chapter 25 nitrogen acquisition and amino acid metabolism biochemistry by reginald garrett and...
TRANSCRIPT

Chapter 25
Nitrogen Acquisition and Amino Acid Metabolism
Biochemistry
by
Reginald Garrett and Charles Grisham

Outline
1. Which Metabolic Pathways Allow Organisms to Live on Inorganic Forms of Nitrogen?
2. What Is The Metabolic Fate of Ammonium?
3. What Regulatory Mechanisms Act on Escherichia coli Glutamine Synthetase?
4. How Do Organisms Synthesize Amino Acids?
5. How Does Amino Acid Catabolism Lead into Pathways of Energy Production?

25.1 – Which Metabolic Pathways Allow Organisms to Live on
Inorganic Forms of Nitrogen?
Nitrogen is cycled between organisms and inanimate enviroment
• The principal inorganic forms of N are in an oxidized state– As N2 in the atmosphere
– As nitrate (NO3-) in the soils and ocean
• All biological compounds contain N in a reduced form (NH4
+)

• Thus, Nitrogen acquisition must involve 1. The Reduction of the oxidized forms (N2 and NO3
-) to NH4
+
2. The incorporation of NH4+ into organic linkage as
amino or amido groups
• The reduction occurs in microorganisms and green plants. But animals gain N through diet.
Figure 25.1 The nitrogen cycle.
(+5) (-3)
(+3)(0)
(+5)
(+3)
(-3)
(+2)(+1)

The Reduction of Nitrogen
Nitrogen assimilation and nitrogen fixation
1. Nitrate assimilation occurs in two steps: – 2e- reduction of nitrate to nitrite – 6e- reduction of nitrite to ammonium (Fig 25.1) • Nitrate assimilation accounts for 99% of N
acquisition by the biosphere
2. Nitrogen fixation involves reduction of N2 in prokaryotes by nitrogenase

Nitrate Assimilation• Nitrate assimilation
– the reduction of nitrate to NH4+ in plants,
various fungi, and certain bacteria– Two steps:
1. Nitrate reductase
NO3- + 2 H+ + 2 e- → NO2
- + H2O
2. Nitrite reductase
NO2- + 8 H+ + 6 e- → NH4
+ + 2 H2O
• Electrons are transferred from NADH to nitrate

Nitrate reductase• Nitrate reductases are cytosolic 210-270 kD
dimeric protein, pathway involve– SH of enzyme– FAD
– Cytochrome b557
– Molybdenum cofactor
• MoCo required both for reductase activity and for assembly of enzyme subunits to active dimer
NADH NO3-
[E-SH →FAD→cytochrome b557 →MoCo]
NAD+ NO2-

Nitrite ReductaseLight drives reduction of ferredoxins and
electrons flow to 4Fe-4S and siroheme and then to nitrite
• Nitrite is reduced to ammonium while still bound to siroheme
• In higher plants, nitrite reductase is in chloroplasts, but nitrate reductase is cytosolic
In higher plants
siroheme

Figure 25.3 Domain organization within the enzymes of nitrate assimilation. The numbers denote residue number along the amino acid sequence of the proteins.

Nitrogen fixation
N2 + 10 H+ + 8 e- → 2 NH4+ + H2
• Only occurs in certain prokaryotes – Rhizobia fix nitrogen in symbiotic association
with leguminous plants – Rhizobia fix N for the plant and plant provides
Rhizobia with carbon substrates
• Fundamental requirements:1. Nitrogenase2. A strong reductant (reduced ferredoxin)3. ATP4. O-free conditions

Nitrogenase Complex
Two metalloprotein components: 1. Nitrogenase reductase2. Nitrogenase

Nitrogenase reductase
• Nitrogenase reductase– Fe-protein– A 60 kD homodimer with a single 4Fe-4S cluster
• Extremely O2-sensitive • Binds MgATP and hydrolyzes 2 ATPs per
electron transferred• Because reduction of N2 to 2NH4
+ + H2 requires 8 electrons, 16 ATP are consumed per N2
reduced

Figure 25.4 The triple bond in N2 must be broken during nitrogen fixation.
• N2 reduction to ammonia is thermodynamically favorable • However, the activation barrier for breaking the N-N triple
bond is enormous • 16 ATP provide the needed activation energy

