolic pathways. Each pathway consists of a sequence of catalytic steps. The product of the first reaction becomes the substrate of the next and so on. Metabolic and other processes are controlled by altering the quantity or the catalytic efficiency of enzymes.

1) Regulation of enzyme activities• A. Proenyme or Zymogen: Certain proteins are
manufactured and secred in the form of inactive precursor proteins known as proproteins. When the proteins are enzymes, the proproteins are termed proenzymes or zymogens. Conversion of a proprotein to the mature protein involves selective proteolysis, a process that converts the proprotein by one or more successive proteolytic “clips” to a form having the characteristic activity of the mature protein ( its enzymatic activity ). Examples include the hormone insulin (proinsulin), pepsinogen, trypsinogen, etc.

• Selective proteolysis of a proenzyme may be viewed as a process that triggers essential conformational changes that “create” the catalytic site.

B. Allosteric enzyme
• Allosteric enzymes are enzymes whose activity at the catalytic site may be modulated by the presence of allosteric effectors at an allosteric site. Allosteric effector could be products, substrate, and so on.
• Feed back inhibition referred to the inhibition of the activity of an enzyme in a biosynthetic pathway by an end product (often as allosteric effectors) of that pathway.

C. Regulatory covalent modification
• Regulatory covalent modifications can be reversible or irreversible. In mammalian cells, the two most commonly used forms of covalent modification are partial proteolysis and phosphorylation. Because cells lack the ability to reunite the two portions of a protein produced following hydrolysis of a peptide bond, the partial proteolysis is considered an irreversible modification.

• Hydrolysis of the phosphoesters formed when a protein is covalently phosphorylated on the side chain of a serine, threonine, or tyrosine residues is both thermodynamically spontaneous and readily catalyzed by enzymes called protein phosphatases. Hence, phosphorylation represents a reversible modification process.

Cyclic phosphorylation and dephosphorylation is a common cellular mechanism for regulating protein activity. In this example, the target protein R (orange) is inactive when phosphorylated and active when dephosphorylated; the opposite pattern occurs in some proteins.

2) Regulation of enzyme quantity
• Rate of synthesis and degradation determine enzyme quantity. The quantity of an enzyme in a cell may be increased either by elevating its rate of synthesis, by decreasing its rate of degradation, or by both. Cells can synthesize specific enzymes in response to changing metabolic needs, a process referred to as enzyme induction. The induction accomplished by genetic control. Although many inducers are substrates for the enzymes they induce, compounds structurally similar to the substrate may be inducers but not substrates. Conversely, a compound may be a substrate but not an inducer.

• The synthesis of certain enzymes may also be specifically inhibited. In a process called repression, the end product of a biochemical pathway may inhibit the synthesis of a key enzyme in the pathway. Both induction and repression involve cis-elements, specific DNA sequences located upstream of genes that encode a given enzyme, and a trans-acting regulatory proteins.
• Regulation of enzyme degradation. The degradation of mammalian proteins by ATP and ubiqitin-dependent pathways and by ATP-independent pathways. It also Related to the nutrition and hormone state.

Compartmentation
• In eukaryotic cells, biochemical pathways are segregated into different organelles. One purpose for this physical separation is that opposing processes are easier to control if the occur in different compartments. For example, fatty acid biosynthesis occurs in the cytoplasm, while the energy-generating reactions of fatty acid oxidation occur within the mitochondria. Another factor is that each organelle can concentrate specific substances such as substrates and coenzymes. In addition, special microenvironments are often created within organelles.