Nitrogenase• MoFe-protein—a 220 kD 22 heterotetramer
• An dimer serve as the functional unit– Contains two types of metal centers
1. P-cluster (figure 25.5a)8Fe-7S center
2. FeMo-cofactor (figure 25.5b)7Fe-1-Mo-9S cluster
• Oxygen labile• Nitrogenase is a rather slow enzyme
– 12 e- pairs per second, i.e., only three molecules of N2 per second
– As much as 5% of cellular protein may be nitrogenase
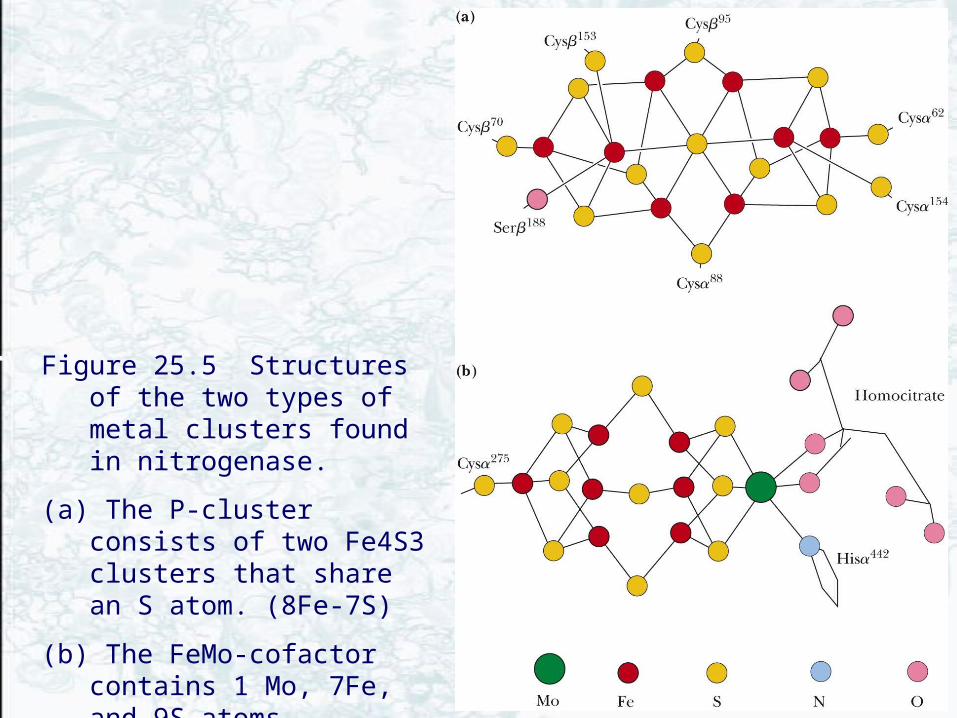
Figure 25.5 Structures of the two types of metal clusters found in nitrogenase.
(a) The P-cluster consists of two Fe4S3 clusters that share an S atom. (8Fe-7S)
(b) The FeMo-cofactor contains 1 Mo, 7Fe, and 9S atoms. Homocitrate provides two oxo ligands to the Mo atom.

The regulation of nitrogen Fixation• Two regulatory controls 1. ADP inhibits the activity of
nitrogenase
2. NH4+ represses the
expression of nif genes• In some organisms, the
nitrogenase complex is regulated by covalent modification. ADP-ribosylation of nitrogenase reductase leads to its inactivation.
Figure 25.8 Regulation of nitrogen fixation.

25.2 – What Is The Metabolic Fate of Ammonium?
NH4+ enters organic linkage via three major
reactions in all cells 1. Glutamate dehydrogenase (GDH)
2. Glutamine synthetase (GS)
3. Carbamoyl-phosphate synthetase I (CPS-I)
• Asparagine synthetase (some microorganisms)

1. Glutamate dehydrogenase (GDH)
• Reductive amination of -ketoglutarate to form glutamate
NH4+ + -ketoglutarate + NADPH + 2 H+ →
glutamate + NADP+ + H2O• Mammalian GDH plays a prominent role in amino
acid catabolism (oxidative amination)

• ATP-dependent amidation of -carboxyl of glutamate to glutamine
NH4+ + glutamate + ATP →
glutamine + ADP + Pi
• Glutamine is a major N donor in the biosynthesis of many organic N compounds, therefore GS activity is tightly regulated
• Glutamine is the most abundant amino acid in human
2. Glutamine synthetase (GS)

Figure 25.10(a) The enzymatic reaction catalyzed by glutamine synthetase.
(b) The reaction proceeds by (a) activation of the -carboxyl group of Glu by ATP, followed by (b) amidation by NH4
+.

Ammonium is converted to carbamoyl-P
• This reaction is an early step in the urea cycle
NH4+ + HCO3
- + 2 ATP →
carbamoyl phosphate + 2 ADP + Pi + 2 H+
• Two ATP required– one to activate bicarbonate
– one to phosphorylate carbamate
3. Carbamoyl-phosphate synthetase I (CPS-I)

The major pathways of Ammonium Assimilation lead to glutamin synthesis
Two principal pathways :1. Principal route: GDH/GS in organisms rich in N
2. Secondary route: GS/GOGAT in organisms confronting N limitation
– GOGAT is glutamate synthase or glutamate:oxo-glutarate amino transferase
– GDH has a higher Km for NH4+ than does GS

The glutamate synthase (GOGAT)reaction, showing the reductants exploited by different organisms in this reductive amination reaction.

25.3 – What Regulatory Mechanisms Act on Glutamine Synthetase
• GS in E. coli is regulated in three ways:1. Feedback inhibition (allosteric regulation)
2. Covalent modification (interconverts between inactive and active forms)
3. Regulation of gene expression and protein synthesis control the amount of GS in cells
• But no such regulation occurs in eukaryotic versions of GS
• E. coli GS is a 12-mer

1. Allosteric Regulation of Glutamine Synthetase
• 9 different feedback inhibitors: Gly, Ala, Ser, His, Trp, CTP, AMP, carbamoyl-P, and glucosamine-6-P– Gly, Ala, Ser are indicators of amino acid metabolism in
cells– Other six are end products of a biochemical pathway
• AMP competes with ATP for binding at the ATP substrate site
• Gly, Ala, and Ser compete with Glu for binding at the active site
• This effectively controls glutamine’s contributions to metabolism

Figure 25.15 The allosteric regulation of glutamine synthetase activity by feedback inhibition.

2. Covalent Modification of Glutamine Synthetase
• Each subunit can be adenylylated at Tyr-397– Adenylylation inactivates GS
• ATP:GS:adenylyl transferase (AT) catalyzes both the adenylylation and deadenylylation– PII (regulatory protein) controls these
– AT:PIIA catalyzes adenylylation
– AT:PIID (PII-UMP) catalyzes deadenylylation
-Ketoglutarate and Gln also affect -Ketoglutarate activates AT:PIID and inhibit AT:PIIA
– Gln activates AT:PIIA and inhibit AT:PIID

Figure 25.16 Covalent modification of GS: Adenylylation of Tyr397 in the glutamine synthetase polypeptide via an ATP-dependent reaction catalyzed by the converter enzyme adenylyl transferase (AT).
From 1 through 12 GS monomers in the GS holoenzyme can be modified, with progressive inactivation as the ratio of [modified]/[unmodified] GS subunits increases.

Figure 25.17 The cyclic cascade system regulating the covalent modification of GS.
(Adenylylation)
(Deadenylylation)

3. Gene Expression regulates GS
Gene GlnA is actively transcribed only if a transcriptional enhancer NRI is in its phosphorylated form, NRI-P
• NRI is phosphorylated by NRII, a protein kinase
• If NRII is complexed with PIIA it acts as a phosphatase, not a kinase
(phosphatase)
(kinase)

25.4 – Amino Acid Biosynthesis
• Organisms show substantial differences in their capacity to synthesize the 20 amino acids common to proteins– Plants and microorganisms can make all 20 amino
acids and all other needed N metabolites – In these organisms, glutamate is the source of N, via
transamination (aminotransferase) reactions
• Amino acids are formed from -keto acids by transamination
Amino acid1 + -keto acid2 → -keto acid1 + Amino acid2

Figure 25.19 Glutamate-dependent transamination of -keto acid carbon skeletons is a primary mechanism for amino acid synthesis.

The Mechanism of the Aminotransferase (Transamination) Reaction

*Arginine and histidine are essential in the diets of juveniles, not adults
• Mammals can make only 10 of the 20 AAs – The others are classed as "essential" amino acids and
must be obtained in the diet

The pathways of amino acid biosynthesis can be organized into families
According to the intermediates that they are made from
1. -ketoglutarate
2. Oxaloacetate
3. Pytuvate
4. 3-phosphoglycerate
5. Phosphoenolpyruvate and erythrose-4-P (aromatic)


1. The -Ketoglutarate FamilyGlu, Gln, Pro, Arg, and sometimes Lys
• The routes for Glu and Gln synthesis were described when we considered pathways of ammonia assimilation – Transamination of -Ketoglutarate gives glutamate – Amidation of glutamate gives glutamine
• Proline is derived from glutamate • Ornithine is also derived from glutamate
– the similarity to the proline pathway
• Arginine are part of the urea cycle