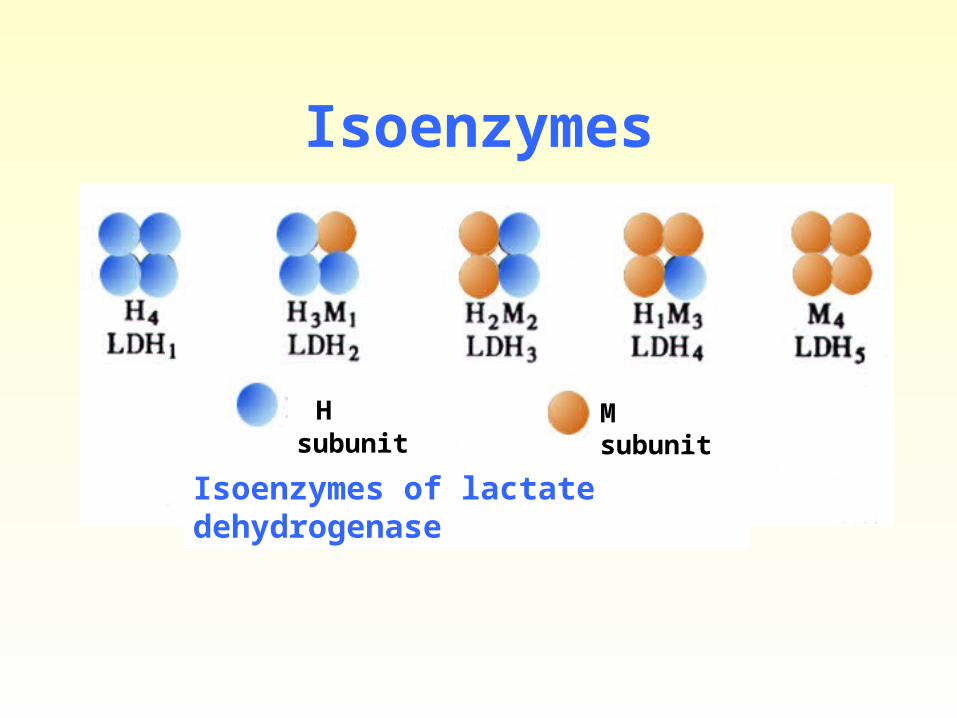
3) Isoenzymes
• The enzymes catalyzing the same biochemical reaction.
• Lactate dehydrogenase (LDH)
Isoenzymes
H subunit M subunit
Isoenzymes of lactate dehydrogenase

5 Nomenclature and classification
• The International Union of Biochemistry (IUB) adopted a complex but unambiguous system of enzyme nomenclature based on reaction mechanism.
• (1) Reactions and the enzymes that catalyzed them form six classes, each having 4-13 subclasses.

• (2) The enzyme name has two parts. The first names the substrate or substrates. The second, ending in –ase, indicates the type of reaction catalyzed.
• (3) Additional information, if needed to clarify the reaction, may follow in parentheses; eg, the enzyme catalyzing
• L-malate + NAD+ pyruvate + CO2 + NADH + H+
is designated 1.1.1.37 L-malate:
NAD+ oxidaoreductase (decarboxylating). • (4) Each enzyme has a code number (EC) that cha
racterizes the reaction type as to class, subclass, and subsubclass.

Classification • Six classes based on reaction mechanism:• (1) Oxidoreductases: LDH, Cytochrome C, etc.• (2) Transferases: methyl transferase.• (3) Hydrolases: amylase• (4) Lyases removing a group to form a double bond,
or reverse reaction.• (5) Isomerase to catalyze the intertransfer of isomers.• (6) Ligase. catalyzing two substrates link to form one
compound.

Relationship between Enzyme and
Medicine
选择题练习酶化学

A .所有蛋白质都有酶的活性
B .其底物都是有机化合物
C .其催化活性都需要特异的辅助因子
D .体内所有具有催化活性的物质都是酶
E .酶是由活细胞合成具有催化作用的蛋白质
1. 关于酶概念的叙述下列哪项是正确的?

A .所有酶的活性中心都有金属离子
B .所有的抑制剂都作用于酶的活性中心
C .所有的必需集团都位于酶的活性中心
D .所有酶的活性中心都含有辅酶
E .所有的酶都有活性中心
2 .关于酶活性中心的叙述下列哪项是正确的?