Figure 25.20 The pathway of proline biosynthesis from glutamate. The enzymes are (1) -glutamyl kinase, (2) glutamate-5-semialdehyde dehydrogenase, and (4) 1-pyrroline-5-carboxylate reductase; reaction (3) occurs nonenzymatically.
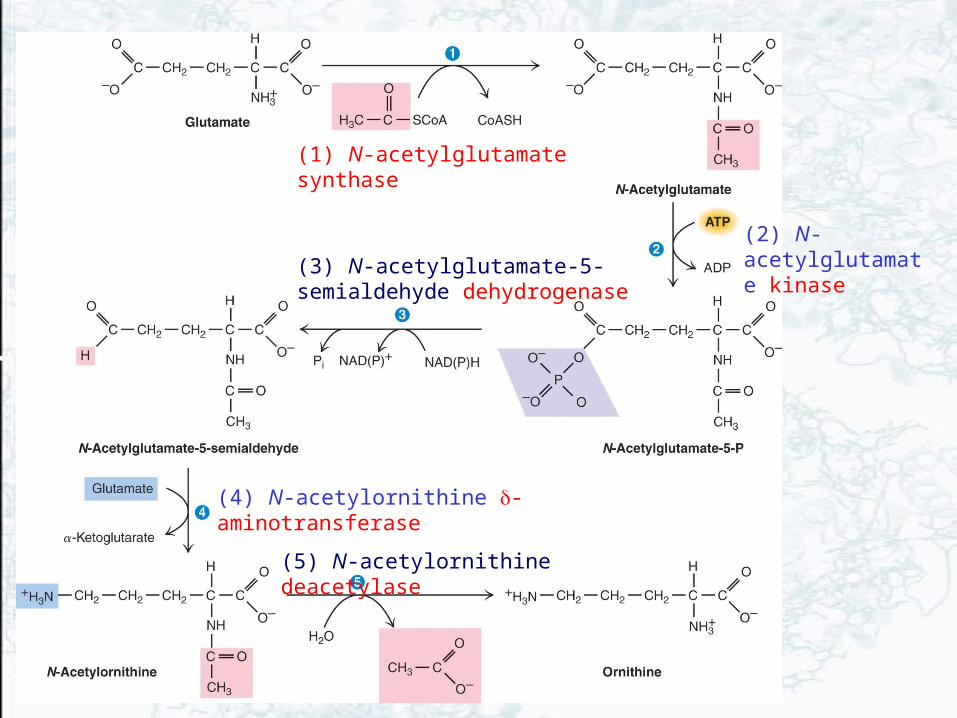
(1) N-acetylglutamate synthase
(5) N-acetylornithine deacetylase
(4) N-acetylornithine -aminotransferase
(3) N-acetylglutamate-5-semialdehyde dehydrogenase
(2) N-acetylglutamate kinase

• Ornithine has three metabolic roles 1. To serve as precursor to arginine
2. To function as an intermediate in the urea cycle
3. To act as an intermediate in arginine degradation
• -NH3+ of ornithine is carbamoylated by
onithine transcarbamoylase in urea cycle

Carbamoyl-phosphate synthetase I
• Carbamoyl-phosphate synthetase I (CPS-I)
– NH3-dependent mitochondrial CPS isozyme
1. HCO3- is activated via an ATP-dependent
phosphorylation
2. Ammonia attacks the carbonyl carbon of carbonyl-P, displacing Pi to form carbamate
3. Carbamate is phosphorylated via a second ATP to give carbamoyl-P

Figure 25.22 The mechanism of action of CPS-I

• CPS-I represents the committed step in urea cycle
• Activated by N-acetylglutamate
– Because N-acetylglutamate is a precursor to orinithine synthesis and essential to the operation of the urea cycle
amino acid catabolism ↑
glutamate level (N-acetylglutamate) ↑
Stimulate CPS-I
Raise overall Urea cycle activity

1. Ornithine transcarbamoylase (OTCase)
2. Argininosuccinate synthetase
3. Argininosuccinase
4. Arginase
Urea Cycle

The Urea Cycle• The carbon skeleton of arginine is derived
from -ketoglutarate (Ornithine)• N and C in the guanidino group of Arg
come from NH4+, HCO3
- (carbamoyl-P), and the -NH2 of Glu and Asp
• Breakdown of Arg in the urea cycle releases two N and one C as urea
• Important N excretion mechanism in livers of terrestrial vertebrates
• Urea cycle is linked to TCA by fumarate

Lysine Biosynthesis
• Two pathways:1. -aminoadipate pathway
2. diaminopimelate pathway (Asp)
• Lysine derived from -ketoglutarate– Reactions 1 through 4 are reminiscent of the first four
reactions in the citric acid cycle -ketooadipate
• Transamination gives -aminoadipate• Adenylylation activates the -COOH for reduction• Reductive amination give saccharopine• Oxidative cleavage yields lysine

Figure 25.24 Lysine biosynthesis in certain fungi and Euglena: the -aminoadipic acid pathway.

2. The Aspartate FamilyAsp, Asn, Lys, Met, Thr, Ile
• Transamination of Oxaloaceate gives Aspartate (aspartate aminotransferase)
• Amidation of Asp gives Asparagine ( asparagine synthetase)
• Met, Thr and Lys are made from Aspartate -Aspartyl semialdehyde and homoserine are branch
points
• Isoleucine, four of its six carbons derived from Asp (via Thr) and two come from pyruvate

Figure 25.25 Aspartate biosynthesis via transamination of oxaloacetate by glutamate.
Figure 25.26 Asparagine biosynthesis from Asp, Gln, and ATP by asparagine synthetase.

Figure 25.27 Biosynthesis of threonine, methionine, and lysine, members of the aspartate family of amino acids.
-Aspartyl-semialdehyde is a common precursor to all three.
It is formed by aspartokinase (reaction 1) and -aspartyl-semialdehyde dehydrogenase (reaction 2).

Figure 25.27 Biosynthesis of threonine, methionine, and lysine, members of the aspartate family of amino acids.

Figure 25.27 Biosynthesis of threonine, methionine, and lysine, members of the aspartate family of amino acids.

-Aspartyl-semialdehyde
Figure 25.27 Biosynthesis of threonine, methionine, and lysine, members of the aspartate family of amino acids.

In E. coli, The first reaction is an ATP-dependent phosphorylation catalyzed by aspartokinase– Three isozymes of aspartokinase (I, II, and III)– Uniquely controlled by one of the three end-
products– Form I is feedback-inhibited by threonine– Form III is feedback-inhibited by lysine


Figure 25.28 The synthesis of S-adenosylmethionine (SAM)
Important role of methionine
• in methylations via S-adenosylmethionine (SAM; S-AdoMet)
• polyamine biosynthesis

3. The Pyruvate FamilyAla, Val, Leu, and Ile
• Transamination of pyruvate gives Alanine• Valine is derived from pyruvate• Ile synthesis from Thr mimics Val synthesis
from pyruvate (Fig. 25.29)– Threonine deaminase (also called threonine
dehydratase or serine dehydratase) is sensitive to Ile– Ile and val pathway employ the same set of enzymes
• Leu synthesis begins with an -keto isovalerate– Isopropylmalate synthase is sensitive to Leu
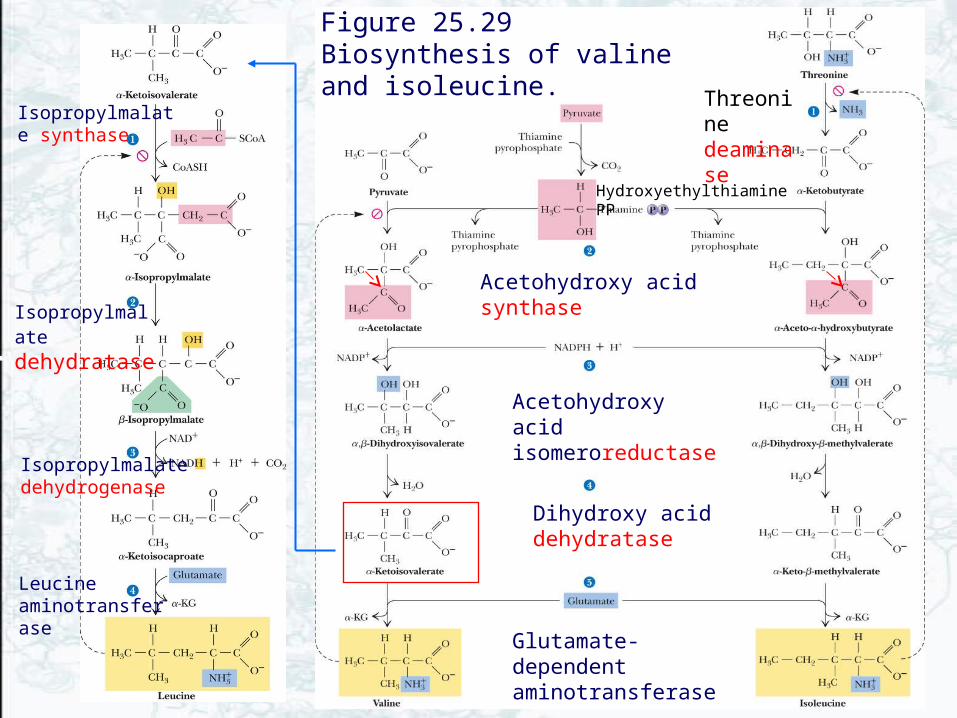
Acetohydroxy acid isomeroreductase
Threonine deaminase
Acetohydroxy acid synthase
Dihydroxy acid dehydratase
Glutamate-dependent aminotransferase
Figure 25.29 Biosynthesis of valine and isoleucine.
HydroxyethylthiaminePP
Isopropylmalate synthase
Isopropylmalate dehydrogenase
Isopropylmalate dehydratase
Leucine aminotransferase

4. 3-Phosphoglycerate FamilySer, Gly, Cys
1. 3-Phosphoglycerate dehydrogenase diverts 3-PG from glycolysis to amino acid synthesis pathways (3-phosphohydroxypyruvate)
2. Transamination by Glu gives 3-phosphoserine (3-phosphoserine aminotransferase)
3. Phosphoserine phosphatase yields serine

• Serine hydroxymethylase (PLP-dependent) transfers the -carbon of Ser to THF to make glycine
Figure 25.32 Biosynthesis of glycine from serine (a) via serine hydroxymethyltransferase and (b) via glycine oxidase.

• A PLP-dependent enzyme makes Cys
Some bacteria
most microorganism and plants
Figure 25.33 Cysteine biosynthesis. (a) Direct sulfhydrylation of serine by H2S. (b) H2S-dependent sulfhydrylation of O-acetylserine.
serine acetyltransferase
O-acetylserine sulfhydrylase

Figure 25.34 Sulfate assimilation and the generation of sulfide for synthesis of organic S compounds.
ATP sulfurylase
Adenosine-5'-phosphosulfate-3'-phosphokinase.
Sulfite oxidase

5. Aromatic Amino AcidsPhe, Tyr, Trp, His
• The aromatic amino acids, Phe, Tyr, and Trp, are derived from shikimate pathway yields chorismate, thence Phe, Tyr, Trp
• Chorismate as a branch point in this pathway (Figs. 25.35)– Chorismate is synthesized from PEP and
erythrose-4-P – Via shikimate pathway – The side chain of chorismate is derived from a
second PEP

Figure 25.35 Some of the aromatic compounds derived from chorismate.