3. 酶加速化学反应的根本原因是 ( )
A. 升高反应温度
B. 增加反应物碰撞频率
C. 降低催化反应的活化能
D. 增加底物浓度
E. 降低产物的自由能

4. Holoenzyme refer to ( )
A. Complex of enzyme with substrate
B. Complex of enzyme with suppressant
C. Complex of enzyme with cofactor
D. Inactive precursor of enzyme
E. Complex of enzyme with allosteric effector

5. 金属离子作为辅助因子的作用错误的是 ( )
A. 作为酶活性中心的催化基团参加反应
B. 与稳定酶的分子构象无关
C. 可提高酶的催化活性
D. 降低反应中的静电排斥
E. 可与酶、底物形成复合物

6. 活化能的概念是指 ( )
A. 底物和产物之间能量的差值
B. 参与反应的分子所需的总能量
C. 分子由一般状态变成活化态所需能量
D. 温度升高时产生的能量
E. 以上都不是

7. 酶促反应动力学所研究的是 ( )
A. 酶的基因来源
B. 酶的电泳行为
C. 酶的诱导契合
D. 酶分子的空间结构
E. 影响酶促反应速度的因素

8. Michaelis-Menten enzyme kinetics diagram of curves is a ( )
A. straight line
B. rectangular hyperbola
C. S shape curve
D. parabola
E. Not above all

9. 关于 Km 的意义正确的是 ( )
A. Km 为酶的比活性
B. 1/Km 越小 , 酶与底物亲和力越大
C. Km 的单位是 mmol/min
D. Km 值是酶的特征性常数之一
E. Km 值与酶的浓度有关

10. 竞争性抑制剂的特点是 ( )
A. 抑制剂以共价键与酶结合
B. 抑制剂的结构与底物不相似
C. 当抑制剂的浓度增加时 , 酶变性失活
D. 当底物浓度增加时 , 抑制作用不减弱
E. 抑制剂和酶活性中心外的部位结合

11. In anticompetitive inhibition of enzyme, the reaction kinetics parameter change as ( )
A. Km↑,Vmax invariably
B. Km↓,Vmax↓
C. Km invariably,Vmax↓
D. Km↓,Vmax invariably
E. Km↓,Vmax↑

12. 有机磷农药与酶活性中心结合的基团是 ( )
A. 组氨酸上的咪唑基
B. 赖氨酸上的 ε- 氨基
C. 丝氨酸上的羟基
D. 半胱氨酸上的巯基
E. 谷氨酸上的 γ- 羧基

13. 关于变构酶的论述错误的是 ( )
A. 变构酶为多亚基组成
B. 如底物与一亚基结合后 , 使其他亚基迅速与底物结合程正协同效应
C. 正协同效应的底物浓度曲线呈矩形双曲线
D. 底物与一亚基结合后 , 使其亚基结合底物能力减少称负协同效应
E. 变构效应剂与一亚基结合后 , 使酶其他亚基迅速与底物结合为异促协同效应

14. –SH is one enzyme’s essential group. Which substance can protect this enzyme from oxidation?A. Cys
B. GSH
C. urea
D. ionic detergent
E. ethanol

15. 快速调节可通过 ( )
A. 磷酸化与去磷酸化
B. 腺苷酸化与腺苷酸化
C. 变构调节
D. 改变酶的合成速度
E. 酶促反应的可调节性

16. The characteristic constants of enzymes include ( )
A. Enzymic optimum temperature
B. Enzymic optimum pH
C. Vmax
D. Km
E. KS

17. 磺胺药的抑菌机理是 ( )
A. 竞争性抑制二氢叶酸合成酶的活性
B. 干扰体内核酸的代谢
C. 结构与二氢叶酸相似
D. 抑制程度强弱取决于药物与酶底物浓度的相对比例
E. 磺胺药是二氢叶酸合成酶的变构抑制剂

18. Cofactors of enzyme are ( )
A. Micromolecule organic compounds
B. metal ion
C. vitamine
D. various kinds of organic and inorganic compounds
E. A kind of conjugated protein

19. 某种酶的活性依赖于酶活性中心的必需基团 -SH, 能保护此酶不被氧化的物质是 ( )
A. GSH
B. 维生素 C
C. 半胱氨酸
D. 维生素 A
E. 两价阳离子

20. 酶分子上必需基团的作用是 ( )
A. 与底物结合
B. 催化底物发生化学反应
C. 含砷的有机化合物
D. 决定辅酶结构
E. 维持酶分子空间结构

Thank you!