(1) 2-keto-3-deoxy-D-arabino-heptulosonate-7-P synthase
(4) shikimate dehydrogenase
(5) shikimate kinase
(6) 3-enolpyruvyl-shikimate-5-phosphate synthase
(7) chorismate synthase.
(2) dehydroquinate synthase
(3) 5-dehydroquinate dehydratase

The Biosynthesis of Phe, Tyr, and Trp
• At chorismate, the pathway separates into three branches, each leading to one of the aromatic amino acids
• Mammals can synthesize tyrosine from phenylalanine by phenylalanine hydroxylase (Phenylalanine-4-monooxygenase)
Figure 25.38 The formation of tyrosine from phenylalanine.

Figure 25.37 The biosynthesis of phenylalanine, tyrosine, and tryptophan from chorismate.
(1) chorismate mutase
(2) prephenate dehydratase
(3) phenylalanine aminotransferase
(4) prephenate dehydrogenase
(5) tyrosine aminotransferase
(6) anthranilate synthase
(7) anthranilate-phosphoribosyl transferase
(8) N-(5'-phosphoribosyl)-anthranilate isomerase
(9) indole-3-glycerol phosphate synthase
(10) tryptophan synthase (-subunit)
(11) tryptophan synthase (-subunit).

Histidine Biosynthesis
• His synthesis, like that of Trp, shares metabolic intermediates (PRPP) with purine biosynthetic pathway
• His operon
• Begin from PRPP and ATP
• The intermediate 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR) is a purine precursor (replenish ATP; Ch 26)

Figure 25.40 The pathway of histidine biosynthesis.
(1) ATP-phosphoribosyl transferase
(2) pyrophosphohydrolase
(3) phosphoribosyl-AMP cyclohydrolase
(4) phosphoribosylformimino-5-aminoimidazole carboxamide ribonucleotide isomerase
(5) glutamine amidotransferase
(6) imidazole glycerol-P dehydratase
(7) L-histidinol phosphate aminotransferase
(8) histidinol phosphate phosphatase
(9) histidinol dehydrogenase.

Amino Acid Biosynthesis Inhibitors as Herbicides
A variety of herbicides have been developed as inhibitors of plant enzymes that synthesize “essential” amino acids
• These substances show no effect on animals
• For example, glyphosate, sold as RoundUp, is a PEP analog that acts as an uncompetitive inhibitor of 3-enolpyruvylshikimate-5-P synthase.
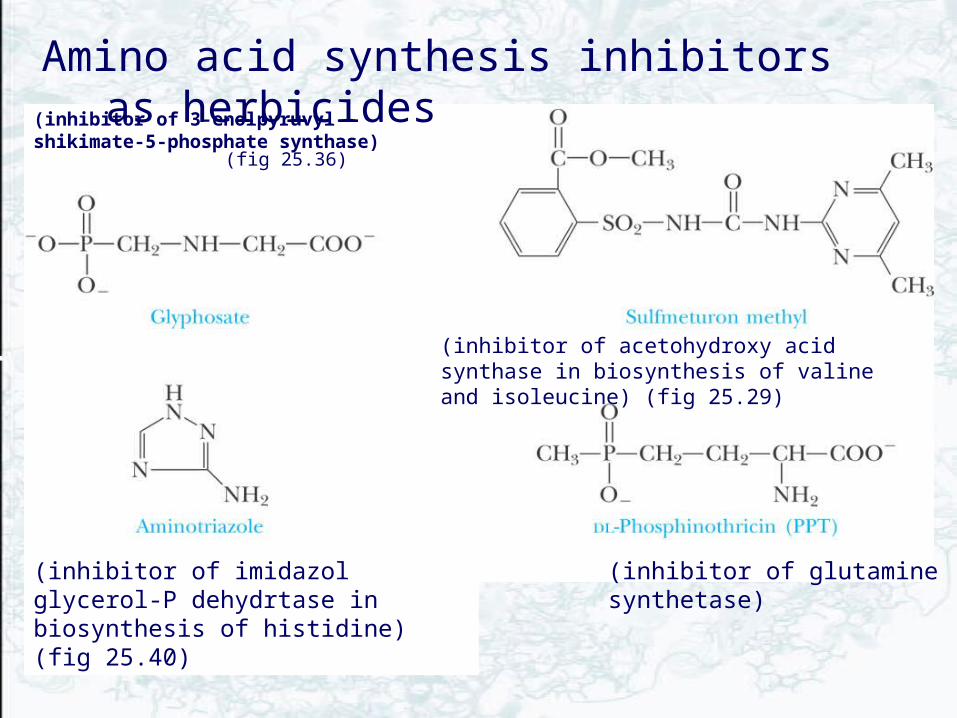
(inhibitor of 3-enolpyruvyl-shikimate-5-phosphate synthase)
(fig 25.36)
(inhibitor of acetohydroxy acid synthase in biosynthesis of valine and isoleucine) (fig 25.29)
(inhibitor of glutamine synthetase)(inhibitor of imidazol glycerol-P dehydrtase in biosynthesis of histidine) (fig 25.40)
Amino acid synthesis inhibitors as herbicides

25.5 – Degradation of Amino Acids
The 20 amino acids are degraded to produce (mostly) TCA intermediates
• The primary physiological purpose of amino acids is to serve as building blocks for protein synthesis
• Energy requirement– 90% from oxidation of carbohydrates and fats– 10% from oxidation of amino acids
• The classifications of amino acids in Fig. 25.41 • Glucogenic and ketogenic
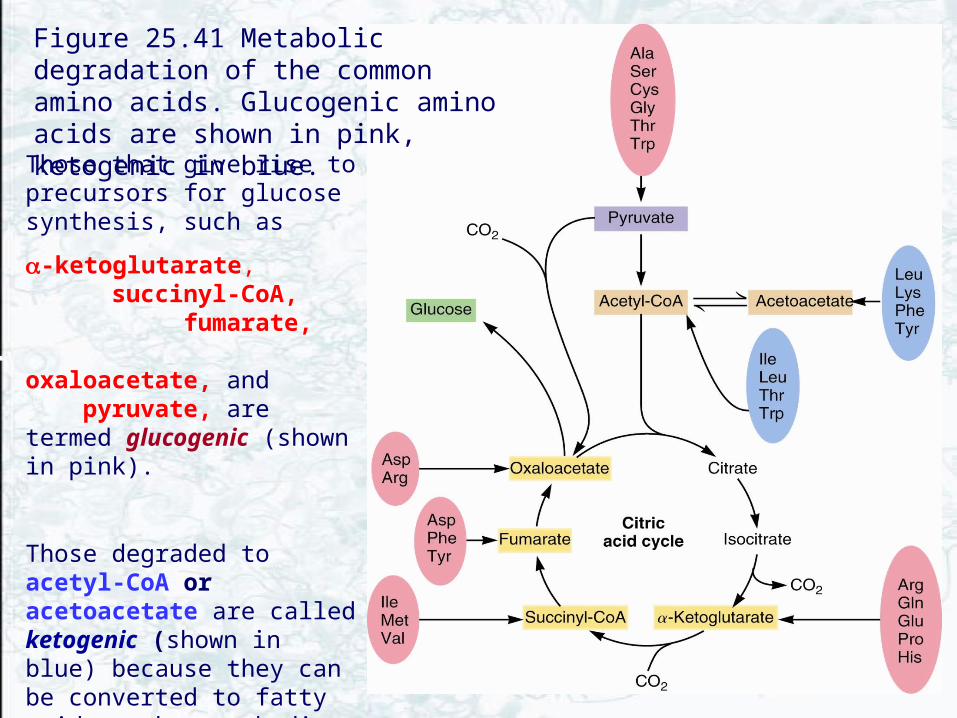
Those that give rise to precursors for glucose synthesis, such as
-ketoglutarate, succinyl-CoA, fumarate, oxaloacetate, and pyruvate, are termed glucogenic (shown in pink).
Those degraded to acetyl-CoA or acetoacetate are called ketogenic (shown in blue) because they can be converted to fatty acids or ketone bodies.
Some amino acids are both glucogenic and ketogenic.
Figure 25.41 Metabolic degradation of the common amino acids. Glucogenic amino acids are shown in pink, ketogenic in blue.

The 20 amino acids are degraded by 20 different pathways that converge to just 7 metabolic intermediates

C-3 family (pyruvate):Ala, Ser, Cys, Gly, Thr, Trp
C-4 family (oxaloaceate & fumarate):Oxaloaceate: Asp, Asn
Fumarate: Asp, Phe, Tyr
C-5 family (-ketoglutarate):Glu, Gln, Arg, Pro, His
Succinyl-CoA:Ile, Met, Val
Acetyl-CoA & acetoacetateIle, Leu, Thr, Trp
Leu, Lys, Phe, Tyr

C-3 family:Ala, Ser, Cys, Gly, Thr,
Trp
Figure 25.42 Formation of pyruvate from alanine, serine, cysteine, glycine, tryptophan, or threonine.

Figure 25.43 The degradation of the C-5 family of amino acids leads to -ketoglutarate via glutamate. The histidine carbons, numbered 1 through 5, become carbons 1 through 5 of glutamate, as indicated.

Figure 25.44 Valine, isoleucine, and methionine are converted via propionyl-CoA to succinyl-CoA for entry into the citric acid cycle.
The shaded carbon atoms of the three amino acids give rise to propionyl-CoA.
All three amino acids lose their -carboxyl group as CO2.
Methionine first becomes S-adenosylmethionine, then homocysteine (see Figure 25.28).
The terminal two carbons of isoleucine become acetyl-CoA.

Figure 25.45 Leucine is one of only two purely ketogenic amino acids; the other is lysine. Deamination of leucine via a transamination reaction yields α-ketoisocaproate, which is oxidatively decarboxylated to isovaleryl-CoA. Subsequent reactions give β-hydroxy-β-methylglutaryl-CoA, which is then cleaved to yield acetyl-CoA and acetoacetate, a ketone body.
Leucine is Degraded to Acetyl-CoA and Acetoacetate

Hereditary defects in BCKDH leads to maple sugar urine disease
Unlike the other 17 amino acids, which are broken down in the liver, Val, Ile, and Leu are also degraded in adipose tissue.

Figure 25.47 Lysine is degraded through saccharopine and α-aminoadipate to α-ketoadipate. Oxidative decarboxylation yields glutaryl-CoA, which can be transformed into acetoacetyl-CoA and then acetoacetate.
The Predominant Pathway of Lysine Degradation is the Saccharopine Pathway

Phenylalanine and Tyrosine Are Degraded to Acetoacetate and Fumarate
• The first reaction in phenylalanine degradation is the hydroxylation reaction of tyrosine biosynthesis• Both these amino acids thus share a common
degradative pathway• Transamination of tyrosine yields p-
hydroxyphenylpyruvate• A vitamin C-dependent dioxygenase then produces
homogentisate• Ring opening and isomerization gives 4-fumaryl-
acetoacetate, which is hydrolyzed to acetoacetate and fumarate
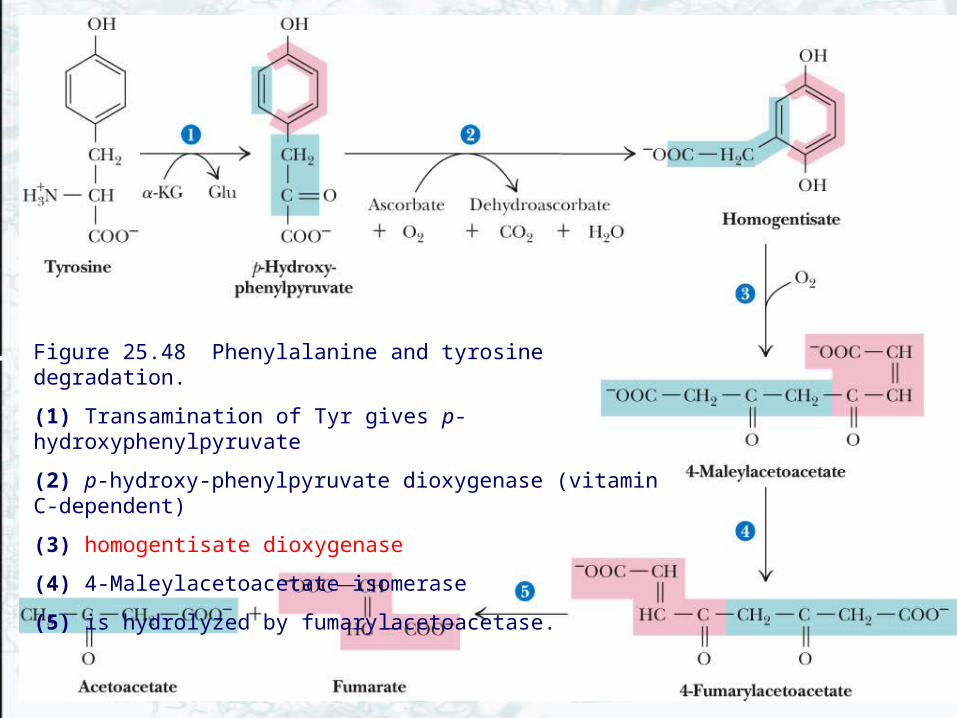
Figure 25.48 Phenylalanine and tyrosine degradation.
(1) Transamination of Tyr gives p-hydroxyphenylpyruvate
(2) p-hydroxy-phenylpyruvate dioxygenase (vitamin C-dependent)
(3) homogentisate dioxygenase
(4) 4-Maleylacetoacetate isomerase
(5) is hydrolyzed by fumarylacetoacetase.

Tryptophan is a crucial precusor for synthesis of a variety of important substances•Serotonin (5-hydroxytryptophan) is a neurotransmitter•Melatonin (N-acetyl-5-methoxytrptophan) is a hormone

Maple syrup urine disease– After the initial step (deamination) to produce -keto
acids– The defect in oxidative decarboxylation of Ile, Leu,
and Val (25.44)
Phenylketonuria– The defect in phenylalanine hydoxylase (25.38)– Accumulation of phenylpyruvate
Alkaptouria – Homogentisate dioxygenase (25.47)
Hereditary defects

Ammonotelic:– Ammonia– Aquatic animals
Ureotelic:– Urea– Terrestrial vetebrates
Uricotelic: – Uric acid– Birds and reptiles
Nitrogen excretion